
Un abonnement à JoVE est nécessaire pour voir ce contenu. Connectez-vous ou commencez votre essai gratuit.
Method Article
L'utilisation du test Ramsay mesurer la sécrétion de liquide et de flux d'ions dans le Tarifs
Dans cet article
Résumé
Ce protocole décrit l'utilisation de l'essai Ramsay pour mesurer les taux de sécrétion de liquide de isolés Malpighian (rénale) tubules de Drosophila melanogaster. En outre, l'utilisation d'électrodes spécifiques des ions pour mesurer les concentrations de sodium et de potassium dans le fluide sécrété, permettant le calcul de flux transépithélial d'ions, est décrit.
Résumé
Modulation de transport d'ions rénale épithéliale organismes permet de maintenir l'homéostasie ionique et osmotique dans la face de la variation des conditions extérieures. Le Drosophila melanogaster Malpighian (rénale) tubule offre une occasion unique d'étudier les mécanismes moléculaires de transport d'ions épithéliale, en raison de la génétique puissants de cet organisme et de l'accessibilité de ses tubules rénaux à l'étude physiologique. Ici, nous décrivons l'utilisation de l'essai Ramsay pour mesurer les taux de sécrétion de liquide de mouche isolé tubules rénaux, avec l'utilisation d'électrodes spécifiques des ions pour mesurer les concentrations de sodium et de potassium dans le liquide sécrété. Ce test permet d'étudier des liquides ioniques et transépithélial flux de ~ 20 tubes à la fois, sans la nécessité de transférer le fluide sécrété à un appareil séparé pour mesurer les concentrations d'ions. Génétiquement tubules distinctes peuvent être analysées pour évaluer le rôle des gènes spécifiques dans les processus de transport. En outre, le bsaline athing peut être modifié pour examiner les effets de ses caractéristiques chimiques, ou des médicaments ou des hormones ajoutées. En résumé, cette technique permet la caractérisation moléculaire des mécanismes de base de transport epithelial d'ions dans le tubule Drosophila, ainsi que la régulation de ces mécanismes de transport.
Introduction
Rénale transport ionique épithéliale sous-tend organismal ionosphère et osmorégulation. Le Drosophila melanogaster Malpighian (rénale) tubule offre une occasion unique d'étudier les mécanismes moléculaires de transport d'ions épithéliale. Cela est dû à la combinaison de la génétique de la drosophile puissants, associé à l'accessibilité de ses tubules rénaux à l'étude physiologique. Le dosage Ramsay, nommé d'après le chercheur pionnier de la technique 1, mesure les taux de sécrétion de fluides à partir de tubes de Malpighi isolées, et a été établie chez la drosophile en 1994 par Dow et ses collègues 2. Cela a ouvert la voie à d'autres études utilisant la drosophile outils génétiques, telles que le système GAL4 UAS 3,4, pour définir les voies de signalisation cellulaires spécifiques régissant la sécrétion de fluide. Un exemple comprend la signalisation du calcium en réponse à une hormone peptidique 5, parmi beaucoup d'autres 6,7.
ve_content "> Une combinaison de techniques génétiques et étude physiologique classique a montré que la production d'urine chez la mouche se produit par la sécrétion d'un chlorure de potassium riche en fluide à partir du segment principal du tube. Ceci se produit par l'intermédiaire du transépithélial sécrétion parallèle de cations, principalement K + mais aussi Na +, à travers la cellule principale, et Cl -. sécrétion à travers la cellule stellaire 8-12 La capacité de mesurer séparément transépithéliaux flux K + et Na + permet une caractérisation plus détaillée des mécanismes de transport que la mesure de la sécrétion de fluide seul. Par exemple, dans les tubules Drosophila non stimulées, la Na + / K + -ATPase ouabaïne n'a aucun effet sur la sécrétion de fluide 2, même lorsque son absorption dans les cellules principales est inhibée par l'inhibiteur de transport anionique organique taurocholate 13. Cependant, Linton et O'Donnell a montré que l'ouabaïne dépolarisele potentiel de la membrane basolatérale, Na + et augmente le flux 9. Comme le montrent les résultats représentatifs, nous avons reproduit ces résultats, et montré que flux de K + est concomitante diminué de 14; l'augmentation du flux de Na + et K + a diminué flux des effets opposés sur la sécrétion de fluide, de sorte qu'aucun changement net de la sécrétion. Ainsi, il ya deux résolutions au «paradoxe ouabaïne», à savoir l'observation initiale que ouabaïne n'a aucun effet sur la sécrétion de fluide dans le tubule Drosophila:. La première, dans les tubules stimulés, l'effet de la ouabaïne sur la sécrétion de fluide est pas évident en raison de son absorption par le système de transport anionique organique 13; et la seconde, dans les tubules non stimulées, ouabaïne a des effets sur transepithelial Na + et K + flux opposés, entraînant aucun changement net de la sécrétion de fluide (voir résultats représentatifs et réf. 9). Par conséquent, le rôle principal de la Na + / K + ATPase dans les tubules non stimulées est de réduire la concentration intracellulaire de Na + à générer un gradient de concentration favorable pour Na + -coupled processus de transport à travers la membrane basolatérale. En effet, en mesurant séparément Na + et K + flux, nous avons démontré que les tubules manque la volée sodium-potassium-2-chlorure cotransporteur (NKCC) ont diminué K transepithelial + flux, sans diminution supplémentaire après addition ouabaïne, et aucun changement dans transepithelial Na + flux 14. Ces résultats ont appuyé notre conclusion que Na + entrant dans la cellule par l'intermédiaire du NKCC est recyclée à travers la Na + / K + -ATPase. Dans un autre exemple, Ianowski et al. Observe que l'abaissement de bain concentration de K + de 10 à 6 mm diminué transepithelial flux de K + et une augmentation transepithelial Na + flux dans les tubules de Rhodnius prolixus, avec aucun changement net de la sécrétion de fluide 15. Les effets différentiels sur Na + et K + flux flux à travers les tubules larvaires ont également été observés dans les tubules de drosophile en réponse à divers régimes de sel 16 et dans deux espèces de moustiques en réponse à l'élevage de la salinité 17.La plus grande difficulté dans la mesure de flux transépithélial d'ions dans la préparation d'essai Ramsay est la détermination des concentrations des ions dans le fluide sécrété. Ce défi a été relevé avec différentes solutions, y compris la flamme photometery 18, l'utilisation d'ions radioactifs 19 et sonde électronique longueur d'onde de la spectroscopie à dispersion 20. Ces techniques nécessitent le transfert de la goutte de fluide sécrété à un instrument de mesure des concentrations d'ions. Etant donné que le volume de fluide sécrété par les tubules Drosophila non stimulé est petite, typiquement environ 0,5 nl / min, ce qui pose un défi technique et introduit également erreur si une partie du fluide est sécrétéperdu au moment du transfert. En revanche, l'utilisation d'électrodes spécifiques des ions permet la mesure de l'activité ionique (à partir de laquelle la concentration en ions peut être calculée) in situ. Le protocole actuel a été adaptée à partir de celle utilisée par les Maddrell et collègues pour mesurer transépithélial flux de K + à travers le tube Rhodnius utilisant la valinomycine comme K + ionophore 21, et décrit également l'utilisation d'un -butylcalix 4- tert [4] d'acides arène-tétraacétique Na tétraéthyle à base d'ester + électrode spécifique-ion spécifique de caractérisé par Messerli et. . 22 al. Électrodes ions spécifiques ont également été utilisés pour mesurer les concentrations d'ions dans le liquide sécrété par les tubules de Malpighi dans le dosage Ramsay chez l'adulte et larvaire 9,23 16 Drosophila melanogaster, la Nouvelle-Zélande Alpine Weta (Hemideina maori) 24 et 17 chez les moustiques.
Ici, nous décrivons en détail l'utilisation de la Ramsay commedire pour mesurer les taux de sécrétion de fluide dans les tubes de Malpighi de Drosophila melanogaster, ainsi que l'utilisation d'électrodes spécifiques des ions afin de déterminer les concentrations de K + et Na + dans le fluide sécrété, et donc le calcul des flux transépithélial d'ions. Une vue d'ensemble de l'essai est fourni à la figure 1.

Figure 1. Schéma de l'Malpighian Pailles et le test Ramsay avec utilisation de l'ion-spécifiques électrodes pour mesurer les concentrations d'ions. Cette figure illustre la configuration pour le dosage Ramsay. (A) Pour chaque volée comporte quatre tubes, une paire de tubules antérieure et une paire de tubules postérieures, qui flottent dans la cavité abdominale entouré d'hémolymphe. Dans chaque paire, les deux tubes se rejoignent à l'uretère, qui se jette ensuite l'urine à la jonction de l'intestin moyen et hindgut. Les tubules sont aveugles-clos. L'urine est généré par le segment principal de fluide sécrétant (en rouge), et coule vers l'uretère et sortir dans l'intestin. Après dissection, la paire de tubule est dissocié à partir de l'intestin par transection l'uretère. (B) La paire de tubes est ensuite transféré dans une gouttelette de solution saline dans un bain bien du plat de dosage. L'un des deux tubes, appelé ici le "tube d'ancrage», est enroulé autour d'une tige de métal et est inerte. L'autre tube est le tube de sécrétion. Le segment initial (qui ne sécrètent pas fluide) et le segment principal de la tubule sécrétant restent dans la gouttelette de solution saline de baignade. Ions et mouvement de l'eau de la solution saline de bain et dans la lumière des tubules du segment principal, puis se déplacent vers l'uretère, comme cela se produirait in vivo. Le segment inférieur (bleu) est en dehors de la solution saline de baignade et donc inerte. Depuis l'uretère est coupé, le fluide sécrété apparaît comme une gouttelette à partir de la fin de l'uretère de coupe. Til agrandit gouttelette de fluide sécrété dans le temps comme la sécrétion continue, et son diamètre est mesuré en utilisant un micromètre oculaire. Une couche d'huile minérale empêche l'évaporation du fluide sécrété. Les électrodes spécifiques référence et d'ions de mesurer la concentration en ions du fluide sécrété. S'il vous plaît cliquer ici pour voir une version plus grande de cette figure.
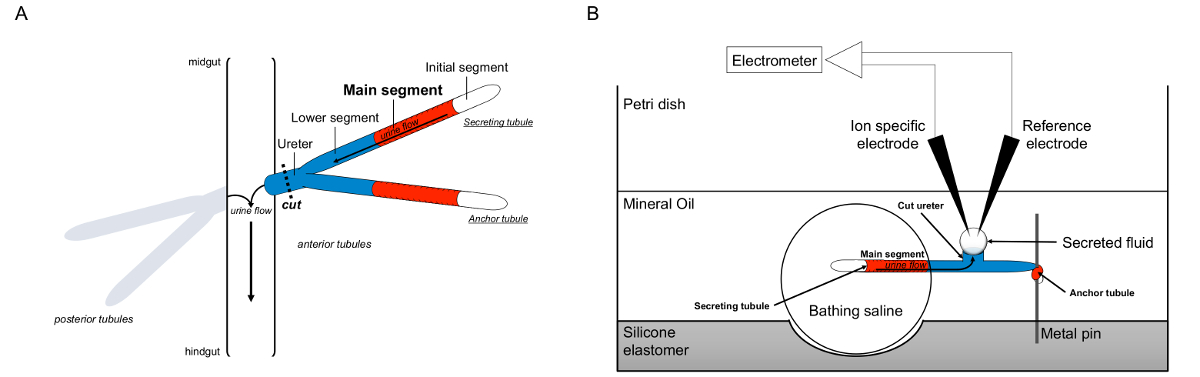{kind=link}
Protocole
1. Préparation de la dissection, le calibrage et le dosage Plats
Remarque: Dans cette étape, trois boîtes de Petri en plastique doublés d'élastomère de silicone sont préparés: l'un pour la dissection, l'un pour la réalisation du test Ramsay ("plat de test"), et l'autre pour effectuer l'étalonnage. Ces plats sont réutilisés d'une expérience à, et donc cette étape ne doit être répété si un plat pauses. Une image de la boîte de test est illustré à la figure 2.

Figure 2. La parabolique de dosage. Le plat utilisé pour le dosage Ramsay est montré ici. Il est une boîte de Pétri de 10 cm qui est bordée de silicone élastomère. Entre 20 et 25 puits sont creusés dans l'élastomère. Une tige de métal minuties, coupé en deux, est placé à la droite de chaque puits (ou vers la gauche, si l'expérimentateur est gaucher).tps: //www.jove.com/files/ftp_upload/53144/53144fig2large.jpg "target =" _ blank "> S'il vous plaît cliquer ici pour voir une version plus grande de cette figure.
- Utiliser des gants, verser ~ 80 g de silicone base élastomère dans un bécher de verre. Ajouter 1/10 du poids (8 g) élastomère de silicone guérir. Remuer avec un agitateur métallique. Placer sur un agitateur orbital avec un sommet plat à une vitesse douce, par exemple 100 tours par minute, pendant plusieurs heures jusqu'à ce que toutes les bulles ont été enlevés.
- Verser l'élastomère de silicone dans des boîtes de Petri en plastique propre: 100 mm x 15 plats mm pour les plats de dissection et d'analyse, et 35 x 10 mm pour le plat de calibrage. L'épaisseur de la couche d'élastomère doit être ~ 6-7 mm pour les plats de 100 mm, et à 5 mm pour le plat de 35 mm.
- Placez les plats sur le banc à la température ambiante (RT) pour guérir (durcir), ~ 24 - 48 h.
- Une fois le plat de dosage a durci, préparer un lot plus petit d'élastomère de silicone comme dans l'étape 1.1, par exemple., 10 g de silicone base élastomère avec 1 g remède. Agiter orbitaleshaker comme dans l'étape 1.1 jusqu'à ce que des bulles d'air ne sont plus présents. Cet élastomère est utilisé dans l'étape 1.5.4 et ne doit pas être autorisé à durcir avant cette étape.
- Avec un scalpel chirurgical et précautions d'objets tranchants standard, faire des puits dans l'une des boîtes de Pétri recouvertes d'élastomère de silicone 100 mm. Ce sera le plat de dosage.
- Ajouter puits ~ 1 cm et d'un diamètre de ~ 3 - 4 mm. 25 puits peuvent facilement se glisser dans un plat de dosage de 100 mm. Wells doit être d'au moins 6 mm éloignées des parois de la boîte. La figure 2 illustre l'espacement des puits. Chaque puits contient une tubule de fluide sécrétant pendant l'expérience. Ainsi, une boîte de 25 puits contenant 25 tubules permettra à analyser.
- Utilisez un marqueur permanent pour marquer la position des puits si nécessaire.
- Faire les parois du puits aussi lisse que possible en plaçant le scalpel dans l'élastomère, puis en utilisant la main opposée pour faire tourner le plat à 360 °.
- Puis, en utilisant la norme sha RPS précautions, trempez une aiguille 30 G dans l'élastomère non durci de l'étape 1.4 et placez une petite goutte au fond de chaque puits. Ceci permet de fluidifier le fond du puits. Laisser durcir (durcir) x 24 - 48 h.
- Préparer les épingles minuties.
- Placez 0,15 mm anodisé noir épingles minuties dans une rangée sur un morceau de TP type étiquetage bande de 1 pouce. Grand axe Les broches de devrait être orthogonale à l'axe de long de la bande. Couper la bande sur toute sa longueur afin de sectionner chaque broche en deux moitiés à peu près égales (Figure 3). Utilisez une demi-broches pour chaque puits.

Figure 3. Couper les Pins minuties. Les broches sont alignés sur un morceau de ruban d'étiquetage en parallèle. Ensuite, les ciseaux sont utilisés pour couper les tiges de moitié.ge.jpg "target =" _ blank "> S'il vous plaît cliquer ici pour voir une version plus grande de cette figure.
- Insérez chaque demi-tige dans l'élastomère de silicone d'environ 1 mm à la droite de chaque puits (si droitier, si gaucher, insérer la tige à la gauche de chaque puits). Ceci est plus facile à faire lors de la visualisation des puits à basse puissance sous un stéréomicroscope à dissection et est aidé par l'utilisation d'une pince franches. La figure 2 illustre le positionnement des broches.
2. Préparation Beaux Rods en verre
Remarque: Dans cette étape, une tige de verre est préparé qui sera utilisé pour transférer les tubules du plat de dissection dans la goutte de baignade. La tige de verre est réutilisé d'une expérience à, cette étape est effectuée une seule fois, sauf si les pauses de tige et un nouveau est nécessaire.
- Obtenir des feuilles de 3 mm (1/8 po) d'épaisseur de verre noir teinté dans un magasin de passe-temps et de verre appropriéemassicot équipements, comme un coupe-verre et des pinces. Utiliser un équipement de sécurité approprié (gants, lunettes épaisses).
- Couper le verre en bandes ~ 6 mm de large x 10 cm de long
- Tenir une bande de verre dans chaque main. Adoucir l'extrémité courte de chaque bande sur la flamme d'un brûleur Bunsen, en utilisant des mesures de sécurité appropriées. Ensuite, appuyez sur les extrémités des deux bandes ensemble et tirer dehors dans un mouvement lisse pour créer une fine tige de verre avec une poignée.
3. Physiologie Setup
Remarque: Dans cette étape, le microscope, le circuit électromètre et électrique est mis en place. Autre que re-chloration périodique (étape 3.2) des fils d'argent et ré-étalonnage de l'électromètre (étape 3.8), cette étape est réalisée une seule fois. La figure 4 illustre la configuration.

Figure 4. Physiologie Setup. La configuration de la physiologie est photographié ici. (A) Vue d'ensemble de la configuration. Le stéréomicroscope est placé à l'intérieur de la cage de Faraday avec les micromanipulateurs de chaque côté. A la lumière de la fibre optique est enfilée à travers un trou dans le côté de la cage de Faraday. L'électromètre est placé en dehors de la cage de Faraday. (B) au chlorure enrober les fils d'argent, le fil est plongé dans l'eau de Javel. (C) Close-up de la configuration. Le titulaire de microélectrodes droite, montré dans cette image sur la gauche, est vissé sur la sonde de l'électromètre. L'électrode à spécificité ionique est ensuite enfilé sur le fil d'argent dans le porte-électrode. Sur la droite, l'électrode de référence est enfilé sur le fil d'argent du titulaire de microélectrodes 45 °. Le circuit doit ensuite être mise à la terre appropriée. Le plat de dosage est représenté comme il sera positionné lors de l'exécution des mesures. S'il vous plaîtcliquez ici pour voir une version plus grande de cette figure.
{kind=link}
- Placez le stereomicrosope avec micromètre oculaire à l'intérieur de la cage de Faraday. Terrain à l'intérieur de la cage de Faraday, qui est ensuite mis à la terre à la masse du châssis de l'électromètre. Placez les micromanipulateurs part et d'autre du microscope (Figure 4A).
- Chlorure de deux fils d'argent par immersion dans l'eau de Javel pendant au moins 1 h. Elargir la nuit (O / N) si nécessaire (figure 4B). Répétez cette étape chaque fois que les fils d'argent doivent être re-chloré, par exemple si elles sont gris en apparence plutôt que noir.
- Enfiler un fil d'argent chlorée dans chacun des détenteurs de microélectrodes.
- Établir le circuit électrique avec mise à la terre appropriée. Par exemple, placer le, titulaire de microélectrodes droite ventilé, qui tiendra l'électrode spécifique-ion (ISE), sur la sonde de l'électromètre, qui est fixé sur le micromanipulateur (figure 4C).
- Fixez le, 45 ° Porte de microélectrodes ventilé, qui tiendra l'électrode de référence, sur l'autre micromanipulateur (figure 4C). Puis la terre à la terre de circuit sur l'électromètre.
- Relier le «AB sur" sortie BNC de l'électromètre à la masse du châssis de l'électromètre.
- Placer la source de lumière à fibres optiques à l'extérieur de la cage de Faraday, avec le tube à col de cygne enfilée à travers un trou dans la cage de Faraday (figure 4A).
- Mettre en place et calibrer l'électromètre selon les instructions du fabricant. Re-calibrer l'électromètre sur une base régulière (tous les 1 - 2 semaines). Une fois terminé, et entre les mesures, laisser l'électromètre en mode "veille" avec la position bascule réglé sur "IN", entrée de compteur réglé sur "A" et la gamme est réglée sur "200 mV."
4. Préparer les solutions à disséquer et à Bath
- Préparer Drosophila solution saline comme détaillé dans le tableau 1. Pour une utilisation dans des expériences, versez 40 ml dans ~ un tube conique de 50 ml et garder à température ambiante. Jeter si il existe des preuves de la croissance bactérienne ou fongique.
- Pour préparer le milieu de bain standard (SBM), mélanger la drosophile saline à 1: 1 avec du milieu de Schneider, et de passer à travers un filtre à seringue de 0,22 um. Préparer en petites portions (~ 10 - 15 ml), conserver à 4 ° C et jeter si il existe des preuves de la croissance bactérienne ou fongique. Les composants du milieu de Schneider sont répertoriés dans le tableau 2.
5. Faire l'électrode Ion spécifique: silanisant Pipettes
Remarque: Dans cette étape, le dichlorodiméthylsilane est utilisé pour "silaner" légèrement l'électrode à spécificité ionique. Cela ajoute une couche hydrophobe à l'intérieur de l'électrode qui lui permet de conserver l'ionophore hydrophobe. Silanisation excessive est évitée pour empêcher l'absorption de l'huile minérale au moment de measurements en gouttes sous l'huile. Électrodes Silanized sont bonnes pour plusieurs semaines. Par conséquent, cette étape est réalisée toutes les quelques semaines.
- Flame-polonais les extrémités de 5 - 6 unfilamented tubes capillaires en verre borosilicate (diamètre extérieur 1,2 mm, diamètre intérieur de 0,69 mm, longueur 10 cm) à feu doux, en utilisant des mesures de sécurité appropriées.
- Placer les tubes capillaires dans le fond d'un bêcher de 1 litre en verre.
- Dans un capot et un équipement de protection individuelle approprié, verser de l'acide nitrique à 70% (ATTENTION: inflammable et corrosif, voir la Fiche de Données de Sécurité pour le stockage sûr et de manutention) sur les tubes capillaires et laisser tremper pendant 5 min.
- Verser l'acide nitrique de nouveau dans une bouteille en verre. Re-utilisation pour le lavage subséquent de l'acide nitrique.
- Ajouter 200 ml de ~ H 2 O désionisée au bêcher. Vider les déchets dans une bouteille de verre dédié pour les déchets de l'acide nitrique. Répéter l'opération avec un 200 ml supplémentaires de H 2 O. déminéralisée Suivez les directives institutionnelles pour ee élimination sûre des déchets acides.
- Faire trois lavages supplémentaires avec de grands volumes d'eau déminéralisée. Vider dans l'évier.
- Dans la hotte, placer les tubes capillaires sur une plaque chaude avec l'ensemble plateau en céramique à 200 ° C et à sec pour un minimum de 20 min, 1 h de manière optimale. Il peut également être laissé pendant des périodes de temps plus longues.
- Sur un extracteur de pipette (figure 5A), tirez pipettes à un diamètre de ~ 1 pointe - 2 pm.
- Placez pipettes ramené sur plaque de cuisson, en prenant soin de ne pas briser conseils, pendant au moins 10 min, 30 min de façon optimale, mais peut être laissé plus longtemps.
- Ajouter 20 pi de dichlorodiméthylsilane (ATTENTION: inflammable, corrosif, toxicité aiguë, voir la Fiche de Données de Sécurité pour le stockage sûr et de manutention) dans une boîte de Pétri en verre de 15 cm et inverser plat sur les pipettes sur la plaque chauffante (figure 5B). Laisser en place pendant au moins 20 min, 2 h de manière optimale. La même boîte de Pétri peut être ré-utilisé dans les expériences suivantes.
- Déterminer l'amou nt de dichlorodiméthylsilane ajouté par essais et erreurs. Après ionophore est ajouté (étape 8.4), veiller à ce que l'interface entre l'ionophore et la solution de remplissage est plat (Figure 5C). Si l'interface est concave, ce qui indique sur-silanisation, et moins de silane doit être utilisé. Si l'interface est convexe, ce qui indique moins de silanisation, en plus du silane doit être utilisé.
Remarque: Le dichlorodiméthylsilane tendance à aller "off" au fil du temps, à savoir, silanisation moins efficace est réalisé avec le même volume de silane.. À ce stade, soit nouvelle silane peut être commandé, ou le montant ajusté pour obtenir silanisation équivalent.
- Déterminer l'amou nt de dichlorodiméthylsilane ajouté par essais et erreurs. Après ionophore est ajouté (étape 8.4), veiller à ce que l'interface entre l'ionophore et la solution de remplissage est plat (Figure 5C). Si l'interface est concave, ce qui indique sur-silanisation, et moins de silane doit être utilisé. Si l'interface est convexe, ce qui indique moins de silanisation, en plus du silane doit être utilisé.
- Éteignez la plaque chaude et laisser refroidir. Retirer le plat de Pétri de verre et de transférer des pipettes à pot de stockage contenant du gel de silice, ce qui maintient la dessiccation. Manipuler avec soin pipettes (forceps) utiles pour prévenir la rupture pointe.
5 "src =" / files / ftp_upload / 53144 / 53144fig5.jpg "/>
Figure 5. Pipettes silanisant. (Liste a) Sous Exemple de pipette extracteur. (B) Image des pipettes tiré sur la plaque chauffante. Le plat de verre contenant une goutte de dichlorodiméthylsilane a été inversée au cours des pipettes tirés. (C) Schéma illustrant l'interface entre le cocktail d'ionophore et la solution de remplissage. Une interface plane indique silanisation optimale. S'il vous plaît cliquez ici pour voir une version plus grande de cette figure.
{kind=link}
6. Préparer le Dispositif d'aspiration négative
Remarque: Dans cette étape, un dispositif d'aspiration négative simple est préparé (figure 6) qui sera utilisée pour remplir l'électrode spécifique-ion. Cette étape est effectuée une seule fois.
- Fixez une seringue de 3 ml à un robinet 3 voies avec luer lock. Àl'extrémité opposée du robinet, qui contient le collier rotatif et garde, visser dans un connecteur de Luer femelle de verrouillage avec l'extrémité de fer barbelé. Ensuite, attacher un tube de silicone, 1/16 de pouce de diamètre interne de diamètre extérieur 8/1 pouce, à l'extrémité cannelée de l'embout. Ensuite, insérer le tuyau en plastique avec 1/32 pouce de diamètre intérieur et diamètre extérieur 3/32 pouces

Figure 6. L'aspiration de l'appareil négative. Image des composants du dispositif d'aspiration négative (3 ml seringue avec luer lock, robinet 3 voies avec collier rotatif et le garde, luer connecteur de verrouillage femelle fin barbelés, tube en silicone, tube en plastique) et le produit final. S'il vous plaît cliquez ici pour voir une version plus grande de cette figure.
{kind=link}
7. Les mouches percevoir pour Dissection < / p>
- Utiliser des techniques d'élevage de mouches standard 25 de mettre en place des croix de mouches et de modifier au besoin. Par exemple, la température d'utilisation (telle que 28 ° C) a augmenté dans des expériences où l'activité a augmenté de GAL4 est souhaitée.
Remarque: Il est important que les mouches ne sont pas élevés dans des conditions trop de monde; le nombre de parents masculins et féminins devraient être réduites dans ce cas. Si différents génotypes sont utilisés, le nombre de parents mâles et femelles doivent être ajustées pour obtenir le numéro approximativement similaire de la progéniture. - Recueillir des mouches pour la dissection (étape 11) en utilisant des techniques d'élevage de mouches standard de 25 dans 1 - 2 jours de l'éclosion. Tubules de mouches femelles sont plus facilement disséqués, mais tubules de mouches mâles peuvent également être utilisés si nécessaire ou souhaité. Lieu vole dans un flacon contenant de la nourriture à la mouche. Placez les flacons à la température souhaitée pendant 3 - 5 jours avant la dissection.
8. Remplissage de l'électrode Ion spécifique (ISE)
e_content "> Remarque:.. Dans cette étape, l'ISE est remblayé avec une solution de sel, puis ionophore est introduit dans la pointe Le ISE peut être réutilisé de jour en jour aussi longtemps que cela fonctionne bien donc, cette étape est effectuée à quelques jours si nécessaire.- Pour faire une électrode spécifique de K +, remblayer une pipette silanisé avec 0,5 M de KCl utilisant une seringue et microfilament 1 ml (réutilisation microfilaments d'une expérience à). Assurez-vous que la solution de remplissage remplit à la pointe de la pipette - pas d'air dans la pointe. En cas de doute, de visualiser sous un microscope composé. Déloger les bulles d'air en tapotant délicatement la pipette.
- Pour une électrode spécifique de Na +, remblayer avec NaCl 150 mM.
- Insérez l'extrémité arrière de l'ISE dans le tube en plastique de l'appareil d'aspiration négative préparée à l'étape 6. Dans la hotte, placer l'ISE sur un plastique de 3,5 cm boîte de Pétri inversé avec un morceau de pâte à modeler pour fixer en place.
- Générer p négativeression à l'aide du dispositif d'aspiration. Tirer sur la seringue avec le "off" de la poignée de robinet d'arrêt dirigée vers l'orifice latéral. La quantité de reculer varie, mais est généralement dans la gamme de 0,6 - 0,7 ml. Puis tournez la poignée du robinet de sorte que le "off" est dirigée vers le tube.
- Dans la hotte et l'utilisation de l'équipement de protection individuelle approprié, tremper un 1 - ul pourboire de 10 pipette dans la solution de ionophore (ATTENTION: toxique Voir Fiche de Données de Sécurité pour le stockage sûr et instructions de manipulation.). Expulser une petite goutte en plaçant un doigt ganté sur la plus grande ouverture de la pointe de la pipette. Ensuite, toucher la goutte de ionophore à la pointe de l'ISE, sans toucher la pointe ISE avec la pointe de la pipette pour éviter de casser la pointe ISE.
- Utilisez le ionophore de potassium figurant dans le tableau des matériaux "comme est." Pour préparer l'ionophore de sodium, faire une solution de (en% w / w) 10% 4-tert -butylcalix [4] tétraéthylester d'acide arène-tétra-acétique, 89Éther nitrophényl octyl 0,75%, et 0,25% du tétraphénylborate de sodium (ATTENTION:. Toxique Voir Fiche de Données de Sécurité pour le stockage sûr et de manutention). Conserver dans un flacon de verre enveloppés dans une feuille pour protéger de la lumière.
- Examinez l'ISE sous un grossissement de 40X en utilisant un microscope composé pour déterminer si l'ionophore a été "repris" dans la pointe ISE et si l'interface de solution ionophore / de remblai est plat (voir l'étape 5.10.1 et la figure 5C).
- Si aucun ionophore a été repris, augmenter la quantité de pression négative générée par le dispositif d'aspiration. Si cela ne réussit pas, l'électrode peut avoir été insuffisamment silanisée. Répétez l'étape 5 en utilisant une plus grande quantité de dichlorodiméthylsilane.
- Placez l'ISE, pointe vers le bas, sur le mur d'un bécher partiellement remplie de KCl 150 mM. Fixez le ISE en plaçant la pâte à modeler sur le côté du bécher. La pointe se trouve dans le KCl 150. Continuer à utiliser le vo ISEer plusieurs jours tant qu'il fonctionne bien (voir l'étape 10.6).
- Pour un Na + ISE, stocker l'électrode dans NaCl 150 mM.
9. Préparer l'électrode de référence
Note: les étapes 9.1 à 9.3 peuvent être effectuées à l'avance. Les étapes 9.4 à 9.6 sont effectuées chaque jour expérimentale.
- Flame-polonais les extrémités de 10 filamenteuses tubes capillaires en verre borosilicate (diamètre extérieur 1,2 mm, diamètre intérieur de 0,69 mm, longueur 10 cm) à feu doux, en utilisant des mesures de sécurité.
- Sur un extracteur de la pipette, tirez pipettes à un diamètre de ~ 1 pointe - 2 pm.
- Pipettes de conserver dans un bocal de stockage de la pipette jusqu'à utilisation. Pipettes peuvent être stockées indéfiniment.
- Le jour de l'expérience, en utilisant un microfilaments et la seringue, remplir la pointe et la tige de la pipette avec de l'acétate de sodium 1 M. Assurer qu'il n'y a pas de bulles d'air et cette solution va à la pointe. Tapotez doucement la pipette si des bulles d'air sont présentes.
- L'utilisation d'un secondmicrofilaments et la seringue, le remblayage de la pipette avec 3 M de KCl. Encore une fois, assurez-bulles d'air ne sont pas présents.
- Stocker l'électrode de référence dans un bécher contenant 150 mM de KCl (voir l'étape 8.6).
10. Calibration de l'ISE
Remarque: Cette étape est effectué trois fois le jour de l'expérience: tôt dans la journée pour vous assurer que l'ISE travaille, puis avant et après les mesures des 20 - 25 gouttes liquide sécrété (tableau 3).
- Pour l'étalonnage du potassium ISE, placez deux 0,6 pi gouttes chacune des quatre concentrations de KCl suivantes sur le 3,5 cm boîte de Pétri d'enduit d'élastomère de silicone (préparé à l'étape 1): 15 mm, 75 mm, 150 mm et 200 mm. Couche avec précaution 2 ml d'huile minérale sur les gouttes.
- Pour le sodium ISE, utiliser des gouttes d'étalonnage de 15 mM et NaCl 150 mM.
- Placer le plat d'étalonnage sur la scène de l'stéréoscopique dans la cage de Faraday et illuminer.
- Threannonce l'électrode ISE et de référence sur les fils d'argent et de le fixer dans les supports de microélectrodes.
- En utilisant les micromanipulateurs, avancer l'ISE et des électrodes de référence dans la goutte mM KCl 15.
- Mettez l'électromètre à "exploiter" en mode. Autoriser la lecture de régler.
- Lecture d'enregistrement dans le carnet. Répéter avec le 75 mm, 150 mm et 200 mM gouttes. Calculer la pente pour déterminer si ISE fonctionne bien (voir l'étape 13.1 et le Tableau 3). Si non, préparer une nouvelle ISE.
Remarque: Les signes que l'ISE ne fonctionne pas bien: l'échec d'obtenir une lecture; lent à équilibrer (plusieurs secondes ou plus); lecture instable; pente <49 mV / changement de décile K + ou Na + concentration. - Entre le premier étalonnage de la journée et les mesures effectuées à l'étape 12, soit au cours de l'exécution de l'étape 11 (dissections tubules), stocker l'ISE et des électrodes de référence en mM KCl 150 (comme décrit dans l'étape 8.6).
11. Tubule Dissection
Remarque: cette étape est effectuée le jour de l'expérience.
- Aliquote une petite quantité (~ 500 - 600 pi) du milieu de bain standard (SBM), préparé à l'étape 4.2, pour une utilisation sur le jour de l'expérience et laisser réchauffer à température ambiante. Cela devrait être fait au moins 30 min avant de dissections, mais peut également être fait plus tôt. Aussi, avoir au moins 20 ml de solution saline RT drosophile (étape 4.1) disponibles avant de commencer dissections.
- Immédiatement avant de commencer dissections: voir le plat de dosage à un grossissement de 10x sous un stéréomicroscope et ajouter suffisamment SBM pour remplir presque chaque puits dans le plat de dosage, généralement entre 10 et 30 pi. Si les médicaments ou peptide vont être ajouté à la mi-SBM expérience, faire une note du volume exact de la SBM ajouté à chaque puits. Éviter de trop remplir le bien, car cela peut conduire à la tubule flotter à la dérive pendant l'expérience.
- Superposer soigneusement ~ 12 - 13 ml d'huile minérale au-dessus de sorte que les puits d'unerétabli. Cela permettra d'éviter l'évaporation des gouttes fluides sécrétés au cours de l'expérience.
- Lieu vole à être disséqué sur le CO 2 pad.
- Ramassez une mouche par l'intermédiaire de sa patte ou une aile avec une pince et les placer sur son dos (ventrale vers le haut) sur le plat de dissection élastomère de silicone revêtu préparé à l'étape 1. Empale le thorax avec une épingle minuties pour garantir la volée en place.
- Ajouter une goutte de solution saline RT drosophile (étape 4.1) pour immerger la volée dans une solution saline.
- Facultatif: clip hors ailes et les pattes. En pratique, cela est généralement pas nécessaire.
- Utilisez la pince à main non dominante à "conserver" l'abdomen de la mouche à la jonction thoraco-abdominale. Utilisez la pince à main dominante pour éplucher la cuticule abdominale distance, à partir de la jonction thoraco-abdominale et le déplacement vers l'extrémité de la queue de la mouche. L'intestin, avec les tubes de Malpighi attachés, doit être exposé à cette manœuvre.
- Sans toucher les tubules, disséquer gratuitement l'intestin moyen /intestin postérieur et les tubules attachés. Maintenez l'intestin dans les pinces à main non dominante et utiliser une aiguille de 30 G de rompre l'uretère de l'intestin, détacher les tubules de l'intestin et libre de la fly.It est essentiel que pas de larmes ou de loyers être introduits dans le tubule, autre qu'à l'uretère.
Remarque: La paire antérieure de tubules est plus facile à disséquer, mais les tubules postérieures peuvent être utilisés aussi bien. - Utilisation de la fine tige de verre (étape 2), ramasser la paire de tubules et de les transférer dans un puits de la boîte de dosage.
- Immédiatement après la paire de tubule a été transféré dans le puits, ramasser la fin de l'un des tubes avec la tige de verre, se retirer de la chute de la baignade jusqu'à la fin de l'uretère de coupe est à mi-chemin entre l'axe et la chute de la baignade, et enveloppement l'extrémité du tube autour de la broche l'aide de la baguette de verre. A la fin de cette manoeuvre, une tubule reste dans le bain de solution saline goutte et sécréter un fluide depuis l'extrémité de l'uretère de coupe, comme illustré sur la figure 1 . L'autre tube, marqué le «tube d'ancrage" dans la figure 1, est enroulé autour de la broche. Il ancre le tubule sécrétant en place, est entouré par de l'huile, et ne sécrète pas fluide.
- Immédiatement après l'étape 11.11, notez le bien, tubule des informations d'identification (par exemple, A, B, C) (par ex., Le génotype ou de l'état), et le temps (ce qui est l'heure de début lorsque le fluide commence à être sécrétée par le tubule dans la gouttelette de solution saline de baignade).
- Procéder à la prochaine dissection. Une fois que l'expérimentateur est l'homme de cette technique, il faut généralement 3 - 4 min à disséquer une paire de tubes, de les transférer à la solution saline de baignade, et envelopper le tubule ancrage autour de la broche. Par conséquent, 20 - 25 tubules peuvent être mis en place dans l'essai à l'intérieur de Ramsay 1,5 h. L'heure de début de chaque tube sera donc d'environ 3 - 4 min après l'heure de début du tubule précédente.
12. Prise de mesures
Remarque: Cette étape est effectuée le jour de l'expérience.
- Calibrer l'ISE (étape 10) environ 20 minutes avant la première mesure. Ce délai permet de faire une nouvelle ISE si nécessaire.
Remarque: Au moment voulu, par exemple après 2 h de sécrétion, la perte de fluide sécrété de chaque tube est prêt pour la mesure. - Noter le temps de mesure. Mesurer le diamètre de la goutte de fluide sécrété en utilisant le micromètre oculaire et fiche. Notez le grossissement, par exemple, 50X.
- Avancer le ISE et l'électrode de référence dans la goutte de liquide. Mettez l'électromètre à "opérer." Laisser la lecture se stabiliser. Enregistrez la valeur.
- Répétez l'opération pour la prochaine chute.
- A la fin de l'expérience, et répéter ces mesures d'étalonnage (étape 10).
13. Calculs
Remarque: Cette étape peut être effectuée soit à la fin de la journée de l'expérience, ou à un moment ultérieur.
- Calculer la moyennepente / changement décile de la concentration de potassium. Voir le tableau 3 pour un exemple.
Remarque: Pour sodium, les valeurs seront à la différence entre la 15 mM et 150 mM de NaCl mesures. - Déterminer la valeur moyenne des deux mesures (avant et après) de 200 mM de KCl (150 mM de NaCl ou).
- Calculer le volume de chaque gouttelette. ND = V 3/6, où d est le diamètre de la gouttelette mesurée avec le micromètre oculaire à l'étape 12.2.
- Calculer le taux de sécrétion = V / heure (Nl / min / tube), où V est le volume de la gouttelette déterminée à l'étape 13.3, et le temps est la durée pendant laquelle le fluide sécrété du tubule (= temps de mesure - durée de l'uretère tirant gouttelette de bain).
- Calculer la concentration d'ions en utilisant la formule [K] = 10e (Av / S) * 200 ou [Na] = 10e (Av / S) * 150, où Av = la différence (en mV) entre le potentiel mesuré du fluide sécrété goutte, et le potentiel de l'étalonnage goutte 200 mM (fou potassium; Goutte 150 mM pour le sodium). S = la pente déterminée à l'étape 13.1.
- Calculer flux d'ions = [ions] x taux de sécrétion de liquide. Pour les tubes de Drosophila, ce sera pmol / min / tubule.
14. Nettoyer
Remarque: Cette étape est réalisée à la fin de la journée de l'expérience.
- Un nettoyage en profondeur des puits est essentiel de veiller à ce que les cristaux de sel résiduels ne restent pas dans les puits, en modifiant la concentration en ions et l'osmolarité dans des expériences ultérieures.
- Laisser l'huile minérale pour égoutter.
- Rincer les puits de la boîte de dosage. Une pointe de pipette 200 pi peut être utilisée pour gratter doucement cristal de sel résiduel. En utilisant des tubes en plastique fixé au robinet, presser le tube pour créer un jet d'eau chaude à haute pression pour rincer abondamment à chaque puits.
- Laisser sécher O / N. En variante, un séchoir peut être utilisé pour sécher les puits.
- Rincez pince avec H 2 O déminéralisée etfaire tremper dans de l'éthanol pendant 15 minutes à plusieurs heures.
- Vidanger l'huile hors de plat d'étalonnage et laver avec de l'eau chaude. Utilisez un savon ainsi tant qu'il est complètement rincé. Effectuer le rinçage final à l'eau distillée H 2 O.
- Rincer les microfilaments et des seringues avec H 2 O. distillée
Résultats
Les figures 7 et 8 démontrent que l'utilisation de l'essai Ramsay avec des électrodes spécifiques des ions pour mesurer les concentrations de K + et Na + peut distinguer génétiquement et pharmacologiquement distinct K + et les flux de Na +, des informations qui ne sont pas capturés par la mesure des taux de sécrétion de liquide seul. Figure La figure 7 montre que diminution de la sécrétion de fluide dans les tubes de vol port...
Discussion
L'utilisation du test Ramsay, avec électrodes spécifiques des ions, permet de mesurer les taux de sécrétion des fluides et des flux d'ions dans insecte isolé Malpighi (rénales) tubules. Vingt ou plusieurs tubes peuvent être testés à la fois, ce qui permet un débit plus élevé par rapport au dosage de l'individu dans les tubules in vitro microperfused. En outre, des électrodes spécifiques des ions permettent la détermination des concentrations ioniques dans le fluide sécrété in s...
Déclarations de divulgation
The authors have nothing to disclose.
Remerciements
The authors wish to thank Drs. Sung-wan An and Mike O’Donnell for practical advice on establishing this assay, Dr. Chih-Jen Cheng for helpful discussions on the use of ion-specific electrodes, and Dr. Chou-Long Huang for his mentorship and support. This work was supported by the National Institutes of Health (K08DK091316 to ARR) and the American Society of Nephrology Gottschalk Award to ARR.
matériels
| Name | Company | Catalog Number | Comments |
| Sylgard 184 Silicone Elastomer Kit | Ellsworth Adhesives | http://www.ellsworth.com/dow-corning-sylgard-184-silicone-encapsulant-0-5kg-kit-clear/ | May be purchased from multiple distributors |
| Petri dish, polystyrene, 100 mm x 15 mm | Fisher | FB0875712 | Specific brand is not important |
| Petri dish, polystyrene, 35 mm x 10 mm | Corning Life Sciences | Fisher 08-757-100A | Specific brand is not important |
| Scalpel Handle #3 | Fine Science Tools | 10003-12 | Specific brand is not important |
| Scalpel Blades #1 | Fine Science Tools | 10011-00 | Specific brand is not important; use appropriate sharps precautions |
| Needle, 30 G x 1/2 | Becton Dickinson | 305106 | Use appropriate sharps precautions |
| Minutien pins, black anodized, 0.15 mm | Fine Science Tools | 26002-15 | |
| Stereomicroscope with ocular micrometer | Nikon | SMZ800 | Specific brand is not important; this is given as an example |
| Sheet of black stained glass, 3 mm (1/8 inch) thick | Hobby shop | Example includes Spectrum Black Opal by Spectrum Glass (http://www.delphiglass.com/spectrum-glass/opalescent/spectrum-black-opal) | |
| Glass cutting tools (glass cutter, glass cutting pliers) | Hobby shop | Examples include the Studio Pro Lightweight Running Pliers by Diamond Tech (http://www.delphiglass.com/glass-cutters-tools/pliers-nippers/studio-pro-lightweight-running-pliers) and the Studio Pro Brass Glass Cutter by Diamond Tech (http://www.delphiglass.com/glass-cutters-tools/glass-cutters/studio-pro-brass-glass-cutter). Use appropriate safety precautions when cutting glass | |
| Borosilicate glass capillary tube, unfilamented, GC120-10, OD 1.2 mm, ID 0.69 mm, length 10 cm | Warner Instruments | 30-0042 | |
| Borosilicate glass capillary tube, filamented, GC120F-10, OD 1.2 mm, ID 0.69 mm, length 10 cm | Warner Instruments | 30-0044 | |
| Nitric acid, 70% | Sigma | 438073 | CAUTION: see Material Data Safety Sheet for appropriate storage and handling guidelines. Specific brand is not important |
| Cimarec 7 in x 7 in hotplate | Fisher | 11675911Q | Specific brand is not important; caution when heated |
| Selectophore dichlorodimethylsilane | Sigma | 40136-1ML | CAUTION: see Material Data Safety Sheet for appropriate storage and handling guidelines |
| Two-step vertical pipet puller | Narishige | PC-10 | Other pipet pullers can be used; this is given as an example |
| Glass petri dish, 150 mm diameter x 15 mm height | Fisher | 08-748E | Specific brand is not important; only one dish needed |
| World Precision Instruments E210 1 mm micropipette storage jar | Fisher | 50-821-852 | May be available from other distributors. Useful to have two jars. Note that although this jar is specified for 1 mm pipets, and the pipets used here are 1.2 mm, in our experience the 1 mm jar works best for the 1.2 mm pipets. |
| Silica Gel, Tel-Tale Desiccant, indicating, 10-18 mesh | Fisher | S161-500 | Indicating silica useful for determining whether silica gel retains desiccating ability |
| World Precision Instruments MicroFil, 34G | Fisher | 50-821-914 | May be available from other distributors. |
| 1 ml syringe with luer lock | Becton Dickinson | 309659 | May be available from other distributors. |
| 3 ml syringe with luer lock | Becton Dickinson | 309657 | May be available from other distributors. |
| D300 3-way stopcock with female luer lock inlet port, male luer outlet port with rotating collar and guard | Cole-Parmer | UX-30600-02 | Specific brand is not important |
| Female Luer Locking Connector | 4 Medical Solutions | ADC 9873-10 | Specific brand is not important; barbed end is ~4 mm at narrowest point and ~7 mm at widest point. |
| Silicone Tubing I.D. x O.D. x Wall: 1/16 x 1/8 x 1/32 in. (1.59 x 3.18 x 0.79 mm) | Fisher | 14-179-110 | Specific brand is not important |
| E-3603 tubing, I.D. x O.D.: 1/32 x 3/32 in | Fisher | 14171208 | Specific brand is not important |
| Modeling clay | Specific brand is not important | ||
| Selectophore potassium ionophore I, cocktail B | Sigma | 99373 | CAUTION: see Material Data Safety Sheet for appropriate storage and handling guidelines |
| Selectophore sodium ionophore X | Sigma | 71747 | Sodium ionophore X = 4-tert-butylcalix[4]arene-tetraacetic acid tetraethylester |
| Selectophore 2-nitrophenyl octyl ether | Sigma | 73732 | |
| Selectophore sodium tetraphenylborate | Sigma | 72018 | CAUTION: see Material Data Safety Sheet for appropriate storage and handling guidelines |
| Schneider's Drosophila medium | Life Technologies | 21720024 | |
| High impedance electrometer | World Precision Instruments | FD223a | |
| Microelectrode holder 1 mm with 45° body, vented, with handle | Warner Instruments | 64-1051 | |
| Microelectrode holder 1 mm with straight body, vented | Warner Instruments | 64-1007 | |
| Silver wire | Warner Instruments | 64-1318 | |
| Micromanipulators, pair | Leitz | Various brands/models will work; this is an example | |
| Faraday cage | Technical Manufacturing Corporation | 81-334-03 | This is an example; any Faraday cage will work |
| Single gooseneck fiberoptic light | Nikon | Specific brand is not important | |
| mineral oil | Fisher | BP-2629 | Specific brand is not important |
| forceps, Dumont #5 with Biologie tip | Fine Science Tool | 11295-10 | May be available from other distributors. |
Références
- Ramsay, J. A. Active Transport of Water by the Malpighian Tubules of the Stick Insect, Dixippus-Morosus (Orthoptera, Phasmidae). J Exp Biol. 31, 104-113 (1954).
- Dow, J. A., et al. The malpighian tubules of Drosophila melanogaster: a novel phenotype for studies of fluid secretion and its control. J Exp Biol. 197, 421-428 (1994).
- Brand, A. H., Perrimon, N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development. 118, 401-415 (1993).
- Sozen, M. A., Armstrong, J. D., Yang, M., Kaiser, K., Dow, J. A. Functional domains are specified to single-cell resolution in a Drosophila epithelium. Proc Natl Acad Sci U S A. 94, 5207-5212 (1997).
- Rosay, P., et al. Cell-type specific calcium signalling in a Drosophila epithelium. J Cell Sci. 110 (15), 1683-1692 (1997).
- Dow, J. T., Davies, S. A. Integrative physiology and functional genomics of epithelial function in a genetic model organism. Physiol Rev. 83, 687-729 (2003).
- Beyenbach, K. W., Skaer, H., Dow, J. A. The developmental, molecular, and transport biology of Malpighian tubules. Annu Rev Entomol. 55, 351-374 (2010).
- Donnell, M. J., et al. Hormonally controlled chloride movement across Drosophila tubules is via ion channels in stellate cells. Am J Physiol. 274, 1039-1049 (1998).
- Linton, S. M., O'Donnell, M. J. Contributions of K+:Cl- cotransport and Na+/K+-ATPase to basolateral ion transport in malpighian tubules of Drosophila melanogaster. J Exp Biol. 202, 1561-1570 (1999).
- Rheault, M. R., O'Donnell, M. J. Analysis of epithelial K(+) transport in Malpighian tubules of Drosophila melanogaster: evidence for spatial and temporal heterogeneity. J Exp Biol. 204, 2289-2299 (2001).
- Donnell, M. J., Dow, J. A., Huesmann, G. R., Tublitz, N. J., Maddrell, S. H. Separate control of anion and cation transport in malpighian tubules of Drosophila Melanogaster. J Exp Biol. 199, 1163-1175 (1996).
- Cabrero, P., et al. Chloride channels in stellate cells are essential for uniquely high secretion rates in neuropeptide-stimulated Drosophila diuresis. Proc Natl Acad Sci U S A. 111, 14301-14306 (2014).
- Torrie, L. S., et al. Resolution of the insect ouabain paradox. Proc Natl Acad Sci U S A. 101, 13689-13693 (2004).
- Rodan, A. R., Baum, M., Huang, C. L. The Drosophila NKCC Ncc69 is required for normal renal tubule function. Am J Physiol Cell Physiol. 303, 883-894 (2012).
- Ianowski, J. P., Christensen, R. J., O'Donnell, M. J. Na+ competes with K+ in bumetanide-sensitive transport by Malpighian tubules of Rhodnius prolixus. J Exp Biol. 207, 3707-3716 (2004).
- Naikkhwah, W., O'Donnell, M. J. Salt stress alters fluid and ion transport by Malpighian tubules of Drosophila melanogaster: evidence for phenotypic plasticity. J Exp Biol. 214, 3443-3454 (2011).
- Donini, A., et al. Secretion of water and ions by malpighian tubules of larval mosquitoes: effects of diuretic factors, second messengers, and salinity. Physiol Biochem Zool. 79, 645-655 (2006).
- Maddrell, S. H. Secretion by Malpighian Tubules of Rhodnius movements of Ions and Water. J Exp Biol. 51, 71-97 (1969).
- Maddrell, S. H., Overton, J. A. Stimulation of sodium transport and fluid secretion by ouabain in an insect malpighian tubule. J Exp Biol. 137, 265-276 (1988).
- Williams, J. C., Beyenbach, K. W. Differential effects of secretagogues on Na and K secretion in the Malpighian tubules of Aedes Aegypti (L). J Comp Physiol. 149, 511-517 (1983).
- Maddrell, S. H., O'Donnell, M. J., Caffrey, R. The regulation of haemolymph potassium activity during initiation and maintenance of diuresis in fed Rhodnius prolixus. J Exp Biol. 177, 273-285 (1993).
- Messerli, M. A., Kurtz, I., Smith, P. J. Characterization of optimized Na+ and Cl- liquid membranes for use with extracellular, self-referencing microelectrodes. Anal Bioanal Chem. 390, 1355-1359 (2008).
- Ianowski, J. P., O'Donnell, M. J. Basolateral ion transport mechanisms during fluid secretion by Drosophila Malpighian tubules: Na+ recycling, Na+:K+:2Cl- cotransport and Cl- conductance. J Exp Biol. 207, 2599-2609 (2004).
- Neufeld, D. S., Leader, J. P. Electrochemical characteristics of ion secretion in malpighian tubules of the New Zealand alpine weta (Hemideina maori). J Insect Physiol. 44, 39-48 (1997).
- Greenspan, R. J. . Fly Pushing: The Theory and Practice of Drosophila Genetics. , (1997).
- Jayakannan, M., Babourina, O., Rengel, Z. Improved measurements of Na+ fluxes in plants using calixarene-based microelectrodes. J Plant Physiol. 168, 1045-1051 (2011).
- Wu, Y., Schellinger, J. N., Huang, C. L., Rodan, A. R. Hypotonicity Stimulates Potassium Flux through the WNK-SPAK/OSR1 Kinase Cascade and the Ncc69 Sodium-Potassium-2-Chloride Cotransporter in the Drosophila Renal Tubule. J Biol Chem. 289, 26131-26142 (2014).
- Blumenthal, E. M. Modulation of tyramine signaling by osmolality in an insect secretory epithelium. Am J Physiol Cell Physiol. 289, 1261-1267 (2005).
- Dow, J. A., Maddrell, S. H., Davies, S. A., Skaer, N. J., Kaiser, K. A novel role for the nitric oxide-cGMP signaling pathway: the control of epithelial function in Drosophila. Am J Physiol. 266, 1716-1719 (1994).
- Dube, K., McDonald, D. G., O'Donnell, M. J. Calcium transport by isolated anterior and posterior Malpighian tubules of Drosophila melanogaster: roles of sequestration and secretion. J Insect Physiol. 46, 1449-1460 (2000).
- Efetova, M., et al. Separate roles of PKA and EPAC in renal function unraveled by the optogenetic control of cAMP levels in vivo. J Cell Sci. 126, 778-788 (2013).
- Rheault, M. R., O'Donnell, M. J. Organic cation transport by Malpighian tubules of Drosophila melanogaster: application of two novel electrophysiological methods. J Exp Biol. 207, 2173-2184 (2004).
- Donnell, M. J. Too much of a good thing: how insects cope with excess ions or toxins in the diet. J Exp Biol. 212, 363-372 (2009).
- Cheng, C. J., Truong, T., Baum, M., Huang, C. L. Kidney-specific WNK1 inhibits sodium reabsorption in the cortical thick ascending limb. Am J Physiol Renal Physiol. 303, 667-673 (2012).
- Cheng, C. J., Yoon, J., Baum, M., Huang, C. L. STE20/SPS1-related Proline/alanine-rich Kinase (SPAK) is Critical for Sodium Reabsorption in Isolated Perfused Thick Ascending Limb. Am J Physiol Renal Physiol. , (2014).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.