
Method Article
Характеристика заживления эпителиальных ран in vivo с использованием книдарийского модельного организма Clytia hemisphaerica
В этой статье
Резюме
В данной работе описан способ создания ран в эпителии живой медузы Clytia hemisphaerica и изображения заживления ран с высоким разрешением in vivo. Дополнительно представлена методика введения красителей и лекарственных препаратов для возмущения сигнальных процессов в эпителиальных клетках и внеклеточном матриксе при заживлении ран.
Аннотация
Все органы животных, от кожи до глаз и кишечника, покрыты листами эпителиальных клеток, которые позволяют им поддерживать гомеостаз, защищая их от инфекции. Поэтому неудивительно, что способность заживлять эпителиальные раны имеет решающее значение для всех многоклеточных. Заживление эпителиальных ран у позвоночных включает перекрывающиеся процессы, включая воспалительные реакции, васкуляризацию и реэпителизацию. Регуляция этих процессов включает сложные взаимодействия между эпителиальными клетками, соседними клетками и внеклеточным матриксом (ECM); ECM содержит структурные белки, регуляторные белки и активные малые молекулы. Эта сложность, наряду с тем фактом, что большинство животных имеют непрозрачные ткани и недоступные ECM, затрудняет изучение заживления ран на живых животных. Поэтому большая работа по заживлению эпителиальных ран выполняется в системах культивирования тканей с одним типом эпителиальной клетки, покрытым монослоем на искусственном матриксе. Clytia hemisphaerica (Clytia) является уникальным и захватывающим дополнением к этим исследованиям, позволяя изучать заживление эпителиальных ран у интактного животного с подлинным ECM. Эктодермальный эпителий Clytia представляет собой один слой крупных плоских эпителиальных клеток, что позволяет получать изображения с высоким разрешением с использованием дифференциально-интерферирующей контрастной (ДВС-микроскопии) у живых животных. Отсутствие мигрирующих фибробластов, сосудистой сети или воспалительных реакций позволяет проанализировать критические события при реэпителизации in vivo. Можно проанализировать заживление различных типов ран, включая одноклеточные микрораны, мелкие и большие эпителиальные раны, а также раны, повреждающие базальную мембрану. В этой системе можно наблюдать образование ламеллиподий, сокращение кисетной нити, растяжение клеток и коллективную миграцию клеток. Кроме того, фармакологические агенты могут быть введены через ECM для модификации клеточных: ECM-взаимодействий и клеточных процессов in vivo. В этой работе показаны методы создания ран у живых клиций, съемки видеороликов о заживлении и зондирования механизмов заживления путем микроинъекций реагентов в ECM.
Введение
Листы эпителиальных клеток покрывают внешнюю поверхность всех многоклеточных, выстилают внутренние органы и делят тело животного на отдельные отсеки. Эпителий также отделяет внутреннее тело от внешней среды и защищает его от повреждений и инфекций. Следовательно, появление эпителиальных слоев было существенной частью эволюции многоклеточных животных, а эпителиальные слои наблюдаются у всех животных от позвоночных до большинства базальных многоклеточных1. Эпителий некоторых органов представляет собой единый монослой, например, в воздушных мешочках легких, кровеносных сосудах и кишечнике2, а также в эпидермисе беспозвоночных, таких как планарии и книдарии3. В других тканях, таких как кожа4 и роговица5 позвоночных, эпителий стратифицирован, что означает, что есть несколько слоев эпителиальных клеток2. Во всех случаях самый базальный эпителиальный слой прикрепляется к базальной мембране, белковому листу, который образует специализированную область внеклеточного матрикса (ECM)6,7,8.
Разрывы в эпителии должны быть быстро восстановлены, чтобы воссоздать непрерывный эпителиальный лист. Повреждение эпителия происходит во время естественных процессов, таких как отшелушивание эпителиальных клеток в кишечнике,9,10 а также в результате воспаления или физической травмы. Когда одна эпителиальная клетка повреждена, она должна либо восстановиться, либо быть устранена, чтобы позволить окружающим клеткам прикреплиться друг к другу и закрыть отверстие11,12. В ранах, превышающих размер одной клетки, эпителиальные клетки должны двигаться, чтобы достичь друг друга и восстановить лист13. Это может быть достигнуто путем распространения клеток, если промежутки малы или может потребоваться миграция эпителиальных клеток с краев раны, чтобы закрыть раневую щель; Этот последний процесс называется реэпителизацией14,15. В эмбриональных тканях эпителиальные клетки распространяются и мигрируют к закрытым ранам или вытягиваются через щель за счет сокращения актомиозиновых кабелей, которые образуются между клетками на краю раны, в механизме, напоминающем кошельковую нить16. Во многих тканях взрослого человека реэпителизация включает миграцию когерентных клеточных листов, где клетки сохраняют свои соединения с соседними клетками14,17,18. В других тканях клеточные связи демонтируются, и эпителиальные клетки ведут себя больше как мезенхимальные клетки, перемещаясь скоординированным, но независимым образом в область раны во время повторной эпителизации 14,19,20,21.
Движения эпителиальных клеток регулируются сложными взаимодействиями между мигрирующими клетками, а также между клетками и ECM. Несмотря на то, что существует огромное количество экспериментальной литературы, посвященной механизмам раневой активации эпителиальных клеток и последующей миграции, многое еще предстоит открыть. Например, первоначальный сигнал, который активирует эпителиальные клетки для миграции в ответ на рану, не был окончательно идентифицирован 22, и не до конца понятно, как актин перераспределяется для создания ламеллиподий на стороне эпителиальных клеток, ближайших к ране 22,23,24,25,26,27. Коллективная миграция клеток требует, чтобы информация от клеток в ране передавалась клеткам дистальнее раны, и путь связи до сих пор неясен28. Ячейка: клеточные соединения и ячейка: ECM насадки должны быть разобраны и преобразованы по мере того, как ячейки в листе перестраиваются, но регулирование этого процесса плохо изучено14,29. Достижение прогресса в этих и других связанных с ними вопросах важно не только как фундаментальная биологическая проблема, но и из-за клинической значимости правильного заживления ран. Заболевания, которые нарушают способность эпителиальных клеток правильно мигрировать, приводят к хроническим ранам; примером может служить генетическое заболевание буллезный эпидермолиз, при котором гены, участвующие в прикреплении эпителиальных клеток к ECM, мутируют, в результате чего кожа становится хрупкой, которая шелушится и покрывается волдырями. Реэпителизация также нарушается в естественно стареющих тканях30,31. Поэтому лучшее понимание имеет важное значение для разработки вмешательств для улучшения результатов заживления ран.
Миграция эпителиальных клеток при заживлении ран была изучена с использованием как подходов in vitro, так и модельных организмов. Большинство исследований заживления ран и механизмов миграции клеток было проведено в культуре тканей, где монослои одного типа эпителиальных клеток выращиваются на субстрате, замещающем ECM. Клеточные монослои либо царапают, либо выращивают с помощью трафаретов для создания промежутков определенных форм и размеров, а затем наблюдают32,33,34. Модель in vitro позволяет идеально визуализировать поведение клеток, а также дает возможность изменять качества субстрата, подвергать клетки воздействию лекарств и абиотических и биотических факторов, а также трансфицировать клетки конструкциями, которые экспрессируют или подавляют различные интересующие гены. Однако этот редукционистский подход может не охватить некоторые важные параметры, участвующие в поведении эпителиальных клеток в контексте in vivo, включая связь между различными типами клеток и сигнальные события, которые происходят в ECM11. Модели in vivo обеспечивают аутентичный контекст раны с несколькими типами клеток, перекрывающимися сигнальными путями и сложным ECM35. Одной из таких моделей для исследований заживления ран является мышь19, в которой последние достижения позволили исследователям наблюдать эпидермальные клетки во время заживления ран на всю толщину у живых животных36. Однако мышь и другие системы in vivo создают проблемы для изучения реэпителизации. Во-первых, большое преимущество наблюдения за поведением клеток в естественном контексте уравновешивается сложностью временно перекрывающихся событий, которые происходят во время заживления ран позвоночных, включая свертывание крови, рекрутирование иммунных клеток и воспаление, рекрутирование фибробластов и дедифференцировку клеток, реваскуляризацию и ремоделирование ECM. Кроме того, непрозрачные ткани затрудняют визуализацию. Системы эпидермиса личинок дрозофилы и рыбок данио-рерио 37,38 преодолели некоторые из этих трудностей из-за их относительной простоты39.
Наша лаборатория недавно представила новую модель для изучения заживления эпителиальных ран: форма медузы (медузы) гидрозоя книдария Clytia hemisphaerica (Clytia)40. Clytia является новым модельным организмом с полностью секвенированным и аннотированным геномом41, транскриптомом 42 одноклеточной РНКseq и протоколами модификации генома (мутагенез и трансгенез)43,44,45. Книдарии являются одной из старейших сохранившихся линий с эпителиальными слоями, поэтому понимание заживления ран книдария дает представление о предковых путях, которые обеспечивали целостность эпителия. Для тех путей, которые были сохранены на протяжении всего древа жизни, Clytia предлагает захватывающую новую систему для изучения динамики эпителиальных клеток и функциональной регуляции заживления ран in vivo.
Эпителий, покрывающий верхнюю поверхность Clytia medusa (exumbrella), представляет собой монослой прозрачных плоских эпителиальных клеток шириной примерно 50 мкм и толщиной 1-2 мкм (рис. 1). Они прикреплены к ECM, называемому мезоглеей — «желе» медузы. Мезоглея композиционно сходна с ECM, обнаруженным у других животных 46,47,48, включая позвоночных, имеет базальную мембрану 40 и полностью прозрачна. Эпителиальный слой у Clytia medusa можно легко поцарапать или ранить (см. Ниже). Простота и прозрачность эпителия и ECM позволяют получать изображения клеток и их движений во время заживления с высоким разрешением. Недавно Kamran et al. подробно охарактеризовали заживление небольших ран в эпителии Clytia40. Было продемонстрировано, что заживление у Clytia происходит за счет ползания клеток на основе ламеллиподий, распространения клеток и коллективной миграции клеток, а также закрытия кошельковых нитей, что более типично для эмбриональных систем (хотя ранее наблюдалось в структурах взрослых животных, таких как роговица49). Заживление ран Clytia происходит чрезвычайно быстро, как это было замечено в других системах, у которых отсутствует воспалительная реакция40,50. Заживление зонтика Clytia полностью зависит от движений существующих эпителиальных клеток — ни одна клетка не размножается и не мигрирует через ECM к месту раны (Дополнительный фильм 1). Все эти данные свидетельствуют о том, что Clytia является полезной модельной системой для изучения заживления эпителиальных ран. Действительно, легкость визуализации эпителиальных клеток в Clytia во время заживления ран привела к открытию, что ламеллиподии эпителиальных клеток расширяются и распространяются по областям открытого ECM до тех пор, пока есть неповрежденная базальная мембрана; Если базальная мембрана повреждена, эпителиальное заживление переключается на механизм40 кисетной струны. Это была первая демонстрация механизма, лежащего в основе решения о закрытии с помощью ползания на основе ламелиподий по сравнению с закрытием кошелька, что подчеркивает важность специфических взаимодействий клеток: ECM в заживлении и наблюдении за клетками в их естественном контексте.
Ниже описаны протоколы для создания и визуализации одноклеточных микроран, небольших ран, которые закрываются в основном за счет распространения клеток, и больших ран, для закрытия которых требуется коллективная миграция клеток. Кроме того, описан протокол введения малых молекул в ECM и эпителиальные клетки, что позволяет экспериментально возмущать предполагаемые регуляторные пути заживления ран.
протокол
1. Животноводство
- Поддерживайте колонии полипов Clytia на предметных стеклах микроскопа и медузах в искусственной морской воде (ASW) при температуре 18 ° C в системе рыбок данио, с 2-литровыми резервуарами для рыбок данио-рерио для колоний полипов и изготовленными на заказ 5-литровыми псевдо-крейзельскими резервуарами для медуз (дополнительный рисунок 1)51. ASW состоит из 4% Instant Ocean в деионизированном (DI) H2O.
- Ежедневно кормите животных 2-3-дневной артемией, как описано51.
ПРИМЕЧАНИЕ: Визуализация заживления ран легче, если животные недавно не были накормлены, так как из кишечника в поле зрения попадает меньше мусора. - Собирайте детенышей медуз из установленных колоний полипов по мере необходимости, помещая колонии в 2-литровый стакан, наполненный 1 л ASW на ночь. Используйте 2-3-недельных медуз женского пола для всех экспериментов по заживлению ран. Размножение Clytia было подробно описано в другом месте51.
2. Ранение
- Создание микроран внутри и между клетками (20-500мкм 2 )
- Создайте модифицированную пипетку для переноса, отрезав кончик ножницами, чтобы сделать отверстие большего размера (диаметр 0,5-0,7 см).
ПРИМЕЧАНИЕ: Отверстие в пипетке должно быть достаточно широким, чтобы не повредить животное. - Используя модифицированную пипетку для переноса, поместите медузу на углубление горкой зонтиком медузы вверх, чтобы покрыть животное ровно настолько, чтобы покрыть животное.
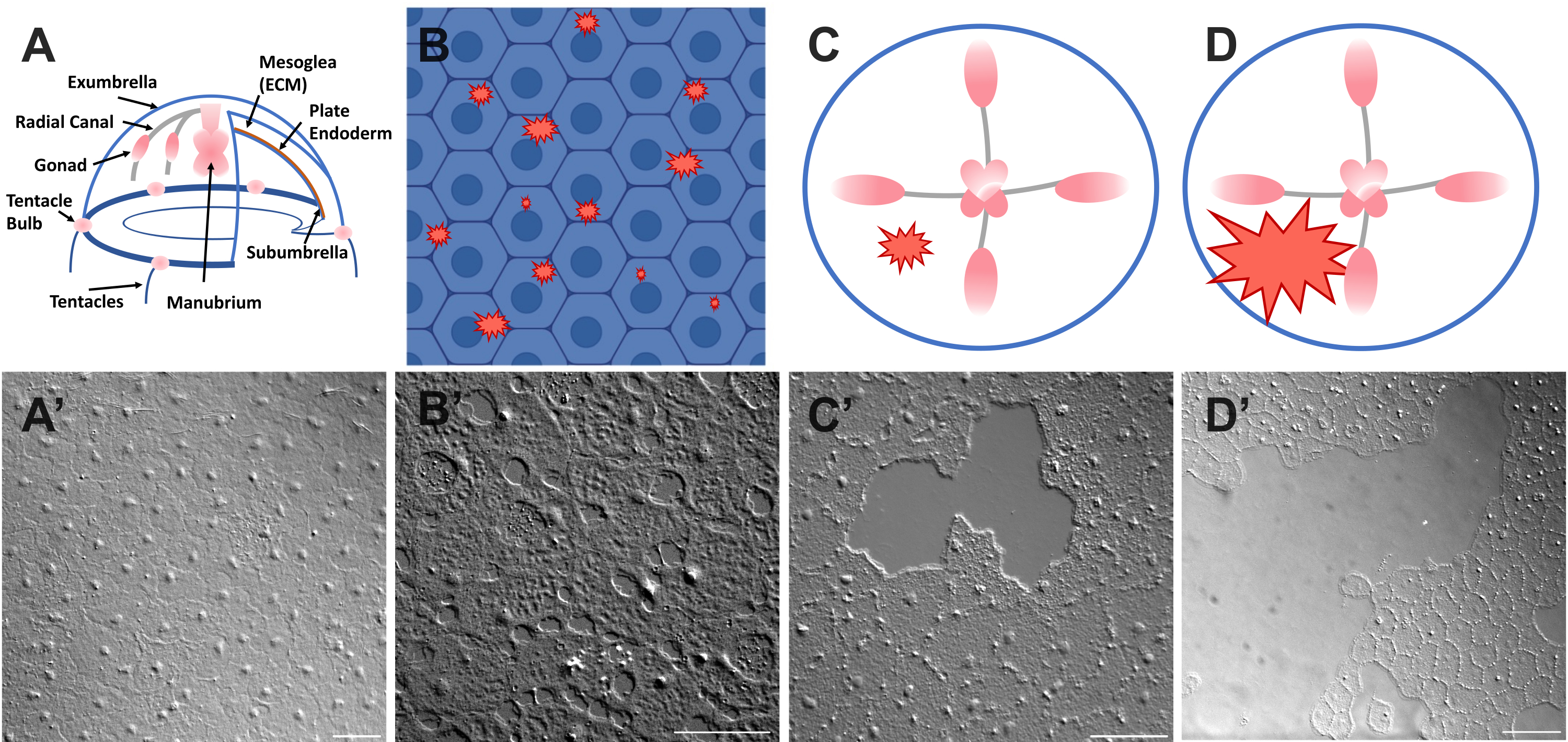
- Немедленно наклейте покровное стекло на животное и изображение (см. Ниже описание изображения). Покровное стекло сжимает мезоглею, и отскок сжатой ткани создает силу, которая слегка раздвигает клетки52. Это сразу же проявляется в виде промежутков между каждой клеткой и повреждений внутри некоторых клеток (рис. 1B, B', рис. 2 и рис. 3A-C).
- Создайте модифицированную пипетку для переноса, отрезав кончик ножницами, чтобы сделать отверстие большего размера (диаметр 0,5-0,7 см).
- Создание небольших эпителиальных ран (0,02-0,125мм2)
- Используя модифицированную пипетку для переноса (как указано выше), поместите медузу на углубление зонтиком медузы вверх.
- Используя наконечник пипетки объемом 200 мкл, аккуратно поцарапайте поверхность медузы. Легкое царапание также может привести к разрывам в базальной мембране, которые легко видны22. Накройте животное покровным стеклом для визуализации. В качестве альтернативы, размещение покровного стекла иногда достаточно для создания небольших эпителиальных ран даже без царапин (рис. 1C, C', рис. 2 и рис. 3A-C).
ПРИМЕЧАНИЕ: Не нажимайте при царапании поверхности медузы, так как это повреждает ECM и создает неровную поверхность — эпителиальные клетки, мигрирующие по неровной поверхности, труднее держать в фокусе.
- Создание больших эпителиальных ран (0,5-0,9мм2)
- Сделайте иглу для микроинъекций, используя съемник микропипетки и стеклянную капиллярную трубку (шаг 5.2). Поместите пустую иглу для микроинъекций в держатель микроинжектора, прикрепленный к микроманипулятору. Отрежьте кончик иглы так, чтобы отверстие составляло примерно 20-40 мкм.
ПРИМЕЧАНИЕ: Разрезанные иглы для больших эпителиальных ран можно хранить и использовать повторно для повышения согласованности между экспериментами. - Установите давление удержания микроинжектора на ноль и установите давление выброса примерно на 20 фунтов на квадратный дюйм. Настройте микроинжектор на подачу импульса воздуха в 2 с.
ПРИМЕЧАНИЕ: Возможно, потребуется отрегулировать давление выталкивания в зависимости от диаметра отверстия иглы (т. Е. Меньшие наконечники будут использовать более высокое давление, а большие наконечники будут использовать более низкое давление). - Поместите медузу зонтиком вверх на углубление на сцене препарирующей прицела, чтобы противолодочная лошадь могла покрыть животное. С помощью микроманипулятора отрегулируйте кончик иглы микроинъекции так, чтобы он находился чуть выше воды. Для этого осторожно опустите наконечник в воду (вода может попасть в наконечник пипетки), затем втяните его так, чтобы он оказался близко к эпителиальной поверхности медузы.
ПРИМЕЧАНИЕ: Наконечник должен быть расположен над одним квадрантом медузы. Радиальные каналы медузы делят колокол медузы на четыре отдельных квадранта. Нацеливание на квадрант приведет к более чистому изображению, так как гонады и лучевые каналы исключаются из области раны. - Нагнетайте воздух, нажимая кнопку «Пуск» на инжекторе. Повторите пульс в одном и том же месте от двух до четырех раз, в зависимости от ширины наконечника. Большие наконечники требуют меньшего количества импульсов.
ПРИМЕЧАНИЕ: Вмятина в воде/медузе, вызванная импульсом воздуха, должна быть видна. - Накройте раненое животное покровным стеклом для визуализации больших ран (рис. 1D, D').
- Выполните следующие действия (раздел 3) для визуализации заживления эпителиальных ран.
- Сделайте иглу для микроинъекций, используя съемник микропипетки и стеклянную капиллярную трубку (шаг 5.2). Поместите пустую иглу для микроинъекций в держатель микроинжектора, прикрепленный к микроманипулятору. Отрежьте кончик иглы так, чтобы отверстие составляло примерно 20-40 мкм.

Рисунок 1: Интактный и раненый эпителиальный слой Exumbrella у Clytia medusa. (А) Карикатурное изображение тела медузы Клития. (А') Интактный эксзонтичный эпителий медузы, вид сверху. (B) Карикатура на одноклеточные микрораны (красные зубчатые формы) с эпителиальными клетками синего цвета. (В') Одноклеточные микрораны. (C) Карикатура на маленькую эпителиальную рану (красная неровная форма). (С') Небольшая эпителиальная рана. (D) Карикатура на большую эпителиальную рану (красная неровная форма). (Д') Большая эпителиальная рана. Все изображения были получены с помощью ДВС-микроскопии. Масштабные линейки в (A'-C'): 50 мкм. Масштабная линейка в (D'): 100 мкм. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

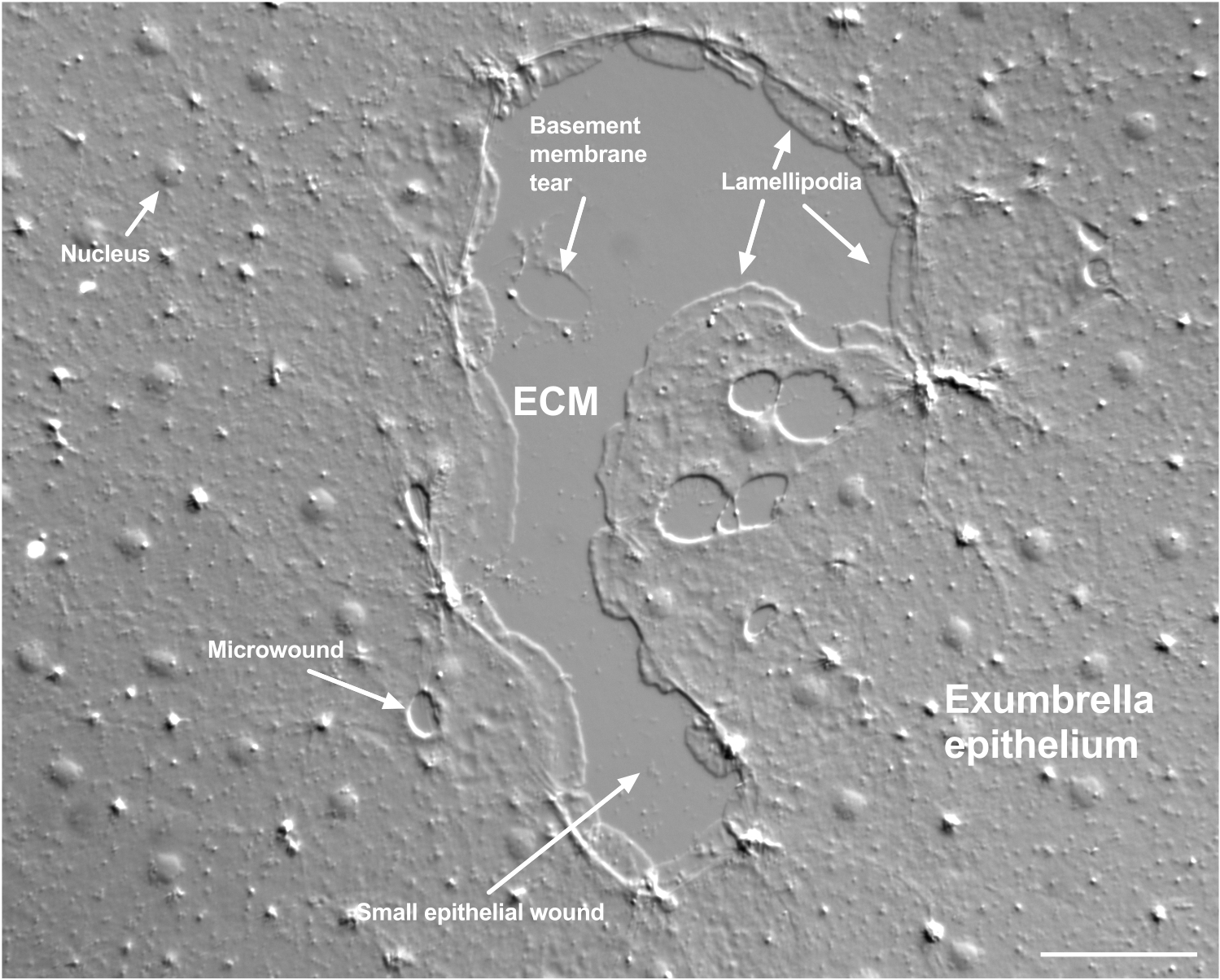
Рисунок 2: Раны разного размера и поврежденная базальная мембрана. Показана типичная маленькая эпителиальная рана с метками, указывающими на ламеллиподии, которые образуются из краевых клеток. Кроме того, видны микрораны внутри и между эпителиальными клетками. Обратите внимание на небольшой разрыв базальной мембраны в верхней части раны. В фильме 4 показано заживление этой раны. Масштабная линейка: 50 мкм. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
3. Визуализация заживления эпителиальных ран
- Удостоверьтесь, что микроскоп выровнен для освещения53 по Кёлеру и что он правильно настроен для дифференциально-интерферирующей контрастной (ДВС-микроскопии)54. Эпителиальные клетки практически незаметны при стандартной оптике (рис. 3D,E).
- Отрегулируйте фокус на зонт. Хотя это тонкий слой, гексагональные клетки должны быть четкими.
ПРИМЕЧАНИЕ: Зонтик и субзонт разделены толстой мезоглеей, которая поддерживается вертикальными волокнами. Субзонтичные клетки находятся в той же фокальной плоскости, что и лучевые каналы. Если изначально фокусироваться на субзонтичном слое, то медленно регулируйте фокус через мезоглею и вертикальные волокна, пока не найдете эксзонт. - Вручную определите рану по изображению. Для больших ран используйте 10-кратный объектив. Для небольших ран и одноклеточных ран используйте 20-кратный объектив.
- Запустите программу, которая собирает изображения в виде фильма в режиме реального времени или серию изображений через равные промежутки времени. Следите за прогрессом, чтобы убедиться, что область раны не выходит из поля зрения и что интересующие клетки остаются в фокусе.
- Одноклеточные раны закрываются в течение минуты; Поэтому представьте их закрытие фильмом.
- Чтобы запечатлеть детали клеточной динамики небольших ран, собирайте изображения примерно каждые 10 секунд. Закрытие небольших ран занимает 20-50 минут в зависимости от размера.
- Не снимайте незапечатанные предметные стекла более 45 мин, так как испарение воды с предметного стекла со временем приводит к гибели животных и разрыву клеток.
- Для более длительного наблюдения запечатайте покровное стекло вазелином, чтобы уменьшить испарение.
ПРИМЕЧАНИЕ: Некоторые медузы могут пульсировать на предметном стекле, что мешает визуализации. В этом случае размещение животных в разведении 1:10 1% этил-3-аминобензоата метансульфоната (трикаина), доведенного до рН 7,5, в ASW служит эффективным анестетиком и не оказывает видимого влияния на заживление в течение 1 часа. Однако животные погибнут, если оставить их на несколько часов в Трикейне.

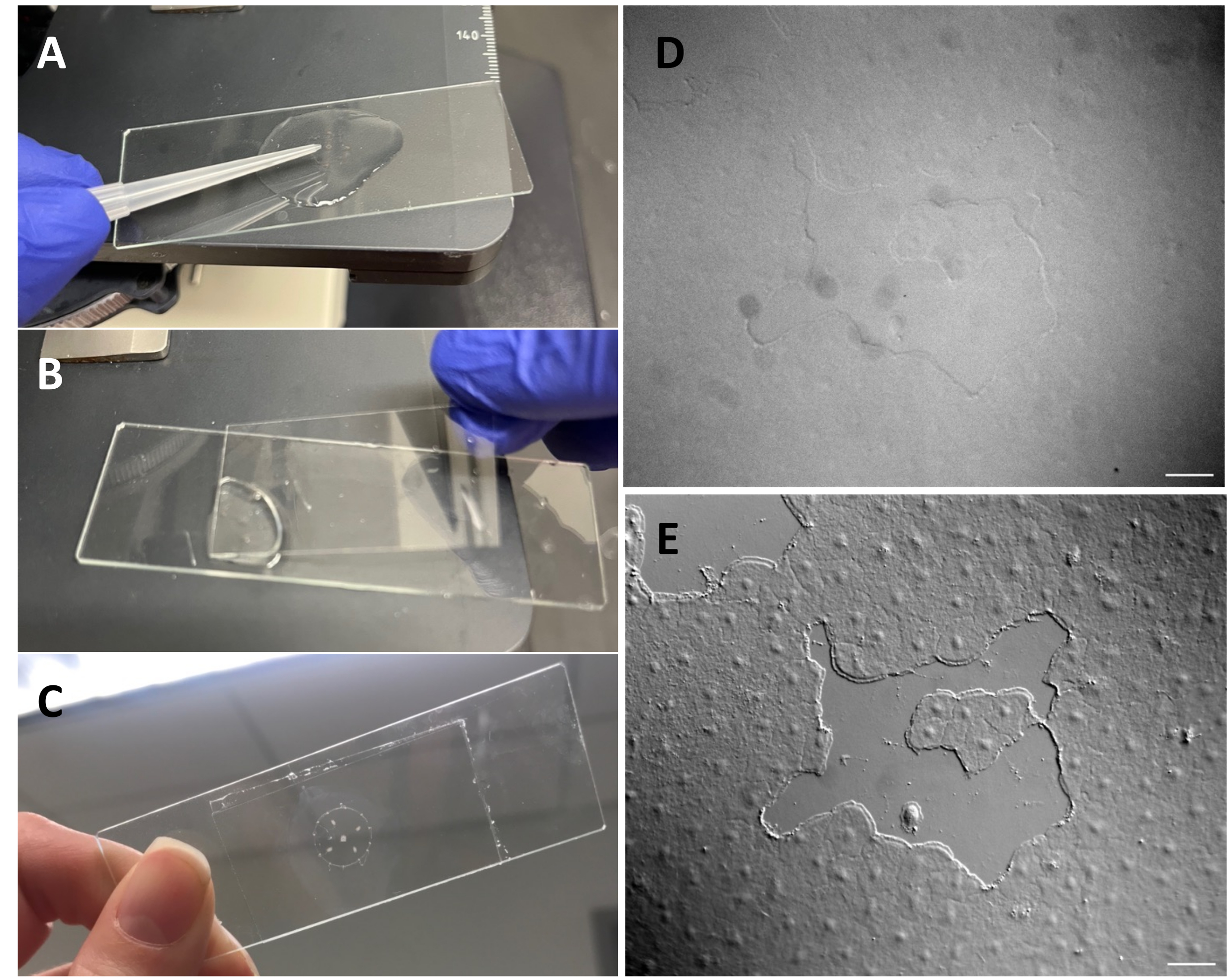
Рисунок 3: Создание небольшой раны в эпителии экзонтика. (A) Осторожное расчесывание зонта наконечником пипетки объемом 200 мкл для создания небольшой эпителиальной раны. (B) Иногда достаточно поместить покровное стекло для создания небольших эпителиальных ран. (C) Медуза, установленная на горке депрессии. (D ) Изображение небольшой эпителиальной раны без ДВС-оптики и (Е) с ДВС-оптикой. Масштабные линейки: 50 мкм Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
4. Анализ
- Подготовка файлов изображений
ПРИМЕЧАНИЕ: Для обработки файлов изображений используйте FIJI/ImageJ с обновленными плагинами BioFormat.- Установите правильное соотношение пикселей на микрон перед регистрацией стека изображений; Проанализируйте > установите масштаб. Это необходимо для извлечения фактических измерений размеров при последующем анализе.
- Часто животное слегка дрейфует по предметному стеклу микроскопа; поэтому, чтобы устранить дрейф в фильмах, зарегистрируйте изображения с помощью плагина FIJI для линейного выравнивания стека с помощью SIFT. Плагины > регистрации > линейного выравнивания стека с помощью SIFT.
- Сохраните зарегистрированный стек в виде файла .avi. Файл > Сохранить как > AVI... Во всплывающем окне установите частоту кадров (анимированные фигуры здесь установлены на 10 кадров в секунду) и нажмите «ОК». Используйте этот вывод для выполнения анализа заживления ран.
- Анализ области раны
- С помощью инструмента «Лассо» в FIJI/ImageJ обведите контур раны, обведя края ячеек. Измерьте область раны, которая была только что обведена с помощью клавиш Command+M или CTRL+M.
- Повторяйте измерение площади раны каждые 10 кадров. Затем измерения с помощью FIJI/ImageJ можно нанести на график с помощью Prism 9 (рис. 4).

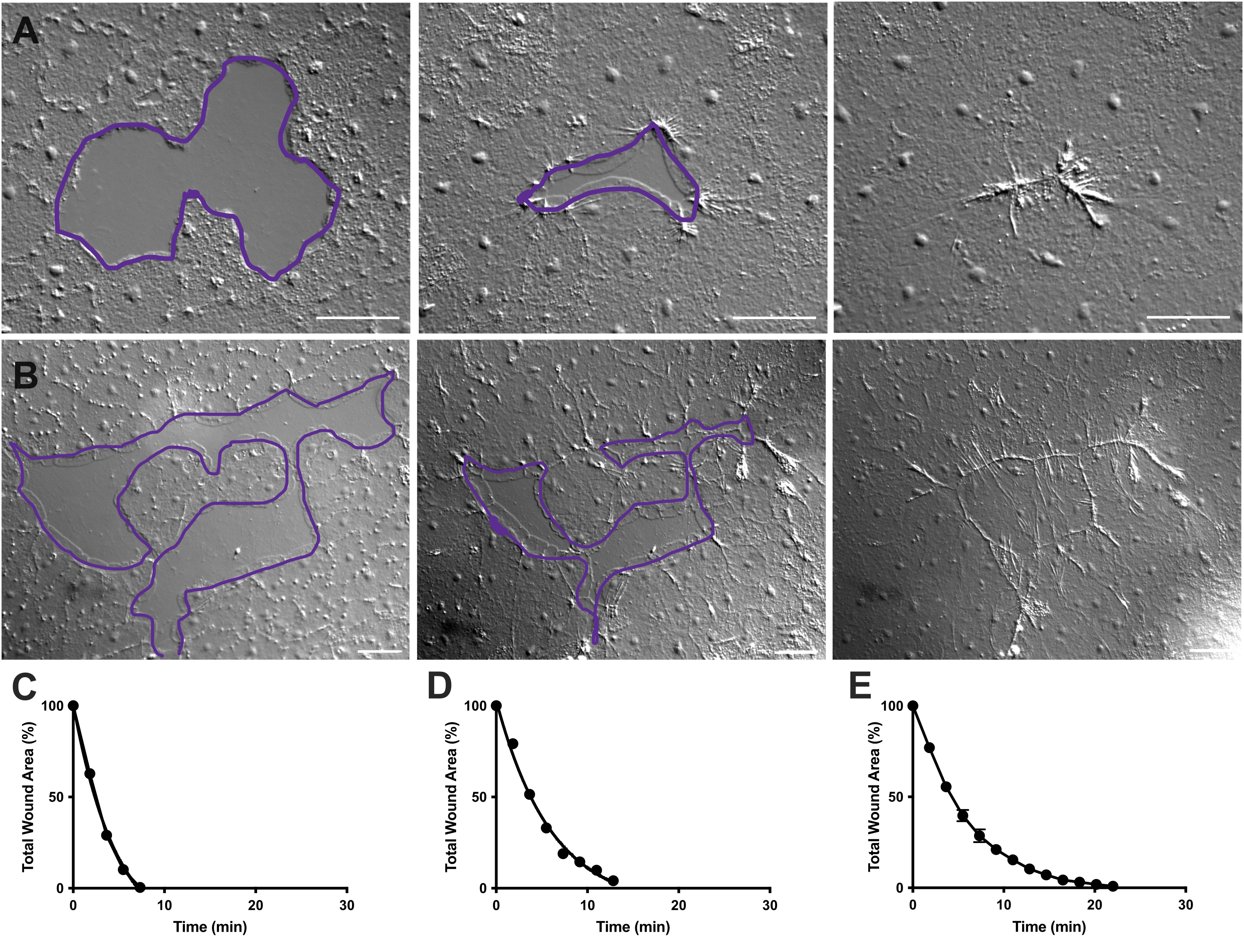
Рисунок 4: Анализ площади раны в небольших эпителиальных ранах. (A ) Пример заживления небольшой эпителиальной раны в течение 10 мин. (B) Пример заживления другой эпителиальной раны в течение 21 мин. Фиолетовые контуры в A,B сопоставимы с измерениями раневых участков с помощью инструмента лассо в FIJI/ImageJ. (C) Нормализованное уменьшение площади раны с течением времени в А. (D) Нормализованное уменьшение площади раны с течением времени в B. (E) Среднее уменьшение площади раны с течением времени для 14 небольших ран. n = 14. Полосы погрешностей, центрированные вокруг среднего ± SEM. Масштабные линейки: 50 мкм Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
5. Мезоглеальные инъекции
- Создание инъекционной чашки
- Получают полидиметилсилоксан (ПДМС), комбинируя основание ПДМС и отвердитель в соотношении 10 частей основания к 1 части отвердителя по весу. Энергично перемешайте, чтобы полностью смешать основу и отвердитель.
- Чтобы удалить пузырьки, поместите смесь в вакуумную камеру на 15 минут. Вылейте смесь в чашку Петри диаметром 60 мм с крышками микроцентрифужных пробирок, чтобы удерживать форму на месте. Немедленно поместите форму на крышки трубок под углом 45° и заклейте ленту на место. Форма представляет собой три сложенных друг на друга смещенных предметных стекла, склеенных вместе для создания выступов в конечной чашке для литья под давлением.
- Поместите всю посуду, форму и смесь в духовку при температуре 60 ° C на 2 часа, чтобы эластомер затвердел. Снимите форму для готовой формы для литья под давлением.
- Вытягивание микропипеток
- Используя съемник микроэлектродов, разработайте программу вытягивания. Используйте одношаговую программу с высокой скоростью. Тепло примерно соответствует результату теста стекла RAMP55,56. Проверьте полученные микропипетки на наличие длинных последовательных конусов.
ПРИМЕЧАНИЕ: Используйте тонкостенные стеклянные боросиликатные капилляры с наружным диаметром 1.0 мм, внутренним диаметром 0.75 мм и длиной 10 см.
- Используя съемник микроэлектродов, разработайте программу вытягивания. Используйте одношаговую программу с высокой скоростью. Тепло примерно соответствует результату теста стекла RAMP55,56. Проверьте полученные микропипетки на наличие длинных последовательных конусов.
- Инъекции красителей и лекарств
- Сделайте иглу для микроинъекций (как указано выше).
- Засыпьте иглу для микроинъекций с помощью длинного наконечника пипетки с избыточным объемом красителя или препарата для инъекции в медузу.
ПРИМЕЧАНИЕ: Для Clytia диметилсульфоксид (ДМСО) следует хранить в разведении <1:100 с ASW, так как более высокие концентрации ДМСО препятствуют заживлению ран. При впрыскивании прозрачного раствора можно добавить раствор Fast Green FCF (разведение 1:100 0,1% Fast Green FCF в ASW) для визуализации вводимой жидкости. - Используя модифицированную пипетку для переноса, как указано выше, поместите медузу субзонтиком вверх в инъекционную чашку PDMS с достаточным количеством ASW, чтобы покрыть животное (рис. 5C). Поставьте блюдо на сцену рассекающего прицела.
ПРИМЕЧАНИЕ: Ограничение избытка ASW не позволяет медузе плавать в тарелке и позволяет делать более успешные инъекции. - Сосредоточьтесь на кончике иглы для микроинъекций и опустите ее в воду рядом с медузой. С помощью микроманипулятора вдавливайте иглу в посуду до тех пор, пока она не согнется и не сломается. Это отверстие наконечника составляет примерно 10-20 мкм.
ПРИМЕЧАНИЕ: Эту иглу можно использовать повторно для одних и тех же инъекций красителя / лекарства в этот день. Рекомендуется использовать свежий наконечник каждый день и для отдельных красителей / лекарств. - С помощью микроманипулятора введите кончик иглы через зонтик в мезоглею, не прокалывая зонт.
ПРИМЕЧАНИЕ: Будет заметна складка/складка эпителия. После того, как игла вставлена в медузу, сгибание/складывание прекращается. - На микроинжекторе установите давление удержания на ноль и давление выброса на ≤20 фунтов на квадратный дюйм. Вводят в один или два квадранта, заполняя каждый пятном красителя или лекарства примерно на 1/4 площади этого квадранта.
ПРИМЕЧАНИЕ: В зависимости от размера медузы в местах однократной инъекции подходят большие или меньшие объемы. Переполнение медузы приводит к сильному повреждению эпителия и даже гибели животного. - В зависимости от того, какой краситель или лекарство вводится, животных помещают в стакан со свежей противолодочной жидкостью, чтобы обеспечить диффузию и инкубацию красителя или лекарства.
- Для визуализации установите медузу на слайд с углублением с помощью модифицированной пипетки, расположив животное так, чтобы зонтик был обращен вверх (рис. 5). Животные могут быть ранены на этом этапе, чтобы проверить действие введенного реагента.

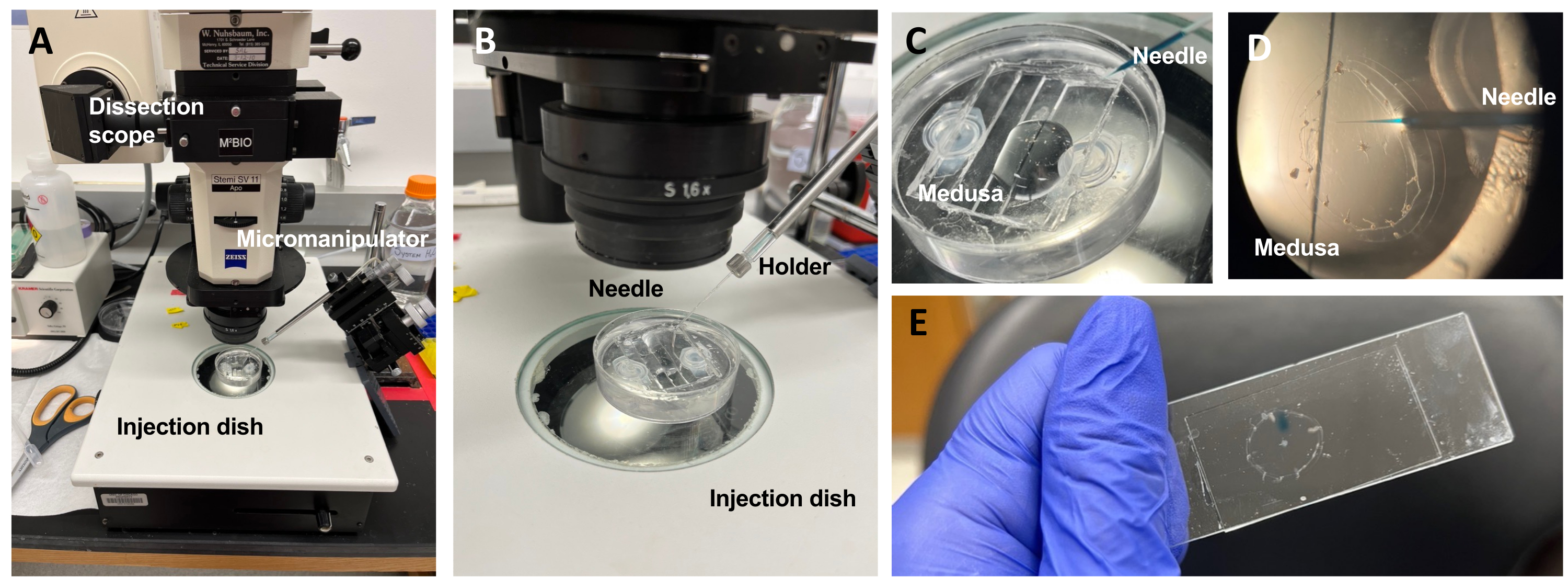
Рисунок 5: Инъекционная установка для введения красителей или лекарств в ECM. (A) Установка впрыска. (B) Крупный план установки инъекции, показывающий ориентацию иглы для микроинъекций (приблизительно 45° под углом относительно животного в чашке). (C) Крупный план силиконовой инъекционной чашки с медузой в небольшом количестве ASW для инъекций. (D) Игла для микроинъекций, загруженная Fast Green FCF, вводится в мезоглею медузы через субзонтик. (E) После инъекции Fast Green FCF в верховую медузу. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
Результаты
В соответствии с приведенными выше протоколами были визуализированы одноклеточные микрораны, небольшие раны и большие раны. Зарегистрированные стопки файлов изображений были сохранены в виде файлов .avi.
В фильме 1 можно увидеть, как микрораны закрываются между клетками и внутри них (рис. 1 и рис. 2). Небольшие ламеллоподии наблюдаются при закрытии с последующим сокращением и заживлением. Мусор исключается и выпускается в воду. Заживление завершается за минуту или меньше.
В фильмах 2 и 3 небольшие раны разной формы заживают за счет образования ламеллиподий, расширения ламеллиподиальных контактов и распространения клеток на краю раны, как описано ранее40 (рис. 1 и рис. 2). Клетки, расположенные ярусами позади маргинальных клеток, не участвуют в заживлении ран такого размера, и коллективной миграции клеток не происходит. Быстрое и прогрессирующее закрытие эпителиальных промежутков сопровождается сокращением тканей вдоль новообразованного раневого шва40. Показана нормализованная скорость заживления этих двух ран, выраженная в процентах от исходной площади с течением времени (рис. 4C, D). Несмотря на некоторую изменчивость динамики закрытия раны, усреднение процентного закрытия площади с течением времени для 14 ран различной формы в диапазоне от 0,02 до 0,125 мм2 позволяет построить среднюю кривую заживления ран у необработанных животных (рис. 4E).
Повреждение базальной мембраны хорошо видно при его возникновении (рис. 2). В фильме 4 клетки на краю небольшой раны, в которой есть повреждение базальной мембраны, распространяются вокруг поврежденного участка, и закрытие щели завершается сокращением кисетной нити.
Если ткань обезвожена или слишком повреждена для восстановления, движение клеток может прекратиться, или весь лист клеток может лопнуть (Фильм 5 и Фильм 6). Обычно это происходит после длительных периодов визуализации (45 минут или дольше). Если разрыв клеток происходит на ранней стадии визуализации, образец отбрасывается.
Как показано в фильме 7, большие раны заживают в несколько этапов. Во-первых, край раны становится гладким и правильным из-за схваток на краю, о чем сообщалось ранее57. Затем видно, что ламеллиподии образуются из клеток на краю раны, при этом ламеллиподии движутся вперед, чтобы максимизировать контакт с соседними ламеллиподиями. Отслеживание ядер в клетках на краю раны и на нескольких ярусах позади маргинальных клеток показывает, что большие промежутки закрываются коллективной миграцией клеток40. Клетки никогда не отделяются, а движутся вместе, как лист.
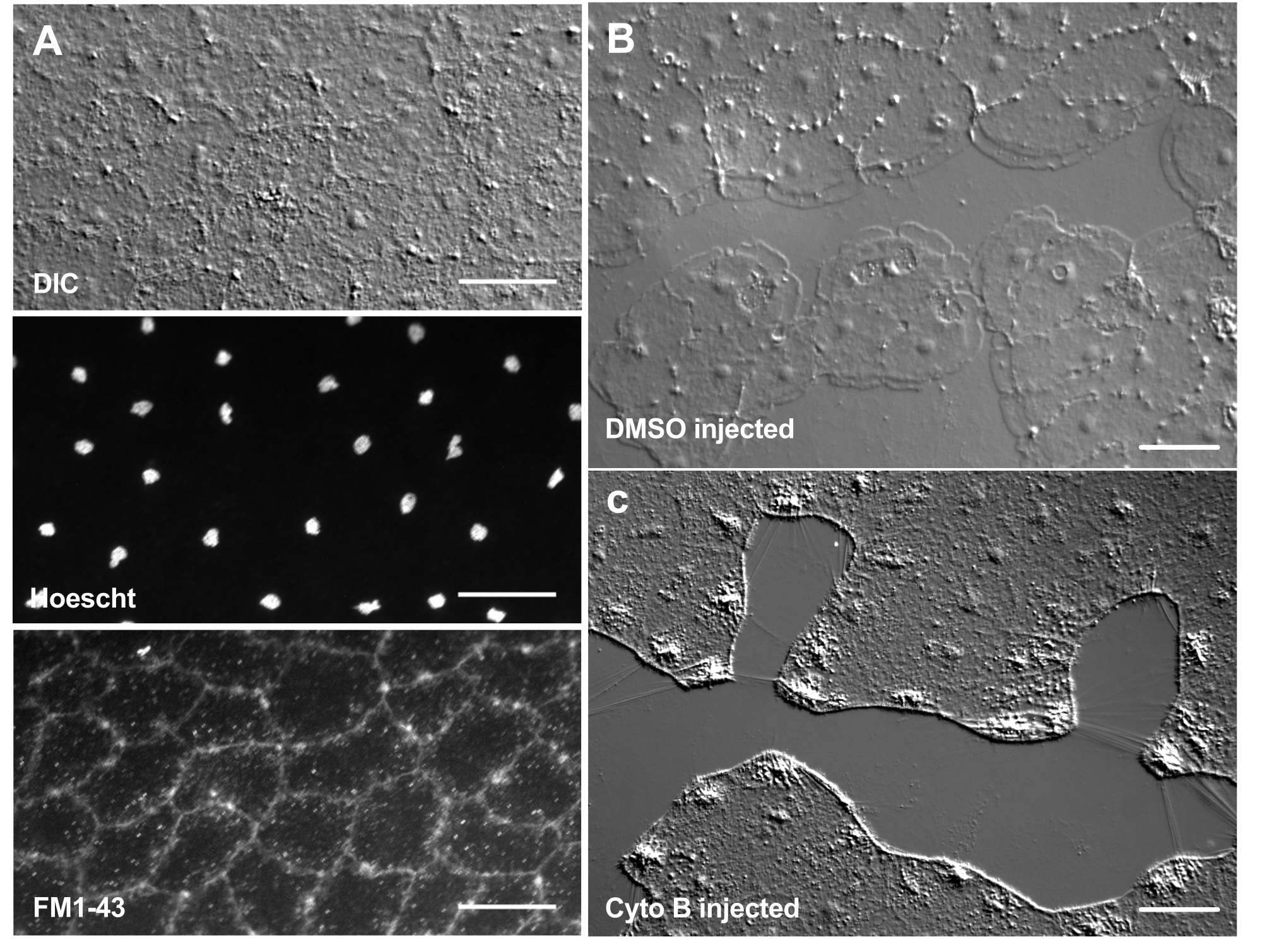
Введение красителей и фармакологических средств может стать мощным инструментом для препарирования биологических механизмов. Многие вещества исключены из Clytia (не показаны), вероятно, из-за слоя слизи, который покрывает поверхность животного. Тем не менее, микроинъекция может быть использована для непосредственного введения молекул в ECM, нарушая структуру ECM или нарушая регуляторную активность в ECM. Кроме того, красители и другие молекулы способны проникать в эпителиальные клетки с базальной стороны. Например, на рисунке 6 показано ядерное окрашивание с помощью Хёхста, окрашивание мембраны с помощью FM1-43 и ингибирование образования ламеллиподий цитохалазином B после микроинъекции этих реагентов в ECM. Введение этих молекул в ECM и эпителиальные клетки перед ранением позволяет проводить эксперименты, которые проверяют влияние фармакологических средств на процесс заживления.

Рисунок 6: Эпителиальные клетки медузы после микроинъекций красителей или фармакологических средств. (A) Эпителиальные клетки, показанные на верхней панели через 5 мин после инъекции с 20 мкМ Hoechst (ядра) и 50 мкМ FM1-43 (мембраны). (В,В) Заживление ран после инъекции с контролем 1:1,000 ДМСО (B) или 100 мкМ цитохалазина B (C). Раны были нанесены через 15 минут после инъекции. Снимки были сделаны через 5 минут после ранения. Образование ламеллиподий ингибируется цитохалазином В. Считается, что кажущиеся «волокна», часто наблюдаемые между клетками в области раны, являются результатом растяжения растяжения базальной мембраны — они не окрашиваются фаллоидином (не показаны). Масштабные линейки: 50 мкм. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
Фильм 1: Покадровая съемка заживления одноклеточных микроран. Прошедшее время: 20 с. Частота кадров: 10 кадров в секунду. Масштабная линейка: 50 мкм. Пожалуйста, нажмите здесь, чтобы скачать этот фильм.
Фильм 2: Покадровая съемка заживления небольшой эпителиальной раны. Хронометраж: 9 мин 54 с. Частота кадров: 10 кадров в секунду. Масштабная линейка: 50 мкм. Пожалуйста, нажмите здесь, чтобы скачать этот фильм.
Фильм 3: Замедленная съемка заживления небольшой эпителиальной раны. Эта рана больше и имеет более неправильную форму, чем рана в фильме 2. Хронометраж: 20 мин 54 с. Частота кадров: 10 кадров в секунду. Масштабная линейка: 50 мкм. Пожалуйста, нажмите здесь, чтобы скачать этот фильм.
Фильм 4: Замедленная съемка небольшой раны и микрораны, заживающей с разрывом базальной мембраны. Ламеллиподии распространяются вокруг разрыва базальной мембраны, хотя они могут распространяться на остальную часть ECM. Как только область раны с повреждением базальной мембраны окружена, сокращение кисетной нити тянет клетки над областью. Хронометраж: 19 мин 4 с. Частота кадров: 10 кадров в секунду. Масштабная линейка: 50 мкм. Пожалуйста, нажмите здесь, чтобы скачать этот фильм.
Фильм 5: Клетки умирают в небольшой эпителиальной ране. Гибель клеток, вероятно, связана с обезвоживанием животного. Хронометраж: 4 мин 24 с. Частота кадров: 10 кадров в секунду. Масштабная линейка: 100 мкм. Пожалуйста, нажмите здесь, чтобы скачать этот фильм.
Фильм 6: Небольшая эпителиальная рана не заживает . Хронометраж: 42 мин 32 с. Частота кадров: 10 кадров в секунду. Масштабная линейка: 50 мкм. Пожалуйста, нажмите здесь, чтобы скачать этот фильм.
Фильм 7: Заживление большой эпителиальной раны. Хронометраж: 25 мин 29 с. Частота кадров: 10 кадров в секунду. Масштабная линейка: 100 мкм. Пожалуйста, нажмите здесь, чтобы скачать этот фильм.
Дополнительный рисунок 1: Схемы размеров резервуара Clytia. 3D визуализация танков Clytia по индивидуальному заказу. (А) Вид спереди и сзади. (B) Вид сбоку. Вырез в изделии, показанном зеленым цветом, покрыт нейлоновой сеткой. Вода поступает в резервуар непосредственно над сеткой, проносится по сетке и создает круговой поток. Вода выходит из системы через отверстие в торцевой части, показанное синим цветом. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Дополнительный фильм 1: Бесклеточный внеклеточный матрикс у Клитии. Z-стек Clytia взят с помощью конфокальной микроскопии. Стек сначала фокусируется на эксзонте, а затем сканирует каждые 10 мкм через ECM до энтодермы и субзонта пластины. Изображения с использованием ДВС-синдрома (слева) и ядерного окрашивания Хёхста (справа) демонстрируют отсутствие клеток в ЭВМ. Масштабная линейка: 100 мкм. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Обсуждение
Здесь представлена методология визуализации ран in vivo у Clytia, относительно нового модельного организма беспозвоночных40,43,58. Есть несколько факторов, которые делают эту систему уникальным и мощным исследовательским инструментом, отличным от других моделей, используемых для изучения заживления ран и реэпителизации. Во-первых, монослойный эпителий прикрепляется к прозрачному ECM, что напоминает анализы культуры тканей in vitro (рис. 1, рис. 2, рис. 3 , рис. 4). Как и в анализах in vitro, клетки могут быть визуализированы с высоким разрешением. Однако, в отличие от культуры тканей, существует подлинная клеточная среда и ECM, так что заживление ран можно рассматривать в контексте сложных сигнальных событий, которые происходят у живого раненого животного. Во-вторых, у Clytia отсутствуют воспалительные реакции, мигрирующие фибробласты, сосудистая сеть и кровь. Это позволяет изучать процесс реэпителизации in vivo при отсутствии перекрывающихся событий, которые происходят у более сложных взрослых животных во время заживления ран59. В-третьих, ECM является бесклеточным (дополнительный фильм 1) и большим, что обеспечивает легкий доступ с помощью иглы для микроинъекций (рис. 5 и рис. 6). Используя этот подход, исследователи могут проверить влияние фармакологических реагентов, которые нарушают структуру ECM или передачу сигналов о заживлении ран in vivo. Реагенты также могут быть введены в эпителиальные клетки, и их влияние на заживление ран in vivo может быть оценено. В-четвертых, существуют протоколы создания мутантов и трансгенных животных в системе Clytia42,43,44,45. Таким образом, заживление ран in vivo может наблюдаться у животных с повышенной/пониженной экспрессией интересующих генов.
В этой технике есть несколько важных шагов. Во-первых, как показано на рисунке 3, необходимо использовать микроскоп, который правильно сконфигурирован для ДВС-микроскопии, поскольку плоские, прозрачные эпителиальные клетки почти невидимы при стандартной световой микроскопии. Также важно выработать навык аккуратно ранить животных, чтобы эпителий был поврежден, не выдолбив ЭКМ. Такое же мягкое прикосновение необходимо для микроинъекций материалов в ECM, так как обширное повреждение животного во время инъекции может поставить под угрозу последующий анализ заживления раны. Несмотря на то, что этим методам нужно учиться, даже начинающие студенты быстро освоили их в лаборатории Малами. Действительно, эти протоколы использовались для демонстрации миграции клеток на лабораторных курсах бакалавриата в Чикагском университете.
Для оптимальной визуализации важно, чтобы животное не двигалось, а выбранная область раны не выходила из поля зрения. Если животные пульсируют, лечение трикаином, как описано, очень эффективно. Для дрейфа часто приходится вручную перемещать образец. Эти движения могут быть удалены из финального фильма с помощью функции регистрации в FIJI/ImageJ.
Ограничением этой системы является то, что невозможно создать идентичные раны, так как раны различаются как по форме, так и по размеру с использованием методов, описанных здесь. Поэтому может быть трудно количественно определить точную скорость закрытия раны или миграции клеток. Позиционные маркеры, такие как углеродные зерна, прилипают к открытому ECM у раненого животного и могут использоваться для измерения скорости коллективной миграции клеток в больших ранах (не показаны). Для анализа закрытия небольших ран, даже с переменным размером и формой раны, существует ограниченный диапазон скоростей закрытия среди ран такого размера (рис. 4). Таким образом, можно количественно обнаружить эффекты стимулирующих или репрессивных фармакологических реагентов.
Хотя в этой работе описывается характеристика заживления ран с использованием только ДВС-микроскопии, те же подходы могут быть использованы для визуализации заживления с использованием флуоресценции или конфокальной микроскопии. Чтобы помочь в этом, существуют протоколы для создания трансгенных животных, у которых различные клеточные и внеклеточные белки флуоресцентно мечены. Одновременная визуализация с ДВС-синдромом и флуоресценцией в сочетании с нарушением заживления ран с использованием фармакологических агентов или мутантных линий станет мощным подходом к пониманию механизмов, лежащих в основе процесса заживления ран в эпителии.
Раскрытие информации
Нечего раскрывать.
Благодарности
E.E.L.L. поддерживается грантом Национального научного фонда PRFB 2011010. Мы хотели бы поблагодарить Цуёси Момосэ и Эвелин Хулистон за помощь в создании наших колоний Клиции, Жана-Батиста Ренье за сбор изображений заживления микроран, Гарри Кириазеса за строительство резервуаров псевдо-крейзеля и Элизабет Бальдо за поддержание среды обитания Клитии. Рисунок 1B был создан с помощью BioRender.com.
Материалы
| Name | Company | Catalog Number | Comments |
| 20500 ACE EKE Microscope Fiber Optic Light Source | Kramer Scientific Corporation | ||
| AxioCam 506 mono | ZEISS | 426557-0000-000-MA285 | |
| Capillary tubes | World Precision Instruments | TW1004 | |
| Cytochalasin B | Abcam | ab143482 | |
| Depression slides | Amscope | BS-C12 | |
| DMR with DIC options and fluorescence halogen lamp | Leica | ||
| Ethyl 3-aminobenzoate methanesulfonate | Sigma Aldrich | E10521-10G | |
| Fast Green FCF | Thermo Scientific | A16520-06 | |
| FM1-43 | Biotium | 70022 | Excitation/Emission: 480/598 nm |
| Hoechst 33342 | Thermo Scientific | 62249 | Excitation/Emission: 361/497 nm |
| imageJ | NIH | ||
| Microloader tips (0.5-10 μL /2-20 μL) | Eppendorf | 930001007 | |
| Micromanipulator | World Precision Instruments | 3301R / M3301L | |
| Microscope Cover Glass (22X40-1.5) | Fisherbrand | 12-544-BP | |
| Petri Dish (60 mm x 15 mm) | Fisherbrand | FB085713A | |
| PicoNozzle v2 | World Precision Instruments | 5430-ALL | |
| Pipette puller | Sutter Instrument Co | P-97 | |
| Pneumatic PicoPump | World Precision Instruments | PV820 | |
| Polycarbonate vacuum, desiccator | Bel-art | F42025-0000 | |
| Prism 9 | GraphPad | ||
| STEMI Sv11 Dissection scope | ZEISS | STEMI SV11 | |
| SYLGARD 184 | Dow Silicones | 1024001 | |
| Transfer pipettes | Fisherbrand | 13-711-7M | |
| Z-Hab mini system | Pentair | ||
| ZEN Microscopy software | Zeiss |
Ссылки
- Tyler, S. Epithelium-the primary building block for metazoan complexity. Integrative and Comparative Biology. 43 (1), 55-63 (2003).
- Kurn, H., Daly, D. T. Histology, Epithelial Cell. StatPearls. , (2022).
- Schempp, C., Emde, M., Wölfle, U. Dermatology in the Darwin anniversary. Part 1: Evolution of the integument. Journal of the German Society of Dermatology. 7 (9), 750-757 (2009).
- Lopez-Ojeda, W., Pandey, A., Alhajj, M., Oakley, A. M. Anatomy, Skin (Integument). StatPearls. , (2022).
- Bukowiecki, A., Hos, D., Cursiefen, C., Eming, S. A. Wound-healing studies in cornea and skin: parallels, differences and opportunities. International Journal of Molecular Sciences. 18 (6), 1257(2017).
- Frantz, C., Stewart, K. M., Weaver, V. M. The extracellular matrix at a glance. Journal of Cell Science. 123 (24), 4195-4200 (2010).
- Hynes, R. O. The evolution of metazoan extracellular matrix. The Journal of Cell Biology. 196 (6), 671-679 (2012).
- Fidler, A. L., et al. Collagen IV and basement membrane at the evolutionary dawn of metazoan tissues. eLife. 6, 24176(2017).
- Bullen, T. F., et al. Characterization of epithelial cell shedding from human small intestine. Laboratory Investigation; a Journal of Technical Methods and Pathology. 86 (10), 1052-1063 (2006).
- Watson, A. J. M., et al. Epithelial barrier function in vivo is sustained despite gaps in epithelial layers. Gastroenterology. 129 (3), 902-912 (2005).
- Sonnemann, K. J., Bement, W. M. Wound repair: toward understanding and integration of single-cell and multicellular wound responses. Annual Review of Cell and Developmental Biology. 27, 237-263 (2011).
- Abreu-Blanco, M. T., Verboon, J. M., Parkhurst, S. M. Single cell wound repair. BioArchitecture. 1 (3), 114-121 (2011).
- Fenteany, G., Janmey, P. A., Stossel, T. P. Signaling pathways and cell mechanics involved in wound closure by epithelial cell sheets. Current Biology. 10 (14), 831-838 (2000).
- Pastar, I., et al. Epithelialization in wound healing: a comprehensive review. Advances in Wound Care. 3 (7), 445-464 (2014).
- Rousselle, P., Braye, F., Dayan, G. Re-epithelialization of adult skin wounds: Cellular mechanisms and therapeutic strategies. Advanced Drug Delivery Reviews. 146, 344-365 (2019).
- Bement, W. M., Forscher, P., Mooseker, M. S. A novel cytoskeletal structure involved in purse string wound closure and cell polarity maintenance. The Journal of Cell Biology. 121 (3), 565-578 (1993).
- Vedula, S. R. K., Ravasio, A., Lim, C. T., Ladoux, B. Collective Cell migration: a mechanistic perspective. Physiology. 28 (6), 370-379 (2013).
- Li, L., He, Y., Zhao, M., Jiang, J. Collective cell migration: Implications for wound healing and cancer invasion. Burns & Trauma. 1 (1), 21-26 (2015).
- Bornes, L., Windoffer, R., Leube, R. E., Morgner, J., van Rheenen, J. Scratch-induced partial skin wounds re-epithelialize by sheets of independently migrating keratinocytes. Life Science Alliance. 4 (1), 202000765(2021).
- Theveneau, E., Mayor, R. Collective cell migration of epithelial and mesenchymal cells. Cellular and Molecular Life Sciences. 70 (19), 3481-3492 (2013).
- Haensel, D., Dai, X. Epithelial-to-mesenchymal transition in cutaneous wound healing: where we are and where we are heading. Developmental Dynamics. 247 (3), 473-480 (2018).
- Cordeiro, J. V., Jacinto, A. The role of transcription-independent damage signals in the initiation of epithelial wound healing. Nature Reviews. Molecular Cell Biology. 14 (4), 249-262 (2013).
- Abreu-Blanco, M. T., Watts, J. J., Verboon, J. M., Parkhurst, S. M. Cytoskeleton responses in wound repair. Cellular and Molecular Life Sciences. 69 (15), 2469-2483 (2012).
- Klarlund, J. K., Block, E. R. Free edges in epithelia as cues for motility. Cell Adhesion & Migration. 5 (2), 106-110 (2011).
- Enyedi, B., Niethammer, P. Mechanisms of epithelial wound detection. Trends in Cell Biology. 25 (7), 398-407 (2015).
- Niethammer, P. The early wound signals. Current Opinion in Genetics & Development. 40, 17-22 (2016).
- Jacinto, A., Martinez-Arias, A., Martin, P. Mechanisms of epithelial fusion and repair. Nature Cell Biology. 3 (5), 117-123 (2001).
- Mayor, R., Etienne-Manneville, S. The front and rear of collective cell migration. Nature reviews. Molecular Cell Biology. 17 (2), 97-109 (2016).
- Gupta, S., Yap, A. S. How adherens junctions move cells during collective migration. Faculty Reviews. 10, 56(2021).
- Blair, M. J., Jones, J. D., Woessner, A. E., Quinn, K. P. Skin structure-function relationships and the wound healing response to intrinsic aging. Advances in Wound Care. 9 (3), 127-143 (2020).
- Falanga, V., et al. Chronic wounds. Nature Reviews. Disease Primers. 8 (1), 50(2022).
- Liang, C. -C., Park, A. Y., Guan, J. -L. In vitro scratch assay: a convenient and inexpensive method for analysis of cell migration in vitro. Nature Protocols. 2 (2), 329-333 (2007).
- Jonkman, J. E. N., et al. An introduction to the wound healing assay using live-cell microscopy. Cell Adhesion & Migration. 8 (5), 440-451 (2014).
- Poujade, M., et al. Collective migration of an epithelial monolayer in response to a model wound. Proceedings of the National Academy of Sciences. 104 (41), 15988-15993 (2007).
- Masson-Meyers, D. S., et al. Experimental models and methods for cutaneous wound healing assessment. International Journal of Experimental Pathology. 101 (1-2), 21-37 (2020).
- Park, S., et al. Tissue-scale coordination of cellular behaviour promotes epidermal wound repair in live mice. Nature Cell Biology. 19 (2), 155-163 (2017).
- Tsai, C. -R., Wang, Y., Galko, M. J. Crawling wounded: molecular genetic insights into wound healing from Drosophila larvae. The International Journal of Developmental Biology. 62 (6-7-8), 479-489 (2018).
- Richardson, R., et al. Adult zebrafish as a model system for cutaneous wound-healing research. The Journal of Investigative Dermatology. 133 (6), 1655-1665 (2013).
- Erickson, J. R., Echeverri, K. Learning from regeneration research organisms: The circuitous road to scar free wound healing. Developmental Biology. 433 (2), 144-154 (2018).
- Kamran, Z., et al. In vivo imaging of epithelial wound healing in the cnidarian Clytia hemisphaerica demonstrates early evolution of purse string and cell crawling closure mechanisms. BMC Developmental Biology. 17 (1), 17(2017).
- Home. MARIMBA. , Available from: http://marimba.obs-vlfr.fr/home (2023).
- Chari, T., et al. Whole-animal multiplexed single-cell RNA-seq reveals transcriptional shifts across Clytia medusa cell types. Science Advances. 7 (48), (2021).
- Weissbourd, B., et al. A genetically tractable jellyfish model for systems and evolutionary neuroscience. Cell. 184 (24), 5854-5868 (2021).
- Momose, T., et al. High doses of CRISPR/Cas9 ribonucleoprotein efficiently induce gene knockout with low mosaicism in the hydrozoan Clytia hemisphaerica through microhomology-mediated deletion. Scientific Reports. 8 (1), 11734(2018).
- Houliston, E., Leclère, L., Munro, C., Copley, R. R., Momose, T. Past, present and future of Clytia hemisphaerica as a laboratory jellyfish. Current Topics in Developmental Biology. 147, 121-151 (2022).
- Schmid, V., et al. The extracellular matrix (mesoglea) of hydrozoan jellyfish and its ability to support cell adhesion and spreading. Hydrobiologia. 216 (1), 3-10 (1991).
- Day, R. M., Lenhoff, H. M. Hydra mesoglea: a model for investigating epithelial cell-basement membrane interactions. Science. 211 (4479), 291-294 (1981).
- Zhang, X., et al. The collagens of hydra provide insight into the evolution of metazoan extracellular matrices. The Journal of Biological Chemistry. 282 (9), 6792-6802 (2007).
- Danjo, Y., Gipson, I. K. Actin 'purse string' filaments are anchored by E-cadherin-mediated adherens junctions at the leading edge of the epithelial wound, providing coordinated cell movement. Journal of Cell Science. 111 (22), 3323-3332 (1998).
- Arenas Gómez, C. M., Sabin, K. Z., Echeverri, K. Wound healing across the animal kingdom: Crosstalk between the immune system and the extracellular matrix. Developmental Dynamics. 249 (7), 834-846 (2020).
- Lechable, M., et al. An improved whole life cycle culture protocol for the hydrozoan genetic model Clytia hemisphaerica. Biology Open. 9 (11), (2020).
- Casares, L., et al. Hydraulic fracture during epithelial stretching. Nature Materials. 14 (3), 343-351 (2015).
- Wayne, R. Chapter 4 - Bright-Field Microscopy. Light and Video Microscopy (Third Edition). , 95-116 (2019).
- Murphy, D. B., Davidson, M. W. Fundamentals of Light Microscopy and Electronic Imaging: Second Edition. , John Wiley and Sons. (2012).
- Micropipette Techniques for Electrophysiology. , Available from: https://www.sutter.com/micropipette/cookbook.html (2022).
- Brown, A. L., Johnson, B. E., Goodman, M. B. Making patch-pipettes and sharp electrodes with a programmable puller). Journal of Visualized Experiments. (20), e939(2008).
- Klarlund, J. K. Dual modes of motility at the leading edge of migrating epithelial cell sheets. Proceedings of the National Academy of Sciences. 109 (39), 15799-15804 (2012).
- Houliston, E., Momose, T., Manuel, M. Clytia hemisphaerica: a jellyfish cousin joins the laboratory. Trends in Genetics. 26 (4), 159-167 (2010).
- Rodrigues, M., Kosaric, N., Bonham, C. A., Gurtner, G. C. Wound healing: a cellular perspective. Physiological Reviews. 99 (1), 665-706 (2019).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены