
Method Article
siRNA אלקטרופורציה כדי לווסת את החלונות האוטומטיים בווירוס הרפס סוג 1-מונוציט הנגזר מונולוייט תאים דנדריטים
In This Article
Summary
במחקר זה, אנו מציגים מעכב-ו siRNA אסטרטגיות מבוססות להתערב השטף האוטומטי הזרע בנגיף הרפס סוג 1 (HSV-1)-הנגועים מונוציט הנגזרים התאים הדנדריטים.
Abstract
וירוס הרפס סוג-1 (HSV-1) מעורר בצורה אוטומטית בשניהם, תאים דנדריטים לא בוגרים (iDCs), כמו גם תאים דנדריטים בוגרת (mDCs), ואילו שטף הזרע האוטומטי נצפה רק ב-iDCs. כדי לזכות בתובנות מכניסטיות, פיתחנו אסטרטגיות יעילות להתערב במחזור האוטומטי של HSV-1 המושרה. אסטרטגיה מבוססת מעכב, כדי לווסת את האוטומטי HSV-1 המושרה, מהווה את הבחירה הראשונה, שכן היא שיטה קלה ומהירה. כדי לעקוף את ההשפעות הפוטנציאליות מחוץ ליעד של תרכובות כאלה, פיתחנו מבוסס siRNA האסטרטגיה החלופית, כדי לווסת מחזור האנדרורגיות אוטומטי ב-iDCs על זיהום HSV-1. אכן, אלקטרופורציה של idcs עם FIP200-ספציפי sirna לפני הזיהום HSV-1 היא שיטה ספציפית מאוד ומוצלחת ביטוי FIP200 חלבון ablate אוחר ובכך לעכב שטף הידרורגיות אוטומטי. שתי שיטות מוצגות כתוצאה עיכוב יעיל של מחזור HSV-1 המושרה אוטומטי ב-iDCs, לפיה הטכניקה מבוססת siRNA הוא יותר ספציפי היעד. גישה נוספת מבוססת siRNA פותחה כדי להשתיק באופן סלקטיבי את ביטוי החלבון של KIF1B ו KIF2A, הקלה על תחלופת מחזור אוטומטי על זיהום HSV-1 ב mDCs. לסיכום, הטכניקה של sirna אלקטרופורציה מייצגת אסטרטגיה מבטיחה, כדי באופן סלקטיבי ביטוי של חלבונים ברורים לנתח את השפעתם על זיהום HSV-1.
Introduction
הדור של התאים הדנדריטים הנגזרים האנושי (DCs) מהווה מודל מתאים בתחום החיצוני כדי ללמוד את הפונקציות ואת הביולוגיה של סוג זה תא החיסון החשוב. בידוד, כמו גם הבדלה של מונוציטים לתוך DCs הוקמה היטב בשנים האחרונות1,2. הזיהום של DCs עם הרפס α-her, וירוס מסוג וירוס-1 (HSV-1) משמש מערכת מודל ללמוד HSV-1-תיווך ומאפדי ביולוגיה DC2,3,4,5,6 . זה חשוב במיוחד כדי להבהיר כיצד היפסח לחלח או לעכב את התגובות החיסונית אנטי וירוס חזק, כדי ליצור השהיה של נישות החיסון המורשה בתוך המארח7,8. במובן זה, הרבה וירוסים הם פתוגנים מוצלחים מאוד, כי הם רחבים מתפשטים ברחבי האוכלוסייה להגיע sero-שכיחות של עד 90% על פי האזור הגיאוגרפי9. כדי להבין ואולי למנוע את זה, תובנות נוספות לתוך המודולוניות HSV-1 בתיווך של המערכת החיסונית של המחשב המארח, ובמיוחד של תאים חיסוניים כגון DCs, נדרשים.
התבוננות חדשה לחלוטין לגבי הגומלין של DCs עם HSV-1 פורסמה לאחרונה על ידי Turan ואח '10. המחברים הפגינו כי ההישג של שכפול HSV-1 תלוי לחלוטין במצב ההבשלה של DCs. ב-iDCs, השכפול השלם של HSV-1 הוא הקלה על ידי מנגנונים התלויים האוטומטית. בעוד HSV1 מעורר האוטומטית בשני, iDCs ו mDCs, שטף ההכולינרגיות האוטומטי נצפה רק ב-iDCs. זה בתורו מקלה על היציאה הגרעינית של הקפידים ויראלי באמצעות השפלה אוטומטית של הלבינים גרעינית ב-iDCs. כדי לקבל תובנות מכניסטיות לתוך מסלול השפלה זה HSV-1 המושרה ב-iDCs לעומת mDCs, אסטרטגיות חדשות ויעילות הם קריטיים כדי לחקור שטף מחדש בצורה אוטומטית.
מקרואוטומטיים האגי (תצוגה מקדימה) הוא תהליך רב שימור היטב מיקוד של חלבונים תאיים או אורגלים שלמים לעיכול ליזוזומא11. במידה מרבית, ניתן לחלק את החלקים האוטומטיים לתוך הייזום (i), (ii) התגרדת ממברנה, (iii) הרחבת שלפוחית, ו (iv) בתצוגה מקדימה של חלק היתוך שלב12. במהלך הייזום (i), רכיבים כגון מתחם ULK1/2 קינאז המופעל, המכיל את החלבון המפעיל של משפחה הדבקה קינאז של 200 kD (FIP200), הם קריטיים להפעלת מורכבות beclin-1-Vps34-AMBRA1. לאחר מכן, הנוקלאוציה ממברנה (ii) יוזם היווצרות phagophore13, אשר בולע את cartoplasmic כאשר מסומנים על ידי מולקולות כגון p6214. במהלך התרחבות שלפוחית לפוחית והתבגרות באופן אוטומטי (iii) שרשרת אור חלבון משויך 3 (LC3)-אני מומר לתוך הטופס lipidated LC3-II כי הוא מוכנס לתוך הקרום האוטומטי הפזומלית. לפיכך, שיעורי ההמרה של LC3-I-II הם מחוון של השראה אוטומטית על-ידי שיקוף היווצרות של הגוזומים האוטומטיים בוגרים15,16. על פי הוראות היתוך (iv), לא רק את המטען האוטומטי, אלא גם הקשורים p62 ו-LC3-II חלבונים עוברים השפלה (למשל, על ידי הידרוליזה). כך, אובדן של p62 ו-LC3-II לשמש סמנים עבור שטף הזרם האוטומטי17. המיזוג של הגוזומים אוטומטיים עם lysosomes, ולכן בעקבות מחזור הנקרא אוטומטית, הוא תלוי מאוד על לוקליזציה ליסוזומתאיים. זה, בין היתר, מוסדר על ידי בני משפחה KIF1B ו KIF2A, אשר הוכחו לרעה להשפיע על האוטומטי מסוימים היתוך כמה-lysosome מיזוג18. מעניין, ביטוי החלבון של KIF1B ו KIF2A הוא המושרה על התבגרות DC והוא אחראי על ובכך את השטף האוטומטי יעיל ה ב HSV-1-הנגועים mDCs, אשר הסלים השלם HSV-1 שכפול10.
ניסיונות ניסיוניים לווסת את התצוגה האוטומטית האוטומטי כוללים את השימוש של תרכובות ידוע לגרום או לעכב את השביל המסוים הזה19,20,21. במחקר זה, אנו מתארים שתי אסטרטגיות מעכבי מבוססי לחסום מחזור הכולינרגיות אוטומטי ב HSV-1-הנגועים iDCs. התרכובת הראשונה המשמשת בניסויים שלנו הוא מעכב האוטומטי והחזק מעכבי-1 (spautin-1), אשר תוארה לקדם beclin-1-Vps34-AMBRA1 מורכבים השפלה במהלך שלב החניכה של התצוגה האוטומטית22. התרכובת השנייה המשמשת במחקר הנוכחי הוא bafilomycin-A1 (BA1), מעכב V-atpase החוסם את האירועים האוטומטיים המאוחרת (כלומר, היתוך האוטומטי-lysosome פיוז'ן, כמו גם את החמצה)23,24. השימוש של אחת משתי מעכבי אלה לפני הזיהום במרכז הבינתחומי עם HSV-1 מעכב את הבחירה האוטומטית, אך אינו מפריע יעיל ביטוי גנים ויראלי. Thus, זו אסטרטגיה מבוססת מעכב לפני הזיהום HSV-1 מציע כלי רב עוצמה כדי לעכב את HSV-1 המושרה שטף ההכולינרגיות זה יכול בקלות להיות מורחבת עבור שפע של סוגים שונים של תאים ווירוסים, אשר גם פוטנציאלי לגרום אוטומטית.
כדי להתגבר על החיסרון העיקרי של גישה מבוססת מעכב (כלומר, תופעות לא ספציפיות מחוץ ליעד), פיתחנו שיטה מבוססת sirna לחסום שטף הכולינרגיות האוטומטי ב (HSV-1-נגוע) idcs. הטכניקה של sirna אלקטרופורציה מייצגת אסטרטגיה חלופית רבת עוצמה, באמצעות אבלציה סלקטיבית של הביטוי של חלבונים ברורים (כלומר, רכיבים אוטומטיים). בניסויים שלנו idcs היו electroporated עם FIP200 ספציפי sirna באמצעות מנגנון אלקטרופורציה I (לראות את לוח החומרים) ופרוטוקול שונה שתוארה על ידי gerer et al. (2017) ו prechtel ואח ' (2007), כדי לעכב את התצוגה האוטומטית במהלך ה שלבהחניכה 25,26. טכניקה זו אפשרה לנו במיוחד להוריד את הביטוי FIP200 ב-iDCs, מבלי להפריע לכדאיות התאים ואת הפנוטיפים בלתי בוגרים שלהם יומיים לאחר ההודעה. לציין, HSV-1 הזיהום הוקמה ב-Idporated אלה שיקוף על ידי יעיל חלבון נגיפי הביטוי. טכניקה זו מבוססת siRNA מציעה יתרון ייחודי (כלומר, כי מגוון של רכיבים שונים הניתנים לבחירה אוטומטית, גם בשילוב), ניתן לפלח במיוחד עבור אבלציה של הביטוי שלהם.
במחקר זה, אנו עוד לתאר שיטה מבוססת siRNA כדי לגרום השטף ההכולינרגיות גם ב HSV-1-הנגועים mDCs. במקרה זה, iDCs היו electroporated עם siRNA ממוקד נגד KIF1B ו KIF2A לפני התבגרות DC באמצעות מנגנון electroporated השני (ראה טבלת חומרים). מכיוון ששני החלבונים הם upregulated במהלך התבגרות DC ידוע שלילי להסדיר פיוז'ן של הגוזומים אוטומטיים עם lysosomes10,18, הנוק שלהם המושרה בחוזקה השטף השני באופן אוטומטי ב mdcs על זיהום HSV-1. כך, הטכניקה מבוססת siRNA אפשרה לנו במיוחד לזרז מחזור המולא אוטומטי באמצעות הפרעה ביטוי החלבון קיף ב mDCs, ובכך יכול ובכך לחקות את רמות הביטוי שלהם ב-iDCs.
לסיכום, אנו מציגים שתי שיטות נפרדות כדי לעכב שטף הכולינרגיות אוטומטי ב HSV-1-מושפע iDCs. בעוד הגישה המבוססת על מעכב הראשון מהווה קל, דרך זולה ומהירה להפריע השפלה האוטומטית, טכניקה מבוססת siRNA השני הוא ספציפי יותר, שיטה מתאימה מאוד לתמוך ולאמת את התוצאות של מעכב מבוססי ניסויים. בנוסף, אנו מתארים שיטה לגרום השטף הדו האוטומטי גם ב HSV-1-הנגועים mDCs, באמצעות הסתרה siRNA-מתווכת של שני חלבונים קיף.
Protocol
מונוציט-בקרי התחום הנגזרים נוצרו ממוצרי ליוקפיזיס של תורמים בריאים. בשל כך, הושגה הצבעה חיובית מוועדת האתיקה המקומית (התייחסות מספר 4556). הניסויים של המחקר הנוכחי נערכו בהתאם להמלצות של ועדת האתיקה של "פרידריך-אלכסנדר-אוניברסיטת ארלנגן-נירנברג" (מספר 4556). כל התורמים אישרו הסכמה מושכלת בכתב, כולל בהתאם להכרזת הלסינקי.
1. הדור והטיפול של תאים דנדריטים בלתי בוגרים (iDCs) ותאים דנדריטים מבוגרים (mDCs)
- בידוד תאים אנושיים היקפיים לתאי מונפקציה (PBMCs) מתאי מערכת של לאוקורחנך (LRSCs) כפי שתוארו בעבר27. הימנע הקפאה של PBMCs ולהשתמש בהם ישירות על בידוד כדי להשיג תשואה DC גבוה יותר.
- ליצור DCs אנושי מ pbmcs של תורמים בריאים שונים ב T175 התרבות תאים מבחנות כמו בעבר תיאר10,27. בקצרה, השתמש 350-400 מיליוני PBMCs ב 30 מ ל של DC בינונית (RPMI 1640 ללא L-גלוטמין, 1% (v/v) AB-סרום, 100 U/mL, 100 mg/mL סטרפטומיצין, 0.4 mM L-גלוטמין, 10 מ"מ HEPES) לכל תרבות התא בקבוקון לבידוד של מונוציטים על ידי דבקות. אחרי 1 h, לשטוף את השבר הלא מחסיד באמצעות RPMI 1640. הוסף מדיום טרי DC שיושלם עם 800 U/mL GM-שדרתי ו 250 U/mL IL-4, ו דגירה עבור 3 ימים.
- ביום 3 פוסט הדבקות, להוסיף 5 מ ל של מדיום DC טרי המכיל GM-שדרתי ו-IL-4 עם ריכוז סופי של 400 U/mL ו 250 U/mL לכל התרבות התא בקבוקון, בהתאמה, עבור DC בידול.
- כדי לקצור את iDCs, לשטוף בעדינות מחסיד באופן רופף iDCs מהחלק התחתון של הבקבוקון תרבות התא, ביום 4 פוסט הדבקות. חזור על שלב זה 2 פעמים. עבור דור של mDCs, להוסיף קוקטייל ההבשלה מורכב כדלקמן: GM-שדרתי (ריכוז סופי: 40 U/mL), IL-4 (ריכוז סופי: 250 U/mL), IL-6 (ריכוז סופי: 1000 U/mL), IL-1β (הריכוז הסופי: 200 U/mL), TNF-α (הריכוז הסופי mL), פרוגלנדין E2 (PGE2; ריכוז סופי: 1 μg/mL).
- שישה ימים לאחר הדבקות (יומיים הודעה השראה של התבגרות באמצעות קוקטייל ציטוקינים), לשטוף mdcs מהחלק התחתון של הבקבוקון תרבות התא. חזור על שלב זה פעמיים.
הערה: בקרי תחום בוגרים ומבוגרים ניתן ליצור באופן רציף מתורמים זהים ב 1 בקבוקון תרבות התא. כדי לעשות זאת, (אני) להפריד את המספר המתאים של idcs ו (ii) לגרום התבגרות של התאים הנותרים מבחנות באמצעות הקוקטייל ציטוקינים המפורטים בשלב 1.2.2.
- העברת iDCs או mDCs ב התרבות המתאימה ביותר בגוף התאים לתוך 50 mL צינורות. לקצור את התאים דרך צנטריפוגה ב 300 x g עבור 5 דקות.
- בעדינות להשעות מחדש (= לשטוף) DCs ב 5-10 mL של RPMI 1640 לכל התרבות התא בקבוקון. שילוב השתלים מתחום DC בצינור אחד.
- הגדר את מספר התא באמצעות תא ספירה או שיטה חלופית. הימנע שינויים בטמפרטורה בעת טיפול ב-iDCs, כדי להפחית את הסיכון לשינויים פנוטיאריים.
2. ניתוח ציטומטלי זרימה כדי לפקח על מצב ההבשלה פנוטימית
- העברת iDCs או mDCs (0.5 x 106) משלב 1.3.1 לתוך צינור 1.5 mL. לקצור את התאים באמצעות צנטריפוגה ב 3390 x g עבור 1.5 min. לשטוף את התאים פעם אחת עם מאגר FACS (PBS בתוספת עם 2% סרום עגל עוברי (fcs)).
- השהה מחדש את התאים ב-100 μL של פתרון מכתים נוגדנים (מאגר FACS) המכיל נוגדנים מבוססי fluorochrome המסומנים נגד מולקולות של משטח מוגדר.
- השתמש בנוגדנים הבאים כדי לאמת את הטוהר (CD3-FITC, CD14-PE) כמו גם התבגרות סטטוס של DCs (CD80-PacBlue/-PE-Cy5, CD11c-PE-Cy5, CCR7-PE-Cy7, CD83-APC, CD86-pe, MHCII-APC-Cy7).
- הכינו דגימה אחת בלתי מוכתמת ב-100 μL של מאגר FACS כפקד.
- כתם את התאים על קרח בחשיכה במשך 30 דקות.
- לאחר מכן לשטוף את התאים פעמיים 1 מ ל של מאגר FACS ו צנטריפוגה ב 3390 x g עבור 1.5 דקות.
- לבסוף, להשעות מחדש את התאים ב-200 μL של מאגר FACS בתוספת 2% בתחתית ולנתח את התאים על ידי הזרמת cy, try. ניתן לאחסן תאים קבועים ב -4 ° C בחשכה עד יומיים.
3. זיהום הליך של DCs עם וירוס הרפס סוג-1 (HSV-1) והפרעה של שטף הנגרמת על ידי הנגיף HSV-1 המושרה באמצעות spautin-1 או bafilomycin-A1
הערה: מאמץ HSV-1/17 +/CMVT-EGFP/UL43 (HSV-1 EGFP) המשמש במחקר זה הושג מאמץ מעבדה HSV-1 זן 17 +. מאמץ HSV-1 EGFP מבטא את חלבון פלורסנט ירוק משופר (EGFP) אשר הוכנס לתוך לוקוס גנים UL43 תחת שליטה של מקדם CMV. EGFP משמש סמן עבור זיהום HSV-1. יתר על כן, את המתח HSV1-RFPVP26 שימש למחקרים זיהום DC (שתוארו קודם לכן Turan ואח ', 2019). וירוס זה מבטא את הקרקע capsid חלבון VP26 התמזגו לחלבון פלורסנט מונומר אדום (mRFP).
- העברת iDCs או mDCs (2 x 106) משלב 1.3 לתוך צינור 2 מ"ל. לאחר מכן, צנטריפוגה את התאים ב 3390 x g עבור 1.5 דקות ולהיפטר supernatant.
- השהה מחדש בעדינות את התאים באמצעי זיהום (RPMI 1640 שיושלם עם 20 מ"מ HEPES).
- כדי לעכב את מסלול השפלה האוטומטי-lysoזומלי, טרום לטפל DCs עם spautin-1 או bafilomycin-A1 1 h לפני זיהום. הוסף גם 10 μM spautin-1 או 1 μM BA1, או DMSO כפקד לא מטופל, למדיום זיהום. מודטה את התאים בבלוק חימום ב 300 rpm רועד ב 37 ° צ' עבור 1 h.
- עבור לימודי זיהום, התחסן את התאים עם הריסונים HSV-1 בריבוי של זיהום (מע) של 2. להוסיף את הנפח המתאים של MNT מאגר (30 מ"מ 2-(N-שורגוולסטינו) חומצה ethanesulfonic (MES), 100 מ"מ הנאל, 20 מ"מ טריס) כפקד מדומה. מודטה את התאים בבלוק חימום ב 300 rpm רועד ב 37 ° צ' עבור 1 h.
- 1 h לאחר זיהום (hpi), לאסוף את התאים בשעה 3390 x g עבור 1.5 דקות. מפחית את התאים בעדינות להשעות את התא בינונית DC המכיל 40 U/ML של GM-שדרתי, 250 U/ML של IL-4 ו 10 μm spautin-1, 1 ΜM BA1, או dmso כפקד. זרעים שטופלו מבוים ו HSV-1 תאים נגועים בריכוז הסופי של 1 x 106/mL לתוך צלחת 6-טוב.
- ב 16-24 hpi, לקצור את התאים על ידי שטיפה (mDCs) או באמצעות מגרד תא (iDCs). העבר את התאים 1.5 לתוך safelock mL.
- לאסוף את התאים דרך צנטריפוגה ב 3390 x g עבור 1.5 דקות ולשטוף את הגלולה פעם אחת על ידי הוספת 1 מ ל של PBS.
- השעיית מחדש במרץ את התאים בתמהיל הליזה המכיל 29 μL של 2x Roti-Load, 1 μL 100 mM MgCl2 ו 12.5 U/mL בנזיל.
- לפירוק תאים והעיכול DNA באמצעות בנזיל, מודתיאת הדגימות ב 37 ° c עבור 10 דקות. לאחר מכן, הטמפרטורה של החלבונים ב 95 ° c עבור 10 דקות.
- בצע ניתוחים של SDS-PAGE וכתמי אבן מערביים כדי לוודא את רמות החלבונים של LC3BI/II, p62, ICP0/5 ו-, ו-.
4. הפרעות של שטף הנגרמת על ידי הזרם HSV-1 המושרה באמצעות אלקטרופורציה של iDCs באמצעות FIP200-siRNA
הערה: הפרוטוקול הנוכחי של ה-sirna אלקטרופורציה השתנה מ-prechtel ואח ' (2007) ו-gerer et al. (2017).
- העברת iDCs (12 x 106) ביום 3.5 לאחר הדבקות לתוך שפופרת 50 mL. לאחר מכן, צנטריפוגה את התאים ב 300 x g עבור 5 דקות ולהיפטר supernatant. במקביל, לבצע ניתוח ציטומטלי זרימה לפקח על מצב ההבשלה כמתואר בשלב 2 (השימוש בחיים/סגול מת במקום CD80-PacBlue).
- בעדינות לשטוף iDCs ב 5 מ ל של OptiMEM ללא פנול אדום צנטריפוגה את התאים ב 300 x g עבור 5 דקות. להיפטר supernatant ולהשהות בעדינות את idcs ב 200 μl של OptiMEM ללא פנול אדום, התאמת ריכוז התא של 6 x 106/100 μl. אין למקם את התאים על הקרח ולהימנע שינויים בטמפרטורה. לנוע במהירות ולהימנע תקופות דגירה ארוכה של iDCs ב OptiMEM ללא פנול אדום.
- העבר או 75 pmol FIP200-ספציפי siRNA או 75 pmol של siRNA מעורבל, כפקד, לתוך 4 מילימטר אלקטרו כימיקלים ולהוסיף 100 μL (6x106 תאים) של ההשעיה התא. הדופק הישיר iDCs באמצעות מנגנון אלקטרופורציה I, החלת ההגדרות הבאות: 500 V עבור 1 ms.
- לפני ההליך הניסיוני, להכין השתלים sirna על פי ההוראה של היצרן, סדרת מחלקים ולאחסן אותם ב-20 ° c. להפשיר ולשמור אותם על הקרח בעת שימוש אלה siRNAs עבור אלקטרופורציה. , לפני שהוא מחשמל את הדגימות. בצע פעימה במבחן
- לאחר electroporation, ישירות להעביר את המרכז לתוך 6-היטב צלחות עם מדיום חדש מחומם מראש DC (שיושלם עם 40 U/mL של GM-שדרתי ו 250 U/mL של IL-4). הזרע את התאים בריכוז הסופי של 1 x 106/ml ולמקם אותם לתוך אינקובטור. אין לשטוף את התאים מתוך קובט אלקטרו.
- לאחר 48 h, בדוק תחילה את המבנה של המיקרו-מחשב האלקטרו-ממדי. לאחר מכן, לקצור את התאים באמצעות מסיק תא ולהעבירם 15 מ ל צינורות. לשטוף את הבארות עם 1 מ ל של PBS בתוספת 0.01% EDTA ולהעביר את הפתרון בצינורות המתאימים.
- לאחר מכן, פצל 6 x 106 מצבי idcs לכל sirna כמתואר בשלבים הבאים.
- השתמש 0.5 x 106 תאים כדי להעריך את מצב ההבשלה ואת הכדאיות התא כפי שמתואר בשלב 2. השתמש בנוגדנים הבאים: CD11c-PE-Cy5, CCR7-PE-Cy7, CD83-APC, MHCII-APC-Cy7 וחיים/סגול מת.
- השתמש 1 x 106 תאים עבור כתמי אבן מערבית ניתוחים כדי לאמת את היעילות FIP200 להוריד ספציפי. להעביר ולקצור את התאים לתוך 1.5 מ"ל safelock mL על ידי צנטריפוגה ב 3390 x g עבור 1.5 דקות. להכין את lysates תא כפי שמתואר בשלב 3.4 ולבצע ניתוחים מערביים כתמי.
- השתמש בתאים הנותרים (4.5 x 106) עבור ניסויים לזיהום HSV-1. עבור כל תנאי ניסיוני, להעביר 2.25 x 106 idcs לתוך 2 צינורות mL, או להדביק אותם עם HSV1 ב-משרד המידע של 2 או להוסיף מאגר mnt כפקד מבוים. בצע את הזיהום כפי שמתואר בשלב 3.3.
- ב 20 hpi, לקצור את התאים באמצעות מגרד התא ולהכין lysates תא לניתוח כתמי אבן המערבי כמתואר בשלב 3.4.
5. אפנון האוטומטי מסלול ליסוזומל ב HSV-1-הנגועים mDCs באמצעות KIF1B/2A-siRNA אלקטרופורציה
- העברת iDCs (24 x 106) ביום 4 הדבקות לתוך צינור 50 mL. לאחר מכן, צנטריפוגה את התאים ב 300 x g עבור 5 דקות ולהיפטר supernatant.
- השעיה מחדש בעדינות (= לשטוף) iDCs ב 8 מ ל של PBS, כדי להתאים ריכוז התא הסופי של 3 x 106/Ml. העברה 3 x 106 תאים לתוך 1.5 מ"ל צינורות והקציר התאים ב 3390 x g עבור 1.5 דקות.
- להשעות את idcs ב 100 μl של מאגר P3 המכיל את התוספת לערבב (על פי ההוראות של היצרן; מנגנון ערכת אלקטרופורציה II) ו (i) 75 pmol של KIF1B-ספציפי sirna, (II) 75 pmol של KIF2A-ספציפי sirna, או (iii) שניהם. השתמש (iv) את הסכום המתאים של siRNA מעורבל כפקד. הכינו שתי שפופרות עבור כל תנאי siRNA (6 x 106 תאים) ולהעביר את השעיות לתוך כימיקלים אלקטרו וכלי. ישר הדופק iDCs החלת הדופק "EH-100" באמצעות מנגנון אלקטרופורציה II.
- לפני ההליך הניסיוני, להכין השתלים sirna על פי ההוראה של היצרן, סדרת מחלקים ולאחסן אותם ב-20 ° c. להפשיר ולשמור אותם על הקרח בעת שימוש אלה siRNAs עבור אלקטרופורציה. אל תמקם iDCs על קרח ולהימנע שינויים בטמפרטורה. מעבר במהירות ולהימנע דגירה ארוכה של iDCs ב-PBS או מאגר P3.
- ישירות לאחר electroporation, להוסיף 500 μL של טרום מחומם RPMI 1640 האלקטרו כימיקלים. דגירה את התאים בחממה עבור 5-10 דקות. העברת iDCs לתוך 6-צלחות לוחות עם מדיום חדש מחומם מראש DC (בתוספת 40 U/mL של GM-שדרתי ו 250 U/mL של IL-4). לשלב את התנאים המתאימים לאחד טוב, זרע את התאים בריכוז הסופי של 1 · 106/ml ולמקם אותם לתוך אינקובטור.
- 4 h לאחר דגירה, להוסיף את קוקטייל ההבשלה המכילה את ציטוקינים המפורטים בשלב 1.2.2.
הערה: הכינו דוגמה אחת מDCs שאינם אלקטרופורטיים (1 x 106) כבקרה לניתוח ציטומטלי הזרימה 2 ימים של אלקטרופורציה. טיפול בתאי בקרה אנלוגיים לדגימות אלקטרופורנות. - יומיים לכתוב אלקטרופורציה, הקציר תאים על ידי השעיית מחדש ולהעביר לתוך 15 מ ל צינורות. לשטוף את הבארות עם 1 מ ל של PBS ולהעביר את השעיות בצינורות המתאימים. פצל 6 · 10בקרי dc לכל תנאי sirna כפי שמתואר בשלבים הבאים:
- השתמש 0.25 x 106 DCs (electroporated ו-לא electroporated) כדי לבדוק את מצב ההבשלה ואת הכדאיות התאית כמתואר בשלב 2. השתמש בנוגדנים הבאים: CD80-PE-Cy5, CD83-APC, CD86-PE, MHCII-APC-Cy7, וחיים/סגול מת.
- השתמש 0.75 x 106 תאים עבור כתמי אבן מערבית ניתוחים כדי להעריך KIF1B/2a-ספציפי להוריד את היעילות. לאסוף את התאים לתוך 1.5 מ"ל safelock mL על ידי צנטריפוגה ב 3390 x g עבור 1.5 דקות. הכינו lysates תא כפי שמתואר בשלב 3.4 ולבצע ניתוחים מערביים כתמי.
- השתמש בתאים הנותרים (5 x 106) מכל מצב sirna לבצע ניסויים לזיהום HSV-1. עבור כל תנאי ניסיוני, להעביר 2.5 x 106 idcs לתוך 2 צינורות mL, או להדביק אותם עם HSV-1 ב-משרד המידע של 2 או להוסיף מאגר mnt כפקד מבוים. בצע את הזיהום כפי שמתואר בשלב 3.3.
- ב 20 h לאחר זיהום, לקצור את התאים על ידי השעיית מחדש ולהכין לlysates תא עבור ניתוחים מערביים כתמי כדי לאמת את אינדוקציה של מחזור HSV-1 המושרה המכונה אוטומטית, כמתואר לעיל בשלב 3.4.
תוצאות
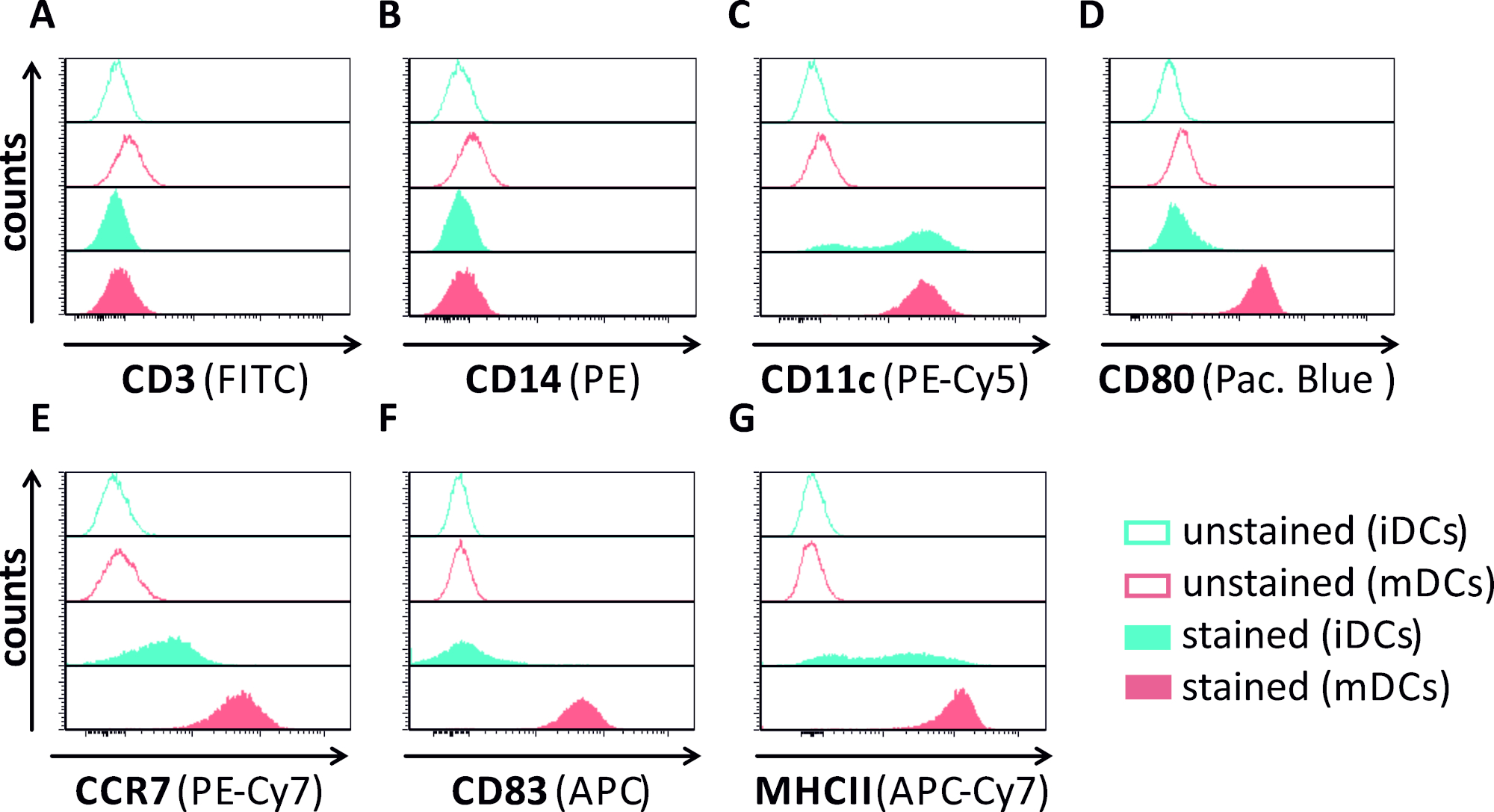
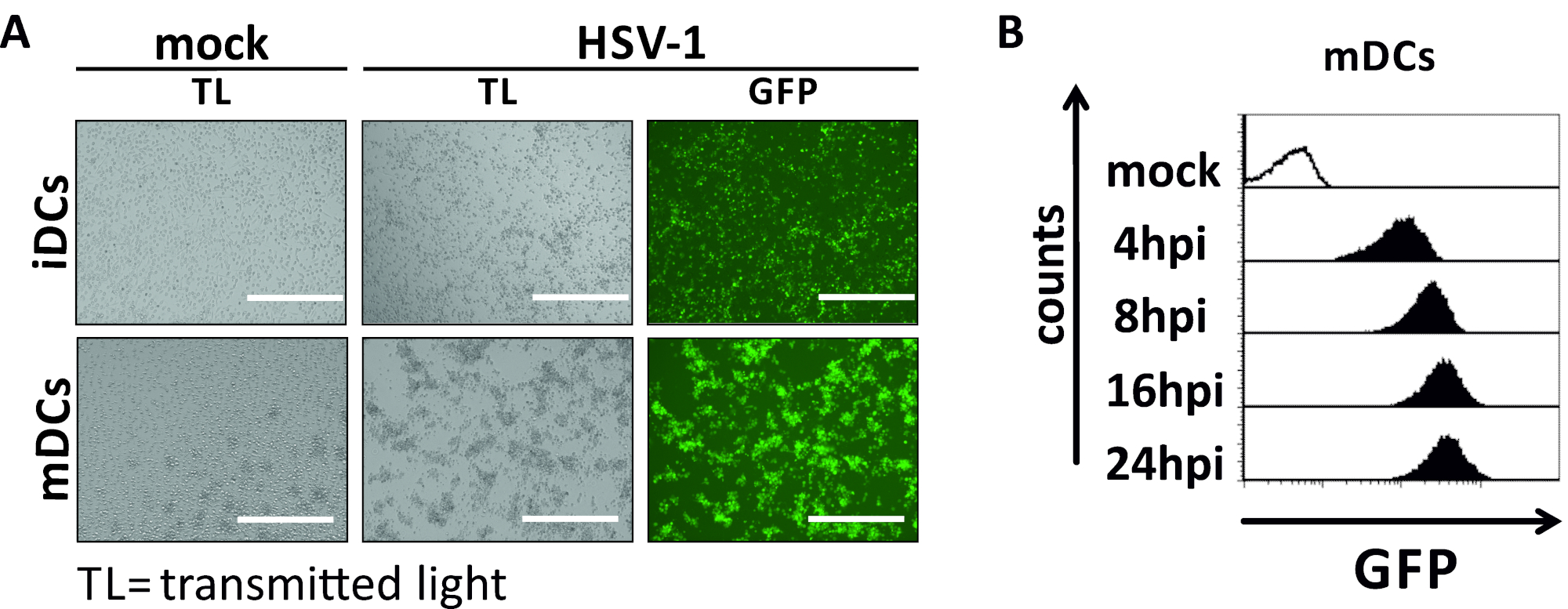
בכתב יד זה, אנו מתארים שיטות להפריע HSV-1-המושרה מכתבים אוטומטיים בתאים דנדריטים. זה כולל את הדור של האדם מונוציט הנגזר iDCs ו mDCs, אשר היו פנופי בדרך כלל שנותחו על ידי זרימה cy, (איור 1). ביום 4 לאחר הדבקות, DCs להראות פנוטיפ בלתי בוגר מאופיין על ידי ביטוי חלש של CD80, CCR7, ו CD83, כמו גם גבוה CD11c וביניים MHCII ביטוי. מאחר שCD3 ואותות CD14 חסרים, תא T ומונוציט זהום ניתנים להוצאה. ביום 6 פוסט הדבקות (כלומר, יום 2 פוסט אינדוקציה של התבגרות), DCs להראות פנוטיפ בוגרת משתקף על ידי עלייה משמעותית CD80, CCR7, CD83, ו MHC-II הביטוי פני השטח. זיהום עם זן eGFP ביטוי HSV-1 (איור 2) תוצאות זיהום כמעט מוחלט של idcs (איור 2הפאנל העליון ) או mdcs (איור 2פאנל תחתון, איור 2ב), מבוסס על אותות GFP חזקים שנותחו על ידי המיקרוסקופיה פלואורסצנטית, כמו גם cy, זרימה הציטונסה.
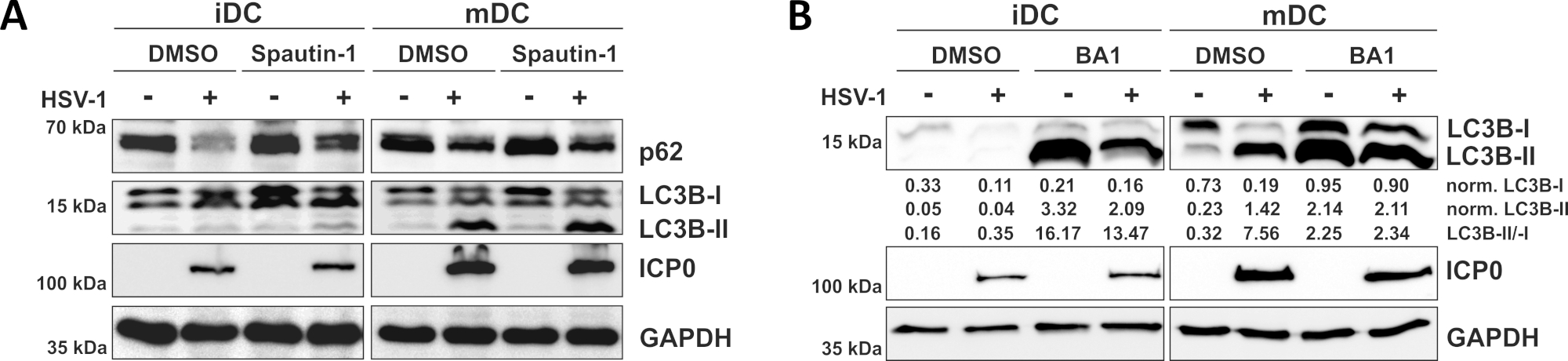
כפי שמתואר הדו ח האחרון שלנו, HSV-1 מעורר באופן אוטומטי שניהם iDCs ו mDCs, אולם, מחזור המכונה אוטומטית מתרחשת ב-iDCs רק10. בגישה הראשונה, התייחסו ל-iDCs ו-mDCs עם spautin-1 (איור 3A)-כדי לחסום את החניכה של התצוגה האוטומטית-או Bafilomycin-A1 (BA1; איור 3ב)-לעכב את מספר האוטומטי הסופי של היתוך. עם הזיהום HSV-1 של iDCs בהעדר spautin-1 ו BA1, השטף הנקרא אוטומטית משתקף על ידי הירידה של p62 ו LC3B ביטוי, בהתאמה. לעומת זאת, הזיהום HSV-1 של mDCs בהיעדר spautin-1 אינו משפיע על ביטוי p62, בעוד spautin-1 ו-BA1 הטיפול לגרום להצטברות של LC3B-II. הדבר משקף את האינדוקציה של הבחירה האוטומטית, אך כישלון במחזור האוטומטי של mDCs. ב-iDCs, מראש הטיפול הראשון spautin משחזר בחוזקה השפלה של הp62 על ידי זיהום HSV-1, עקב עיכוב של התצוגה האוטומטית במהלך שלב החניכה. עם טרום טיפול עם BA1, ללעוג-ו HSV-1-נגועים iDCs להראות הצטברות חזקה של LC3B-II רמות החלבון, המציין עיכוב מוצלח של מחזור אוטומטי ה, באמצעות חסימת מאוחר יותר האוטומטית כמה-lysosome היתוך. בהתאם לזה, spautin-1 ו BA1 טרום טיפול של mDCs גם תוצאות p62 יציב ומוגברת LC3B-II רמות החלבון, בהתאמה.
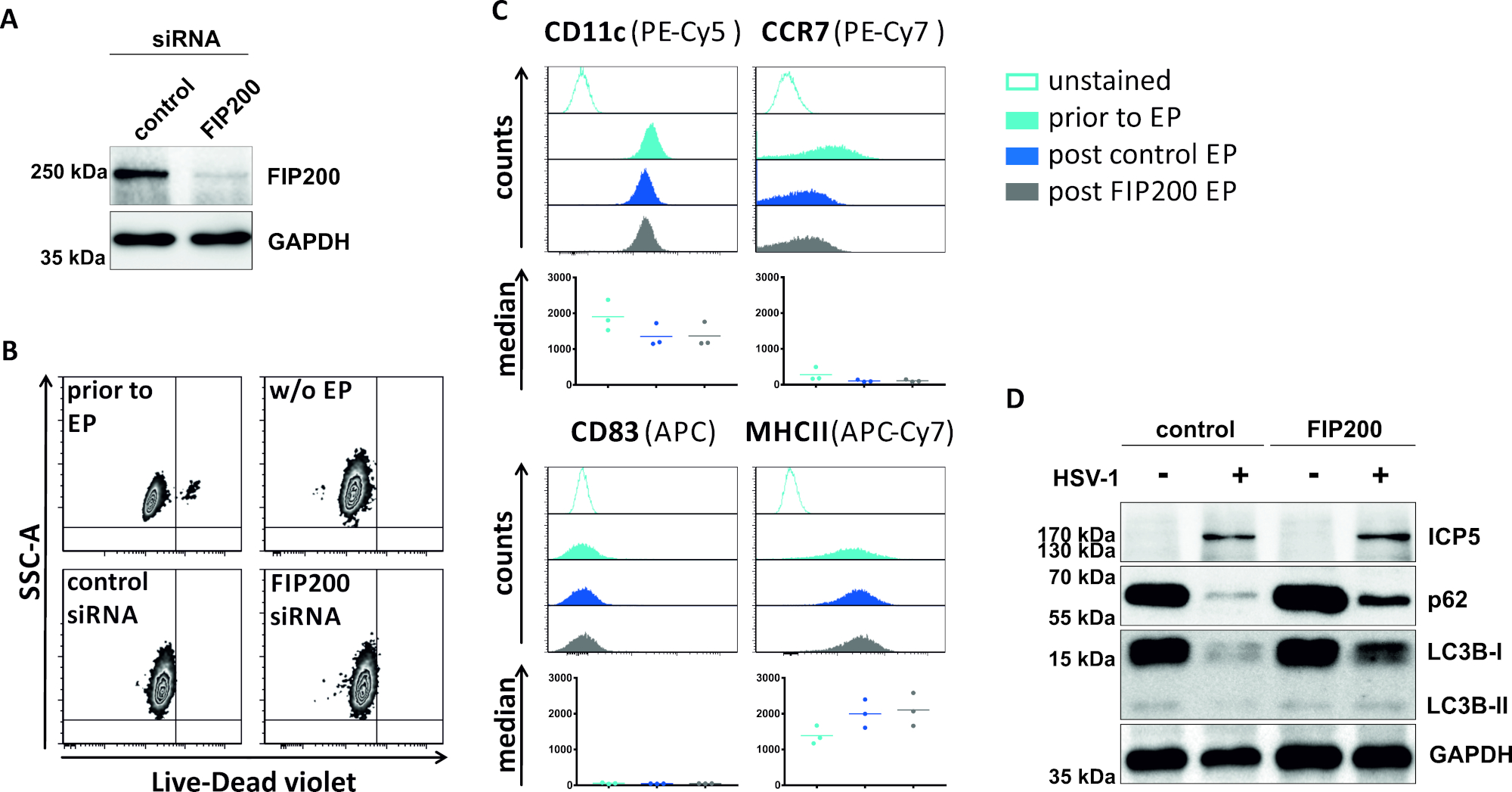
בשיטה השנייה כדי לפגוע השטף האוטומטי השני, sirna אלקטרופורציה מיקוד FIP200 נבדק לגבי יכולתו לחסום שטף הולכה אוטומטי ב HSV-1-נגוע idcs. כפי שמוצג באיור 4A, מופחתת בחריפות FIP200 רמות החלבון זוהו ב-idcs 48 h פוסט אלקטרופורציה, לעומת sirna שליטה. בשלב זה, iDCs לא להראות סימנים של מוות תא (איור 4ב) ולשמור על שלהם פנוטיפ בלתי מפותחים (איור 4ג). זיהום של FIP200-מושתק iDCs עם HSV-1 חושף ירידה חזקה השטף ה, האוטומטי לעומת שלהם שליטה שלהם הטיפול המקבילים (איור 4ד). זה מלווה ברמות חלבונים מוגברת של LC3B, כמו גם p62 כאשר FIP200 מושתקים HSV-1-הנגועים iDCs.
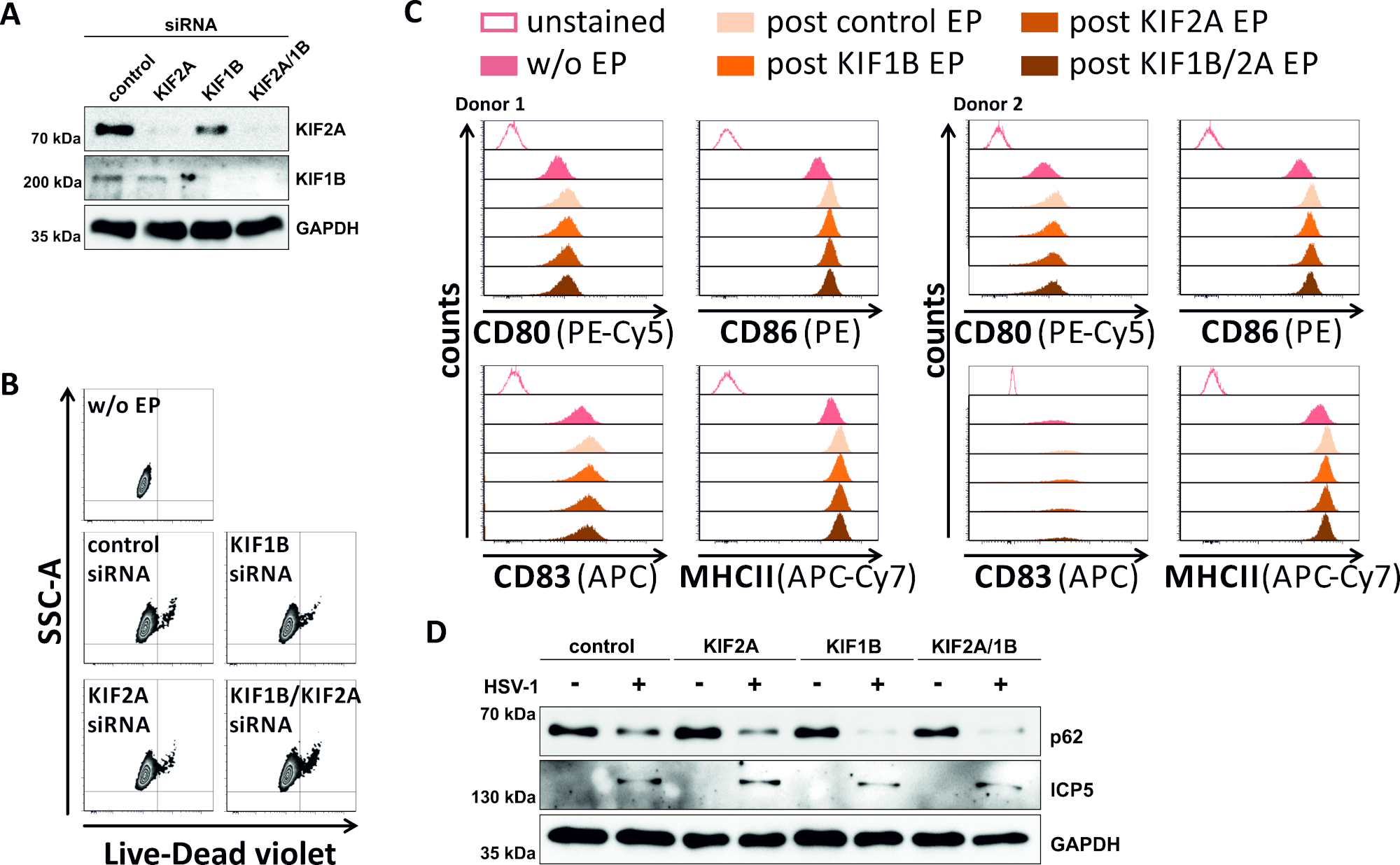
בניסיון הפוך, למדנו אם siRNA-תיווך אבלציה של KIF1B ו KIF2A ביטוי חלבון מאפשר מחזור אוטומטי-lysoזומיום גם ב HSV-1-הנגועים mDCs. לפיכך, iDCs היו electroporated באמצעות מיקוד מסוים siRNAs אחד או שניהם של חלבונים אלה, ותאים הובשילו (איור 5). 2 יומיים אלקטרופורציה, mDCs מראים ירידה חזקה KIF1B ו/או KIF2A חלבון ביטוי, כאשר siRNAs ספציפיים שימשו (איור 5א). שיטה זו גם לא הובילה למוות של תאים בולטים (איור 5ב') וגם לא לשינויים במצב ההבשלה הפנוטיתית שלהם (איור 5ג). תמיכה בחשיבות של KIF1B ו KIF2A במהלך השפלה האוטומטית ליסוזומלית, דלדול שלהם לפני הזיהום HSV-1 מקלה שטף מוגבר מוגברת ב mDCs. זה משתקף על ידי ירידה שיורית רמות החלבון p62, בניגוד למצב שליטה בהתאמה (איור 5ד).

איור 1: איפיון פנוטיאני של האדם הנגזר מסוג idcs ו mdcs באמצעות הזרימה cy, try. DCs נוצרו ומוכתמים בנוגדנים ספציפיים כדי לאמת את הטוהר שלהם: (א) CD3 להוציא תא T זהום, (ב) CD14 להוציא מזהמים עם מונוציטים, ו (ג) CD11c כסמן עבור DCs. כדי להעריך את המצב התבגרותם שלהם בשימוש הנוגדנים הבאים שימשו: (D) CD80, (E) CCR7, (F) CD83, ו (ז) mhcii. מולקולות אלה מבוטאים מאוד על mDCs ובכך לאפשר את האפליה בין הפנוטיפים בוגרים בוגרת DC. הנתונים נותחו באמצעות FCS express 5.0. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 2: מיקרוסקופיים, כמו גם ניתוח הציטוטומטלי של הזרימה של HSV-1-הנגועים idcs ו mdcs. iDCs ו mDCs נדבקו זן HSV-1 המבטא EGFP (HSV-1 EGFP), כדי לאפשר כימות של שיעור זיהום מבוסס על האות GFP. (א) ניתוחים מיקרוסקופיים של gfp-חיובית HSV-1-הנגועים idcs ו mdcs נגועים ב משרד הבית של 2, לעומת עמיתיהם נגוע שלהם, ב 24 hpi. כדי להמחיש את התאים הנגועים, הקרינה הפלואורסצנטית GFP היה מפוקח. סרגל שינוי קנה מידה מייצג 400 μm. (ב) המדידה cy, מדידה של מדגם-או HSV-1-נגוע mdcs במהלך זיהום קינטיקה. לוחות העליון (שחור מרופדת היסטגרמות) להראות מצב מבוים, נמוך לוחות (שחור מלא היסטגרמות) הצג HSV-1 תאים נגועים לאחר נקודות הזמן המצוין לכתוב זיהום. הנתונים נותחו באמצעות FCS express 5.0. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 3: Spautin-1 ו-b afilomycin-A1 לווסת את שטף התלת-מידי האוטומטי ב HSV-1-הנגועים iDCs. iDCs ו mDCs טופלו עם (א) spautin-1 או (ב) BAFILOMYCIN-A1 (BA1) עבור 1 h לפני זיהום. תאים היו לאחר מכן ללעוג-או HSV-1 נגועים (HSV1-RFPVP26) באמצעות משרד הבין 2. לאחר 16-18 h, DCs היו קוצרים וחלבון lysates היו חשופים לחסום המערבי כדי לקבוע את הביטוי של p62 או LC3B-I/-II כסמנים האוטומטיים אוטומטי, ICP0 כמו בקרת זיהום, ו-"העברת בקרת העמסה". רמות החלבון LC3B-I ו-LC3B-II הינן מנורמלות לחלבון הייחוס, באמצעות Bio1D (צפיפות אופטית). היחס של מנורמל LC3B-II לאותות מנורמלים LC3B-I מוצג. דמות זו שונתה והותאמה מ © 2019 Turan ואח ' פורסם במקור ב-JCB. . https://doi.org/10.1083/jcb.20180115110 אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 4: ניתוח של שטף הידרורגיות אוטומטי ב HSV-1-הנגועים iDCs על FIP200-siRNA electroporation. iDCs היו electroporated עם siRNA שליטה או FIP200-ספציפי siRNA באמצעות מנגנון אלקטרופורציה I. (A) DCs נותחו לגבי היעילות של FIP200 הפוסט 48 h הודעה לאחר ההודעה, על ידי ביצוע ניתוחים מערביים כתמי. (ב) הכדאיות התא כמו גם (ג) מעמד התבגרות נותחו לפני אלקטרופורציה (היסטגרמות כחולות בהירים) ו 48 h הצבה (היסטולגרם כחול כהה ואפור) אלקטרופורציה על ידי הזרם cy, לנסות. ערכים חציון עבור שלושה תורמים שונים מוצגים. לאחר המאשרת הנוק-אין יעיל של FIP200 ואת הפנוטיפים בלתי בוגרים, תאים היו HSV-1 נגוע (HSV-1 EGFP) באמצעות משרד המשתמש השני של 2. הנתונים נותחו באמצעות FCS express 5.0. (D) ב 20 h לאחר ההדבקה, תאים היו חשופים ניתוח כתמי אבן המערבי כדי לקבוע את הביטוי של LC3B-I/-II ו p62 כמו סמנים אוטומטיים. ICP5 זוהתה כבקרת זיהום, ו-"העברת שליטה" בתור בקרת הטעינה. פאנלים A ו-D שונו והותאמו מ © 2019 Turan ואח ' פורסם במקור ב-JCB. . https://doi.org/10.1083/jcb.20180115110 אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 5: מחזור siRNA-בתיווך של KIF1B ו/או KIF2A מודוללי מודולים האוטומטיים האוטומטי ב HSV-1-הנגועים mDCs. iDCs היו electroporated עם KIF1B ספציפי ו/או KIF2A-ספציפי siRNA, כמו גם שליטה siRNA, באמצעות מנגנון electroporated השני. ב 4 הפוסט electroporation, התבגרות המושרה באמצעות הוספת קוקטייל התבגרות. ב 48 h לפוסט אלקטרופורציה, בקרי Dc נותחו לגבי (א) היעילות של הסתרה קיף באמצעות בלוק המערבי, (ב) הכדאיות תא כמו גם שלהם (C) פנוטיic מצב התבגרות באמצעות ניתוח cy, שונות תורמים מוצגים). "w/o EP" פירושו ללא אלקטרופורציה, אבל פוסט אינדוקציה של התבגרות; "post בקרת EP" פירושו הודעה פוסט באמצעות siRNA פקד; "post KIF1B, KIF2A, KIF1B/2A EP" פירושו לכתוב אלקטרופורציה באמצעות KIF1B ו/או KIF2A-מיוחד siRNA. לאחר המאשרת מיקוד יעיל של KIF1B ו/או KIF2A ואת הפנוטיפ בוגרת, התאים HSV-1 נגוע (HSV-1 EGFP) באמצעות משרד המשתמש של 2. הנתונים נותחו באמצעות FCS express 5.0. (ד) תאים היו חשופים לכתם מערבי מנתח 20 h לאחר ההדבקה, על מנת להעריך את הביטוי של p62 כסמן המכונה אוטומטית. ICP5 שימש כבקרת מדבקה, ו-"העברת שליטה". דמויות A ו-D שונו והותאמו מ © 2019 Turan ואח ' פורסם במקור ב-JCB. . https://doi.org/10.1083/jcb.20180115110 אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}
Discussion
היקף הפרוטוקול הנוכחי כולל (i) הטיפול של האדם מונוציט הנגזר iDCs, כמו גם mDCs, (ii) הזיהום שלהם עם HSV-1, (iii) הטיפול שלהם עם תרכובות ידוע לעכב את התצוגה האוטומטית, ו (iv) האלקטרופורציה שלהם עם siRNA באמצעות שני הגדרות טכניות שונות. באמצעות הפרוטוקול הנוכחי, השטף הניתן לשימוש אוטומטי יכול להיות חסום HSV-1-נגועים iDCs או המושרה ב HSV-1-הנגועים mDCs.
מאז DCs, ובמיוחד iDCs, הם תאים פגיעים מאוד, עבודה עם תאים אלה כרוך צעדים עדינים למדי. עבור דור DC, אנו ממליצים להשתמש PBMCs מבודדים טריים, וכדי להימנע הקפאת שלהם, כדי לקבל התשואות תאים גבוהים יותר. יתרה מזאת, בעת טיפול ב-iDCs במהלך ניסויים, כולל הטיפוח-תרגול הבאים, מונעים שינויים בטמפרטורה קשה או ממושכת. אחרת, iDCs יכול לעבור שינויים פנוטיפ ולכן יש צורך לאמת הפנוטיפים בלתי בוגרים שלהם על ידי זרימה cy, לנסות. הערה, בניגוד לעמיתיהם הבוגרים, idcs חסרים סמנים שונים, כגון CD80, CD83, ו CD8628,29. הזיהום של idcs ו mdcs עם HSV-1 היא שיטה מבוססת היטב2,3,4,5,6,10. אנו ואחרים הראו כי DCs הם רגישים מאוד עבור זיהום HSV-1, כאשר משרד משנה של 1 או 2 נעשה שימוש (איור 2). בידינו, שמירה על עוצמת הקול של המדיום זיהום ברמות נמוכות (1-3 x 106 תאים ב 250-350 μl) יוביל יעילות זיהום טוב יותר.
גישה קלאסית להתערב עם מסלול סלולרי נתון ברורים הוא השימוש של תרכובות ספציפיות. מגוון של מודולטורים שונים של הבחירה האוטומטית, כלומר , הפעלות כמו גם מעכבי, זמינים כעת30. לגבי HSV-1-המושרה אוטומטי הנגרמת ב DCs, Turan ואח ', (2019) לאחרונה הראה את ההשפעות המעעכבות של spautin-1 ו-bafilomycin-A1 (BA1) במחזור האוטומטי מחזור iDCs10. טכניקה זו עבור עיכוב משני השלבים מתאים לשילוב עם זיהום HSV-1 לאחר מכן, מאז לא שיעור זיהום ולא מעמד התבגרות של DCs (במיוחד iDCs) הוא לקוי. ביישומים עתידיים, הגישה המבוססת על מעכב זה יכולה להיות מיושמת גם בשילוב עם גורמים מדבקים אחרים, תנאי לחץ, כגון רעב, כמו גם עבור סוגי תאים שונים. עם זאת, בעת שימוש במעכבי, מגבלות עולות בקביעת הריכוז המתאים לעיכוב יעיל באופן אוטומטי של המערכת, מבלי להשפיע באופן חמור על הכדאיות התאית. המגבלה העיקרית בעת שימוש מעכבי היא, עם זאת, התרחשות של פוטנציאל מחוץ ליעד או תופעות לוואי, אשר עלול להוביל לתוצאות מטעה31,32.
הגישה השנייה כדי להפריע האוטומטית, מכוסה בפרוטוקול הנוכחי, הוא הנוק הספציפי באמצעות sirna33,34,35. מצד אחד, השתמשנו מנגנון אלקטרופורציה אני במיוחד לאחר הביטוי של FIP200, ובכך מעכב HSV-1 המושרה מחזור השני באופן אוטומטי ב-iDCs. מצד שני, השתקתי שני חלבונים שונים של הקיף (כלומר, KIF1B ו KIF2A), באמצעות מנגנון אלקטרופורציה II, כדי להקל על השטף הדו האוטומטי ב HSV-1 הנגועים mDCs. הפרוטוקולים של אלקטרופורציה הובילו לצריבה כמעט מלאה של FIP200 ב-iDCs, ו-KIF1B/KIF2A ב-mDCs, אשר אומת באמצעות ניתוחיכתמי אבן מערביים(איור 4א, איור 5א). בניגוד למנגנון אלקטרופורציה I, אשר אינו משפיע על הכדאיות של DCs, אלקטרופורציה של mDCs באמצעות מנגנון אלקטרופורציה השני מביא שיעור גבוה יותר של תאים מתים (איור 4ב', איור 5ב ). מכאן, ביישומים עתידיים, מנגנון האלקטרופורציה אני צריך להיות מעדיף בשימוש עבור שניהם, iDCs ו-mDCs. באופן מדהים, שניהם siRNA מבוססי טכניקות, כדי לווסת שטף האנטי אוטומטי, הם תואמים HSV-1 בעקבות הזיהום של iDCs או mDCs. יתרה מזאת, לא הפנוטיפ הילדותי של iDCs או הפנוטיפים הבוגרים של mDCs שינו את הפוסט-אלקטרופורציה.
אלקטרופורציה של iDCs באמצעות FIP200-ספציפי siRNA היא שיטה יעילה וספציפית מאוד עבור הסתרה גנטית, כמו גם עיכוב של שטף הכולינרגיות אוטומטית על זיהום HSV-1. בנוסף להשתקה הספציפית של FIP200, ניתן להתאים פרוטוקול זה כדי להשתיק רכיבים אוטומטיים אחרים, המשתתפים בשלבים שונים במהלך המפל האוטומטי. עם זאת, זיהוי היעד המתאים לעיכוב יעיל siRNA-בתיווך של התצוגה האוטומטית כוללת מספר היבטים של דאגה. ראשית, מנומת יעילות של גנים הקשורים לתצוגה מקדימה (ATG) אינו בהכרח לתאם חיובי עם עיכוב יעיל של התצוגה האוטומטית והוא תלוי מאוד בחלבון ATG ספציפי כי הוא מושתק36. שנית, חלבונים atg ברורים מעורבים בנוסף מסלולים ברורים מפני התצוגה האוטומטית, ולכן אבלציה שלהם יכול גם להוביל תופעות לוואי שליליות37,38,39. שלישית, ATGs שונים עשויים להיות פונקציות עודפות, ולכן מנוק של רכיב אחד לא יכול להיות מספיק כדי לעכב את הבחירה האוטומטית (g., beclin-1 ו-beclin-2)40.
בנוסף, פרוטוקול האלקטרופורציה המבוססת על מנגנוני החשמל של DCs מתאים גם ל-mRNAs, וניתן להשתמש בו למגוון סוגי תאים ראשיים נוספים, כגון PBMCs25. מערכת זו מספקת ובכך אסטרטגיה כללית כדי לספק מינים שונים של RNA לתוך סוגי תאים הראשי השונים. לסיכום, אנו מציגים שני פרוטוקולים כדי לעכב שטף הכולינרגיות אוטומטי, על ידי שימוש בגישה מעכב או siRNA מבוססי בשילוב עם זיהום HSV-1 הבאים של iDCs. יתר על כן, אנו מתארים גישה sirna אלקטרופורציה כדי לזרז שטף הכולינרגיות ב mdcs על זיהום HSV-1.
Disclosures
. למחברים אין מה לגלות
Acknowledgements
עבודה זו נתמכת על ידי מועצת המחקר הגרמנית (DFG) באמצעות הפרויקט STE 432/11-1 הוענק כמו ובאמצעות תוכנית ELAN מהפקולטה לרפואה (פרידריך-אלכסנדר-אוניברסיטת ארלנגן-נירנברג) דרך הפרויקט 18-12-21-1, שהוענקו LG.
Materials
| Name | Company | Catalog Number | Comments |
| 4D-Nucleofector Core Unit (electroporation apparatus II) | Lonza (Basel, Switzerland) | AAF-1002B | |
| AB-Serum | Sigma Aldrich Chemie GmbH (Steinheim, Germany) | H4522 | Dendritic cell cultivation |
| ACD-A | Sigma-Aldrich Chemie GmbH (Steinheim, Germany) | 9007281 | |
| Amaxa P3 Primary Cell 4D-Nucleofector X Kit L (electroporation kit apparatus II) | Lonza (Basel, Switzerland) | V4XP-3024 | |
| Amersham ECL Prime Western Blotting Detection Reagent | GE Healthcare (Solingen, Germany) | RPN2232 | Western Blot Detection |
| Ammonium persulfate (APS) | Sigma Aldrich Chemie GmbH (Steinheim, Germany) | A3678 | |
| anti-mouse-IgG (mouse, polyclonal, HRP) | Cell Signaling (Leiden, Netherlands) | 7076 | Western Blot detection |
| anti-rabbit-IgG (goat, polyclonal, HRP) | Cell Signaling (Leiden, Netherlands) | 7074 | Western Blot detection |
| Bafilomycin A1 | Sigma-Aldrich Chemie GmbH (Steinheim, Germany) | tlrl-baf1 | inhibition of autophagy and lysosomal degradation |
| BD FACS Canto II Flow Cytometer | BD Biosciences (Heidelberg, Germany) | 338962 | |
| Benzonase | Sigma-Aldrich Chemie GmbH (Steinheim, Germany) | E1014 | |
| Blotting Chamber Fastblot B44 | Biometra (Göttingen, Germany) | 846-015-100 | |
| CCR7 (mouse, Pe-Cy7) | BioLegend (Fell, Germany) | 557648 | Flow cytometry Dilution: 1:100 Clone: G043H7 |
| CD11c (mouse, Pe-Cy5) | BD Biosciences (Heidelberg, Germany) | 561692 | Flow cytometry Dilution: 1:100 Clone: B-ly6 |
| CD14 (mouse, PE) | BD Biosciences (Heidelberg, Germany) | 555398 | Flow cytometry Dilution: 1:100 Clone: M5E2 |
| CD3 (mouse, FITC) | BD Biosciences (Heidelberg, Germany) | 555332 | Flow cytometry Dilution: 1:100 Clone: UCHT1 |
| CD80 (mouse, V450) | BD Biosciences (Heidelberg, Germany) | 560442 | Flow cytometry Dilution: 1:100 Clone: L307.4 |
| CD83 (mouse, APC) | eBioscience Thermo Fisher Scientific (Langenselbold, Germany) | 17-0839-41 | Flow cytometry Dilution: 1:200 Clone: HB15e |
| CD86 (mouse, PE) | BD Biosciences (Heidelberg, Germany) | 553692 | Flow cytometry Dilution: 1:100 |
| EVOS FL Cell Imaging | System AMG/Life Technologies (Carlsbad, USA) | AMF4300 | |
| FIP200 (rabbit) | Cell Signaling (Leiden, Netherlands) | 12436 | Western Blot detection Dilution: 1:1000 Clone: D10D11 |
| GAPDH (mouse) | Merck Millipore (Massachusetts, USA) | AB2302 | Western Blot detection Dilution: 1:5000 Clone: MAB374 |
| Gene Pulser II apparatus (electroporation apparatus I) | BioRad Laboratories GmbH (München, Germany) | 165-2112 | |
| GM-CSF (4x104 U/mL) | Miltenyi Biotec (Bergisch Gladbach, Germany) | 130-093-868 | |
| HLA?DR (mouse, APC-Cy7) | BioLegend (Fell, Germany) | 307618 | Flow cytometry Dilution: 1:200 Clone: L243 |
| HSV-1/17+/CMV-EGFP/UL43 | BioVex | DC infection | |
| ICP0 (mouse) | Santa Cruz Biotechnology (St. Cruz; Dallas, Texas, USA) | sc-53070 | Western Blot detection Dilution: 1:1000 Clone: 11060 |
| ICP5 (mouse) | Santa Cruz Biotechnology (St. Cruz; Dallas, Texas, USA) | sc-56989 | Western Blot detection Dilution: 1:1000 Clone: 3B6 |
| IL-1β (0.1x106 U/mL) | Cell Genix GmbH (Freiburg, Germany) | 1411-050 | |
| IL-4 (1x106 U/mL) | Miltenyi Biotec (Bergisch Gladbach, Germany) | 130-093-924 | |
| IL-6 (1x106 U/mL) | Cell Genix GmbH (Freiburg, Germany) | 1404-050 | |
| ImageQuant LAS 4000 | GE Healthcare (Solingen, Germany) | 28955810 | |
| KIF1B (mouse) | Santa Cruz Biotechnology (St. Cruz; Dallas, Texas, USA) | sc-376246 | Western Blot detection Dilution: 1:1000 Clone: E-12 |
| KIF2A (mouse) | Santa Cruz Biotechnology (St. Cruz; Dallas, Texas, USA) | sc-271471 | Western Blot detection Dilution: 1:1000 Clone: D-7 |
| LC3B (rabbit) | Cell Signaling (Leiden, Netherlands) | 3868 | Western Blot detection Dilution: 1:1000 Clone: D11 |
| L-glutamine | Lonza (Basel, Switzerland) | 17-605E | |
| LIVE/DEAD Fixable Violet dead cell stain kit | Life Technologies (Carlsbad, CA, USA) | L34964 | L/D staining in Flow cytometry |
| Lymphoprep | Alere Technologies AS (Oslo, Norway) | 04-03-9391/01 | |
| Magnesiumchloride | Carl Roth GmbH (Karlsruhe, Germany) | A537.1 | |
| Megafuge 2.0 RS | Heraeus (Hanau, Germany) | 75015505 | |
| N, N, N', N'-Tetramethylethylendiamine (TEMED) | Sigma-Aldrich Chemie GmbH (Steinheim, Germany) | T9281 | |
| Neubauer counting chamber | Brand (Wertheim, Germany) | 717805 | |
| Nunc Cell culture flasks (175.0 cm2) | Thermo Scientific (Rockford, USA) | 159910 | |
| p62 (rabbit) | Cell Signaling (Leiden, Netherlands) | 88588 | Western Blot detection Dilution: 1:1000 Clone: D5L7G |
| PageRuler prestained protein ladder | Thermo Fisher Scientific (Langenselbold, Germany) | 26616 | |
| Paraformaldehyde, 16 % | Alfa Aesar, Haverhill, USA | 43368.9M | |
| PerfectSpin 24 Plus | Peqlab (Erlangen, Germany) | C2500-R-PL | |
| PGE2 (1 mg/mL) | Pfizer (Berlin, Germany) | BE130681 | |
| Phosphate buffered saline (PBS) | Lonza (Basel, Switzerland) | 17-512F | |
| Protein gel system MiniProtean II | Bio-Rad Laboratories GmbH (München, Germany) | 1652960 | |
| RestoreTM Western Blot Stripping Buffer | Thermo Scientific, Rockford, USA | 21059 | |
| Rocking Platform wt 15 | Biometra (Göttingen, Germany) | 042-590 | |
| RotiBlock | Carl Roth GmbH (Karlsruhe, Germany) | A151.4 | |
| Roti-Load 1 (4x) | Carl Roth GmbH (Karlsruhe, Germany) | K929.3 | |
| Rotiphorese Gel 30 (37.5:1) | Carl Roth GmbH (Karlsruhe, Germany) | 3029.1 | |
| RPMI 1640 | Lonza (Basel, Switzerland) | 12-167F | |
| Sodium dodecyl Sulfate (SDS) | Carl Roth GmbH (Karlsruhe, Germany) | 2326.2 | |
| Thermomixer comfort | Eppendorf (Hamburg, Germany) | 5355 000.011 | |
| TNF-α (10 μg/mL) | Peprotech (Hamburg, Germany) | 300-01A | |
| Tris | Carl Roth GmbH (Karlsruhe, Germany) | 4855.3 | |
| Trypan blue solution (0.4 %) | Sigma-Aldrich Chemie GmbH (Steinheim, Germany) | T8154 | |
| Tween 20 | Carl Roth GmbH (Karlsruhe, Germany) | 9127.1 | |
| Whatman 0.2 μm nitrocellulose membrane | GE Healthcare (Solingen, Germany) | 10600001 | |
| WhatmanTM Chromatography Paper 3 mm Chr | Fisher Scientific GmbH (Schwerte, Germany) | 3030917 |
References
- Chapuis, F., Rosenzwajg, M., Yagello, M., Ekman, M., Biberfeld, P., Gluckman, J. C. Differentiation of human dendritic cells from monocytes in vitro. European Journal of Immunology. 27 (2), 431-441 (1997).
- Kummer, M., et al. Herpes simplex virus type 1 induces CD83 degradation in mature dendritic cells with immediate-early kinetics via the cellular proteasome. Journal of Virology. 81 (12), 6326-6338 (2007).
- Kruse, M., et al. Mature dendritic cells infected with herpes simplex virus type 1 exhibit inhibited T-cell stimulatory capacity. Journal of Virology. 74 (15), 7127-7136 (2000).
- Salio, M., Cella, M., Suter, M., Lanzavecchia, A. Inhibition of dendritic cell maturation by herpes simplex virus. European Journal of Immunology. 29 (10), 3245-3253 (1999).
- Prechtel, A. T., et al. Infection of mature dendritic cells with herpes simplex virus type 1 dramatically reduces lymphoid chemokine-mediated migration. Journal of General Virology. 86, 1645-1657 (2005).
- Theodoridis, A. A., Eich, C., Figdor, C. G., Steinkasserer, A. Infection of dendritic cells with herpes simplex virus type 1 induces rapid degradation of CYTIP, thereby modulating adhesion and migration. Blood. 118 (1), 107-115 (2011).
- Cohrs, R. J., Gilden, D. H. Human herpesvirus latency. Brain Pathology. 11 (4), 465-474 (2001).
- Grinde, B. Herpesviruses: latency and reactivation - viral strategies and host response. Journal of Oral Microbiology. 5, (2013).
- Whitley, R. J., Roizman, B. Herpes simplex virus infections. The Lancet. 357 (9267), 1513-1518 (2001).
- Turan, A., et al. Autophagic degradation of lamins facilitates the nuclear egress of herpes simplex virus type 1. The Journal of Cell Biology. 218 (2), 508-523 (2019).
- Takeshige, K., Baba, M., Tsuboi, S., Noda, T., Ohsumi, Y. Autophagy in yeast demonstrated with proteinase-deficient mutants and conditions for its induction. The Journal of Cell Biology. 119 (2), 301-311 (1992).
- Yin, Z., Pascual, C., Klionsky, D. J. Autophagy: machinery and regulation. Microbial Cell. 3 (12), 588-596 (2016).
- Bodemann, B. O., et al. RalB and the exocyst mediate the cellular starvation response by direct activation of autophagosome assembly. Cell. 144 (2), 253-267 (2011).
- Bjorkoy, G., et al. p62/SQSTM1 forms protein aggregates degraded by autophagy and has a protective effect on huntingtin-induced cell death. The Journal of Cell Biology. 171 (4), 603-614 (2005).
- Kabeya, Y., et al. LC3, a mammalian homologue of yeast Apg8p, is localized in autophagosome membranes after processing. The EMBO Journal. 19 (21), 5720-5728 (2000).
- Kabeya, Y., Mizushima, N., Yamamoto, A., Oshitani-Okamoto, S., Ohsumi, Y., Yoshimori, T. LC3, GABARAP and GATE16 localize to autophagosomal membrane depending on form-II formation. Journal of Cell Science. 117, 2805-2812 (2004).
- Pankiv, S., et al. p62/SQSTM1 binds directly to Atg8/LC3 to facilitate degradation of ubiquitinated protein aggregates by autophagy. The Journal of Biological Chemistry. 282 (33), 24131-24145 (2007).
- Korolchuk, V. I., Rubinsztein, D. C. Regulation of autophagy by lysosomal positioning. Autophagy. 7 (8), 927-928 (2011).
- Li, Y., et al. A cell-based quantitative high-throughput image screening identified novel autophagy modulators. Pharmacological Research. 110, 35-49 (2016).
- Pampaloni, F., et al. A Novel Cellular Spheroid-Based Autophagy Screen Applying Live Fluorescence Microscopy Identifies Nonactin as a Strong Inducer of Autophagosomal Turnover. SLAS Discovery. 22 (5), 558-570 (2017).
- Deng, Y., Zhu, L., Cai, H., Wang, G., Liu, B. Autophagic compound database: A resource connecting autophagy-modulating compounds, their potential targets and relevant diseases. Cell Proliferation. 51 (3), 12403 (2018).
- Liu, J., et al. Beclin1 controls the levels of p53 by regulating the deubiquitination activity of USP10 and USP13. Cell. 147 (1), 223-234 (2011).
- Mauvezin, C., Neufeld, T. P. Bafilomycin A1 disrupts autophagic flux by inhibiting both V-ATPase-dependent acidification and Ca-P60A/SERCA-dependent autophagosome-lysosome fusion. Autophagy. 11 (8), 1437-1438 (2015).
- Yoshimori, T., Yamamoto, A., Moriyama, Y., Futai, M., Tashiro, Y. Bafilomycin A1, a specific inhibitor of vacuolar-type H(+)-ATPase, inhibits acidification and protein degradation in lysosomes of cultured cells. Journal of Biological Chemistry. 266 (26), 17707-17712 (1991).
- Gerer, K. F., Hoyer, S., Dorrie, J., Schaft, N. Electroporation of mRNA as Universal Technology Platform to Transfect a Variety of Primary Cells with Antigens and Functional Proteins. Methods in Molecular Biology. 1499, 165-178 (2017).
- Prechtel, A. T., Turza, N. M., Theodoridis, A. A., Steinkasserer, A. CD83 knockdown in monocyte-derived dendritic cells by small interfering RNA leads to a diminished T cell stimulation. The Journal of Immunology. 178 (9), 5454-5464 (2007).
- Pfeiffer, I. A., et al. Leukoreduction system chambers are an efficient, valid, and economic source of functional monocyte-derived dendritic cells and lymphocytes. Immunobiology. 218 (11), 1392-1401 (2013).
- Villadangos, J. A., Heath, W. R. Life cycle, migration and antigen presenting functions of spleen and lymph node dendritic cells: limitations of the Langerhans cells paradigm. Seminars in Immunology. 17 (4), 262-272 (2005).
- Lechmann, M., Berchtold, S., Hauber, J., Steinkasserer, A. CD83 on dendritic cells: more than just a marker for maturation. Trends in Immunology. 23 (6), 273-275 (2002).
- Yang, Y. P., et al. Application and interpretation of current autophagy inhibitors and activators. Acta Pharmacologica Sinica. 34 (5), 625-635 (2013).
- Redmann, M., et al. Inhibition of autophagy with bafilomycin and chloroquine decreases mitochondrial quality and bioenergetic function in primary neurons. Redox Biology. 11, 73-81 (2017).
- Yan, Y., et al. Bafilomycin A1 induces caspase-independent cell death in hepatocellular carcinoma cells via targeting of autophagy and MAPK pathways. Scientific Reports. 6, 37052 (2016).
- Hale, C. M., et al. Identification of modulators of autophagic flux in an image-based high content siRNA screen. Autophagy. 12 (4), 713-726 (2016).
- Lipinski, M. M., et al. A genome-wide siRNA screen reveals multiple mTORC1 independent signaling pathways regulating autophagy under normal nutritional conditions. Developmental Cell. 18 (6), 1041-1052 (2010).
- Orvedahl, A., et al. Image-based genome-wide siRNA screen identifies selective autophagy factors. Nature. 480 (7375), 113-117 (2011).
- Staskiewicz, L., Thorburn, J., Morgan, M. J., Thorburn, A. Inhibiting autophagy by shRNA knockdown: cautions and recommendations. Autophagy. 9 (10), 1449-1450 (2013).
- Lee, I. H., et al. Atg7 modulates p53 activity to regulate cell cycle and survival during metabolic stress. Science. 336 (6078), 225-228 (2012).
- Fremont, S., Gerard, A., Galloux, M., Janvier, K., Karess, R. E., Berlioz-Torrent, C. Beclin-1 is required for chromosome congression and proper outer kinetochore assembly. EMBO Reports. 14 (4), 364-372 (2013).
- Bestebroer, J., V'kovski, P., Mauthe, M., Reggiori, F. Hidden behind autophagy: the unconventional roles of ATG proteins. Traffic. 14 (10), 1029-1041 (2013).
- Galluzzi, L., Kroemer, G. Common and divergent functions of Beclin 1 and Beclin 2. Cell Research. 23 (12), 1341-1342 (2013).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved