
Method Article
Cultivo primario de baja densidad fácil y reproducible utilizando existencias congeladas de neuronas embrionarias del hipocampo
En este artículo
Resumen
Un stock congelado de neuronas listo para usar es una herramienta poderosa para evaluar las funciones sinápticas. Aquí, introducimos un cultivo primario fácil de baja densidad a partir de material congelado utilizando una placa de 96 pocillos.
Resumen
El cultivo neuronal es un sistema valioso para evaluar las funciones sinápticas y los exámenes de detección de fármacos. En particular, un cultivo de baja densidad de neuronas primarias del hipocampo permite el estudio de neuronas individuales o componentes subcelulares. Hemos demostrado la localización de proteínas subcelulares dentro de una neurona mediante inmunocitoquímica, polaridad neuronal, morfología sináptica y su cambio en el desarrollo utilizando un cultivo primario del hipocampo de baja densidad. Recientemente, las reservas congeladas de neuronas listas para usar se han vuelto comercialmente disponibles. Estas reservas congeladas de neuronas reducen el tiempo necesario para preparar experimentos con animales y también contribuyen a la reducción del número de animales utilizados. Aquí, introducimos un método de cultivo primario de baja densidad reproducible utilizando una placa de 96 pocillos. Utilizamos un stock congelado comercialmente disponible de neuronas del hipocampo embrionario de rata. Las neuronas se pueden cultivar de manera estable a largo plazo sin cambios en los medios al reducir el crecimiento de células gliales en puntos de tiempo particulares. Este ensayo de alto rendimiento que utiliza cultivo de baja densidad permite evaluaciones reproducibles basadas en imágenes de la plasticidad sináptica.
Introducción
El desarrollo de un sistema experimental in vitro que pueda evaluar las funciones sinápticas involucradas en el aprendizaje y la memoria es importante. El cultivo neuronal es un sistema valioso para evaluar las funciones sinápticas in vitro. La técnica de cultivo neuronal se utilizó por primera vez en la década de 1980, y en la década de 1990, se desarrolló un cultivo de baja densidad de neuronas primarias del hipocampo 1,2,3 para el estudio de neuronas individuales en términos de localización subcelular de componentes proteicos, tráfico de proteínas, polaridad neuronal, morfología de la columna vertebral, desarrollo de sinapsis y plasticidad 4,5,6,7,8 . Sin embargo, hay muchos pasos involucrados en esta técnica: apareamiento de animales, disección de embriones, preparación de vasos de cultivo y cultivo de células durante 3 semanas con cambios de medios una vez a la semana. Además, requiere técnicas avanzadas3.
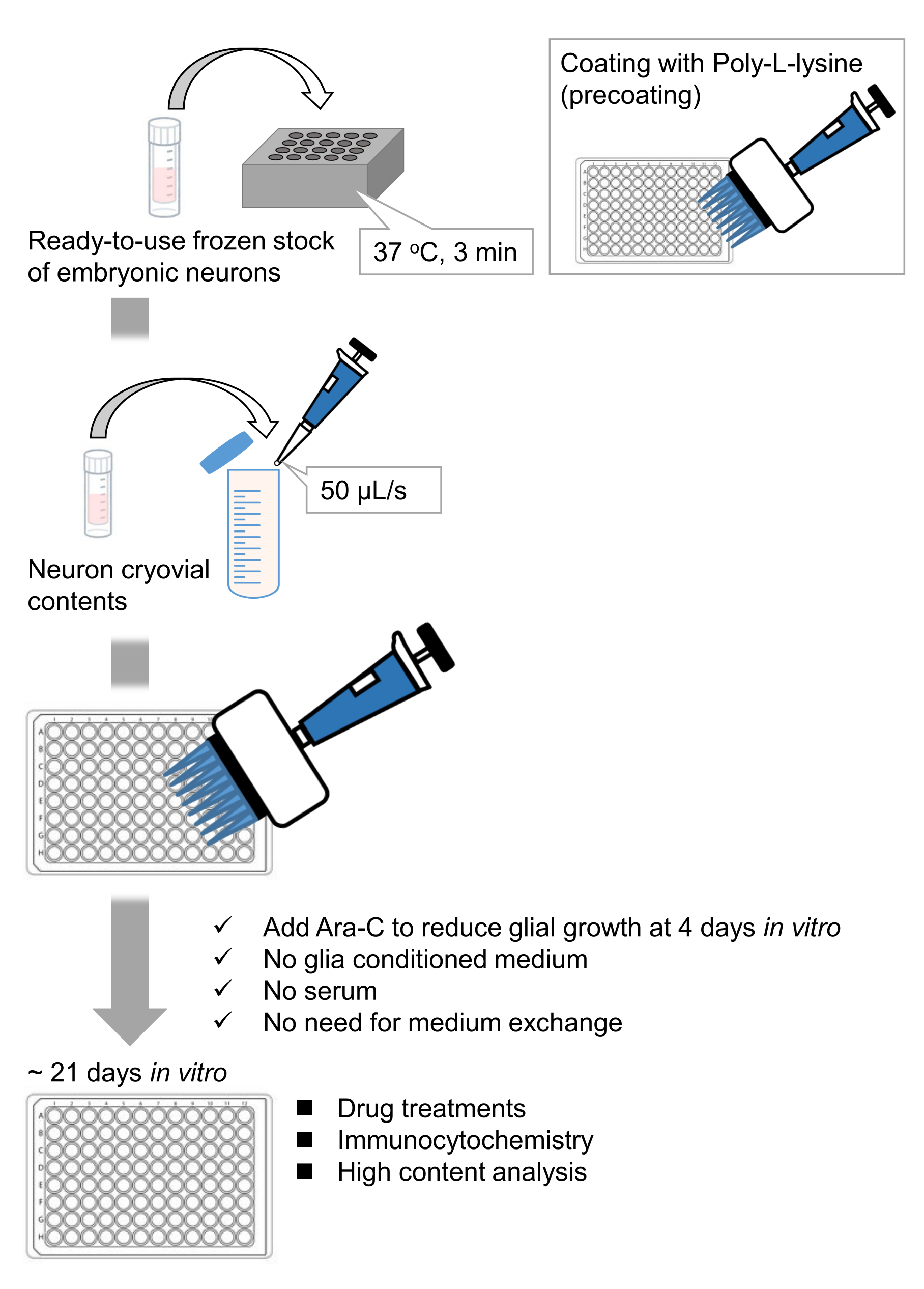
Hemos desarrollado reservas congeladas de neuronas disociadas del hipocampo a partir de embriones de rata 9,10. Las reservas congeladas de neuronas están listas para usar, y no se requieren técnicas avanzadas para cultivar las células11,12. En otras palabras, cultivar las neuronas a partir de existencias congeladas no depende de la técnica de un experimentador. Elimina la necesidad de experimentos con animales (por ejemplo, permiso para experimentos con animales, organización de animales preñados cronometrados y disección de embriones de rata), reduciendo así el número de animales utilizados. Recientemente, las reservas congeladas de neuronas de alta calidad y listas para usar se han vuelto comercialmente disponibles. Aquí, utilizamos existencias congeladas disponibles comercialmente del hipocampo de rata embrionario (E)18 13,14,15. El cultivo de las neuronas a partir de un material congelado no requiere medios acondicionados por la glía o cocultivo con células gliales. Se pueden utilizar medios de cultivo primarios ordinarios sin suero adicional para cultivar las células; por lo tanto, podemos adquirir datos reproducibles. Además, no hay necesidad de intercambio de medios durante 3 semanas después de la siembra celular ya que se reduce el crecimiento de las células gliales (Figura 1).
Las espinas dendríticas son el compartimiento postsináptico de la mayoría de las sinapsis excitatorias. Contienen proteínas receptoras, proteínas de andamio postsináptico y proteínas citoesqueléticas de actina. Nos centramos en una proteína de unión a actinadrebrina 5,6,7,16,17,18. La drebrina se acumula en la cabeza de la columna vertebral en las neuronas maduras19, y reportamos drebrina como marcador del estado sináptico 15,17,20,21,22,23. Al realizar un análisis de alto contenido utilizando drebrina como lectura, hemos reportado recientemente los efectos inhibitorios de los análogos de fenciclidina sobre los receptores de glutamato tipo ácido N-metil-D-aspártico (NMDARs)10 y los efectos dependientes de NMDAR de compuestos naturales y drogas crudas en estados sinápticos15.
Aquí, detallamos cómo cultivar reservas congeladas de neuronas a baja densidad. Además, mostramos una evaluación basada en imágenes de drebrina del estado sináptico utilizando placas de 96 pocillos.
Protocolo
1. Revestimiento de la placa
- Cubrir una microplaca de 96 pocillos con poli-L-lisina (1 mg/ml, diluido en tampón de borato 0,1 M [pH: 8,5]; 100 μL/pocillo) e incubar durante la noche a 37 °C.
NOTA: Solo cubra los pozos que necesitan ser utilizados. En los experimentos realizados aquí, se utilizan los 60 pozos intermedios. El tampón de borato se prepara mezclando 50 mM de ácido bórico y 12 mM de borato en agua esterilizada. - Lave el plato dos veces con agua esterilizada (250 μL/pocillo).
- Lavar el plato una vez con un medio de cultivo fresco sin suplementos (250 μL/pocillo).
- Seque el plato en un banco limpio durante 20 minutos.
- Envuelva la placa con papel de aluminio y manténgala a 4 °C hasta su uso (válido durante 1 mes).
2. Siembra celular
- Añadir 50 μL/pocillo del medio de cultivo a la placa recubierta y mantenerla en una incubadora deCO2 al 5% a 37 °C durante 30 min a 1 h. Llenar los pocillos periféricos con agua esterilizada (200 μL/pocillo).
NOTA: El medio de cultivo se prepara agregando 50x B-27, 400x Glutamax y 100 U/mL de penicilina/estreptomicina al medio neurobasal (consulte la Tabla de materiales para obtener más detalles). - Retire la neurona criovial del tanque de nitrógeno líquido. Las neuronas utilizadas aquí fueron neuronas criopreservadas DMSO11.
- Sumerja el criovial en un bloque de calor a 37 °C durante un máximo de 3 minutos y descongele parcialmente el contenido. No caliente el criovial durante demasiado tiempo. Transfiera el contenido a un tubo de 50 ml tan pronto como se descongele.
- Transfiera lentamente el contenido criovial de la neurona a un tubo estéril de 50 ml en sentido gota (50 μL/s) utilizando una pipeta de 1 ml con una punta de poro ancho.
- Enjuague el criovial vacío con 1 ml del medio de cultivo (temperatura ambiente; RT). Transfiera este 1 ml del medio de cultivo del criovial en forma de gota (50 μL/s) al tubo de 50 ml que contiene la suspensión celular.
- Agregue 9 ml del medio de cultivo (RT) al tubo de 50 ml en forma de gota (0,5 ml/s) y enrasar el volumen a 11 ml. No repita el pipeteo, pero mezcle la suspensión celular lentamente.
- Cuente el número de células (use un contador de células o un hemocitómetro).
- Transfiera toda la suspensión celular a un depósito y dispense la suspensión celular a la placa de 96 pocillos utilizando una pipeta multicanal con puntas de poro ancho (1,0 x 104 células/pocillo). Para reducir la evaporación del medio de cultivo, llene los pocillos periféricos con agua esterilizada (paso 2.1).
NOTA: Este estudio confirma que la evaporación del medio de cultivo es pequeña para un cultivo de 3 semanas sin intercambio de medios. La tasa de reducción del medio es del 3,6% (n = 120 pozos). Por lo tanto, el cambio de osmolalidad no será drástico durante el período de incubación de 3 semanas. - Incubar las neuronas durante 1-2 h en una incubadora deCO2 al 5% a 37 °C.
- Sustituir el medio de cultivo por 100 μL de medio de cultivo precalentado (37 °C) por pocillo y volver a colocarlo en una incubadora deCO2 al 5 % a 37 °C (no se requiere cambio de medio durante el cultivo).
3. Tratamiento Ara-C
- A los 4 días in vitro (DIV), agregue citosina β-D-arabino-furanósido (Ara-C) a una concentración final de 0,2 μM por pocillo para reducir el crecimiento de las células gliales.
4. Tratamientos farmacológicos
- A los 21 días in vitro, tratar las células con los fármacos de interés.
- Mantener la temperatura de la placa a 37 °C durante los tratamientos farmacológicos.
- Para un control positivo, tratar las células con 100 μM de glutamato (por pocillo para la concentración final) durante 10 minutos antes de la fijación.
5. Fijación
- Para la fijación, utilizar paraformaldehído al 4% en tampón fosfato 0,1 M (100 μL/pocillo).
- Después de ~20 min de fijación, lave los pocillos con solución salina tamponada con fosfato (PBS; 250 μL/pocillo) 2x durante 5 min cada uno.
6. Inmunocitoquímica
- Lave las células con PBS (250 μL/pocillo) 1x durante 5 min.
- Permeabilizar las células con Triton X-100 al 0,1% (100 μL/pocillo) en PBS durante 5 min.
- Lave las células con PBS (250 μL/pocillo) 3 veces durante 5 minutos cada una.
- Para el bloqueo, use albúmina sérica bovina al 3% en PBS (PBSA; 100 μL/pocillo) durante 1 h en RT.
- Incubar las células con anticuerpos anti-drebrina (1:1) y anti-microtúbulos asociados a la proteína 2 (MAP2) (1:000) (60 μL/pocillo) a 4 °C durante la noche.
- Lave las células con PBS (250 μL/pocillo) 4 veces durante 5 minutos cada una.
- Incubar las células con anticuerpos secundarios apropiados y 4′,6-diamidino-2-fenilindol, diclorhidrato (DAPI; 1:1000) en PBSA (60 μL/pocillo) durante 2 h a RT.
- Lave las células con PBS (250 μL/pocillo) 4 veces durante 5 minutos cada una.
- Almacene las células en PBS que contengan 0,1% de azida sódica (150 μL/pocillo).
7. Adquisición y análisis de imágenes
- Para adquirir las imágenes, utilice un microscopio apropiado.
- Para identificar los cuerpos celulares de las neuronas, utilice regiones MAP2 positivas y DAPI positivas.
- Para identificar las dendritas de las neuronas, use señales positivas para MAP2 sin cuerpos celulares.
- Para identificar grupos de drebrina, use señales positivas de drebrina a lo largo de dendritas positivas para MAP2.
Resultados
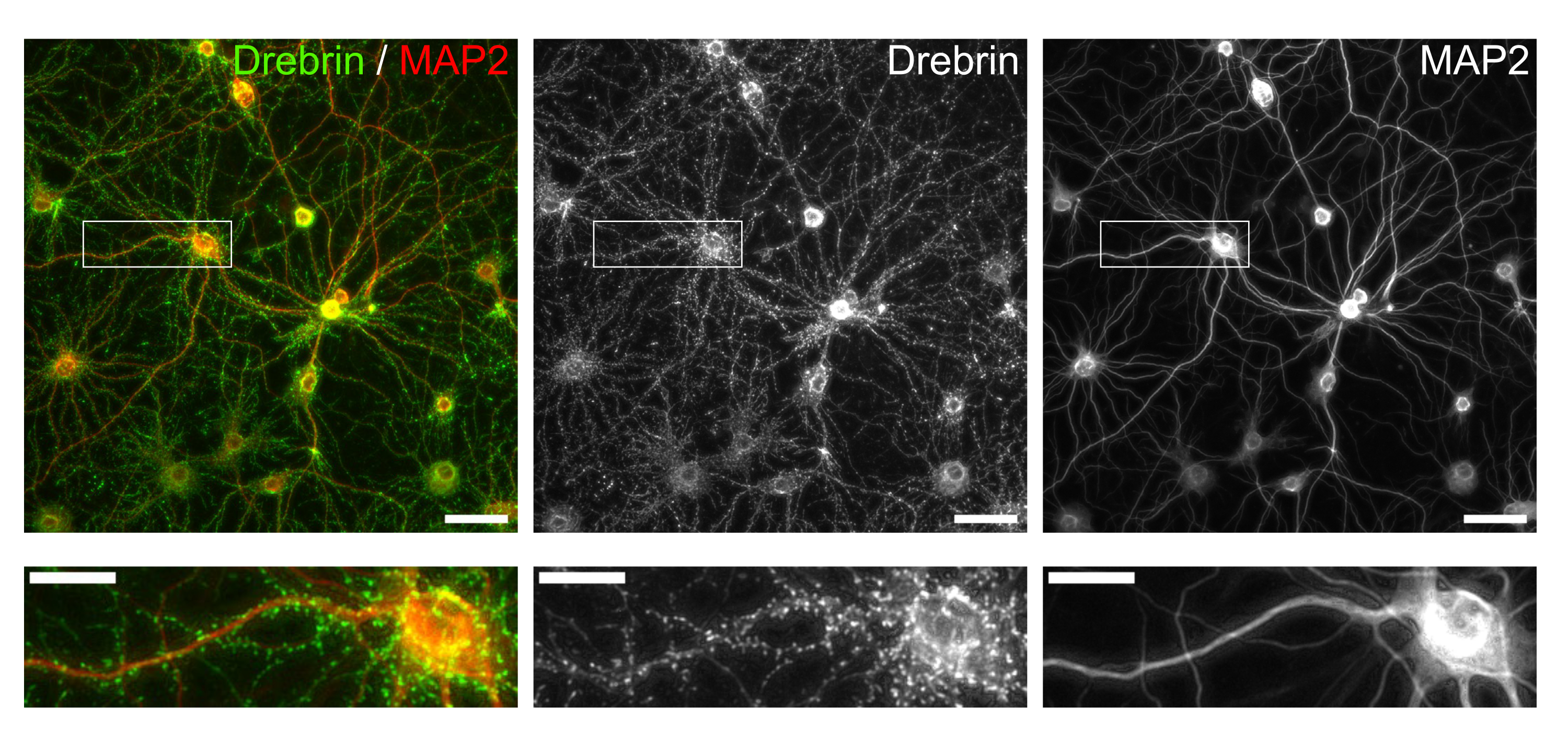
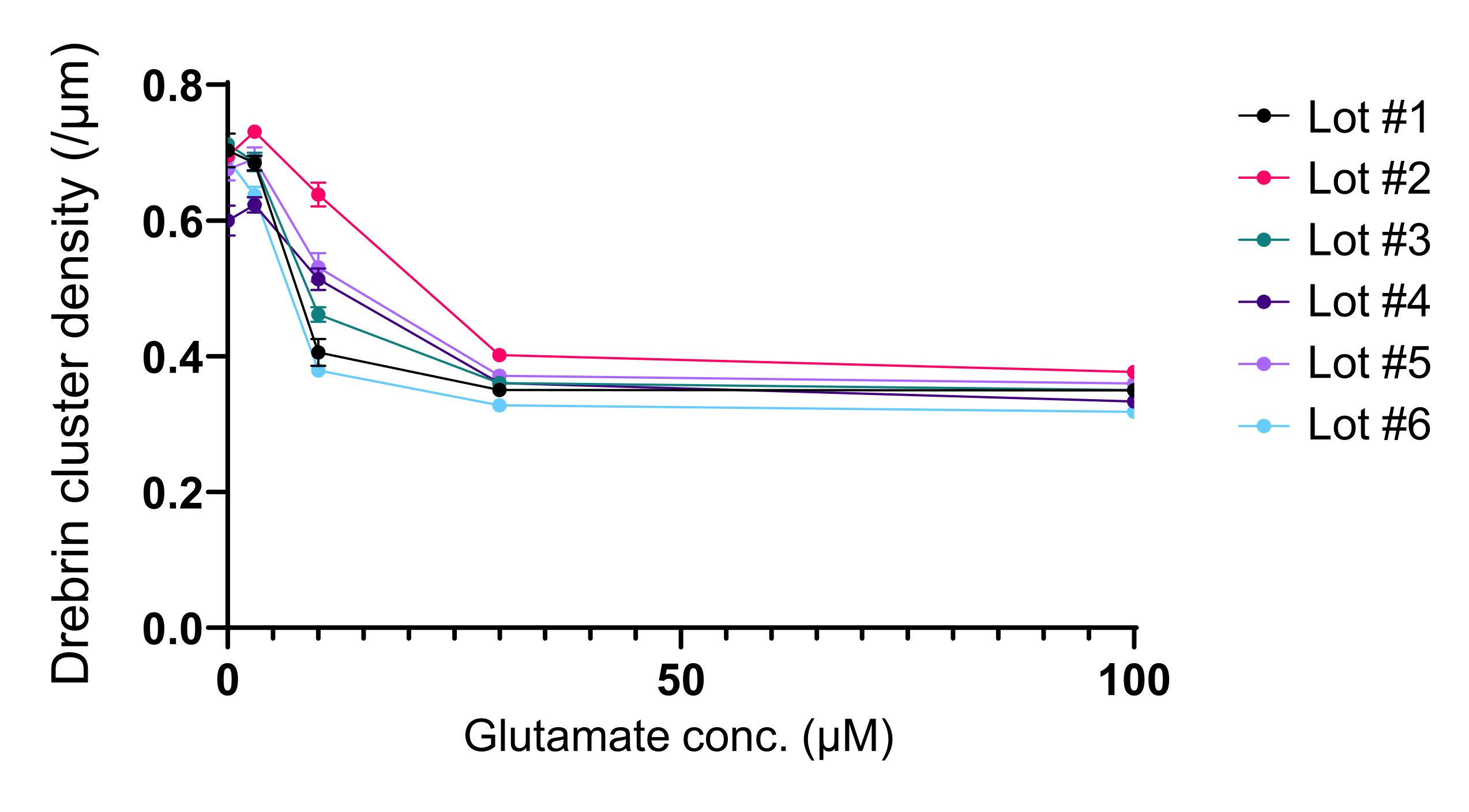
Siguiendo el protocolo, las neuronas fueron cultivadas en una placa de 96 pocillos durante 21 días, y luego tratadas con glutamato (Figura 1). Las neuronas se desarrollaron normalmente sin un intercambio del medio de cultivo durante 3 semanas (Figura 2). Tratamos las células con varias concentraciones de glutamato (1 μM, 3 μM, 10 μM, 30 μM y 100 μM diluidas en agua esterilizada) durante 10 min y las fijamos. Se realizó inmunocitoquímica y se adquirieron imágenes de fluorescencia de drebrina y MAP2 utilizando un microscopio de fluorescencia automatizado con una cámara sCMOS. Como se muestra en la Figura 3, las espinas dendríticas positivas para drebrina se observan claramente a lo largo de las dendritas positivas para MAP2. Se ha demostrado que la estimulación con glutamato provoca la afluencia de Ca2+ a través de NMDAR, lo que provoca el éxodo de drebrina de las espinas dendríticas, lo que resulta en una reducción de las densidades de racimo de drebrina 5,17. En consecuencia, observamos la reducción dependiente de la dosis de las densidades de racimos de drebrina frente a la estimulación del glutamato10 (Figura 4). Como se muestra en la Figura 5, este método es altamente reproducible si se utiliza drebrina como marcador para los estados sinápticos.

Figura 1: Esquema del método. Las neuronas se cultivaron en una placa de 96 pocillos durante 21 días y luego se trataron con glutamato. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 2: Imágenes de campo brillante de neuronas cultivadas usando una placa de 96 pocillos. Las imágenes de contraste de fase se obtuvieron de cada etapa de desarrollo (DIV 1, 7, 14, 21) utilizando un citómetro de imagen cuantitativa confocal. Barra de escala: 50 μm. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 3: Imágenes representativas de neuronas cultivadas inmunoteñidas. (Izquierda) Imágenes de fluorescencia combinadas de drebrina (verde) y MAP2 (rojo). Cada imagen de fluorescencia de drebrin y MAP2 se muestra en los paneles central y derecho, respectivamente. Los rectángulos blancos muestran el área ampliada debajo. Barras de escala; Paneles superiores: 50 μm, Paneles inferiores: 20 μm. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 4: Cambios dosis-respuesta dependientes del glutamato en la densidad normalizada del grupo de drebrina. (A) Imágenes representativas de fluorescencia inmunoteñidas con drebrina (verde) y MAP2 (rojo) del pocillo tratado con glutamato de 0 μM, 10 μM y 100 μM (de izquierda a derecha). Barra de escala: 50 μm. (B) La densidad del conglomerado de Drebrin se normalizó por el promedio de control (0 μM). 0 μM, N = 58 pocillos; 1 μM, N = 46; 3 μM, N = 54; 10 μM, N = 45; 30 μM, N = 54; 100 μM, N = 55, de 13 experimentos utilizando diferentes lotes. ** P < 0,01 versus control (0 μM) según la prueba de comparaciones múltiples de Dunnett después de ANOVA. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 5: Cambios dosis-respuesta dependientes del glutamato en la densidad del grupo de drebrina. Los datos brutos de seis experimentos utilizando diferentes lotes. N = 4 pocillos para cada concentración (0 μM, 3 μM, 10 μM, 30 μM y 100 μM). Los valores se expresan como media ± SEM. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

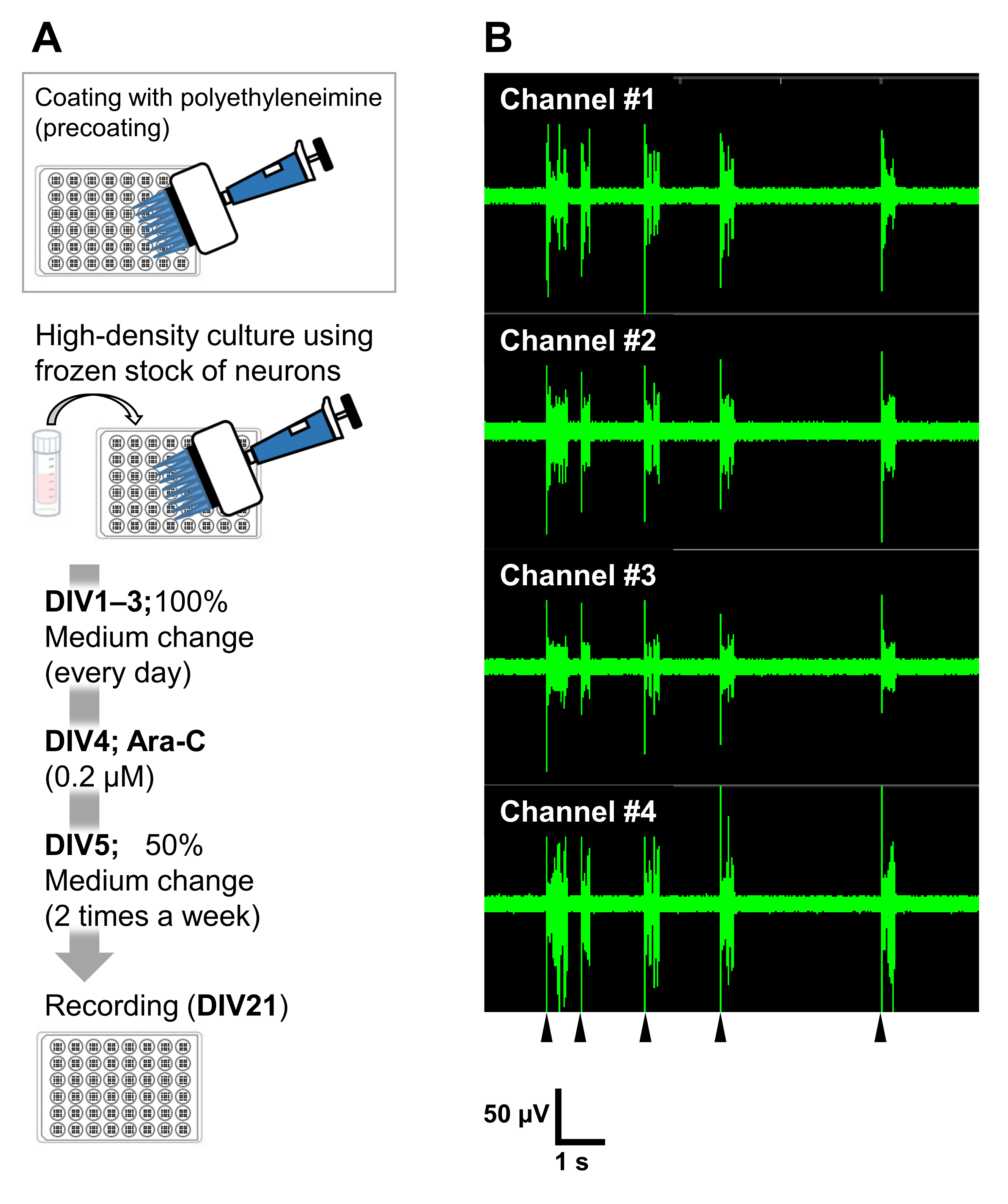
Figura 6: Aplicación de las reservas congeladas de neuronas a experimentos electrofisiológicos. (A) Un protocolo para experimentos electrofisiológicos utilizando placas de matriz de microelectrodos (MEA). Recubrimiento: Un día antes de enchapar las células, cada placa MEA de 48 pocillos se recubrió previamente con una solución de polietileniena (PEI: 0,1%) y se incubó durante 1 h a 37 °C. La placa MEA se lavó 3 veces con agua esterilizada y se secó durante 1 h. Luego, la placa MEA se mantuvo a 4 ° C durante la noche. Cultivo de alta densidad: 50.000 células/pocillo de las neuronas se colocaron en placas MEA de 48 pocillos. El paso de siembra celular se realizó como se describe en la sección 2 del protocolo descrito anteriormente. Se utilizó laminina (20 μg/ml) medio de cultivo añadido (añadir 2 v/v% B-27, 2,5 mM Glutamax y 100 μg/ml de penicilina/estreptomicina al medio neurobasal) para placar las neuronas. Posteriormente, las neuronas se cultivaron a 37 °C, 5% deCO2 en el medio de cultivo. Los medios se intercambiaron completamente en DIV 1 con el medio de cultivo hasta DIV 3. Ara-C se añadió en DIV 4 (0,2 μM finales). Desde DIV 5 en adelante y 2 veces por semana, el 50% de los medios se cambió con el medio de cultivo. La actividad de las neuronas en cada pocillo de la placa MEA se registró con un sistema MEA. (B) La actividad neuronal espontánea se adquirió a 37 °C bajo una atmósfera de CO2 al 5% utilizando un sistema MEA a una frecuencia de muestreo de12,5 kHz/canal en DIV 21. Se muestran grabaciones de 4 canales de los 16 canales dentro de un pozo. Para todas las grabaciones, se aplicó un filtro de paso de banda Butterworth (200-3.000 Hz). Las puntas de flecha muestran el tiempo de disparo sincronizado de ráfaga. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
Discusión
Un paso crítico en este método es descongelar la suspensión celular. La transferencia de la suspensión celular antes de que se caliente demasiado es muy importante. Sin embargo, para evitar el cambio rápido de osmolalidad, no transfiera la suspensión celular a un gran volumen del medio a la vez. La adición gota a gota del medio de cultivo también es crucial para evitar cambios repentinos en la presión osmótica.
Las neuronas se pueden cultivar en otros vasos de cultivo: placas de 24 pocillos, cámaras de 8 pocillos, platos de 60 mm o sondas MED. En esos casos, sin embargo, la concentración final y el momento de la adición de Ara-C deben ajustarse. Además, la densidad de las neuronas necesita ser optimizada en diferentes tipos de experimentos. Por ejemplo, se requiere un cultivo de alta densidad para experimentos electrofisiológicos, y en ese caso, se necesita el intercambio de medios dos veces por semana (Figura 6). Por lo tanto, una cultura de baja densidad requiere menos pasos que una cultura de alta densidad.
El cultivo neuronal de baja densidad a menudo requiere técnicas avanzadas; Sin embargo, el uso de material congelado listo para usar resuelve este problema. El método descrito no depende de la habilidad de un experimentador. La calidad del material congelado es estable y se puede cultivar de forma estable siempre que se almacenen en nitrógeno líquido y eviten cambios de temperatura hasta por 4 años.
El cultivo de las células durante 3 semanas sin intercambio de medios plantea la cuestión de si hay cambios significativos en la osmolalidad o evaporación de los medios de cultivo. Sin embargo, hemos confirmado que la evaporación de los medios de cultivo es pequeña (tasa de reducción del 3,6%). La localización de las proteínas sinápticas y la morfología de las neuronas parecen normales después de 3 semanas. Por lo tanto, el cultivo de 3 semanas sin intercambio de medios no causa grandes cambios de osmolalidad que afecten las condiciones de las neuronas cultivadas. Mantener la placa en una incubadora después del tratamiento Ara-C también es un punto importante que minimiza la evaporación.
No hay limitación con respecto al uso de las neuronas de stock congeladas. Sin embargo, existen algunas limitaciones del método de cultivo de baja densidad. Confirmamos que el cultivo de baja densidad podría aplicarse para la observación morfológica de neuronas, la evaluación de la función sináptica y la transfección de GFP. Sin embargo, no hemos examinado las imágenes de células vivas. Además, como se mencionó anteriormente, se requiere un cultivo de alta densidad para realizar electrofisiología.
La maduración de la sinapsis suele tardar 3 semanas7, y no podemos confirmar que las neuronas cultivadas tengan sinapsis adecuadas hasta el final. Si la maduración de la sinapsis no es buena después de 3 semanas, tendríamos que volver a cultivar. Al conocer la calidad de las neuronas antes de comenzar los experimentos, podemos ahorrar estas 3 semanas. Por lo tanto, para realizar experimentos de manera eficiente, es mejor verificar la calidad de las neuronas con anticipación. Las existencias congeladas permiten verificar la calidad de las neuronas de antemano. Cada lote de existencias congeladas se genera a partir de una camada de ratas, y podemos utilizar una de las existencias de cada lote para un control de calidad. La drebrina es un buen marcador para el control de calidad de las neuronas. Como se ha descrito, la drebrina se acumula en la cabeza de la columna vertebral en las neuronas maduras y reacciona a la estimulación sináptica. Por lo tanto, podemos verificar la calidad de las neuronas en las existencias congeladas utilizando drebrina como marcador.
Este método se puede aplicar para evaluar el efecto de los fármacos en el estado sináptico. El éxodo de drebrina de las espinas dendríticas ocurre durante las etapas iniciales de la plasticidad sináptica22. Por lo tanto, la detección de la reducción del grupo de drebrina provocada por el tratamiento farmacológico muestra que el fármaco estimula la sinapsis y causa plasticidad sináptica. Además, para identificar si la reducción es dependiente de NMDAR, es útil un experimento con ácido 2-amino-5-fosfonovalérico (APV, un antagonista de NMDAR). Usando drebrin como marcador, incluso una dependencia NMDAR está claramente determinada10,15. El método descrito es útil en exámenes de detección de fármacos, estudios farmacológicos de seguridad y evaluación de la función sináptica.
Divulgaciones
Tomoaki Shirao es el CEO de AlzMed, Inc. El estudio fue financiado por AlzMed, Inc. (500,000 JPY a NK para el proyecto titulado "Análisis de alto rendimiento de la función sináptica").
Agradecimientos
Agradecemos a Kazumi Kamiyama y Manami Kawada por su ayuda con los experimentos. Este trabajo fue apoyado por JSPS KAKENHI (Número de subvención 19K08010 a N.K.) y la Agencia Japonesa para la Investigación y el Desarrollo Médico (AMED) (Número de subvención JP19bk0104077 y JP22bm0804024 a T.S.).
Materiales
| Name | Company | Catalog Number | Comments |
| 96 well plate | Zeon Corporation | Gifted | |
| 96 well plate | greiner | 655986 | |
| Anti-drebrin antibody (M2F6) | MBL | D029-3 | Mouse monoclonal (dilution 1:1) |
| Anti-MAP2 antibody | Millipore | AB5622 | Rabbit (dilution 1:1000) |
| Anti-mouse Alexa Fluor 568 | Thermo Fisher Scientific | A11031 | Dilution 1: 500 |
| Anti-rabbit Alexa Fluor | Thermo Fisher Scientific | A11008 | Dilution 1: 500 |
| B-27 | Gibco | 17504-044 | 2 v/v% for MEA plates; 50x for normal plates |
| Borax | Sigma | B-9876 | Final concentration 12 mM |
| Boric acid | WAKO | 021-02195 | Final concentration 50 mM |
| Bovine serum albumin | Millipore | 12659-100G | Final concentration: 3% in PBS |
| Confocal quantitative image cytometer CellVoyager CQ1 | YOKOGAWA | Phase contrast images | |
| Cytosine β-D-arabino-furanoside (Ara-C) | Sigma | C-6645 | Diluted in dH2O (final concentration: 0.2 µM) |
| DAPI | FUJIFILM | 340-07971 | Dilution 1:1000 |
| GlutaMAX | Gibco | 35050-061 | 2.5 mM for MEA plates; 400x for normal plates |
| In Cell Analyzer 2200 | Cytiva | Fluorescence images | |
| Laminin | Sigma | 114956-81-9 | Final concentration: 20 µg/mL |
| Maestro | Axion Biosystems | MEA recordings | |
| MEA plate | Axion Biosystems | M768-tMEA-48W | |
| Neurobasal | Gibco | 21103-049 | |
| Paraformaldehyde | nacalai tesque | 26126-25 | Final concentration: 4% in PBS |
| Penicillin/Streptomycin | Gibco | 15140-122 | 100 U/mL for normal plates |
| Penicillin/Streptomycin | nacalai tesque | 26253-84 | 100 µg/mL for MEA plates |
| polyethyleimine | Sigma | 9002-98-6 | Final concentration: 0.1% |
| Poly-L-lysine | Sigma | P2636 | Diluted in the borate buffer (final concentration: 1 mg/mL) |
| SKY Neuron | AlzMed , Inc. | ARH001 | 1.0 x 106 cells/tube |
| Sodium azide | FUJIFILM | 195-11092 | 0.1% |
| SodiumL(+)-Glutamate monohydrate | WAKO | 194-02032 | Diluted in dH2O (final concentrations: 1 µM, 3 µM, 10 µM, 30 µM, 100 µM) |
Referencias
- Banker, G. A., Cowan, W. M. Rat hippocampal neurons in dispersed cell culture. Brain Research. 126 (3), 397-342 (1977).
- Kaech, S., Banker, G. Culturing hippocampal neurons. Nature Protocols. 1 (5), 2406-2415 (2006).
- Roppongi, R. T., Champagne-Jorgensen, K. P., Siddiqui, T. J. Low-density primary hippocampal neuron culture. Journal of Visualized Experiments. (122), e55000 (2017).
- Mizui, T., et al. Drebrin E is involved in the regulation of axonal growth through actin-myosin interactions. Journal of Neurochemistry. 109 (2), 611-622 (2009).
- Mizui, T., et al. Myosin II ATPase activity mediates the long-term potentiation-induced exodus of stable F-actin bound by drebrin A from dendritic spines. PLoS One. 9 (1), 85367 (2014).
- Takahashi, H., Mizui, T., Shirao, T. Down-regulation of drebrin A expression suppresses synaptic targeting of NMDA receptors in developing hippocampal neurons. Journal of Neurochemistry. 97, 110-115 (2006).
- Takahashi, H., et al. Drebrin-dependent actin clustering in dendritic filopodia governs synaptic targeting of postsynaptic density-95 and dendritic spine morphogenesis. The Journal of Neuroscience. 23 (16), 6586-6595 (2003).
- Yamazaki, H., Sasagawa, Y., Yamamoto, H., Bito, H., Shirao, T. CaMKIIbeta is localized in dendritic spines as both drebrin-dependent and drebrin-independent pools. Journal of Neurochemistry. 146 (2), 145-159 (2018).
- Hanamura, K., et al. High-content imaging analysis for detecting the loss of drebrin clusters along dendrites in cultured hippocampal neurons. Journal of Pharmacological and Toxicological Methods. 99, 106607 (2019).
- Mitsuoka, T., et al. Assessment of NMDA receptor inhibition of phencyclidine analogues using a high-throughput drebrin immunocytochemical assay. Journal of Pharmacological and Toxicological Methods. 99, 106583 (2019).
- Ishizuka, Y., Bramham, C. R. A simple DMSO-based method for cryopreservation of primary hippocampal and cortical neurons. Journal of Neuroscience Methods. 333, 108578 (2020).
- Pischedda, F., et al. Cryopreservation of primary mouse neurons: The benefit of neurostore cryoprotective medium. Frontiers in Cellular Neuroscience. 12, 81 (2018).
- Kobayashi, Y., et al. Impairment of ciliary dynamics in an APP knock-in mouse model of Alzheimer's disease. Biochemical and Biophysical Research Communications. 610, 85-91 (2022).
- Kobayashi, Y., et al. Properties of primary cilia in melanin-concentrating hormone receptor 1-bearing hippocampal neurons in vivo and in vitro. Neurochemistry International. 142, 104902 (2021).
- Koganezawa, N., et al. NMDA receptor-dependent and -independent effects of natural compounds and crude drugs on synaptic states as revealed by drebrin imaging analysis. The European Journal of Neuroscience. 53 (11), 3548-3560 (2021).
- Mizui, T., Takahashi, H., Sekino, Y., Shirao, T. Overexpression of drebrin A in immature neurons induces the accumulation of F-actin and PSD-95 into dendritic filopodia, and the formation of large abnormal protrusions. Molecular and Cellular Neurosciences. 30 (1), 149-157 (2005).
- Sekino, Y., et al. Activation of N-methyl-D-aspartate receptor induces a shift of drebrin distribution: disappearance from dendritic spines and appearance in dendritic shafts. Molecular and Cellular Neurosciences. 31 (3), 493-504 (2006).
- Takahashi, H., Yamazaki, H., Hanamura, K., Sekino, Y., Shirao, T. Activity of the AMPA receptor regulates drebrin stabilization in dendritic spine morphogenesis. Journal of Cell Science. 122, 1211-1219 (2009).
- Aoki, C., et al. Drebrin A is a postsynaptic protein that localizes in vivo to the submembranous surface of dendritic sites forming excitatory synapses. The Journal of Comparative Neurology. 483 (4), 383-402 (2005).
- Koganezawa, N., Hanamura, K., Sekino, Y., Shirao, T. The role of drebrin in dendritic spines. Molecular and Cellular Neurosciences. 84, 85-92 (2017).
- Shirao, T., et al. The role of drebrin in neurons. Journal of Neurochemistry. 141 (6), 819-834 (2017).
- Sekino, Y., Koganezawa, N., Mizui, T., Shirao, T. Role of drebrin in synaptic plasticity. Advances in Experimental Medicine and Biology. 1006, 183-201 (2017).
- Togo, K., et al. Postsynaptic structure formation of human iPS cell-derived neurons takes longer than presynaptic formation during neural differentiation in vitro. Molecular Brain. 14 (1), 149 (2021).
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoExplorar más artículos
This article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados