
Method Article
Microscopia a super-risoluzione del complesso sinaptonemale all'interno della linea germinale di Caenorhabditis elegans
In questo articolo
Riepilogo
La microscopia a super-risoluzione può fornire una visione dettagliata dell'organizzazione dei componenti all'interno del complesso sinaptonemale nella meiosi. Qui, dimostriamo un protocollo per risolvere le singole proteine del complesso sinaptonemale Caenorhabditis elegans .
Abstract
Durante la meiosi, i cromosomi omologhi devono riconoscersi e aderire l'uno all'altro per consentire la loro corretta segregazione. Uno degli eventi chiave che assicura l'interazione dei cromosomi omologhi è l'assemblaggio del complesso sinaptonemale (SC) nella profase meiotica I. Anche se c'è poca omologia di sequenza tra i componenti proteici all'interno della SC tra le diverse specie, la struttura generale della SC è stata altamente conservata durante l'evoluzione. Nelle micrografie elettroniche, la SC appare come una struttura tripartita, simile a una scala, composta da elementi laterali o assi, filamenti trasversali e un elemento centrale.
Tuttavia, identificare con precisione la localizzazione dei singoli componenti all'interno del complesso mediante microscopia elettronica per determinare la struttura molecolare della SC rimane difficile. Al contrario, la microscopia a fluorescenza consente l'identificazione di singoli componenti proteici all'interno del complesso. Tuttavia, poiché l'SC è largo solo ~ 100 nm, la sua sottostruttura non può essere risolta dalla microscopia a fluorescenza convenzionale limitata dalla diffrazione. Pertanto, la determinazione dell'architettura molecolare della SC richiede tecniche di microscopia ottica a super-risoluzione come la microscopia a illuminazione strutturata (SIM), la microscopia a deplezione a emissione stimolata (STED) o la microscopia a localizzazione a singola molecola (SMLM).
Per mantenere la struttura e le interazioni dei singoli componenti all'interno della SC, è importante osservare il complesso in un ambiente vicino al suo ambiente nativo nelle cellule germinali. Pertanto, dimostriamo un protocollo di immunoistochimica e imaging che consente lo studio della sottostruttura della SC in tessuto germinale intatto ed estruso di Caenorhabditis elegans con microscopia SMLM e STED. Il fissaggio diretto del tessuto al vetrino riduce il movimento dei campioni durante l'imaging e riduce al minimo le aberrazioni nel campione per ottenere l'alta risoluzione necessaria per visualizzare la sottostruttura del SC nel suo contesto biologico.
Introduzione
Ridurre il numero di cromosomi della metà durante la meiosi è la chiave per generare una progenie sana negli organismi che si riproducono sessualmente. Per ottenere questa riduzione del numero di cromosomi, i cromosomi omologhi devono accoppiarsi e segregare durante la meiosi I. Per garantire l'accurata segregazione dei cromosomi omologhi, le cellule germinali subiscono una profase I estesa, durante la quale i cromosomi omologhi si accoppiano, sinapsi e si ricombinano per generare collegamenti fisici tra omologhi1. La SC è emersa come la struttura centrale che è la chiave per regolare la corretta progressione attraverso la profase meiotica2.
La SC è un complesso la cui struttura generale è evolutivamente conservata, anche se c'è poca omologia tra le sue componenti proteiche. La SC è stata identificata per la prima volta nelle micrografie elettroniche come una struttura tripartita, simile a una scala, costituita da due elementi laterali o assi, una regione centrale formata da filamenti trasversali e un elemento centrale 3,4. Determinare l'organizzazione dei singoli componenti all'interno del complesso è la chiave per far progredire la nostra comprensione del ruolo del SC durante la profase meiotica.
L'organismo modello C. elegans è ideale per studiare la struttura e la funzione della SC poiché le sue linee germinali contengono un gran numero di nuclei meiotici con SC 5 completamente assemblati. Studi genetici e biochimici hanno rivelato che gli assi cromosomici sono formati da tre distinti complessi di coesina6,7 e quattro proteine del dominio HORMA chiamate HTP-1/2/3 e HIM-3 7,8,9,10,11 in C. elegans. Nella regione centrale del SC, sei proteine contenenti domini a spirale sono state identificate fino ad oggi 12,13,14,15,16,17. Per colmare la distanza tra i due assi, SYP-1, -5 e -6 dimerizzano in modo testa a testa (Figura 1), mentre tre proteine aggiuntive stabilizzano la loro interazione nell'elemento centrale16,17,18,19.
Ottenere informazioni dettagliate sull'organizzazione di queste proteine è essenziale per comprendere le molte funzioni della SC durante la meiosi. Poiché la larghezza della regione centrale del SC è solo ~ 100 nm, la sua sottostruttura non può essere risolta mediante microscopia a fluorescenza limitata dalla diffrazione. Tuttavia, la visualizzazione di componenti all'interno di una struttura di queste dimensioni è facilmente realizzabile con la microscopia a super-risoluzione. Infatti, la microscopia a illuminazione strutturata (SIM), la microscopia ad espansione 20, la microscopia a deplezione a emissione stimolata (STED)21 e la microscopia a localizzazione a singola molecola (SMLM)22,23 sono emerse come strumenti essenziali per studiare l'architettura molecolare della SC tra le specie 16,24,25,26,27,28,29, 30.
Per superare il limite di risoluzione, la microscopia STED si basa sulla sovrapposizione del punto limitato dalla diffrazione della luce di emissione con un raggio a forma di ciambella dal laser STED, che teoricamente restringe la funzione di diffusione del punto fino alle dimensioni molecolari31,32. Tuttavia, la risoluzione che è praticamente raggiungibile da STED all'interno di campioni biologici rimane nell'intervallo di poche decine di nanometri in xy33.
Una risoluzione ancora più elevata nei campioni biologici può essere ottenuta con le tecniche SMLM. SMLM sfrutta le proprietà lampeggianti di specifici fluorofori per risolvere oggetti a livello di sub-diffrazione separando nel tempo i fluorofori spazialmente sovrapposti. Il campione viene quindi ripreso ripetutamente per catturare diversi sottoinsiemi di fluorofori. La posizione dei fluorofori all'interno del campione viene quindi determinata adattando la funzione di diffusione puntuale (PSF) ai segnali ottenuti in tutte le immagini, che possono risolvere strutture fino a 15 nm 23,34.
Nel loro insieme, le immagini localizzate codificano le posizioni di tutti i fluorofori. La risoluzione di SMLM è determinata dalla densità di etichettatura e dalle caratteristiche lampeggianti del fluoroforo. Secondo il criterio di Nyquist-Shannon, è impossibile risolvere in modo affidabile oggetti che sono meno del doppio della distanza media da etichetta a etichetta. Pertanto, è necessaria un'elevata densità di etichettatura per l'imaging ad alta risoluzione. Per la SC in C. elegans, un'alta densità di marcatura può essere ottenuta utilizzando tag epitopi attaccati a siti specifici di proteine endogene utilizzando l'editing del genoma. Le etichette epitopiche possono quindi essere colorate ad alta densità utilizzando anticorpi monoclonali specifici con alta affinità19,30. Allo stesso tempo, il ciclo di accensione dei singoli fluorofori deve essere abbastanza breve da garantire che i fluorofori spazialmente sovrapposti non vengano catturati contemporaneamente35.
A causa di questi due requisiti, risolvere la struttura di grandi complessi macromolecolari come la SC richiede l'imaging di un numero sufficientemente grande di immagini e può quindi richiedere diverse ore. La trappola dei lunghi tempi di imaging è che i campioni tendono ad andare alla deriva a causa del movimento dello stadio o di piccole correnti all'interno del buffer del campione; Anche piccoli movimenti dell'ordine di 10 nm sono dannosi alla risoluzione nm e devono essere corretti. Tuttavia, i metodi di correzione della deriva comunemente usati non sono abbastanza robusti da sovrapporre accuratamente le immagini di due canali ripresi in sequenza36. Questo è problematico perché le domande biologiche spesso richiedono una rilevazione precisa e la localizzazione di più bersagli all'interno dello stesso campione. Per aggirare questi problemi, sono stati sviluppati metodi come l'imaging raziometrico. L'imaging raziometrico consente l'imaging simultaneo di più fluorofori con spettri di eccitazione ed emissione sovrapposti, con una successiva assegnazione di ciascun segnale rilevato al rispettivo fluoroforo in base al rapporto di intensità in canali spettralmente distinti37,38.
Inoltre, lo studio dell'organizzazione di complessi macromolecolari come la SC richiede informazioni tridimensionali (3D). Per ottenere una super-risoluzione in tre dimensioni (3D-SMLM), una lente cilindrica è incorporata nel percorso ottico della luce emessa che distorce la forma della PSF di un fluoroforo a seconda della sua distanza dal piano focale. Quindi, la posizione precisa di un fluoroforo nel piano z può essere estrapolata analizzando la forma del suo segnale di emissione35,39. La combinazione di questi progressi in SMLM consente l'imaging dell'organizzazione 3D di complessi macromolecolari, incluso il SC.
Protocollo
1. Preparazione di soluzioni e vetrini
NOTA: Vedere la Tabella dei materiali per i dettagli relativi a tutti i materiali e reagenti e la Tabella 1 per la composizione delle soluzioni utilizzate in questo protocollo.
- Coprivetrini rivestiti in poli-L-lisina
- Preparare lo 0,01% (p/v) di poli-L-lisina (vedere Tabella 1).
- Lavare un coprislip di precisione (diametro 24 mm; 0,17 ± 0,005 mm, n. 1,5) in etanolo per 10-30 minuti. Risciacquare il coprivetrino con ddH2O per rimuovere l'etanolo e lasciare asciugare il coprislip a temperatura ambiente.
- Pulire al plasma il vetrino usando un detergente al plasma.
NOTA: La pulizia al plasma aumenta l'idrofilia del coprislip e facilita i passaggi successivi. Se non è disponibile un detergente al plasma, questo passaggio può essere saltato, anche se ciò potrebbe richiedere la regolazione del volume e/o della concentrazione della soluzione di poli-L-lisina. Questa modifica non è stata testata. - Introdurre una goccia (120 μL) di poli-L-lisina allo 0,01% (p/v) sul vetrino. Incubare per 10 minuti a temperatura ambiente.
- Dopo l'incubazione, sciacquare il vetrino di copertura in ddH2O e asciugarlo a temperatura ambiente. Conservare a 4 °C fino a 1 mese.
- Frammenti di F(ab')2 coniugati con coloranti organici fluorescenti
- Aggiungere il seguente ordine a una provetta PCR: 10 μL di 0,6-0,7 mg/mL di frammento di F(ab')2 in PBS, 1 μL di 0,1 M NaHCO 3 (pH8,3 ) e 1 μL di 1 mM di fluoroforo reattivo all'estere succinimidil (NHS) nel DMSO (rapporto molare di F(ab')2:colorante è ~1:17). Mescolare bene pipettando su e giù.
- Incubare per 1 ora a temperatura ambiente.
- Separare il frammento F(ab')2 dal colorante reattivo libero rimanente utilizzando una colonna di desalinizzazione (7K MWCO) seguendo le specifiche del produttore. Utilizzare 1x PBS per l'equilibrio della colonna e l'eluizione del frammento F(ab')2 marcato.
- Conservare il frammento F(ab')2 marcato a 4 °C per un massimo di 3 mesi.
NOTA: i tempi di conservazione superiori a 3 mesi non sono stati testati.
2. Dissezione e fissazione
NOTA: Le procedure di dissezione e fissazione sono modificate rispetto alle procedure precedentemente raccomandate16,40 per ottenere campioni ottimali per la microscopia a super-risoluzione.
- Dissezione
- Prelevare i vermi C. elegans di pari età (cresciuti a 20 °C per questo studio) in una goccia di 30 μL di EBTT (1x tampone per uova41 con detergente non ionico allo 0,2%, Tabella 1) su un vetrino (22 mm x 22 mm, n. 1). Posizionare il vetrino su un vetrino per facilitarne la manipolazione. Lavare con 30 μL di EBTT mediante pipettaggio su e giù più volte. Rimuovere 30 μL della soluzione per lasciare una goccia di 30 μL sul vetrino.
NOTA: Piccole quantità di detergente non ionico devono essere aggiunte a tutte le soluzioni in cui i vermi sono pipettati per evitare che i vermi si attacchino alle punte di plastica. - Utilizzare una lama di bisturi per tagliare le teste e/o le code dei vermi per estrudere la gonade (Figura 2A).
- Prelevare i vermi C. elegans di pari età (cresciuti a 20 °C per questo studio) in una goccia di 30 μL di EBTT (1x tampone per uova41 con detergente non ionico allo 0,2%, Tabella 1) su un vetrino (22 mm x 22 mm, n. 1). Posizionare il vetrino su un vetrino per facilitarne la manipolazione. Lavare con 30 μL di EBTT mediante pipettaggio su e giù più volte. Rimuovere 30 μL della soluzione per lasciare una goccia di 30 μL sul vetrino.
- Fissazione
- Pipet 30 μL di soluzione fissativa (Tabella 1) nella goccia dei vermi sezionati e pipettare su e giù per mescolare.
NOTA: Il pipettaggio su e giù alcune volte può aiutare a rilasciare più gonadi. - Fissare esattamente per 1 minuto dopo aver aggiunto la soluzione fissativa.
- Interrompere la fissazione trasferendo i vermi in un tubo PCR riempito con TBST (Tabella 1). Trasferire i vermi nel minor volume possibile (~15 μL).
- Far girare il tubo PCR su una mini centrifuga da banco (2.000 × g, 10 s). Rimuovere il surnatante e lavare 2 volte con 200 μL di TBST ciascuno.
- Lavare con 200 μL di PBST (Tabella 1) per 5-10 min. Ripetere i passaggi da 2.1.1 a 2.2.5 per un massimo di quattro campioni mantenendo i campioni sezionati sul ghiaccio.
NOTA: se si elaborano più di quattro campioni, procedere con i passaggi da 2.2.6 a 2.2.7 dopo ogni quattro campioni per garantire che la fissazione del campione rimanga coerente in tutti i campioni. - Far girare i campioni sezionati su una mini centrifuga da banco (2.000 × g, 10 s), rimuovere il PBST e aggiungere 50-100 μL di metanolo freddo (-20 °C).
ATTENZIONE: Il metanolo è tossico. Indossare dispositivi di protezione ed evitare l'inalazione. - Mescolare mediante pipettaggio su e giù e lasciare i campioni in metanolo per 30-60 s. Lavare i campioni 2x in 200 μL di PBST.
NOTA: se si elaborano più di quattro campioni, procedere con la dissezione dei campioni rimanenti (punti da 2.1.1 a 2.2.7). - Lavare i campioni una terza volta con 200 μL di PBST.
- Pipet 30 μL di soluzione fissativa (Tabella 1) nella goccia dei vermi sezionati e pipettare su e giù per mescolare.
3. Incubazioni di anticorpi
- Inceppamento
- Bloccare i campioni in 1x Soluzione di blocco (Tabella 1) per 45-60 minuti a temperatura ambiente.
NOTA: Il tempo di incubazione può variare da 30 minuti a temperatura ambiente a diversi giorni a 4 °C (il test è stato effettuato fino a 3 giorni).
- Bloccare i campioni in 1x Soluzione di blocco (Tabella 1) per 45-60 minuti a temperatura ambiente.
- Soluzione anticorpale primaria
- Diluire gli anticorpi anti-HTP-3 (pollo42) e anti-HA (topo) (o gli anticorpi di scelta) alle soluzioni di lavoro (1:250 per SMLM e 1:1.000 per campioni di microscopia STED) in 1x Soluzione di blocco.
NOTA: Gli anticorpi utilizzati per marcare i campioni SMLM sono più concentrati rispetto ai campioni STED poiché per la microscopia SMLM è raccomandata una densità di etichettatura più elevata. - Centrifugare i campioni su una mini centrifuga da banco (2.000 × g, 10 s), rimuovere il tampone bloccante e aggiungere 30-50 μL della soluzione anticorpale primaria. Incubare per una notte a 4 °C (preferibile) o per 1-2 ore a temperatura ambiente.
- Dopo l'incubazione, lavare 3 x 5-15 minuti con PBST.
- Diluire gli anticorpi anti-HTP-3 (pollo42) e anti-HA (topo) (o gli anticorpi di scelta) alle soluzioni di lavoro (1:250 per SMLM e 1:1.000 per campioni di microscopia STED) in 1x Soluzione di blocco.
- Soluzione di lavoro di frammenti F(ab')2 coniugati a colorante fluorescente
- Diluire i frammenti F(ab')2 marcati (fase 1.2.4) nelle soluzioni di lavoro (1:100 per SMLM e 1:1.000 per campioni di microscopia STED) in 1x soluzione bloccante.
NOTA: Per entrambe le tecniche di super-risoluzione, sono state utilizzate coppie di fluorofori precedentemente riportate, ovvero AlexaFluor647/CF680 per SMLM e AlexaFluor594/Abberior STAR635P per STED. AlexaFluor647 e STAR645P sono stati utilizzati per marcare frammenti anti-topo (Fab')2 per colpire il C-terminus di SYP-5 e CF680/AlexaFluor594-marcati anti-pollo (Fab')2 frammenti per colpire HTP-3. - Far girare i campioni su una mini centrifuga da banco (2.000 × g, 10 s), rimuovere il PBST e aggiungere 30-50 μL di soluzione anticorpale secondaria. Incubare per 30 minuti a 2 ore a temperatura ambiente (preferito) o per una notte a 4 °C. Lavare 3 x 5-15 minuti con PBST.
- Diluire i frammenti F(ab')2 marcati (fase 1.2.4) nelle soluzioni di lavoro (1:100 per SMLM e 1:1.000 per campioni di microscopia STED) in 1x soluzione bloccante.
4. Montaggio dei campioni su un vetrino
- Postfissazione
NOTA: Processare i campioni singolarmente attraverso i passaggi 4.1.1-4.2.1.- Ruotare i campioni macchiati e rimuovere il surnatante. Aggiungere 50 μL di PBST0,2 e trasferire i vermi colorati su un coprivetrino n. 1 da 22 x 22 mm.
NOTA: Utilizzare PBST 0,2 fresco con detergente non ionico allo0,2 % (Tabella 1) per questo passaggio per evitare che i vermi aderiscano al vetrino. - Pipet 5,7-6,3 μL di soluzione postfissativa su un coprivetrino di poli-L-lisina.
NOTA: I vetrini di poli-L-lisina conservati a 4 °C devono essere portati prima a temperatura ambiente. - Pipet rimuove i vermi sezionati nello stesso volume (5,7-6,3 μL) e trasferiscono nella goccia di fissativo sul coprislip di poli-L-lisina (Figura 2B).
NOTA: In questa e nella fase successiva, è molto importante mantenere il tessuto sezionato al centro del coprislip rivestito di poli-L-lisina. Ciò è particolarmente importante se si montano i campioni in un supporto personalizzato per adattarsi al microscopio SMLM personalizzato utilizzato qui (vedere il passaggio 5.1, Figura 2B). - Coprire il campione con un piccolo vetrino di copertura (diametro 12 mm, figura 2B). Rimuovere il liquido in eccesso utilizzando un piccolo pezzo di carta da filtro (Figura 2B). Fissare per 3-5 minuti in una camera buia.
- Ruotare i campioni macchiati e rimuovere il surnatante. Aggiungere 50 μL di PBST0,2 e trasferire i vermi colorati su un coprivetrino n. 1 da 22 x 22 mm.
- "Freeze-cracking"
- Congelare i campioni posizionandoli su un blocco di alluminio in ghiaccio secco (Figura 2B).
NOTA: Il blocco di alluminio deve essere ben raffreddato nel ghiaccio secco prima di posizionare i campioni su di esso. Procedere con la postfissazione dei campioni rimanenti (punti da 4.1.1 a 4.2.1). Il campione deve essere su ghiaccio secco per almeno 20 minuti o fino a 1 ora prima della fase successiva (4.2.2). - Rimuovere il coprislip più piccolo con un rasoio (Figura 2B).
NOTA: per STED, procedere al passaggio 5.2.1. Per SMLM, continuare con il passaggio 4.2.3. - Immergere il foglietto in un tubo conico da 50 mL contenente PBS ghiacciato (preferito) o metanolo a -20 °C per circa 10 s.
NOTA: La temperatura è un fattore molto importante per questo passaggio. Pertanto, utilizzare PBS appena scongelato o conservato in un bagno di ghiaccio / etanolo. - Posizionare il coprislip in un pozzetto di una piastra a sei pozzetti riempita con tampone PBST. Rimuovere il PBST dai pozzetti e aggiungere PBS fresco. Lasciare i campioni in PBS per 5 minuti.
NOTA: Pipet il PBS sul lato del pozzo per evitare di danneggiare e staccare i campioni. - Lavare con PBS fresco e lasciare i campioni a 4 °C fino all'imaging.
NOTA: i campioni sono stabili per un massimo di 2 settimane, ma i migliori risultati si ottengono se i campioni vengono ripresi entro 2 giorni. - Prima dell'imaging, valutare la qualità del montaggio del campione al microscopio stereo.
NOTA: le linee germinali montate con successo sono fissate stabilmente senza alcun movimento visibile rispetto al vetrino. Le linee germinali scarsamente attaccate si sbattono nella soluzione tampone.
- Congelare i campioni posizionandoli su un blocco di alluminio in ghiaccio secco (Figura 2B).
5. Imaging
- Microscopia di localizzazione a singola molecola
NOTA: Le immagini sono state acquisite presso l'EMBL Imaging Centre utilizzando un microscopio di localizzazione a singola molecola costruito su misura che è stato costruito attorno a un corpo personalizzato, come precedentemente riportato38,43, con le caratteristiche specifiche specificate nella Tabella dei materiali; fare riferimento a https://www.embl.org/about/info/imaging-centre- Acquisizione della calibrazione 3D delle perline
- Preparare un coprislip di precisione (diametro 24 mm; 0,17 ± 0,005 mm, n. 1,5) con perline fluorescenti aderenti da 100 nm come descritto in precedenza38,44.
- Posizionare il campione di taratura del punto 5.1.1.1 su un portacampioni.
- Aggiungere una goccia di olio ad immersione sull'obiettivo pulito 100x/1,5 e montare il campione di calibrazione sul microscopio.
- All'interno di MicroManager 2 45,46, specificare15-20 posizioni nel campione di calibrazione.
- All'interno della finestra47 del plugin EMU, impostare l'acquisizione di un'immagine z-stack per ciascuna delle posizioni dal punto 5.1.1.5.
NOTA: Qui, una lente cilindrica composta fornisce l'astigmatismo richiesto per l'imaging 3D e sono state acquisite 201 sezioni z per ogni posizione che copre l'intervallo compreso tra -1 μm e 1 μm, con un incremento di 10 nm. È stata utilizzata un'illuminazione laser da 2 kW/cm2 640 nm per 25 ms per ogni z-slice. - Acquisire le immagini z-stack delle sfere fluorescenti da 100 nm attraverso un percorso ottico identico che verrà utilizzato per acquisire immagini campione nel passaggio 5.1.11.
- Utilizzando la piattaforma di analisi al microscopio a super-risoluzione (SMAP48), generare un modello cspline della funzione di diffusione del punto sperimentale (PSF) che verrà utilizzato per adattare i dati 3D-SMLM nella fase 5.1.13.
- Preparare il portacampioni. Per il supporto personalizzato utilizzato qui che utilizza un anello magnetico per creare la camera di imaging (Figura 2B), avvolgere l'anello magnetico con parafilm.
NOTA: In alternativa, un vetrino per microscopio con una cavità di depressione concava può essere utilizzato per montare campioni per microscopi con supporti per vetrino. - Preparare 1 mL di tampone di imaging44 (Tabella 1).
- Prendete un foglietto del punto 4.2.6 e mettetelo nel supporto personalizzato. Fissare il vetrino di copertura nel supporto con l'anello magnetico avvolto nel parafilm (punto 5.1.2).
- Pipetare delicatamente il tampone di imaging (punto 5.1.3) nella camera creata dall'anello magnetico sulla parte superiore del campione (Figura 2B). Sigillare la camera con un pezzo di parafilm.
- Per montare il campione, aggiungere una goccia di olio ad immersione sull'obiettivo dell'olio pulito 100x/1,5. Senza introdurre aria nell'olio per immersione, posizionare delicatamente il portacampioni con il campione montato (fase 5.1.5) sul palco del microscopio.
NOTA: Prima di posizionare il campione sul microscopio, pulire il fondo del vetrino con fazzoletto e etanolo al 70%. - Utilizzando la finestra47 del plugin EMU all'interno di MicroManager 245,46, spostare lo stadio piezoelettrico fino a quando il segnale del laser di blocco della messa a fuoco non viene rilevato al fotodiodo del quadrante (QPD).
NOTA: per mantenere una messa a fuoco fissa per tutto il tempo di imaging, il blocco della messa a fuoco si ottiene mediante la riflessione interna totale di un laser accoppiato a fibre nel vicino infrarosso dal coprislip e il successivo rilevamento sensibile all'altezza su un fotodiodo a quadrante (QPD). Il segnale QPD forniva il controllo ad anello chiuso dell'attacco piezoelettrico dell'obiettivo. - Acquisire un'immagine del piano focale posteriore con un laser di eccitazione a 640 nm a bassa potenza (cioè 1-5%) per confermare l'assenza di bolle d'aria nell'olio di immersione.
NOTA: rimuovere il campione dallo stage se viene rilevata una bolla d'aria. Pulire la parte inferiore del copricostume e l'obiettivo e ripetere i passaggi 5.1.6-5.1.8. In caso contrario, procedere a bloccare lo stato attivo all'interno del software EMU47. - Localizzare il tessuto gonadico utilizzando l'illuminazione a campo chiaro. Utilizzando un'illuminazione a bassa intensità a 640 nm, concentrarsi sulla sezione del tessuto che contiene molti tratti SC.
NOTA: Non mettere a fuoco strutture che si trovano a più di 2 μm dal coprivetrino. Non utilizzare una potenza laser superiore per localizzare il campione, poiché ciò potrebbe trasformare alcuni fluorofori in uno stato lampeggiante prematuramente. Qui, 1 kW / cm2 è stato utilizzato in modalità crescente con un impulso impostato su 1.000. - Procedere all'esposizione del campione con un'illuminazione di 640 nm ad alto irraggiamento (27 kW/cm2) per ~30 s fino a raggiungere una velocità di lampeggio appropriata (Video supplementare 1).
- Acquisite 200.000 fotogrammi con un tempo di esposizione di 20 ms utilizzando lo strumento di acquisizione multidimensionale di MicroManager 245,46.
- Nel frattempo, imposta l'attivazione UV utilizzando l'opzione di attivazione del plug-in EMU38,47 per mantenere la velocità di lampeggio desiderata.
NOTA: utilizzare il laser UV con un'irradianza di 3 kW/cm2 in modalità crescente con lunghezza massima dell'impulso impostata su 10.000. - Eseguire la ricostruzione e la post-elaborazione delle immagini SMLM.
NOTA: per ricostruire immagini da dati SMLM non elaborati, fare riferimento ai metodi pubblicati. I dati qui presentati sono stati elaborati utilizzando il software SMAP48,49. Nel software SMAP48 sono stati eseguiti la ricostruzione delle immagini super-risolte, l'assegnazione dei canali, la correzione della deriva e il filtraggio delle localizzazioni con scarsa precisione di localizzazione e un filtro di massima verosimiglianza.
- Acquisizione della calibrazione 3D delle perline
- Microscopia a deplezione ad emissione stimolata
NOTA: Le immagini sono state acquisite sul sistema microscopico STED integrato equipaggiato con un laser a luce bianca, un laser STED pulsato a 775 nm e il modulo di invecchiamento FALCON Fluorescence Lifetime IM(Table of Materials) presso l'EMBL Imaging Centre (https://www.embl.org/about/info/imaging-centre).- Posizionare una goccia da 20 μL di supporto di montaggio (Tabella dei materiali) su un vetrino da microscopio. Prendere un foglietto di copertina dal punto 4.2.2 e posizionare delicatamente il campione sul vetrino rivolto verso il supporto di montaggio (Figura 2B).
NOTA: Evitare di introdurre sacche d'aria all'interno del mezzo di montaggio. - Lasciare che il mezzo di montaggio polimerizzi durante la notte.
NOTA: Visualizzare i campioni il giorno successivo o mantenerli a 4 °C fino all'imaging. - Per montare il campione, aggiungere una goccia di olio ad immersione sul coperchio del campione dal punto 5.2.2. Posizionare delicatamente il campione sul palco del microscopio utilizzando un obiettivo per olio 100x/1,40.
- Concentrarsi sul campione e localizzare il tessuto germinale utilizzando l'illuminazione a campo luminoso.
- Utilizzando il software del microscopio, specificare la regione di interesse per la quale verrà acquisita l'immagine TauSTED.
- Selezionare i laser di eccitazione e la loro potenza appropriata utilizzata per eccitare i fluorofori utilizzati nel campione.
NOTA: Qui, il laser a 580 nm al 4% di potenza è stato utilizzato per visualizzare frammenti di anticorpi secondari coniugati AlexaFluor 594 F(ab')2 e 635 nm al 3% di potenza per visualizzare frammenti F(ab')2 coniugati con STAR 635P. - Utilizzando il software del microscopio, selezionare una potenza laser di esaurimento STED appropriata e impostare il rilevamento dell'immagine.
NOTA: Qui, la potenza del laser a esaurimento STED da 775 nm è stata impostata al 40%. Il rivelatore è stato utilizzato in modalità di conteggio con un valore di guadagno di 10 per il rilevamento di fotoni, con una frequenza di scansione di 100 Hz e con una dimensione dei pixel di 17 nm. L'accumulo a quattro linee è stato utilizzato per l'acquisizione di TauSTED.
- Posizionare una goccia da 20 μL di supporto di montaggio (Tabella dei materiali) su un vetrino da microscopio. Prendere un foglietto di copertina dal punto 4.2.2 e posizionare delicatamente il campione sul vetrino rivolto verso il supporto di montaggio (Figura 2B).
Risultati
Per visualizzare la SC all'interno del tessuto germinale di C. elegans mediante SMLM, abbiamo impiegato 3D-SMLM raziometrico a 2 colori per localizzare HTP-3, un componente degli assi cromosomici, e il C-terminale del filamento trasversale SYP-5 etichettato endogenamente con un tag emoagglutinina (HA). La posizione di entrambe le proteine all'interno della SC di C. elegans è stata precedentemente determinata da altri studi16,30.
Per ridurre al minimo la diffusione della luce e le aberrazioni ottiche inerenti ai campioni biologici spessi, abbiamo ripreso la sezione z più in basso dei nuclei meiotici che contengono gli SC (Figura 3, linee gialle). Per ogni immagine acquisita, la posizione dello stadio piezoelettrico del piano di imaging è stata contrassegnata rispetto alla posizione dello stadio piezoelettrico quando l'obiettivo è stato focalizzato sulla vetrina. Ciò ha permesso il calcolo della distanza piezoelettrica dal coprifoglio. I campioni montati con successo sono fissati stabilmente vicino al vetrino di copertura e mantengono la forma gonade (cioè, il tessuto non viene schiacciato tra i due vetrini di copertura durante la fase di postfissazione). La qualità del montaggio del campione può essere facilmente valutata al microscopio stereoscopico poiché le gonadi ben fissate non mostrano alcun movimento in soluzione (punto 4.2.6). Tuttavia, a causa della stocasticità del processo di montaggio, il tessuto gonadico non sarà necessariamente disposto completamente piatto sul vetrino. Pertanto, il piano inferiore dei nuclei contenenti SC può essere trovato a distanze variabili rispetto al coprivetrino all'interno della stessa gonade.
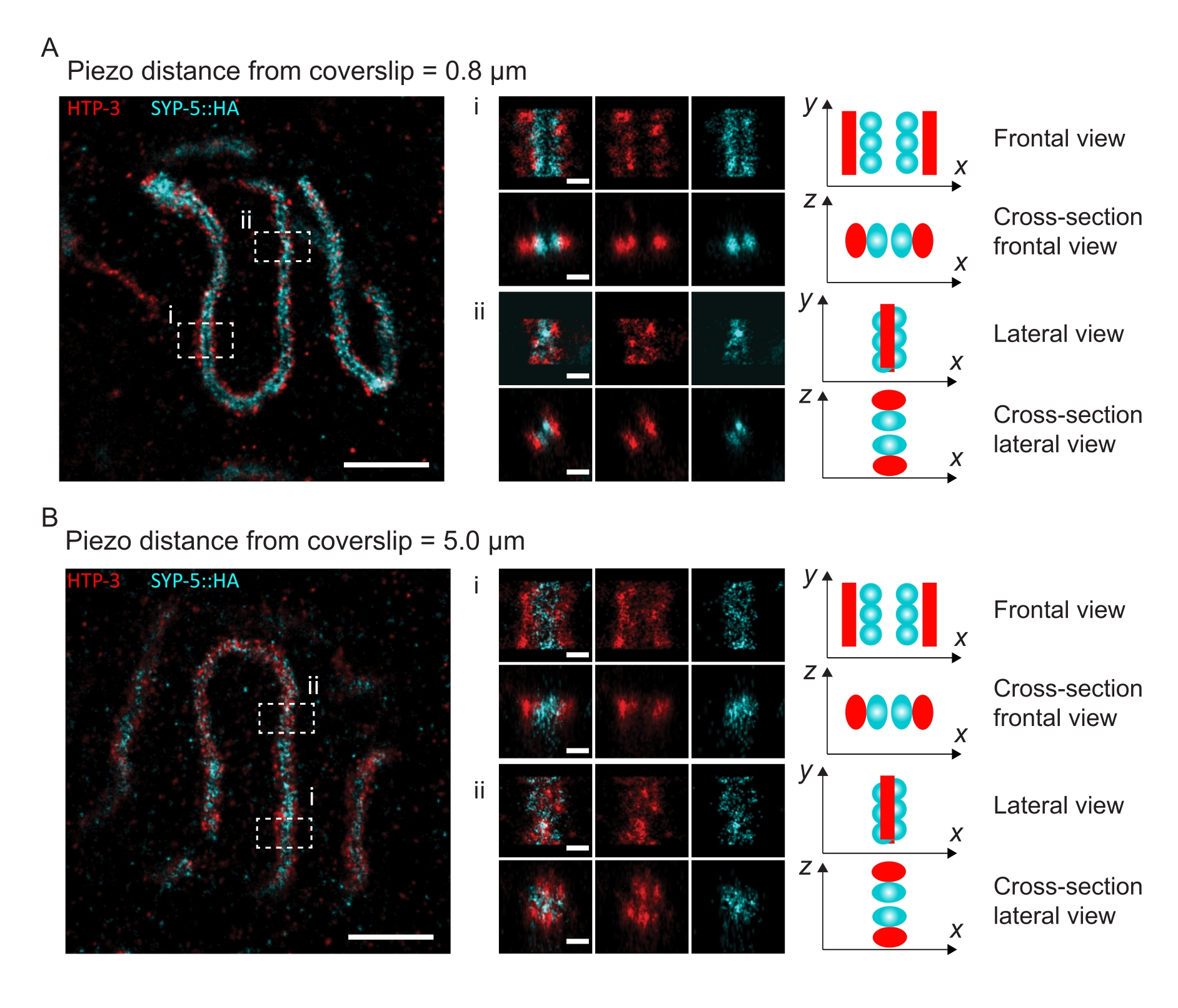
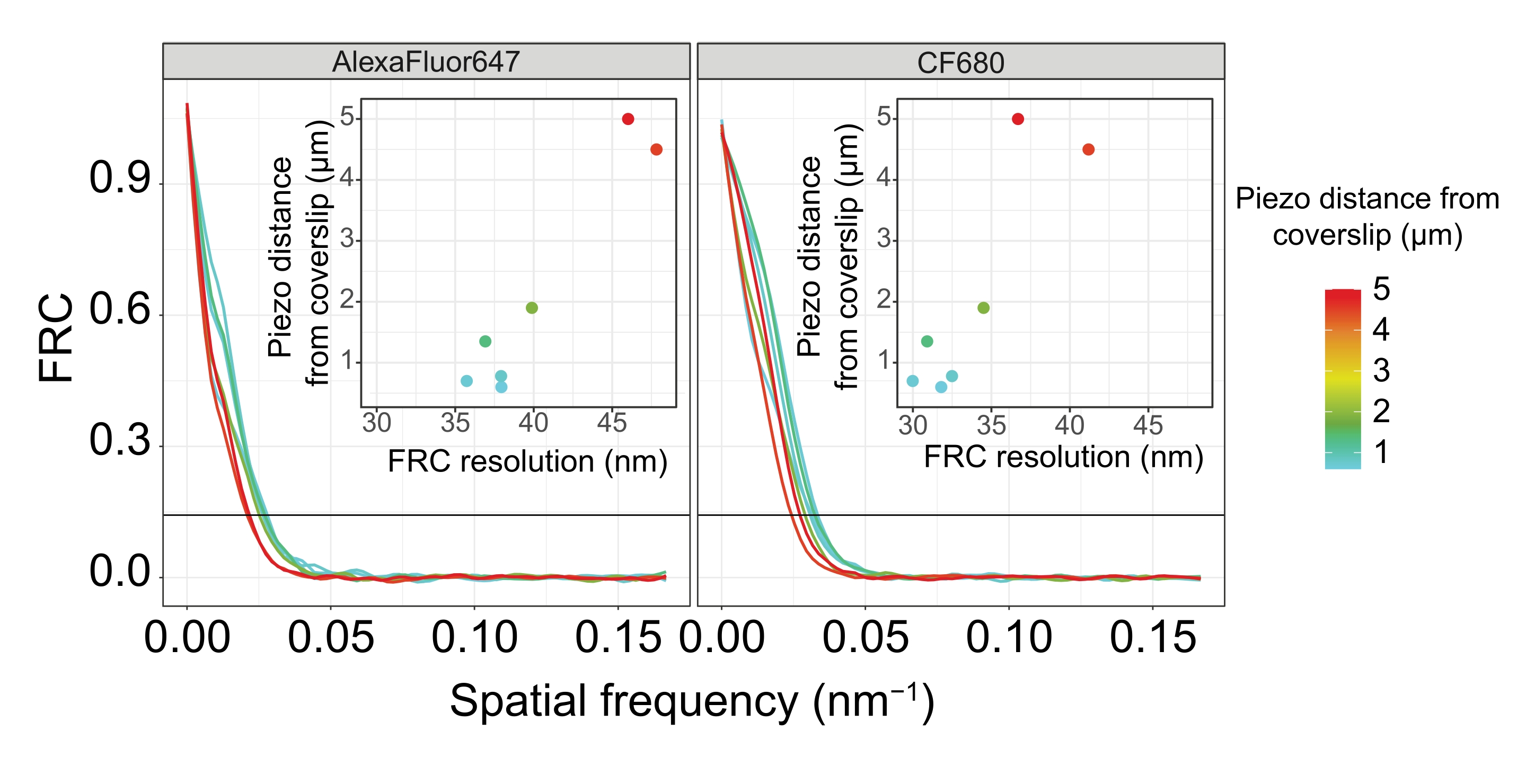
Per illustrare come la risoluzione cambia a seconda dell'attacco del tessuto al vetrino, abbiamo acquisito immagini a diverse distanze piezoelettriche dal vetrino. Per valutare la qualità di una singola immagine, sono state calcolate le curve di correlazione dell'anello di Fourier (FRC)50,51 e la risoluzione è stata determinata utilizzando il plugin FRCResolution all'interno del software SMAP48. Due nuclei rappresentativi estratti da due immagini 3D-SMLM separate prese a distanze diverse dal coprivetrino sono mostrati nella Figura 4. Nelle SC situate vicino al coprivetrino, gli assi cromosomici e il C-terminale di SYP-5::HA sono ben risolti in tutte e tre le dimensioni (Figura 4A, 0,8 μm dal coprifoglio). Per risolvere due strutture separate da una data distanza, la risoluzione FRC ottenuta deve generalmente essere inferiore alla metà di questa distanza nella risoluzione assiale.
Per separare lateralmente le stesse strutture, è necessario ottenere valori di risoluzione FRC ancora più piccoli. Infatti, nei campioni che si trovano in prossimità del coverslip, la risoluzione FRC è di 38 nm per il canale AlexaFluor 647 e 34 nm per il canale CF680, e quindi ben al di sotto della distanza prevista di 84 nm tra i termini C di SYP-516. Questa risoluzione risolve quindi prontamente l'organizzazione del CS non solo in vista frontale ma anche laterale (Figura 4B i,ii). Al contrario, la risoluzione si deteriora negli SC situati a una distanza di 5 μm dal vetrino di copertura a causa della diffusione della luce e delle aberrazioni sferiche (Figura 4B). Le risoluzioni FRC a questa distanza scendono a 47 nm (AlexaFluor 647) e 41 nm (CF680), che non possono risolvere completamente il C-termini di SYP-5. Poiché le aberrazioni ottiche compromettono la risoluzione laterale in modo più grave rispetto alla risoluzione assiale, le bande HTP-3 e SYP-5 non sono più chiaramente risolte nella sezione trasversale della vista laterale nei campioni situati a una distanza di 5 μm dal vetrino di copertura (Figura 4B ii). Il confronto della risoluzione FRC delle immagini acquisite a diverse distanze piezoelettriche dal coprivetrino ha rivelato che il tessuto ripreso non deve trovarsi più di 2 μm dal coprivetrino (Figura 5). Questo risultato evidenzia l'importanza della corretta esecuzione della fase di postfissazione, durante la quale il tessuto deve essere reticolato con successo al rivestimento in poli-L-lisina del vetrino.
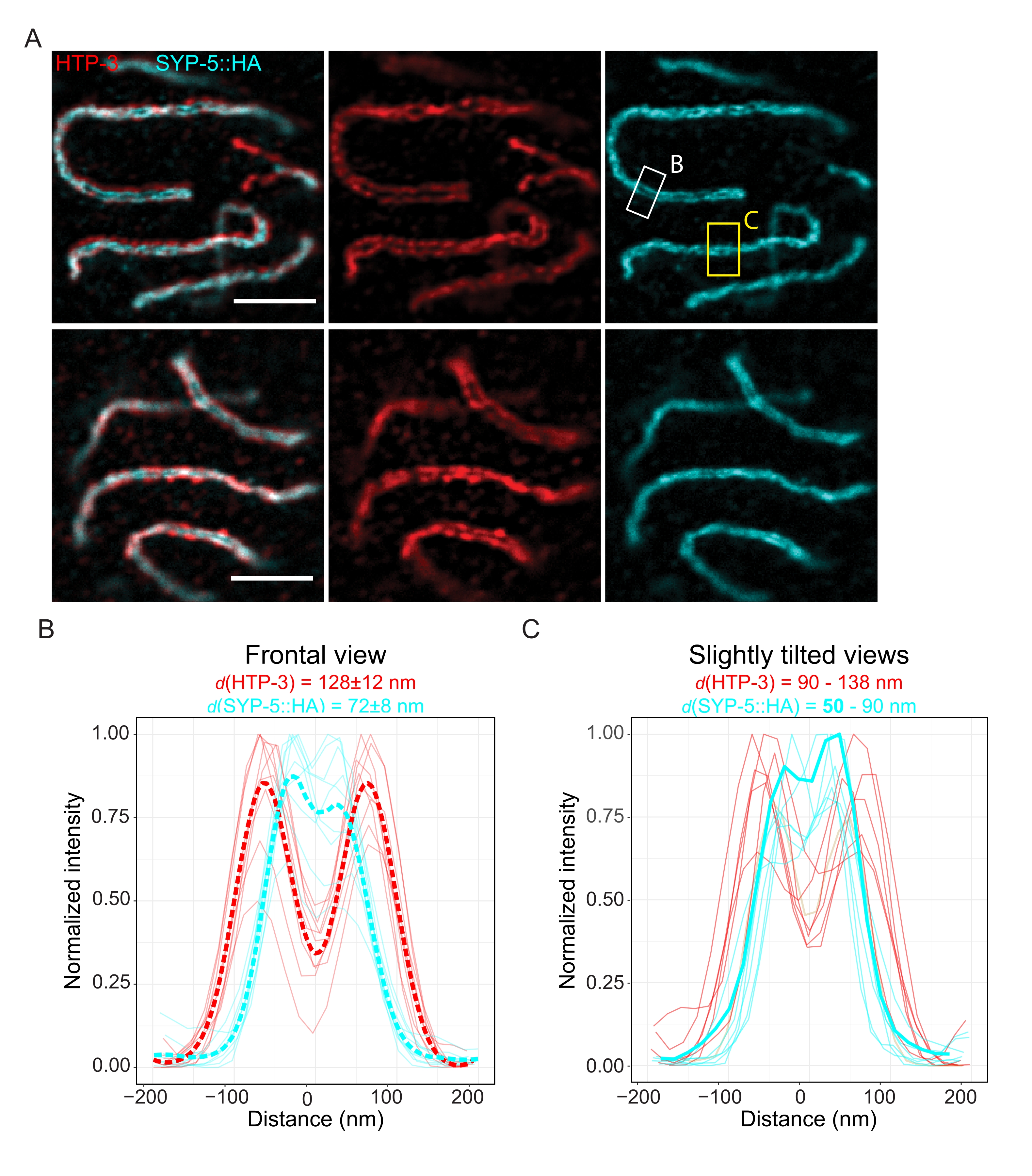
Per dimostrare la risoluzione ottenibile con un'altra tecnica di super-risoluzione, abbiamo anche ripreso SC in tessuto germinale intatto fisso con microscopia TauSTED. La Figura 6A mostra le immagini TauSTED con la risoluzione più alta e più bassa raggiunta nell'ambito di questo studio, come stimato dai profili lineari del SC in vista frontale (Figura 6B). In entrambi i nuclei, abbiamo potuto risolvere le due bande di localizzazione di HTP-3 negli assi cromosomici e i C-termini di SYP-5 nella regione centrale, dimostrando che la risoluzione ottenibile in TauSTED utilizzando questo protocollo ottimizzato è inferiore a 84 nm. In condizioni ottimali (Figura 6A, in alto), abbiamo potuto risolvere i C-termini in viste leggermente inclinate della SC che erano separate da soli 50 nm (Figura 6A, rettangolo giallo e 6C).

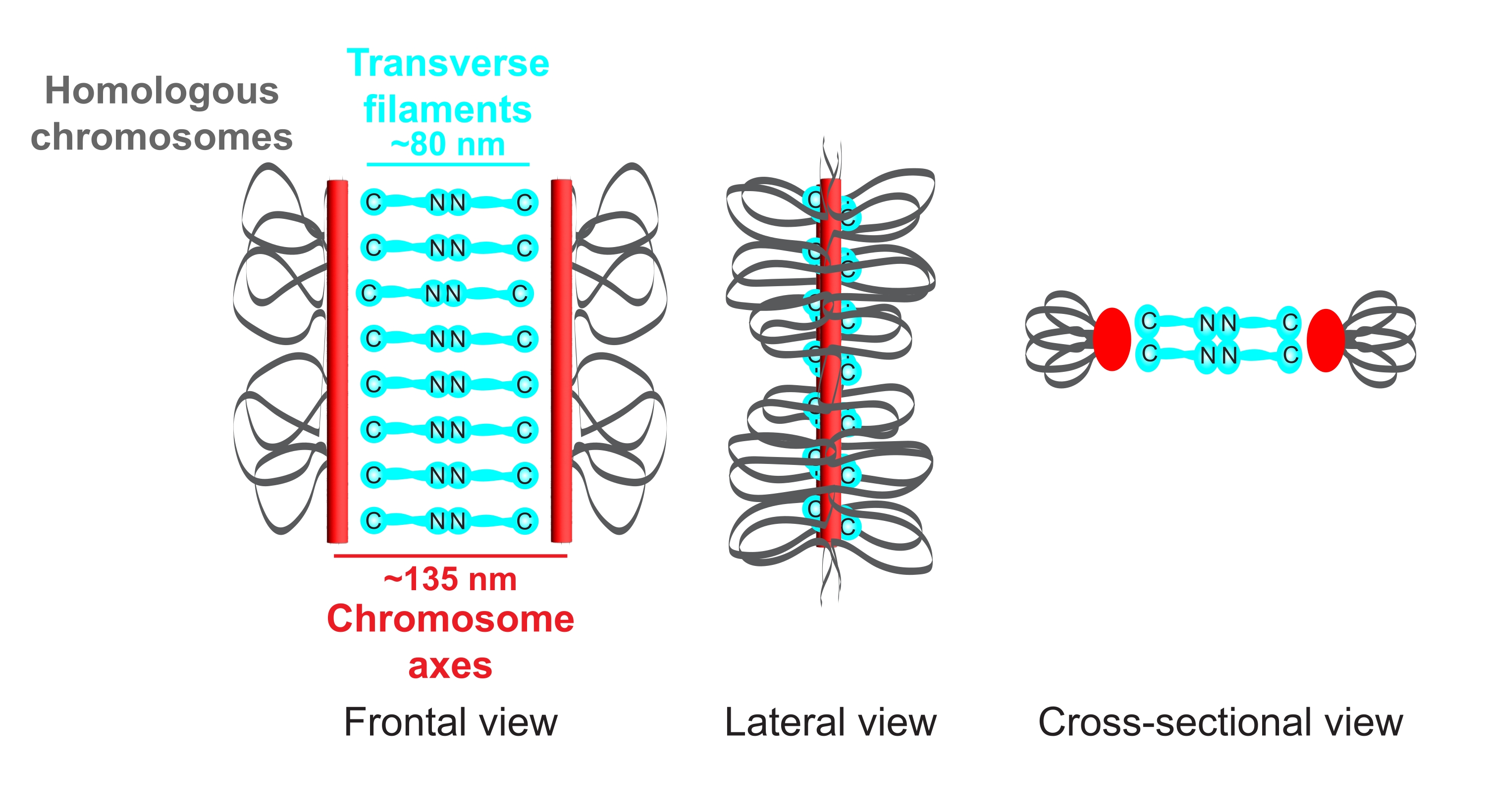
Figura 1: Schema dell'organizzazione del complesso sinaptonemale in Caenorhabditis elegans. Il fumetto mostra una struttura semplificata della SC in C. elegans che collega due cromosomi omologhi (grigio). La struttura è mostrata in viste frontali, laterali e trasversali. Gli assi cromosomici sono visualizzati come barre rosse mentre i filamenti trasversali sono mostrati in ciano. Le proteine del filamento trasversale (SYP-1, 5, 6 in C. elegans) sono orientate testa a testa (grafica ciano a sfera a bastone) nella regione centrale per colmare la distanza tra i due assi. Sono indicate le distanze previste tra gli assi e i termini C dei filamenti trasversali. Abbreviazione: SC = complesso sinaptonemale. Fare clic qui per visualizzare una versione ingrandita di questa figura.
{kind=link}

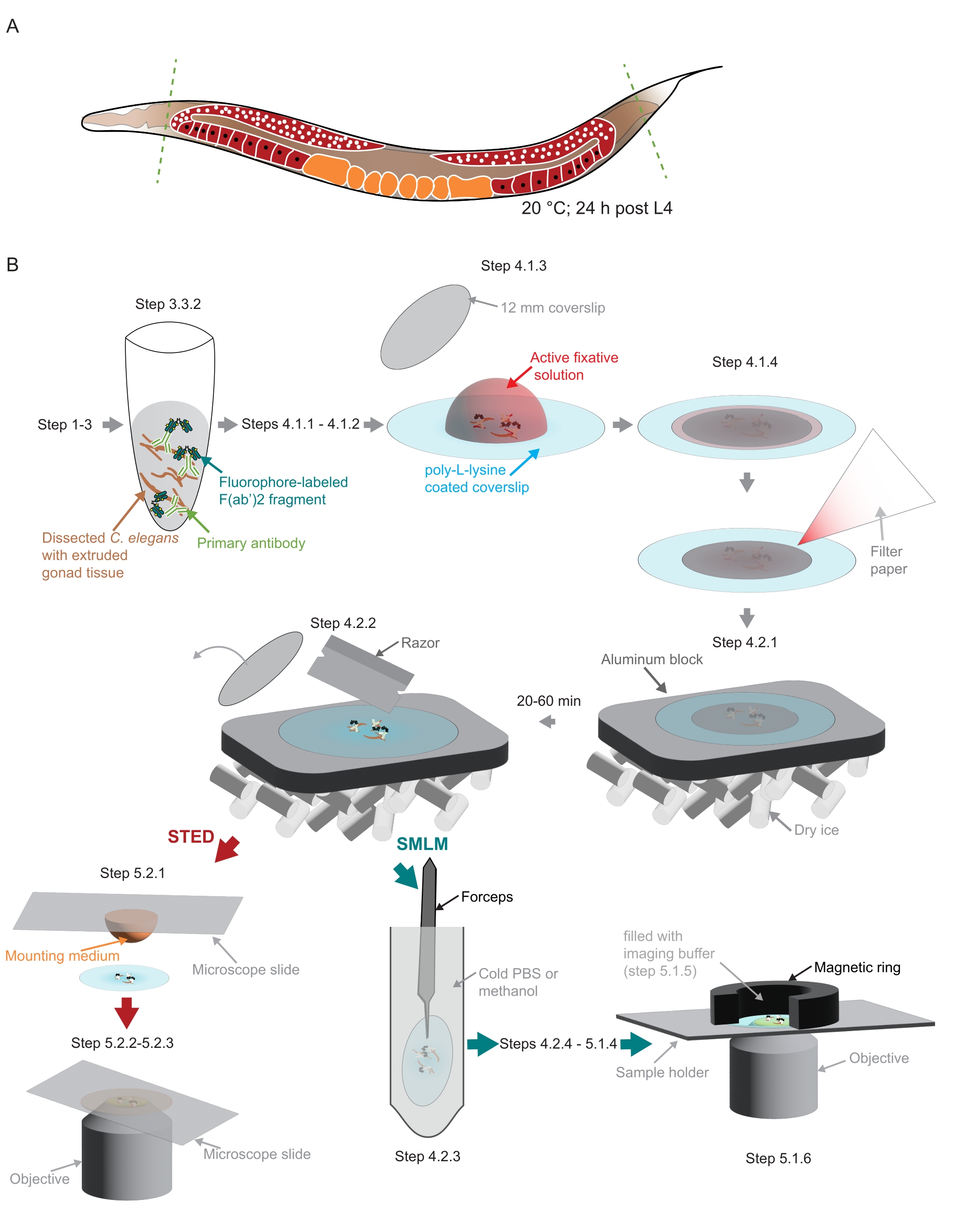
Figura 2: Illustrazione della preparazione del campione utilizzata nello studio . (A) I giovani adulti di C. elegans vengono sezionati alla testa o alla coda (linee tratteggiate verdi) e trattati come descritto nel protocollo. (B) I singoli passaggi del metodo sono indicati con grafici collegati a frecce grigie. Abbreviazioni: STED = esaurimento delle emissioni stimolate; SMLM = microscopia di localizzazione a singola molecola; PBS = soluzione salina tamponata con fosfato. Fare clic qui per visualizzare una versione ingrandita di questa figura.
{kind=link}

Figura 3: Posizione della sezione di tessuto che può essere osservata mediante microscopia di localizzazione di una singola molecola. MIP di un'immagine confocale del disco rotante di un'intera gonade di C. elegans. Il tessuto è stato colorato per HTP-3 e il C-terminale di SYP-5 (SYP-5::HA), e il segnale combinato è mostrato in grigio. Le singole immagini confocali sono state cucite utilizzando il plugin52 delle Fiji per creare un'immagine dell'intera gonade. L'inserto mostra una vista xy del piano z più in basso contenente gli SC. La localizzazione di questo piano è mostrata in viste ortogonali della sezione di tessuto indicate da un rettangolo nell'immagine MIP della gonade (linee gialle). Barre della scala = 10 μm. Abbreviazioni: MIP = proiezione di intensità massima; SCs = complessi sinaptonemali. Fare clic qui per visualizzare una versione ingrandita di questa figura.
{kind=link}

Figura 4: Microscopia di localizzazione a singola molecola di HTP-3 e C-termini di SYP-5. (A,B) A sinistra: immagini SMLM che mostrano nuclei di pachitene colorati per HTP-3 (rosso) e il C-terminale di SYP-5 (SYP-5::HA, ciano) (barra di scala = 1 μm). Centro: Immagini ingrandite delle regioni di interesse indicate in A e B con corrispondenti viste in sezione trasversale visualizzate sotto ogni immagine (i, ii; barra di scala = 100 nm). I tratti della SC all'interno delle immagini ingrandite vengono ruotati per orientare gli assi cromosomici parallelamente all'asse y. A destra: Rappresentazione grafica della localizzazione delle proteine di interesse all'interno del SC che rappresenta l'orientamento della SC nelle regioni ingrandite visualizzate al centro della figura. Abbreviazioni: SMLM = microscopia di localizzazione a singola molecola; SC = complesso sinaptonemale. I dati grezzi per ricostruire le immagini SMLM sono disponibili attraverso il database BioStudies60 (ID adesione: S-BIAD504). Fare clic qui per visualizzare una versione ingrandita di questa figura.
{kind=link}

Figura 5: La risoluzione di correlazione dell'anello di Fourier delle immagini al microscopio di localizzazione di singole molecole dipende dalla distanza del piano z ripreso dal piano del vetrino. Le linee colorate mostrano le curve FRC delle immagini acquisite a diverse distanze (come raffigurato dalla barra dei colori) dalla copertina. La soglia di 1/7 utilizzata per determinare la risoluzione FRC è indicata da una linea orizzontale nera. Gli inserti mostrano la dipendenza della risoluzione FRC dalla distanza piezoelettrica dal coprifoglio. Il plottaggio è stato eseguito da uno script R scritto su misura (versione 4.1.2, file supplementare 1) in cui le curve originali sono state smussate con funzioni del pacchetto "ggplot2". Abbreviazioni: FRC = correlazione dell'anello di Fourier; SMLM = microscopia di localizzazione a singola molecola; SC = complesso sinaptonemale. I dati per le curve FRC e i dati SMLM sono disponibili attraverso il database BioStudies60 (ID adesione: S-BIAD504). Fare clic qui per visualizzare una versione ingrandita di questa figura.
{kind=link}

Figura 6: La microscopia a deplezione ad emissione stimolata potenziata da informazioni basate sulla vita di fluorescenza (TauSTED) risolve due bande di localizzazione sia per HTP-3 che per il C-terminale di SYP-5. (A) Due immagini rappresentative di TauSTED mostrano nuclei di pachitene colorati per HTP-3 (rosso) e il C-terminale di SYP-5 (SYP-5::HA, ciano) con definizione strutturale superiore (superiore) e inferiore (inferiore) (barra di scala = 1 μm). I rettangoli segnano le regioni con i termini C risolti di SYP-5 in frontale (bianco) e una vista leggermente inclinata (giallo) del SC. (B,C) Distribuzione dell'HTP-3 (rosso) e del C-terminale del segnale SYP-5 (ciano) risolto da TauSTED. I profili delle linee delle regioni di interesse che contengono la SC nelle viste frontali (B) o leggermente inclinate (C) sono mostrati come linee complete con intensità normalizzata al valore massimo. I profili di linea sono stati generati utilizzando Fiji ImageJ. Le linee tratteggiate in B mostrano i dati medi per ciascuna proteina. La linea ciano spessa in C corrisponde al profilo della linea con la distanza risolta più breve tra i termini C di SYP-5. Per determinare le distanze tra gli anticorpi che colpiscono proteine specifiche, i profili di linea (n = 9 (B), n = 7 (C)) sono stati dotati di doppi gaussiani utilizzando uno script R scritto su misura (versione 4.1.2, file supplementare 1). La distanza media ± la deviazione standard (B) e l'intervallo con il valore minimo evidenziato in grassetto (C) sono indicati rispettivamente nella parte superiore di ciascun grafico. Abbreviazioni: STED = microscopia a deplezione ad emissione stimolata; SC = complesso sinaptonemale. Le immagini visualizzate e i punti dati dei profili delle linee tracciate sono disponibili attraverso il database BioStudies60 (ID adesione: S-BIAD504). Fare clic qui per visualizzare una versione ingrandita di questa figura.
{kind=link}
Tabella 1: Composizione dei buffer e delle soluzioni utilizzate in questo protocollo. Clicca qui per scaricare questa tabella.
Video supplementare 1: Acquisizione microscopica di localizzazione di singole molecole. Video che mostra fluorofori che lampeggiano a una velocità appropriata (vengono mostrati 50 fotogrammi, barra di scala = 5 μm, 20 ms / fotogramma). Clicca qui per scaricare questo video.
File supplementare 1: script di analisi dei dati. Clicca qui per scaricare questo file.
Discussione
L'organizzazione a scala della SC, che è essenziale per la corretta ricombinazione e segregazione dei cromosomi omologhi, è stata osservata per la prima volta quasi 70 anni fa nella microscopia elettronica 3,4. Mentre l'organizzazione complessiva della SC è facilmente risolvibile al microscopio elettronico, la localizzazione dei singoli componenti all'interno di questo complesso richiede un approccio più mirato. Con la sua larghezza di soli ~ 100 nm, la sottostruttura della SC non può essere risolta dalla microscopia a fluorescenza convenzionale. Tuttavia, la microscopia a super-risoluzione è diventata un importante motore per nuove scoperte sulla struttura e la funzione del complesso sinaptonemale 16,19,24,25,26,27,28,29,30. Per facilitare questa ricerca, abbiamo dimostrato una procedura di montaggio che consente di studiare l'architettura della SC all'interno del tessuto gonadico di C. elegans con microscopia SMLM e STED.
Un passo fondamentale per ottimizzare la risoluzione nell'imaging SMLM è la reticolazione diretta del tessuto germinale a un coprivetrino rivestito di poli-L-lisina (fase 4). L'attacco covalente del tessuto al vetrino di copertura è essenziale per ridurre i movimenti all'interno del campione che si tradurrebbero in grandi derive e renderebbero impossibile l'imaging per lunghi periodi di tempo per SMLM. Inoltre, anche un attacco non ottimale che lascia i nuclei contenenti SC a una certa distanza dal vetrino di copertura porta ad un calo significativo della risoluzione ottenibile derivante da aberrazioni sferiche (Figura 4). In alternativa all'attacco covalente qui utilizzato, il tessuto germinale colorato può anche essere immobilizzato tra due vetrini sigillati in una piccola goccia di tampone di imaging19,30. Tuttavia, questo metodo di immobilizzazione riduce notevolmente il volume del tampone di imaging nel campione da 1 mL utilizzato nel protocollo ottimizzato qui a pochi μL, il che comporterà un'acidificazione del tampone di imaging e ridurrà gravemente il tempo per il quale il campione può essere ripreso 38,53,54.
I lunghi tempi di acquisizione sia per la microscopia SMLM che per la microscopia STED limitano l'uso di questi metodi all'imaging di campioni fissati chimicamente. Qui, la fissazione della paraformaldeide assicura che la struttura del SC sia preservata durante la preparazione del campione e l'imaging. Tuttavia, nonostante le precauzioni prese qui per visualizzare il SC all'interno di tessuto intatto, la struttura risultante della SC dopo la fissazione non è necessariamente identica alla struttura nel suo stato nativo all'interno di un organismo vivente. Inoltre, poiché una singola immagine del SC fisso rappresenta una singola "istantanea" della struttura biologica, questo approccio rimane cieco alla dinamica della struttura nativa in vivo.
Tuttavia, le informazioni sulla dinamica e la variabilità delle strutture macromolecolari possono essere ottenute anche acquisendo non una singola ma molte "istantanee". Mentre questo approccio può risolvere i cambiamenti nella struttura del SC durante il pachitene19, ci sono diversi fattori che limitano il numero di immagini che possono essere acquisite da un singolo campione preparato utilizzando questo protocollo. In primo luogo, le elevate potenze laser utilizzate durante l'acquisizione delle immagini portano allo sbiancamento permanente dei fluorofori e precludono l'imaging di regioni adiacenti di interesse o piani z multipli, riducendo così significativamente il numero di immagini che possono essere acquisite da un singolo campione. In secondo luogo, la densità del campione/tessuto sul vetrino preparato con questo metodo è bassa, il che limita significativamente il numero di immagini che possono essere acquisite da un singolo vetrino. La bassa densità del campione vieta anche l'uso di pipeline di acquisizione automatica delle immagini che hanno contribuito a far luce su altre questioni biologiche 34,55,56,57,58,59. Tuttavia, la densità del campione può essere leggermente aumentata da un utente esperto.
Il protocollo qui presentato è ottimizzato per ottenere un'elevata densità di etichettatura necessaria per ottenere una risoluzione ottimale in SMLM35. Mentre i protocolli precedenti attaccano covalentemente il tessuto al vetrino prima dell'immunocolorazione16, questo nuovo protocollo collega il tessuto al vetrino solo dopo che i campioni sono stati colorati in soluzione. Questa modifica consente agli anticorpi utilizzati per l'immunomarcatura di accedere liberamente al tessuto da tutti i lati, mentre l'attaccamento covalente del tessuto al coprislip può impedire agli anticorpi di raggiungere i nuclei più vicini al vetrino, riducendo così il grado di etichettatura. Insieme, le modifiche qui descritte migliorano la risoluzione da 40-50 nm (risoluzione FRC)16 a 30-40 nm (questo protocollo).
È importante sottolineare che, mentre un'alta densità di etichettatura e un'alta concentrazione di anticorpi sono essenziali per SMLM, abbiamo scoperto che migliori immagini di microscopia STED sono ottenute utilizzando concentrazioni di anticorpi più basse (fase 3). Con una risoluzione di decine di nanometri, la dimensione delle molecole utilizzate per etichettare la proteina di interesse diventa sempre più importante. Abbiamo quindi impiegato frammenti F(ab')2 che sono la metà delle dimensioni degli anticorpi a lunghezza intera. Il miglioramento del contrasto locale dovuto ad una sorgente di segnale più piccola, e quindi la risoluzione ottenuta da questa modifica rispetto all'utilizzo di anticorpi secondari full-length, ha permesso la risoluzione dei due C-termini di SYP-5 all'interno della regione centrale da parte di TauSTED, che non sono risolti da STED convenzionali utilizzando anticorpi full-length (16 e dati non mostrati). Prevediamo che questo protocollo ottimizzato per l'imaging di SC in linee germinali intatte di C. eleganfaciliterà lo studio della relazione struttura-funzione della SC durante la meiosi.
Divulgazioni
Gli autori non dichiarano conflitti di interesse.
Riconoscimenti
Vorremmo ringraziare Jonas Ries e il laboratorio Ries per aver condiviso i buffer di imaging per l'imaging SMLM. Ringraziamo anche Yumi Kim per il ceppo C. elegans utilizzato in questo protocollo e Abby F. Dernburg per l'anticorpo pollo-anti-HTP-3. Ringraziamo Marko Lampe e Stefan Terjung dell'Advanced Light Microscopy Facility di EMBL Heidelberg per il loro supporto nell'utilizzo del microscopio confocale Olympus iXplore SPIN SR. Questo lavoro è stato sostenuto dal Laboratorio europeo di biologia molecolare e dalla Deutsche Forschungsgemeinschaft (DFG, German Research Foundation - 452616889, SK). Riconosciamo l'accesso e i servizi forniti dal Centro di Imaging presso il Laboratorio Europeo di Biologia Molecolare (EMBL IC), generosamente sostenuto dalla Fondazione Boehringer Ingelheim.
Materiali
| Name | Company | Catalog Number | Comments |
| 100x/1.5 oil objective | Olympus | UPLAPO100XOHR | UPLAPO100XOHR |
| 2-mercaptoethylamine (MEA) | Sigma-Aldrich | 30070-10G | Dissolved in MilliQ water to 5 M solution, pH 8.7 adjusted with HCl. Aliquoted to a single-use volume, frozen, and kept at -80 °C. |
| Additional 640 nm booster laser | Toptica | IBEAM-SMART-640-S-HP | |
| AlexaFluor 594, NHS ester | ThermoFischer Scientific | A37572 | Dissolved in DMSO to 1 mM solution, aliquoted to single use volume, frozen and kept at -80 °C |
| AlexaFluor 647, NHS ester | ThermoFischer Scientific | A37573 | Dissolved in DMSO to 1 mM solution, aliquoted to single use volume, frozen and kept at -80 °C |
| anti-HA | Thermo Fisher Scientific | 2-2.2.14 | Mouse monoclonal, 1:250 (SMLM), 1:1,000 (STED Microscopy) |
| anti-HTP-3 | a gift from Abby F. Dernburg | MacQueen et al., 2005 | Chicken polyclonal, 1:250 (SMLM), 1:1,000 (STED Microscopy) |
| Caenorhabditis elegans strain YKM349 | a gift from Yumi Kim | Hurlock et al., 2020 | syp-5(kim9[syp-5::HA]) I; meIs8[pie-1p::GFP::cosa-1, unc-119(+)] II |
| CF6680, NHS ester | Biotium | 92139 | Dissolved in DMSO to 1mM solution, aliquoted to single use volume, frozen and kept at -80 °C |
| Circular cover glass 12 mm No. 1 | Menzel-Gläser; VWR | 631-0713 | |
| Circular cover glass 24 mm No. 1.5 | Carl Roth | PK26.1 | |
| Cylindrical lenses | Thorlabs | LJ1516RM-A, LK1002RM-A | |
| Egg Buffer (10x) | Edgar 1995 | 250 mM HEPES, 1.18 M NaCl, 480 mM KCl, 20 mM EDTA, 5 mM EGTA, pH 7.4 | |
| Ethanol (absolute for analysis) | Merck | 64-17-5 | |
| F(ab’)2 fragment anti-chicken IgY | Jackson Immunoresearch | AB_2340347 | Donkey polyclonal, 1:100 (SMLM), 1:1,000 (STED Microscopy) |
| F(ab’)2 fragment anti-mouse IgG | Jackson Immunoresearch | AB_2340761 | Donkey polyclonal, 1:100 (SMLM), 1:1,000 (STED Microscopy) |
| Fisherbrand Microscope slides T/F Ground 0.8-1.0 mm thick | Fisher scientific | 7107 | |
| Gauge Worm Pick 30 diameter 0.254 mm - Iridium 10% | Kisker | 789265 | |
| Glucose oxidase/Catalase enzyme mix (GlOX/Cat ) | a gift from Jonas Ries | Hoess, Mund, Reitberger, & Ries, 2018 | 20x, 1916 U/mL glucose oxidase (Sigma G7141), 42350 U/mL catalase (Sigma C3155), 50 mM Tris-HCl pH 8.0, 51% glycerol, MilliQ water. Stored at -20 °C. |
| Imaging buffer base | a gift from Jonas Ries | Hoess, Mund, Reitberger, & Ries, 2018 | 50 mM Tris-HCl, pH 8.0, 10 mM NaCl, 10% D-Glucose. Aliquoted to a single-use volume (950 μL), frozen, and kept at -80 °C. |
| Invitrogen ProLong Glass Antifade Mountant | ThermoFischer Scientific | P36982 | |
| Leica Stellaris 8 STED FALCON | Leica | N/A | The microscope is equiped with the latest generation white light laser, a 775nm pulsed STED laser, the FALCON Fluorescence Lifetime IMaging module, HC PL APO CS2 100x/1.40 oil objective, and Leica HyD X detector. The system is capable of FLIM module enhanced Tau-STED which measures the specific fluorescence lifetime of a dye and is therefore capable of removing background signal based on differences in fluorescence lifetimes of the dyes, and dye conditions in the sample. Additionally, the resolution is increased by accounting for the variation of fluorescence lifetimes in different areas of the depletion donut. |
| Longwave channel emission filter | AHF Analysentechnik | F47-702 | 700/100 nm bandpass |
| Methanol (absolute for analysis) | Merck | 67-56-1 | |
| NaHCO3 | Sigma-Aldrich/Merck | S5761-500G | 100 mM NaHCO3, pH 8.3 |
| Near-infrared fiber-coupled laser | Toptica | IBEAM-SMART-PT-CD | Custom Design, 808 nm - 75mW |
| Objective lens piezo mount (PIFOC ) | Physik Instrumente | P-726.1.CD | 100 µm travel range |
| Orca Fusion BT sCMOS camera | Hamamatsu | C15440-20UP | |
| PCR tubes | Greiner Bio-One | 673283 | 0.2 mL |
| Phosphate Saline Buffer (PBS 10x) | N/A | 137 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, 1.8 mM KH2PO4, pH 7.4 | |
| Pierce 16% formaldehyde (w/v), methanol free | ThermoFischer Scientific | 28906 | 16% formaldehyde is transferred from the original glass ampule into 1.5 mL tube and kept at room temperature. |
| PlasmaPrep2 plasma cleaner | GaLa Instrumente GmbH | N/A | |
| Poly-L-lysine hydrobromide | Sigma-Aldrich/Merck | P2636-25MG | 0.1% w/v solution was prepared in Milli-Q water, and stored in aliquots at -20 °C. |
| Primary dichroic (illumination reflecting) | AHF Analysentechnik | F73-866S | quad bandpass @ 405, 488, 561, 640 |
| Quadnotch filter | AHF Analysentechnik | F40-072 | 405/488/561/640 nm |
| Quadrant photodiode (QPD) | Laser Components | SD197-23-21-041, LC301DQD-PV | |
| Razor blades | Apollo Herkenrath Solinger | N/A | |
| Refractive beam shaper | AdlOptica | PiShaper 6_6_VIS | |
| Roche Blocking Reagent | Roche | 11096176001 | 10x solution was prepared according to recommendation. Frozen aliquots were stored at -20 °C. |
| Scalpel blade (Feather brand #11, No. 3) | Heinz Herenz Medizinalbedarf GmbH | 1110911 | |
| Scalpel removal box | Fisher scientific | 10002-50 | |
| Secondary dichroic (emission reflecting) | AHF Analysentechnik | F38-785S | 750 nm longpass |
| Shortpass filter | Semrock | BSP01-785R-25 | 750 nm |
| Shortwave channel emission filter | AHF Analysentechnik | F37-677 | 676/37 nm bandpass |
| Single molecule localization microscope | EMBL Imaging Centre | Diekmann et al., 2020 with modifications | The microscope provides widefield epi-illumination via a single-mode fiber-coupled laser engine, additional booster laser, and refractive beam shaper to provide a uniform illumination field (Stehr et al, 2019). Widefield images are captured on a sCMOS camera and appropriate relay optics for a system magnification of 61x and a pixel size of 106 nm. For ratiometric imaging of spectrally overlapping far-red dyes, an image splitter produces two spectrally distinct images on the camera (splitting dichroic: 665 nm long pass, shortwave channel emission filter: 676/37 nm bandpass, longwave emission filter: 700/100 nm bandpass. An additional 405/488/561/640 nm quadnotch filter and 750 nm shortpass filter are common to the two paths and provide additional laser blocking). A compound cylindrical lens provides the astigmatism required for 3D imaging. To maintain a fixed focus across acquisitions exceeding 2 hours in time (comprising 200 000 - 250 000 images), focus locking is achieved by total internal reflection of a near-infrared fiber-coupled laser from the coverslip and subsequent height sensitive detection on a quadrant photodiode (QPD). The QPD signal provided closed-loop control of the objective lens piezo mount. For access to this microscope, refer to https://www.embl.org/about/info/imaging-centre or contact ic-contact@embl.de |
| Single-mode fiber-coupled multi-laser engine | Toptica | iCHROME MLE-LFA-HP | Provides widefield epi-illumination of 100 mW at 405, 488, 561, 640 nm |
| Splitting dichroic | AHF Analysentechnik | F48-665SG | 665 nm long pass |
| Square cover glass 22 x 22 mm No.1 | Menzel-Gläser; VWR | 630-2882 | |
| STAR 635P, NHS ester | Abberior | ST635P-0002-1MG | Dissolved in DMSO to 1 mM solution, aliquoted to single use volume, frozen and kept at -80 °C |
| Stereo microscope Stemi 305 Stand K LAB | Zeiss | N/A | |
| Tetramisole hydrochloride | Sigma-Aldrich/Merck | T1512-2G | 1% (w/v) solution was prepared in Milli-Q water. Frozen aliquots were stored at -20 °C. Thawed aliquot was kept at 4 °C and used for several months. |
| TetraSpeck Microspheres | ThermoFischer Scientific | T7279 | 0.1 µm, fluorescent blue/green/orange/dark red |
| Tris Saline Buffer (TBS 10x) | N/A | 200 mM Tris-HCl, 1.5 M NaCl, pH 7.5 | |
| TWEEN 20 | Sigma-Aldrich/Merck | P9416-50ML | Kept at room temperature in original packaging. |
| WormStuff worm pick | Kisker | 789277 | |
| XY microscope stage | Smaract | N/A | Custom Design |
| Zeba Micro Spin Desalting Column | ThermoFischer Scientific | 89877 | 7K MWCO, 75 µL |
Riferimenti
- Zickler, D., Kleckner, N. Meiotic chromosomes: integrating structure and function. Annual Review of Genetics. 33, 603(1999).
- Ur, S. N., Corbett, K. D. Architecture and dynamics of meiotic chromosomes. Annual Review of Genetics. 55, 497-526 (2021).
- Fawcett, D. W. The fine structure ot chromosomes in the meiotic prophase of vertebrate spermatocytes. The Journal of Biophysical and Biochemical Cytology. 2 (4), 403-406 (1956).
- Moses, M. J. Chromosomal structures in crayfish spermatocytes. The Journal of Biophysical and Biochemical Cytology. 2 (2), 215-218 (1956).
- Hillers, K. J., Jantsch, V., Martinez-Perez, E., Yanowitz, J. L. Meiosis. WormBook. , 433-434 (2017).
- Pasierbek, P., et al. A Caenorhabditis elegans cohesion protein with functions in meiotic chromosome pairing and disjunction. Genes & Development. 15 (11), 1349-1360 (2001).
- Severson, A. F., Ling, L., Van Zuylen, V., Meyer, B. J. The axial element protein HTP-3 promotes cohesin loading and meiotic axis assembly in C. elegans to implement the meiotic program of chromosome segregation. Genes & Development. 23 (15), 1763-1778 (2009).
- Zetka, M. C., Kawasaki, I., Strome, S., Müller, F. Synapsis and chiasma formation in Caenorhabditis elegans require HIM-3, a meiotic chromosome core component that functions in chromosome segregation. Genes & Development. 13 (17), 2258-2270 (1999).
- Martinez-Perez, E. HTP-1-dependent constraints coordinate homolog pairing and synapsis and promote chiasma formation during C. elegans meiosis. Genes & Development. 19 (22), 2727-2743 (2005).
- Couteau, F., Zetka, M. HTP-1 coordinates synaptonemal complex assembly with homolog alignment during meiosis in C. elegans. Genes & Development. 19 (22), 2744-2756 (2005).
- Goodyer, W., et al. HTP-3 Links DSB Formation with Homolog Pairing and Crossing Over during C. elegans Meiosis. Developmental Cell. 14 (2), 263-274 (2008).
- Colaiácovo, M. P., et al. Synaptonemal complex assembly in C. elegans is dispensable for loading strand-exchange proteins but critical for proper completion of recombination. Developmental Cell. 5 (3), 463-474 (2003).
- MacQueen, A. J., Colaiácovo, M. P., McDonald, K., Villeneuve, A. M. Synapsis-dependent and -independent mechanisms stabilize homolog pairing during meiotic prophase in C. elegans. Genes & Development. 16 (18), 2428-2442 (2002).
- Smolikov, S., et al. Synapsis-defective mutants reveal a correlation between chromosome conformation and the mode of double-strand break repair during Caenorhabditis elegans meiosis. Genetics. 176 (4), 2027-2033 (2007).
- Smolikov, S., Schild-Prüfert, K., Colaiácovo, M. P. A yeast two-hybrid screen for SYP-3 interactors identifies SYP-4, a component required for synaptonemal complex assembly and chiasma formation in Caenorhabditis elegans meiosis. PLoS Genetics. 5 (10), 1000669(2009).
- Hurlock, M. E., et al. Identification of novel synaptonemal complex components in C. Elegants. The Journal of Cell Biology. 219 (5), (2020).
- Zhang, Z., et al. Multivalent weak interactions between assembly units drive synaptonemal complex formation. The Journal of Cell Biology. 219 (5), (2020).
- Schild-Prüfert, K., et al. Organization of the synaptonemal complex during meiosis in Caenorhabditis elegans. Genetics. 189 (2), 411-421 (2011).
- Köhler, S., Wojcik, M., Xu, K., Dernburg, A. F. The interaction of crossover formation and the dynamic architecture of the synaptonemal complex during meiosis. bioRxiv. , (2020).
- Chen, F., Tillberg, P. W., Boyden, E. S. Expansion microscopy. Science. 347 (6621), 543-548 (2015).
- Klar, T. A., Jakobs, S., Dyba, M., Egner, A., Hell, S. W. Fluorescence microscopy with diffraction resolution barrier broken by stimulated emission. Proceedings of the National Academy of Sciences. 97 (15), 8206-8210 (2000).
- Rust, M. J., Bates, M., Zhuang, X. Sub-diffraction-limit imaging by stochastic optical reconstruction microscopy (STORM). Nature Methods. 3 (10), 793-796 (2006).
- Betzig, E., et al. Imaging intracellular fluorescent proteins at nanometer resolution. Science. 313 (5793), 1642-1645 (2006).
- Schücker, K., Holm, T., Franke, C., Sauer, M., Benavente, R. Elucidation of synaptonemal complex organization by super-resolution imaging with isotropic resolution. Proceedings of the National Academy of Sciences. 112 (7), 2029-2033 (2015).
- Cahoon, C. K., et al. Superresolution expansion microscopy reveals the three-dimensional organization of the Drosophila synaptonemal complex. Proceedings of the National Academy of Sciences. 114 (33), 6857-6866 (2017).
- Zwettler, F. U., et al. Tracking down the molecular architecture of the synaptonemal complex by expansion microscopy. Nature Communications. 11 (1), 1-11 (2020).
- Yoon, S., Choi, E. H., Kim, J. W., Kim, K. P. Structured illumination microscopy imaging reveals localization of replication protein A between chromosome lateral elements during mammalian meiosis. Experimental & Molecular Medicine. 50 (8), 1-12 (2018).
- Prakash, K., et al. Superresolution imaging reveals structurally distinct periodic patterns of chromatin along pachytene chromosomes. Proceedings of the National Academy of Sciences. 112 (47), 14635-14640 (2015).
- Xu, H., et al. Molecular organization of mammalian meiotic chromosome axis revealed by expansion STORM microscopy. Proceedings of the National Academy of Sciences. 116 (37), 18423-18428 (2019).
- Köhler, S., Wojcik, M., Xu, K., Dernburg, A. F. Superresolution microscopy reveals the three-dimensional organization of meiotic chromosome axes in intact Caenorhabditis elegans tissue. Proceedings of the National Academy of Sciences. 114 (24), 4734-4743 (2017).
- Hell, S. W. Far-field optical nanoscopy. Science. 316 (5828), 1153-1158 (2007).
- Hein, B., Willig, K. I., Hell, S. W. Stimulated emission depletion (STED) nanoscopy of a fluorescent protein-labeled organelle inside a living cell. Proceedings of the National Academy of Sciences. 105 (38), 14271-14276 (2008).
- Jahr, W., Velicky, P., Danzl, J. G. Strategies to maximize performance in STimulated Emission Depletion (STED) nanoscopy of biological specimens. Methods. 174, 27-41 (2019).
- Thevathasan, J. V., et al. Nuclear pores as versatile reference standards for quantitative superresolution microscopy. Nature Methods. 16 (10), 1045-1053 (2019).
- Xu, K., Shim, S. -H., Zhuang, X. Super-resolution imaging through stochastic switching and localization of single molecules: an overview. Far-Field Optical Nanoscopy. , 27-64 (2013).
- Wang, Y., et al. Localization events-based sample drift correction for localization microscopy with redundant cross-correlation algorithm. Optics Express. 22 (13), 15982(2014).
- Winterflood, C. M., Platonova, E., Albrecht, D., Ewers, H. Dual-color 3D superresolution microscopy by combined spectral-demixing and biplane imaging. Biophysical Journal. 109 (1), 3-6 (2015).
- Diekmann, R., et al. Optimizing imaging speed and excitation intensity for single molecule localization microscopy. Nature Methods. 17 (9), 909(2020).
- Huang, B., Wang, W., Bates, M., Zhuang, X. Three-dimensional super-resolution imaging by stochastic optical reconstruction microscopy. Science. 319 (5864), 810-813 (2008).
- Phillips, C. M., McDonald, K. L., Dernburg, A. F. Cytological analysis of meiosis in Caenorhabditis elegans. Methods in Molecular Biology. 558, 171-195 (2009).
- Edgar, L. G. Blastomere culture and analysis. Methods in Cell Biology. 48, 303-321 (1995).
- MacQueen, A. J., et al. Chromosome sites play dual roles to establish homologous synapsis during meiosis in C. elegans. Cell. 123 (6), 1037-1050 (2005).
- Stehr, F., Stein, J., Schueder, F., Schwille, P., Jungmann, R. Flat-top TIRF illumination boosts DNA-PAINT imaging and quantification. Nature Communications. 10 (1), 1-8 (2019).
- Hoess, P., Mund, M., Reitberger, M., Ries, J. Dual-color and 3D super-resolution microscopy of multi-protein assemblies. Methods in Molecular Biology. 1764, 237-251 (2018).
- Edelstein, A., Amodaj, N., Hoover, K., Vale, R., Stuurman, N. Computer Control of microscopes using µManager. Current Protocols in Molecular Biology. 92 (1), 14-20 (2010).
- Edelstein, A. D., et al. Advanced methods of microscope control using µManager software. Journal of Biological Methods. 1 (2), 10(2014).
- Deschamps, J., Ries, J. EMU: reconfigurable graphical user interfaces for Micro-Manager. BMC Bioinformatics. 21 (1), 1-13 (2020).
- Ries, J. SMAP: a modular super-resolution microscopy analysis platform for SMLM data. Nature Methods. 17 (9), 870-872 (2020).
- Li, Y., et al. Global fitting for high-accuracy multi-channel single-molecule localization. Nature Communications. 13 (1), 1-11 (2022).
- Nieuwenhuizen, R. P. J., et al. Measuring image resolution in optical nanoscopy. Nature Methods. 10 (6), 557-562 (2013).
- Banterle, N., Bui, K. H., Lemke, E. A., Beck, M. Fourier ring correlation as a resolution criterion for super-resolution microscopy. Journal of Structural Biology. 183 (3), 363-367 (2013).
- Preibisch, S., Saalfeld, S., Tomancak, P. Globally optimal stitching of tiled 3D microscopic image acquisitions. Bioinformatics. 25 (11), 1463-1465 (2009).
- Shi, X., Lim, J., Ha, T. Acidification of the oxygen scavenging system in single-molecule fluorescence studies: in situ sensing with a ratiometric dual-emission probe. Analytical Chemistry. 82 (14), 6132-6138 (2010).
- Olivier, N., Keller, D., Rajan, V. S., Gönczy, P., Manley, S. Simple buffers for 3D STORM microscopy. Biomedical Optics Express. 4 (6), 885-899 (2013).
- Mund, M., et al. Superresolution microscopy reveals partial preassembly and subsequent bending of the clathrin coat during endocytosis. bioRxiv. , (2022).
- Mund, M., et al. Systematic nanoscale analysis of endocytosis links efficient vesicle formation to patterned actin nucleation. Cell. 174 (4), 884-896 (2018).
- Sabinina, V. J., et al. Three-dimensional superresolution fluorescence microscopy maps the variable molecular architecture of the nuclear pore complex. Molecular Biology of the Cell. 32 (17), 1523-1533 (2021).
- Cieslinski, K., et al. Nanoscale structural organization and stoichiometry of the budding yeast kinetochore. bioRxiv. , (2021).
- Sieben, C., Banterle, N., Douglass, K. M., Gönczy, P., Manley, S. Multicolor single-particle reconstruction of protein complexes. Nature Methods. 15 (10), 777-780 (2018).
- Sarkans, U., et al. The BioStudies database—one stop shop for all data supporting a life sciences study. Nucleic Acids Research. 46, (2018).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneThis article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati