
A subscription to JoVE is required to view this content. Sign in or start your free trial.
Method Article
הפעלה מחדש של מודלים של תאים מדולדלים בכלמידומונס ריינהרדטי
In This Article
Summary
הפעלה מחדש במבחנה של תאי תנועתיות היא ניסוי מכריע בהבנת המנגנונים של תנועתיות התאים. הפרוטוקול מתאר הפעלה מחדש של מודלי התאים המדולדלים של Chlamydomonas reinhardtii, אורגניזם מודל לחקר ריסונים/פלגלה.
Abstract
מאז הניסוי ההיסטורי על התכווצות שריר גליצרין על ידי הוספת ATP, אותו הדגים Szent-Györgyi באמצע המאה ה-20, הפעלה מחדש במבחנה של תאים שעברו דה-ממבראן הייתה דרך מסורתית ורבת עוצמה לבחון את תנועתיות התאים. היתרון הבסיסי של שיטת ניסוי זו הוא כי הרכב של פתרון ההפעלה מחדש עשוי להשתנות בקלות. לדוגמה, ניתן לשכפל במעבדה סביבת ריכוז גבוהה של Ca2+ המתרחשת רק באופן זמני עקב עירור ממברנה in vivo . ריסונים אאוקריוטיים (באנגלית: Eukaryotic cilia) הם מכונות תנועתיות משוכללות שמנגנוני הוויסות שלהן עדיין לא ברורים. האצה הירוקה החד-תאית Chlamydomonas reinhardtii היא אורגניזם מודל מצוין בתחום המחקר של הריסונים. ניסויי ההפעלה מחדש באמצעות מודלים של תאים שעברו דה-ממברניזציה של C. reinhardtii ונגזרותיהם, כגון אקסונמות מדולדלות של ריסונים מבודדים, תרמו באופן משמעותי להבנת המנגנונים המולקולריים של תנועתיות סילארית. ניסויים אלה הבהירו כי ATP ממריץ תנועתיות ציליארית וכי אותות תאיים שונים, כולל Ca2+, cAMP ומיני חמצן תגובתי, מווסתים תנועות סילאריות. השיטה המדויקת לדה-ממברנציה של תאי C. reinhardtii ולהפעלה מחדש של מודלי התאים מתוארת כאן.
Introduction
הפעלה מחדש במבחנה של תאי תנועתיות שעברו דה-ממברנות היא כלי רב ערך לחקר הבסיס המולקולרי למנגנון הוויסות של תנועתיות התא. Szent-Györgyi הדגים לראשונה התכווצות במבחנה של סיבי שרירי השלד של ארנב המופקים מ-50% גליצרול על ידי הוספת אדנוזין טריפוספט (ATP)1. ניסוי זה היה הראשון שהוכיח כי ATP ממריץ התכווצות שרירים. המתודולוגיה יושמה במהרה בחקר תנועתיות סילארית/פלאגלרית המופעלת על ידי ATP, כגון פלגלה זרע2, ריסונים Paramecium 3, ו- Chlamydomonas reinhardtii cilia (הנקראת גם פלגלה)4 תוך שימוש בדטרגנטים לא יוניים לדה-ממברנציה.
האצה הירוקה החד-תאית C. reinhardtii היא אורגניזם מודל לחקר ריסונים: היא שוחה עם שני ריסונים על ידי הכאתם כמו מכת חזה של אדם5. תנועה סילארית מונעת על ידי dynein, חלבון מוטורי מבוסס מיקרוטובול מכוון קצה מינוס 6,7. ניתן לסווג את הדיינינים הסילאריים לדיינינים בעלי זרוע חיצונית ולדיינינים בעלי זרוע פנימית. מוטנטים שחסרים להם כל סוג של דיינין בודדו כמוטנטים שוחים לאט עם הפרעות שונות בתנועתיות. ניתוח תנועתיות מבחנה מפורט של מוטנטים אלה קידם באופן משמעותי את מחקר הדיינין8.
ממצאים חשובים רבים הושגו תוך שימוש בשיטה זו ובנגזרותיה מאז הוקם ניסוי ההפעלה מחדש במבחנה של תאי C. reinhardtii (מודלים של תאים) שעברו דה-ממברנטציה. הפעלה מחדש של מודלים של תאים בסדרה של מאגרי Ca2+, למשל, הראתה9 ששתי ריסונים מווסתים באופן שונה על ידי תת-מיקרומולאר Ca2+, ובקרת ריסונים אסימטרית זו מאפשרת את הכיוון הפוטוטקטי של C. reinhardtii10. יתר על כן, שתי הריסונים מראים המרה של צורת גל ממצב השחייה הקדמית (הנקראת צורת גל אסימטרית) למצב השחייה לאחור (הנקרא צורת גל סימטרית המופיעה לתקופה קצרה כאשר התאים הם בהלם פוטו או מכני)11,12. המרת צורת גל זו מווסתת על ידי תת-מילימולאר Ca2+, אשר הודגם על ידי הפעלה מחדש של מה שמכונה מנגנון נוקלאופלגלר (קומפלקס המכיל שני ריסונים, הגופים הבסיסיים, המבנים המקשרים בין הגופים הבסיסיים לגרעין, ושריד הגרעין)11 או אקסונמים מתפרקים של ריסונים מבודדים13. מלבד Ca2+, חמצון-חיזור (חמצון-הפחתה) הוא אות המווסת את תדירות הפעימה הציליארית, אשר הוכחה על ידי הפעלה מחדש של מודלים תאיים במאגרי חמצון-חיזור המכילים יחסים שונים של גלוטתיון מופחת לעומת גלוטתיון מחומצן14. בנוסף, אדנוזין מונופוספט מחזורי (cAMP) מווסת באופן אסימטרי שני ריסונים, אשר הודגם על ידי הפעלה מחדש של אקסונמים עם cAMP15 כלוב פוטו-מחשוף. ממצאים אלה במבחנה, בשילוב עם ממצאים גנטיים, הובילו להבנה מעמיקה יותר של המנגנונים המולקולריים של ויסות הריסונים ב- C. reinhardtii.
פרוטוקול להפעלה מחדש של מודלי התאים מתואר כאן. השיטה היא פשוטה, מאפשרת שינויים שונים, וניתן ליישם אותה על אורגניזמים מרובים הנעים עם ריסונים. עם זאת, מאחר שתאים שעברו דה-ממברניזציה הם שבריריים, זה דורש כמה טיפים כדי להפעיל מחדש את התנועתיות של מודלים של תאים ביעילות טובה תוך מניעת דצילציה.
Protocol
זן פראי של Chlamydomonas reinhardtii, CC-125, שימש למחקר הנוכחי. CC-125 התקבל ממרכז המשאבים של כלמידומונס (ראה טבלת חומרים) ונשמר על טריס-אצטט-פוספט (TAP)16, 1.5% בינוני אגרוז ב 20-25 מעלות צלזיוס.
1. תרבית תאים
- תרבית Chlamydomonas reinhardtii (CC-125) במדיום TAP16 בתקופה של 12 שעות/12 שעות של אור-חושך (תנאי אור לתקופת האור: ~50 μmol פוטונים m−2 s−1 אור לבן) בטמפרטורה של 20-25 מעלות צלזיוס למשך יומיים (איור 1).
הערה: התאים חייבים להיות בשלב גדילה לוגריתמי באמצע (סרט 1). תרבית ארוכה (>4 ימים, בצמיחה הלוגריתמית המאוחרת או בשלב הנייח) מפחיתה את יעילות ההפעלה מחדש של מודלים של תאים שעברו דה-ממברנות.

איור 1: תרבית נוזלית לאחר התבגרות של יומיים. מתוך צלחת אגר TAP-1.5%, נתח של תאים מסוג בר למילוי לולאת הפלטינה חוסן לכ-150 מ"ל ל-TAP נוזלי בינוני בבקבוקון. צפיפות התא לאחר תרבית של יומיים הייתה 2.3 × 106 תאים/מ"ל. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
{kind=link}
סרט 1: שחייה של תאים חיים. התאים נצפו תחת מיקרוסקופ עם עדשה אובייקטיבית פי 10 ומעבה שדה כהה טבילת שמן. סרגל קנה מידה = 100 μm. אנא לחץ כאן כדי להוריד סרט זה.
2. הכנת מודלים של תאים שעברו דה-ממברנות
הערה: לפני שתתחיל בניסוי, שמור את חיץ הכביסה בטמפרטורת החדר ואת הדה-ממברנציה, הדילול, מאגרי ההפעלה מחדש ופתרון ה-ATP על הקרח. ההרכב של מאגרים אלה מובא בטבלה משלימה 1.
- צנטריפוגה ~ 10 מ"ל של תרבית נוזלית ב 1000 × גרם ב 20 מעלות צלזיוס למשך 3 דקות.
הערה: לאורך כל הפרוטוקול משלב זה, אין להשתמש בכלי פלסטיק אוטומטיים (צינורות, טיפים לפיפטה וכו '), ובכך להפחית את יעילות ההפעלה מחדש. עבור צינורות חרוטיים, אלה בשימוש חוזר עדיפים. צפיפות התא לאחר תרבית של יומיים עשויה להיות בדרך כלל 1.0-5.0 × 106 תאים/מ"ל. כאשר צפיפות התאים נמוכה מזו, קח מספיק נפח תרבית כדי להכיל ~ 5 × 107 תאים. - יש להשליך את הסופרנאטנט, תחילה על ידי דקאנטציה ולאחר מכן על ידי פיפטה של פסטר, ולהחזיר את המשקעים לכ-5 מ"ל של חיץ כביסה.
- תאי צנטריפוגה במאגר הכביסה ב 1000 × גרם במשך 3 דקות ב 20 מעלות צלזיוס.
- השליכו את ה-supernatant בזהירות באמצעות פיפטה (איור 2).
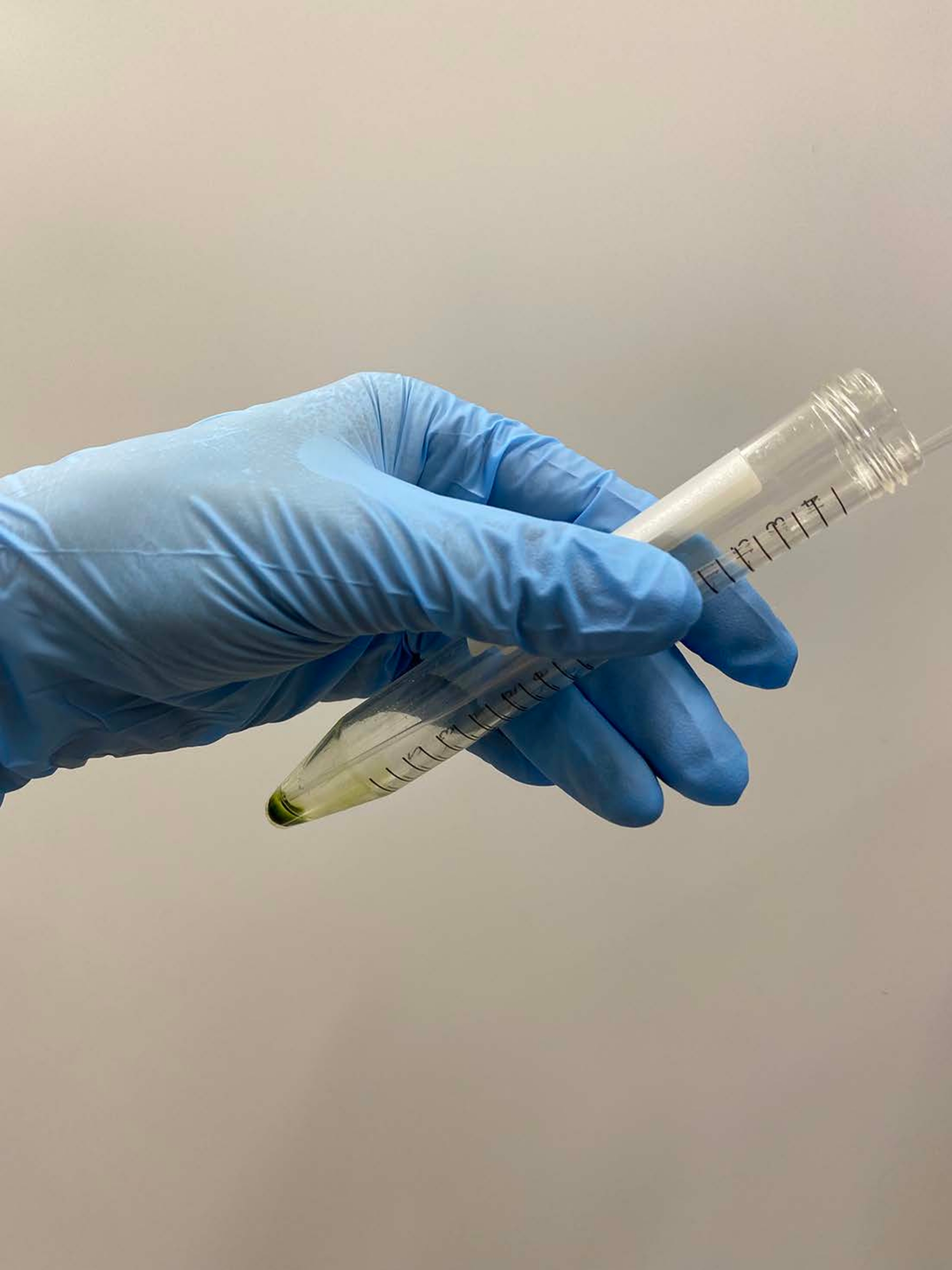
הערה: פיפטות פסטר עדיפות. בעת שימוש במיקרופיפטות, הימנעו משימוש בקצות פיפטה שעברו אוטוקלאב. - שכבה של כ-0.5 מ"ל של חיץ דה-ממברנציה על כדור תא, מנענעים את הצינור בעדינות ביד כדי להשעות בערך את התאים במאגר, ומניחים את הצינור על הקרח (איור 3A).
הערה: אין צורך להשעות לחלוטין את הכדור ברגע זה. העלו את הריכוז של MgSO4 עד 15 mM במאגרי הדה-ממברנציה וההפעלה מחדש כאשר מודלים של תאים מופעלים מחדש, עם ריכוז ATP סופי >1 mM להפעלה מחדש יציבה17. - השעו את הכדור הנותר של התא בעדינות באמצעות פיפטה, והניחו שוב את הצינור על הקרח (איור 3B).
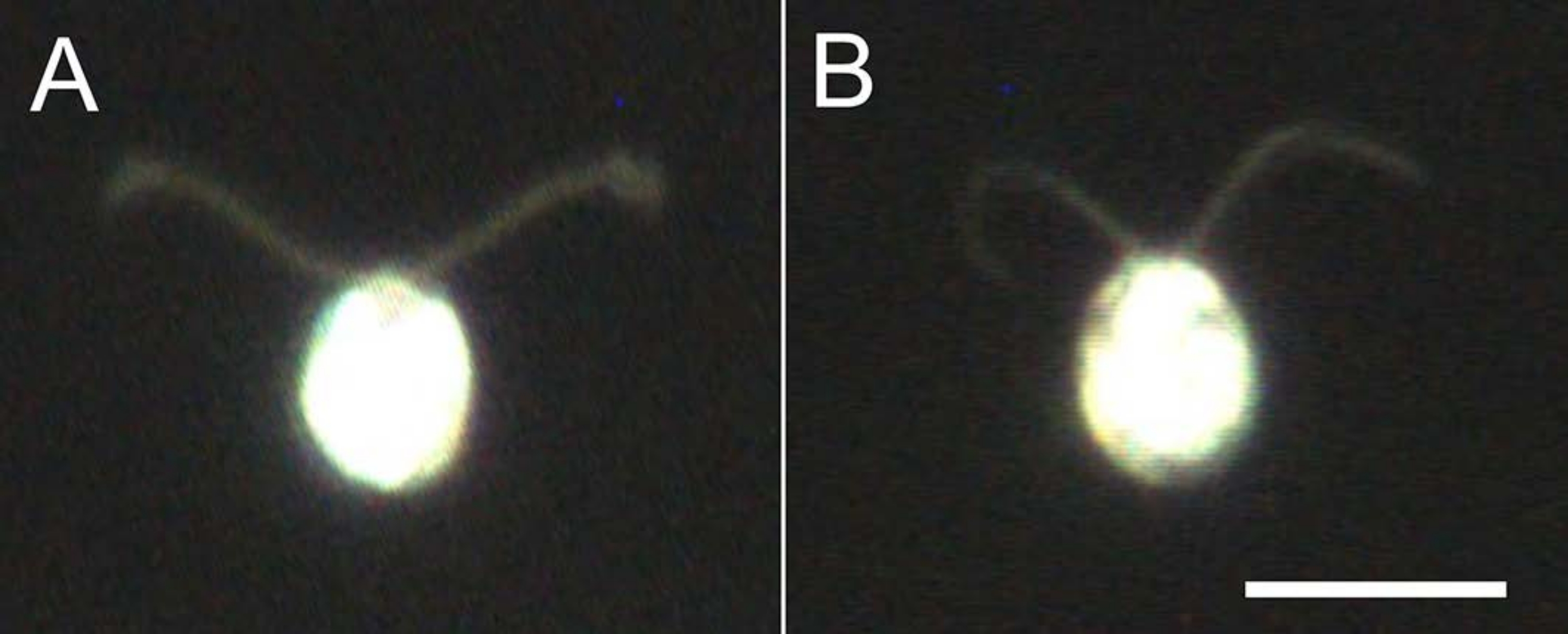
- קחו 5-10 מיקרול' מהמודלים של התאים, דיללו פי 10 עם מאגר הדילול, והתבוננו מתחת למיקרוסקופ (סרט 2) כדי לוודא שכל המודלים של התאים מתפרקים ולא שוחים (איור 4).
הערה: אם תאים מסוימים עדיין שוחים (בחיים), הוסף את חומר הניקוי הלא-יוני המשמש במאגר הדה-ממברנציה ישירות לתמיסת מודל התא לריכוז הסופי של כ-0.15%. לחלופין, חזור על שלבים 2.1-2.5 כאשר תמיסת הדה-ממברנציה מכילה 0.15% של חומר ניקוי.

איור 2: השלכת הסופרנאטנט. שאר הסופרנטנט הוסר בזהירות עם פיפטת פסטר לאחר שהסופרנטנט הוסר על ידי דקאנטציה של הצינור. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
{kind=link}

איור 3: Demembranation. (A) לאחר שכבה של 0.5 מ"ל של תמיסת דה-ממברנציה על גבי גלולת התא, התמיסה הייתה מעורבבת ביד כדי להשעות את התאים בערך. (B) לאחר הערבוב, שאר כדור התא הושעה לחלוטין בתמיסה על ידי פיפטה של פסטר. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
{kind=link}

איור 4: השפעות של דה-ממברנציה. (B) דגם תא תקוע על מגלשת זכוכית. שימו לב שבמודל התא, הריסונים נעשו מעט דקים יותר. התמונות נצפו תחת מיקרוסקופ עם עדשה אובייקטיבית פי 20 ומעבה שדה כהה טבילת שמן. סרגל קנה מידה = 10 μm. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
{kind=link}
סרט 2: אישור על דה-ממברנציה. תרחיף מודל התא נצפה תחת מיקרוסקופ עם עדשה אובייקטיבית פי 10 ומעבה שדה כהה טבילת שמן. אף תא לא שוחה. סרגל קנה מידה = 100 μm. אנא לחץ כאן כדי להוריד סרט זה.
3. הפעלה מחדש של מודלים של תאים שעברו דה-ממברנים
- ערבבו 80 μL של תמיסת הפעלה מחדש, 10 μL של תמיסת ATP ו-10 μL של מודלים תאיים בצינור של 0.5 מ"ל על-ידי הקשה על הצינור (איור 5).
הערה: ריכוז ה-ATP הסופי חייב להיות <3 mM מכיוון שתדירות הפעימה הציליארית, פרמטר לתנועה סילארית, עולה עם ריכוז ה-ATP ומרווית ב-2-3 mM של ATP17.
אזהרה: ערבוב על ידי צנרת או מערבולת גורם לדצילציה ומקטין את יעילות ההפעלה מחדש.- להפעלה מחדש עם <0.2 mM של ATP, הוסיפו מערכת התחדשות ATP, כגון 70 U/mL של קריאטין קינאז ו-5 mM של קריאטין פוספט (ראו טבלת חומרים).
הערה: תמיסת ההפעלה מחדש מוכנה בריכוז גבוה יותר (1.125x, טבלה משלימה 1) מאשר תמיסת הדילול כך שלאחר ערבוב ה-ATP המומס במים, התכולה מגיעה לריכוזים הסופיים הבאים: 30 mM של Hepes (pH 7.4), 5 mM של MgSO4, 1 mM של dithiothreitol (DTT), 1 mM של EGTA ו-50 mM של אשלגן אצטט (ראה טבלת חומרים).
- להפעלה מחדש עם <0.2 mM של ATP, הוסיפו מערכת התחדשות ATP, כגון 70 U/mL של קריאטין קינאז ו-5 mM של קריאטין פוספט (ראו טבלת חומרים).
- הניחו כ-30 μL של התמיסה המעורבת על מגלשת זכוכית והניחו עליה בעדינות מכסה עם ספייסרים כדי למנוע הלם מכני למודלים של התאים (איור 6).
הערה: ניתן להשתמש בסרטי נפט לבנים או סרטי דבק דו-צדדיים כדי לייצר את המרווחים. - התבונן במודלים של תאים שהופעלו מחדש תחת מיקרוסקופ (סרט 3).

איור 5: תמיסת ערבוב על ידי הקשה על הצינור. (A) עד 80 μL של תמיסת ההפעלה מחדש, 10 μL של תמיסת ATP ו-10 μL של מודלים תאיים נוספו בסדר רציף. (B) התמיסה הייתה מעורבת על ידי הקשה על הצינור באצבע. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
{kind=link}

איור 6: יצירת ספייסרים על קצוות כיסויים. (A) שכבה דקה של נפט לבן הוחלה על גב היד. (B) כמות קטנה של נפט לבן נשרטה עם קצה של מכסה. (C) ספייסר נוצר על קצהו של כיסוי. (D) ספייסר נוסף נוצר בקצה הנגדי. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
{kind=link}
סרט 3: שחייה של מודלים של תאים שהופעלו מחדש. התנועתיות של מודלי התאים הופעלה מחדש על ידי הוספת ATP בריכוז סופי של 1 mM, ונצפתה תחת מיקרוסקופ עם עדשה אובייקטיבית פי 10 ומעבה שדה כהה טבילת שמן. סרגל קנה מידה = 100 μm. אנא לחץ כאן כדי להוריד סרט זה.
תוצאות
תהליך הדה-ממברנציה וההפעלה מחדש בזן הבר C. reinhardtii (CC-125) מוצג כאן. התרבות יומיים לאחר החיסון הפכה לצבע ירוק בהיר (שלב 1.1) (איור 1). התאים נאספו (שלב 2.1), נשטפו (שלב 2.2) והושמדו (שלב 2.5). לאחר הדה-ממברנציה, כל המודלים התאיים הפכו לאימוטיליים (שלב 2.7). הריסונים המדולדלים (הנקראים אקס?...
Discussion
ישנם שני שלבים קריטיים בפרוטוקול זה. הראשון הוא תהליך המכונה demembranation, אשר צריך להתבצע בעדינות אך ביסודיות. דציליה (כלומר, התנתקות של ריסונים מגוף התא) נגרמת על ידי צנרת נמרצת או מערבולת, מה שהופך את מודלי התאים לבלתי משותקים גם לאחר הוספת ATP. בדרך כלל, 5 × 107 תאים מושעים בכ-0.5 מ"ל של מאגר ד?...
Disclosures
למחברים אין מה לחשוף.
Acknowledgements
מחקר זה נתמך על ידי מענקים של האגודה היפנית לקידום המדע KAKENHI (https://www.jsps.go.jp/english/index.html) ל- N.U. (19K23758, 21K06295) ו- K.W. (19H03242, 20K21420, 21H00420), מקרן המדע של Ohsumi Frontier Foundation (https://www.ofsf.or.jp/en/) ל- K.W., ומהברית הדינמית לחדשנות פתוחה המגשרת בין האדם, הסביבה והחומרים (http://alliance.tagen.tohoku.ac.jp/english/) ל- N.U. ו- K.W. אנו מודים לגב' מיוקי שינוהארה (Hosei Univ.) על עזרתה בהכנת הדמויות.
Materials
| Name | Company | Catalog Number | Comments |
| 0.5 mL plastic tube | QSP | 502-PLN-Q | |
| 15 mL conical tube | SARSTEDT | 62.554.502 | |
| Adenosine 5'-triphosphate disodium salt hydrate (ATP) | Sima-Aldrich | A2383 | |
| Centrifuge | KUBOTA | 2800 | |
| Chlamydomonas strain CC-125 | Chlamydomonas Resource Center | https://www.chlamycollection.org/ | |
| Creatine kinase | Merck | CK-RO | |
| Creatine phosphate | Merck | CRPHO-RO | |
| Dithiothreitol (DTT) | Nakalai tesque | 14128-46 | |
| GEDTA(EGTA) | Dojindo | G002 | |
| Hepes | Dojindo | GB70 | |
| Igepal CA-630 | Sigma-Aldrich | I8896 | IUPAC name is octylphenoxypolyethoxyethanol: IGEPAL CA-630 is a substitute for Nonidet P-40 (NP-40); NP-40 is no longer available in Sigma-Aldrich. |
| MgSO4-7H2O | Nakalai tesque | 21002-85 | |
| Microscope | Olympus | BX-53 | |
| Pasteur pipette | fisher scientific | 13-678-20C | |
| Polyethylene glycol, Mr 20,000 | Merck | 8.18897.1000 | |
| Pottasium acetate | Nakalai tesque | 28434-25 | |
| Sodium Hydroxide | Nacalai | 31511-05 | |
| Sucrose | FUJIFILM Wako Pure Chemical Corporation | 196-00015 |
References
- Szent-Gyorgyi, A. Free-energy relations and contraction of actomyosin. Biological Bulletin. 96 (2), 140-161 (1949).
- Hoffman-Berling, H. Adenosintriphosphat als betriebsstoff von zellbewegungen. Biochimica et Biophysica Acta. 14, 182-194 (1954).
- Naitoh, Y., Kaneko, H. Reactivated Triton-extracted models of Paramecium: modification of ciliary movement by calcium ions. Science. 176 (4034), 523-524 (1972).
- Witman, G. B., Plummer, J., Sander, G. Chlamydomonas flagellar mutants lacking radial spokes and central tubules. Structure, composition, and function of specific axonemal components. Journal of Cell Biology. 76 (3), 729-747 (1978).
- Rüffer, U., Nultsch, W. Flagellar coordination in Chlamydomonas cells held on micropipettes. Cell Motility and the Cytoskeleton. 41 (4), 297-307 (1998).
- Sale, W. S., Satir, P. Direction of active sliding of microtubules in Tetrahymena cilia. Proceedings of the National Academy of Sciences of the United States of America. 74 (5), 2045-2049 (1977).
- Fox, L. A., Sale, W. S. Direction of force generated by the inner row of dynein arms on flagellar microtubules. Journal of Cell Biology. 105 (4), 1781-1787 (1987).
- Kamiya, R., Yagi, T. Functional diversity of axonemal dyneins as assessed by in vitro and in vivo motility assays of Chlamydomonas mutants. Zoolog Science. 31 (10), 633-644 (2014).
- Kamiya, R., Witman, G. B. Submicromolar levels of calcium control the balance of beating between the two flagella in demembranated models of Chlamydomonas. Journal of Cell Biology. 98 (1), 97-107 (1984).
- Okita, N., Isogai, N., Hirono, M., Kamiya, R., Yoshimura, K. Phototactic activity in Chlamydomonas 'non-phototactic' mutants deficient in Ca2+-dependent control of flagellar dominance or in inner-arm dynein. Journal of Cell Science. 118, 529-537 (2005).
- Hyams, J. S., Borisy, G. G. Isolated flagellar apparatus of Chlamydomonas: characterization of forward swimming and alteration of waveform and reversal of motion by calcium ions in vitro. Journal of Cell Science. 33, 235-253 (1978).
- Fujiu, K., Nakayama, Y., Yanagisawa, A., Sokabe, M., Yoshimura, K. Chlamydomonas CAV2 encodes a voltage- dependent calcium channel required for the flagellar waveform conversion. Current Biology. 19 (2), 133-139 (2009).
- Bessen, M., Fay, R. B., Witman, G. B. Calcium control of waveform in isolated flagellar axonemes of Chlamydomonas. Journal of Cell Biology. 86 (2), 446-455 (1980).
- Wakabayashi, K., King, S. M. Modulation of Chlamydomonas reinhardtii flagellar motility by redox poise. Journal of Cell Biology. 173 (5), 743-754 (2006).
- Saegusa, Y., Yoshimura, K. cAMP controls the balance of the propulsive forces generated by the two flagella of Chlamydomonas. Cytoskeleton. 72 (8), 412-421 (2015).
- Gorman, D. S., Levine, R. P. Cytochrome f and plastocyanin: their sequence in the photosynthetic electron transport chain of Chlamydomonas reinhardi. Proceedings of the National Academy of Sciences of the United States of America. 54 (6), 1665-1669 (1965).
- Takano, W., Hisabori, T., Wakabayashi, K. Rapid estimation of cytosolic ATP concentration from the ciliary beating frequency in the green alga Chlamydomonas reinhardtii. Journal of Biological Chemistry. 296, 100156 (2021).
- Wakabayashi, K., Yagi, T., Kamiya, R. Ca2+-dependent waveform conversion in the flagellar axoneme of Chlamydomonas mutants lacking the central-pair/radial spoke system. Cell Motility and the Cytoskeleton. 38 (1), 22-28 (1997).
- Yueh, Y. G., Crain, R. C. Deflagellation of Chlamydomonas reinhardtii follows a rapid transitory accumulation of inositol 1,4,5-trisphosphate and requires Ca2+ entry. Journal of Cell Biology. 123 (4), 869-875 (1993).
- Wakabayashi, K., Ide, T., Kamiya, R. Calcium-dependent flagellar motility activation in Chlamydomonas reinhardtii in response to mechanical agitation. Cell Motility and the Cytoskeleton. 66 (9), 736-742 (2009).
- Ueki, N., Wakabayashi, K. Detergent-extracted Volvox model exhibits an anterior-posterior gradient in flagellar Ca2. Proceedings of the National Academy of Sciences of the United States of America. 115 (5), 1061-1068 (2018).
- Tanno, A., et al. The four-celled Volvocales green alga Tetrabaena socialis exhibits weak photobehavior and high-photoprotection ability. PLoS One. 16 (10), 0259138 (2021).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved