Method Article
Isolement et imagerie vivants, régions intactes du stimulateur cardiaque du bassinetet du rein de souris par section vibratome
Dans cet article
Résumé
L’objectif de ce protocole est d’isoler les régions intactes du stimulateur cardiaque du bassinet du rein de la souris à l’aide d’une section vibratome. Ces sections peuvent ensuite être utilisées pour l’imagerie in situ du Ca2+ afin d’élucider les propriétés transitoires du Ca2+ des cellules du stimulateur cardiaque et d’autres cellules interstitielles dans des tranches de vibratome.
Résumé
Le bassinet du rein (RP) est une structure musculaire lisse en forme d’entonnoir qui facilite le transport normal de l’urine du rein à l’uretère par des contractions régulières et propulsives. Les contractions régulières de la RP reposent sur l’activité du stimulateur cardiaque, qui provient de la région la plus proximale de la RP à la jonction bassin-rein (PKJ). En raison de la difficulté d’accéder et de préserver les préparations intactes de la PKJ, la plupart des recherches sur le pacemaking RP se sont concentrées sur l’électrophysiologie unicellulaire et les expériences d’imagerie Ca2+. Bien que d’importantes révélations sur le pacemaking RP aient émergé de ces travaux, ces expériences ont plusieurs limites intrinsèques, notamment l’incapacité de déterminer avec précision l’identité cellulaire dans des suspensions mixtes et le manque d’imagerie in situ de l’activité du stimulateur cardiaque RP. Ces facteurs ont entraîné une compréhension limitée des mécanismes qui sous-tendent les contractions rythmiques normales de la RP. Dans cet article, un protocole est décrit pour préparer des segments intacts de PKJ de souris en utilisant une technique de sectionnement vibratome. En combinant cette approche avec des souris exprimant des rapporteurs spécifiques aux cellules et des indicateurs Ca2+ génétiquement codés, les chercheurs pourraient être en mesure d’étudier plus précisément les types de cellules spécifiques et les mécanismes responsables des contractions péristaltiques de la RP qui sont vitaux pour le transport normal de l’urine.
Introduction
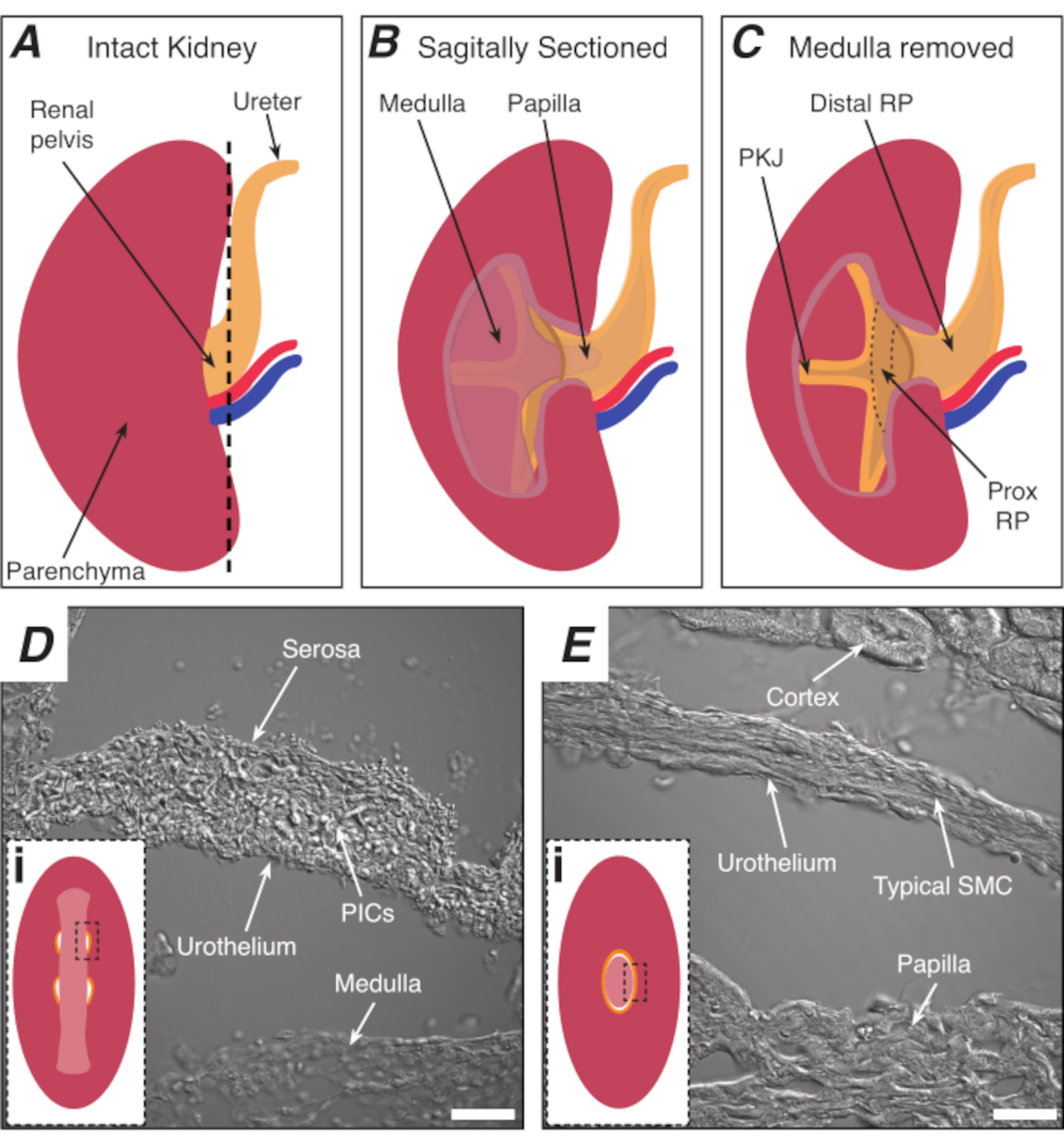
Le bassinet du rein (RP) est une structure musculaire lisse en forme d’entonnoir qui transporte l’urine du rein à l’uretère. Le RP transporte l’urine en générant des contractions rythmiques régulières (péristaltisme)1,2,3,4,5, qui propulse un bolus d’urine du rein distalement vers l’uretère et finalement vers la vessie, où il est stocké jusqu’à ce que la miction se produise6,7. La perte de cette activité régulière a des conséquences désastreuses, notamment l’hydronéphrose et l’insuffisance rénale1,3,8; par conséquent, il est essentiel d’étudier les mécanismes sous-jacents aux contractions régulières et rythmiques de la RP. Les contractions péristaltiques proviennent de la région la plus proximale de la RP-dans la jonction bassin-rein (PKJ)9,10,11,12,13,14,15 ( Figure1A-C) et se propagent distalement pour pousser l’urine de la papille dans la RP ( Figure1B). L’activité du stimulateur cardiaque électrique est enregistrée dans la PKJ sous forme de dépolarisations transitoires spontanées10,11, 12,13,15,16,17, qui proviendrant de cellules de stimulateur cardiaque spécialisées. On pense que ces cellules de stimulateurs cardiaques, auparavant appelées cellules musculaires lisses atypiques (ASMC), génèrent ou coordonnent l’activité des stimulateurs cardiaques et entraînent les contractions des cellules musculaires lisses « typiques » (SPC)9,10,11,18 , 19,20,21,22,23.
Les ASMC sont les plus abondants dans la RP proximale, au PKJ(Figure 1A-C),où les contractions péristaltiques et l’activité du stimulateur cardiaque électrique prennent naissanceà 5,7,8,9,12,13,14,16,17,18,19,20,21 ,22. Une étude récemment publiée par ce groupe a identifié le récepteur alpha du facteur de croissance dérivé des plaquettes (PDGFRα), en combinaison avec la chaîne lourde de myosine musculaire lisse (smMHC), comme un biomarqueur unique pour ces cellules interstitielles (CI)24, une découverte qui a été corroborée par d’autres groupes25. Sur la base de leur morphologie et de leur profil d’expression protéique, ces cellules ont été appelées PDGFRα+ IC de type 1 (PIC1)24,26. Les PIC1 résident dans la couche musculaire de la PKJ où ils présentent des transitoires Ca2+ à haute fréquence et de courte durée, censés sous-tendre la génération de potentiels de stimulateurs cardiaques24. Cependant, d’autres types de cellules existent dans la PKJ, y compris les PDGFRα+ CI (PIC2) non exprimant smMHC dans la couche adventitielle. Des rapports antérieurs ont suggéré que les CSM non-SMHC pourraient participer à la réglementation de l’activité des stimulateurs cardiaques19. Cependant, l’étude plus approfondie des CSM non smMHC est entravée par une mauvaise distinction au cours des études d’imagerie Ca2+. En règle générale, les types de cellules hétérogènes dans les préparations RP sont chargés sans discernement de colorants sensibles au Ca2+(par exemple, Fluo-4). Pour surmonter ces défis et étudier une variété de types de cellules dans la RP, des indicateurs Ca2+ (GECI) génétiquement codés peuvent être utilisés pour exprimer sélectivement les fluorophores sensibles au Ca2+dans les types cellulaires d’intérêt.
La majorité des études élucidant les propriétés transitoires du Ca2+ dans les PIC1/ASMC ont été réalisées en imageant des préparations tissulaires de RP à feuille plate19,21,27. Comme les PIC1 sont le seul type de cellule dans la PKJ à exprimer smMHC, l’expression conditionnelle du GECI, GCaMP, dans les cellules smMHC+ est appropriée pour étudier les PIC1 dans cette configuration. Cependant, comme les PIC1 et les PIC2 expriment tous deux PDGFRα, l’expression conditionnelle des variantes GCaMP dans les cellules PDGFRα+ interdit la distinction cellulaire dans les préparations à feuilles plates. Pour contourner ce problème, une approche de sectionnement vibratome a été utilisée pour distinguer les PIC1 et les PIC2 à travers la paroi tissulaire PKJ24. Pour révéler ces populations cellulaires discrètes, le RP a été sectionné coronalement, ce qui a permis d’identifier les PIC2 dans l’adventitia et les PIC1 dans la paroi musculaire sur la base d’un étiquetage immunohistochimique connu et de modèles d’expression GECI. À la suite de cette nouvelle approche d’imagerie PKJ, les PIC1 et PIC2 présentaient des propriétés de signalisation Ca2+ distinctes24. De plus, en isolant les sections les plus proximales de la région PKJ(Figure 2),la région du stimulateur cardiaque de la RP a été préservée d’une manière qui n’avait pas été accomplie auparavant. Ici, un protocole est décrit pour montrer comment isoler les préparations PKJ du rein de souris en utilisant la section vibratome, comment configurer ces préparations pour les expériences d’imagerie Ca2+ et comment distinguer les différents types de cellules à travers la paroi PKJ.
Protocole
Toutes les souris utilisées et les protocoles décrits dans cette étude étaient conformes au National Institutes of Health Guide for the Care and Use of Laboratory Animals et au Institutional Animal Use and Care Committee de l’Université du Nevada, Reno, NV. Les expériences et l’utilisation des animaux étaient également conformes aux principes et règlements décrits par Grundy28.
1. Générer des souris PDGFRα-GCaMP6f et SMCGCaMP3
- Croisez des souris GCaMP6flox/+ (B6; 129S-Gt(ROSA)26Sortm95.1(CAG-GCaMP6f)Hze/J) avec des souris PDGFRαCre (C57BL/6-Tg(Pdgfra-cre)1Clc/J) pour générer des souris PDGFRα-GCaMP6f.
- Croisez des souris GCaMP3lox/+ (B6.129S-Gt(ROSA)26Sortm38(CAG-GCaMP3)Hze/J)avec des souris mâles smMHCCreERT2 (B6. FVB-Tg(Myh11-cre/ERT2)1Soff/J) pour générer des souris SMC-GCaMP3.
REMARQUE: Seules les souris mâles du croisement (sourisGCaMP3lox / + et souris smMHCCreERT2) peuvent être utilisées car l’expression de Cre est pilotée à partir du chromosome Y. Les souris smMHCCreERT2 peuvent également être croisées avec des souris GCaMP6flox/+ pour un rapport signal/bruit Ca2+ amélioré et des propriétés temporelles plus rapides du signal Ca2+.
2. Préparer et injecter du tamoxifène à des souris transgéniques pour induire une expression conditionnelle de GCaMP
- Activer la Cre recombinase inductible pour l’expression de GCaMP spécifiques à une cellule dans des types de cellules spécifiques, comme décrit précédemment29.
3. Préparer des solutions
- Préparer 1 L de solution de bicarbonate de Krebs-Ringer (KRB) contenant 120,35 mM de NaCl, 5,9 mM de KCl, 15,5 mM deNaHCO3,1,2 mM de Na2HPO4,1,2 mM de MgCl2, 11,5 mM de glucose et 2,5 mM deCaCl2. Le jour de l’utilisation, maintenir la solution KRB sur la glace.
REMARQUE: KRB peut être conservé à 4 ° C jusqu’à une semaine et doit être pré-bulle avec un mélange de 97% O2 et 3% co2 pendant au moins 10 min avant utilisation.
4. Préparer des plats de dissection et d’imagerie au microscope revêtus d’élastomère de silicium
- Mélangez les composants en élastomère de silicium conformément aux instructions du fabricant. Remplissez une boîte de Petri de 35 mm x 10 mm et une boîte de Petri de 60 mm x 15 mm remplies d’environ un quart d’élastomère de silicium liquide pour les expériences d’imagerie et la dissection, respectivement. Polymériser l’élastomère de silicium à 37 °C pendant 1 jour avant utilisation.
REMARQUE: Pour améliorer le contraste des sections de reins minces, appliquez un petit cercle noir de papier à la base de la boîte de Petri d’imagerie avant de remplir avec de l’élastomère de silicium.
5. Dissection rénale
- Anesthésier les souris par inhalation de 3-4% d’isoflurane dans une hotte ventilée. Confirmer l’induction d’une anesthésie profonde par perte du réflexe de pincement des orteils et/ou de la queue, puis euthanasier les souris par luxation cervicale.
- Appliquez 70% d’éthanol sur la poitrine pour humidifier la fourrure. À l’aide de ciseaux à dissection externe, ouvrez la cavité abdominale via une incision longitudinale, avec des lames de ciseaux inclinées loin de l’animal pour éviter d’endommager les organes internes.
- À l’aide de pinces tissulaires internes et de ciseaux à dissection interne, pincez les intestins et éloignez-les de la paroi abdominale. Tout en soulevant les intestins, coupez le dessous des intestins libre du corps au niveau du duodénum proximal et du côlon distal pour accéder à l’espace rétropéritonéal contenant les reins.
- Une fois les reins exposés, extrayez-les individuellement. Pincez doucement et soulevez l’extrémité distale de l’uretère (~ 4 mm du rein) avec une pince à tissu. À l’aide des ciseaux à dissection, coupez sous l’uretère pincé vers le rein. Continuez à couper sous le rein jusqu’à ce qu’il soit libéré du tissu conjonctif environnant.
REMARQUE: Pour maximiser l’intégrité des tissus et la consistance de coupe pendant la sectionnement du vibratome, le rein doit être aussi intact que possible. Pour ce faire, évitez de pincer ou de couper le rein avec une pince et des ciseaux à dissection. - Placez le rein avec l’uretère attaché dans une solution de KRB glacée. Répétez les étapes 5.4 et 5.5 avec le rein controlatéral. Maintenir les reins en solution de KRB sur de la glace.
REMARQUE: Passez immédiatement à la section suivante du protocole pour préserver la viabilité des tissus PKJ. En raison de son emplacement anatomique profondément dans le parenchyme rénal, le PKJ est privé de contact avec la solution de KRB.
6. Préparer le rein pour la section vibratome
- Transférer le rein dans une boîte de dissection recouverte d’élastomère de silicium (60 mm x 15 mm) et le remplir de solution de KRB glacée jusqu’à ce que le rein soit complètement submergé.
- Sous un microscope à dissection, ancrer le rein à la base du plat en insérant des broches minutien dans l’uretère proximal et à travers la fine capsule rénale antérieure ou le tissu adipeux environnant.
REMARQUE: Veillez à ne pas percer le tissu du parenchyme rénal. - Utilisez des ciseaux à ressort fins et une pince interne pour enlever le tissu adipeux de la base du rein afin d’exposer la RP distale et l’uretère proximal.
- Retirez l’uretère proximal et une partie du RP distal de la base du RP à l’aide de ciseaux à ressort fins.
REMARQUE: Veillez à ne pas couper le parenchyme rénal environnant. La ligne pointillée noire de la figure 1A indique la position approximative de cette coupe. Cette coupe crée une base plate sur le rein pour une section tissulaire plus uniforme. Lors de la dissection du rein, il faut être conscient de la position anatomique de la région PKJ. La figure 1B montre que le rein intact peut être coupé le long d’un plan sagittal pour exposer la moelle, la papille (médullaire distale où convergent les canaux collecteurs) et la RP proximale et distale. Si la papille devait être complètement exposée, comme dans la figure 1C, la PKJ et la RP proximale (RP prox) peuvent être visualisées. Cependant, cela ne devrait pas être fait pour la technique du vibratome; cette description vise à orienter le lecteur vers l’emplacement de la PKJ en général, soulignée par la différence anatomique montrée dans les images lumineuses transmises de la région PKJ et de la région mi-RP de la Figure 1D,E. - Percez la capsule rénale externe avec une pince à pointe fine, en écartant les extrémités du corps rénal. À l’aide de pinces avec chaque main, pincez les extrémités lâches de la capsule et pelez-les. Continuez à peler la membrane restante de la capsule rénale jusqu’à ce qu’elle soit complètement enlevée.
REMARQUE: La capsule rénale est une couche fibreuse dure qui entoure le rein. Il doit être retiré avant le sectionnement du vibratome pour une coupe optimale.
7. Préparer et calibrer l’instrument vibratome
- Insérez une lame de rasoir dans le support de lame de l’instrument vibratome et ajustez l’angle de jeu de la lame à ~18°.
REMARQUE: En option pour une section de meilleure qualité, un bloc d’étalonnage (fourni avec certains instruments vibratome) doit être utilisé pour ajuster la position de la lame pour chaque nouvelle lame utilisée conformément aux instructions du fabricant. Cela assurera un positionnement optimal de la lame et minimisera les vibrations verticales. - Ajustez la vitesse d’avancement de la lame à 0,2 mm/s, la vibration horizontale / l’évent de la lame à une amplitude de 2,00 mm et la taille en Z de la lame à ~ 100-150 μm. Assurez-vous que l’épaisseur de la section rénale ne dépasse pas 150 μm, car cela aura un impact négatif sur les expériences d’imagerie Ca2+ (car la paroi PKJ roule et se replie sur elle-même si elle est trop épaisse).
REMARQUE: L’utilisateur doit affiner ces paramètres de coupe, car les paramètres varient entre les préparations rénales individuelles et les instruments vibratomes. Un réglage fin doit avoir lieu pendant le sectionnement lorsque l’utilisateur peut inspecter visuellement les sections au microscope, comme décrit à l’étape 7.2. - Installez un bain de glace et un plateau tampon sur l’instrument vibratome. Remplissez le bain de glace avec de la glace pilée et remplissez la zone intérieure de la scène avec une solution KRB glacée (remplissez à peu près à moitié plein). Pendant le sectionnement vibratome, surveillez et remplacez la glace pilée qui a fondu.
8. Vibratome sectionnant le rein
- Utilisez des pinces à extrémité émoussée pour saisir doucement et retirer le rein préparé de la solution de KRB glacée. Placez immédiatement le rein sur du papier absorbant pendant environ 2 à 4 s pour éliminer l’excès d’humidité externe. Rouler doucement le rein sur le papier absorbant pour s’assurer que tous les côtés du parenchyme ont séché afin qu’il y ait une adhérence optimale du rein au stade vibratome.
REMARQUE: Comme le RP est situé à l’intérieur du rein et donc protégé par le parenchyme externe, cette courte période de séchage ne serait pas préjudiciable à l’intégrité des tissus. - Appliquez immédiatement une fine couche de colle cyanoacrylate (~1 cm2)sur la base de la plaque d’échantillon de vibratome et utilisez une pince à extrémité émoussée pour placer le rein, côté uretère vers le bas, sur la zone recouverte de colle. Appliquez doucement une pression vers le bas sur le dessus du rein avec le bord plat de la pince pendant environ 10 à 20 s pour sécher la colle.
REMARQUE: Pour stabiliser le rein pendant cette procédure, utilisez une paire supplémentaire de pinces pour garder le rein droit pendant que la colle sèche. Il est essentiel que le rein adhère à la plaque d’échantillon en position verticale afin que les sections soient coupées droites. Pour vous assurer que le rein a adhéré avec succès à la plaque d’échantillon, poussez doucement le côté du rein. Si le rein a adhéré avec succès à la plaque, la base du rein doit rester fixée à la plaque. - Fixez fermement la plaque d’échantillon au bas du bac tampon. Ajustez le niveau de la solution KRB afin que le haut du rein soit complètement immergé. Pendant les étapes de sectionnement, lorsque la lame vibratome s’enfonce plus profondément dans le bac tampon, retirez la solution KRB afin que le support de lame ne soit pas immergé dans la solution.
REMARQUE: Si la colle n’a pas complètement séché avant de procéder à cette étape, la colle remontera souvent de la base et se déplacera vers le parenchyme rénal. Cet excès de colle rendra le sectionnement plus variable. Si cela se produit, retirez le rein de la plaque et procédez à une préparation rénale fraîche. - Pour le sectionnement automatique du vibratome, sélectionnez les positions de début et d’extrémité du cycle de coupe de la lame du vibratome. Vérifiez que ces positions sont d’environ 0,5 à 1 cm du rein pour vous assurer qu’à chaque avancement de la lame, tout le plan rénal est sectionné.
- Préparer une plaque multi-puits (24 ou 48 puits) en remplissant les puits avec une solution KRB et placer la plaque sur de la glace.
REMARQUE: Lorsqu’elles sont générées, les sections individuelles doivent être placées dans des puits séparés pour garder une trace de la profondeur de la section. - Démarrez le processus de découpe automatique. Lors du passage initial de la lame, assurez-vous que la lame entre en contact avec le sommet du rein. Si aucun contact n’est établi, ajustez la position Z de départ de la lame.
- À l’aide de pinces, collectez les sections libérées du rein. Transférez immédiatement les sections dans des puits individuels et notez la profondeur Z des sections pour évaluer l’emplacement approximatif de la PKJ dans les sections rénales.
REMARQUE: Selon les paramètres de coupe, certaines sections peuvent ne pas être coupées sans bloc rénal. Si cela se produit, utilisez soigneusement des ciseaux à ressort fins pour couper des sections du bloc rénal. Les utilisateurs sont également encouragés à visualiser activement les sections sous un microscope optique tout en flottant librement dans des puits individuels pour assurer des réglages de coupe et un emplacement des tissus optimaux. Les sections contenant la PKJ seront généralement dérivées à environ 1000-1500 μm du haut du rein. - Poursuivez le protocole de sectionnement jusqu’à ce que les régions PKJ deviennent plus apparentes. Reportez-vous à la section des résultats représentatifs pour obtenir une description des régions PKJ au fur et à mesure que la section se poursuit.
- À ce stade, optimisez les paramètres de sectionnement pour vous assurer que les sections sont libérées du bloc rénal uniformément et sont intactes. De plus, assurez-vous que les régions PKJ sont continues et ininterrompues, car les parois PKJ brisées ne permettront pas une imagerie adéquate des cellules à l’intérieur de la paroi en raison de l’effondrement. Si les parois se brisent, diminuez la vitesse de section et augmentez l’épaisseur de la section, et continuez à observer les sections au microscope optique pour affiner les paramètres de coupe.
- Conserver les sections à 4 °C dans une solution KRB jusqu’au début de l’expérimentation.
9. Acquisition d’image de tranche de rein Ca2+
- Transférer une tranche de rein individuelle dans une boîte d’imagerie recouverte d’élastomère de silicium (35 mm x 10 mm) et remplir immédiatement la boîte avec une solution de KRB fraîche et glacée.
- Insérez des broches minutien autour de la périphérie d’une tranche de rein pour fixer la section à la base de la boîte d’imagerie.
REMARQUE: Cette étape est essentielle pour empêcher la section de se déplacer lorsque des solutions physiologiques sont perfusées sur la tranche pendant l’imagerie. - Placez la parabole d’imagerie sur la scène d’un microscope confocal à disque rotatif vertical et commencez immédiatement à infuser avec la solution KRB.
REMARQUE: Dans ce protocole, un microscope vertical équipé d’une unité de scanner confocal à disque rotatif Nipkow à grande vitesse a été utilisé. - Maintenir le débit de perfusion à 3 mL/min et la température de la solution KRB à 36 ± 1 °C. Avant l’imagerie, laissez la tranche s’équilibrer pendant 1 h.
- Sélectionnez le cube d’imagerie dichroïque et les lasers appropriés. Acquérez des images avec un dispositif à couplage de charge multiplicateur d’électrons (EMCCD) ou une caméra scientifique complémentaire métal-oxyde-semi-conducteur.
REMARQUE: Le système confocal de ce protocole est équipé d’un laser de 488 nm pour exciter GCaMP6f ou GCaMP3. Les images ont été acquises à l’aide d’une caméra EMCCD de 512 pixels x 512 pixels. - Utilisez une lentille d’objectif à faible grossissement et à immersion dans l’eau (4x ou 10x) pour localiser la tranche de rein. Centrez le champ d’imagerie sur les zones de la tranche où le PKJ est présent. Identifiez les points de repère, comme illustré à la figure 2D,pour localiser la PKJ (c.-à-d. les demi-cercles de tissus musculaires suspendus entre le tissu parenchymateux).
- Une fois le PKJ localisé, utilisez un objectif à immersion dans l’eau à grossissement plus élevé (20x, 40x ou 60x) pour agrandir la zone d’intérêt.
REMARQUE: Dans ce protocole, l’ouverture numérique de l’objectif 20x (NA) était de 1,0, l’objectif NA de 40x était de 0,8 et l’objectif NA de 60x était de 1,0. - Distinguer différentes cellules d’intérêt dans la paroi PKJ en utilisant des souris transgéniques exprimant GCaMP6f ou GCaMP3 dans des cellules PDGFRα+ ou des SMC, respectivement. Observez les différents types de durées transitoires de Ca2+ dans les cellules PDGFRα+ dans la paroi PKJ dans PDGFRα+ GCaMP6f+ tranches rénales (Figure 3C, voir les résultats représentatifs pour la description) et dans GCaMP3+ SMC dans SMC GCaMP3+ tranches rénales (Figure 3D, voir les résultats représentatifs pour la description).
- Une fois que les cellules d’intérêt dans la paroi PKJ ont été identifiées, ajustez l’intensité du laser pour obtenir un bon rapport signal/bruit. Enregistrez des images à une fréquence d’échantillonnage temporelle comprise entre 16 Hz et 32 Hz à l’aide d’un logiciel d’acquisition approprié.
REMARQUE: Selon l’intensité du laser utilisé lors de l’imagerie, il est recommandé de limiter le nombre d’enregistrements en raison des effets de blanchiment sur les fluorophores Ca2+. L’utilisateur doit choisir la durée d’enregistrement en fonction des objectifs expérimentaux spécifiques.
10. Analyse d’imagerie Ca2+
- Effectuer une analyse d’imagerie Ca2+ de différents types de cellules dans les régions des stimulateurs cardiaques PKJ par cartographie spatio-temporelle comme décrit précédemment pour d’autres préparations de stimulateurs cardiaques intacts dans le tractus gastro-intestinal29,30.
Résultats
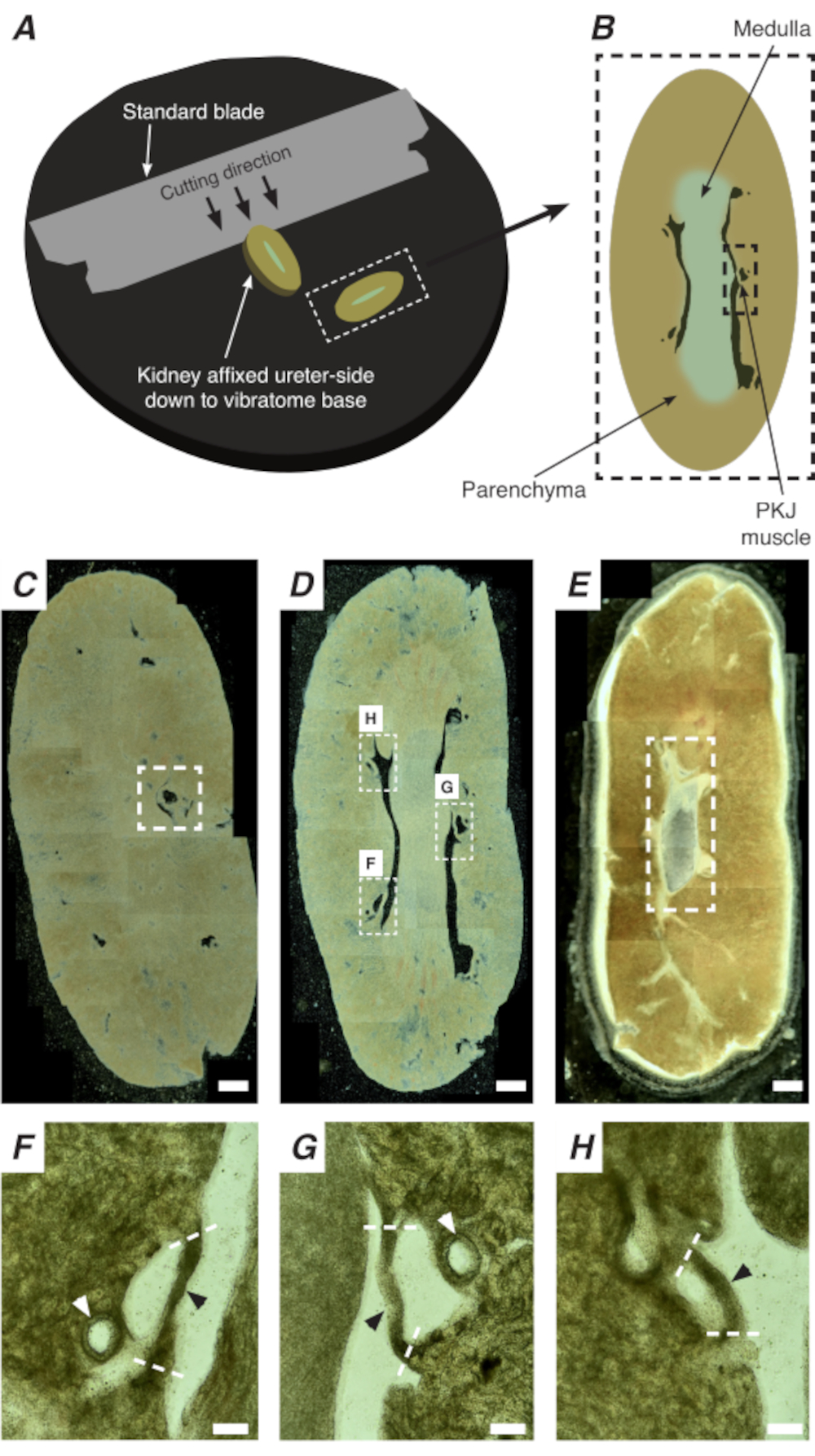
L’imagerie In situ Ca2+ de la PKJ peut révéler une activité cellulaire importante des cellules du stimulateur cardiaque RP. En utilisant des souris qui expriment des indicateurs ca2+ codés génétiquement (tels que GCaMP), pilotés par des promoteurs spécifiques aux cellules, des informations sur le rythme RP peuvent être obtenues avec une précision et des détails qui ne sont pas possibles à partir d’expériences d’imagerie Ca2+ à partir de préparations RP à feuille plate. Le début de la PKJ se distingue par l’apparition soudaine de demi-cercles de muscles suspendus entre le tissu parenchymateux du rein(Figure 2C; PKJ proximale enfermée dans une boîte en pointillés). Au cours des cycles de sectionnement suivants, la moelle interne se distingue du tissu cortical environnant. Au microscope optique, la moelle interne apparaît striée par régions, de couleur plus claire par rapport au tissu cortical et discontinue sur son long axe avec le reste du rein (Figure 2B, D). À ce stade, d’autres régions PKJ commenceront à apparaître. Des exemples de ceci sont montrés dans la figure 2D (rectangles pointillés, H et G) où 3 demi-cercles de muscle sont suspendus par du tissu parenchymateux. Ces bandes musculaires seront étroitement apposées à la papille interne et seront généralement voisines d’une artériole rénale(Figure 2D,rectangles pointillés; Figure 2F-H, pointes de flèches noires). Au fur et à mesure que d’autres sections distales sont dérivées, ces bandes de muscle s’intégreront pour former une structure plus complète et unifiée, indiquant la fin de la région PKJ(Figure 2E).
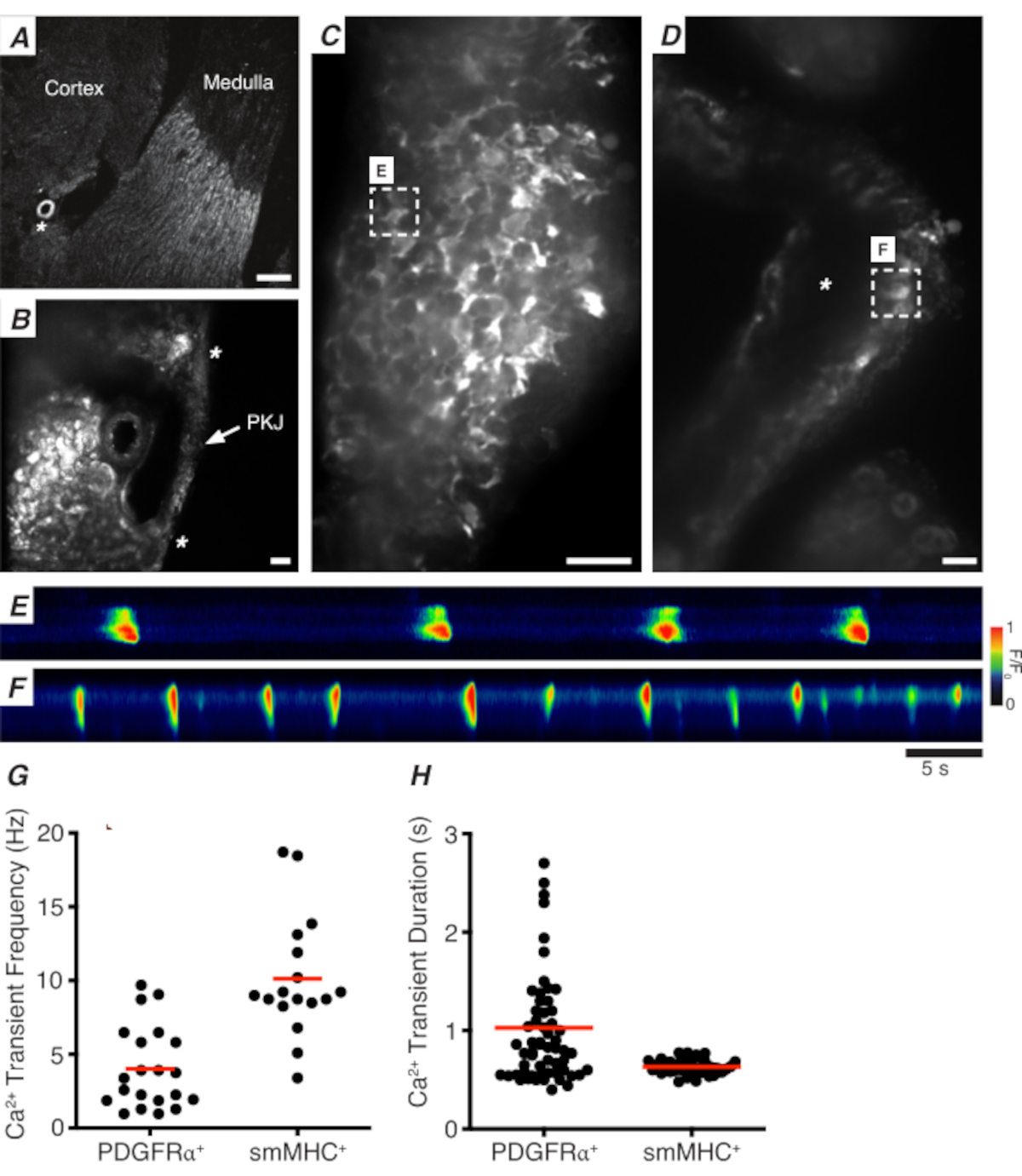
La figure 3A,B montre une section PKJ à faible puissance (4-10x) d’une souris exprimant GCaMP dans des cellules PDGFRα+ (GCaMP6f exprimée par la Cre-recombinase inductible entraînée par Pdgfra). En utilisant des repères tels que l’artériole rénale (Figure 3A; astérisque), les expérimentateurs devraient être en mesure de distinguer facilement la fine paroi PKJ suspendue entre le tissu parenchymateux (Figure 3B; astérisques). L’expression de GCaMP6f dans ce tissu transgénique spécifique est répartie sur toute la largeur de la PKJ, à la fois sur les couches musculaires et adventitielles(Figure 3C).
Dans les tranches de REIN PDGFRα+ GCaMP6f+, un réseau de cellules qui s’étend généralement sur la largeur de la paroi PKJ sera fluorescent(Figure 3C)et affichera des transitoires Ca2+ oscillants de différentes durées et fréquences. Les cellules PDGFRα+ de la paroi PKJ affichent deux types différents de durées transitoires ca2+. Dans la couche adventitielle (orientée plus près du cortex), les cellules PDGFRα+ présentent sous la forme d’un réseau de cellules et leurs processus sont définis. Les cellules ADVENTICES PDGFRα+ présentent des transitoires Ca 2+ à basse fréquence (4 ± 2,7 Hz) et de longue durée (1 ±0,67 s). La deuxième couche de cellules PDGFRα+, présente dans la couche musculaire (orientée plus près de la moelle), présente des fréquences et des durées transitoires Ca2+ similaires à celles des cellules SMC GCaMP3+ (décrites ci-dessous) car elles sont du même type de cellule.
Dans SMC GCaMP3+ tranches de rein, une couche de cellules GCaMP3+ est présente dans la couche musculaire ( Figure3D). Il n’y aura pas de signal fluorescent dans la couche adventitielle (Figure 3D; astérisque). Les GCaMP3+ SMC dans la couche musculaire présentent généralement des transitoires Ca2+ à haute fréquence (10 ± 4 Hz) et de courte durée (632 ± 74 s). Les cellules PDGFRα+ situées dans l’adventice PKJ provoquent des transitoires Ca2+ de longue durée et à basse fréquence (Figure 3E, Vidéo 1). Cependant, les expériences d’imagerie Ca2+ à partir de tissus exprimant GCaMP3 entraînés par le promoteur Myh11 sont limitées à l’aspect musculaire de la PKJ(Figure 3D). Par rapport aux cellules PDGFRα+ dans l’adventice, les SNC ont tiré plus fréquemment des transitoires Ca2+ de plus courte durée (Figure 3F, Vidéo 2).
En plus de comprendre les propriétés de signalisation dans les PKJ PDGFRα+ CI, l’application de cette technique pour étudier d’autres types de cellules dans le rein sectionné vibratome a été démontrée dans cet article. Après un examen attentif de la médusullaire rénale (chez la souris exprimant GCaMP6f dans les cellules PDGFRα+), un réseau de signaux fluorescents Ca2+ à l’intérieur et autour des conduits collecteurs (Vidéo 3) a été observé. Les cellules médullaires PDGFRα+ ont déclenché des transitoires spontanés de Ca2+ de fréquence et de durée variables. Ces études d’imagerie Ca2+ de sections de vibratomes rénaux pourraient également être étendues à l’étude des artérioles rénales (~ 50-80 mm de diamètre) qui sont souvent voisines des segments musculaires PKJ(Figure 2F,G; flèches blanches). L’imagerie Ca2+ des artérioles rénales (à partir de tissus exprimant GCaMP dans les cellules musculaires lisses) démontre des transitoires Ca2+ oscillants dans les SMC (Vidéo 4).

Figure 1: Anatomie rénale de base et emplacement de la région du stimulateur cardiaque PKJ. (A) Diagramme du rein intact montrant l’orientation de la RP et de l’uretère. L’artère rénale et la veine rénale sont affichées en rouge et en bleu, respectivement. (B) Le rein intact peut être coupé le long d’un plan sagittal pour exposer l’aspect interne du rein, y compris la moelle, la papille (médullaire distale où convergent les canaux collecteurs) et la RP proximale et distale. (C) La moelle et la papille peuvent être excisées pour exposer complètement la PKJ et la RP prox. (D et E) représentent des images lumineuses transmises de la région du stimulateur cardiaque PKJ et de la RP distale, respectivement. La coupe séquentielle vers l’extrémité distale du bassin entraîne la combinaison des demi-cercles musculaires dans la région PKJ(Di)en un seul anneau musculaire plus épais(Ei) qui encapsule toute la papille. Les rectangles noirs en pointillés dans Di et Ei montrent des zones approximatives dans les sections des reins coronaires où des images de lumière transmise ont été acquises. L’orientation des images D et E est de 90° dans le sens inverse des aiguilles d’une montre par rapport aux encarts respectifs(Di et Ei). Barres d’échelle en D et E = 20 μm. Abréviations : RP = bassinet du rein; prox RP = bassinet du rein proximal; PKJ = jonction pelvienne-rénale; PICs = cellules interstitielles alpha-positives du facteur de croissance dérivé des plaquettes; SMC = cellule musculaire lisse. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 2: Sectionnement vibratomique de reins entiers pour générer des sections minces. (A) Les reins sont montés du côté de l’uretère jusqu’à la base de l’instrument vibratomique, et une lame standard (attachée à la tête du vibratome) est utilisée pour couper des sections séquentielles de l’extrémité proximale à l’extrémité distale du rein. (B) Représentation schématique d’une section mince coupée de tout le rein avec des repères annotés. Les segments musculaires PKJ (rectangle pointillé noir) sont souvent en suspension entre le tissu parenchymateux. (C) Image microscopique légère d’une section rénale proximale. L’apparition de bandes musculaires suspendues entre le tissu parenchymateux indique le début des projections proximales de PKJ (indiquées à l’intérieur du rectangle pointillé blanc). (D) Image microscopique lumineuse représentant la région optimale où plusieurs (2-3) segments PKJ peuvent être trouvés (zones dans des rectangles pointillés blancs). Les bandelettes musculaires minces de PKJ sont suspendues entre le parenchyme rénal et s’alignent étroitement avec les artérioles rénales et la médulla. (E) Image microscopique lumineuse d’une section rénale distale. Des segments musculaires individuels ont fusionné pour former une seule bande musculaire continue (rectangle en pointillés blancs) qui entoure la papille interne (non présente sur cette image). Barres d’échelle C-E = 500 μm. F-H Les régions zoomées (20x) du panneau D indiquent l’emplacement du PKJ (pointes de flèches noires), des artérioles rénales (pointes de flèches blanches) et des sites de coupe pour isoler le PKJ (lignes blanches pointillées). Barres d’échelle F-H = 100 μm. Abréviation : PKJ = jonction pelvienne-rénale. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 3: Imagerie Ca2+ des sections de vibratome. (A) Image représentative de faible puissance (4x) d’une section vibratome indiquant l’emplacement de l’artériole rénale (astérisque). Barre d’échelle = 200 μm. (B) Image représentative zoomée (20x) de la PKJ (étiquetée) suspendue entre le tissu parenchymateux rénal indiquant les emplacements du muscle PKJ (pointe de flèche blanche), artériole rénale (astérisque). Barre d’échelle = 50 μm. (C) Image haute puissance (40x) du PKJ exprimant GCaMP dansles cellules PDGFRα +. Barre d’échelle = 20 μm. (D) Image de haute puissance (20x) de la PKJ exprimant GCaMP dans les cellules musculaires lisses. Barre d’échelle = 20 μm. (E) Carte spatio-temporelle des transitoires Ca2+ échantillonnés à partir d’une cellule GCaMP+ PDGFRα+ indiquée dans le panneau C. Rechercher une table codée pour F/F0. (F) Carte spatio-temporelle des transitoires ca2+ prélevés à partir d’une cellule GCaMP+ PDGFRα+ indiquée dans le panneau D. Rechercher une table codée pour F/F0. (G) Données représentatives de la fréquence transitoire Ca2+ (Hz) pour les cellules GCaMP+ PDGFRα+ et les cellules GCaMP+ smMHC. (H) Données représentatives pour la durée transitoire Ca2+ pour les cellules GCaMP+ PDGFRα+ et les cellules GCaMP+ smMHC. Abréviations: PKJ = jonction pelvienne-rénale; PDGFRα+ = récepteur alpha-positif du facteur de croissance dérivé des plaquettes; smMHC = chaîne lourde de myosine musculaire lisse. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
Vidéo 1 : Transitoires spontanés de Ca2+ dans les cellules GCaMP+ PDGFRα+ dans les sections de vibratome PKJ. Abréviations: PKJ = jonction pelvienne-rénale; PDGFRα+ = récepteur alpha-positif du facteur de croissance dérivé des plaquettes. Veuillez cliquer ici pour visionner cette vidéo. (Cliquez avec le bouton droit de la souris pour télécharger.)
Vidéo 2 : Transitoires spontanés de Ca2+ dans les cellules musculaires lisses GCaMP+ dans les sections vibratomes de la jonction pelvienne-rénale. Veuillez cliquer ici pour visionner cette vidéo. (Cliquez avec le bouton droit de la souris pour télécharger.)
Vidéo 3 : Transitoires spontanés de Ca2+ dans les cellules GCaMP+ PDGFRα+ dans les sections de vibratome médullaire rénale. Abréviation : PDGFRα+ = récepteur alpha-positif du facteur de croissance dérivé des plaquettes. Veuillez cliquer ici pour visionner cette vidéo. (Cliquez avec le bouton droit de la souris pour télécharger.)
Vidéo 4 : Activité transitoire Ca2+ de faible amplitude dans les sections vibratomes de l’artériole rénale. Veuillez cliquer ici pour visionner cette vidéo. (Cliquez avec le bouton droit de la souris pour télécharger.)
Discussion
La RP est constituée d’une population hétérogène de cellules avec des densités cellulaires différentielles observées dans diverses régions RP. Les PIC1 (précédemment appelés ASMC) sont les plus abondants dans la PKJ, où l’activité du stimulateur cardiaque prend naissance24. Le protocole, décrit ici, permet aux chercheurs d’isoler la région du stimulateur cardiaque du reste du rein de la souris. En coupant des sections de PKJ à l’aide d’un vibratome, les régions du stimulateur cardiaque de la RP (identifiées comme des bandes musculaires attachées au parenchyme) sont maintenues intactes, ce qui permet d’utiliser l’imagerie in situ pour étudier avec précision les cellules du stimulateur cardiaque RP lorsqu’elles sont combinées avec des rapporteurs de fluorescence spécifiques aux cellules.
Bien que cette approche puisse fournir de nouvelles informations sur le rythme de la RP, les expérimentateurs devraient connaître certaines considérations pour améliorer les résultats d’imagerie et l’efficacité de la section. Pour un utilisateur non formé, cette méthode d’isolement et d’imagerie PKJ est plus facile à apprendre que la dissection nette typique des préparations RP à feuille plate. Une dissection nette de la RP à partir de reins entiers nécessite des semaines de pratique constante pour isoler avec succès des tissus viables pour des expériences de physiologie. Comme ce protocole de sectionnement vibratome nécessite peu de connaissances pointues en dissection, il est accessible aux utilisateurs qui n’ont pas d’expérience dans la dissection d’autres structures musculaires lisses.
Cependant, il y a quelques points critiques à noter pour ce protocole. Réussir à adhérer les reins à la plaque d’échantillon de vibratome nécessite de la dextérité et de la patience. Si le rein est mal orienté et penche d’un côté, des sections obliques plutôt que droites seront coupées. En raison de la nature délicate de la PKJ, l’angle oblique peut souvent détruire les bandes musculaires de la région du stimulateur cardiaque. En outre, l’imagerie des sections obliques entraîne une mauvaise acquisition de l’imagerie car les réseaux cellulaires ne sont généralement pas dans le même plan focal. La procédure prend également beaucoup de temps, la section d’un seul rein prenant souvent jusqu’à une heure, au cours de laquelle la configuration nécessite une surveillance.
Alors que le mouvement du vibratome peut être accéléré, si la vitesse est trop augmentée (>20% que ce qui est recommandé dans le protocole), la lame déchiquetera plutôt que de couper proprement le rein, entraînant la perte de structures PKJ délicates. De même, une vitesse de coupe trop faible peut entraîner un dentelage de la section. L’optimisation de la vitesse de coupe et de l’amplitude de la lame est essentielle. Des précautions doivent également être prises lors de la manipulation des sections vibratomes. En raison de leur nature délicate, les muscles PKJ sont facilement perturbés lors de la manipulation et peuvent se déchirer. Un utilisateur bien formé sera en mesure de récolter environ 1 à 2 régions PKJ par tranches de rein qui conviennent aux expériences d’imagerie Ca2+. En règle générale, les sections PKJ qui ne répondent pas aux critères d’imagerie Ca2+ ont : 1) une mauvaise expression de GCaMP, 2) une paroi PKJ contorsionné ou 3) une paroi PKJ cassée. Pour l’analyse des données, environ 3 à 4 cellules par champ de vision (FOV) ont pu être échantillonnées.
Bien qu’il y ait de nombreuses cellules dans le champ de vision des sections PDGFRα-GCaMP6f et SMC-GCaMP3 PKJ, les petits mouvements tissulaires excluent souvent les cellules de l’analyse. Cela peut généralement être résolu en appliquant un protocole de stabilisation aux images. Dans des conditions où les préparations ne bougent pas, au moins 3 à 5 cellules peuvent être échantillonnées à partir de sections PDGFRα-GCaMP6f et 5 à 6 cellules de sections SMC-GCaMP3. En règle générale, le temps nécessaire entre le sacrifice d’animaux (l’âge optimal pour les souris est de 8 à 16 semaines) et la réalisation d’expériences d’imagerie Ca2+ est de 2 à 3 h, ce qui est suffisant pour assurer l’intégrité des tissus, si les tissus sont incubés dans des solutions glacées tout au long de la procédure si nécessaire. En résumé, un protocole de coupe de vibratome a été décrit ici pour générer des préparations intactes de régions RP PKJ à partir du rein de la souris. Cette technique permet de préserver les régions des stimulateurs cardiaques RP pour des études d’imagerie in situ Ca2+ afin d’étudier les mécanismes des stimulateurs cardiaques RP.
Déclarations de divulgation
Les auteurs n’ont rien à divulguer.
Remerciements
Ce projet a été financé par R01 DK124509 de NIDDK.
matériels
| Name | Company | Catalog Number | Comments |
| 24-well culture plate | ThermoFisher Scientific | 142485 | To store kidney slices in |
| 35 mm x 10 mm Petri dishes | Sigma Aldrich | CLS430165 | Kidney slice calcium imaging dish |
| 48-well culture plate | ThermoFisher Scientific | 152640 | To store kidney slices in |
| 60 mm x 15 mm Petri dish | Sigma Aldrich | P5481 | Kidney sharp dissection dish |
| Absorbent paper | Fisher Scientific | 06-666A | To dry the kidney before applying glue |
| B6;129S-Gt(ROSA)26Sor/J | The Jackson Laboratory | 13148 | GCaMP3 Mice |
| B6;129S-Gt(ROSA)26Sor/J | The Jackson Laboratory | 24105 | GCaMP6f Mice |
| B6.FVB-Tg(Myh11-cre/ERT2)1Soff/J | The Jackson Laboratory | 19079 | smMHC-CRE Mice |
| C57BL/6-Tg(Pdgfra-cre)1Clc/J | The Jackson Laboratory | 13148 | PDGFRa-CRE Mice |
| Cyanoacrylate glue | Amazon | B001PILFVY | For adhering the kidney to the specimen plate |
| Ethanol | Phamco-Aaper | SDA 2B-6 | For dissection |
| Extra-Fine Bonn Scissors | Fine Science Tools | 14083-08 | Used for internal dissecting scissors |
| Fine scissors | Fine Science Tools | 14060-09 | Used for external dissecting scissors |
| Fine-tip forceps | Fine Science Tools | 11254-20 | Used for fine dissection of kidney |
| Gillette Silver Blue double-edge blades | Amazon | B009XHQGYO | For insertion into blade holder of vibratome |
| ImageJ | NIH | For calcium imaging analysis | |
| Isoflurane | Baxter | NDC 1001936060 | For anesthesia |
| Minutien pins | Fisher Scientific | NC9677548 | Pins were cut in half to reduce their length |
| Silicon elastomer | Fisher Scientific | NC9285739 | Sylgard 184 |
| Student Adson Forceps | Fine Science Tools | 91106-12 | For gently holding and moving the kidney |
| Student Dumont Forceps | Fine Science Tools | 91150-20 | Used for internal dissecting forceps |
| Vannas spring scissors | Fine Science Tools | 15000-03 | For sharp dissection and cleanup of isolated kidney |
| Vibrocheck | Leica | 14048142075 | Optional component for calibrating blade movement during cutting |
| VT1200 S Vibrating Blade Microtome | Leica | 14912000001 | Configuration 1 is used in our protocol |
Références
- Constantinou, C. E., Djurhuus, J. C. Pyeloureteral dynamics in the intact and chronically obstructed multicalyceal kidney. The American Journal of Physiology. 241 (5), 398-411 (1981).
- Constantinou, C. E., Yamaguchi, O. Multiple-coupled pacemaker system in renal pelvis of the unicalyceal kidney. TheAmerican Journal of Physiology. 241 (5), 412-418 (1981).
- Constantinou, C. E., Hrynczuk, J. R. Urodynamics of the upper urinary tract. Investigative Urology. 14 (3), 233-240 (1976).
- Schmidt-Nielsen, B., Schmidt-Nielsen, B. On the function of the mammalian renal papilla and the peristalsis of the surrounding pelvis. Acta Physiologica. 202 (3), 379-385 (2011).
- Dwyer, T. M., Schmidt-Nielsen, B. The renal pelvis: machinery that concentrates urine in the papilla. Physiology. 18 (1), 1-6 (2003).
- Hill, W. G. Control of urinary drainage and voiding. Clinical Journal of the American Society of Nephrology. 10 (3), 480-492 (2015).
- Brading, A. F. The physiology of the mammalian urinary outflow tract. Experimental Physiology. 84 (1), 215-221 (1999).
- Dixon, J. S., Gosling, J. A. The musculature of the human renal calices, pelvis and upper ureter. Journal of Anatomy. 135, 129-137 (1982).
- Lang, R. J., et al. Pyeloureteric peristalsis: role of atypical smooth muscle cells and interstitial cells of Cajal-like cells as pacemakers. The Journal of Physiology. 576, 695-705 (2006).
- Lang, R. J., Takano, H., Davidson, M. E., Suzuki, H., Klemm, M. F. Characterization of the spontaneous electrical and contractile activity of smooth muscle cells in the rat upper urinary tract. Journal of Urology. 166 (1), 329-334 (2001).
- Lang, R. J., Exintaris, B., Teele, M. E., Harvey, J., Klemm, M. F. Electrical basis of peristalsis in the mammalian upper urinary tract. Clinical and Experimental Pharmacology & Physiology. 25 (5), 310-321 (1998).
- Morita, T., Ishizuka, G., Tsuchida, S. Initiation and propagation of stimulus from the renal pelvic pacemaker in pig kidney. Investigative Urology. 19 (3), 157-160 (1981).
- Tsuchida, S., Morita, T., Harada, T., Kimura, Y. Initiation and propagation of canine renal pelvic peristalsis. Urologia Internationalis. 36 (5), 307-314 (1981).
- Yamaguchi, O. A., Constantinou, C. E. Renal calyceal and pelvic contraction rhythms. American Journal of Physiology - Regulatory Integrative and Comparative Physiology. 257 (4), 788-795 (1989).
- Klemm, M. F., Exintaris, B., Lang, R. J. Identification of the cells underlying pacemaker activity in the guinea-pig upper urinary tract. Journal of Physiology. 519 (3), 867-884 (1999).
- Lang, R. J., et al. Spontaneous electrical and Ca2+ signals in the mouse renal pelvis that drive pyeloureteric peristalsis. Clinical and Experimental Pharmacology & Physiology. 37 (4), 509-515 (2010).
- Lutzeyer, W. Pacemaker process of ureteral peristalsis in multicalyceal kidneys. Urologia Internationalis. 37 (4), 240-246 (1982).
- Lang, R. J., Hashitani, H. Pacemaker mechanisms driving pyeloureteric peristalsis: modulatory role of interstitial cells. Advances in Experimental Medicine and Biology. 1124, 77-101 (2019).
- Hashitani, H., et al. Interstitial cell modulation of pyeloureteric peristalsis in the mouse renal pelvis examined using FIBSEM tomography and calcium indicators. Pflugers Archiv: European Journal of Physiology. 469 (5-6), 797-813 (2017).
- Lang, R. J., Hashitani, H., Tonta, M. A., Suzuki, H., Parkington, H. C. Role of Ca2+ entry and Ca2+ stores in atypical smooth muscle cell autorhythmicity in the mouse renal pelvis. British Journal of Pharmacology. 152 (8), 1248-1259 (2007).
- Lang, R. J., Hashitani, H., Tonta, M. A., Parkington, H. C., Suzuki, H. Spontaneous electrical and Ca2+ signals in typical and atypical smooth muscle cells and interstitial cell of Cajal-like cells of mouse renal pelvis. The Journal of Physiology. 583, 1049-1068 (2007).
- Hashitani, H., Lang, R. J., Mitsui, R., Mabuchi, Y., Suzuki, H. Distinct effects of CGRP on typical and atypical smooth muscle cells involved in generating spontaneous contractions in the mouse renal pelvis. British Journal of Pharmacology. 158 (8), 2030-2045 (2009).
- Iqbal, J., et al. Potassium and ANO1/ TMEM16A chloride channel profiles distinguish atypical and typical smooth muscle cells from interstitial cells in the mouse renal pelvis. British Journal of Pharmacology. 165 (7), 2389-2408 (2012).
- Grainger, N., et al. Identification and classification of interstitial cells in the mouse renal pelvis. Journal of Physiology. 598 (15), 3283-3307 (2020).
- Hashitani, H., Mitsui, R., Lang, R. Functional heterogeneity of PDGFRα (+) cells in spontaneously active urogenital tissues. Neurourology and Urodynamics. 39 (6), 1667-1678 (2020).
- Hashitani, H., Lang, R. J. ATYPICAL or INTERSTITIAL, take your PIC. Journal of Physiology. 598 (15), 3061-3062 (2020).
- Lang, R. J., Hashitani, H., Tonta, M. A., Suzuki, H., Parkington, H. C. Role of Ca2+ entry and Ca2+ stores in atypical smooth muscle cell autorhythmicity in the mouse renal pelvis. British Journal of Pharmacology. 152 (8), 1248-1259 (2007).
- Grundy, D. Principles and standards for reporting animal experiments in The Journal of Physiology and Experimental Physiology. Experimental Physiology. 100 (7), 755-758 (2015).
- Drumm, B. T., Hennig, G. W., Baker, S. A., Sanders, K. M. Applications of spatio-temporal mapping and particle analysis techniques to quantify intracellular Ca 2+ signaling in situ. Journal of Visualized Experiments. 2019 (143), 1-13 (2019).
- Leigh, W. A., et al. A high throughput machine-learning driven analysis of Ca2+ spatio-temporal maps. Cell Calcium. 91, 102260 (2020).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.