
Method Article
Microscopie confocale à balayage laser de la dynamique du calcium dans les tranches de tissu pancréatique aigu de souris
Dans cet article
Résumé
Nous présentons la préparation de tranches de tissu pancréatique aigu et leur utilisation en microscopie confocale à balayage laser pour étudier simultanément la dynamique du calcium dans un grand nombre de cellules vivantes, sur de longues périodes et avec une résolution spatio-temporelle élevée.
Résumé
La tranche aiguë de tissu pancréatique de souris est une préparation in situ unique avec une communication intercellulaire préservée et une architecture tissulaire qui implique beaucoup moins de changements induits par la préparation que les îlots isolés, les acini, les canaux ou les cellules dispersées décrits dans des études in vitro typiques. En combinant la tranche de tissu pancréatique aigu avec l’imagerie calcique à cellules vivantes en microscopie confocale à balayage laser (CLSM), les signaux calciques peuvent être étudiés simultanément dans un grand nombre de cellules endocriniennes et exocrines, avec une résolution unicellulaire ou même subcellulaire. La sensibilité permet la détection des changements et permet l’étude des ondes intercellulaires et de la connectivité fonctionnelle ainsi que l’étude de la dépendance des réponses physiologiques des cellules sur leur localisation dans l’îlot et la relation paracrine avec d’autres cellules. Enfin, du point de vue du bien-être animal, l’enregistrement des signaux provenant d’un grand nombre de cellules à la fois réduit le nombre d’animaux requis dans les expériences, contribuant ainsi au principe de remplacement, de réduction et de raffinement des 3R.
Introduction
Le pancréas des mammifères est une grande glande exocrine et endocrine. La partie exocrine représente 96 à 99% du volume total du pancréas et se compose d’acini et de conduits. La partie endocrinienne est composée d’un grand nombre d’îlots de Langerhans représentant les 1 à 4% restants du volume total du pancréas1. La partie exocrine sécrète des enzymes digestives majeures qui décomposent les polymères riches en énergie dans les aliments, ainsi qu’un liquide riche en bicarbonate, qui se combine avec d’autres sécrétions gastro-intestinales pour fournir un environnement propice à l’action des enzymes. La partie endocrinienne sécrète des hormones qui régulent la distribution postprandiale, le stockage et la libération interprandiale de nutriments riches en énergie. Bien que le tissu exocrine soit relativement sous-développé et que le système endocrinien soit relativement bien développé à la naissance, le premier dépasse rapidement le second au sevrage 2,3,4. Les premières études de la fonction pancréatique ont marqué la naissance de la physiologie moderne, et les avancées méthodologiques majeures dans le domaine ont été suivies par des ruptures scientifiques majeures5. Travailler avec le pancréas est techniquement difficile en raison de la structure complexe de la glande, mais c’est aussi une grande motivation en raison de maladies telles que le cancer du pancréas, la pancréatite et le diabète qui présentent des menaces majeures pour la santé publique et pour lesquelles de nouvelles approches thérapeutiques sont nécessaires.
Les îlotsisolés 6, acini 7,8 et les fragments canalaires avaient été développés et utilisés pendant des décennies comme méthodes de référence en raison de leurs avantages par rapport aux lignées cellulaires et aux cellules endocriniennes, acineuses et canalaires primaires dispersées 9,10. Malgré la fonction nettement améliorée des collectifs de cellules isolées, ces méthodes impliquent toujours un stress mécanique et enzymatique considérable, isolent les cellules du tissu environnant et manquent donc d’interactions paracrines et de soutien mécanique, et surtout, s’accompagnent de modifications significatives de la physiologie normale 11,12,13 . La tranche aiguë de tissu pancréatique de souris a été développée en 2001 à partir d’un besoin perçu de développer une plate-forme expérimentale similaire aux tranches cérébrales, hypophysaires et surrénales avec des contacts intercellulaires préservés, des interactions paracrines, du mésenchyme et de l’architecture tissulaire, ainsi que sans certaines des lacunes les plus importantes de la méthode de référence dans la recherche sur les îlots de cette époque - les îlots isolés12, 14. Parmi ces lacunes figurent les dommages aux couches les plus externes, le manque d’accessibilité des zones centrales des îlots et la nécessité d’une culture ayant des effets potentiellement importants sur l’identité et la physiologie cellulaires12,15. De plus, la méthode des tranches de tissu permet des études sur des modèles animaux avec une architecture d’îlots grossièrement dérangée où il est impossible d’isoler les îlots, ou lorsque le rendement des îlots est extrêmement faible par isolement traditionnel 16,17,18,19,20,21.
De plus, la tranche est plus adaptée à l’étude des changements morphologiques au cours du développement du diabète et de la pancréatite, par exemple, car elle permet une meilleure vue d’ensemble de l’ensemble du tissu et est également compatible avec l’étude des différences régionales. Il est important de noter que, malgré l’accent mis au début sur la partie endocrinienne, la méthode des tranches de tissu permet intrinsèquement l’étude des composants exocrines 9,22,23. Au cours de la première décennie après son introduction, la méthode a été utilisée pour des études électrophysiologiques de cellules bêta 14,24,25,26,27,28,29 et alpha 30,31 ainsi que pour examiner la maturation morphologique et fonctionnelle du pancréas 2,3 . Une décennie plus tard, en 2013, la méthode a été adaptée avec succès pour l’imagerie calcique des cellules calciques des cellules des îlots vivants à l’aide du CLSM pour caractériser leurs réponses au glucose32, leurs modèles de connectivité fonctionnelle33 et la relation entre le potentiel membranaire et le calcium intracellulaire en combinant un colorant de calcium fluorescent avec un colorant potentiel membranaire34. Plus tard dans la même année, la méthode a également été utilisée pour évaluer la dynamique du calcium dans les cellules acineuses22,35. Au cours des années suivantes, les tranches de tissu pancréatique ont été utilisées dans un certain nombre d’études différentes et adaptées avec succès aux tissus porcins et humains 9,36,37,38,39,40,41. Cependant, prises ensemble, l’imagerie calcique - dans les tranches de tissu pancréatique de souris en général et dans les îlots en particulier - est encore principalement effectuée par ce groupe. L’une des principales raisons à cela peut résider dans la combinaison d’une préparation de tranches de tissu techniquement difficile, de la nécessité d’un microscope confocal et d’une analyse de données plutôt complexe. L’objectif principal du présent document est de rendre cette méthode puissante plus accessible aux autres utilisateurs potentiels.
Il existe déjà d’excellents articles méthodologiques traitant en détail de la préparation des tranches de tissu et de l’utilisation des tranches pour les études structurelles et de sécrétion, mais pas pour l’imagerie confocaledu calcium 9,42,43. Par conséquent, cet article se concentre sur quelques conseils et astuces supplémentaires lors de la préparation des tranches, sur les étapes essentielles pour un chargement réussi de colorant, l’acquisition d’images, ainsi que sur les principales étapes de l’analyse de base des données calciques. Par conséquent, cette contribution devrait être considérée comme complémentaire à la méthode susmentionnée plutôt que comme une alternative à celle-ci. De même, l’imagerie calcique dans les tranches de tissu pancréatique de souris doit être considérée comme une approche expérimentale à utiliser pour répondre à des questions spécifiques et est donc complémentaire plutôt qu’une alternative absolue à d’autres approches d’imagerie calcique en physiologie pancréatique telles que les canaux isolés ou acini, les îlots isolés, les organoïdes, les îlots transplantés dans la chambre antérieure de l’œil, et enregistrements in vivo 11,44,45,46,47,48. La promesse de l’imagerie calcique dans les tranches de tissu pancréatique de souris est probablement mieux illustrée par les récents enregistrements réussis de la dynamique du calcium dans les cellules mésenchymateuses des îlots tels que les péricytes49 et les macrophages50, ainsi que dans les cellules canalaires23.
Protocole
NOTE: Toutes les expériences ont été réalisées en stricte conformité avec les directives institutionnelles pour le soin et l’utilisation des animaux dans la recherche. Le protocole a été approuvé par l’Administration de la République de Slovénie pour la sécurité alimentaire, le secteur vétérinaire et la protection des végétaux (numéro de permis: 34401-35-2018/2).
1. Préparation des tranches de tissu pancréatique
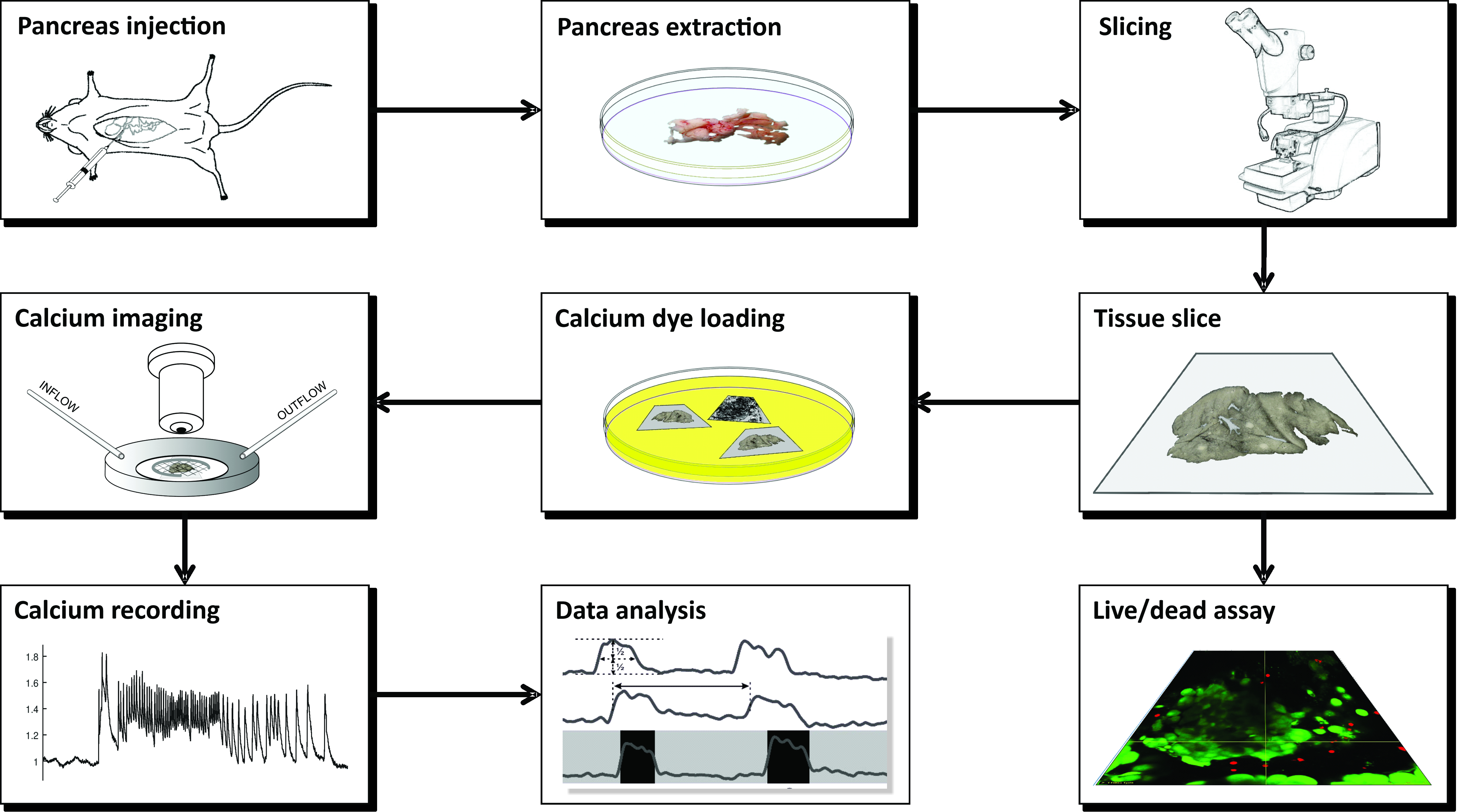
REMARQUE: La préparation de tranches de tissu pancréatique aigu de souris pour l’imagerie calcique à l’aide de CLSM nécessite un certain nombre d’instruments, de solutions différentes et se déroule dans une série d’étapes critiques qui sont schématiquement présentées à la figure 1 et décrites en détail ci-dessous.

Figure 1 : Diagramme de flux de travail. Représentation schématique de toutes les étapes du processus de préparation des tranches de tissu pancréatique, en commençant par l’injection d’agarose dans le canal biliaire commun, suivie de l’extraction du pancréas et du tranchage. Les tranches préparées peuvent être utilisées pour évaluer la viabilité du tissu avec un kit Live/Dead ou colorées avec un capteur de calcium. Une fois tachés, ils sont prêts pour l’imagerie. Les enregistrements obtenus à partir du processus d’imagerie sont ensuite utilisés pour l’analyse des données. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
- Préparation de solutions
REMARQUE: Toutes les solutions doivent être préparées à l’avance et peuvent être conservées au réfrigérateur à 4-8 ° C pendant un mois maximum. Pour la préparation et le stockage des tranches de tissu, environ 0,5 L de solution extracellulaire (SEC) avec 6 mM de glucose et 0,3 L de tampon d’acide 4-(2-hydroxyéthyl)-1-pipérazineéthanesulfonique (HEPES) est nécessaire. Pour 1 jour d’imagerie calcique avec le système de périfusion réglé à un débit de 1-2 mL / min, environ 0,5 L de SEC est nécessaire.- Solution extracellulaire avec 6 mM de glucose
- Préparer 1 L de SEC contenant 125 mM de NaCl, 26 mM de NaHCO3, 6 mM de glucose, 6 mM d’acide lactique, 3 mM de myo-inositol, 2,5 mM de KCl, 2 mM de na pyruvate, 2 mM de CaCl2, 1,25 mM de NaH2PO4, 1 mM deMgCl2 et 0,5 mM d’acide ascorbique. Bien mélanger jusqu’à ce que tous les ingrédients se dissolvent complètement. Prenez 50 μL de SEC dans un tube de microcentrifugation de 0,5 mL, placez-le sur l’osmomètre conformément aux instructions du fabricant et vérifiez l’osmolarité.
REMARQUE: L’osmolarité doit être de 300-320 mOsm. Pour la stimulation des cellules bêta, utilisez des solutions avec des concentrations de glucose plus élevées. Pour assurer une valeur de pH physiologique de 7,4 pendant le tranchage et les expériences, faites constamment buller le SEC avec du carbogène (c’est-à-dire un mélange gazeux de 95% O2 et 5% de CO2) à la pression barométrique. Un système de bouillonnement simple peut être mis en place en attachant une extrémité d’un tube en silicium de 5 mm à la source de carbogène (c’est-à-dire une bouteille de gaz sous pression) et l’autre extrémité du tube placé directement dans la bouteille contenant ECS. - Alternativement, préparez un bouillon 10x contenant 1250 mM de NaCl, 260 mM de NaHCO3, 30 mM de myo-inositol, 25 mM de KCl, 20 mM de pyruvate de Na, 12,5 mM de NaH2PO4 et 5 mM d’acide ascorbique. Lorsque le SEC contenant 6 mM de glucose est nécessaire, mélanger 100 mL du bouillon avec 2 mL de CaCl2 M, 1 mL de 1 MMgCl2, 0,455 mL d’acide lactique 13,2 M et 1,08 g de glucose, et remplir d’eau double distillée jusqu’à 1 L. Si nécessaire, utilisez différentes quantités de glucose pour obtenir d’autres concentrations de glucose.
- Préparer 1 L de SEC contenant 125 mM de NaCl, 26 mM de NaHCO3, 6 mM de glucose, 6 mM d’acide lactique, 3 mM de myo-inositol, 2,5 mM de KCl, 2 mM de na pyruvate, 2 mM de CaCl2, 1,25 mM de NaH2PO4, 1 mM deMgCl2 et 0,5 mM d’acide ascorbique. Bien mélanger jusqu’à ce que tous les ingrédients se dissolvent complètement. Prenez 50 μL de SEC dans un tube de microcentrifugation de 0,5 mL, placez-le sur l’osmomètre conformément aux instructions du fabricant et vérifiez l’osmolarité.
- Tampon HEPES avec 6 mM de glucose
- Préparer 0,5 L de solution tamponnée HEPES (HBS) contenant 150 mM de NaCl, 10 mM de HEPES, 6 mM de glucose, 5 mM de KCl, 2 mM de CaCl2 et 1 mM deMgCl2; titrer au pH = 7,4 avec 1 M de NaOH.
REMARQUE: Si le carbogène n’est pas disponible, ce tampon peut être utilisé pour toutes les étapes au lieu de ECS.
- Préparer 0,5 L de solution tamponnée HEPES (HBS) contenant 150 mM de NaCl, 10 mM de HEPES, 6 mM de glucose, 5 mM de KCl, 2 mM de CaCl2 et 1 mM deMgCl2; titrer au pH = 7,4 avec 1 M de NaOH.
- Agarose (1,9 % p/p)
- Préchauffer un bain-marie à 40 °C.
- Ajouter 0,475 g d’agarose à point de fusion bas et 25 mL d’ECS contenant 6 mM de glucose dans une fiole d’Erlenmeyer et placer la fiole dans un four à micro-ondes à puissance maximale pendant quelques secondes jusqu’à ce qu’elle commence à bouillir. Sortez la fiole du four et faites-la tourbillonner plusieurs fois, jusqu’à ce que l’agarose se dissolve complètement. Transférer la fiole avec l’agarose liquide au bain-marie préavertissé à 40 °C pour refroidir l’agarose à la température souhaitée et la garder liquide jusqu’à l’injection. Fixez la fiole à l’aide d’un anneau de plomb stabilisateur.
REMARQUE: L’agarose peut être préparée à l’avance et conservée au réfrigérateur. Avant utilisation, réchauffer l’agarose au four à micro-ondes jusqu’à ce qu’elle se liquéfie et transférer la fiole d’Erlenmeyer dans un bain-marie préavertissé à 40 °C. L’agarose peut être réutilisé jusqu’à 5x. S’il est réutilisé au-delà de 5x, il deviendra dense et plus difficile à injecter.
- Solution extracellulaire avec 6 mM de glucose
- Injection de pancréas avec de l’agarose
NOTE : Les sections 1.2 et 1.3 expliquent la préparation de tranches de tissu qui peuvent être utilisées à différentes fins expérimentales telles que l’imagerie calcique, l’électrophysiologie, l’immunohistochimie, les études de sécrétion et les études structurelles / microanatomiques.- Remplissez une seringue de 5 mL avec l’agarose liquide de la fiole d’Erlenmeyer dans le bain-marie de l’étape 1.1.3.2, retirez les bulles et montez une aiguille de 30 G. Protégez l’aiguille avec un capuchon et gardez la seringue remplie dans le bain-marie avec l’aiguille tournée vers le bas et tout le volume d’agarose sous la surface de l’eau. Fixez la seringue avec un anneau de plomb stabilisateur de manière à ce que l’anneau appuie la seringue contre la paroi du bain-marie.
REMARQUE: Veillez à ne pas pousser d’agarose dans l’aiguille car elle durcira rapidement et bloquera l’aiguille. Si la température ambiante est basse et si l’injection est effectuée par une personne moins expérimentée, augmentez la température du bain-marie jusqu’à 42 ° C pour gagner du temps supplémentaire pour l’injection. - Remplissez un seau à glace avec de la glace et placez-y la bouteille contenant du SEC. Faites bouillonner le SEC en permanence à 1,5 mL/min avec du carbogène à pression barométrique et à température ambiante pour assurer l’oxygénation et un pH de 7,4.
- Sacrifiez une souris en administrant une forte concentration de CO2 suivie d’une luxation cervicale. Faites tous les efforts pour minimiser la souffrance animale.
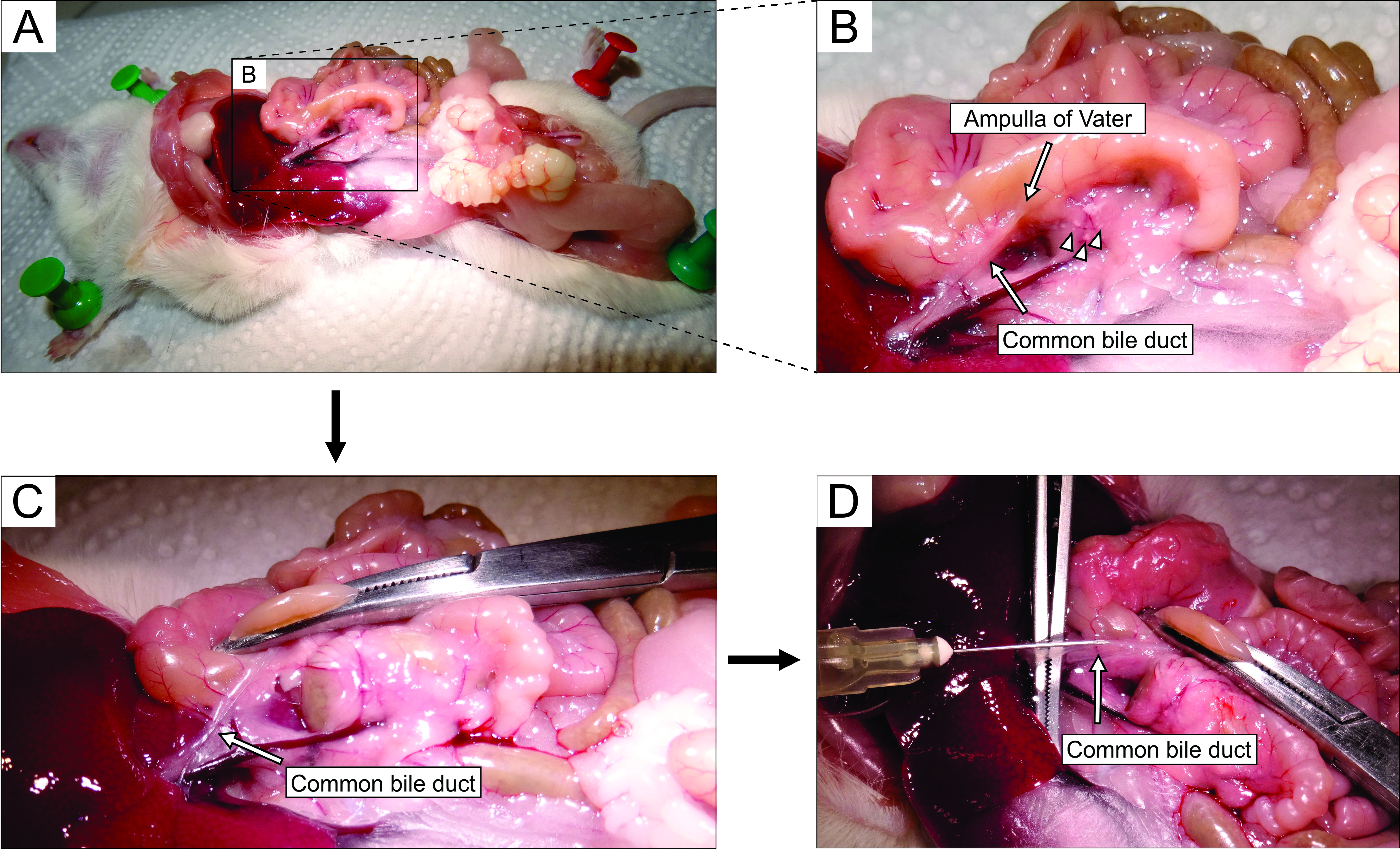
- En travaillant sous stéréomicroscope, accédez à l’abdomen par laparotomie (Figure 2A). Retournez doucement l’intestin vers le côté gauche de la souris (du point de vue anatomique de la souris) pour exposer le canal biliaire commun. Utilisez des pinces pour soulever légèrement la partie duodénale et trouvez la papille-papille duodénale majeure de Vater. Serrez le canal biliaire commun au niveau de la papille duodénale à l’aide d’un hémostat (figure 2B, C) pour éviter les fuites d’agarose du canal dans le duodénum.
REMARQUE: Pour éviter les fuites d’agarose dans le duodénum et plus haut et bas dans le tractus gastro-intestinal, placez l’hémostat de manière à ce qu’il serre également le duodénum à la fois proximalement et distalement de la papille. Il est préférable d’utiliser un hémostat incurvé à cette fin. - Avec de petites pinces pointues, atteignez sous le canal biliaire commun et brisez la membrane qui attache le canal au tissu pancréatique. Pour un meilleur contrôle visuel et une injection plus facile, éliminez autant de graisse et de tissu conjonctif que possible du canal.
- Placez le conduit perpendiculairement sur de grandes pinces (figure 2D) et injectez l’agarose liquide préparée dans la partie proximale du canal biliaire commun (figure 2D). Assurez-vous de presser la seringue fort car l’agarose est visqueuse. Continuez à remplir le pancréas jusqu’à ce qu’il devienne blanchâtre et légèrement distendu ou pendant au moins 20-30 s.
REMARQUE: Il s’agit de l’étape la plus critique dans la préparation des tranches. S’il y a des plis dans l’arbre canalaire du pancréas, soulevez doucement ou éloignez le pancréas de la seringue pour les niveler. Ne décidez pas quand arrêter l’injection en fonction du volume injecté à partir de la seringue, car le refoulement au point d’injection et la fuite vers l’avant dans le duodénum sont généralement beaucoup plus élevés que le volume injecté dans l’arbre canalaire du pancréas. Il est important de noter que les injections réussies peuvent être effectuées avec des changements pratiquement imperceptibles du volume de la seringue. - Retirez la seringue et versez lentement 20 mL du SEC glacé à bulles à 0-4 °C du flacon sur le pancréas pour refroidir le tissu et durcir l’agarose.
- Extrayez doucement le pancréas à l’aide de pinces et de ciseaux fins et durs. Placez le pancréas extrait dans une boîte de Petri de 100 mm contenant environ 40 mL de SEC glacé et déplacez-le doucement pour le laver. Transférer le pancréas dans une boîte de Petri fraîche de 100 mm contenant environ 40 mL de SEC glacé.
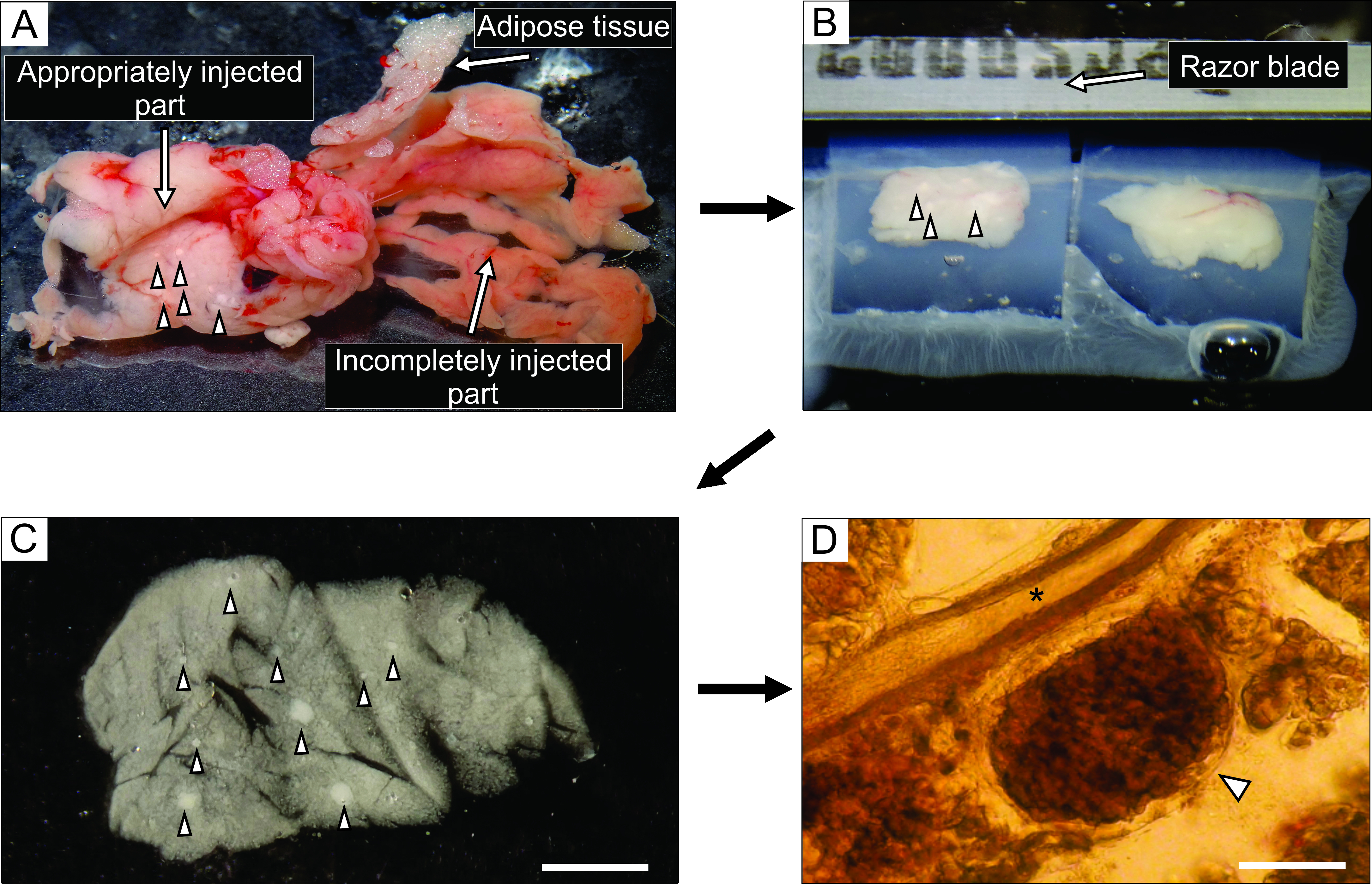
- À partir de la partie bien injectée du pancréas, qui semble blanchâtre (Figure 3A), coupez jusqu’à 6 blocs de tissu, de 0,1 à 0,2 cm3 , à l’aide d’une pince et de ciseaux coupés durement. Débarrassez-les de tout tissu conjonctif et adipeux.
- Remplissez une boîte de Petri inférieure non collante de 35 mm avec environ 5 mL d’agarose liquide à 40 °C, transférez-y les blocs de tissu et mettez immédiatement la boîte de Petri sur de la glace pour la refroidir et durcir l’agarose.
REMARQUE: La façon dont les blocs du pancréas sont piégés dans l’agarose détermine la façon dont ils sont coupés pendant le tranchage. Les expérimentateurs expérimentés peuvent essayer d’affiner la position des blocs pendant les quelques instants avant que l’agarose ne durcisse lorsqu’elle est placée sur la glace. - Une fois que l’agarose avec les blocs de tissu durcit, tournez la boîte de Petri à l’envers sur une surface plane et lisse telle que le couvercle d’une boîte de Petri de 100 mm et retirez l’agarose en coupant doucement avec la moitié d’une lame de rasoir dans la marge entre la paroi latérale de la boîte de Petri et l’agarose. Avec une lame de rasoir, coupez des cubes d’agarose individuels, chacun contenant un bloc de tissu, en veillant à ce que chaque bloc de tissu soit entouré d’agarose. Collez les blocs d’agarose sur la plaque d’échantillonnage du vibratome avec de la colle cyanoacrylate (Figure 3B).
- Remplissez une seringue de 5 mL avec l’agarose liquide de la fiole d’Erlenmeyer dans le bain-marie de l’étape 1.1.3.2, retirez les bulles et montez une aiguille de 30 G. Protégez l’aiguille avec un capuchon et gardez la seringue remplie dans le bain-marie avec l’aiguille tournée vers le bas et tout le volume d’agarose sous la surface de l’eau. Fixez la seringue avec un anneau de plomb stabilisateur de manière à ce que l’anneau appuie la seringue contre la paroi du bain-marie.

Figure 2 : Injection d’agarose dans le canal biliaire commun. (A) Ouvrez la cavité abdominale et exposez les organes de la cavité péritonéale. (B) La partie agrandie de la zone délimitée par le rectangle dans le panneau A. La tache blanche sur le duodénum (indiquée par la flèche) indique l’ampoule de Vater. Les îlots de Langerhans sont désignés par des pointes de flèches. (C) Serrez l’ampoule de Vater par un hémostat incurvé et soulevez-la légèrement pour exposer et étirer doucement le canal biliaire commun (flèche). D) Canulation du canal biliaire commun et injection d’une solution d’agarose à 1,9 % à l’aide d’une seringue de 5 mL et d’une aiguille de 30 G. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
- Trancher
- Remplissez la chambre de coupe du vibratome avec ~ 0,15 L de SEC glacé et faites-buller constamment avec du carbogène. Entourez la chambre de coupe de glace et ajoutez 2 glaçons (~ 10 mL chacun) en SEC avec 6 mM de glucose dans la chambre de coupe. Montez la lame de rasoir pour couper sur le vibratome et fixez la plaque d’échantillonnage avec des blocs d’agarose à sa place.
- Réglez la trancheuse pour couper des blocs d’agarose à 0,05 à 1 mm/s et 70 Hz en tranches de 140 μm d’épaisseur d’une surface de 20 à 100 mm2. Pour les paramètres de la trancheuse, suivez les instructions du fabricant.
- Immédiatement après chaque étape de coupe, mettez la trancheuse en pause, ramassez doucement les tranches avec un pinceau fin et transférez-les dans une boîte de Petri de 100 mm remplie de 40 mL de tampon HEPES avec 6 mM de glucose à température ambiante (Figure 3C).
REMARQUE: Les tranches peuvent être conservées dans un tampon HEPES à température ambiante pendant au moins 12 h, et le tampon doit être échangé toutes les 2 h.

Figure 3 : Préparation et tranchage du tissu pancréatique. (A) Le pancréas de souris extrait après injection d’agarose. Le tissu blanc à gauche indique une partie bien injectée (partie duodénale), tandis que la partie plus rougeâtre à droite montre la partie insuffisamment injectée du pancréas (partie splénique). (B) Tranchage vibratome de deux blocs de tissu pancréatique incorporés dans l’agarose. (C) Tranche de tissu pancréatique aigu avec îlots de Langerhans indiqués par des pointes de flèches. Barre d’échelle = 3000 μm. (D) Tranche aiguë de tissu pancréatique au microscope optique avec l’îlot de Langerhans indiqué par une pointe de flèche, l’astérisque indique un canal pancréatique. Barre d’échelle = 100 μm. Veuillez cliquer ici pour afficher une version agrandie de cette figure.
{kind=link}
2. Essai vivant/mort à l’aide du kit de viabilité/cytotoxicité VIVANT/MORT pour les cellules de mammifères
REMARQUE: Pour certaines expériences, il est utile de vérifier la viabilité des cellules dans les tranches (Figure 4) par le test vivant/ mort comme suit.
- Suivez les instructions du fabricant pour décongeler les flacons avec les réactifs du kit de viabilité / cytotoxicité LIVE / DEAD et préparez des solutions de travail de calcéine AM juste avant utilisation. Utilisez les solutions en une journée.
- Dans un tube centrifuge de 15 mL, mélanger 5 μL de 4 mM de calccéine AM (composant A), 20 μL d’homodimère d’éthidium homodimère-1 de 2 mM (EthD-1, composant B) et 10 mL de solution saline tamponnée au phosphate (D-PBS) de Dulbecco pour préparer une solution de travail contenant environ 2 μM de calcéine AM et 4 μM d’EthD-1. Vortex à fond.
- À l’aide d’un pinceau fin, transférez délicatement les tranches de tissu dans une boîte de Petri de 3 mL avec un tampon HEPES frais pour diluer l’activité sérique de l’estérase. Retirez le tampon HEPES et couvrez les tranches de 100 à 200 μL (ou plus si nécessaire) de la solution de travail à partir de l’étape 2.2.
- Incuber les tranches pendant 30-45 min à température ambiante dans une boîte de Petri fermée. Imagez les tranches de tissu à l’aide de filtres d’excitation/émission recommandés par le fabricant.
3. Charge en colorant de calcium
REMARQUE: Les colorants fluorescents doivent être protégés de l’exposition à la lumière pendant tout le processus de préparation et de chargement du colorant, ainsi que lors de la manipulation des tranches de tissu colorées. Le papier d’étain peut être utilisé pour recouvrir des tubes ou des boîtes de Petri contenant le colorant calcique.
- Préparation de colorant
- Dissoudre le contenu d’un flacon (50 μg) du colorant indicateur Ca2+ perméable à la cellule (excitation/émission 495/523 nm; voir la table des matériaux), 7,5 μL de diméthylsulfoxyde (DMSO) et 2,5 μL de polaxamer (solution à 20 % dans du DMSO; Tableau des matériaux) dans 6,667 mL de HBS contenant 6 mM de glucose dans un tube à bouchon à vis de 15 mL.
REMARQUE: Cette solution finale contient 6 μM du colorant indicateur Ca2+ , 0,11% de DMSO et 0,037% de polaxamer. - Aspirer et expulser la solution dans le tube du bouchon à vis à plusieurs reprises avec une pipette pendant 20 s; immerger le tube dans une chambre de bain à ultrasons pendant 30 s et vortex pendant 30 s pour améliorer la solubilisation. Aliquote 3,333 mL de la solution finale de colorant indicateur Ca2+ préparée à l’étape 3.1.1 dans des boîtes de Petri de 5 mL.
- Dissoudre le contenu d’un flacon (50 μg) du colorant indicateur Ca2+ perméable à la cellule (excitation/émission 495/523 nm; voir la table des matériaux), 7,5 μL de diméthylsulfoxyde (DMSO) et 2,5 μL de polaxamer (solution à 20 % dans du DMSO; Tableau des matériaux) dans 6,667 mL de HBS contenant 6 mM de glucose dans un tube à bouchon à vis de 15 mL.
- Chargement de colorant
- Transférer les tranches de tissu préparées de la boîte de Petri de 60 mL avec HBS dans des boîtes de Petri de 5 mL remplies de la solution de colorant en soulevant doucement chaque tranche de tissu à l’aide d’un pinceau mince et doux et en la plaçant dans la solution de colorant. Incuber jusqu’à 10 tranches de tissu par boîte de Pétri.
- Placez la boîte de Petri chargée en tranches sur un agitateur orbital à température ambiante réglé sur le mouvement orbital à 40 tours par minute pendant 50 minutes. Incuber les tranches dans la solution de colorant exposée à l’air ambiant à température ambiante, mais à l’abri de la lumière en recouvrant la boîte de Pétri d’une feuille d’étain.
- Stockage des tranches
- Transférer les tranches de tissu coloré de la boîte de Petri de 5 mL dans une boîte de Petri de 60 mL remplie de HBS sans colorant en les soulevant doucement à l’aide d’un pinceau fin et doux. Conservez jusqu’à 20 tranches par boîte de Pétri.
REMARQUE: Utilisez les tranches de tissu pour l’imagerie à ce stade. Les tranches de tissu retiendront le colorant indicateur Ca2+ pendant plusieurs heures. La survie des tranches et la rétention du colorant peuvent être améliorées en plaçant la boîte de Petri dans un récipient isolé, entouré de glace. Ceci est particulièrement important si les tranches chargées de colorant doivent être transportées. De plus, échangez le HBS toutes les 2 h.
- Transférer les tranches de tissu coloré de la boîte de Petri de 5 mL dans une boîte de Petri de 60 mL remplie de HBS sans colorant en les soulevant doucement à l’aide d’un pinceau fin et doux. Conservez jusqu’à 20 tranches par boîte de Pétri.
4. Imagerie calcique
- Installation du microscope confocal
- Choisissez un grossissement objectif approprié en fonction de l’intérêt de l’étude. Sélectionnez 20x et 25x (ouverture numérique [NA] 0,77-1,00) pour visualiser un îlot entier, plusieurs acini simultanément ou des conduits plus grands. Sélectionnez des grossissements plus élevés pour étudier la dynamique intracellulaire.
- Choisissez le mode d’acquisition pour l’imagerie time-lapse (par exemple, time-lapse, xyt ou mode similaire). Réglez le sténopé sur 100-200 μm.
- Définissez le trajet de la lumière pour les fluorophores verts: excitation à 488 nm et collecte des émissions à 500-700 nm. Choisissez de préférence des détecteurs à haute efficacité quantique (par exemple, le phosphure d’arséniure de gallium) plutôt que des détecteurs photomultiplicateurs.
- Configuration de la chambre d’enregistrement et du système de périfusion
- Montez la chambre d’enregistrement sur l’étage à température contrôlée du microscope et du système de périfusion (configuration par gravité ou par pompe péristaltique, volume 1 mL). Placez l’entrée et la sortie sur les bords éloignés de la chambre d’enregistrement pour éviter les méandres du perfusat dans la chambre et réglez l’entrée et la sortie à des valeurs égales (1-2 mL / min). Évitez la dérive de la hauteur du ménisque liquide et des gouttelettes dans le périfusat.
- Réglez le contrôle de la température du système de périfusion à 37 °C.Initiez la périfusion avec la solution non stimulante et préparez les solutions stimulantes. Changez les solutions via des vannes motorisées ou en changeant manuellement les solutions qui alimentent le système de périfusion.
- Enregistrer la dynamique du calcium
- Transférer une seule tranche de tissu dans la chambre d’enregistrement. Immobilisez la tranche de tissu avec un poids en platine en forme de U avec une maille de nylon tendue (par exemple, à partir de bas en nylon). Évitez de positionner des fils de nylon sur la structure qui vous intéresse.
- Localisez un îlot/acinus/conduit à l’aide de l’option de champ lumineux. Exécutez l’imagerie en direct pour positionner les structures étudiées dans le champ de vision et configurez les paramètres d’imagerie. Optimisez le rapport signal/bruit en ajustant la puissance du laser, l’amplification du détecteur et la moyenne/binning de ligne pour permettre la visualisation des cellules tout en maintenant la puissance laser minimale.
- Ajustez le plan focal de l’enregistrement à environ 15 μm sous la surface coupée (Figure 5) pour éviter l’enregistrement à partir de cellules potentiellement endommagées sur la surface coupée.
- Acquérir des images. Réglez la fréquence d’échantillonnage sur 1-2 Hz pour détecter les oscillations individuelles initialement, et utilisez un scanner résonant capable de faire la moyenne rapide des lignes (8-20) à un taux d’acquisition plus élevé (>10 Hz) pour enregistrer l’activité intracellulaire Ca2+ ([Ca2+]IC). Prévoir un intervalle (p. ex., 30 % du temps d’échantillonnage total) entre les éclairements ponctuels consécutifs pour prévenir la phototoxicité. Enregistrez une image haute résolution (p. ex., 1024 x 1024 pixels, une moyenne de lignes > 50) avant l’acquisition de la série chronologique (voir la section 5).
REMARQUE: La fréquence d’échantillonnage de 1-2 Hz est inférieure au critère de Nyquist pour la fréquence d’acquisition pour la plupart des cellules, et la forme du signal sera sous-échantillonnée par défaut. - Reportez-vous à un tableau en ligne, s’il est disponible dans le logiciel d’imagerie, pour obtenir un retour instantané sur la réponse de préparation, le suréclairage, le photoblanchiment et la dérive mécanique. En cas de taux élevé de blanchiment lors de l’acquisition, arrêtez l’enregistrement et diminuez la puissance du laser tout en augmentant le gain du détecteur pour maintenir le rapport signal/bruit. En cas de dérive mécanique, vérifiez la tension entre les tubes/câbles et l’étage du microscope ainsi que les fuites de liquide ou les changements de volume dans la chambre d’enregistrement. Si vous le souhaitez, essayez de corriger manuellement la dérive lors de l’acquisition; cependant, notez que cela donnera intrinsèquement des résultats limités.
REMARQUE: Les cellules endocriniennes sont très hétérogènes à des concentrations proches du seuil. Une durée suffisante de stimulation est nécessaire pour détecter la gamme des retards d’activation/désactivation dans les cellules individuelles. Ceci est particulièrement important pour la détection précise des réponses off-réponses suivant des protocoles hautement stimulateurs. - Utilisez l’imagerie calcique pour discriminer fonctionnellement entre les cellules endo- et exocrines (Figure 6). Pour enregistrer l’activité transitoire pendant l’activation et la désactivation, appliquez des stimuli sans arrêter l’enregistrement.
- Enregistrez les données après la fin de l’expérimentation (envisagez d’utiliser une fonction d’enregistrement automatique). Prévoyez une période de refroidissement avant d’éteindre l’alimentation du laser afin de ne pas endommager les lasers pendant la procédure d’arrêt.
5. Analyse des données
- Inspectez visuellement l’enregistrement qualitativement en relisant la vidéo time-lapse. Vérifiez les dérives des cellules du champ de vision ou du plan optique. Si une dérive dans le plan optique s’est produite, utilisez le plugin de correction de dérive dans ImageJ.
- Sélectionnez les régions d’intérêt (ROI) à l’aide d’un logiciel de microscope ou d’un logiciel tiers. Utilisez l’image haute résolution, la projection maximale ou la moyenne d’image comme référence pour sélectionner les retours sur investissement. Relisez l’imagerie time-lapse pour visualiser les cellules répondantes qui ne sont pas visibles dans les images de référence. Positionnez les ROI de manière à ce que la zone sélectionnée d’un ROI ne se chevauche pas avec les cellules voisines pour éviter la diaphonie du signal entre les ROI.
- Exportez les données de la série chronologique en tant que valeur moyenne du retour sur investissement par image. Exporter les coordonnées du retour sur investissement.
- Corriger les données de série chronologique pour le blanchiment (figure 7A) en utilisant une combinaison d’un ajustement exponentiel et linéaire, comme décrit par
 (1)
(1)
où x(t) désigne le signal de fluorescence à un point temporel t; xcorr(t) le signal corrigé aux points temporels correspondants; et a, b et c les paramètres de l’ajustement calculés comme la plus petite somme des carrés entre le corr(t) et x(t). - Analyser la phase d’activation et de désactivation de la réponse (Figure 7B). Calculez la première dérivée des données de la série chronologique et déterminez le zénith et le nadir de la dérivée correspondant respectivement à l’activation et à la désactivation. Vous pouvez également sélectionner manuellement le début de l’augmentation phasique. Enregistrez et exportez les temps d’activation/désactivation et les coordonnées de cellule correspondantes.
- Analyser la phase de plateau (Figure 7C). Détectez les oscillations individuelles en seuillant les données brutes ou en seuillant la première dérivée des données de séries chronologiques. Définissez le début et la fin d’une oscillation individuelle comme le temps correspondant à la demi-amplitude de l’oscillation.
- Calculer la durée et la fréquence des oscillations individuelles pour chaque cellule. Calculez la valeur inverse de l’intervalle interspike (adapté aux modèles d’activité réguliers). Alternativement, divisez le nombre d’oscillations par l’intervalle de temps de l’enregistrement (adapté aux modèles d’activité irréguliers).
- Calculez le temps actif. Exprimez le temps actif comme la somme des durées et divisez cette valeur par l’intervalle de temps. Alternativement, multipliez la fréquence et la durée qui correspondent à une oscillation.
REMARQUE: La division de la somme des durées par l’intervalle de temps fournit des résultats robustes, mais présente une faible discrimination statistique car un seul point de données par cellule est obtenu. La multiplication de la fréquence et de la durée d’une oscillation fournit une résolution temporelle d’oscillation à oscillation.
Résultats
L’injection de la solution d’agarose dans le canal pancréatique est l’étape la plus critique de la préparation des tranches de tissu pancréatique. Une injection réussie peut être reconnue par un blanchiment du tissu pancréatique, comme on le voit sur le côté gauche de la figure 3A, tandis qu’une partie incomplètement injectée du pancréas est présentée sur le côté droit de la figure 3A. Les îlots de Langerhans peuvent être reconnus à l’œil nu ou sous un stéréomicroscope, ce qui aide à couper les parties appropriées du pancréas pour une intégration ultérieure dans des blocs d’agarose (Figure 3B). Dans une tranche de tissu pancréatique de souris fraîchement coupée, les îlots de Langerhans peuvent être facilement distingués du tissu exocrine et du mésenchyme environnants sous forme de taches blanches sous le stéréomicroscope (Figure 3C) ou de structures brunâtres sous le microscope optique (Figure 3D). Les tranches de tissu pancréatique peuvent être utilisées pour des types d’expériences distincts pendant au moins 12 heures après le tranchage. En plus de l’évaluation morphologique globale sous stéréomicroscope, au microscope optique et des réponses fonctionnelles des cellules lors de l’imagerie calcique, la viabilité des tranches de tissu pancréatique peut être évaluée (figure 4).

Figure 4 : Viabilité des cellules à l’intérieur de la tranche de tissu. La viabilité des cellules a été déterminée à l’aide du test Vivant/Mort. Les cellules vivantes sont colorées par Calcein AM (en vert), tandis que les cellules mortes sont colorées avec de l’homodimère-1 à l’éthidium (en rouge). Les lignes jaunes indiquent la position de la section X-Y de la pile Z affichée en bas et à droite. La profondeur totale de la pile Z est de 88 μm. Veuillez cliquer ici pour voir une version plus grande de cette figure.
{kind=link}
Pour les expériences d’imagerie calcique, l’indicateur de calcium fluorescent doit pénétrer à travers quelques couches de cellules. La figure 5A présente le chargement réussi du colorant indicateur Ca2+ perméable aux cellules dans la tranche de tissu pancréatique dans laquelle des îlots individuels et des cellules acineuses peuvent être reconnus. En revanche, les tranches de la figure 5B-D ne sont pas optimales en raison de la pénétration infructueuse du colorant (figure 5B), de l’absence de cellules d’îlots (figure 5C) et de beaucoup de tissu nécrotique à la surface (figure 5D). Ces tranches peuvent être jetées, vérifiées pour la présence d’îlots supplémentaires qui sont mieux coupés ou colorés (voir le tableau 1 pour le dépannage), ou utilisées pour enregistrer les réponses des cellules exocrines.

Figure 5: Exemples de préparations utilisables et inutilisables. (A) Exemple d’une préparation réussie de la tranche de tissu pancréatique avec des cellules bien colorées dans les îlots de Langerhans, ainsi que des cellules canalaires et du tissu acineux environnant. (B) Un exemple de tranche de tissu mal colorée. (C) Exemple d’îlot de Langerhans avec des arrêts structurels. (D) Un exemple d’îlot de Langerhans contenant de nombreuses cellules mortes et beaucoup de débris. La table de recherche « glow-over, glow-under » à droite affiche l’intensité 0 en vert et la saturation en bleu. Barre d’échelle = 100 μm. Veuillez cliquer ici pour afficher une version agrandie de cette figure.
{kind=link}
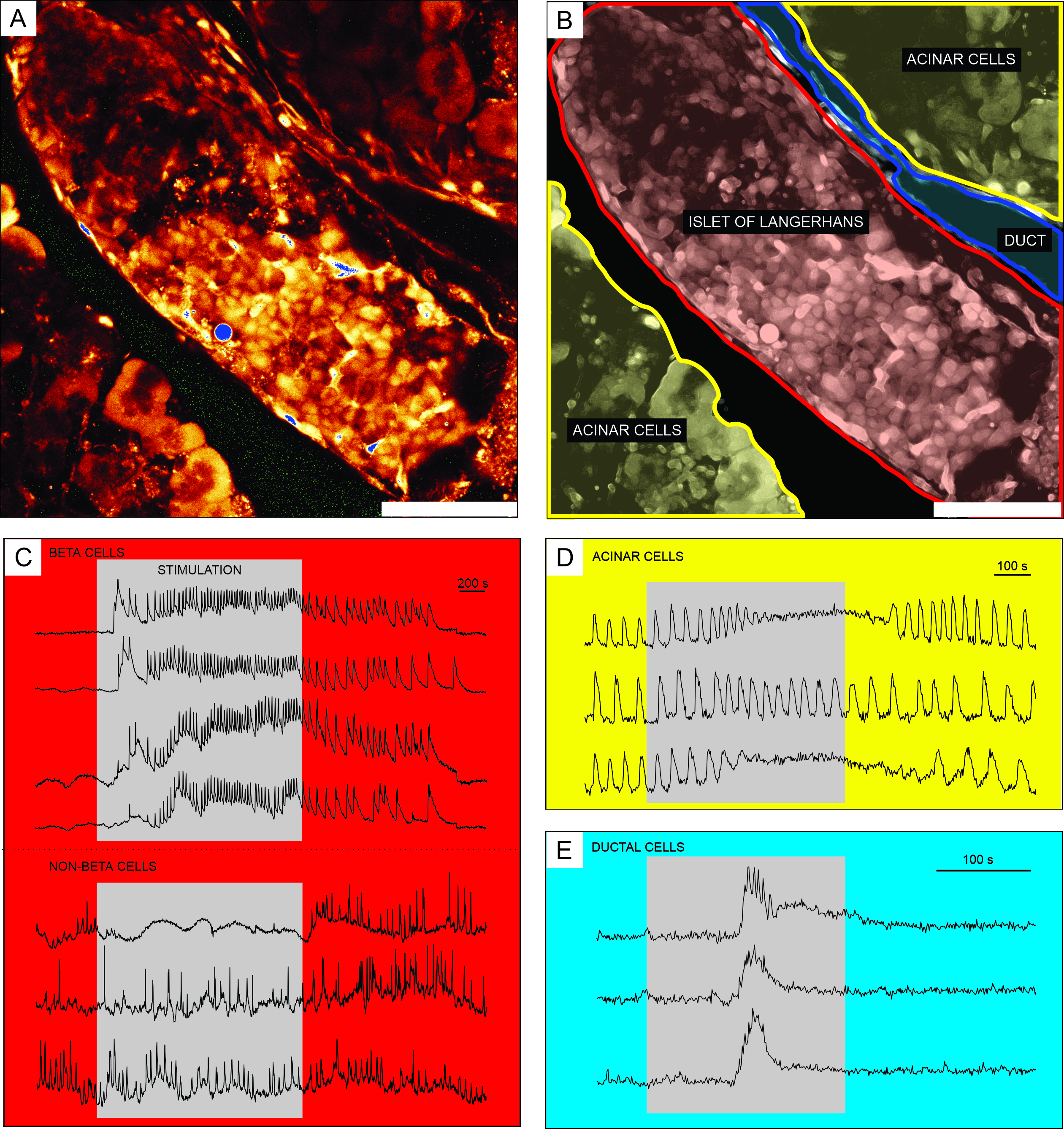
Les résultats représentatifs de l’imagerie calcique à l’aide du colorant indicateur Ca2+ perméable aux cellules sont présentés à la figure 6. Dans la figure 6A, une image à haute résolution d’une tranche de tissu pancréatique est présentée, contenant un îlot de Langerhans, un tissu acineux et un canal pancréatique. Pour une meilleure distinction, les parties endocrinienne, exocrine et canalaire de la tranche de tissu pancréatique présentée à la figure 6A sont colorées à la figure 6B. L’utilisation de stimuli appropriés peut discriminer fonctionnellement entre différentes cellules d’îlots, ou cellules d’îlots et non-îlots51. Les cellules bêta répondent généralement à une stimulation de l’impulsion carrée par le glucose avec une augmentation transitoire de [Ca2+]IC suivie d’oscillations rapides du calcium sur un plateau soutenu (Figure 6C, panneau supérieur).
Comme toutes les cellules bêta sont couplées en un seul grand syncytium fonctionnel, ces oscillations sont également très bien synchronisées entre différentes cellules au moyen de la propagation des ondes [Ca2+]IC 32,34,52,53,54 (Figure 7C). Des oscillations plus lentes [Ca2+]IC avec une période de 5 à 15 minutes peuvent sous-tendre les oscillations rapides ou même être le type prédominant de réponse55,56. Le même protocole simple peut révéler d’autres types de réponses, en particulier à la périphérie des îlots (Figure 6C, panneau inférieur). Comme ces cellules ne sont pas synchronisées avec les cellules bêta et répondent avec des oscillations plus rapides et plus irrégulières qui sont déjà présentes dans des conditions de faible glycémie ou avec une diminution de l’activité, de telles réponses sont très suggestives des cellules non bêta 21,32,57,58. Cependant, leur caractérisation fonctionnelle définitive nécessite des protocoles plus complexes avec des étapes de stimulation supplémentaires ou des approches alternatives, qui sont discutées ci-dessous. Les réponses typiques des cellules acineuses et canalaires sont présentées dans les figures 6D et 6E, respectivement. Reportez-vous à la littérature pour plus de détails sur les cellules acineuses et canalaires 22,23,35.

Figure 6: Résultats représentatifs de la dynamique du calcium dans des types distincts de cellules pancréatiques. (A) Image à haute résolution d’un îlot de Langerhans avec les tissus environnants. Barre d’échelle = 100 μm. (B) Délimitation de parties distinctes du tissu pancréatique avec un tissu acineux représenté en jaune, un îlot de Langerhans en rouge et un segment de l’arbre canalaire en bleu. Barre d’échelle = 100 μm. (C) Traces typiques de dynamique du calcium dans les cellules bêta et putatives non bêta pendant la stimulation avec 12 mM de glucose; 3 mM de glucose a été utilisé pour des conditions non stimulantes. Les protocoles qui peuvent être utilisés pour une discrimination plus spécifique des cellules non bêta sont décrits dans la section de discussion. (D) Une trace typique de la dynamique calcique des cellules acineuses stimulée par 25 nM d’acétylcholine. (E) Une trace typique de dynamique calcique des cellules canalaires stimulée par 1 mM d’acide chénodésoxycholique. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
Après une imagerie calcique réussie, les données sont d’abord exportées et corrigées pour le blanchiment par une combinaison d’un ajustement exponentiel et linéaire, comme décrit dans la section du protocole. Une série chronologique avant et après la correction du blanchiment est présentée à la figure 7A. Par la suite, plusieurs paramètres de la phase d’activation et de désactivation de la réponse ainsi que de la phase de plateau peuvent être analysés. Un retard dans l’apparition de l’augmentation de [Ca2+]IC après la stimulation peut être mesuré tel que représenté par le retardA dans la figure 7B et l’hétérogénéité des retards entre les cellules individuelles (retardA1). Les mêmes paramètres (délaiD et retardD1) peuvent être utilisés pour décrire la phase de désactivation. Après l’augmentation transitoire initiale [Ca2+]IC , la phase de plateau dans la plupart des cellules bêta pancréatiques d’un îlot est caractérisée par des oscillations relativement régulières à haute fréquence [Ca2+]IC . La phase de plateau peut être décrite en analysant les paramètres fonctionnels classiques. La présentation schématique de la durée, de la fréquence et du pourcentage de temps actif des oscillations [Ca2+]IC est présentée à la figure 7C. En imagerie calcique avec des taux d’acquisition supérieurs à 10 Hz, les ondes de calcium se propageant à plusieurs reprises sur l’îlot peuvent également être clairement reconnues (Figure 7C).

Figure 7: Analyse des données des séries chronologiques. (A) Correction des données des séries chronologiques pour le photoblanchiment. (B) Analyse des retards d’activation après stimulation et de désactivation après arrêt de la stimulation avec 12 mM de glucose. La durée de la stimulation est indiquée par la barre gris clair et ombragée de l’image. (C) Analyse de plusieurs paramètres de la phase de plateau: I) Durée de l’oscillation déterminée à mi-hauteur, II) fréquence des oscillations déterminée par des intervalles d’oscillation inter-oscillation. III) le temps actif en tant que produit de la fréquence et de la durée des oscillations. I-IV) Délais entre les oscillations dans une onde donnée d’oscillations qui se propagent à travers l’îlot de Langerhans déterminés par les retards (Δt) dans le temps auquel une seule cellule atteint la moitié de la hauteur de l’oscillation. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
Tableau 1. Veuillez cliquer ici pour télécharger ce tableau.
Discussion
La méthode de la tranche de tissu pancréatique est une méthode expérimentale rapide pour étudier la morphologie et la physiologie des parties endocrinienne et exocrine du pancréas dans une préparation in situ plus conservée. Bon nombre des avantages ont déjà été soulignés dans l’introduction. Il convient de souligner qu’en général (c’est-à-dire pas seulement pour l’imagerie calcique), l’approche par tranches pour étudier la physiologie pancréatique permet de gagner du temps car elle n’implique pas de période de récupération après l’isolement. Ce dernier n’est pas absolument nécessaire avec tous les types d’expériences et d’utilisations d’îlots isolés de différentes espèces, mais est généralement utilisé pour augmenter la pureté, restaurer la viabilité et la fonctionnalité, et parfois pour collecter des îlots de plusieurs donneurs 59,60,61,62,63,64 . Cependant, dans le contexte de l’imagerie calcique, les réponses des cellules bêta dépendent de la durée et des conditions de culture, ce qui constitue une source importante de variation qui doit être prise en compte lors de l’utilisation d’îlots isolés15,65. La même question devrait être envisagée pour les tranches de tissu si leur culture à long terme devient une option largement utilisée à l’avenir22,36. La méthode des tranches de tissu a également un rendement élevé et réduit ainsi potentiellement la souffrance animale et augmente la puissance statistique. De plus, autant de tranches peuvent être préparées à partir d’un seul animal et parce que les tranches survivent pendant de longues périodes, il devient possible d’inclure le même animal ou même le même îlot dans les groupes expérimental et témoin.
Comme l’architecture originale et les communications de cellule à cellule sont préservées, et parce qu’elle est compatible avec un certain nombre d’analyses structurelles, d’électrophysiologies, de méthodes d’imagerie et de tests de sécrétion d’hormones, cette méthode est particulièrement utile pour étudier les fonctions pancréatiques qui dépendent d’interactions non perturbées entre les cellules individuelles, par exemple la sensibilité aux sécrétagogues, la paracrine et les interactions immunitaires entre différents types de cellules, les modèles d’activité électrique, les propriétés de la dynamique du calcium et la sécrétion de différentes hormones. Pour l’imagerie calcique en particulier, les principaux avantages de l’utilisation de tranches sont l’exposition du noyau de l’îlot et la possibilité d’acquérir des signaux de nombreux types de cellules différents avec une haute résolution. Selon les exigences de l’expérience et l’âge des animaux, l’épaisseur peut varier, les tranches peuvent être transfectées ou obtenues à partir d’animaux avec des rapporteurs génétiquement codés. Comme expliqué plus en détail ci-dessous, les deux dernières approches permettent également l’identification fonctionnelle spécifique et la caractérisation des réponses des cellules non bêta 31,66. De plus, les îlots provenant de parties bien définies de l’organe peuvent être étudiés pour déterminer les différences de réactivité ou de susceptibilité à la maladie. Bien qu’ils ne nécessitent pas de période d’incubation de récupération, ils peuvent facilement être incubés avec différents agents pharmacologiques, acides gras, glucose élevé et cytokines.
Plus important encore, comme la haute résolution est réalisable en combinaison avec une résolution unicellulaire ou même subcellulaire, l’imagerie confocale du calcium en tranches est l’une des méthodes les plus appropriées pour analyser les ondes de calcium, la connectivité fonctionnelle et les différents rôles fonctionnels des cellules dans des parties distinctes d’un îlot54,67. Malgré un certain nombre d’avantages, l’approche des tranches de tissu présente d’importantes limites. Tout d’abord, il perturbe encore au moins en partie l’architecture des îlots et des exocrines, en particulier à la surface de coupe, et des précautions, telles que des températures basses, des échanges fréquents de solutions et une manipulation douce et rapide, sont nécessaires pendant la préparation pour éviter des dommages enzymatiques mécaniques et endogènes supplémentaires. Deuxièmement, les schémas d’administration des nutriments et des sécrétagogues sont encore inférieurs à la voie in vivo, la préparation est détachée de l’innervation systémique et la rétroaction inter-organes, par exemple entre l’îlot et ses tissus cibles, est impossible, contrairement aux approches in vivo. Troisièmement, l’épaisseur maximale de la tranche est limitée par l’oxygénation, l’apport de nutriments et la régulation du pH à ~200 μm9. En outre, la préparation des tranches et l’imagerie nécessitent beaucoup de formation, et les analyses approfondies des données sur le calcium provenant de longues séries chronologiques et de nombreuses cellules nécessitent des connaissances spécialisées qui ne sont souvent pas incluses dans la boîte à outils d’un physiologiste classique et nécessitent l’aide de physiciens ou de scientifiques des données. L’avantage que les interactions homo- et hétérotypiques sont préservées peut également compliquer l’analyse des échantillons en raison de la présence de signaux provenant d’autres cellules dans les régions d’intérêt. Selon les protocoles, l’activation d’autres cellules peut entraîner une stimulation ou une inhibition supplémentaire indirecte d’une cellule observée.
Cela ne peut être résolu de manière concluante que par des approches de déconvolution, par des protocoles de stimulation plus complexes, y compris des substances qui bloquent certains des effets indirects, en utilisant des animaux knock-out spécifiques et en comparant soigneusement les résultats avec les résultats d’autres études utilisant des méthodologies plus réductionnistes. De plus, si des mesures de sécrétion sont nécessaires, il convient de garder à l’esprit que certaines tranches peuvent manquer d’îlots et que la masse totale de tissu endocrinien dans une seule tranche est généralement faible. La préparation de tranches de tissu pancréatique aigu pour l’imagerie implique plusieurs étapes critiques discutées dans les sections suivantes et résumées dans le tableau 1, où le lecteur peut également trouver des conseils courts mais importants pour le dépannage. Tout d’abord, lors de la préparation de la solution d’agarose, la poudre d’agarose doit se dissoudre complètement, sinon les particules non dissoutes peuvent obstruer l’injection. Conserver la solution homogène d’agarose à 37-45 °C pour éviter le durcissement de l’agarose dû à une température trop basse d’une part et pour éviter les dommages tissulaires dus à des températures trop élevées d’autre part. Après utilisation, l’agarose restante peut être stockée à 4 °C et réchauffée, bien qu’un réchauffage répété puisse entraîner une augmentation de la densité due à l’évaporation de l’eau, ce qui rend éventuellement l’injection difficile ou impossible.
La prochaine étape critique de la préparation consiste à serrer correctement la papille duodénale majeure. Une tache blanche sur le duodénum indique la jonction du canal biliaire commun et du duodénum. Une pince placée trop proximalement entraînera une obstruction de certaines branches pancréatiques latérales du canal commun, désactivant l’injection de ces parties, tandis qu’une pince placée trop distalement entraînera une fuite d’agarose à travers le chemin de résistance inférieur directement dans le duodénum. Avant la canulation du canal biliaire commun, le tissu adipeux environnant peut être soigneusement retiré pour une meilleure visualisation du canal et un meilleur contrôle pendant l’injection. Une précision insuffisante lors de l’ablation du tissu environnant peut entraîner une perforation du conduit. Le choix du diamètre de l’aiguille utilisée pour l’injection d’agarose est également important. Chez la souris, une aiguille de 30 G est de préférence utilisée ; les aiguilles plus petites (32 ou 33 G) nécessitent plus d’efforts en raison de la viscosité élevée de la solution d’agarose et sont plus sujettes à l’obstruction. Cependant, s’ils sont utilisés en combinaison avec une solution d’agarose de densité inférieure, ils peuvent être très utiles chez les souches de souris plus petites et les animaux plus jeunes. Au cours des premiers jours postnatals, l’agarose peut également être injectée par voie sous-capsulaire plutôt que par voie intracanalaire2. L’utilisation d’aiguilles de plus grand diamètre chez la souris entraînera très probablement l’endommagement du canal biliaire commun. Cela peut également se produire avec le bon diamètre d’aiguille, et une pince peut aider à maintenir l’aiguille en place pendant l’injection. Des aiguilles de plus grand diamètre peuvent être la seule solution dans le cas de conduits plus grands, comme on en trouve chez les rats. Si l’aiguille est trop étroite pour assurer une étanchéité étanche empêchant les fuites arrière, une ligature peut être placée autour d’elle lors de l’entrée réussie dans le conduit.
L’injection d’agarose demande un certain effort en raison de la viscosité de la solution, et une fois que le processus d’injection a commencé, il ne doit pas être interrompu car la solution d’agarose à point de fusion bas peut se solidifier dans l’aiguille ou les plus grandes parties de l’arbre canalaire avant la fin de l’injection. Cela entraînera une mauvaise pénétration des tissus et un soutien moindre pendant la coupe. Le canal doit toujours être canulé au point où le canal hépatique gauche et le canal kystique se rejoignent pour former le canal biliaire commun. Si le canal biliaire commun est perforé, essayez à plusieurs reprises de canuler plus près du duodénum. Lorsque le pancréas est suffisamment stabilisé avec une solution d’agarose et extrait de la cavité péritonéale, de petits morceaux de tissu bien injecté sont coupés. Avant de les incorporer dans l’agarose, il est crucial d’enlever tous les tissus adipeux et conjonctifs, car leurs résidus rendent le tranchage plus difficile. Il en va de même pour les vaisseaux sanguins et les résidus de conduits, sauf lorsqu’ils sont au centre de l’expérience. Dans ce cas, assurez-vous de les positionner de manière à obtenir la section transversale souhaitée. Lors de l’incorporation du tissu dans l’agarose, assurez-vous que la température est appropriée (37 ° C) et que le tissu est complètement entouré d’agarose, car les forces exercées lors du tranchage du vibratome peuvent arracher le tissu pancréatique des blocs d’agarose.
Sécher rapidement les blocs de tissus avant de les placer dans de l’agarose en les plaçant brièvement sur un mouchoir en papier peut aider à prévenir un mauvais contact entre les tissus et l’agarose au cours de cette étape. Pendant la solidification des blocs d’agarose, placez la boîte de Petri horizontalement et empêchez le contact entre le tissu pancréatique et le fond de la boîte de Pétri. Si le pancréas n’est pas complètement injecté, le processus de coupe sera difficile. Par conséquent, essayez de réduire la vitesse de coupe pour obtenir des tranches de tissu. Pour minimiser les dommages cellulaires lors du tranchage du vibratome, remplacez régulièrement le SEC (et les glaçons en SEC) dans la chambre de tranchage. Ce dernier réduira l’activité des enzymes pancréatiques libérées par le tissu acineux lors du tranchage. L’épaisseur des tranches est également d’une importance cruciale. Pour les expériences de dynamique du calcium et d’électrophysiologie, des tranches de 140 μm sont généralement coupées; cependant, selon l’objectif de l’étude, l’épaisseur des tranches peut varier de 90 μm à 200 μm. Gardez à l’esprit que dans les tranches plus épaisses, la diffusion de l’oxygène et des nutriments sera limitée, mais ils incluront plus de tissu. De plus, on peut s’attendre à ce que la proportion d’îlots non coupés augmente avec l’augmentation de l’épaisseur des tranches. Les tranches peuvent être stockées dans un SEC régulièrement échangé à température ambiante pendant plusieurs heures ou même cultivées dans un milieu cellulaire approprié pendant plusieurs jours; cependant, cela peut éventuellement affecter la physiologie normale des cellules des îlots 3,22.
Lors de la préparation de la solution de colorant, assurez-vous d’un mélange minutieux de tous les composants et évitez l’exposition à la lumière ambiante. La tranche pancréatique est composée de nombreuses couches cellulaires, et l’absorption du colorant calcique est limitée aux premières couches cellulaires les plus superficielles, comme décrit précédemment pour les îlots isolés58,68 et les tranches hypophysaires69. Cependant, contrairement aux îlots isolés où la capsule environnante et les couches cellulaires externes entravent la pénétration du colorant dans les couches plus profondes, les tranches de tissu permettent d’accéder à toute la surface transversale de l’îlot, permettant la mesure simultanée de la dynamique du calcium dans des centaines de cellules de toutes les couches d’un îlot. Les indicateurs fluorescents Ca2+ sont les plus largement utilisés pour mesurer la dynamique du calcium et, avec CLSM, ils permettent des enregistrements à haute résolution temporelle, atteignant plusieurs centaines de Hertz. Lors de la sélection de l’indicateur fluorescent Ca2+ le plus approprié, tenez compte de différents facteurs, y compris la forme de l’indicateur, qui influence la méthode de charge cellulaire, le mode de mesure (qualitatif ou quantitatif) et la constante de dissociation (Kd) qui doit se situer dans la plage de concentration Ca2+ et dépend du pH, de la température, de la présence de Mg2+ et d’autres ions, ainsi que la liaison aux protéines. Comme les signaux cellulaires Ca2+ sont généralement transitoires, la constante de taux de liaison Ca2+ doit également être prise en compte. Pour mesurer la dynamique [Ca2+]IC dans les cellules pancréatiques, ce groupe utilise principalement le colorant indicateur Ca2+ perméable aux cellules décrit dans ce protocole (Table des matériaux) car il s’agit d’un indicateur de longue longueur d’onde avec les longueurs d’onde d’émission dans le spectre où l’autofluorescence cellulaire est généralement moins problématique et l’énergie de la lumière d’excitation est faible, ce qui réduit le risque de photodommages cellulaires. Parce que ce colorant est fluorescent à de faibles concentrations de Ca2+, cela facilite la détermination de la ligne de base [Ca2+]IC et augmente la visibilité cellulaire avant la stimulation. Après avoir lié le Ca2+, l’intensité de fluorescence du colorant est multipliée par 14, ce qui permet de détecter même de légers changements dans le [Ca2+]IC.
Pour une imagerie calcique réussie des cellules vivantes, plusieurs paramètres matériels cruciaux doivent être pris en compte, comme décrit dans la section du protocole. Pour l’imagerie de cellules vivantes dans laquelle les amplitudes du signal sont faibles et les risques de phototoxicité élevés, des objectifs avec un NA plus élevé sont de préférence utilisés pour recueillir plus de lumière de l’échantillon. Si la dynamique du calcium doit être enregistrée avec une résolution temporelle élevée, utilisez le scanner résonant au lieu de galvanomètres linéaires. Outre le choix du bon objectif, l’utilisation de détecteurs très sensibles, tels que les détecteurs hybrides nécessitant moins de puissance laser, évite la phototoxicité et le photoblanchiment. Ceci est d’une importance particulière pour l’imagerie calcique de longue durée. D’autres étapes importantes de l’imagerie calcique sont les paramètres de qualité d’image pour les acquisitions de séries chronologiques. Les plus importants sont la résolution temporelle et spatiale. Comme la dynamique du calcium en soi détermine la résolution temporelle acceptable la plus faible, le taux d’échantillonnage doit être au moins deux fois plus élevé que la fréquence de signal attendue pour détecter le signal ou même 10 fois plus élevé pour détecter la forme du signal de manière fiable. Dans les tranches de tissu pancréatique aigu, la dynamique du calcium peut être mesurée simultanément dans des centaines de cellules et, par conséquent, la résolution spatiale est également importante. Cela peut être amélioré en augmentant le nombre de pixels ou en augmentant la moyenne des lignes lors de l’acquisition en direct. Cependant, en raison de la relation inverse entre la résolution spatiale et la résolution temporelle, un compromis entre les deux paramètres est nécessaire.
Si l’imagerie calcique doit être réalisée dans une population cellulaire spécifique du pancréas, un stimulus capable de différencier fonctionnellement les cellules à l’intérieur de la tranche est nécessaire. Un taux élevé de glucose active de manière fiable et rapide les cellules bêta selon un schéma oscillatoire qui se superpose à un taux de calcium élevé et est hautement synchronisé entre toutes les cellules d’un îlot 32,58,70. Les cellules bêta sont le type de cellule le plus nombreux dans un îlot et sont situées principalement dans le noyau de l’îlot chez la souris. Le même protocole de stimulation diminue et parfois ne modifie pas sensiblement l’éclatement des cellules alpha 30,32,58,70,71,72. Pour discriminer fonctionnellement les cellules alpha, un faible taux (3 mM) de glucose, de glutamate ou d’adrénaline peut être utilisé pour augmenter leur fréquence ou basale [Ca2+]IC 21,72,73,74,75. Ils représentent 10 à 20% des cellules des îlots et seront détectés à la périphérie des îlots1. Les cellules Delta se trouvent également à la périphérie. Ils ne représentent que ~ 5% du nombre total de cellules endocriniennes dans un îlot et sont généralement actifs dans 6 mM de glucose et répondent à la stimulation du glucose avec une activité d’éclatement irrégulière accrue par rapport à l’inclusion ou un niveau de calcium légèrement élevé 1,32,71,76. La ghréline peut être utilisée pour la stimulation spécifique des cellules delta 21,77,78,79 dans les expériences d’imagerie calcique. Cependant, des protocoles pour l’identification fonctionnelle spécifique des cellules PP et epsilon restent à définir. En outre, 25 nM acétylcholine active de manière fiable les cellules acineuses dans l’activité d’éclatement 35,80,81. En outre, un certain nombre d’autres sécrétagogues, tels que la céruléine, la cholécystokinine et la carbamylcholine, peuvent être utilisés pour évoquer les réponses calciques dans les cellules acineuses 22,40,82,83.
Enfin, l’acide chénodésoxycholique de 1 mM évoque de manière fiable les réponses calciques dans les cellules canalaires dans les tranches de tissu; l’angiotensine II, l’ATP et d’autres sécrétagogues peuvent également être utilisés 11,23,84,85. Chaque fois qu’une identification fonctionnelle basée sur des réponses caractéristiques à des sécrétagogues et à des inhibiteurs spécifiques n’est pas suffisante, des animaux génétiquement marqués31, des cellules transfectées73 ou l’immunocytochimie peuvent être utilisés pour l’identification de différents types de cellules 9,22,71,86 . Au cours des deux dernières années, la méthode des tranches de tissu a été adaptée avec succès aux tissus humains, ouvrant de nombreuses nouvelles voies de recherche importantes à la fois dans l’exocrine41 et la physiologie endocrinienne 9,36,37,39. Fait intéressant, une évaluation détaillée de la dynamique du calcium dans les îlots humains a été notoirement difficile et reste à étudier plus en détail87. Combinée à la microscopie confocale avancée, la méthode de la tranche de tissu pancréatique a permis de nombreuses nouvelles connaissances sur la dynamique du calcium chez la souris et, espérons-le, fera de même pour les tissus humains.
Déclarations de divulgation
Les auteurs déclarent que la recherche a été menée en l’absence de tout intérêt commercial ou financier.
Remerciements
Les travaux présentés dans cette étude ont été soutenus financièrement par l’Agence slovène de la recherche (financement de base de la recherche no. P3-0396 et I0-0029, ainsi que des projets de recherche nos. J3-9289, N3-0048 et N3-0133) et par le Fonds autrichien pour la science / Fonds zur Förderung der Wissenschaftlichen Forschung (subventions bilatérales I3562--B27 et I4319--B30). Nous remercions Maruša Rošer, Maša Čater et Rudi Mlakar pour leur excellente assistance technique.
matériels
| Name | Company | Catalog Number | Comments |
| Equipment | |||
| Analytical balance KERN ALJ 120-4 | KERN & SOHN GmbH | ALJ 160-4A | |
| Confocal microscope Leica TCS SP5 II Upright setup | Leica | 5100001578 | |
| Confocal microscope Leica TCS SP5 AOBS Tandem II setup | Leica | ||
| Cork pad 15 cm x 15 cm | |||
| Corning 15 mL centrifuge tubes | Merck KGaA, Darmstadt, Germany | CLS430790 | |
| Corning Round Ice Bucket with Lid, 4 L | Fischer Scientific, Leicestershire, UK | 432124 | |
| Double edge razor blade | Personna, USA | ||
| Dumont #5 - Fine Forceps | FST, Germany | 11254-20 | |
| Eppendorf Safe-Lock Tubes 0.5 mL | Eppendorf | 0030 121.023 | |
| Erlenmeyer flask 200 mL | IsoLab, Germany | 027.01.100 | |
| Fine Scissors - ToughCut | FST, Germany | 14058-11 | |
| Flat orbital shaker IKA KS 260 basic | IKA | Ident. No.: 0002980200 | |
| Glass lab bottle 1000 mL | IsoLab, Germany | 091.01.901 | |
| Hartman Hemostat, curved | FST, Germany | 13003-10 | |
| HCX APO L 20x/1.00 W HCX APO L (water immersion objective, 20x, NA 1.0) | Leica | 15507701 | |
| Measuring cylinder 25 mL | IsoLab, Germany | 015.01.025 | |
| Micromanipulator Control box SM-7, Keypad SM-7 | Luigs & Neumann | 200-100 900 7311, 200-100 900 9050 | |
| Microwave owen | Gorenje, Slovenia | MO20MW | |
| Osmometer Gonotec 010 | Gonotec, Berlin, Germany | OSMOMAT 010 Nr. 01-02-20 | |
| Paint brush | Faber-Castell, No.2 | Any thin soft round paint brush No.2, preferably black | |
| Paper towels | |||
| Perifusion pumps | Ismatec | ISM 827 | Reglo Analog MS - 4/8 |
| Petri dish 100/20 mm | Sarstedt | 83.3902 | |
| Petri dish 35/10 mm | Greiner bio-one | 627102 | |
| Petri dish 35 x 10 mm Nunclon Delta | Thermo Fischer Scientific, Waltham, MA USA | 153066 | NON-STICKY for agarose blocks |
| pH meter inoLab pH Level 1 | WTW, Weilheim, Germany | E163694 | |
| Pipette 1000 mL | Eppendorf | 3121 000.120 | |
| Pipette 50 mL | Eppendorf | 3121 000.066 | |
| Push pins 23 mm | Deli, Ningbo, China | E0021 | |
| Screw cap tube, 15 mL | Sarstedt | 62.554.502 | |
| Semken Forceps | FST, Germany | 11008-13 | |
| Stabilizing ring for Erlenmeyer flask | IsoLab, Germany | 027.11.048 | |
| Stereomicroscope Nikon SMZ 745 | Nikon, Melville, NY USA | ||
| Syringe Injekt Solo 5 mL | Braun, Melsungen, Germany | 4606051V | |
| Syringe needle 0.30 x 12 mm (30 G x 1/2") | Braun, Melsungen, Germany | 4656300 | |
| Temperature controller | Luigs & Neumann | 200-100 500 0150, 200-150-500-145 | Slice mini chamber, Temperature controller TC 07 |
| Tubings for perifusion system | Ismatec | SC0310 | Ismatec Pharmed 1.14 mm(ID) + silicone tubing 1.0 (ID) x 1.8 mm(OD) |
| Ultrasonic bath Studio GT-7810A | Globaltronics | ||
| Vibrotome Leica VT 1000 S | Leica, Nussloch, Germany | 14047235613 | |
| Volumetric flask 1000 mL | IsoLab, Germany | 013.01.910 | |
| Vortex mixer Neolab 7-2020 | Neolab | 7-2020 | |
| Water bath Thermo Haake open-bath circulator | Thermo Fisher Scientific | Z527912 | |
| Material/Reagent | |||
| Calcium chloride dihydrate - CaCl2.2H2O | Sigma Aldrich, Germany | C5080-500G | |
| D-(+)-glucose | Sigma Aldrich, Germany | G8270-1KG | |
| Dimethyl sulfoxide | Sigma Aldrich | D4540-100ML | |
| DL-lactic acid | Sigma Aldrich, Germany | L1250-500ML | |
| Dulbecco’s Phosphate Buffered Saline | Merck KGaA, Darmstadt, Germany | D8662-500ML | |
| Gas mixture containing 95% O2 and 5% CO2 at barometric pressure | |||
| Glue Wekem sekundenkleber WK-110 | Wekem GmbH, Bergkamen, Germany | WK 110-020 | |
| HEPES | Sigma Aldrich, Germany | H3375-250G | |
| L-(+)-ascorbic acid | Sigma Aldrich, Germany | A9,290-2 | |
| LIVE/DEAD Viability/Cytotoxicity Kit, for mammalian cells | Thermo Fischer Scientific, Waltham, MA USA | L3224 | |
| Magnesium chloride hexahydrate - MgCl2.2H2O | Sigma Aldrich, Germany | M2670-500G | |
| Myo-inositol | Sigma Aldrich, Germany | I5125-100G | |
| Oregon Green 488 BAPTA-1, AM | Invitrogen (Thermo FisherScientific) | O6807 | cell-permeable Ca2+ indicator (excitation/emission: 495/523 nm) |
| Pluronic F-127 (20% Solution in DMSO) | Invitrogen (Thermo Fisher Scientific) | P3000MP | polaxamer: nonionic triblock copolymer |
| Potassium chloride - KCl | Sigma Aldrich, Germany | 31248 | |
| SeaPlaque GTG agarose | Lonza, Rockland, USA | 50111 | |
| Sodium bicarbonate - NaHCO3 | Honeywell, Germany | 31437-500G | |
| Sodium chloride - NaCl | Honeywell, Germany | 31434-1KG | |
| Sodium hydroxide - NaOH | Sigma Aldrich, Germany | 30620 | |
| Sodium phosphate monobasic- NaH2PO4 | Sigma Aldrich, Germany | S0751-500G | |
| Sodium pyruvate | Sigma Aldrich, Germany | 15990-100G | |
| Software | |||
| FIJI | FIJI is an open source project | ||
| LASAF | Leica microsystems, Inc. | ||
| Matlab | Mathworks | ||
| Python | Python Software Foundation | Python is an open source project |
Références
- Dolensek, J., Rupnik, M. S., Stozer, A. Structural similarities and differences between the human and the mouse pancreas. Islets. 7 (1), e1024405(2015).
- Meneghel-Rozzo, T., Rozzo, A., Poppi, L., Rupnik, M. In vivo and in vitro development of mouse pancreatic ß-cells in organotypic slices. Cell and Tissue Research. 316 (3), 295-303 (2004).
- Rozzo, A., Meneghel-Rozzo, T., Delakorda, S. L., Yang, S. B., Rupnik, M. Exocytosis of insulin: in vivo maturation of mouse endocrine pancreas. Annals of the New York Academy of the Sciences. 1152, 53-62 (2009).
- Dolenšek, J., Pohorec, V., Rupnik, M. S., Stožer, A. Pancreas physiology, challenges in pancreatic pathology. IntechOpen. Seicean, A. , (2017).
- Williams, J. A. The nobel pancreas: a historical perspective. Gastroenterology. 144 (6), 1166-1169 (2013).
- Lacy, P. E., Kostianovsky, M. Method for the isolation of intact islets of Langerhans from the rat pancreas. Diabetes. 16 (1), 35-39 (1967).
- Williams, J. A., Korc, M., Dormer, R. L. Action of secretagogues on a new preparation of functionally intact, isolated pancreatic acini. American Journal of Physiology. 235 (5), 517-524 (1978).
- Peikin, S. R., Rottman, A. J., Batzri, S., Gardner, J. D. Kinetics of amylase release by dispersed acini prepared from guinea pig pancreas. American Journal of Physiology. 235 (6), E743-E749 (1978).
- Marciniak, A., et al. Using pancreas tissue slices for in situ studies of islet of Langerhans and acinar cell biology. Nature Protocols. 9 (12), 2809-2822 (2014).
- Skelin, M., Rupnik, M., Cencic, A. Pancreatic beta cell lines and their applications in diabetes mellitus research. Altex-Alternatives to Animal Experimentation. 27 (2), 105-113 (2010).
- Molnar, R., et al. Mouse pancreatic ductal organoid culture as a relevant model to study exocrine pancreatic ion secretion. Laboratory Investigation. 100 (1), 84-97 (2020).
- Rupnik, M. The physiology of rodent beta-cells in pancreas slices. Acta Physiologica (Oxford, England). 195 (1), 123-138 (2009).
- Blinman, T. A., et al. Activation of pancreatic acinar cells on isolation from tissue: cytokine upregulation via p38 MAP kinase. American Journal of Physiology. Cell Physiology. 279 (6), C1993-C2003 (2000).
- Speier, S., Rupnik, M. A novel approach to in situ characterization of pancreatic ß-cells. Pflügers Archive: European Journal of Physiology. 446 (5), 553-558 (2003).
- Gilon, P., Jonas, J., Henquin, J. Culture duration and conditions affect the oscillations of cytoplasmic calcium concentration induced by glucose in mouse pancreatic islets. Diabetologia. 37 (10), 1007-1014 (1994).
- Huang, C., Gu, G. Effective isolation of functional islets from neonatal mouse pancreas. Journal of Visualized Experiments: JoVE. (119), e55160(2017).
- Szot, G. L., Koudria, P., Bluestone, J. A. Murine pancreatic islet isolation. Journal of Visualized Experiments: JoVE. (7), e255(2007).
- Qi, M., et al. Human pancreatic islet isolation: Part I: digestion and collection of pancreatic tissue. Journal of Visualized Experiments: JoVE. (27), e1125(2009).
- Qi, M., et al. Human pancreatic islet isolation: Part II: purification and culture of human islets. Journal of Visualized Experiments: JoVE. (27), e1343(2009).
- Stull, N. D., Breite, A., McCarthy, R., Tersey, S. A., Mirmira, R. G. Mouse islet of Langerhans isolation using a combination of purified collagenase and neutral protease. Journal of Visualized Experiments: JoVE. (67), e4137(2012).
- Hamilton, A., Vergari, E., Miranda, C., Tarasov, A. I. Imaging calcium dynamics in subpopulations of mouse pancreatic islet cells. Journal of Visualized Experiments: JoVE. (153), (2019).
- Marciniak, A., Selck, C., Friedrich, B., Speier, S. Mouse pancreas tissue slice culture facilitates long-term studies of exocrine and endocrine cell physiology in situ. PLoS ONE. 8 (11), e78706(2013).
- Gal, E., et al. A Novel in situ approach to studying pancreatic ducts in mice. Frontiers in Physiology. 10, 938(2019).
- Speier, S., Yang, S. B., Sroka, K., Rose, T., Rupnik, M. KATP-channels in beta-cells in tissue slices are directly modulated by millimolar ATP. Molecular and Cellular Endocrinology. 230 (1-2), 51-58 (2005).
- Speier, S., Gjinovci, A., Charollais, A., Meda, P., Rupnik, M. Cx36-mediated coupling reduces β-cell heterogeneity, confines the stimulating glucose concentration range, and affects insulin release kinetics. Diabetes. 56 (4), 1078-1086 (2007).
- Rose, T., Efendic, S., Rupnik, M. Ca2+-secretion coupling is impaired in diabetic Goto Kakizaki rats. The Journal of General Physiology. 129 (6), 493-508 (2007).
- Paulmann, N., et al. Intracellular serotonin modulates insulin secretion from pancreatic β-cells by protein serotonylation. PLoS Biology. 7 (10), e1000229(2009).
- Mandic, S. A., et al. Munc18-1 and Munc18-2 proteins modulate β-cell Ca2+ sensitivity and kinetics of insulin exocytosis differently. Journal of Biological Chemistry. 286 (32), 28026-28040 (2011).
- Dolensek, J., Skelin, M., Rupnik, M. S. Calcium dependencies of regulated exocytosis in different endocrine cells. Physiological Research. 60, S29-S38 (2011).
- Huang, Y. C., Rupnik, M., Gaisano, H. Y. Unperturbed islet α-cell function examined in mouse pancreas tissue slices. Journal of Physiology. 589 (2), 395-408 (2011).
- Huang, Y. C., et al. In situ electrophysiological examination of pancreatic α cells in the streptozotocin-induced diabetes model, revealing the cellular basis of glucagon hypersecretion. Diabetes. 62 (2), 519-530 (2013).
- Stožer, A., Dolenšek, J., Rupnik, M. S. Glucose-stimulated calcium dynamics in islets of Langerhans in acute mouse pancreas tissue slices. PLoS ONE. 8 (1), e54638(2013).
- Stožer, A., et al. Functional connectivity in islets of Langerhans from mouse pancreas tissue slices. PLoS Computational Biology. 9 (2), e1002923(2013).
- Dolenšek, J., Stožer, A., Skelin Klemen, M., Miller, E. W., Slak Rupnik, M. The relationship between membrane potential and calcium dynamics in glucose-stimulated beta cell syncytium in acute mouse pancreas tissue slices. PLoS ONE. 8 (12), e82374(2013).
- Perc, M., Rupnik, M., Gosak, M., Marhl, M. Prevalence of stochasticity in experimentally observed responses of pancreatic acinar cells to acetylcholine. Chaos. 19 (3), 037113(2009).
- Qadir, M. M. F., et al. Long-term culture of human pancreatic slices as a model to study real-time islet regeneration. Nature Communications. 11 (1), 3265-3265 (2020).
- Panzer, J. K., et al. Pancreas tissue slices from organ donors enable in situ analysis of type 1 diabetes pathogenesis. JCI Insight. 5 (8), e134525(2020).
- Cohrs, C. M., et al. Vessel network architecture of adult human islets promotes distinct cell-cell interactions in situ and is altered after transplantation. Endocrinology. 158 (5), 1373-1385 (2017).
- Cohrs, C. M., et al. Dysfunction of persisting beta cells is a key feature of early type 2 diabetes pathogenesis. Cell Reports. 31 (1), 107469(2020).
- Dolai, S., et al. Pancreatitis-induced depletion of syntaxin 2 promotes autophagy and increases basolateral exocytosis. Gastroenterology. 154 (6), 1805-1821 (2018).
- Liang, T., et al. Ex vivo human pancreatic slice preparations offer a valuable model for studying pancreatic exocrine biology. Journal of Biological Chemistry. 292 (14), 5957-5969 (2017).
- Panzer, J. K., Cohrs, C. M., Speier, S. Using pancreas tissue slices for the study of islet physiology. Methods in Molecular Biology. 2128, 301-312 (2020).
- Klemen, M., Dolenšek, J., Stožer, A., Rupnik, M. Exocytosis Methods. Thorn, P. 7, Humana Press. 127-146 (2014).
- Speier, S. Experimental approaches for high-resolution in vivo imaging of islet of Langerhans biology. Current Diabetes Reports. 11 (5), 420-425 (2011).
- Leibiger, I. B., Berggren, P. O. Intraocular in vivo imaging of pancreatic islet cell physiology/pathology. Molecular Metabolism. 6 (9), 1002-1009 (2017).
- Reissaus, C. A., et al. A Versatile, portable intravital microscopy platform for studying beta-cell biology in vivo. Scientific Reports. 9 (1), 8449(2019).
- Jacob, S., et al. In vivo Ca(2+) dynamics in single pancreatic beta cells. FASEB Journal. 34 (1), 945-959 (2020).
- Fernandez, J., Valdeolmillos, M. Synchronous glucose-dependent [Ca2+]i oscillations in mouse pancreatic islets of Langerhans recorded in vivo. FEBS Letters. 477 (1-2), 33-36 (2000).
- Almaca, J., Weitz, J., Rodriguez-Diaz, R., Pereira, E., Caicedo, A. The pericyte of the pancreatic islet regulates capillary diameter and local blood flow. Cell Metabolism. 27 (3), 630-644 (2018).
- Weitz, J. R., et al. Mouse pancreatic islet macrophages use locally released ATP to monitor beta cell activity. Diabetologia. 61 (1), 182-192 (2018).
- Tian, G., Sandler, S., Gylfe, E., Tengholm, A. Glucose- and hormone-induced cAMP oscillations in α- and β-cells within intact pancreatic islets. Diabetes. 60 (5), 1535-1543 (2011).
- Benninger, R. K., Zhang, M., Head, W. S., Satin, L. S., Piston, D. W. Gap junction coupling and calcium waves in the pancreatic islet. Biophysical Journal. 95 (11), 5048-5061 (2008).
- Santos, R. M., et al. Widespread synchronous Ca oscillations due to bursting electrical activity in single pancreatic islets. Pflügers Archive: European Journal of Physiology. 418 (4), 417-422 (1991).
- terk, M., et al. Assessing the origin and velocity of Ca2+ waves in three-dimensional tissue: Insights from a mathematical model and confocal imaging in mouse pancreas tissue slices. Communications in Nonlinear Science and Numerical Simulation. 93, 105495(2021).
- Gosak, M., et al. Critical and supercritical spatiotemporal calcium dynamics in beta cells. Frontiers in Physiology. 8, 1106(2017).
- Satin, L. S., Butler, P. C., Ha, J., Sherman, A. S. Pulsatile insulin secretion, impaired glucose tolerance and type 2 diabetes. Molecular Aspects in Medicine. 42, 61-77 (2015).
- Tengholm, A., Gylfe, E. Oscillatory control of insulin secretion. Molecular and Cellular Endocrinology. 297 (1-2), 58-72 (2009).
- Quesada, I., et al. Glucose induces opposite intracellular Ca2+ concentration oscillatory patterns in identified α- and β-cells within intact human islets of Langerhans. Diabetes. 55 (9), 2463-2469 (2006).
- Ferguson, J., Allsopp, R. H., Taylor, R. M., Johnston, I. D. Isolation and long term preservation of pancreatic islets from mouse, rat and guinea pig. Diabetologia. 12 (2), 115-121 (1976).
- Andersson, A. Isolated mouse pancreatic islets in culture: effects of serum and different culture media on the insulin production of the islets. Diabetologia. 14 (6), 397-404 (1978).
- Ramirez-Dominguez, M. Isolation of mouse pancreatic islets of Langerhans. Advances in Experimental Medicine and Biology. 938, 25-34 (2016).
- Carter, J., Dula, S., Corbin, K., Wu, R., Nunemaker, C. A practical guide to rodent islet isolation and assessment. Biological Procedures Online. 11 (1), 3-31 (2009).
- Daoud, J., Rosenberg, L., Tabrizian, M. Pancreatic islet culture and preservation strategies: advances, challenges, and future outlook. Cell Transplantation. 19 (12), 1523-1535 (2010).
- Zawalich, W. S., Yamazaki, H., Zawalich, K. C. Biphasic insulin secretion from freshly isolated or cultured, perifused rodent islets: comparative studies with rats and mice. Metabolism. 57 (1), 30-39 (2008).
- Roe, M. W., et al. Absence of effect of culture duration on glucose-activated alterations in intracellular calcium concentration in mouse pancreatic islets. Diabetologia. 38, 876-877 (1995).
- Shuai, H., Xu, Y., Yu, Q., Gylfe, E., Tengholm, A. Fluorescent protein vectors for pancreatic islet cell identification in live-cell imaging. Pflügers Archive. European Journal of Physiology. 468 (10), 1765-1777 (2016).
- Dolenšek, J., et al. Glucose-dependent activation, activity, and deactivation of beta cell networks in acute mouse pancreas tissue slices. bioRxiv. , (2020).
- QZhang, Q., et al. Cell coupling in mouse pancreatic beta-cells measured in intact islets of Langerhans. Philosophical Transactions. Series A, Mathematical, Physical, and Engineering Sciences. 366 (1880), 3503-3523 (2008).
- Sánchez-Cárdenas, C., Hernández-Cruz, A. GnRH-induced Ca2+-signalling patterns in mouse gonadotrophs recorded from acute pituitary slices in vitro. Neuroendocrinology. 91 (3), 239-255 (2010).
- Asada, N., Shibuya, I., Iwanaga, T., Niwa, K., Kanno, T. Identification of alpha- and beta-cells in intact isolated islets of Langerhans by their characteristic cytoplasmic Ca2+ concentration dynamics and immunocytochemical staining. Diabetes. 47 (5), 751-757 (1998).
- Nadal, A., Quesada, I., Soria, B. Homologous and heterologous asynchronicity between identified α-, β- and δ-cells within intact islets of Langerhans in the mouse. Journal of Physiology. 517 (1), 85-93 (1999).
- Shuai, H., Xu, Y., Yu, Q., Gylfe, E., Tengholm, A. Fluorescent protein vectors for pancreatic islet cell identification in live-cell imaging. Pflugers Archive: European Journal of Physiology. 468 (10), 1765-1777 (2016).
- Cabrera, O., et al. Glutamate is a positive autocrine signal for glucagon release. Cell Metabolism. 7 (6), 545-554 (2008).
- Li, J., et al. Submembrane ATP and Ca2+ kinetics in α-cells: unexpected signaling for glucagon secretion. FASEB Journal. 29 (8), 3379-3388 (2015).
- Hamilton, A., et al. Adrenaline stimulates glucagon secretion by Tpc2-dependent Ca(2+) mobilization from acidic stores in pancreatic α-cells. Diabetes. 67 (6), 1128-1139 (2018).
- Arrojo, E. D. R., et al. Structural basis for delta cell paracrine regulation in pancreatic islets. Nature Communications. 10 (1), (2019).
- DiGruccio, M. R., et al. Comprehensive alpha, beta and delta cell transcriptomes reveal that ghrelin selectively activates delta cells and promotes somatostatin release from pancreatic islets. Molecular Metabolism. 5 (7), 449-458 (2016).
- Adriaenssens, A. E., et al. Transcriptomic profiling of pancreatic alpha, beta and delta cell populations identifies delta cells as a principal target for ghrelin in mouse islets. Diabetologia. 59 (10), 2156-2165 (2016).
- Rorsman, P., Huising, M. O. The somatostatin-secreting pancreatic δ-cell in health and disease. Nature reviews. Endocrinology. 14 (7), 404-414 (2018).
- Petersen, C. C., Toescu, E. C., Petersen, O. H. Different patterns of receptor-activated cytoplasmic Ca2+ oscillations in single pancreatic acinar cells: dependence on receptor type, agonist concentration and intracellular Ca2+ buffering. The EMBO journal. 10 (3), 527-533 (1991).
- Thorn, P., Lawrie, A. M., Smith, P. M., Gallacher, D. V., Petersen, O. H. Local and global cytosolic Ca2+ oscillations in exocrine cells evoked by agonists and inositol trisphosphate. Cell. 74 (4), 661-668 (1993).
- Behrendorff, N., Floetenmeyer, M., Schwiening, C., Thorn, P. Protons released during pancreatic acinar cell secretion acidify the lumen and contribute to pancreatitis in mice. Gastroenterology. 139 (5), e1711-e1715 (2010).
- Criddle, D. N., et al. Cholecystokinin-58 and cholecystokinin-8 exhibit similar actions on calcium signaling, zymogen secretion, and cell fate in murine pancreatic acinar cells. American Journal of Physiology. Gastrointestinal and Liver Physiology. 297 (6), G1085-G1092 (2009).
- Venglovecz, V., et al. Effects of bile acids on pancreatic ductal bicarbonate secretion in guinea pig. Gut. 57 (8), 1468-3288 (2008).
- Maleth, J., Hegyi, P. Calcium signaling in pancreatic ductal epithelial cells: an old friend and a nasty enemy. Cell Calcium. 55 (6), 337-345 (2014).
- Nadal, A., Quesada, I., Soria, B. Homologous and heterologous asynchronicity between identified alpha-, beta- and delta-cells within intact islets of Langerhans in the mouse. Journal of Physiology. 517 (Pt 1), 85-93 (1999).
- Skelin Klemen, M., Dolenšek, J., Slak Rupnik, M., Stožer, A. The triggering pathway to insulin secretion: Functional similarities and differences between the human and the mouse β cells and their translational relevance. Islets. 9 (6), 109-139 (2017).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.