
Method Article
Ensamblaje de tendones de ingeniería para sondear la diafonía celular en la enfermedad y la reparación
En este artículo
Resumen
Aquí, presentamos un sistema modelo ensamblado para imitar la diafonía celular del tendón entre el tejido central del tendón que soporta la carga y un compartimento extrínseco que contiene poblaciones celulares activadas por enfermedades y lesiones. Como caso de uso importante, demostramos cómo se puede implementar el sistema para investigar la activación relevante de la enfermedad de las células endoteliales extrínsecas.
Resumen
Los tendones permiten la locomoción transfiriendo fuerzas musculares a los huesos. Se basan en un núcleo tendinoso resistente que comprende fibras de colágeno y poblaciones de células estromalas. Este núcleo de carga está abarcado, nutrido y reparado por una capa de tejido similar a la sinovial que comprende el compartimento del tendón extrínseco. A pesar de este sofisticado diseño, las lesiones tendinosas son comunes y el tratamiento clínico sigue basándose en la fisioterapia y la cirugía. Las limitaciones de los sistemas modelo experimentales disponibles han ralentizado el desarrollo de nuevos tratamientos modificadores de la enfermedad y regímenes clínicos de prevención de recaídas.
Los estudios in vivo en humanos se limitan a comparar tendones sanos con tejidos enfermos o rotos en etapa terminal muestreados durante la cirugía de reparación y no permiten el estudio longitudinal de la enfermedad tendinosa subyacente. Los modelos animales in vivo también presentan límites importantes en cuanto a la complejidad fisiológica opaca, la carga ética sobre los animales y los grandes costes económicos asociados a su uso. Además, los modelos animales in vivo no son adecuados para el sondeo sistemático de fármacos y vías de interacción multicelulares y multitisulares. Los sistemas de modelos in vitro más simples también se han quedado cortos. Una de las principales razones es la incapacidad de replicar adecuadamente la carga mecánica tridimensional necesaria para estudiar de manera significativa las células tendinosas y su función.
El nuevo sistema de modelos 3D presentado aquí alivia algunos de estos problemas mediante la explotación de explantes del núcleo del tendón de la cola murina. Es importante destacar que estos explantes son fácilmente accesibles en grandes cantidades desde un solo ratón, conservan patrones de carga in situ en 3D a nivel celular y cuentan con una matriz extracelular similar a la in vivo. En este protocolo, se dan instrucciones paso a paso sobre cómo aumentar los explantes del núcleo del tendón con hidrogeles de colágeno cargados de células endoteliales derivadas del músculo, fibroblastos derivados del tendón y macrófagos derivados de la médula ósea para sustituir las poblaciones celulares activadas por enfermedades y lesiones dentro del compartimento del tendón extrínseco. Se demuestra cómo los ensamblajes tendinosos resultantes pueden ser desafiados mecánicamente o a través de estímulos microambientales definidos para investigar la diafonía multicelular emergente durante enfermedades y lesiones.
Introducción
En su función de transferir fuerzas musculares a los huesos para permitir el movimiento, los tendones se enfrentan a algunas de las tensiones mecánicas más extremas que ocurren en el cuerpo humano 1,2,3. Debido al envejecimiento de las sociedades, el aumento de la prevalencia de la obesidad y la creciente popularidad de las actividades deportivas mecánicamente exigentes, se prevé que la prevalencia de enfermedades y lesiones tendinosas aumente en los países desarrollados 4,5,6. El desarrollo de nuevos regímenes de tratamiento basados en la evidencia y modificadores de la enfermedad para combatir este aumento se ha visto obstaculizado por las limitaciones de los sistemas modelo actualmente disponibles 1,7,8.
Idealmente, los modelos de reparación de enfermedades y lesiones permitirían estudiar cómo el órgano diana procesa un conjunto definido de parámetros de entrada (imitando los desencadenantes de la enfermedad, Tabla 1) en parámetros de salida medibles (que representan las características distintivas de la enfermedad, Tabla 2) mientras se controlan los factores de confusión. Los estudios que utilicen estos sistemas modelo podrían entonces identificar los procesos fisiológicos (patológicos) que subyacen a la reparación de enfermedades y lesiones y obtener conocimientos que podrían aprovecharse para prevenir o reducir las características distintivas de las enfermedades y lesiones en las clínicas. Aplicando este principio a los tendones, un sistema modelo útil debería recapitular las partes centrales de la respuesta tendinosa in vivo a la enfermedad y la lesión, que abarcan las siguientes características distintivas: microdaño, inflamación, neovascularización, hipercelularidad, recambio acelerado de la matriz y descompartimentación 9,10,11,12,13,14,15 . Utilizando estas características como base, se pueden inferir los siguientes requisitos para un sistema modelo exitoso de reparación de enfermedades y lesiones tendinosas.
Se plantea la hipótesis de que la sobrecarga mecánica es un factor central en la lesión tendinosa y la patogénesis de la enfermedad y, por lo tanto, es un enfoque experimental comúnmente utilizado para crear microdaño16. La capacidad de carga mecánica controlable es, por lo tanto, un requisito previo primordial para los modelos de reparación de enfermedades y lesiones tendinosas. Idealmente, el sistema de modelos permite tres modos principales: carga de estiramiento simple para dañar, carga de fatiga y descarga 8,17,18. Tras la deformación mecánica, las células residentes en los tejidos experimentan una combinación compleja de fuerzas de tensión, fuerzas de cizallamiento (debido al deslizamiento de las fibras de colágeno que rodean las células) y fuerzas de compresión que ocurren durante la descarga o cerca de la entesis19,20. Los sistemas modelo deben recrear estos patrones de carga complejos con la mayor precisión posible.
Una forma alternativa de introducir microdaño en la matriz es aprovechar los estresores bioquímicos que imitan las predisposiciones sistémicas a la enfermedad y lesión de los tendones, como las citocinas (pro)inflamatorias, el estrés oxidativo o las altas concentraciones de glucosa 21,22,23. En consecuencia, un microambiente de nicho controlable es ventajoso para un sistema modelo de reparación de enfermedades y lesiones tendinosas.
Un requisito previo común para que los sistemas modelo puedan recapitular la inflamación, la neovascularización y la hipercelularidad es la presencia selectiva de poblaciones celulares que impulsan estos procesos24. Para los procesos inflamatorios, estas poblaciones incluyen neutrófilos, linfocitos T y macrófagos, mientras que las células endoteliales y los pericitos serían necesarios para estudiar la neovascularización 25,26,27,28,29. Los fibroblastos tendinosos no solo son vitales para la reparación de los tendones, sino que, como células proliferativas y migratorias, también son parcialmente responsables de la hipercelularidad local observada en la enfermedad tendinosa 30,31,32,33,34,35,36.
Además de los cambios en las poblaciones celulares residentes, la composición de la matriz tendinosa también está alterada en la enfermedad y lesión del tendón 7,37,38,39,40. Para presentar las señales microambientales correctas relevantes para la enfermedad, los sistemas modelo deben ser capaces de integrar una composición de matriz extracelular que coincida con la enfermedad o la etapa de lesión objetivo, por ejemplo, permitiendo combinaciones proporcionales relevantes de colágeno-1, colágeno-3 y fibronectina celular41.
La compartimentación de los tendones sanos en el núcleo del tendón y los compartimentos extrínsecos (es decir, endotenón, epitenón y paratenón) es fundamental para su función y, a menudo, se altera en tendones enfermos o lesionados 1,42,43,44,45,46,47 . Por lo tanto, la incorporación de la compartimentación de tendones en 3D en los sistemas modelo de tendones no solo es necesaria para simular más de cerca los procesos subyacentes a la descompartimentación y recompartimentación, sino que también ayuda a establecer los gradientes espacio-temporales correctos de citocinas y nutrientes48,49.
Finalmente, la modularidad es otro activo central de los sistemas modelo, ya que permite a los investigadores combinar la contribución relativa correcta y la interacción entre los estresores previamente descritos durante los procesos investigados 8,17.
Además de seleccionar las modalidades de entrada óptimas, un paso importante es poder medir, observar y realizar un seguimiento de los cambios en la salida resultante. Las propiedades mecánicas del sistema modelo (es decir, la longitud de la región del dedo, el módulo elástico lineal, la tensión máxima por tracción, la tensión máxima por tracción, la resistencia a la fatiga y la relajación de la tensión) son fundamentales aquí, ya que caracterizan la función principal del tendón 50,51,52. Para vincular estos cambios funcionales con los cambios a nivel tisular, es importante habilitar métodos que detecten el daño estructural de la matriz (colágeno) y rastreen la proliferación y el reclutamiento de poblaciones celulares relevantes para la enfermedad y la reparación 30,53,54,55,56,57,58,59,60.
Para estudiar la diafonía emergente célula-célula y célula-matriz, se debe ser capaz de aislar o marcar proteínas en cantidades adecuadas para su cuantificación (es decir, ELISA, proteómica, inmunohistoquímica, citometría de flujo)14,21,61,62. También debería ser posible el análisis de la expresión génica específica de la población o al menos del compartimento (es decir, la clasificación celular activada por fluorescencia [FACS], la secuenciación de ARN de una sola célula/a granel y la reacción en cadena de la polimerasa cuantitativa en tiempo real (RT-qPCR))21,24,27,63. El sistema modelo debe permitir medir la mayor cantidad posible de los parámetros de salida antes mencionados en la misma muestra y en varias muestras de una manera lo suficientemente rápida como para desbloquear estudios de alto rendimiento.
Entre los sistemas modelo disponibles actualmente para estudiar la reparación de enfermedades y lesiones de los tendones humanos, el propio cuerpo humano es, por supuesto, el más representativo. También es el menos compatible con la intervención experimental. Mientras que los pacientes con lesiones agudas de los tendones están abundantemente disponibles para estudios clínicos, los pacientes con tendinopatía temprana (la enfermedad tendinosa más común) están en gran medida libres de síntomas y, a menudo, no se detectan clínicamente hasta que se manifiestan cambios más graves 14,64,65. Esto hace que sea difícil precisar el momento crítico en el que la homeostasis del tendón se descarrila y los mecanismos detrás de este descarrilamiento 16,66,67,68,69. Además, la extracción de biopsias de tendones sanos es un desafío ético, ya que puede provocar daños persistentes. Los restos del tendón isquiotibial de la cirugía de reconstrucción del ligamento cruzado anterior se utilizan a menudo como controles sanos, pero podría decirse que difieren en función, propiedades mecánicas, poblaciones celulares y composición de la matriz en comparación con el manguito rotador, el tendón de Aquiles y los tendones rotulianos comúnmente afectados por enfermedades y lesiones tendinosas 70,71,72,73.
Los modelos animales in vivo son más accesibles y manejables, pero su uso impone una carga ética significativa a los animales y un costo económico a los investigadores. Además, la mayoría de los animales modelo populares no desarrollan lesiones tendinopáticas espontáneamente (es decir, ratas, ratones, conejos) o carecen de los cebadores y cepas modificadas genéticamente necesarias para rastrear las vías de comunicación multicelular involucradas en ella (es decir, caballos, conejos).
Los sistemas simples de modelos in vitro en 2D se encuentran en el otro lado del espectro de complejidad/trazabilidad y permiten un estudio controlado y eficiente en el tiempo de vías de comunicación intercelulares específicas en respuesta a un conjunto más controlable de desencadenantes 8,74. Sin embargo, estos sistemas simplificados comúnmente no recapitulan la carga mecánica multidimensional (es decir, tensión, compresión y cizallamiento) que es fundamental para la funcionalidad del tendón. Además, las (demasiado) altas rigideces del plástico de cultivo de tejidos tienden a anular cualquier señal de matriz proporcionada por los recubrimientos destinados a imitar el estado de interés de la enfermedad75,76.
Para superar este inconveniente, se han desarrollado sistemas de modelos 3D de ingeniería tisular cada vez más sofisticados para proporcionar una matriz cargable cuya composición puede coincidir al menos parcialmente con el estado deseado de la enfermedad 77,78,79. Sin embargo, estos sistemas no solo luchan por replicar con precisión las complejas composiciones de la matriz extracelular in vivo y los patrones de carga celular, sino que generalmente carecen de capacidad de carga a largo plazo y de las interfaces compartimentales necesarias para estudiar las vías de comunicación intercompartimental que coordinan la reparación de enfermedades y lesiones tendinosas 48,49,80.
Los sistemas modelo de explante de tendones ex vivo tienen la clara ventaja de una composición matricial similar a la de in vivo incorporada que comprende nichos pericelulares, barreras intercompartimentales, así como gradientes espaciotemporales de citocinas/nutrientes y recapitula patrones de carga complejos cuando se estira8. Como resultado de los límites de difusión de nutrientes dependientes del tamaño, los explantes de modelos animales más grandes (es decir, caballos) son difíciles de mantener vivos para el estudio a largo plazo de la reparación de enfermedades y lesiones tendinosas 81,82,83. Mientras tanto, los explantes más pequeños de especies murinas (es decir, el tendón de Aquiles, el tendón rotuliano) son difíciles de sujetar de manera reproducible y cargar mecánicamente. Su tamaño también restringe la cantidad de material que se puede recolectar para las lecturas a nivel de células, proteínas y genes sin agrupar muestras y disminuir el rendimiento. En este sentido, los fascículos del tendón de la cola murina ofrecen el potencial de desbloquear el estudio de alto rendimiento de la reparación de enfermedades y lesiones de los tendones, ya que están fácilmente disponibles en grandes cantidades de un solo ratón, preservan la compleja composición de la matriz pericelular in vivo y recapitulan los patrones de carga celular. Sin embargo, durante el proceso de extracción, pierden la mayor parte de su compartimento extrínseco y las poblaciones vasculares, inmunitarias y fibroblásticas que contienen, que ahora se considera que impulsan la enfermedad tendinosa y la reparación 8,18.
Para cerrar esta brecha, se ha desarrollado un sistema modelo que combina las ventajas de los explantes de núcleo derivados del tendón de la cola murina con las ventajas de los sistemas modelo basados en hidrogel 3D. Este sistema modelo consiste en un hidrogel cargado de células (colágeno-1) moldeado alrededor de los explantes del tendón de la cola84,85. En este artículo, se proporcionan en detalle los pasos de fabricación necesarios junto con lecturas útiles que se pueden obtener mediante el cocultivo de explantes centrales (compartimento intrínseco) dentro de un hidrogel de colágeno tipo 1 cargado de células endoteliales (compartimento extrínseco).
Protocolo
Todos los métodos descritos aquí fueron aprobados por las autoridades responsables (números de licencia del cantón de Zúrich ZH104-18 y ZH058-21). En la Figura 1 se presenta una descripción general.
1. Aislamiento de componentes del ensamblaje de tendones de ratones de 12 a 15 semanas de edad (es decir, B6/J-Rj)
- Sacrificar a los ratones mediante asfixia inducida por gas CO2 . Para maximizar el rendimiento, no procese más de 3 ratones a la vez y proceda con el aislamiento celular inmediatamente después de la eutanasia.
- Asegurar la muerte por inducción bilateral de neumotórax.
- Esterilizar la piel del ratón con etanol al 80% y trasladar el ratón a una capucha de bioseguridad estéril.
- Aísle los explantes del núcleo del tendón de la cola.
- Usa un bisturí (No. 21) para separar la cola del ratón cortándola en su base.
- Comenzando por la punta de la cola, agárrala con las pinzas y muévela para romper la piel. Luego, retira suavemente las pinzas de la cola para exponer los explantes del núcleo del tendón.
- Colocar los explantes del núcleo del tendón en el medio de cultivo estándar (DMEM/F12 + 10% FBS + 1% penicilina/estreptomicina + 1% de anfotericina + 1% de aminoácidos no esenciales) y separarlos de la parte de la cola arrancada con una hoja de bisturí nueva (n.º 21).
- Repita los pasos 1.4.2. y 1.4.3. hasta que se procesa toda la cola y los explantes del tendón se acortan más de 25 mm.
- Cortar los explantes de núcleo aislado en trozos de 25 mm de largo con una hoja de bisturí nueva (n.º 21).
- Mida el diámetro medio de los explantes de núcleos con un microscopio óptico conectado a un software de análisis de imágenes a través de una cámara digital acoplable con montura C.
- Use apuntar y hacer clic para seleccionar la herramienta de medición de línea en el lado derecho.
- Mida el diámetro del explante en tres lugares diferentes y calcule su diámetro medio.
- Para facilitar la sujeción y las pruebas mecánicas posteriores, proceda únicamente con explantes de núcleos que tengan un diámetro medio superior a 100 μm.
- La descarga combinada con la exposición a condiciones de cultivo estándar (37 °C, 20%O2, suplementación sérica) cambia la expresión génica dentro de las 6 h posteriores al aislamiento y resulta en degradación dentro de los 7 días21. Para comenzar con un estado cuasi-homeostático, produzca los ensamblajes de tendones y comience los experimentos inmediatamente después del aislamiento del núcleo del tendón.
- Dependiendo de la configuración experimental, se requieren explantes de núcleo de tendón desvitalizados como grupo de control. Para desvitalizar los explantes del núcleo del tendón, congélelos en un recipiente pequeño lleno de nitrógeno líquido durante 5 s con pinzas y luego descongélelos durante 5 s a temperatura ambiente (RT). Repita este ciclo de congelación-descongelación 3 veces y continúe con el paso 4 ("Sujeción de los explantes de núcleo").
PRECAUCIÓN: El nitrógeno líquido puede causar quemaduras por frío, asfixia y fragilizar muchos materiales ordinarios. Solo use recipientes diseñados para líquidos de baja temperatura y use ropa protectora (es decir, protector facial, guantes apropiados, zapatos cerrados).
- Aislar los fibroblastos tendinosos.
- Utiliza un bisturí (n.º 21) para hacer una incisión transversal en el centro de la pata del ratón. Haga dos cortes perpendiculares al pie a lo largo de los lados de las patas traseras y hasta las caderas desde cada extremo de esta incisión.
- Use las pinzas para fijar el colgajo de piel recortado en el pie y retire la piel que cubre los músculos de la pantorrilla. Al aislar células endoteliales del mismo ratón, retire toda la piel.
- Separe el tendón de Aquiles del hueso calcáneo con una hoja de bisturí nueva (n.º 21). Fije el extremo suelto del tendón de Aquiles con las pinzas y separe el otro extremo del músculo gastrocnemio.
- Lave el tendón de Aquiles una vez en PBS y use el bisturí (No. 21) para eliminar todo el tejido muscular restante hasta que solo quede el tejido tendinoso blanco. Si se aíslan células endoteliales del mismo ratón, deje el tendón de Aquiles en PBS y continúe con el paso 1.6. Primero.
- Agrupar los tendones de Aquiles de un animal en un tubo de plástico de 15 ml que contenga 10 ml de medio digestivo tendinoso (DMEM/F12 + 1% de penicilina/estreptomicina + 1% de anfotericina + 2 mg/ml de colagenasa 1) y digerir durante 6-8 h a 37 °C bajo agitación lenta y constante utilizando un agitador orbital de baja velocidad a 15 rpm.
- Centrifugar la solución tendinosa digerida a 500 x g durante 5 min a RT, aspirar el sobrenadante y volver a suspender en 8 mL de medio de cultivo estándar (DMEM/F12 + 10% FBS + 1% penicilina/estreptomicina + 1% anfotericina + 1% aminoácidos no esenciales), y cultivar en un matraz de cultivo T25 en condiciones de cultivo estándar (37 °C, 20%O2) durante 7 días sin cambio de medio. Después de eso, cambie los medios una vez por semana.
- Divida las células al 80% de confluencia en un matraz de cultivo T150 (1:6). Congele las células en el paso 2 en 2 ml de medio de congelación filtrado estéril (70% DMEM/F12 + 20% FBS + 10% DMSO) distribuidas en dos criotubos de 1,5 ml y manténgalas a -80 °C hasta su uso posterior. Utilice tripsina para eliminar las células del plástico de cultivo de tejidos.
- Aislar las células endoteliales derivadas del músculo.
- Si no se aíslan fibroblastos tendinosos del mismo ratón, comience con los pasos 1.5.1 y 1.5.2.
- Use tijeras para separar las patas traseras del cuerpo cortando la articulación de la cadera.
- Lavar las patas traseras una vez en PBS frío (~ 4 °C), retirar los músculos (cuádriceps femoral, extensor largo de los dedos, sóleo y gastrocnemio) con un bisturí (n.º 21) y colocar los músculos en una placa de Petri con hielo.
- Utilice una hoja de bisturí nueva (n.º 21) para picar el tejido muscular en trozos de menos de 1mm3 mientras mantiene la placa de Petri en hielo.
- Agrupe el tejido muscular picado de ambas patas traseras en un tubo de plástico de 50 ml que contenga 12,5 ml de medio de digestión muscular (PBS + 2 mg/ml de colagenasa IV + 2 mg/ml de dispasa II + 2 mM de CaCl2).
- Coloque el tubo de plástico en un baño de agua a 37 °C durante 10 min. Agite vigorosamente la solución y vuelva a colocarla durante otros 10 minutos. Repita hasta que la solución parezca homogénea y solo queden trozos (blancos) de tendón y fascia (aprox. 4 x 10 min). Mientras tanto, continúe con el aislamiento de fibroblastos tendinosos o el aislamiento de macrófagos.
- Agregue 12,5 ml de PBS frío + 10% de FBS al tubo de plástico para detener la digestión.
- Utilice un soporte de pipeta a batería equipado con una pipeta de 50 ml para aspirar la suspensión del tubo de plástico. Equipe el tubo de plástico con un filtro de celdas de 400 μm y filtre la suspensión para eliminar los residuos. Repita el proceso con un colador de células de 100 μm.
- Centrifugar la suspensión filtrada a 400 x g durante 5 min a RT. Volver a suspender en 10 mL de PBS frío + FBS al 10% y volver a centrifugar.
- Resuspender en 8 mL de medio de cultivo endotelial (mezcla 1:1 de DMEM/F12 y el kit endopan 3 + 10 U/mL de heparina + 20% FBS + 1% de penicilina/estreptomicina + 1% de anfotericina + 30 mg/mL de suplemento de crecimiento endotelial) suplementado con puromicina (4 mg/mL) para la selección de la población.
- Siembre las células de un ratón en un matraz de cultivo T25 que previamente se recubrió con 2 ml de una solución estéril de gelatina al 0,2% durante 2 h a 37 °C y luego se secó durante la noche a RT después de eliminar el exceso de solución. Preparar los frascos el día anterior al aislamiento.
- Después de 24 h en condiciones de cultivo estándar (37 °C, 20%Ø2), retirar el medio suplementario de puromicina, lavar las células adheridas una vez con PBS y cultivarlas en 8 mL de medio de cultivo endotelial.
- Pasar las células 1:5 al 80% de confluencia en matraces recubiertos de gelatina y utilizarlos en experimentos hasta P2. Utilice una solución de desprendimiento celular que no sea tripsina (Tabla de materiales) para eliminar las células del plástico de cultivo de tejidos y no las congele.
- Aislar macrófagos derivados de la médula ósea.
- Si no se aíslan fibroblastos tendinosos o células endoteliales del mismo ratón, realice primero los pasos 1.5.1, 1.5.2, 1.6.2 y 1.6.3.
- Después de retirar la piel, el tendón y los tejidos musculares, lavar los huesos sobrantes (fémur y tibia) una vez en PBS frío (~4 °C).
- Coloque los huesos en PBS fresco y frío (~ 4 ° C) y use un bisturí (No. 21) para cortar gradualmente las epífisis hasta que la médula ósea quede expuesta. Aparece como un punto rojo en ambos lados del hueso.
- Equipar una jeringa con una aguja de inyección de 0,4 mm x 25 mm (G27) y llenarla con 10 ml de medio de cultivo de macrófagos (DMEM/F12 + 10% FBS + 1% penicilina/estreptomicina + 1% anfotericina + 1% aminoácidos no esenciales).
- Sostenga un hueso tras otro sobre un tubo de plástico de 50 ml, inserte la aguja de inyección a aproximadamente 1 mm de profundidad en la médula ósea expuesta en la parte superior y enjuague la médula ósea vaciando la jeringa. La médula ósea enrojecida aparece como una estructura rojiza en forma de tubo cuando se suspende en el medio.
- Disuelva la médula ósea pipeteándola suavemente hacia arriba y hacia abajo repetidamente con una punta de pipeta de 1 ml. Utilice un soporte de pipeta a batería equipado con una pipeta de 50 ml para filtrar la suspensión de celdas a través de un filtro de celdas de 100 μm de nuevo en el tubo de plástico de 50 ml y centrifuérrelo a 350 x g durante 5 min en RT.
- Retire el sobrenadante, vuelva a suspender el gránulo en 10 ml de tampón de lisis de glóbulos rojos (RBC) y vuelva a centrifugar a 350 x g durante 10 min a temperatura ambiente.
- Resuspender el pellet en 5 mL de medio de cultivo de macrófagos (DMEM/F12 + 10% FBS + 1% penicilina/estreptomicina + 1% anfotericina + 1% aminoácidos no esenciales) y sembrarlo en placas de Petri no tratadas de 100 mm de diámetro (5-8 x 10,6 células por placa).
- Después de 4 h, añadir 5 ml de medio de cultivo de macrófagos suplementado con 40 ng/ml de factor estimulante de colonias de macrófagos (m-CSF) al medio de cultivo celular sin m-CSF (mezcla 1:1) para llegar a una concentración final de 20 ng/mL de m-CSF.
- Después de 6 días, use las células en experimentos o congélelas hasta que se vuelvan a usar. Utilice una solución de desprendimiento celular que no sea tripsina (Tabla de materiales) para eliminar las células del plástico de cultivo de tejidos.
NOTA: Una vez aisladas, las células ya no se expanden. Los métodos de aislamiento celular descritos aquí también funcionan con ratones y ratas fuera del rango de edad indicado.
- Para verificar el fenotipo de las poblaciones celulares aisladas con citometría de flujo, continúe con el paso 6.3.4.
2. Aislamiento de colágeno de ratas Wistar o Sprague-Dawley
- Seguir el protocolo de aislamiento descrito en detalle en otro lugar86. También funciona con ratones, aunque con un rendimiento mucho menor.
- Determinar la concentración de la solución resultante con un ensayo de hidroxiprolina, evaluar la pureza con SDS-page y almacenar la solución a 4 °C hasta su uso en los experimentos.
3. Producción de los componentes del sistema de cultivo
- Imprime en 3D los soportes de las abrazaderas, la estación de montaje y los moldes de la cámara.
- Cargue el archivo .stl adjunto (Archivo complementario 1) para los soportes de la abrazadera, la estación de montaje y los moldes de la cámara en el software de corte. Para adaptar los números de objeto según sea necesario, utilice apuntar y hacer clic para seleccionar objetos y copiar y pegar para multiplicarlos.
- Presione Exportar código G (Ctrl-R) para generar el código G y, a continuación, exportarlo (Ctrl-G).
- Cargue el código G en una impresora 3D.
- Utilice filamentos biocompatibles sin color para el proceso de impresión (es decir, ácido poliláctico).
- Corte roscas de 3 mm en los orificios del soporte de la abrazadera que llevará los tornillos con un cortador de roscas (Archivo Suplementario 2 y Archivo Suplementario 3, orificios 1 y 3).
- Coloque pasadores de acero inoxidable en el orificio en la parte posterior del soporte de la abrazadera (Archivo Suplementario 2 y Archivo Suplementario 3, orificio 4).
- Esterilice los soportes de la abrazadera y la estación de montaje con luz ultravioleta durante al menos 1 h antes de su uso. No reutilice los soportes de abrazadera impresos en 3D.
- Alternativamente, produzca los soportes de las abrazaderas y la estación de montaje con polieterimida utilizando los planos adjuntos (Archivo Suplementario 2, Archivo Suplementario 3 y Archivo Suplementario 4), que es más costoso pero permite mejores métodos de esterilización (es decir, autoclave) y uso repetido.
- Fundir las cámaras utilizando los moldes impresos en 3D.
- Llene los moldes de la cámara con silicona.
- Desgasificar la silicona en una cámara de vacío (90 mbar) durante 30 min.
- Deje que la solución polimerice a RT durante la noche o en una placa calefactora a 70 °C durante 1 h, dependiendo de la resistencia al calor de los filamentos utilizados para los moldes.
- Retire con cuidado las cámaras polimerizadas de los moldes y corte la silicona superflua con un bisturí (n.º 21).
- OPCIONAL: Si los conjuntos y, por lo tanto, las cámaras circundantes se van a cargar mecánicamente, refuerce los orificios de las cámaras de silicona con tubos huecos de acero inoxidable.
- Mecanizar las abrazaderas metálicas de acero inoxidable utilizando el plano adjunto (Archivo complementario 5).
- Antes de cada uso, lave las abrazaderas de acero inoxidable, los soportes de las abrazaderas de polieterimida, los tornillos y la cámara de silicona.
- Sonicato durante 10 min en etanol al 80% (EtOH) y agua de ósmosis inversa al 20% (ROW).
- Sonicato durante 10 min en 50% de EtOH y 50% de Isopropanol.
- Enjuague 3 veces con ROW.
- Sonicato durante 10 min en concentrado limpiador alcalino al 0,5% (es decir, 3 mL en 600 mL de ROW).
- Sonicato durante 10 min en concentrado limpiador alcalino al 0,5%.
- Dejar actuar el concentrado limpiador alcalino al 0,5% agitando durante 1 h 50 min.
- Enjuague 3 veces con ROW.
- Sonicar durante 10 min en ROW.
- Secar los componentes al aire y esterilizarlos en autoclave.
4. Sujeción de los explantes del núcleo
- Coloque los soportes de abrazadera correspondientes junto con una abrazadera de metal cada uno en la estación de montaje.
- Coloque trozos de papel húmedos esterilizados en autoclave (4 mm x 25 mm) encima de las pinzas metálicas y luego corte el papel a lo largo de los bordes interiores de las pinzas con un bisturí (n.º 21). Corta 2 trozos de papel adicionales más pequeños (4 mm x 1,5 mm) de otro trozo de papel y mantenlos húmedos.
- Con unas pinzas puntiagudas, coloque 8 explantes de núcleo en el papel entre las abrazaderas metálicas con sus extremos en las abrazaderas metálicas.
- Cubra los extremos de los explantes de núcleo con los trozos de papel más pequeños (4 mm x 1,5 mm) preparados y luego coloque abrazaderas de metal encima de ellos. Utilice un destornillador y los tornillos pequeños (M3 x 6 mm) para fijar los explantes de núcleo entre las abrazaderas metálicas y el soporte de la abrazadera.
- Transfiera con cuidado los explantes del núcleo sujetados a las cámaras de cultivo de silicona y llene estas cámaras con 2 ml de medio de cultivo celular estándar (DMEM/F12 + 10% FBS + 1% de penicilina/estreptomicina + 1% de anfotericina + 1% de aminoácidos no esenciales).
- OPCIONAL: Si los conjuntos/las cámaras circundantes se van a cargar mecánicamente, fíjelos con tornillos adicionales (M3 x 16 mm) en el orificio 3 (Archivo suplementario 2 y Archivo suplementario 3, orificio 3).
5. Preparación y fundición de hidrogel de colágeno
- Retirar las células diana del plástico de cultivo de tejidos con solución de desprendimiento celular, centrifugarlas a 400 x g durante 5 min a RT y volver a suspenderlas en 1 mL de medio de cultivo estándar.
- Para un ensamble, se requieren 10 μL de PBS (20x), 1,28 μL de NaOH 1 M (125x), 8,72 μL de agua bidestilada (ddH2O, 23x), 80 μL de colágeno-1 (2,5x o 1,6 mg/mL final) y 100 μL (2x) de medio de cultivo estándar (para ensamblajes sin núcleo // células) o suspensión celular. Prepare estos componentes en dos soluciones separadas y mézclelos solo inmediatamente antes de la fundición.
- Solución de reticulación: Agrupe el PBS, el NaOH, el ddH20 y la suspensión celular de hasta 12 ensamblajes (+10% de margen de seguridad) en una solución de reticulación y manténgala en un tubo de plástico de 15 ml sobre hielo. Ajuste la concentración de la suspensión celular para lograr las siguientes concentraciones finales después de mezclar las dos soluciones: 250.000 células/ml de fibroblastos tendinosos, 500.000 células/ml de células endoteliales derivadas del músculo o 370.000 células/ml de macrófagos derivados de la médula ósea.
- Solución de colágeno-1: Agrupe la solución de colágeno-1 necesaria para hasta 12 ensamblajes (+10% de margen de seguridad) en otro tubo de plástico de 15 ml y manténgalo en hielo.
- Una vez que la solución de reticulación y la solución de colágeno-1 estén listas en hielo, aspire el medio de cultivo celular de las cámaras de cultivo que contienen los explantes del núcleo sujeto.
- Agregue la solución de colágeno-1 a la solución de reticulación con una pipeta de 1000 μL y mezcle las dos soluciones pipeteando hacia arriba y hacia abajo rápidamente sin crear burbujas. Cubra los explantes individuales del núcleo del tendón con 200 μL de la solución mezclada pipeteándola en las ranuras proporcionadas por las cámaras de silicona.
- Deje que los hidrogeles polimericen durante 50 min a 37 °C.
- Llene cuidadosamente las cámaras de cultivo de silicona con 1,5 ml del medio de cocultivo respectivo pipeteándolo en las esquinas de las cámaras.
- Para el cocultivo de núcleos // fibroblastos, llene DMEM/F12, 10% de FBS, 1% de aminoácidos no esenciales, 1% de penicilina/estreptomicina, 1% de anfotericina, 200 μM de ácido L-ascórbico, 20 ng/mL de factor estimulante de colonias de macrófagos.
- Para el cocultivo de núcleos // macrófagos, llene DMEM/F12, 10% de FBS, 1% de aminoácidos no esenciales, 1% de penicilina/estreptomicina, 1% de anfotericina, 200 μM de ácido L-ascórbico, 20 ng/mL de factor estimulante de colonias de macrófagos.
- Para el cocultivo de células endoteliales // núcleo, llene la mezcla 1:1 de DMEM/F12 y el kit endopan 3 + 10 U/ml de heparina + 20% FBS + 1% de penicilina/estreptomicina + 1% de anfotericina + 30 mg/ml de suplemento de crecimiento endotelial.
- Cultive los ensambles en las condiciones de cultivo apropiadas para la hipótesis. Para imitar un entorno de nicho similar a una lesión, por ejemplo, cultívelos a 37 °C y 20% de O2. Cambie el medio de cultivo dos veces en 1 semana. Para prevenir infecciones, coloque las cámaras en una placa de Petri grande o en una caja estéril antes de colocarlas en una incubadora.
NOTA: El tiempo de cultivo depende de la hipótesis y de la configuración del cocultivo. Por ejemplo, los ensamblajes de fibroblastos del núcleo // en un entorno de nicho similar a una lesión se vuelven mecánicamente inestables después de aproximadamente 3 semanas.
6. Métodos de lectura disponibles
- Realizar microscopía de fluorescencia, incluyendo ensayos de viabilidad y morfología.
- En general, los ensamblajes se pueden visualizar como montajes completos. Para hacerlo, retire los ensamblajes de las abrazaderas cortándolos con unas tijeras cerca de las abrazaderas y transfiriéndolos a una placa de 12 pocillos.
- Lave los ensamblajes una vez con PBS.
- Si se realiza un análisis de viabilidad, tiñir cada conjunto con 100 μL de homodímero de etidio 4 x 10−6 M en PBS (EthD-1) durante 20 min a 37 °C en la oscuridad.
- Lave los conjuntos 3 veces con PBS, luego fíjelos con 500 μL de formaldehído al 4% cada uno durante 20 minutos a RT.
PRECAUCIÓN: El formaldehído al 4% tiene efectos alergénicos, cancerígenos y mutagénicos, es tóxico para la reproducción y puede causar toxicidad para el desarrollo (tóxico para la reproducción) o daño a los órganos. Use ropa protectora y guantes, protección para los ojos y mascarilla u otra protección respiratoria. - Lave los ensamblajes 3 veces con PBS y continúe con el protocolo de tinción de su elección. Previamente se ha descrito una selección de tinciones84,85.
NOTA: Evite las tinciones que utilizan fluoróforos con una longitud de onda de emisión cercana a la de la autofluorescencia de colágeno (alrededor de 480 nm).
- De acuerdo con las instrucciones del fabricante, realice el aislamiento de ARN específico del compartimento para RT-qPCR o secuenciación de ARN de todo el genoma.
- Retire los ensamblajes de las abrazaderas con unas tijeras.
- OPCIONAL: Utilice pinzas para separar los explantes del núcleo del compartimento de hidrogel extrínseco.
- Agrupe 20-24 explantes de núcleo de 20 mm o 2 hidrogeles de colágeno cargados de células para aislar cantidades suficientes de ARN.
- Use 1 ml de trizol frío y disrupción mecánica (es decir, perlas de metal o molienda criogénica) para destruir la matriz extracelular de los explantes del núcleo agrupado o los hidrogeles de colágeno combinados.
PRECAUCIÓN: Toxicidad oral, dérmica y por inhalación. Causa irritación de la piel y los ojos. Solo manipule con guantes y en un gabinete de seguridad química. - Continuar con el aislamiento del ARN del lisado celular utilizando kits de extracción de ARN estándar como se describió anteriormente o como se describe en las instrucciones del fabricante 84,85.
- Citometría de flujo específica del compartimento.
- Retire los ensamblajes de las abrazaderas con unas tijeras.
- OPCIONAL: Utilice las pinzas para separar los explantes del núcleo del compartimento de hidrogel extrínseco.
- Digiera los compartimentos en 1 mL de PBS con colagenasa I (3 mg/mL) y dispasa II (4 mg/mL) durante 4 h a 37 °C bajo agitación constante.
- Centrifugar la solución digerida a 500 x g durante 5 min a RT y aspirar el sobrenadante.
- Vuelva a suspender el gránulo en 100 μL de tampón FACS (1% de FBS en PBS) que contenga los anticuerpos conjugados con fluoróforos de elección. Previamente se ha descrito una selección de anticuerpos conjugados con fluoróforos funcionales84,85.
- Incubar la solución de tinción durante 30 min en RT.
- Diluir la solución de tinción con 1,4 ml de tampón FACS y centrifugarla durante 5 min a 500 x g a RT.
- Vuelva a suspender el gránulo en 350 μL de tampón FACS y filtre la solución a través de una tapa coladora de malla de nailon de 100 μm antes de analizarla con el citómetro de flujo de su elección de acuerdo con las instrucciones del fabricante.
- Analiza el sobrenadante.
- Sustituya el medio de cocultivo por un medio de cocultivo sin suero 3 días antes de la recogida del sobrenadante.
- Realice análisis inmediatos y diferidos del sobrenadante enriquecido y sin diluir con kits de ensayo de inmunoabsorción ligado a enzimas (ELISA) y kits de ensayo de descubrimiento a escala meso (MSD). Para el análisis diferido, almacenar el sobrenadante en tubos de plástico de 1,5 ml a -80 °C.
- Evaluar las propiedades mecánicas del conjunto.
- Utilice un dispositivo de estiramiento hecho a medida para aplicar fuerzas mecánicas y medir las propiedades mecánicas22. Los pasadores de acero inoxidable y los tornillos de acero inoxidable hacen que las abrazaderas también se puedan acoplar a otros dispositivos de estiramiento.
- Como las propiedades mecánicas del ensamblaje están determinadas en gran medida por las del explante 18 del núcleo incrustado, mida las propiedades mecánicas del explante del núcleo antes de incrustarlo en un hidrogel para disminuir el riesgo de destruir el hidrogel recién fundido en el proceso de medición.
Resultados
Aislamiento de componentes (Figura 1 y Figura 2)
Antes de utilizar los explantes centrales y las poblaciones celulares en el cocultivo de ensamble, estos componentes deben verificarse bajo el microscopio (Figura 1). Los explantes del núcleo deben tener un diámetro uniforme (100-200 μm) y no tener torceduras ni arrugas visibles. Las células endoteliales deben presentar una forma alargada en contacto con otras células, lo que no ocurre cuando se siembran a una densidad demasiado baja debido a un bajo rendimiento inicial del aislamiento. En este caso, las células endoteliales asumen una forma más redondeada con extensiones citoesqueléticas y proliferan marcadamente más lentamente. Divídelos 1:5 después de 7-10 días. Los fibroblastos tendinosos aislados de los tendones de Aquiles asumen una morfología más redondeada en comparación con sus homólogos humanos en 1-2 pasos (10-14 días cada uno) cuando se dividieron 1:6. Los macrófagos son mucho más pequeños que los fibroblastos o las células endoteliales y no proliferan después del aislamiento. Dependiendo del lote, su forma puede variar de piramidal a redonda.
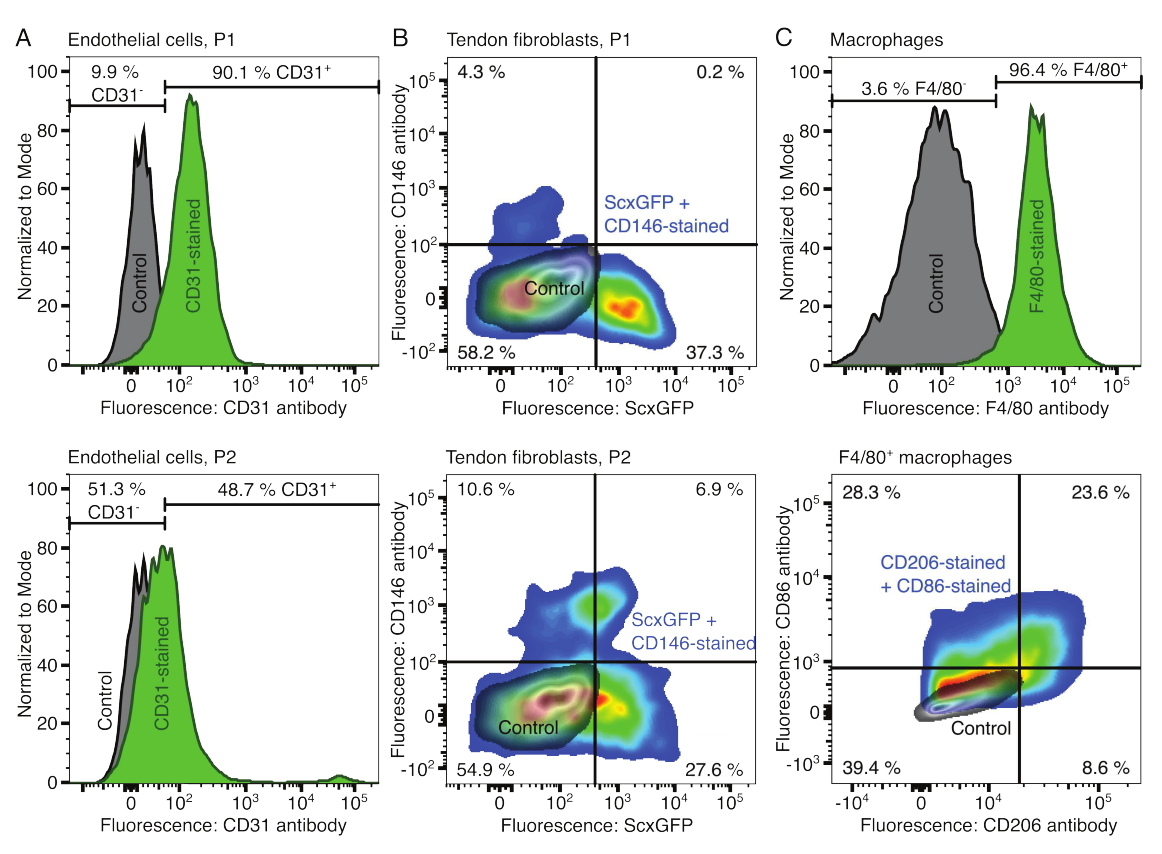
Los fenotipos de los componentes celulares se verificaron con citometría de flujo. Se utilizó un anticuerpo CD31 conjugado como marcador de células endoteliales (Figura 2A). Al establecer el umbral de fluorescencia en función de una muestra de control no teñida (gris), se identificó que el 90,1 % de las células endoteliales del paso 1 (P1) y el 48,7 % del pasaje 2 (P2) eran CD31 positivas. Para caracterizar los fibroblastos tendinosos se utilizó una línea de ratón modificada genéticamente que coexpresa el marcador de fibroblastos tendinosos Scleraxis junto con una proteína verde fluorescente (ScxGFP) y un anticuerpo CD146 conjugado (Figura 2B)35,60. Después de un paso (P1), el 37,3% de los fibroblastos eran ScxGFP+CD146-, el 0,2% ScxGFP+CD146+, el 4,3% ScxGFP-CD146+ y el 58% ScxGFP-CD146-. Después de dos pasadas (P2), el porcentaje de células ScxGFP+CD146- disminuyó al 27,6%, el porcentaje de células ScxGFP+CD146+ aumentó al 6,9%, el porcentaje de células ScxGFP-CD146+ aumentó al 10,6% y el porcentaje de células ScxGFP-CD146- disminuyó al 54,9%. Para identificar y caracterizar los macrófagos, se utilizó un anticuerpo F4/80 en combinación con un anticuerpo CD86 y un anticuerpo CD206 (Figura 2C). Tras el aislamiento y el cultivo, el 96,4% de las células derivadas de la médula ósea fueron positivas para F4/80. Entre estas células positivas para F4/80, el 8,6% eran CD206+CD86-, el 23,6% CD206+CD86+, el 28,3% CD206-CD86+ y el 39,4% CD206-CD86-. La velocidad de reticulación del colágeno puede variar de un lote a otro y debe probarse antes de comenzar los experimentos.
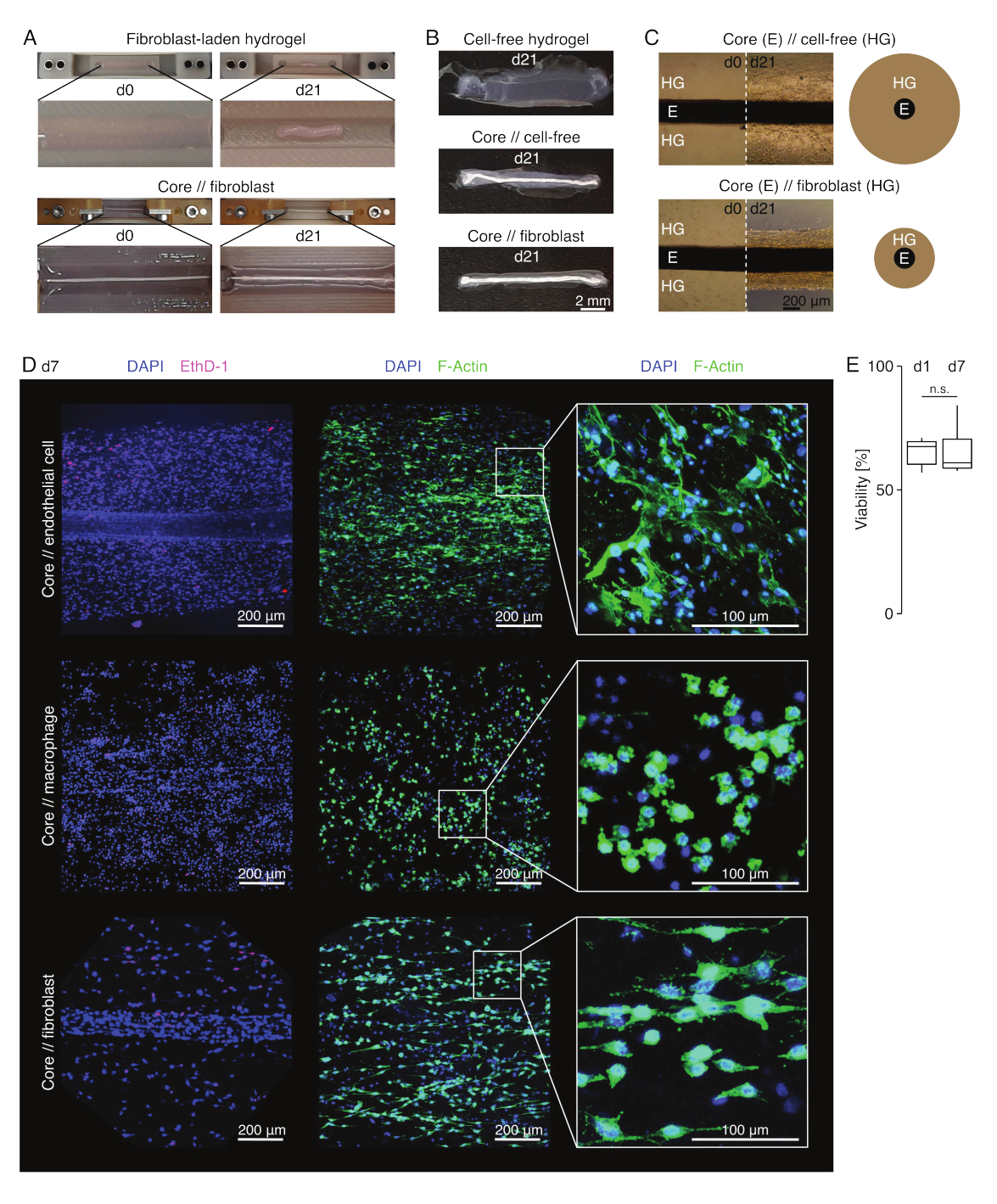
Apariencia del ensamblaje (Figura 3)
En condiciones de cultivo similares a las de una lesión (36 °C, 20%Ø2), el explante del núcleo permaneció mecánicamente estirable, no cambió de apariencia y continuó siendo visualmente distinguible y físicamente separable del hidrogel circundante durante al menos 21 días (Figura 3A, B). El hidrogel circundante se compactó con el tiempo, y la velocidad de compactación dependió de la población celular sembrada en él. Los fibroblastos derivados del tendón de Aquiles contrajeron el hidrogel circundante más rápidamente y lo hicieron radialmente cuando estaban en un hidrogel moldeado alrededor de un explante central y en todas las direcciones cuando no lo estaban (Figura 3B, C). Inicialmente, los hidrogeles libres de células colocados alrededor de un explante de núcleo también se compactaron. Esta contracción probablemente fue causada por la migración de células desde el explante central, lo que indica una interfaz dinámica entre compartimentos. Como los hidrogeles libres de células sin un explante de núcleo incrustado no se compactaron de manera detectable, la contribución de la contracción inducida por la pérdida de agua parece ser insignificante (Figura 3B y Archivo Suplementario 6).
Por lo tanto, la falta de compactación de hidrogel puede utilizarse para detectar errores en el ensamblaje del ensamblaje (es decir, bajas concentraciones de células) y debe verificarse antes de continuar con métodos de lectura más costosos. Al establecer este método, los errores comunes que reducían la concentración celular incluían la muerte de las células en el hidrogel extrínseco porque se dejaban durante demasiado tiempo en la solución de reticulación relativamente dura (pH alto, baja temperatura) y el secado de los explantes del núcleo porque el tiempo entre la aspiración del medio y la inyección del hidrogel era demasiado largo, o porque el explante del núcleo estaba sujetado demasiado alto para incrustarse en el colágeno.
Microscopía de fluorescencia confocal: análisis de viabilidad y morfología (Figura 3)
Una vez retirados de las pinzas con tijeras (Figura 3B), los ensamblajes se pueden fijar, teñir y obtener imágenes con un microscopio confocal en su conjunto sin necesidad de seccionarlos. Aquí, los ensamblajes de núcleo // célula endotelial, núcleo // macrófago y núcleo // fibroblastos se tiñeron con DAPI (NucBlue) y homodímero de etidio (EthD-1) para analizar la viabilidad y DAPI y F-actina para analizar la morfología y la propagación celular en el hidrogel de colágeno 3D (Figura 3D). Se cuantificó la viabilidad de los ensamblajes de células endoteliales del núcleo (Figura 3E) y se encontró que era generalmente menor después del ensamblaje del ensamblaje de los ensambloides que lo informado previamente para los ensamblajes de núcleos // macrófagos y núcleos // fibroblastos84. Sin embargo, la viabilidad se mantuvo estable durante el cultivo ensamblado hasta al menos el día 7.
Microdaño inducido mecánicamente y medición de propiedades mecánicas (Figura 4)
Los tornillos y pasadores unidos a los soportes de abrazadera permiten la fijación de conjuntos sujetados a dispositivos de estiramiento uniaxiales. El dispositivo de estiramiento hecho a medida utilizado aquí está equipado con una célula de carga de 10 N y ha sido descrito en publicaciones anteriores (Figura 4A)22. Todas las muestras se acondicionaron previamente con cinco ciclos de estiramiento al 1% de deformación antes de las mediciones.
El registro de la curva completa de tensión-deformación de los explantes o ensamblajes de núcleos (Figura 4B) permitiría cuantificar el módulo elástico lineal (α), la tensión máxima (β) y la deformación máxima (у). Sin embargo, también daña irreversiblemente el explante del núcleo o el ensamblaje, lo que hace imposible evaluar el desarrollo longitudinal de la tensión máxima (β) y la deformación máxima (у) para las mismas muestras (Figura 4B). Aquí, el módulo elástico lineal se utilizó como medida de la capacidad de la muestra para soportar fuerzas, ya que esta medición requiere estirar la muestra a solo un 2% de deformación, lo que se ha demostrado previamente que no causa reducciones permanentes en el módulo elástico lineal18. En particular, los ensamblajes de células centrales // endoteliales se expusieron al procedimiento de pinzamiento a una deformación del 2% (aproximadamente el final de la región elástica lineal) o al 6% de deformación (aproximadamente la deformación máxima). El microdaño resultante se evaluó midiendo el módulo elástico lineal antes y después del procedimiento (Figura 4C).
De acuerdo con los experimentos realizados previamente con explantes de núcleo monocultivados, los ensamblajes de células endoteliales del núcleo retuvieron su módulo elástico lineal durante al menos 14 días cuando se cultivaron en condiciones de nicho cuasi-homeostático (29 °C, 3%O2) y se expusieron a cepas no superiores al 2%18,21. Con respecto a la estimulación mecánica basal, el estiramiento estático aplicado a través de las pinzas pareció imitar suficientemente los niveles de deformación nativa experimentados por las unidades del núcleo tendinoso in vivo para prevenir procesos catabólicos generalmente asociados con la descarga de la matriz87. De hecho, la disminución progresiva y estadísticamente significativa del módulo elástico lineal observada en los ensamblajes de células endoteliales del núcleo expuestos a una tensión del 6% podría atribuirse a la descarga de la matriz derivada del microdaño de la matriz inducido mecánicamente.
Al realizar estos experimentos, es importante evitar que el conjunto se seque. En este caso, se envolvían en papel esterilizado en autoclave y mojado, pero también podían ser viables otros métodos en función de su compatibilidad con el dispositivo de estiramiento utilizado. Como la fricción entre las abrazaderas metálicas y el explante del núcleo es limitada, agregue pequeños trozos de papel entre el metal y el explante del núcleo durante la sujeción para evitar el deslizamiento y supervise de cerca el proceso de estiramiento para detectar y excluir los explantes y ensamblajes del núcleo deslizados.
Análisis del transcriptoma específico del compartimento y del secretoma específico del conjunto (Figura 5 y Figura 6)
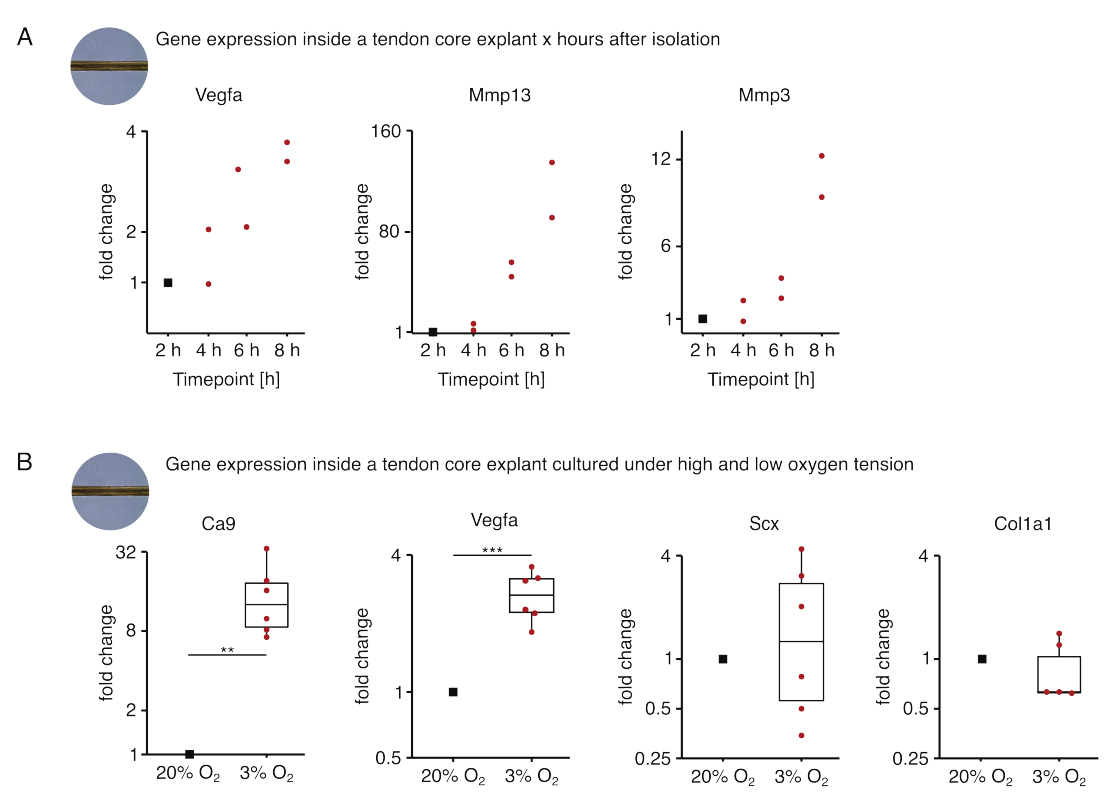
En el primer conjunto de experimentos de monocultivo del núcleo que se presenta aquí, se evaluó la estabilidad de la expresión génica del núcleo después del aislamiento del explante para desacoplar el aislamiento de los efectos experimentales (Figura 5A). Aunque se necesita un mayor número de réplicas para obtener conclusiones precisas, la expresión de Vegfa y Mmps aumentó fuertemente en los explantes de núcleo recién aislados a las pocas horas del aislamiento del explante cuando se cultivaron en condiciones de nicho similares a lesiones (37 °C, 20% O2).
La neovascularización es un sello distintivo central de la enfermedad y la reparación del tendón que podría, en parte, ser impulsada por células endoteliales activadas por factores proangiogénicos (es decir, el factor de crecimiento endotelial vascular, Vegfa) secretado por el núcleo del tendón bajo hipoxia88. Examinando el primer paso de esta posible diafonía (Figura 5B), se encontró que la expresión tanto de Vegfa como del marcador de hipoxia anhidrasa carbónica 9 (Ca9) aumentó de manera estadísticamente significativa en los explantes monocultivados bajo baja tensión de oxígeno (3% O2) en contraste con los monocultivados bajo alta tensión de oxígeno (20% O2). Mientras tanto, la menor tensión de oxígeno no pareció causar cambios en la expresión de marcadores de fibroblastos tendinosos como Scleraxis (Scx) y colágeno-1 (Col1a1). En conjunto, estos resultados identifican a las células residentes en el núcleo como contribuyentes plausibles a la señalización proangiogénica en un nicho hipóxico.
A continuación, se evaluó la activación de las células endoteliales por la señalización proangiogénica del núcleo en el cocultivo del núcleo // ensamblaje de células endoteliales bajo alta (20% O2) y baja (3% O2) tensión de oxígeno. Afortunadamente, la composición modular de los ensambles permite el análisis del transcriptoma específico del compartimento después del cultivo al separar físicamente el explante central del hidrogel de colágeno extrínseco (Figura 6A). En el explante del núcleo (Figura 6D), se confirmó de nuevo que la expresión de Vegfa aumentaba bajo baja tensión de oxígeno, aunque el efecto sobre otros marcadores hipóxicos como Fgf2 fue menos claro y requiere un mayor número de réplicas para obtener conclusiones precisas. Además, la expresión de marcadores proinflamatorios como el Tnf-α y los marcadores de degradación de la matriz extracelular como el Mmp3 disminuyeron en el núcleo bajo baja tensión de oxígeno. En el hidrogel extrínseco sembrado inicialmente con células endoteliales (Figura 6E), la presencia de un explante de núcleo vivo (aC) disminuyó la expresión de Vegfa bajo baja tensión de oxígeno, pero no bajo alta tensión de oxígeno. Además, la presencia de un explante de núcleo desvitalizado (dC) bajo baja tensión de oxígeno tampoco disminuyó la expresión de Vegfa . Bajo baja tensión de oxígeno, la expresión de Tnf-α en el hidrogel extrínseco fue comparable alrededor de aC/dC, pero aumentó bajo alta tensión de oxígeno alrededor de los explantes de núcleo vivo. La expresión de Fgf2 disminuyó en todas las condiciones en comparación con el hidrogel extrínseco cargado de células endoteliales cultivado alrededor de un explante de núcleo desvitalizado bajo alta tensión de oxígeno, pero la mayoría bajo baja tensión de oxígeno. La expresión de Mmp3 fue más alta alrededor de los explantes de núcleo vivo bajo alta tensión de oxígeno y más baja alrededor de los explantes de núcleo desvitalizados bajo baja tensión de oxígeno. En general, las células endoteliales cocultivadas parecen responder tanto al explante del núcleo activo, que es capaz de iniciar la diafonía como a las variaciones en los niveles de oxígeno. Un análisis más exhaustivo del transcriptoma facilitaría la elucidación de sus respectivas contribuciones.
La modularidad del sistema ensamblado permite la integración de células modificadas genéticamente que contienen genes reporteros fluorescentes. Aquí, las células endoteliales aisladas de ratones Pdgfb-iCreER mG89 se sembraron en el compartimento del hidrogel. Estas células coexpresan la subunidad b del factor de crecimiento derivado de plaquetas (Pdgfb) del marcador de células endoteliales junto con la proteína fluorescente verde mejorada (EGFP), que hace que las células endoteliales que expresan Pdgfb aparezcan verdes bajo la microscopía (Figura 6C). Utilizando este método, se confirmó que la presencia de células endoteliales que expresan Pdgfb se mantuvo durante 7 días en cultivo (37 °C) y pareció ser independiente de la tensión de oxígeno (20%O2 frente a 3%O2).
Para analizar el secretoma de los ensambles, el medio de cultivo utilizado respectivamente para el cocultivo de células libres de células y el núcleo // fibroblastos, el núcleo // macrófagos o el cocultivo de células endoteliales centrales // se reemplazó por su contraparte libre de suero tres días antes de aspirar y congelar el sobrenadante ahora enriquecido con el secretoma (Figura 6A). Este tiempo de enriquecimiento fue suficiente para detectar citocinas como el factor de crecimiento endotelial vascular (VEGF) con un ensayo de MSD, como se muestra aquí para explantes de núcleos y ensamblajes de fibroblastos de núcleos // cultivados en condiciones de nicho similares a lesiones (Figura 6B).
Las consideraciones importantes al analizar los secretomas y transcriptomas de los explantes y ensambles de núcleos se refieren al uso de controles adecuados. Los explantes de núcleo recién aislados tienen un valor limitado, ya que especialmente su expresión de Vegfa y Mmps aumenta fuertemente a las pocas horas del aislamiento (Figura 5A). Los explantes emparejados en el tiempo rodeados por un hidrogel inicialmente libre de células son más adecuados como controles para la expresión génica del compartimento central. Para el hidrogel extrínseco, los hidrogeles cargados de células cultivados sin un explante de núcleo son controles inferiores en comparación con los hidrogeles cargados de células cultivados alrededor de explantes de núcleo desvitalizados (Archivo Suplementario 7), principalmente porque se compactan en formas redondeadas en lugar de hidrogeles alargados, lo que cambia en gran medida la morfología celular (Figura 3A).

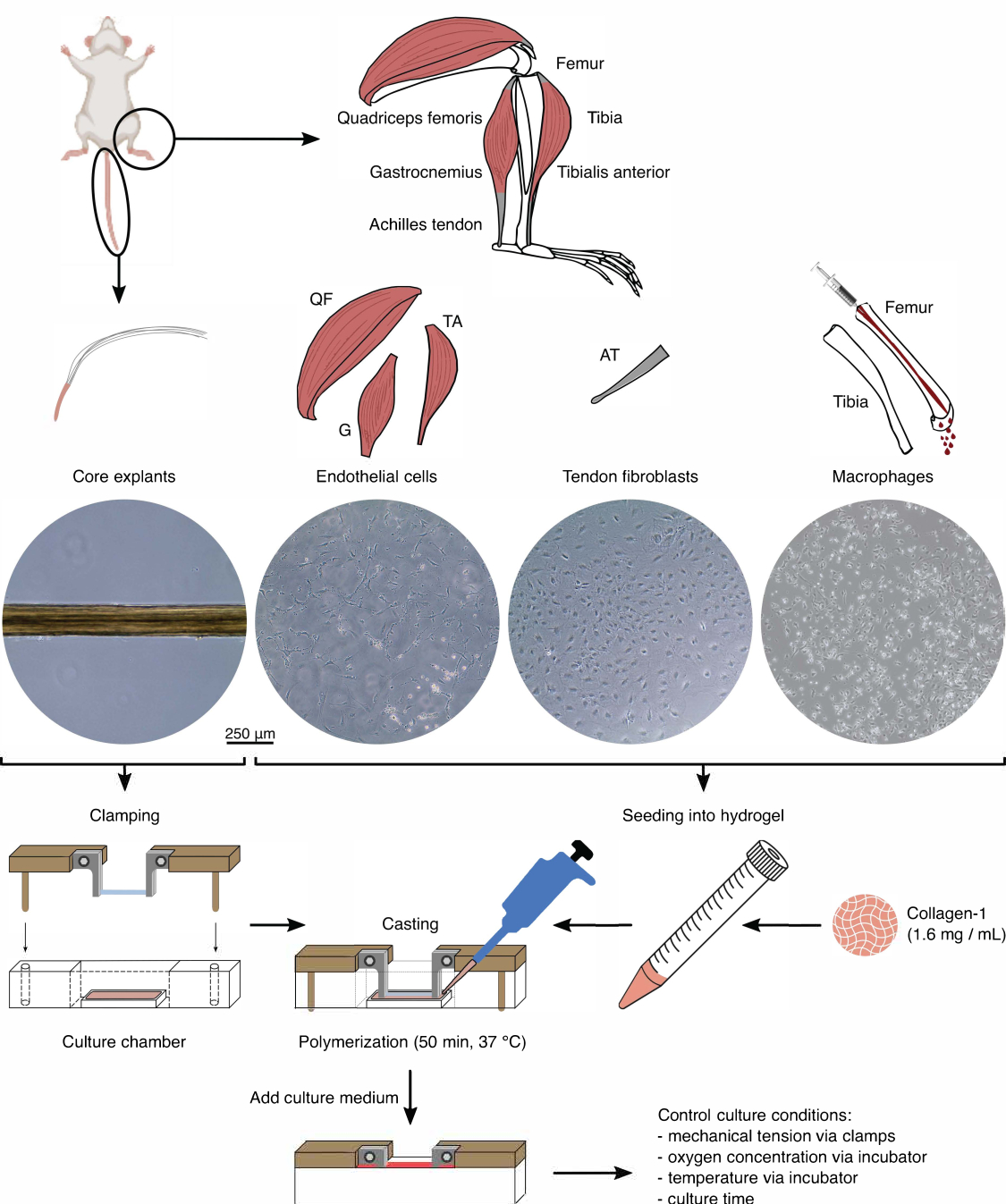
Figura 1: Aislamiento de componentes de ensamblaje y ensamblaje para modelar diafonía in vivo . Se extrajeron explantes de núcleos tendinosos de colas de ratón, se cortaron y se sujetaron. Los músculos de las patas de ratón (es decir, el cuádriceps femoral (QF), el gastrocnemio (G) y el tibial anterior (TA)) se digierieron para aislar células endoteliales que luego se cultivaron en plástico de cultivo de tejidos. Los tendones de Aquiles (AT) también se digierieron para aislar los fibroblastos tendinosos, que luego se cultivaron en plástico de cultivo de tejidos. La médula ósea de la tibia y el fémur fue expulsada de los huesos. A continuación, los monocitos aislados se cultivaron en plástico de cultivo de tejidos y se diferenciaron en macrófagos naïf. Las imágenes de microscopía óptica (10x) muestran la apariencia de los explantes centrales, las células endoteliales, los fibroblastos tendinosos y los macrófagos inmediatamente antes de su integración en los ensambles. Durante el ensamblaje, las células cultivadas en plástico se pusieron en suspensión y luego se sembraron en una solución de colágeno-1 (1,6 mg/mL). A continuación, la mezcla de célula e hidrogel se moldeó alrededor del explante del núcleo sujetado y se polimerizó durante 50 min a 37 °C antes de añadir el medio de cultivo. Las condiciones de cultivo se controlaron a través de las pinzas (tensión mecánica) y los ajustes de la incubadora (concentración de oxígeno, temperatura). Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 2: Caracterización de los componentes del ensamblaje celular. (A) Análisis citométrico de flujo representativo de células endoteliales derivadas del músculo después de un paso (P1, fila superior) y dos pases (P2, fila inferior). Los recuentos de células no teñidas (gris) y teñidas con CD31 (verde) se normalizaron a modal. Los porcentajes se dan para el grupo teñido con CD31. (B) Análisis citométrico de flujo representativo de fibroblastos derivados del tendón de Aquiles después de un paso (P1, fila superior) y dos pasajes (P2, fila inferior). Los ejes informan de las intensidades de fluorescencia de las células no teñidas (gris) y de las células que expresan ScxGFP y teñidas con anticuerpos CD146 (colores del arco iris). (C) Análisis citométrico de flujo representativo de macrófagos derivados de la médula ósea después del cultivo. En la fila superior, los recuentos de células no teñidas (gris) y teñidas con F4/80 (verde) se normalizaron a modal. Los porcentajes se dan para el grupo teñido con F4/80. El gráfico de la fila inferior informa de las intensidades de fluorescencia de las células no teñidas (gris) y del subconjunto F4/80+ de células teñidas con anticuerpos CD206 y anticuerpos CD86 (colores del arco iris). Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 3: Imágenes y apariencia del ensamblado. (A) Las fotografías representativas tomadas en el día 0 (d0) y el día 21 (d21) de cultivo (37 °C, 20%Ø2) muestran una contracción multidimensional de un hidrogel que contiene fibroblastos extrínsecos sin un explante de núcleo incrustado y una fuerte compactación radial de un hidrogel que contiene fibroblastos extrínsecos alrededor de un explante de núcleo. (B) Las fotografías representativas tomadas en el día 21 (d21) de cultivo (37 °C, 20%O2) muestran diferencias en la velocidad de compactación entre los hidrogeles libres de células, los hidrogeles libres de células fundidos alrededor de un explante de núcleo y los hidrogeles cargados de fibroblastos tendinosos moldeados alrededor de un explante de núcleo. (C) Las imágenes representativas de microscopía óptica (10x) tomadas en el día 0 (d0) y el día 21 (d21) de cultivo (37 °C, 20%Ø2) indican cambios longitudinales en la presencia de poblaciones celulares y la velocidad de compactación del hidrogel de colágeno (HG) alrededor del explante del núcleo (E) en el cocultivo de núcleos // libres de células y núcleo// fibroblastos. La representación esquemática muestra las diferencias en la compactación de hidrogel entre el núcleo // ensamblaje libre de células y el co-cultivo de núcleo // ensamble de fibroblastos. (D) Imágenes representativas de microscopía confocal tomadas en el día 7 (d7) del cocultivo de núcleos // células endoteliales, núcleo// macrófagos y núcleo// fibroblastos (37 °C, 20% Ø2). Las imágenes de la fila izquierda muestran ensamblajes con núcleos celulares teñidos en azul (DAPI) y células muertas teñidas en rosa (Ethidium homodimer-1). Las otras dos filas representan ensamblajes con núcleos celulares teñidos en azul (DAPI) y filamentos de actina en verde (F-actina). (E) Diagramas de caja que representan la viabilidad cuantificada de los ensamblajes de células centrales // endoteliales en el día 1 (d1) y el día 7 (d7) de cocultivo. N = 5. Las bisagras superior e inferior corresponden al primer y tercer cuartil (percentiles 25y 75) y la del medio a la mediana. Los bigotes se extienden desde la bisagra superior/inferior hasta el valor más grande/más pequeño no más allá de 1,5 veces el rango intercuartílico. Valores p: n.s.p > 0,05. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 4: Estimulación mecánica de ensamblajes y medición de las propiedades mecánicas de ensamblajes. (A) Representación gráfica del dispositivo de estiramiento hecho a medida que comprende las plataformas de soporte de abrazaderas, un sensor de fuerza y un motor paso a paso. La imagen fotográfica muestra un conjunto montado en el dispositivo de estiramiento con abrazaderas. La tapa de un tubo de plástico de 15 ml (Ø: 17 mm) utilizado para la báscula. (B) Gráfico que muestra curvas representativas de tensión/deformación para explantes de núcleo (azul claro) y ensambles (rojo claro). El módulo elástico lineal (α), la tensión máxima (β) y la deformación máxima (у) se pueden extraer de los datos para caracterizar mecánicamente el explante o ensamblaje del núcleo. (C) Gráfico que muestra el módulo elástico lineal (Emod) de los ensamblajes de células endoteliales del núcleo // co-cultivados (29 °C, 3% O2) durante un curso de tiempo de 14 días después de ser sujetados (línea continua), sujetados y estirados al 2% L0 deformación (línea punteada), o sujetados y estirados al 6% L0 deformación (línea discontinua) al inicio del experimento. N = 5. Los puntos de datos se normalizaron al módulo elástico lineal inicial antes del estiramiento y todos se muestran como media (±sem). Valores p: *p < 0,05, **p < 0,01. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 5: Cambios en el transcriptoma central después del aislamiento y cultivo en diferentes condiciones de nicho. (A) Diagrama de dispersión que representa los cambios de plegamiento en la expresión génica de Vegfa, Mmp13 y Mmp3 en explantes de núcleo murino monocultivados (37 °C, 20% O2) 2 h, 4 h, 6 h y 8 h después de aislarlos de la cola. Los cambios de plegamiento en los respectivos puntos de tiempo se normalizaron a la expresión génica 2 horas después del aislamiento. N = 2. (B) Diagramas de caja que representan los cambios de plegamiento en la expresión génica de Ca9, Vegfa, Scx y Col1a1 en explantes centrales monocultivados bajo baja tensión de oxígeno (3% O2) normalizados y comparados con los monocultivados bajo alta tensión de oxígeno (20% O2). N = 5-6. Las bisagras superior e inferior de los diagramas de caja corresponden al primer y tercer cuartil (percentiles25 y 75) y la del medio a la mediana. Los bigotes se extienden desde la bisagra superior/inferior hasta el valor más grande/más pequeño no más allá de 1,5 veces el rango intercuartílico. Los puntos de datos utilizados para la normalización se representan como puntos negros y los puntos de datos individuales como puntos rojos. Valores p: **p < 0,01, ***p < 0,001. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 6: Análisis del secretoma específico del ensamble y del transcriptoma específico del compartimento. (A) Fotografía representativa que muestre el ensamblador en el día 7 (d7), cuando se tomaron muestras del secretoma y el transcriptoma, y representación del flujo de trabajo subyacente. (B) Concentración de VEGF (pg/mL) en el sobrenadante de los ensambles de núcleos // libres de células y núcleos // fibroblastos después de 7 días de cocultivo (37 °C, 20%O2) representados como diagramas de caja. N = 6. (C) Imágenes representativas de microscopía confocal de ensamblajes de células endoteliales después de 7 días de cocultivo (37 °C) bajo alta tensión de oxígeno (20% O2) y baja tensión de oxígeno (3% O2). Los núcleos celulares se tiñen en azul (DAPI) y las células endoteliales incrustadas coexpresan la proteína verde fluorescente mejorada (EGFP) junto con la subunidad b del factor de crecimiento derivado de plaquetas (Pdgfb) del marcador de células endoteliales. La línea punteada indica la interfaz compartimental entre el explante central (E) y el hidrogel cargado de células endoteliales (HG). (D) Diagrama de dispersión que representa los cambios de plegamiento en la expresión génica de Vegfa, Tnf-α, Fgf2 y Mmp3 en el compartimento central de ensamblajes de células centrales // endoteliales cocultivados bajo baja tensión de oxígeno (3% O2) normalizados y comparados con aquellos cultivados bajo alta tensión de oxígeno (20% O2). N = 2. (E) Diagrama de dispersión que representa los cambios de plegamiento en la expresión génica de Vegfa, Tnf-α, Fgf2 y Mmp3 en el compartimento extrínseco del núcleo // ensamblajes de células endoteliales con un núcleo vivo (aC) o un núcleo desvitalizado (dC) cocultivado bajo alta tensión de oxígeno (20% O2) y baja tensión de oxígeno (3% O2). Los cambios de plegamiento en las condiciones respectivas se normalizaron en el compartimento extrínseco de un núcleo // conjunto de células endoteliales con un núcleo desvitalizado (dC) co-cultivado bajo alta tensión de oxígeno (20%O2). N = 3-4. En B, las bisagras superior e inferior de los diagramas de caja corresponden al primer y tercer cuartil(percentiles 25y 75) y la intermedia a la mediana. Los bigotes se extienden desde la bisagra superior / inferior hasta el valor más grande / más pequeño no más allá de 1,5 veces el rango intercuartílico. Los valores atípicos se representan como puntos negros. Valores p: *p < 0,05. En D y E, los puntos de datos utilizados para la normalización se representan como puntos negros y los puntos de datos individuales se representan como puntos rojos. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
Tabla 1: Requisitos de entrada para los sistemas modelo de enfermedades y lesiones tendinosas. Una lista de desencadenantes de enfermedades tendinosas primarias y factores secundarios coincidentes con una selección de parámetros de entrada cuya manejabilidad es fundamental para modelar enfermedades y lesiones tendinosas. Haga clic aquí para descargar esta tabla.
Tabla 2: Requisitos de salida para los sistemas modelo de enfermedades y lesiones tendinosas. Una selección de las características distintivas de la enfermedad tendinosa coincidió con una selección de parámetros de salida cuya cuantificabilidad es central para la interpretación del comportamiento del modelo de enfermedad y lesión del tendón. Haga clic aquí para descargar esta tabla.
Archivo suplementario 1: archivo .stl para los soportes de las abrazaderas, la estación de montaje y los moldes de la cámara. Haga clic aquí para descargar este archivo.
Archivo complementario 2: Plano del soporte de la abrazadera derecha. Haga clic aquí para descargar este archivo.
Archivo complementario 3: Plano del soporte de la abrazadera izquierda. Haga clic aquí para descargar este archivo.
Archivo complementario 4: Plano de la plataforma de montaje Haga clic aquí para descargar este archivo.
Ficha complementaria 5: Plano de abrazaderas metálicas. Haga clic aquí para descargar este archivo.
Archivo complementario 6: Imagen que muestra la contracción del hidrogel libre de células. Haga clic aquí para descargar este archivo.
Archivo Suplementario 7: Imagen que muestra un explante de núcleo desvitalizado. Haga clic aquí para descargar este archivo.
Discusión
En general, el sistema de modelos ensamblados que se presenta aquí tiene varios pasos críticos que destacar. En primer lugar, el sistema de modelos es tan bueno como la calidad de sus componentes. Es vital comprobar el explante central y las poblaciones celulares que se van a sembrar bajo el microscopio antes de iniciar el proceso de ensamblaje. Es igualmente importante verificar el fenotipo de las poblaciones celulares aisladas al menos una vez con citometría de flujo. Especialmente cuando se utiliza un nuevo lote de colágeno-1 por primera vez, es ventajoso comprobar la velocidad de reticulación en una prueba antes de incrustar células en él. El ensamblaje requiere una gran manipulación manual, lo que aumenta el riesgo de infecciones. Para minimizar el riesgo de infecciones, trabaje en una campana de bioseguridad estéril con flujo de aire laminar, cambie los guantes con frecuencia y descontamine los guantes y el espacio de trabajo con etanol al 80%. Por razones similares, no utilice los soportes de abrazadera impresos en 3D más de una vez. Antes del proceso de inclusión en sí, es importante mantener todos los componentes del hidrogel (solución de reticulación, solución de colágeno-1) en hielo para evitar la reticulación prematura. En consecuencia, se debe trabajar rápidamente una vez que las células se agregan a la solución de reticulación para limitar la muerte celular debido al alto pH y la baja temperatura de la solución de reticulación. Para evitar la muerte celular relacionada con la desecación en el explante del núcleo, aspire el medio que cubre los explantes del núcleo sujetado inmediatamente antes de mezclar la solución de reticulación con la solución de colágeno-1. Para garantizar la colocación central del explante del núcleo dentro del hidrogel, lo ideal es fundir el hidrogel alrededor de un explante de núcleo sujetado que esté ligeramente tensado. Para ello, utilice el pasador de la clavija y el tornillo de perno M3 x 16 mm para fijar los soportes de la abrazadera a un juego de placas (impresas en 3D) con agujeros de las longitudes adecuadas. Después del tiempo de polimerización de 50 minutos, el explante del núcleo incrustado se puede destensar nuevamente dependiendo de las condiciones de cultivo deseadas. La cantidad de tensión que experimenta el ensamble durante el cultivo tiene un profundo impacto en los resultados experimentales y debe mantenerse uniforme en todas las muestras y condiciones21.
Sin embargo, el gran impacto de la (des)carga mecánica en los resultados experimentales es una de las principales ventajas del modelo ensamblado sobre la mayoría de las alternativas de ingeniería tisular, especialmente porque la composición matricial mantenida del explante central también debería recrear los complejos patrones de carga in vivo a nivel celular90. Si bien en la práctica, hasta ahora solo se ha demostrado la medición del módulo elástico lineal, la deformación máxima por tracción y la tensión máxima por tracción de los conjuntos, se han descrito protocolos para las mediciones de resistencia a la fatiga y relajación de la tensión para los explantes del núcleo del tendón en otros lugares y deberían ser aplicables a los conjuntos 91,92. Además de los patrones de carga similares a los de in vivo, la modularidad multinivel del ensamblaje es probablemente su mayor ventaja. Gracias a las cámaras de cultivo individuales, se puede establecer un conjunto controlable de condiciones de nicho para cada muestra por separado (es decir, temperatura, tensión de oxígeno, concentración de glucosa, suplementación, estimuladores, inhibidores y estiramiento estático con una placa). A continuación, la rigidez de la matriz y la composición de la matriz del compartimento extrínseco son personalizables a través de la composición del hidrogel y permitirían, por ejemplo, estudiar el impacto de un microambiente tisular cada vez más enfermo mediante la incorporación de más colágeno-3 y fibronectina celular 93,94,95. Las poblaciones celulares evaluadas en el compartimento extrínseco son fácilmente adaptables mediante la selección de qué células sembrar, pero también pueden modificarse en el explante del núcleo del tendón aprovechando las líneas celulares modificadas genéticamente establecidas y las líneas de ratón (es decir, el agotamiento de las células ScxLin)96. La diferente composición de la matriz y la célula de los dos compartimentos proporciona además una estructura 3D compartimentada única que es otro sello distintivo del tendón central 1,30,46.
Al utilizar este sistema, es importante tener en cuenta las consecuencias de la modularidad del sistema para la granularidad de los parámetros de resultado. Si bien la proliferación y el reclutamiento celular se pueden evaluar para cada compartimento por separado, las propiedades mecánicas, los componentes del secretoma y los productos de degradación actualmente solo se pueden medir para el conjunto completo. En cuanto al rendimiento, una persona debidamente capacitada puede preparar hasta 50 ensamblajes en un día de trabajo normal, siendo el principal cuello de botella el procedimiento de sujeción. Si bien algunos de los métodos de lectura son mutuamente excluyentes, es posible evaluar las propiedades mecánicas y los componentes del secretoma de forma repetitiva en la misma muestra, así como la composición de la población celular (citometría de flujo), el transcriptoma celular (RT-qPCR, secuenciación de ARN) o la distribución de la matriz y la célula (inmunocitoquímica/microscopía de fluorescencia) en los puntos finales. En publicaciones anteriores, estos métodos se han utilizado para caracterizar ampliamente las interacciones intercelulares y compartimentales en ensambles de núcleos // fibroblastos y núcleos // macrófagos expuestos a un nicho similar a una lesión84,85. En este trabajo, se ha explorado la capacidad del sistema modelo ensamblado para sondear la interacción intercompartimental entre las células endoteliales centrales y extrínsecas bajo diferentes estímulos microambientales.
La modularidad del sistema de modelos permite el refinamiento futuro del método, que es necesario para superar las siguientes limitaciones de la iteración de diseño actual. El análisis de citometría de flujo presentado en este trabajo y los datos de secuenciación de ARN unicelular publicados recientemente revelaron que los tenocitos residentes en el núcleo del tendón y las poblaciones derivadas del tendón de Aquiles son más heterogéneas de lo que se suponía anteriormente 24,34,59,84,97. Además, el comportamiento migratorio de las poblaciones celulares inicialmente residentes en el núcleo o en hidrogel difumina la compartimentación de los ensamblajes durante el cultivo. Ambos factores juntos hacen que sea difícil atribuir diferencias transcriptómicas a tipos celulares específicos y separar los procesos basados en la proliferación de los basados en la migración. Esta limitación podría superarse refinando la población de entrada con la clasificación celular activada por fluorescencia (FACS) basada en la composición celular de tendones sanos o enfermos caracterizados en estudios in vivo recientes, mejorando la lectura mediante la implementación de la secuenciación de ARN de una sola célula e integrando marcadores de proliferación como una tinción de EdU (5-etinil-2'-desoxiuridina) durante la microscopía.
Los ensamblajes aquí presentados también comparten una debilidad con la mayoría de los sistemas in vitro disponibles actualmente que simulan órganos enfermos desconectados del resto del cuerpo98,99. Sin embargo, la plataforma basada en cámaras de cultivo utilizada aquí posiciona bien el sistema modelo para su integración en una plataforma multiorgánica en la que se conectan ensamblajes que imitan diferentes órganos y se pueden estudiar las interacciones interorgánicas.
En esencia, el sistema modelo se basa en tendones posicionales de roedores, lo que da como resultado su propio conjunto único de inconvenientes. En primer lugar, la traducibilidad de los resultados se ve obstaculizada por el hecho de que los ratones de tipo salvaje no desarrollan ni sufren enfermedades tendinosas 8,100,101. La integración de tejidos y células de humanos o de cepas de ratón recientemente desarrolladas que exhiben aspectos de la enfermedad de los tendones podría aliviar este problema102. El cambio hacia un ensamblaje basado en humanos es particularmente interesante, ya que permitiría estudios con tejidos derivados de pacientes de tendones con diferentes enfermedades (es decir, tendinitis, tendinosis o peritendinitis) e incluso donantes resistentes al tratamiento que podrían desbloquear programas de tratamiento más personalizados. En segundo lugar, los explantes murinos del tendón de la cola no manejan particularmente bien el microdaño inducido por sobrecarga, lo que limita la aplicabilidad del sistema modelo para el estudio del daño agudo del tendón.
Por todas estas razones, los ensamblajes de explantes // hidrogeles se encuentran en una posición privilegiada para estudiar la biología del núcleo tendinoso, las interacciones estructura-función de la matriz y las interacciones intercompartimentales entre poblaciones celulares específicas en respuesta al microdaño inducido por nichos. Los conocimientos recopilados a partir de estos estudios de alto rendimiento podrían orientar la investigación in vivo y el desarrollo de tratamientos.
Divulgaciones
Los autores no tienen nada que revelar.
Agradecimientos
Este trabajo fue financiado por la Beca ETH 1-005733
Materiales
| Name | Company | Catalog Number | Comments |
| 0.4 mm x 25 mm injection needle (G27) | Sterican | 9186174 | |
| 3D printing filament: Clear polylactic acid prusament | Prusa | NA | |
| 4% formaldehyde | Roti-Histofix | P087.4 | |
| Accutase cell detachment solution | Sigma-Aldrich | A6964-100ML | |
| Amphotericin | VWR | L0009-100 | |
| Attachable digital C-mount camera: Moticam 2 | Motic | NA | |
| Bolt screw M3 x 16 mm, stainless steel | RS PRO | 1871235 | |
| Bolt screw M3 x 6 mm, stainless steel | RS PRO | 1871207 | |
| CaCl2 | Sigma-Aldrich | C5670 | |
| CD146 antibody: PE anti-mouse | BioLegend | 134703 | |
| CD206 antibody: Alexa Fluor 488 anti-mouse | BioLegend | 141709 | |
| CD31 antibody: Alexa Fluor 488 anti-mouse | BioLegend | 102413 | |
| CD86 antibody: PE anti-mouse | BioLegend | 105007 | |
| Collagenase I | Thermo Fisher Scientific | 17100017 | |
| Collagenase IV | Gibco | 17104-019 | |
| Dialyzed Fetal Bovine Serum (FBS) | Sigma-Aldrich | F0392-100ML | |
| Dimethyl sulfoxide (DMSO) | Sigma-Aldrich | 7000183 | |
| Dispase II | Sigma-Aldrich | D4693-1G | |
| DMEM/F12 | Sigma | 7002211 | |
| Dowel Pin, 3 mm x 16 mm, stainless steel | Accu | HDP-3-16-A1 | |
| Dragon Skin 10 Slow/1 silicone | KauPO | 09301-004-000001 | |
| Endopan 3 Kit | Pan-Biotech | P04-0010K | |
| Endothelial cell growth supplement | Lonza | CC-3162 | |
| Eppendorf safe-lock plastic tubes (1.5 mL) | Eppendorf | 30121023 | |
| Ethidium homodimer, EthD-1, 2 mM stock in DMSO | Sigma-Aldrich | 46043-1MG-F | |
| F4/80 antibody: Apc/fire 750 anti-mouse | BioLegend | 123151 | |
| Falcon plastic tube (15 mL) | Corning | 352096 | |
| Falcon plastic tube (50 mL) | Corning | 352070 | |
| Flow cytometer: LSR II Fortessa | BD Bioscience | 23-11617-02 | |
| Gelatin | Invitrogen | D12054 | |
| Hellmanex III alkaline cleaning concentrate | Sigma | Z805939-1EA | |
| Heparin | Sigma-Aldrich | H3149-10KU | |
| Hydroxyproline assay | Sigma-Aldrich | MAK008 | |
| Image analysis software: Motic Images Plus 3.0 ML | Motic | NA | |
| L-Ascorbic Acid Phosphate Magnesium Salt n-Hydrate | Wako Chemicals | 013-19641 | |
| LSE Low Speed Orbital Shaker | Corning | 6780-FP | |
| MEM non-essential amino acids | Sigma | 7002231 | |
| Mouse macrophage-stimulating factor (m-CSF) | PeproTech | 315-02-50ug | |
| MSD assay | Mesoscale Discovery | various | |
| NucBlue | Thermo Fisher Scientific | R37605 | |
| Nylon mesh strainer cap, 100 µm | Corning | 734-2761 | |
| Original Prusa i3 MK3S 3D printer | Prusa | i3 MK3S | |
| Penicillin-Streptomycin | Sigma-Aldrich | P4333 | |
| Phosphate-buffered saline (PBS), ph 7.4, sterile, 10 L | Gibco | 10010001 | |
| Puromycin | Gibco | A1113803 | |
| RBC lysis buffer | VWR | 786-650 | |
| recombinant m-CSF | PeproTech | 315-02 | |
| RNA extraction kit: Rneasy plus Micro | Qiagen | 74034 | |
| Slicing software: PrusaSlicer | Prusa | NA | Version 2.6.0 or higher |
| Sterile Cell Strainer 100 µm | Fisherbrand | 22363549 | |
| Surgical scalpel blade No. 21 | Swann-Morton | 307 | |
| Trizol reagent | Thermo Fisher Scientific | 15596018 | |
| Trypsin-EDTA (0.5 %) | Gibco | 15400054 |
Referencias
- Snedeker, J. G., Foolen, J. Tendon injury and repair - A perspective on the basic mechanisms of tendon disease and future clinical therapy. Acta Biomaterialia. 63, 18-36 (2017).
- Wang, J. H. C. Mechanobiology of tendon. Journal of Biomechanics. 39 (9), 1563-1582 (2006).
- Kirkendall, D. T., Garrett, W. E. Function and biomechanics of tendons. Scandinavian Journal of Medicine and Science in Sports. 7 (2), 62-66 (1997).
- Götmark, F., Cafaro, P., O'Sullivan, J. Aging human populations: Good for us, good for the earth. Trends in Ecology and Evolution. 33 (11), 851-862 (2018).
- Maffulli, N., Wong, J., Almekinders, L. C. Types and epidemiology of tendinopathy. Clinics in Sports Medicine. 22 (4), 675-692 (2003).
- Renström, P. A. F. H., Woo, S. L. -. Y. Tendinopathy: A major medical problem in sport. Tendinopathy in Athletes. , (2007).
- Screen, H. R. C., Birk, D. E., Kadler, K. E., Ramirez, F., Young, M. Tendon functional extracellular matrix. Journal of Orthopaedic Research. 33 (6), 793-799 (2016).
- Wunderli, S. L., Blache, U., Snedeker, J. G., Wunderli, S. L., Blache, U., Tendon, J. G. S. Tendon explant models for physiologically relevant in vitro study of tissue biology - a perspective. Connective Tissue Research. 61 (3-4), 262-277 (2020).
- Magnusson, S. P., Langberg, H., Kjaer, M. The pathogenesis of tendinopathy: balancing the response to loading. Nature Reviews Rheumatology. 6 (5), 262-268 (2010).
- Heinemeier, K. M., Schjerling, P., Øhlenschlæger, T. F., Eismark, C., Olsen, J., Kjær, M. Carbon-14 bomb pulse dating shows that tendinopathy is preceded by years of abnormally high collagen turnover. FASEB Journal. 32 (9), 4763-4775 (2018).
- Andersson, G., Backman, L. J., Scott, A., Lorentzon, R., Forsgren, S., Danielson, P. Substance P accelerates hypercellularity and angiogenesis in tendon tissue and enhances paratendinitis in response to Achilles tendon overuse in a tendinopathy model. British Journal of Sports Medicine. 45 (13), 1017-1022 (2011).
- Rolf, C. G., Fu, B. S. C., Pau, A., Wang, W., Chan, B. Increased cell proliferation and associated expression of PDGFRβ causing hypercellularity in patellar tendinosis. Rheumatology. 40 (3), 256-261 (2001).
- Riley, G. The pathogenesis of tendinopathy. A molecular perspective. Rheumatology. 43 (2), 131-142 (2004).
- Jarvinen, M., Jozsa, L., Kannus, P., Järvinen, T. L., Kvist, M., Leadbetter, W. Histopathological findings in chronic tendon disorders. Scandinavian Journal of Medicine & Science in Sports. 7 (2), 86-95 (1997).
- Soslowsky, L. J., et al. Overuse activity injures the supraspinatus tendon in an animal model: A histologic and biomechanical study. Journal of Shoulder and Elbow Surgery. 9 (2), 79-84 (2000).
- Tran, P. H. T., et al. Early development of tendinopathy in humans: Sequence of pathological changes in structure and tissue turnover signaling. FASEB Journal. 34 (1), 776-788 (2020).
- Theodossiou, S. K., Schiele, N. R. Models of tendon development and injury. BMC Biomedical Engineering. 1 (1), 1-24 (2019).
- Stauber, T., Blache, U., Snedeker, J. G. Tendon tissue microdamage and the limits of intrinsic repair. Matrix Biology. 85-86, 68-79 (2020).
- Wang, T., et al. In vitro loading models for tendon mechanobiology. Journal of Orthopaedic Research. 36 (2), 566-575 (2018).
- Fang, F., Sawhney, A. S., Lake, S. P. Different regions of bovine deep digital flexor tendon exhibit distinct elastic, but not viscous, mechanical properties under both compression and shear loading. Journal of Biomechanics. 47 (12), 2869-2877 (2014).
- Wunderli, S. L., et al. Tendon response to matrix unloading is determined by the patho-physiological niche. Matrix Biology. 89, 11-26 (2020).
- Wunderli, S. L., et al. Minimal mechanical load and tissue culture conditions preserve native cell phenotype and morphology in tendon - A novel ex vivo mouse explant model. Journal of Orthopaedic Research. 36 (5), 1383-1390 (2017).
- Arnoczky, S. P., Lavagnino, M., Egerbacher, M., Caballero, O., Gardner, K. Matrix metalloproteinase inhibitors prevent a decrease in the mechanical properties of stress-deprived tendons. The American Journal of Sports Medicine. 35 (5), 763-769 (2007).
- de Micheli, A. J., et al. Single-cell transcriptomic analysis identifies extensive heterogeneity in the cellular composition of mouse Achilles tendons. American Journal of Physiology - Cell Physiology. 319 (5), C885-C894 (2020).
- Arvind, V., Huang, A. H. Reparative and maladaptive inflammation in tendon healing. Frontiers in Bioengineering and Biotechnology. 9 (July), 1-16 (2021).
- Marsolais, D., Côté, C. H., Frenette, J. Neutrophils and macrophages accumulate sequentially following Achilles tendon injury. Journal of Orthopaedic Research. 19 (6), 1203-1209 (2001).
- Garcia-Melchor, E., et al. Novel self-amplificatory loop between T cells and tenocytes as a driver of chronicity in tendon disease. Annals of the Rheumatic Diseases. 80 (8), 1075-1085 (2021).
- Stolk, M., Klatte-Schulz, F., Schmock, A., Minkwitz, S., Wildemann, B., Seifert, M. New insights into tenocyte-immune cell interplay in an in vitro model of inflammation. Scientific Reports. 7 (1), 9801 (2017).
- Tempfer, H., Traweger, A. Tendon vasculature in health and disease. Frontiers in Physiology. 6, 330 (2015).
- Mienaltowski, M. J., Adams, S. M., Birk, D. E. Regional differences in stem cell/progenitor cell populations from the mouse achilles tendon. Tissue Engineering - Part A. 19 (1-2), 199-210 (2013).
- Sakabe, T., et al. Transcription factor scleraxis vitally contributes to progenitor lineage direction in wound healing of adult tendon in mice. Journal of Biological Chemistry. 293, (2018).
- Dyment, N. A., Hagiwara, Y., Matthews, B. G., Li, Y., Kalajzic, I., Rowe, D. W. Lineage tracing of resident tendon progenitor cells during growth and natural healing. PLoS One. 9 (4), e96113 (2014).
- Harvey, T., Flamenco, S., Fan, C. M. A Tppp3 + Pdgfra + tendon stem cell population contributes to regeneration and reveals a shared role for PDGF signalling in regeneration and fibrosis. Nature Cell Biology. 21 (12), 1490-1503 (2019).
- Zhang, J., et al. Characterization of the structure, vascularity, and stem/progenitor cell populations in porcine Achilles tendon (PAT). Cell and Tissue Research. 384 (2), 367-387 (2021).
- Tarafder, S., et al. Tendon stem/progenitor cells regulate inflammation in tendon healing via JNK and STAT3 signaling. FASEB Journal. 31 (9), 3991-3998 (2017).
- Lee, C. H., et al. Harnessing endogenous stem/progenitor cells for tendon regeneration Find the latest version Harnessing endogenous stem/progenitor cells for tendon regeneration. J Clin Invest. 125 (7), 2690-2701 (2015).
- Lui, P. P. Y., Chan, L. S., Cheuk, Y. C., Lee, Y. W., Chan, K. M. Expression of bone morphogenetic protein-2 in the chondrogenic and ossifying sites of calcific tendinopathy and traumatic tendon injury rat models. Journal of Orthopaedic Surgery and Research. 4, 27 (2009).
- Takeuchi, E., et al. Localization and expression of osteopontin in the rotator cuff tendons in patients with calcifying tendinitis. Virchows Archiv. 438 (6), 612-617 (2001).
- Kadler, K. E., Hill, A., Canty-Laird, E. G. Collagen fibrillogenesis: fibronectin, integrins, and minor collagens as organizers and nucleators. Current Opinion in Cell Biology. 20 (5), 495-501 (2008).
- Millar, N. L., et al. MicroRNA29a regulates IL-33-mediated tissue remodelling in tendon disease. Nature Communications. 6, 6774 (2015).
- Riley, G. P., Harrall, R. L., Constant, C. R., Chard, M. D., Cawston, T. E., Hazleman, B. L. Tendon degeneration and chronic shoulder pain: changes in the collagen composition of the human rotator cuff tendons in rotator cuff tendinitis. Annals of the Rheumatic Diseases. 53 (6), 359-366 (1994).
- Thorpe, C. T., Peffers, M. J., Simpson, D., Halliwell, E., Screen, H. R. C., Clegg, P. D. Anatomical heterogeneity of tendon: Fascicular and interfascicular tendon compartments have distinct proteomic composition. Scientific Reports. 6, 20455 (2016).
- Thorpe, C. T., et al. Distribution of proteins within different compartments of tendon varies according to tendon type. Journal of Anatomy. 229 (3), 450-458 (2016).
- Choi, H., et al. Heterogeneity of proteome dynamics between connective tissue phases of adult Tendon. eLife. 9, e55262 (2020).
- Spiesz, E. M., et al. Tendon extracellular matrix damage, degradation and inflammation in response to in vitro overload exercise. Journal of Orthopaedic Research. 33 (6), 889-897 (2015).
- Dyment, N. A., et al. The paratenon contributes to scleraxis-expressing cells during patellar tendon healing. PLoS One. 8 (3), e59944 (2013).
- Cadby, J. A., Buehler, E., Godbout, C., Van Weeren, P. R., Snedeker, J. G. Differences between the cell populations from the peritenon and the tendon core with regard to their potential implication in tendon repair. PLoS One. 9 (3), e92474 (2014).
- Huh, D., Hamilton, G. A., Ingber, D. E. From three-dimensional cell culture to organs-on-chips. Trends in Cell Biology. 21 (12), 745-754 (2011).
- Duval, K., et al. Modeling physiological events in 2D vs. 3D cell culture. Physiology. 32 (4), 266 (2017).
- Wang, J. H. C. Mechanobiology of tendon. Journal of Biomechanics. 39 (9), 1563-1582 (2006).
- Derwin, K. A., Soslowsky, L. J. A quantitative investigation of structure-function relationships in a tendon fascicle model. Journal of Biomechanical Engineering. 121 (6), 598-604 (1999).
- Herod, T. W., Veres, S. P. Development of overuse tendinopathy: A new descriptive model for the initiation of tendon damage during cyclic loading. Journal of Orthopaedic Research. 36 (1), 467-476 (2017).
- Andersen, M. B., Pingel, J., Kjær, M., Langberg, H. Interleukin-6: A growth factor stimulating collagen synthesis in human tendon. Journal of Applied Physiology. 110 (6), 1549-1554 (2011).
- Langberg, H., Rosendal, L., Kjær, M. Training-induced changes in peritendinous type I collagen turnover determined by microdialysis in humans. Journal of Physiology. 534 (1), 297-302 (2001).
- Zitnay, J. L., et al. Molecular level detection and localization of mechanical damage in collagen enabled by collagen hybridizing peptides. Nature Communications. 8, 14913 (2017).
- Hwang, J., et al. Molecular assessment of collagen denaturation in decellularized tissues using a collagen hybridizing peptide. Acta Biomaterialia. 53, 268-278 (2016).
- Lin, A. H., Zitnay, J. L., Li, Y., Yu, S. M., Weiss, J. A. Microplate assay for denatured collagen using collagen hybridizing peptides. Journal of Orthopaedic Research. 37 (2), 431-438 (2019).
- Blomgran, P., Blomgran, R., Ernerudh, J., Aspenberg, P. A possible link between loading, inflammation and healing: Immune cell populations during tendon healing in the rat. Scientific Reports. 6, 29824 (2016).
- Harvey, T., Flamenco, S., Fan, C. -. M. A Tppp3+Pdgfra+ tendon stem cell population contributes to regeneration and reveals a shared role for PDGF signalling in regeneration and fibrosis. Nature Cell Biology. 21 (12), 1490-1503 (2019).
- Pryce, B. A., Brent, A. E., Murchison, N. D., Tabin, C. J., Schweitzer, R. Generation of transgenic tendon reporters, ScxGFP and ScxAP, using regulatory elements of the scleraxis gene. Developmental Dynamics. 236 (6), 1677-1682 (2007).
- Godinho, M. S. C., Thorpe, C. T., Greenwald, S. E., Screen, H. R. C. Elastin is localised to the interfascicular matrix of energy storing tendons and becomes increasingly disorganised with ageing. Scientific Reports. 7 (1), 9713 (2017).
- Smith, M. M., et al. Modulation of aggrecan and ADAMTS expression in ovine tendinopathy induced by altered strain. Arthritis and Rheumatism. 58 (4), 1055-1066 (2008).
- Asundi, K. R., Rempel, D. M. MMP-1, IL-1β, and COX-2 mRNA expression is modulated by static load in rabbit flexor tendons. Annals of Biomedical Engineering. 36 (2), 237-243 (2008).
- Millar, N. L., et al. Inflammation is present in early human tendinopathy. American Journal of Sports Medicine. 38 (10), 2085-2091 (2010).
- Dakin, S. G., et al. Inflammation activation and resolution in human tendon disease. Science Translational Medicine. 7 (311), 311ra173 (2015).
- Jelinsky, S. A., Rodeo, S. A., Li, J., Gulotta, L. V., Archambault, J. M., Seeherman, H. J. Regulation of gene expression in human tendinopathy. BMC Musculoskeletal Disorders. 12, 86 (2011).
- Kannus, P., Józsa, L. Histopathological changes preceding spontaneous rupture of a tendon. A controlled study of 891 patients. The Journal of Bone and Joint Surgery. American. 73 (10), 1507-1525 (1991).
- Comin, J., et al. The prevalence and clinical significance of sonographic tendon abnormalities in asymptomatic ballet dancers: A 24-month longitudinal study. British Journal of Sports Medicine. 47 (2), 89-92 (2013).
- Schubert, T. E. O., Weidler, C., Lerch, K., Hofstädter, F., Straub, R. H. Achilles tendinosis is associated with sprouting of substance P positive nerve fibres. Annals of the Rheumatic Diseases. 64 (7), 1083-1086 (2005).
- Quigley, A. S., Bancelin, S., Deska-Gauthier, D., Légaré, F., Kreplak, L., Veres, S. P. In tendons, differing physiological requirements lead to functionally distinct nanostructures. Scientific Reports. 8 (1), 4409 (2018).
- Herod, T. W., Chambers, N. C., Veres, S. P. Collagen fibrils in functionally distinct tendons have differing structural responses to tendon rupture and fatigue loading. Acta Biomaterialia. 42, 296-307 (2016).
- Shepherd, J. H., Riley, G. P., Screen, H. R. C. Early stage fatigue damage occurs in bovine tendon fascicles in the absence of changes in mechanics at either the gross or micro-structural level. Journal of the Mechanical Behavior of Biomedical Materials. 38, 163-172 (2014).
- Birch, H. L. Tendon matrix composition and turnover in relation to functional requirements. International Journal of Experimental Pathology. 88 (4), 241-248 (2007).
- Kapałczyńska, M., et al. 2D and 3D cell cultures - a comparison of different types of cancer cell cultures. Archives of Medical Science. 14 (4), 910-919 (2018).
- Hussien, A. A., Niederoest, B., Bollhalder, M., Goedecke, N., Snedeker, J. G. The stiffness-sensitive transcriptome of human tendon stromal cells. Advanced Healthcare Materials. 12 (7), 2101216 (2023).
- Heo, S. J., et al. Aberrant chromatin reorganization in cells from diseased fibrous connective tissue in response to altered chemomechanical cues. Nature Biomedical Engineering. 7 (2), 177-191 (2023).
- Liu, C. -. F., Aschbacher-Smith, L., Barthelery, N. J., Dyment, N., Butler, D., Wylie, C. What we should know before using tissue engineering techniques to repair injured tendons: A developmental biology perspective. Tissue Engineering Part B: Reviews. 17 (3), 165-176 (2011).
- Ruiz-Alonso, S., Lafuente-Merchan, M., Ciriza, J., Saenz-del-Burgo, L., Pedraz, J. L. Tendon tissue engineering: Cells, growth factors, scaffolds and production techniques. Journal of Controlled Release. 333, 448-486 (2021).
- Rinoldi, C., et al. Tendon tissue engineering: Effects of mechanical and biochemical stimulation on stem cell alignment on cell-laden hydrogel yarns. Advanced Healthcare Materials. 8 (7), e1801218 (2019).
- Walia, B., Huang, A. H. Tendon stem progenitor cells: Understanding the biology to inform therapeutic strategies for tendon repair. Journal of Orthopaedic Research. 37 (6), 1270-1280 (2018).
- Thorpe, C. T., Riley, G. P., Birch, H. L., Clegg, P. D., Screen, H. R. C. Fascicles from energy-storing tendons show an age-specific response to cyclic fatigue loading. Journal of The Royal Society Interface. 11 (92), 20131058-20131058 (2014).
- Thorpe, C. T., Riley, G. P., Birch, H. L., Clegg, P. D., Screen, H. R. C. Fascicles and the interfascicular matrix show decreased fatigue life with ageing in energy storing tendons. Acta Biomaterialia. 56, 58-64 (2017).
- Youngstrom, D. W., Rajpar, I., Kaplan, D. L., Barrett, J. G. A bioreactor system for in vitro tendon differentiation and tendon tissue engineering. Journal of Orthopaedic Research. 33 (6), 911-918 (2015).
- Stauber, T., et al. Extrinsic macrophages protect while tendon progenitors degrade: Insights from a tissue engineered model of tendon compartmental crosstalk. Advanced Healthcare Materials. 10 (20), e2100741 (2021).
- Stauber, T., Moschini, G., Hussien, A. A., Jaeger, P. K., De Bock, K., Snedeker, J. G. IL-6 signaling exacerbates hallmarks of chronic tendon disease by stimulating progenitor proliferation & migration to damage. eLife. 12, RP87092 (2023).
- Doillon, C. J., Mantovani, D., Rajan, N., Habermehl, J., Cote, M. Preparation of ready-to-use storable and reconstituted type I collagen from rat tail tendon for tissue engineering applications. Nature Protocols. 1 (6), 2753-2758 (2007).
- Blache, U., et al. Inhibition of ERK 1/2 kinases prevents tendon matrix breakdown. Scientific Reports. 11 (1), 6838 (2021).
- Liu, X., et al. The role of vascular endothelial growth factor in tendon healing. Frontiers in Physiology. 12, 766080 (2021).
- Claxton, S., Kostourou, V., Jadeja, S., Chambon, P., Hodivala-dilke, K., Fruttiger, M. Efficient, inducible Cre-recombinase activation in vascular endothelium. Genesis. 46 (2), 74-80 (2008).
- Passini, F. S., et al. Shear-stress sensing by PIEZO1 regulates tendon stiffness in rodents and influences jumping performance in humans. Nature Biomedical Engineering. 5 (12), 1457-1471 (2021).
- Lee, A. H., Szczesny, S. E., Santare, M. H., Elliott, D. M. Investigating mechanisms of tendon damage by measuring multi-scale recovery following tensile loading. Acta Biomaterialia. 57, 363-372 (2017).
- Lee, A. H., Elliott, D. M. Multi-scale loading and damage mechanisms of plantaris and rat tail tendons. Journal of Orthopaedic Research. 37 (8), 1827-1837 (2019).
- Williams, I. F., Heaton, A., McCullagh, K. G. Cell morphology and collagen types in equine tendon scar. Research in Veterinary Science. 28 (3), 302-310 (1980).
- Maffulli, N., Ewen, S. W. B., Waterston, S. W., Reaper, J., Barrass, V. Tenocytes from ruptured and tendinopathic achilles tendons produce greater quantities of type III collagen than tenocytes from normal achilles tendons: An in vitro model of human tendon healing. American Journal of Sports Medicine. 28 (4), 499-505 (2000).
- Pajala, A., Melkko, J., Leppilahti, J., Ohtonen, P., Soini, Y., Risteli, J. Tenascin-C and type I and III collagen expression in total Achilles tendon rupture. An immunohistochemical study. Histology and Histopathology. 24 (10), 1207-1211 (2009).
- Best, K. T., et al. Scleraxis-lineage cell depletion improves tendon healing and disrupts adult tendon homeostasis. eLife. 10, e62203 (2021).
- Lehner, C., et al. Tenophages: A novel macrophage-like tendon cell population expressing CX3CL1 and CX3CR1. DMM Disease Models and Mechanisms. 12 (12), dmm041384 (2019).
- Ajalik, R. E., et al. Human organ-on-a-chip microphysiological systems to model musculoskeletal pathologies and accelerate therapeutic discovery. Frontiers in Bioengineering and Biotechnology. 10, 846230 (2022).
- Ingber, D. E. Human organs-on-chips for disease modelling, drug development and personalized medicine. Nature Reviews Genetics. 23 (8), 467-491 (2022).
- Brehm, M. A., Shultz, L. D., Luban, J., Greiner, D. L. Overcoming current limitations in humanized mouse research. The Journal of Infectious Diseases. 208 Suppl (Suppl 2), 125-130 (2013).
- Nunan, R., Harding, K. G., Martin, P. Clinical challenges of chronic wounds: searching for an optimal animal model to recapitulate their complexity. Disease Models & Mechanisms. 7 (11), 1205-1213 (2014).
- Costa-Almeida, R., Calejo, I., Reis, R. L., Gomes, M. E. Crosstalk between adipose stem cells and tendon cells reveals a temporal regulation of tenogenesis by matrix deposition and remodeling. Journal of Cellular Physiology. 233 (7), 5383-5395 (2018).
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoExplorar más artículos
This article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados