
Method Article
Caractérisation de la cicatrisation épithéliale in vivo à l’aide de l’organisme modèle cnidaire Clytia hemisphaerica
Dans cet article
Résumé
Cet article décrit une méthode pour créer des plaies dans l’épithélium d’une Clytia hemisphaerica medusa vivante et imager la cicatrisation des plaies à haute résolution in vivo. En outre, une technique pour introduire des colorants et des médicaments pour perturber les processus de signalisation dans les cellules épithéliales et la matrice extracellulaire pendant la cicatrisation des plaies est présentée.
Résumé
Tous les organes animaux, de la peau aux yeux en passant par les intestins, sont recouverts de feuilles de cellules épithéliales qui leur permettent de maintenir l’homéostasie tout en les protégeant des infections. Par conséquent, il n’est pas surprenant que la capacité de réparer les plaies épithéliales soit essentielle pour tous les métazoaires. La cicatrisation épithéliale des plaies chez les vertébrés implique des processus qui se chevauchent, y compris les réponses inflammatoires, la vascularisation et la réépithélialisation. La régulation de ces processus implique des interactions complexes entre les cellules épithéliales, les cellules voisines et la matrice extracellulaire (ECM); l’ECM contient des protéines structurelles, des protéines régulatrices et de petites molécules actives. Cette complexité, ainsi que le fait que la plupart des animaux ont des tissus opaques et des ECM inaccessibles, rend la cicatrisation des plaies difficile à étudier chez les animaux vivants. Une grande partie du travail sur la cicatrisation des plaies épithéliales est donc effectuée dans des systèmes de culture tissulaire, avec un seul type de cellule épithéliale plaqué comme une monocouche sur une matrice artificielle. Clytia hemisphaerica (Clytia) fournit un complément unique et passionnant à ces études, permettant d’étudier la cicatrisation des plaies épithéliales chez un animal intact avec une ECM authentique. L’épithélium ectodermique de Clytia est une couche unique de grandes cellules épithéliales squameuses, permettant une imagerie à haute résolution utilisant la microscopie à contraste interférent différentiel (CIVD) chez les animaux vivants. L’absence de fibroblastes migrateurs, de réponses vasculaires ou inflammatoires permet de disséquer les événements critiques en réépithélialisation in vivo. La cicatrisation de divers types de plaies peut être analysée, y compris les microplaies unicellulaires, les petites et grandes plaies épithéliales et les plaies qui endommagent la membrane basale. La formation de lamellipodes, la contraction de la corde coulissante, l’étirement cellulaire et la migration cellulaire collective peuvent tous être observés dans ce système. De plus, des agents pharmacologiques peuvent être introduits via l’ECM pour modifier les interactions cellule:ECM et les processus cellulaires in vivo. Ce travail montre des méthodes pour créer des plaies dans Clytia vivant, capturer des films de guérison et sonder les mécanismes de guérison en microinjectant des réactifs dans l’ECM.
Introduction
Des feuilles de cellules épithéliales recouvrent la surface externe de tous les métazoaires, tapissent les organes internes et divisent le corps de l’animal en compartiments discrets. L’épithélium sépare également le corps interne de l’environnement extérieur et le protège des dommages et des infections. Par conséquent, l’avènement des couches épithéliales a été une partie essentielle de l’évolution des animaux multicellulaires, et les couches épithéliales sont observées chez tous les animaux, des vertébrés aux métazoaires les plus basaux1. L’épithélium de certains organes est une monocouche unique, comme dans les sacs aériens pulmonaires, les vaisseaux sanguins et l’intestin2, ainsi que dans l’épiderme d’invertébrés tels que les planaires et les cnidaires3. Dans d’autres tissus, tels que la peau4 et la cornée5 des vertébrés, l’épithélium est stratifié, ce qui signifie qu’il existe plusieurs couches de cellules épithéliales2. Dans tous les cas, la couche épithéliale la plus basale est fixée à la membrane basale, une feuille de protéines qui forme une région spécialisée de la matrice extracellulaire (MEC)6,7,8.
Les brèches dans l’épithélium doivent être rapidement réparées pour recréer une feuille épithéliale continue. Les dommages à l’épithélium se produisent au cours de processus naturels, tels que l’excrétion de cellules épithéliales dans l’intestin,9,10 et à la suite d’une inflammation ou d’un traumatisme physique. Lorsqu’une seule cellule épithéliale est endommagée, elle doit soit se réparer, soit être éliminée pour permettre aux cellules environnantes de se fixer les unes aux autres et de fermer le trou11,12. Dans les plaies plus grandes que la taille d’une seule cellule, les cellules épithéliales doivent se déplacer pour se rejoindre et réparer la feuille13. Ceci peut être réalisé par propagation cellulaire si les espaces sont petits ou peut nécessiter la migration des cellules épithéliales à partir des bords d’une plaie pour fermer l’espace de la plaie; Ce dernier processus est appelé réépithélialisation14,15. Dans les tissus embryonnaires, les cellules épithéliales se propagent et migrent vers des plaies rapprochées ou sont tirées à travers l’espace par la contraction des câbles d’actomyosine qui se forment entre les cellules au bord de la plaie, dans un mécanisme ressemblant à un cordon de bourse16. Dans de nombreux tissus adultes, la réépithélialisation implique la migration de feuillets cellulaires cohérents, où les cellules maintiennent leurs jonctions avec les cellules voisines14,17,18. Dans d’autres tissus, les connexions cellule-cellule sont démantelées et les cellules épithéliales se comportent davantage comme des cellules mésenchymateuses, se déplaçant de manière coordonnée mais indépendante dans la région de la plaie lors de la réépithélialisation 14,19,20,21.
Les mouvements des cellules épithéliales sont régulés par des interactions complexes entre les cellules migrantes et entre les cellules et l’ECM. Bien qu’il existe une énorme quantité de littérature expérimentale sur les mécanismes d’activation des cellules épithéliales et de migration ultérieure, il reste encore beaucoup à découvrir. Par exemple, le signal initial qui active les cellules épithéliales à migrer en réponse à une plaie n’a pas été définitivement identifié 22, ni ne comprend complètement comment l’actine est redéployée pour créer des lamellipodes du côté des cellules épithéliales les plus proches de la plaie 22,23,24,25,26,27. La migration cellulaire collective nécessite que les informations provenant des cellules de la plaie soient partagées avec les cellules distales de la plaie, et la voie de communication n’est toujours pas claire28. Les jonctions cellule:cellule et cellule:ECM doivent être désassemblées et reformées au fur et à mesure que les cellules de la feuille se réorganisent, mais la régulation de ce processus est mal comprise14,29. Faire des progrès sur ces questions et d’autres questions connexes est non seulement important en tant que problème biologique fondamental, mais aussi en raison de l’importance clinique d’une cicatrisation correcte des plaies. Les maladies qui compromettent la capacité des cellules épithéliales à migrer correctement entraînent des plaies chroniques; un exemple est la maladie génétique épidermolyse bulleuse, où les gènes impliqués dans la fixation des cellules épithéliales à l’ECM sont mutés, ce qui entraîne une peau fragile qui pèle et cloque. La réépithélialisation est également compromise dans les tissus naturellement vieillissants30,31. Une meilleure compréhension est donc essentielle pour développer des interventions visant à améliorer les résultats de cicatrisation des plaies.
La migration des cellules épithéliales dans la cicatrisation des plaies a été étudiée à l’aide d’approches in vitro et d’organismes modèles. La majorité des études sur la cicatrisation des plaies et les mécanismes de migration cellulaire ont été réalisées en culture tissulaire, où des monocouches d’un seul type de cellules épithéliales sont cultivées sur un substrat qui se substitue à l’ECM. Les monocouches cellulaires sont soit rayées, soit cultivées avec des pochoirs pour créer des espaces de formes et de tailles spécifiques, puis observées32,33,34. Le modèle in vitro permet une visualisation idéale du comportement cellulaire, ainsi que la possibilité de modifier les qualités du substrat, d’exposer les cellules à des médicaments et à des facteurs abiotiques et biotiques, et de transfecter des cellules avec des constructions qui expriment ou suppriment divers gènes d’intérêt. Cependant, cette approche réductionniste peut ne pas saisir certains des paramètres importants impliqués dans le comportement des cellules épithéliales dans un contexte in vivo, y compris la communication entre divers types de cellules et les événements de signalisation qui se produisent dans l’ECM11. Les modèles in vivo fournissent le contexte authentique d’une plaie, avec plusieurs types de cellules, des voies de signalisation qui se chevauchent et un ECMcomplexe 35. L’un de ces modèles pour les études sur la cicatrisation des plaies est la souris19, dans laquelle des progrès récents ont permis aux chercheurs d’observer les cellules épidermiques pendant la cicatrisation des plaies de pleine épaisseur chez les animaux vivants36. Cependant, la souris et d’autres systèmes in vivo présentent des défis pour étudier la réépithélialisation. Premièrement, le grand avantage d’observer le comportement cellulaire dans un contexte naturel est contrebalancé par la complexité des événements qui se chevauchent temporellement pendant la cicatrisation des plaies des vertébrés, y compris la coagulation du sang, le recrutement des cellules immunitaires et l’inflammation, le recrutement des fibroblastes et la dédifférenciation cellulaire, la revascularisation et le remodelage de l’ECM. De plus, les tissus opaques rendent l’imagerie difficile. Les systèmes de larve de drosophile et d’épiderme de poisson zèbre 37,38 ont surmonté certaines de ces difficultés en raison de leur relative simplicité39.
Notre laboratoire a récemment introduit un nouveau modèle pour étudier la cicatrisation des plaies épithéliales : la forme méduse (méduse) de l’hydrozoaire cnidaire Clytia hemisphaerica (Clytia)40. Clytia est un organisme modèle émergent avec un génome 41 entièrement séquencé et annoté, un transcriptome RNAseq unicellulaire42 et des protocoles en place pour la modification du génome (mutagénèse et transgénèse)43,44,45. Les cnidaires sont l’une des plus anciennes lignées existantes à avoir des couches épithéliales, de sorte que la compréhension de la cicatrisation des plaies cnidaires donne un aperçu des voies ancestrales qui assuraient l’intégrité épithéliale. Pour les voies qui ont été conservées tout au long de l’arbre de vie, Clytia propose un nouveau système passionnant pour étudier la dynamique des cellules épithéliales et la régulation fonctionnelle de la cicatrisation des plaies in vivo.
L’épithélium recouvrant la surface supérieure de la Clytia medusa (exumbrella) est une monocouche de cellules épithéliales épithéliales squameuses transparentes d’environ 50 μm de large sur 1-2 μm d’épaisseur (Figure 1). Ils sont attachés à un ECM appelé la mésogle - la « gelée » de la méduse. La composition de la mésogle est similaire à celle de l’ECM trouvée chez d’autres animaux 46,47,48, y compris les vertébrés, a une membrane basale 40 et est complètement transparente. La couche épithéliale de la Clytia medusa peut être facilement rayée ou blessée (voir ci-dessous). La simplicité et la transparence de l’épithélium et de l’ECM permettent une imagerie haute résolution des cellules et de leurs mouvements pendant la cicatrisation. Récemment, Kamran et al. ont caractérisé la cicatrisation de petites plaies dans l’épithélium Clytia en détail40. Il a été démontré que la guérison chez Clytia se produit par le rampement cellulaire à base de lamellipodes, la propagation cellulaire et la migration cellulaire collective, ainsi que par la fermeture du cordon de la bourse qui est plus typique des systèmes embryonnaires (bien que vu précédemment dans les structures animales adultes telles que la cornée49). La cicatrisation des plaies de Clytia est extrêmement rapide, comme on l’a vu dans d’autres systèmes qui n’ont pas de réponse inflammatoire40,50. La cicatrisation dans l’exumbrella de Clytia dépend entièrement des mouvements des cellules épithéliales existantes – aucune cellule ne prolifère ou ne migre à travers l’ECM vers le site de la plaie (film supplémentaire 1). Tous ces résultats suggèrent que Clytia est un système modèle utile pour étudier la cicatrisation des plaies épithéliales. En effet, la facilité d’imagerie des cellules épithéliales chez Clytia pendant la cicatrisation des plaies a conduit à la découverte que les lamellipodes des cellules épithéliales s’étendent et se propagent sur les zones d’ECM exposées tant qu’il y a une membrane basale intacte; Si la membrane basale est endommagée, la cicatrisation épithéliale passe à un mécanisme de cordon de bourse40. Il s’agissait de la première démonstration d’un mécanisme sous-jacent à la décision de fermer par rampement à base de lamellipodes plutôt que par fermeture de cordon de bourse, soulignant l’importance des interactions cellules:ECM spécifiques dans la guérison et de l’observation des cellules dans leur contexte naturel.
Ci-dessous, les protocoles sont décrits pour la création et l’imagerie de microplaies unicellulaires, de petites plaies qui se ferment principalement par propagation cellulaire et de grandes plaies qui nécessitent une migration cellulaire collective pour se fermer. En outre, un protocole est décrit pour l’introduction de petites molécules dans l’ECM et les cellules épithéliales, permettant des perturbations expérimentales des voies de régulation putatives de la cicatrisation des plaies.
Protocole
1. Culture animale
- Maintenir les colonies de polypes de Clytia sur des lames de microscope et des méduses dans de l’eau de mer artificielle (ASW) à 18 °C dans un système de poisson zèbre, avec des réservoirs de poisson-zèbre de 2 L pour les colonies de polypes et des réservoirs pseudo-kreisel de 5 L sur mesure pour les méduses (figure supplémentaire 1)51. ASW se compose de 4% d’Instant Ocean dans désionisé (DI) H2O.
- Nourrissez les animaux quotidiennement avec de l’artémie de 2-3 jours comme décrit51.
REMARQUE: L’imagerie de cicatrisation est plus facile si les animaux n’ont pas été nourris récemment, car il y a moins de débris libérés de l’intestin dans le champ de vision. - Recueillir les bébés méduses des colonies de polypes établies au besoin en plaçant les colonies dans un bécher de 2 L rempli de 1 L d’ASW pendant la nuit. Utilisez des méduses femelles âgées de 2 à 3 semaines pour toutes les expériences de cicatrisation. La propagation de Clytia a été décrite en détail ailleurs51.
2. Blessure
- Création de microplaies à l’intérieur et entre les cellules (20-500 μm2)
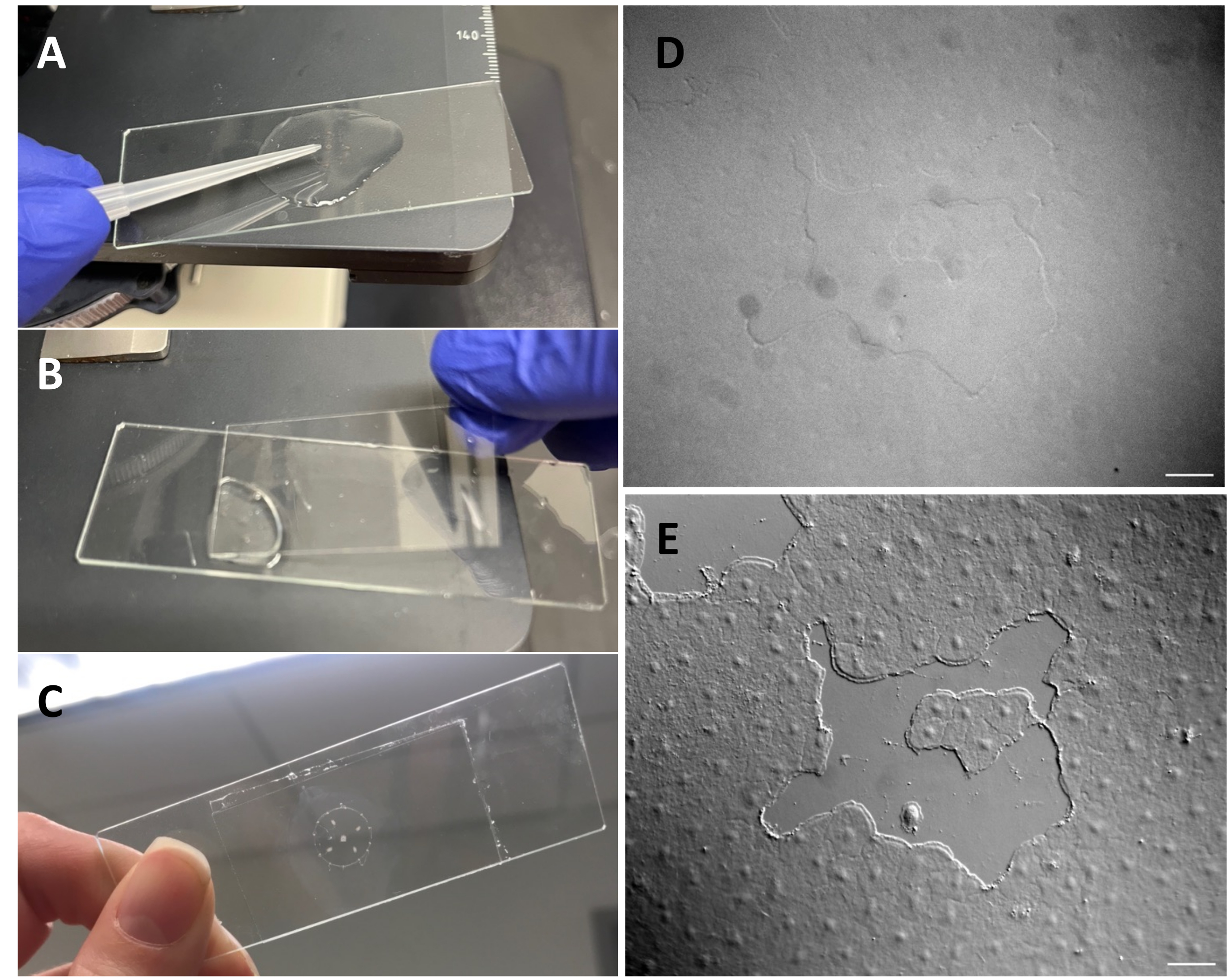
- Créez une pipette de transfert modifiée en coupant la pointe avec des ciseaux pour obtenir une ouverture plus grande (0,5-0,7 cm de diamètre).
REMARQUE : L’ouverture de la pipette doit être suffisamment large pour éviter tout dommage à l’animal. - À l’aide de la pipette de transfert modifiée, placez la méduse sur une lame de dépression avec l’exparapluie de méduse vers le haut, avec juste assez d’ASW pour couvrir l’animal.
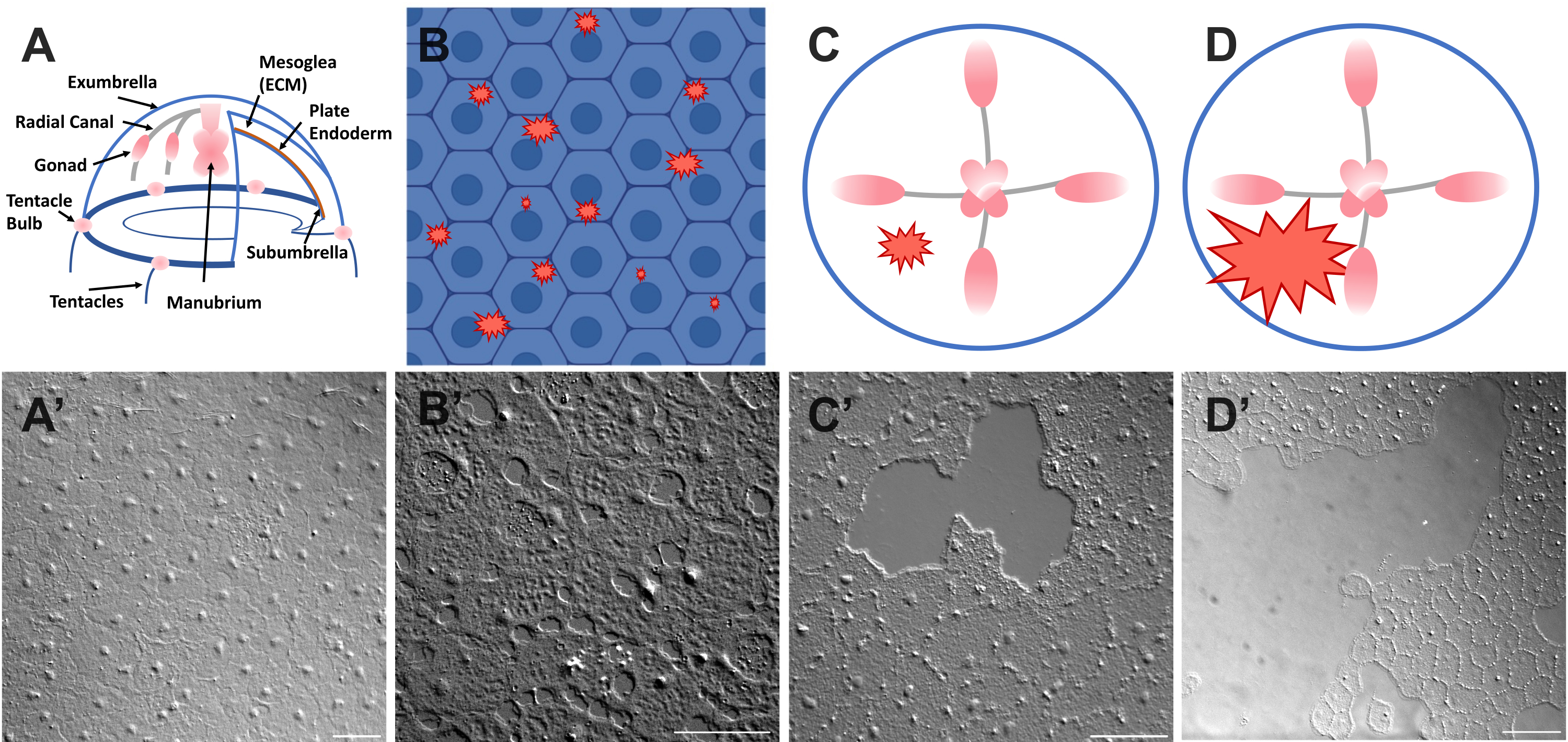
- Placez immédiatement un capseau sur l’animal et l’image (voir ci-dessous pour la description de l’imagerie). La lamelle de couverture comprime la mésoglée, et le rebond du tissu comprimé crée une force qui pousse les cellules légèrement à l’écart52. Cela apparaît immédiatement comme des espaces entre chaque cellule et des dommages à l’intérieur de certaines cellules (Figure 1B, B', Figure 2 et Figure 3A-C).
- Créez une pipette de transfert modifiée en coupant la pointe avec des ciseaux pour obtenir une ouverture plus grande (0,5-0,7 cm de diamètre).
- Création de petites plaies épithéliales (0,02-0,125 mm2)
- À l’aide d’une pipette de transfert modifiée (comme ci-dessus), placez la méduse sur une lame de dépression avec l’exparapluie de méduse vers le haut.
- À l’aide d’un embout de pipette de 200 μL, grattez doucement la surface de la méduse. Un léger grattage peut également créer des déchirures dans la membrane basale, qui sont facilement apparentes22. Couvrez l’animal d’un bordereau de couverture pour l’imagerie. Alternativement, le placement de la lamelle de couverture est parfois suffisant pour créer de petites plaies épithéliales même sans rayures (Figure 1C, C', Figure 2 et Figure 3A-C).
REMARQUE: N’appuyez pas vers le bas lorsque vous grattez la surface de la méduse, car cela endommage l’ECM et crée une surface irrégulière - les cellules épithéliales migrant sur une surface irrégulière sont plus difficiles à garder au point.
- Création de grandes plaies épithéliales (0,5-0,9 mm2)
- Fabriquer une aiguille de microinjection à l’aide d’un extracteur de micropipette et d’un tube capillaire en verre (étape 5.2). Placez l’aiguille de microinjection vide dans un porte-micro-injecteur fixé à un micromanipulateur. Coupez la pointe de l’aiguille de manière à ce que l’ouverture soit d’environ 20-40 μm.
REMARQUE: Les aiguilles coupées pour les grandes plaies épithéliales peuvent être stockées et réutilisées pour augmenter la cohérence entre les expériences. - Réglez la pression de maintien sur le micro-injecteur à zéro et réglez la pression d’éjection à environ 20 PSI. Réglez le micro-injecteur pour qu’il délivre une impulsion d’air de 2 s.
REMARQUE : Il peut être nécessaire d’ajuster la pression d’éjection en fonction du diamètre de l’ouverture de l’aiguille (c.-à-d. que les extrémités plus petites utiliseront une pression plus élevée, tandis que les extrémités plus grandes utiliseront une pression plus faible). - Placez la méduse avec l’exumbrella vers le haut sur une lame de dépression sur la scène d’une lunette de dissection, avec juste assez d’ASW pour couvrir l’animal. À l’aide du micromanipulateur, ajustez la pointe de l’aiguille de micro-injection de manière à ce qu’elle soit juste au-dessus de l’eau. Pour ce faire, plongez soigneusement l’embout dans l’eau (de l’eau peut pénétrer dans l’embout de la pipette), puis rétractez-le de manière à ce qu’il soit proche de la surface épithéliale de la méduse.
REMARQUE: La pointe doit être positionnée sur un quadrant de la méduse. Les canaux radiaux de la méduse divisent la cloche de la méduse en quatre quadrants distincts. Le ciblage d’un quadrant permettra d’obtenir une imagerie plus propre, car les gonades et les canaux radiaux sont exclus de la zone de la plaie. - Pulsez de l’air en appuyant sur start sur l’injecteur. Répétez l’impulsion au même endroit deux à quatre fois, selon la largeur de la pointe. Les pointes plus grandes nécessitent moins d’impulsions.
REMARQUE: Un retrait dans l’eau / méduse causé par l’impulsion d’air doit être visible. - Couvrir l’animal blessé avec une lamelle de couverture pour l’imagerie des grandes plaies (Figure 1D,D').
- Suivez les étapes ci-dessous (section 3) pour l’imagerie de la cicatrisation épithéliale.
- Fabriquer une aiguille de microinjection à l’aide d’un extracteur de micropipette et d’un tube capillaire en verre (étape 5.2). Placez l’aiguille de microinjection vide dans un porte-micro-injecteur fixé à un micromanipulateur. Coupez la pointe de l’aiguille de manière à ce que l’ouverture soit d’environ 20-40 μm.

Figure 1 : Couche épithéliale exumbrella intacte et blessée chez Clytia medusa. (A) Dessin animé du corps de Clytia medusa. (A') Méduse exumbrella épithélium intact vu d’en haut. (B) Caricature de microplaies unicellulaires (formes dentelées rouges) avec des cellules épithéliales en bleu. (B') Microplaies unicellulaires. (C) Caricature d’une petite plaie épithéliale (forme dentelée rouge). (C') Petite plaie épithéliale. (D) Caricature d’une grande plaie épithéliale (forme dentelée rouge). (D') Grande plaie épithéliale. Les images ont toutes été obtenues par microscopie DIC. Barres d’échelle en (A'-C'): 50 μm. Barre d’échelle en (D'): 100 μm. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

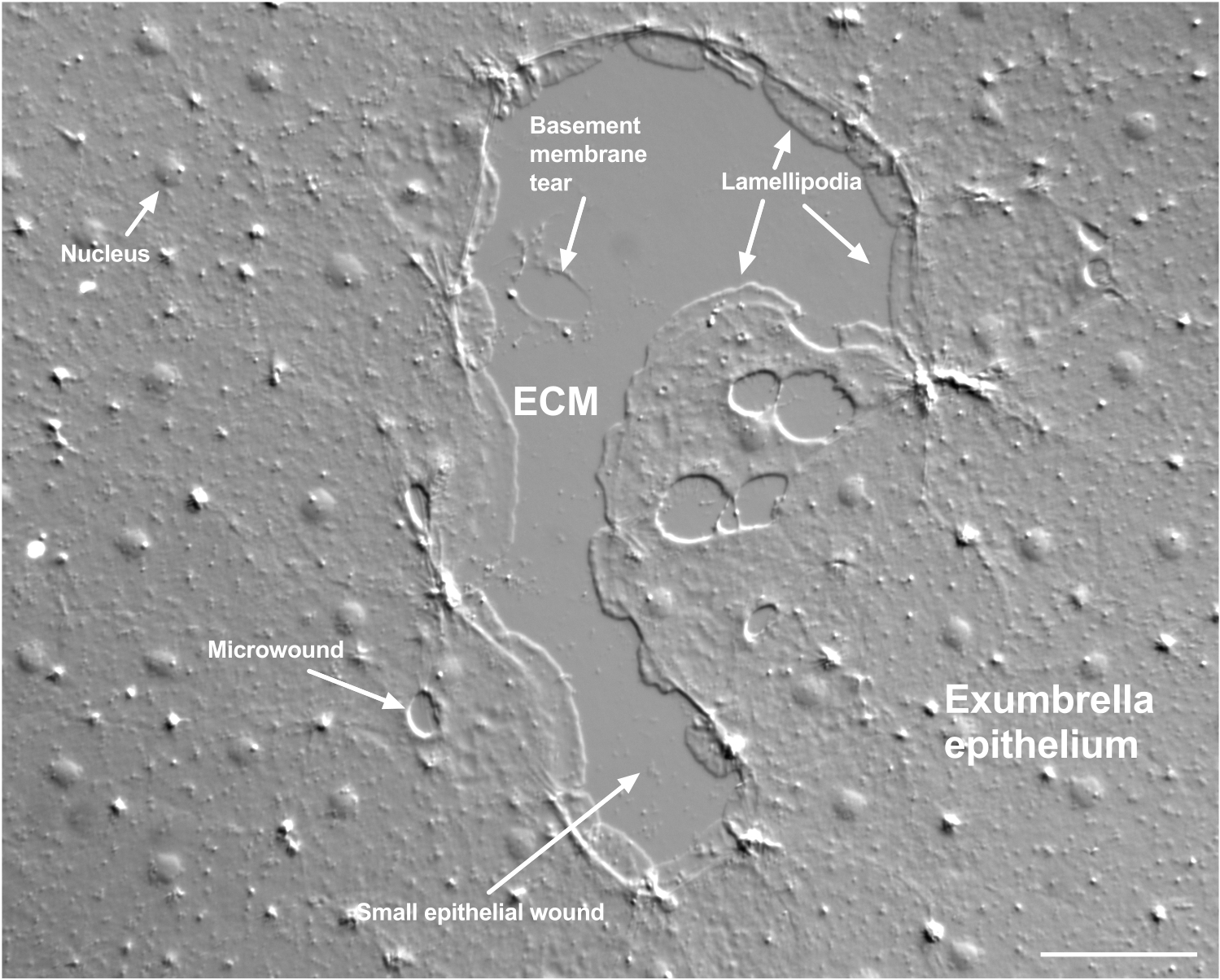
Figure 2 : Plaies de tailles multiples et membrane basale endommagée. Une petite plaie épithéliale exparapluie typique est montrée, avec des étiquettes indiquant des lamellipodes qui se forment à partir de cellules marginales. En outre, des microplaies à l’intérieur et entre les cellules épithéliales sont observées. Notez la petite déchirure de la membrane basale dans la partie supérieure de la plaie. Le film 4 montre la guérison de cette blessure. Barre d’échelle: 50 μm. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
3. Imagerie de la cicatrisation épithéliale
- Assurez-vous que le microscope a été aligné pour l’éclairage Köhler53 et qu’il a été correctement configuré pour la microscopie à contraste interférent différentiel (CIVD)54. Les cellules épithéliales sont presque invisibles avec l’optique standard (Figure 3D,E).
- Ajustez la mise au point sur l’exumbrella. Bien qu’il s’agisse d’une couche mince, les cellules hexagonales doivent être claires.
REMARQUE: L’exumbrella et le sous-parasol sont séparés par une mésogle épaisse qui est soutenue par des fibres verticales. Les cellules sous-parapluies sont dans le même plan focal que les canaux radiaux. Si vous êtes initialement concentré sur la couche de sous-parapluie, ajustez lentement la mise au point à travers la mésogle et les fibres verticales jusqu’à ce que vous trouviez l’exumbrella. - Identifiez manuellement une plaie à imager. Pour les grandes plaies, utilisez un objectif 10x. Pour les plaies plus petites et les plaies unicellulaires, utilisez un objectif 20x.
- Démarrez un programme qui collecte des images sous forme de film en temps réel ou qui collecte une série d’images à intervalles réguliers. Surveillez les progrès pour vous assurer que la zone de la plaie ne dérive pas hors du champ de vision et que les cellules d’intérêt restent concentrées.
- Les plaies unicellulaires se referment en une minute; Par conséquent, imaginez leur fermeture avec un film.
- Pour capturer les détails de la dynamique cellulaire des petites plaies, collectez des images environ toutes les 10 s. La fermeture des petites plaies prend 20-50 minutes selon la taille.
- N’imagez pas les lames non scellées pendant plus de 45 minutes, car l’évaporation de l’eau de la lame au fil du temps entraîne la mort des animaux et la rupture des cellules.
- Pour une observation plus longue, scellez autour de la lamelle de couverture avec de la vaseline pour réduire l’évaporation.
REMARQUE: Certaines méduses peuvent pulser sur la lame, ce qui interfère avec l’imagerie. Dans ce cas, le montage d’animaux dans une dilution 1:10 de méthanesulfonate de 3-aminobenzoate d’éthyle à 1 % (tricaïne), ajustée à un pH de 7,5, dans de l’ASW sert d’anesthésique efficace et n’a aucun effet apparent sur la guérison dans un délai de 1 heure. Cependant, les animaux mourront s’ils sont laissés pendant plusieurs heures à Tricaine.

Figure 3: Création d’une petite plaie dans l’épithélium exumbrellar. (A) Grattage doux de l’exumbrella avec un embout de pipette de 200 μL pour créer une petite plaie épithéliale. (B) La mise en place de la lamelle de couverture est parfois suffisante pour créer de petites plaies épithéliales. (C) Méduse montée sur une glissière de dépression. (D) Petite image de plaie épithéliale sans optique DIC et (E) avec optique DIC. Barres d’échelle : 50 μm Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
4. Analyse
- Préparation des fichiers image
REMARQUE: Pour traiter les fichiers image, utilisez FIJI / ImageJ avec les plugins BioFormat mis à jour.- Réglez l’échelle sur le rapport pixel par micron correct avant d’enregistrer la pile d’images ; Analysez > définissez l’échelle. Ceci est nécessaire pour extraire les mesures de taille réelles dans les analyses en aval.
- Souvent, l’animal dérive légèrement sur la lame du microscope; par conséquent, pour éliminer la dérive dans les films, enregistrez les images à l’aide de l’alignement linéaire de la pile du plugin FIJI avec SIFT. Plugins > l’enregistrement > l’alignement linéaire de la pile avec SIFT.
- Enregistrez la pile enregistrée en tant que fichier .avi. Fichier > enregistrer sous > AVI... Dans la fenêtre contextuelle, définissez la fréquence d’images (les figures animées sont définies ici sur 10 ips) et cliquez sur OK. Utilisez cette sortie pour effectuer une analyse de cicatrisation.
- Analyse de la zone de la plaie
- À l’aide de l’outil lasso de FIJI/ImageJ, définissez le contour de la plaie en traçant les bords de la cellule. Mesurez la zone de la plaie qui vient d’être délimitée avec Commande + M ou CTRL + M.
- Répétez la mesure de la surface de la plaie toutes les 10 images. Les mesures de FIJI/ImageJ peuvent ensuite être tracées à l’aide du Prism 9 (Figure 4).

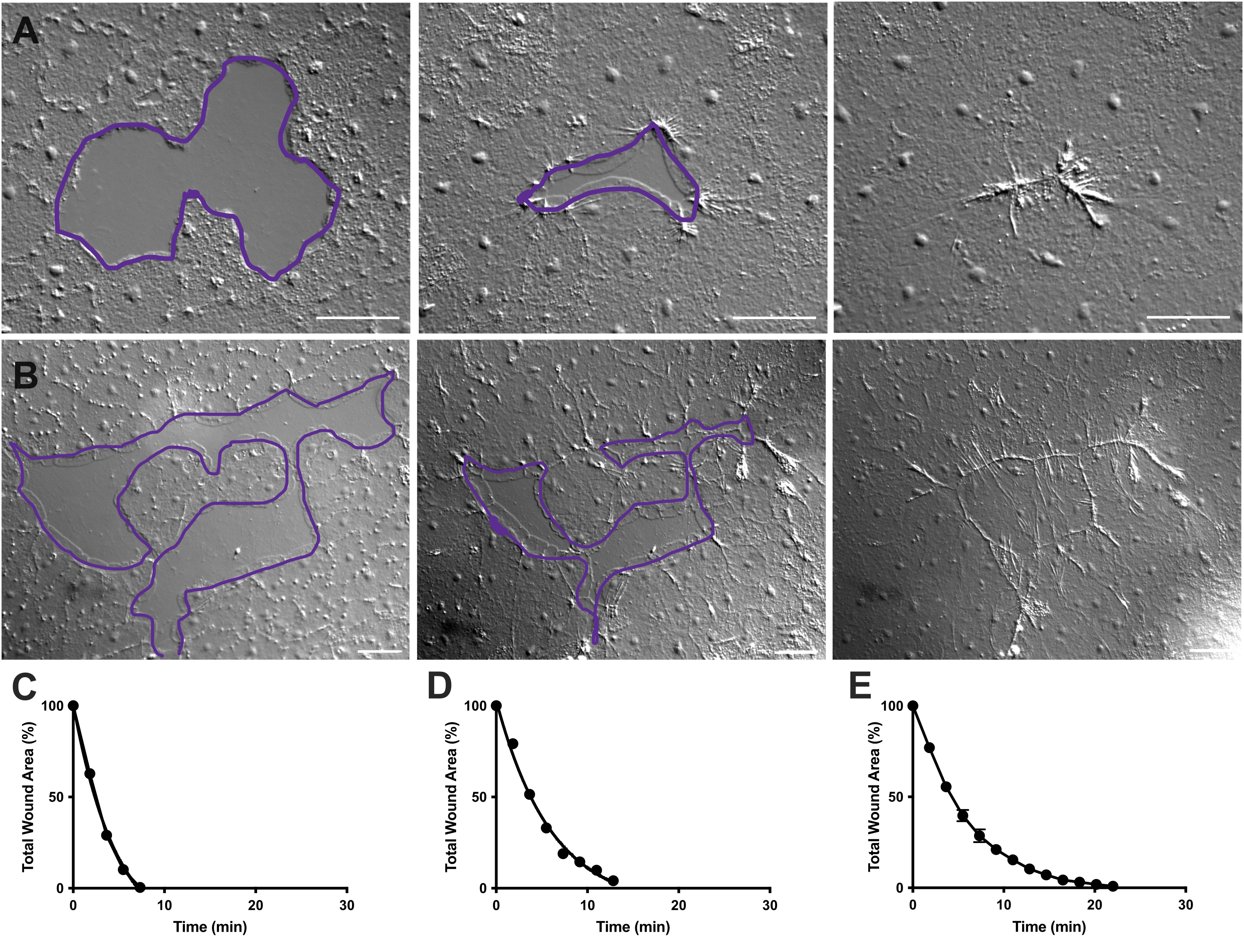
Figure 4 : Analyse de la surface de la plaie dans les petites plaies épithéliales. (A) Exemple d’une petite plaie épithéliale cicatrisant sur 10 min. (B) Exemple d’une plaie épithéliale différente cicatrisant sur 21 min. Les contours violets en A,B sont comparables aux mesures des zones de plaie à l’aide de l’outil lasso dans FIJI/ImageJ. (C) Réduction normalisée de la surface de la plaie au fil du temps en A. (D) Réduction normalisée de la surface de la plaie au fil du temps chez B. (E) Réduction moyenne de la surface de la plaie au fil du temps pour 14 petites plaies. n = 14. Barres d’erreur centrées autour de la moyenne ± SEM. Barres d’échelle : 50 μm Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
5. Injections mésogliennes
- Création d’un plat d’injection
- Préparer le polydiméthylsiloxane (PDMS) en combinant une base PDMS et un agent de durcissement, dans un rapport de 10 parties de base pour 1 partie d’agent de durcissement en poids. Remuer vigoureusement pour mélanger complètement la base et l’agent de durcissement.
- Pour éliminer les bulles, mettez le mélange dans une chambre à vide pendant 15 min. Verser le mélange dans une boîte de Petri de 60 mm avec des bouchons de tubes microcentrifugeuses pour maintenir le moule en place. Placez immédiatement le moule sur les capuchons de tube à une inclinaison de 45° et le ruban adhésif en place. Le moule est constitué de trois lames de verre décalées empilées collées ensemble pour créer des crêtes dans le plat d’injection final.
- Placer la capsule entière, le moule et le mélange dans une étuve à 60 °C pendant 2 h pour durcir l’élastomère. Retirez le moule pour un plat d’injection terminé.
- Traction de micropipettes
- À l’aide d’un extracteur de microélectrodes, concevez un programme de traction. Utilisez un programme en une étape avec une vitesse élevée. La chaleur est approximativement le résultat du test RAMP en verre55,56. Vérifiez les micropipettes résultantes pour de longues conicités cohérentes.
REMARQUE : Utilisez des capillaires en verre borosilicaté à paroi mince d’un diamètre extérieur de 1,0 mm, d’un diamètre intérieur de 0,75 mm et d’une longueur de 10 cm.
- À l’aide d’un extracteur de microélectrodes, concevez un programme de traction. Utilisez un programme en une étape avec une vitesse élevée. La chaleur est approximativement le résultat du test RAMP en verre55,56. Vérifiez les micropipettes résultantes pour de longues conicités cohérentes.
- Injection de colorants et de médicaments
- Faites une aiguille de micro-injection (comme ci-dessus).
- Remplissez l’aiguille de micro-injection à l’aide d’une longue pointe de pipette avec un volume excessif de colorant ou de médicament pour injection dans la méduse.
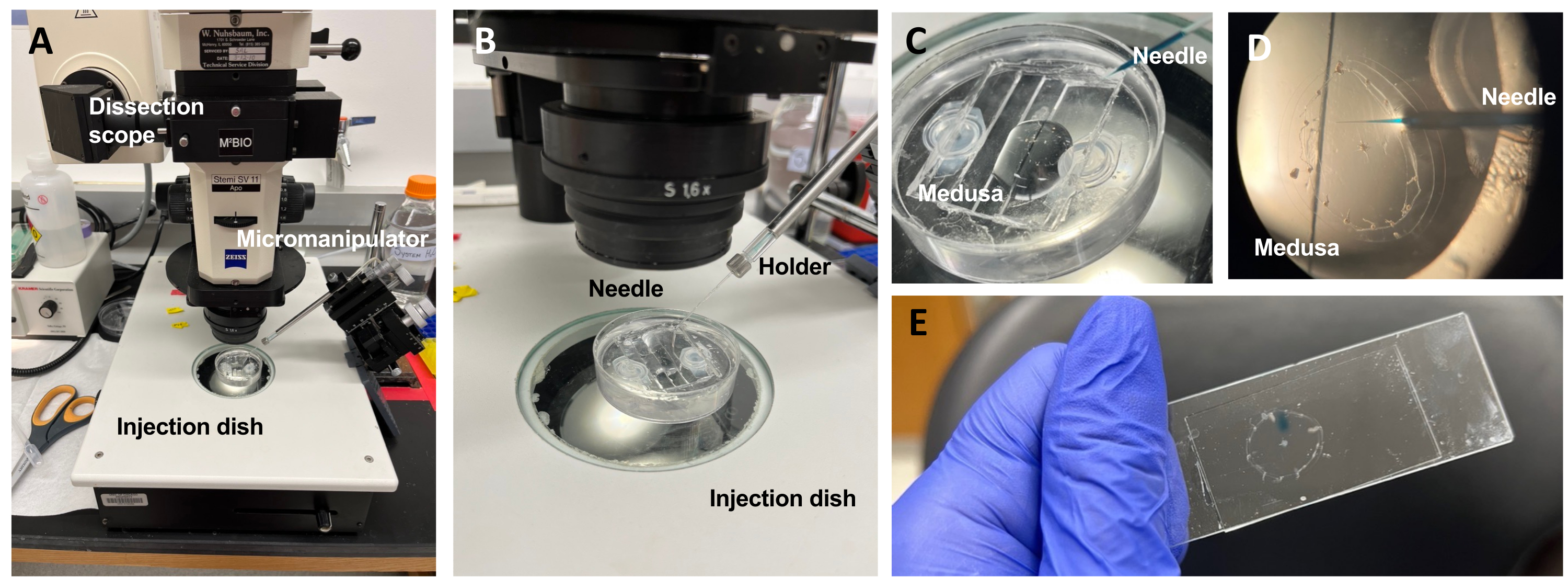
NOTE: Pour Clytia, le diméthylsulfoxyde (DMSO) doit être maintenu à une dilution de <1:100 avec ASW, car des concentrations plus élevées de DMSO empêchent la cicatrisation des plaies. En cas d’injection d’une solution claire, une solution de FCF vert rapide (dilution de 1:100 de FCF vert rapide à 0,1% dans de l’ASW) peut être ajoutée pour visualiser le liquide injecté. - À l’aide d’une pipette de transfert modifiée comme ci-dessus, placer une méduse avec le sous-parasol vers le haut dans une boîte d’injection PDMS avec juste assez d’ASW pour couvrir l’animal (figure 5C). Placez le plat sur la scène d’une lunette de dissection.
REMARQUE: Limiter l’excès de ASW empêche la méduse de nager dans le plat et permet des injections plus réussies. - Concentrez-vous sur la pointe de l’aiguille de micro-injection et avancez-la dans l’eau près de la méduse. Avec le micromanipulateur, enfoncez l’aiguille dans le plat jusqu’à ce qu’il se plie et se casse. Cette ouverture de pointe est d’environ 10-20 μm.
REMARQUE: Cette aiguille peut être utilisée à plusieurs reprises pour les mêmes injections de colorant / médicament ce jour-là. Il est recommandé d’utiliser un embout frais chaque jour et pour des colorants / médicaments séparés. - À l’aide du micromanipulateur, insérez la pointe de l’aiguille à travers le sous-parasol dans la mésogle sans perforer l’exumbrella.
REMARQUE: Un pli / pli de l’épithélium sera perceptible. Une fois l’aiguille insérée dans la méduse, le pli/pliage cesse. - Sur le micro-injecteur, réglez la pression de maintien à zéro et la pression d’éjection sur ≤20 PSI. Injecter dans un ou deux quadrants, en remplissant chacun avec une tache de colorant ou de médicament environ 1/4 de la surface de ce quadrant.
REMARQUE: Selon la taille de la méduse, des volumes plus ou moins grands sont appropriés dans les points d’injection simples. Le remplissage excessif de la méduse entraîne des dommages extrêmes à l’épithélium et même la mort de l’animal. - Selon le colorant ou le médicament injecté, les animaux sont placés dans un bécher d’ASW frais pour permettre la diffusion et l’incubation du colorant ou du médicament.
- Pour l’imagerie, montez la méduse sur une lame de dépression à l’aide d’une pipette de transfert modifiée, en positionnant l’animal de manière à ce que l’exumbrella soit tourné vers le haut (Figure 5). Les animaux peuvent être blessés à ce stade pour tester l’effet d’un réactif injecté.

Figure 5 : Configuration de l’injection pour l’introduction de colorants ou de médicaments dans l’ECM. (A) Configuration de l’injection. (B) Gros plan de la configuration d’injection montrant l’orientation de l’aiguille de microinjection (angle d’environ 45° par rapport à l’animal dans le plat). (C) Gros plan de la capsule d’injection de silicone avec la méduse dans une petite quantité d’ASW pour injection. D) Une aiguille de micro-injection chargée de Fast Green FCF pénétrant dans la mésogle de la méduse par le sous-parasol. E) Post-injection de FCF vert rapide dans une méduse montée. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
Résultats
En suivant les protocoles ci-dessus, des microplaies unicellulaires, de petites plaies et de grandes plaies ont été imagées. Les piles enregistrées de fichiers image ont été enregistrées en tant que fichiers .avi.
Dans le film 1, on peut voir des microplaies se refermer entre et à l’intérieur des cellules (Figure 1 et Figure 2). De petits lamellipodes sont observés pendant la fermeture, suivie d’une contraction et d’une cicatrisation. Les débris sont exclus et rejetés dans l’eau. La guérison est terminée en une minute ou moins.
Dans les films 2 et 3, de petites plaies de formes différentes guérissent par la formation de lamellipodes, l’extension des contacts lamellipodiens et la propagation des cellules au bord de la plaie, comme décrit précédemment40 (Figure 1 et Figure 2). Les cellules situées en rangs derrière les cellules marginales ne participent pas à la cicatrisation des plaies de cette taille et il n’y a pas non plus de migration cellulaire collective. La fermeture rapide et progressive des lacunes épithéliales est suivie d’une contraction tissulaire le long de la couture de la plaie nouvellement formée40. Le taux normalisé de cicatrisation de ces deux plaies, exprimé en pourcentage de la surface d’origine au fil du temps, est présenté (figure 4C,D). Bien qu’il existe une certaine variabilité dans la dynamique de la fermeture des plaies, la moyenne du pourcentage de fermeture de la zone au fil du temps pour 14 plaies de formes variées allant de 0,02 à 0,125 mm2 permet d’établir une courbe moyenne pour la cicatrisation des plaies chez les animaux non traités (figure 4E).
Les dommages à la membrane basale sont clairement visibles lorsqu’ils se produisent (figure 2). Dans le film 4, les cellules à la marge d’une petite plaie dans laquelle il y a des dommages à la membrane basale répartis autour de la zone endommagée, et la fermeture de l’espace est complétée par une contraction de la corde de la bourse.
Si le tissu est déshydraté ou trop endommagé pour être réparé, les mouvements cellulaires peuvent s’arrêter ou toute la feuille de cellules peut éclater (film 5 et film 6). Cela se produit généralement après de longues périodes d’imagerie (45 minutes ou plus). Si l’éclatement cellulaire se produit tôt dans l’imagerie, l’échantillon est jeté.
Comme le montre le film 7, les grandes blessures guérissent en plusieurs étapes. Tout d’abord, le bord de la plaie devient lisse et régulier en raison des contractions à la marge, comme indiqué précédemment57. Ensuite, on voit la formation de lamellipodes à partir des cellules au bord de la plaie, les lamellipodes se déplaçant vers l’avant pour maximiser le contact avec les lamellipodes adjacents. Le suivi des noyaux dans les cellules au bord de la plaie et à plusieurs niveaux derrière les cellules marginales montre que de grandes lacunes se comblent par la migration cellulaire collective40. Les cellules ne se détachent jamais mais se déplacent ensemble comme une feuille.
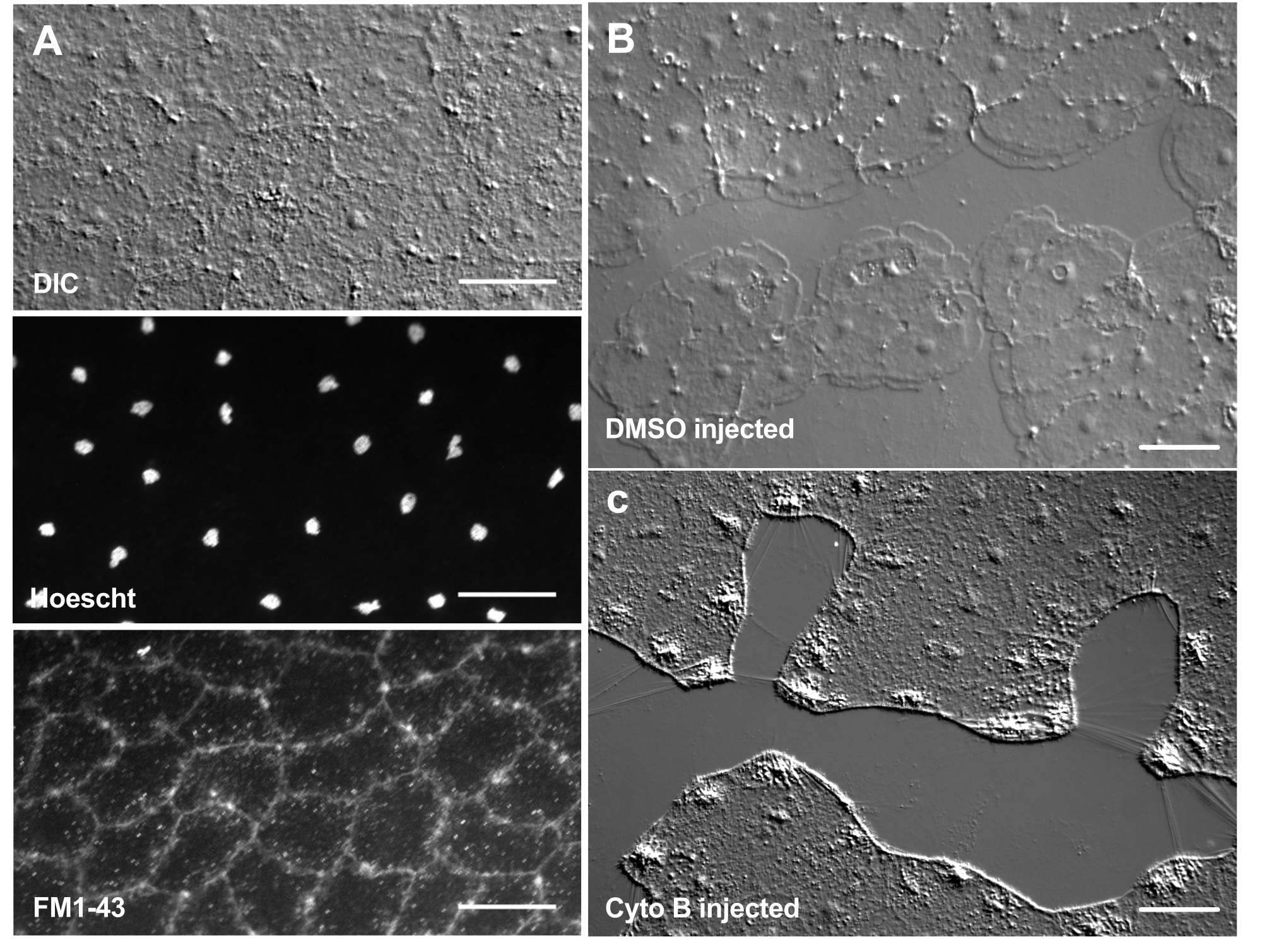
L’introduction de colorants et d’agents pharmacologiques peut être un outil puissant pour disséquer les mécanismes biologiques. De nombreuses substances sont exclues de Clytia (non représentées), probablement en raison de la couche de mucus qui recouvre la surface de l’animal. Cependant, la micro-injection peut être utilisée pour introduire directement des molécules dans l’ECM, perturbant la structure de l’ECM ou perturbant les activités de régulation dans l’ECM. De plus, les colorants et autres molécules sont capables de pénétrer dans les cellules épithéliales par la face basale. Par exemple, la figure 6 montre la coloration nucléaire avec Hoechst, la coloration membranaire avec FM1-43 et l’inhibition de la formation de lamellipodes par la cytochalasine B après que ces réactifs ont été microinjectés dans l’ECM. L’introduction de ces molécules dans l’ECM et les cellules épithéliales avant la blessure permet des expériences qui testent l’effet des outils pharmacologiques sur le processus de guérison.

Figure 6 : Cellules épithéliales de la méduse après micro-injection de colorants ou d’agents pharmacologiques. (A) Cellules épithéliales montrées dans le panneau supérieur 5 minutes après injection avec 20 μM Hoechst (noyaux) et 50 μM FM1-43 (membranes). (B,C) Cicatrisation après injection avec 1:1 000 DMSO témoin (B) ou 100 μM de cytochalasine B (C). Les plaies ont été faites 15 minutes après l’injection. Les images ont été prises 5 minutes après la blessure. La formation de lamellipodes est inhibée par la cytochalasine B. On pense que les « fibres » apparentes souvent observées entre les cellules de la zone de la plaie sont le résultat de la tension qui étire la membrane basale – elles ne se colorent pas avec la phalloïdine (non représentée). Barres d’échelle: 50 μm. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
Film 1: Film accéléré de cicatrisation de microplaies unicellulaires. Temps écoulé : 20 s. Fréquence d’images: 10 ips. Barre d’échelle: 50 μm. Veuillez cliquer ici pour télécharger ce film.
Film 2: Film accéléré d’une petite plaie épithéliale cicatrisant. Temps écoulé : 9 min 54 s. Fréquence d’images: 10 ips. Barre d’échelle: 50 μm. Veuillez cliquer ici pour télécharger ce film.
Film 3: Film en accéléré d’une petite plaie épithéliale cicatrisant. Cette blessure est plus grande et de forme plus irrégulière que celle du film 2. Temps écoulé : 20 min 54 s. Fréquence d’images: 10 ips. Barre d’échelle: 50 μm. Veuillez cliquer ici pour télécharger ce film.
Film 4: Film en accéléré d’une petite plaie et d’une cicatrisation de microplaie avec une déchirure de la membrane basale. Les lamellipodes se propagent autour de la déchirure de la membrane basale, bien qu’ils puissent progresser sur le reste de l’ECM. Une fois que la région de la plaie avec les dommages à la membrane basale est entourée, une contraction de cordon de bourse tire les cellules sur la région. Temps écoulé : 19 min 4 s. Fréquence d’images: 10 ips. Barre d’échelle: 50 μm. Veuillez cliquer ici pour télécharger ce film.
Film 5: Cellules mourant dans une petite plaie épithéliale. La mort cellulaire est probablement due à la déshydratation de l’animal. Temps écoulé : 4 min 24 s. Fréquence d’images: 10 ips. Barre d’échelle: 100 μm. Veuillez cliquer ici pour télécharger ce film.
Film 6: Une petite blessure épithéliale ne parvient pas à compléter la guérison. Temps écoulé : 42 min 32 s. Fréquence d’images: 10 ips. Barre d’échelle: 50 μm. Veuillez cliquer ici pour télécharger ce film.
Film 7: Cicatrisation des grandes plaies épithéliales. Temps écoulé : 25 min 29 s. Fréquence d’images: 10 ips. Barre d’échelle: 100 μm. Veuillez cliquer ici pour télécharger ce film.
Figure supplémentaire 1 : Schémas des dimensions du réservoir Clytia. Visualisation 3D des chars Clytia sur mesure. (A) Vue avant et arrière. (B) Vue latérale. La découpe dans la pièce montrée en vert est recouverte d’une maille de nylon. L’eau pénètre dans le réservoir directement sur le treillis, balaie le treillis et crée un courant circulaire. L’eau sort du système par le trou dans la pièce d’extrémité représentée en bleu. Veuillez cliquer ici pour télécharger ce fichier.
Film supplémentaire 1: Matrice extracellulaire acellulaire dans Clytia. Z-stack de Clytia prélevé par microscopie confocale. La pile se concentre d’abord sur l’exumbrella, puis balaye tous les 10 μm à travers l’ECM jusqu’à l’endoderme et le sous-parapluie de la plaque. Les images utilisant la DIC (à gauche) et la coloration nucléaire de Hoechst (à droite) montrent l’absence de cellules dans l’ECM. Barre d’échelle: 100 μm. Veuillez cliquer ici pour télécharger ce fichier.
Discussion
Ici, la méthodologie est présentée pour l’imagerie des plaies in vivo chez Clytia, un organisme modèle d’invertébrés relativement nouveau40,43,58. Plusieurs facteurs font de ce système un outil de recherche unique et puissant, distinct des autres modèles utilisés pour étudier la cicatrisation et la réépithélialisation des plaies. Tout d’abord, l’épithélium monocouche est attaché à une ECM transparente, ressemblant ainsi à des tests de culture tissulaire in vitro (Figure 1, Figure 2, Figure 3, Figure 4). Comme dans les essais in vitro, les cellules peuvent être imagées à haute résolution. Cependant, contrairement à la culture tissulaire, il existe un environnement cellulaire et une MCE authentiques, de sorte que la cicatrisation des plaies peut être considérée dans le contexte des événements de signalisation complexes qui se produisent chez un animal blessé vivant. Deuxièmement, Clytia manque de réponses inflammatoires, de fibroblastes migrateurs, de système vasculaire et de sang. Cela permet d’étudier le processus de réépithélialisation in vivo en l’absence des événements de chevauchement qui se produisent chez les animaux adultes plus complexes pendant la cicatrisation des plaies59. Troisièmement, l’ECM est acellulaire (film supplémentaire 1) et de grande taille, ce qui permet un accès facile avec une aiguille de micro-injection (Figure 5 et Figure 6). En utilisant cette approche, les chercheurs peuvent tester l’effet des réactifs pharmacologiques qui perturbent la structure ou la signalisation ECM sur la cicatrisation des plaies in vivo. Les réactifs peuvent également être introduits dans les cellules épithéliales et leurs effets sur la cicatrisation in vivo des plaies peuvent être évalués. Quatrièmement, il existe des protocoles pour créer des mutants et des animaux transgéniques dans le système Clytia42,43,44,45. La cicatrisation in vivo peut donc être observée chez les animaux présentant une expression accrue/diminuée des gènes d’intérêt.
Il y a plusieurs étapes critiques dans cette technique. Tout d’abord, comme le montre la figure 3, il est nécessaire d’utiliser un microscope correctement configuré pour la microscopie DIC car les cellules épithéliales plates et transparentes sont presque invisibles avec la microscopie optique standard. Il est également important de développer l’habileté de blesser doucement les animaux afin que l’épithélium soit endommagé sans arracher l’ECM. Un toucher tout aussi doux est nécessaire pour la micro-injection de matériel dans l’ECM, car des dommages importants à l’animal pendant l’injection pourraient compromettre une analyse ultérieure de la cicatrisation de la plaie. Bien qu’il y ait une courbe d’apprentissage à ces techniques, même les étudiants débutants les ont maîtrisées rapidement dans le laboratoire Malamy. En effet, ces protocoles ont été utilisés pour démontrer la migration cellulaire dans les cours de laboratoire de premier cycle à l’Université de Chicago.
Pour une imagerie optimale, il est important que l’animal ne bouge pas et que la zone de plaie choisie ne dérive pas hors du champ de vision. Si les animaux pulsent, le traitement avec Tricaine tel que décrit est très efficace. Pour la dérive, il est souvent nécessaire de repositionner manuellement l’échantillon. Ces mouvements peuvent être éliminés du film final à l’aide de la fonction d’enregistrement dans FIJI/ImageJ.
Une limitation de ce système est qu’il n’est pas possible de créer des plaies identiques, car les plaies varient en forme et en taille en utilisant les méthodes décrites ici. Par conséquent, il peut être difficile de quantifier le taux exact de fermeture de la plaie ou de migration cellulaire. Des marqueurs positionnels tels que les grains de carbone adhèrent à l’ECM exposé chez un animal blessé et peuvent être utilisés pour mesurer le taux de migration cellulaire collective dans les grandes plaies (non représenté). Pour l’analyse de la fermeture des petites plaies, même avec une taille et une forme variables, il existe une gamme limitée de taux de fermeture parmi les plaies de cette taille (figure 4). Il est donc possible de détecter quantitativement les effets des réactifs pharmacologiques promotifs ou répressifs.
Bien que ce travail décrive la caractérisation de la cicatrisation des plaies en utilisant uniquement la microscopie DIC, les mêmes approches peuvent être utilisées pour imager la cicatrisation en utilisant la fluorescence ou la microscopie confocale. Pour aider à cela, des protocoles sont en place pour générer des animaux transgéniques dans lesquels diverses protéines cellulaires et extracellulaires sont marquées par fluorescence. L’imagerie simultanée avec CIVD et fluorescence, combinée à la perturbation de la cicatrisation des plaies à l’aide d’agents pharmacologiques ou de lignées mutantes, constituera une approche puissante pour comprendre les mécanismes qui sous-tendent le processus de cicatrisation des plaies dans l’épithélium.
Déclarations de divulgation
Rien à divulguer.
Remerciements
E.E.L.L. est soutenu par une subvention de la National Science Foundation PRFB 2011010. Nous tenons à remercier Tsuyoshi Momose et Evelyn Houliston pour nous avoir aidés à établir nos colonies Clytia, Jean-Baptiste Reynier pour la collecte des images de guérison des microplaies, Harry Kyriazes pour la construction des réservoirs pseudo-kreisel et Elizabeth Baldo pour le maintien de l’habitat de Clytia. La figure 1B a été créée avec BioRender.com.
matériels
| Name | Company | Catalog Number | Comments |
| 20500 ACE EKE Microscope Fiber Optic Light Source | Kramer Scientific Corporation | ||
| AxioCam 506 mono | ZEISS | 426557-0000-000-MA285 | |
| Capillary tubes | World Precision Instruments | TW1004 | |
| Cytochalasin B | Abcam | ab143482 | |
| Depression slides | Amscope | BS-C12 | |
| DMR with DIC options and fluorescence halogen lamp | Leica | ||
| Ethyl 3-aminobenzoate methanesulfonate | Sigma Aldrich | E10521-10G | |
| Fast Green FCF | Thermo Scientific | A16520-06 | |
| FM1-43 | Biotium | 70022 | Excitation/Emission: 480/598 nm |
| Hoechst 33342 | Thermo Scientific | 62249 | Excitation/Emission: 361/497 nm |
| imageJ | NIH | ||
| Microloader tips (0.5-10 μL /2-20 μL) | Eppendorf | 930001007 | |
| Micromanipulator | World Precision Instruments | 3301R / M3301L | |
| Microscope Cover Glass (22X40-1.5) | Fisherbrand | 12-544-BP | |
| Petri Dish (60 mm x 15 mm) | Fisherbrand | FB085713A | |
| PicoNozzle v2 | World Precision Instruments | 5430-ALL | |
| Pipette puller | Sutter Instrument Co | P-97 | |
| Pneumatic PicoPump | World Precision Instruments | PV820 | |
| Polycarbonate vacuum, desiccator | Bel-art | F42025-0000 | |
| Prism 9 | GraphPad | ||
| STEMI Sv11 Dissection scope | ZEISS | STEMI SV11 | |
| SYLGARD 184 | Dow Silicones | 1024001 | |
| Transfer pipettes | Fisherbrand | 13-711-7M | |
| Z-Hab mini system | Pentair | ||
| ZEN Microscopy software | Zeiss |
Références
- Tyler, S. Epithelium-the primary building block for metazoan complexity. Integrative and Comparative Biology. 43 (1), 55-63 (2003).
- Kurn, H., Daly, D. T. Histology, Epithelial Cell. StatPearls. , (2022).
- Schempp, C., Emde, M., Wölfle, U. Dermatology in the Darwin anniversary. Part 1: Evolution of the integument. Journal of the German Society of Dermatology. 7 (9), 750-757 (2009).
- Lopez-Ojeda, W., Pandey, A., Alhajj, M., Oakley, A. M. Anatomy, Skin (Integument). StatPearls. , (2022).
- Bukowiecki, A., Hos, D., Cursiefen, C., Eming, S. A. Wound-healing studies in cornea and skin: parallels, differences and opportunities. International Journal of Molecular Sciences. 18 (6), 1257(2017).
- Frantz, C., Stewart, K. M., Weaver, V. M. The extracellular matrix at a glance. Journal of Cell Science. 123 (24), 4195-4200 (2010).
- Hynes, R. O. The evolution of metazoan extracellular matrix. The Journal of Cell Biology. 196 (6), 671-679 (2012).
- Fidler, A. L., et al. Collagen IV and basement membrane at the evolutionary dawn of metazoan tissues. eLife. 6, 24176(2017).
- Bullen, T. F., et al. Characterization of epithelial cell shedding from human small intestine. Laboratory Investigation; a Journal of Technical Methods and Pathology. 86 (10), 1052-1063 (2006).
- Watson, A. J. M., et al. Epithelial barrier function in vivo is sustained despite gaps in epithelial layers. Gastroenterology. 129 (3), 902-912 (2005).
- Sonnemann, K. J., Bement, W. M. Wound repair: toward understanding and integration of single-cell and multicellular wound responses. Annual Review of Cell and Developmental Biology. 27, 237-263 (2011).
- Abreu-Blanco, M. T., Verboon, J. M., Parkhurst, S. M. Single cell wound repair. BioArchitecture. 1 (3), 114-121 (2011).
- Fenteany, G., Janmey, P. A., Stossel, T. P. Signaling pathways and cell mechanics involved in wound closure by epithelial cell sheets. Current Biology. 10 (14), 831-838 (2000).
- Pastar, I., et al. Epithelialization in wound healing: a comprehensive review. Advances in Wound Care. 3 (7), 445-464 (2014).
- Rousselle, P., Braye, F., Dayan, G. Re-epithelialization of adult skin wounds: Cellular mechanisms and therapeutic strategies. Advanced Drug Delivery Reviews. 146, 344-365 (2019).
- Bement, W. M., Forscher, P., Mooseker, M. S. A novel cytoskeletal structure involved in purse string wound closure and cell polarity maintenance. The Journal of Cell Biology. 121 (3), 565-578 (1993).
- Vedula, S. R. K., Ravasio, A., Lim, C. T., Ladoux, B. Collective Cell migration: a mechanistic perspective. Physiology. 28 (6), 370-379 (2013).
- Li, L., He, Y., Zhao, M., Jiang, J. Collective cell migration: Implications for wound healing and cancer invasion. Burns & Trauma. 1 (1), 21-26 (2015).
- Bornes, L., Windoffer, R., Leube, R. E., Morgner, J., van Rheenen, J. Scratch-induced partial skin wounds re-epithelialize by sheets of independently migrating keratinocytes. Life Science Alliance. 4 (1), 202000765(2021).
- Theveneau, E., Mayor, R. Collective cell migration of epithelial and mesenchymal cells. Cellular and Molecular Life Sciences. 70 (19), 3481-3492 (2013).
- Haensel, D., Dai, X. Epithelial-to-mesenchymal transition in cutaneous wound healing: where we are and where we are heading. Developmental Dynamics. 247 (3), 473-480 (2018).
- Cordeiro, J. V., Jacinto, A. The role of transcription-independent damage signals in the initiation of epithelial wound healing. Nature Reviews. Molecular Cell Biology. 14 (4), 249-262 (2013).
- Abreu-Blanco, M. T., Watts, J. J., Verboon, J. M., Parkhurst, S. M. Cytoskeleton responses in wound repair. Cellular and Molecular Life Sciences. 69 (15), 2469-2483 (2012).
- Klarlund, J. K., Block, E. R. Free edges in epithelia as cues for motility. Cell Adhesion & Migration. 5 (2), 106-110 (2011).
- Enyedi, B., Niethammer, P. Mechanisms of epithelial wound detection. Trends in Cell Biology. 25 (7), 398-407 (2015).
- Niethammer, P. The early wound signals. Current Opinion in Genetics & Development. 40, 17-22 (2016).
- Jacinto, A., Martinez-Arias, A., Martin, P. Mechanisms of epithelial fusion and repair. Nature Cell Biology. 3 (5), 117-123 (2001).
- Mayor, R., Etienne-Manneville, S. The front and rear of collective cell migration. Nature reviews. Molecular Cell Biology. 17 (2), 97-109 (2016).
- Gupta, S., Yap, A. S. How adherens junctions move cells during collective migration. Faculty Reviews. 10, 56(2021).
- Blair, M. J., Jones, J. D., Woessner, A. E., Quinn, K. P. Skin structure-function relationships and the wound healing response to intrinsic aging. Advances in Wound Care. 9 (3), 127-143 (2020).
- Falanga, V., et al. Chronic wounds. Nature Reviews. Disease Primers. 8 (1), 50(2022).
- Liang, C. -C., Park, A. Y., Guan, J. -L. In vitro scratch assay: a convenient and inexpensive method for analysis of cell migration in vitro. Nature Protocols. 2 (2), 329-333 (2007).
- Jonkman, J. E. N., et al. An introduction to the wound healing assay using live-cell microscopy. Cell Adhesion & Migration. 8 (5), 440-451 (2014).
- Poujade, M., et al. Collective migration of an epithelial monolayer in response to a model wound. Proceedings of the National Academy of Sciences. 104 (41), 15988-15993 (2007).
- Masson-Meyers, D. S., et al. Experimental models and methods for cutaneous wound healing assessment. International Journal of Experimental Pathology. 101 (1-2), 21-37 (2020).
- Park, S., et al. Tissue-scale coordination of cellular behaviour promotes epidermal wound repair in live mice. Nature Cell Biology. 19 (2), 155-163 (2017).
- Tsai, C. -R., Wang, Y., Galko, M. J. Crawling wounded: molecular genetic insights into wound healing from Drosophila larvae. The International Journal of Developmental Biology. 62 (6-7-8), 479-489 (2018).
- Richardson, R., et al. Adult zebrafish as a model system for cutaneous wound-healing research. The Journal of Investigative Dermatology. 133 (6), 1655-1665 (2013).
- Erickson, J. R., Echeverri, K. Learning from regeneration research organisms: The circuitous road to scar free wound healing. Developmental Biology. 433 (2), 144-154 (2018).
- Kamran, Z., et al. In vivo imaging of epithelial wound healing in the cnidarian Clytia hemisphaerica demonstrates early evolution of purse string and cell crawling closure mechanisms. BMC Developmental Biology. 17 (1), 17(2017).
- Home. MARIMBA. , Available from: http://marimba.obs-vlfr.fr/home (2023).
- Chari, T., et al. Whole-animal multiplexed single-cell RNA-seq reveals transcriptional shifts across Clytia medusa cell types. Science Advances. 7 (48), (2021).
- Weissbourd, B., et al. A genetically tractable jellyfish model for systems and evolutionary neuroscience. Cell. 184 (24), 5854-5868 (2021).
- Momose, T., et al. High doses of CRISPR/Cas9 ribonucleoprotein efficiently induce gene knockout with low mosaicism in the hydrozoan Clytia hemisphaerica through microhomology-mediated deletion. Scientific Reports. 8 (1), 11734(2018).
- Houliston, E., Leclère, L., Munro, C., Copley, R. R., Momose, T. Past, present and future of Clytia hemisphaerica as a laboratory jellyfish. Current Topics in Developmental Biology. 147, 121-151 (2022).
- Schmid, V., et al. The extracellular matrix (mesoglea) of hydrozoan jellyfish and its ability to support cell adhesion and spreading. Hydrobiologia. 216 (1), 3-10 (1991).
- Day, R. M., Lenhoff, H. M. Hydra mesoglea: a model for investigating epithelial cell-basement membrane interactions. Science. 211 (4479), 291-294 (1981).
- Zhang, X., et al. The collagens of hydra provide insight into the evolution of metazoan extracellular matrices. The Journal of Biological Chemistry. 282 (9), 6792-6802 (2007).
- Danjo, Y., Gipson, I. K. Actin 'purse string' filaments are anchored by E-cadherin-mediated adherens junctions at the leading edge of the epithelial wound, providing coordinated cell movement. Journal of Cell Science. 111 (22), 3323-3332 (1998).
- Arenas Gómez, C. M., Sabin, K. Z., Echeverri, K. Wound healing across the animal kingdom: Crosstalk between the immune system and the extracellular matrix. Developmental Dynamics. 249 (7), 834-846 (2020).
- Lechable, M., et al. An improved whole life cycle culture protocol for the hydrozoan genetic model Clytia hemisphaerica. Biology Open. 9 (11), (2020).
- Casares, L., et al. Hydraulic fracture during epithelial stretching. Nature Materials. 14 (3), 343-351 (2015).
- Wayne, R. Chapter 4 - Bright-Field Microscopy. Light and Video Microscopy (Third Edition). , 95-116 (2019).
- Murphy, D. B., Davidson, M. W. Fundamentals of Light Microscopy and Electronic Imaging: Second Edition. , John Wiley and Sons. (2012).
- Micropipette Techniques for Electrophysiology. , Available from: https://www.sutter.com/micropipette/cookbook.html (2022).
- Brown, A. L., Johnson, B. E., Goodman, M. B. Making patch-pipettes and sharp electrodes with a programmable puller). Journal of Visualized Experiments. (20), e939(2008).
- Klarlund, J. K. Dual modes of motility at the leading edge of migrating epithelial cell sheets. Proceedings of the National Academy of Sciences. 109 (39), 15799-15804 (2012).
- Houliston, E., Momose, T., Manuel, M. Clytia hemisphaerica: a jellyfish cousin joins the laboratory. Trends in Genetics. 26 (4), 159-167 (2010).
- Rodrigues, M., Kosaric, N., Bonham, C. A., Gurtner, G. C. Wound healing: a cellular perspective. Physiological Reviews. 99 (1), 665-706 (2019).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationExplorer plus d’articles
This article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.