
Method Article
ゼブラフィッシュを用いた病的ヒトゲノムの in vivoモデルで
要約
ここでは、生理的に、関連する高感度で特異的な開発のための体系的なアプローチを提示ヒトの病理の変化を解釈するためのアッセイ。 WTおよび変異ヒトmRNAおよびモルホリノ(MO)アンチセンスオリゴヌクレオチドのマイクロインジェクション経由過渡遺伝子操作は、人間の発達障害との関連で急速にアッセイ病原特に変異が、排他的ではないが、開発にゼブラフィッシュ胚の扱いやすを活用。
要約
ここでは、ゼブラフィッシュにおけるin vivo相補での使用可能性の臨床的に重要な非同義変更を照会するためのアッセイの開発のための手法を提示する。これらの側面は、胚発生の分析において重要な進歩のために許可している胚は急速な発展を ex vivoで受ける、容易な閲覧を可能にするために透明であり、遺伝的に操作することができます1、;。彼らの実験的な扱いやすさのために有用な動物系であるゼブラフィッシュ( ゼブラフィッシュ )分子プロセス、および形態形成シグナル。まとめると、この脊椎動物モデルの利点は、小児疾患、およびいくつかのケースでは、成人発症性疾患で発達障害をモデルにゼブラフィッシュは非常に従順する。ゼブラフィッシュのゲノムは非常に人間(オーソロガス〜70%)のそれと保存されているので、ゼブラフィッシュ、ヒトの疾患状態を要約することができる。これは、変異ヒトMの注入のいずれかを介して行われます関数変異体の損失を模倣する遺伝子を抑制する機能対立遺伝子、又はモルホリノ(MO)アンチセンスオリゴヌクレオチドの使用率のドミナントネガティブ又は利得を誘導するためにRNA。かぶった人間のmRNAとMO-誘発表現型の相補性を通じ、我々のアプローチは、測定可能な、生理学的に関連する表現型を救出するための変異体mRNAの能力に基づいて、人間のタンパク質配列上の変異の有害な影響の解釈を可能にします。ヒトの疾患対立遺伝子のモデリング1-4細胞段階におけるMOおよび/またはヒトのmRNAとゼブラフィッシュ胚のマイクロインジェクションを介して行われ、7日後に受精(DPF)まで表現型。以下のプロトコルで示すように、この一般的な方法は、病気の表現型の広い範囲に拡張することができる。私たちは、形態形成シグナル、頭蓋顔面、心臓、血管の完全性、腎機能、および骨格筋疾患表現型だけでなく、他人のために我々の確立したモデルを提示する。
概要
遺伝子型の予測臨床的価値の遺伝情報と割り当ての機能的解釈は遺伝医学の主要な問題を表しており、ゲノムワイドなシーケンシングの加速技術的および経済的実現可能性とますます痛烈になってきている。したがって、患者で検出された未知の重大さの変種(VUS)の病原性をテストするための新しいパラダイムを開発し、実装することが必要である。これらのアッセイは、時間とコスト効率の高い、正確でなければならず、潜在的な港は臨床的有用性への移行を触媒する。
マウスは伝統的に人間の病気のモデリングの分野で最適なツールであったが、ゼブラフィッシュは、科学的、経済的に有利な代理として浮上している。マウスと違って、ゼブラフィッシュ生物学は発展途上病態のリアルタイムイメージングを可能にした胚の光学的透明性に助けられ、すべての発達段階に簡単かつタイムリーにアクセスすることができます。 1の比較的最近の世代は、機能的研究の多くで採用され、追加のテストおよびモデリングオプションを提供してきましたが、この技術は、(1,38で検討)が制限され続けています。達成する骨の折れる特定変異のノックインと遺伝的変異体であるだけでなく、彼らはまた、単一の遺伝子内の変異の範囲をテストするための中·高スループットの分析に適していません。重要なのは、テストのシングルスイートは遺伝様式を知らせるために重要である、対立遺伝子の病原性の可能性のためだけでなく、細胞レベル(関数の機能対利得の損失など )に効果の方向だけでなく情報を提供することができます家族で、特に小さな人間の家系は、遺伝的伝達のモードに限定された情報を抱い。利用可能なマウスおよびゼブラフィッシュモデルの用途のさらなる比較のため、 表1を参照されたい 。
また、あることに注意再ゼブラフィッシュモデルシステムに固有の限界があります。 D.もののゼブラフィッシュは、器官系の急速な初期の開発を持って、性的成熟は約3ヶ月が必要です。このため、出生前および小児期発症疾患は、この一過性発現モデルに最も適している。大化合物画面を行うための理想的つつ、遺伝子変異体の使用は、非同義に寄与する変異体は数千もの体系的試験のため現実的ではない、および小児疾患において検出され続ける。
ここで説明する相補性試験では、保存された発達過程のために必要な分子のために特にそう、この実験的な扱いやすさ、高度の相同性、そして人間とゼブラフィッシュのタンパク質間の機能の保全を活用する。 図1は 、様々な対立遺伝子の影響のためのテストと識別戦略を概説します。両方の機能喪失(LOF)と優性アッセイを行うことができる。 LOFは、実験はモルホリノノックダウンで目的の遺伝子の抑制から始まり、調査中の臨床表現型に関連するかもしれない表現型をアッセイする。通常の包含どちらを誘導、またはスプライスジャンクションにMOを置くことによって、スプライシングを阻害することによって、抑制がまたはその近くにゼブラフィッシュの遺伝子座の翻訳開始部位(tbMO翻訳ブロッカーモルホリノ)MOを標的とすることで翻訳をブロックすることにより、どちらか達成することができますイントロンまたはスキップ異常エクソン(スプライスブロッキングモルホリノ; sbMO)。
その後、オーソログ人間転写からかぶったmRNAが導入され、表現型の定量救助を測定する。アッセイが確立されると、人間のメッセージ内の候補突然変異が導入され、WTヒトmRNAのと同じ効率でMO-誘導性表現型を救出する能力についてアッセイすることができる。逆に、候補者のために支配的な対立、人間のmRNAが(ただし、MO)導入です支配的な影響を及ぼす試験変異の導入がヒト臨床状態で観察されたものと類似の表現型を誘発するのに対し、WT人間のmRNAが著しく、ゼブラフィッシュ解剖学、生理学に影響を与えないことを期待してED。この実験は、支配的な効果は関数のゲイン(GOF)またはWTおよび変異ヒトのmRNAを配合することによりドミナントネガティブメカニズムによって発生するかどうかを解剖するきめ細かいさらになることができます。GOFイベントの、WT人間のmRNAの追加があると予想される無関係、ドミナントネガティブ対立遺伝子のために、一方、WTおよび変異mRNAのブレンドしてミュータントメッセージによって誘導される表現型の重症度を変更する必要があります。 :すべてのケースでは、我々は注射のすべての組み合わせは、(変異ヒトのmRNAなど( 図1を参照)は、好ましくは胚の同じクラッチ内で、実行されるとモルホリノ対WT人間のmRNAとMO解釈以下のとおりであることをお勧めします
LOFテスト用:
- ノックダウンした場合変異体およびWTのmRNAによって同等に救出することができます表現型を生成し、対立遺伝子は、おそらく良性です。
- ノックダウンの表現型の変異救助ノックダウン表現型そのものと区別がつかない場合は、対立遺伝子は、おそらく機能nullです。実験では、真のヌル(無機能性タンパク質)と全く救助能力を持っていない超低タンパク質活性レベルを区別することはできません。
- ノックダウンの表現型の変異救助MOよりも統計的に良いですが、WTよりも悪い場合は、この結果は、関数の部分的な喪失を示しとして、対立遺伝子はおそらく低次形態です。
支配的なテストのために:
- そこにはノックダウンの表現型がありませんが、WT mRNAの注入は表現型を作り出す実験を続行する場合、コンティンジェンシー·プランは、(下記参照)を使用する必要があります。場合
- WT mRNAのないノックダウン表現型と注入がない場合はない表現型を生成しませんが、実験では、いつものように進行する。
- 注射した場合変異体mRNAの野生型mRNAのそれに相当し、対立遺伝子は、機能の良性または損失のどちらであってもよいし、アッセイは失敗した可能性がある。これは、さらなる実験がこれらのオプションを区別する必要があります。
- 変異体mRNAの注入がMOノックダウンと区別がつかない場合は、遺伝子産物の機能は、おそらく何らかの方法で変更される。機能の変化を識別する、野生型mRNAを有する変異体mRNAの滴定を行うべきである。
- この滴定の結果は、単独で、野生型mRNAに区別できない場合には、その効果が滴の量に応じて変化される変異タンパク質製品は、基質として野生型タンパク質を使用することが示されている。これは、ドミナントネガティブ表現型を示します。
- この滴定の結果は単独変異体mRNAに区別できない場合には、変異タンパク質産物がもはや野生型と同じ機能を持っていないことが示されているので、野生型タンパク質製品の事前の量によって影響されない送った。これは対立遺伝子が機能の可能性が高いゲインであることを示している。
コンティンジェンシー·プラン:
- ない表現型はMOのノックダウンから提示しませんが、WTのmRNAと共に存在しない場合、我々はこのような状況が理想的ではないことを強調するものの、さらなる実験が、発生する可能性があります。 WTヒトmRNAは、表現型を最小化するために滴定されるべきであり、また、新たな設定点として使用することができる。 WTおよび変異ヒトmRNAの更なる同時注入は変異体の救出能力に基づいて評価することができます。
プロトコル
1。バイオインフォマティクス解析
- 興味のある人間の遺伝子がゼブラフィッシュオルソログを持っているかどうかを判断し、そうだとすれば、どのように多くのコピー。我々は相互BLAST(お勧めhttp://blast.ncbi.nlm.nih.gov/ゼブラフィッシュゲノムに対するヒトのタンパク質)、およびヒトゲノムに対してヒットベストゼブラフィッシュのその後のBLASTを。真のオルソログは、各インスタンスで最高のヒットとなる。
- ヒト遺伝子のオープンリーディングフレーム(ORF)のサイズを決定する。 6キロバイトよりも長い場合、このモデルは長いため、テンプレートのインビトロ転写高品質の限界を現時点で難治性である。
- PCS2 +ベクター骨格(または5 'SP6転写サイトおよび3'ポリA信号と同等のベクトル)で人間のORFを含むコンストラクトを入手または生成します。
- ターゲットを絞ったゼブラフィッシュ遺伝子のスプライシングまたは翻訳をブロックするMOを設計します。複数のコピーは、ゼブラフィッシュのゲノム、熱に存在する場合電子つのオプションです:追加のMOの)設計は、aまたはb)完全コピーの間で保存スプライス部位の同定は、単一のMOは、有効であることができ、それに対して。一部公開MOシーケンスがでご利用いただけますwww.zfin.org 。
2。開発ゼブラフィッシュ胚における発現解析
- ゼブラフィッシュのオルソログが表現型の読み出しに関連時空文脈で表現されているかどうかを判断。全く発現データは、目的の遺伝子のために利用できない場合、全体のゼブラフィッシュの胚から、またはin situハイブリダイゼーションを用いてcDNA(RT)-PCR逆転写を行っています。 (2,3を参照)。
3。部位特異的突然変異誘発法
- 中央に所望の変異を持つ長さでデザイン変異導入プライマー25-45塩基、。プライマーの融解温度は、以上78℃以下でなければなりません反対にアニールする順方向および逆方向の突然変異誘発プライマーを設計プラスミドの鎖。
- ORFポスト突然変異生成のシーケンス確認用プライマーを入手し、これら全体のORFをカバーするために〜300塩基対のセクションを越えタイルべき。
- 次のように高忠実度ポリメラーゼとサイクルで変異誘発反応を組み立てる(1:95℃30秒、2:95℃30秒、3:55℃で1分、4:68℃で6分、5:行く2 18X、6:4℃永遠に、7:終了)。
- 2時間37℃でインキュベートし;ダムメチル化テンプレートを除去する反応あたり1μlのDpnIを制限エンドヌクレアーゼを追加します。
- 標準プロトコールに従って20μlのコンピテント細胞に突然変異誘発反応2μlのを変換します。
- 3-4コロニーを選び、適切な抗生物質を含むLB培地を5mlに接種。 225 rpmで37℃で一晩振とうする。
- ミニプレップは、DNAと濃度を決定。
- 配列変異部位は、関心のある突然変異の存在を確認する。
- シーケンスの整合性を確認するためのシーケンス全体ORF。
(4)in vitroでの mRNAの転写
- 線形化PCS2 +テンプレートを使用して、mMessage mMachine SP6キット(Ambion社)を使用して、かぶったmRNAを生成します。私たちは、反応成分量の半分を使用することをお勧めします。
- キットの説明書に記載されているようにLiClを沈殿またはフェノールクロロホルムでRNAサンプルを精製する。
- 吸光度を使用してmRNAの濃度を測定し、ゲル電気泳動を用いてmRNAの整合性を確保し、使用するまで-80℃で三つ以上のアリコートでサンプルを保存します。我々は、mRNAのアリコートの複数の凍結融解サイクルをお勧めしません。
5。関数のバリアントの損失のin vivoアッセイ
- 天然のゼブラフィッシュの交配からゼブラフィッシュ胚を取得し、28でそれらを維持°6または10センチ皿に胚水中のC。
- 表現型特異性、MO効率及びMOの毒性を評価するために、モルホリノ用量応答曲線を行う。での用量曲線を注入する少なくとも50〜100に1-10 ngのMO間の3つの異なる濃度(1-4細胞期)ゼブラフィッシュの胚/バッチ。効率的なMOSがバッチで影響を受けた胚の割合の用量依存的増加を生じさせるべきである。
- 適切な発達段階での表現型の胚は、金利、関連表現型が観察された段階の目標とゼブラフィッシュの遺伝子の発現に基づいて。これは、どちらか(標準化された表現型の基準に基づいて)定量(例えば解剖学的構造との間に測定など)、または定性的かもしれません。すべての注射用> 24 HPFを獲得:胚はメラニン形成の最大の削減に24 HPFでPTU(胚のメディアで0.003%1 - フェニル-2 - チオ尿素)で処理されるべきである。
- 表現型のスコアの時点で全胚溶解物からトータルRNAを抽出しスプライスブロッキングMOは、テストMO効率のために、cDNAを生成し、MOの標的部位に隣接するプライマーを用いて標的遺伝子のRT-PCRを行う。
- 抑制efficを確認するにはTB-MO、収穫全胚タンパク質溶解物と標的タンパク質に対する制御のレベルを比較するためにイムノ行動iency。ゼブラフィッシュタンパク質に対する抗体が多くの商業用の限られた交差反応性があるので、このアプローチは、すべての標的遺伝子に従順ではない。 MO特異性を示す二つの間接的な方法には、次のとおり)表現型に対する用量依存効果があることを示すと、b)TB-MOを有する野生型mRNAの同時注入が効率的に表現型を救出することを示す。可能な場合に効率が直接モニタすることができるので、これらの理由から、我々は、スプライスブロッカーを推薦する。
- 表現型は5.7に進みますが観察されている場合はない表現型が観察されていない場合は、6.1に進みます。
- 定性的な表現型の場合、胚50〜75%が影響を受けているMO用量を選択し、定量的な表現型のために、表現型の指標が(P <0.001)、野生型とは大きく異なっているのMo量を選択します。ゼブラフィッシュEMBの新しいバッチを注入するMOの "検定"線量と人間WT mRNAの(10-200 PGのmRNAからまでの投与量曲線を含むカクテル; ryos(N = 50-100/batch 1-4細胞期)、これらの用量は、上記の実質的な過剰発現を確保ゼブラフィッシュ胚における総ポリA mRNAの0.25から0.5パーセントを表す任意の単一の転写産物のベースライン)。4
- 注入バッチのマスクされた得点を行って、MO、単独と比較して最も重要な救助とWTのmRNA量を選ぶ、これはmRNAの "検定"用量である。
- MOの検定量と変異ヒトmRNAのアッセイ用量で、新しいバッチを(N = 50-100/batch 1-4細胞期)を注入。適切な段階での表現型の胚と適切な統計的検定(t検定やカイ二乗)を使用して、WT人間のmRNAの救助に結果を比較。成果については、図1を参照し、手順7に進みます。 mRNAの毒性の影響を制御するだけではWTと変異mRNAのアッセイ線量を注入します。
6。バリアントドムのin vivoアッセイ機能効果の負inantまたはゲイン
- 機能表現型の損失が認められなかった(ステップ5.5)または変異mRNAがMO単独(ステップ5.7)と有意に異ならない表現型を生じさせる場合には、10から200 pgのWT人間のmRNAから至る用量曲線を注入されている場合は(私たちは25、50を推奨50〜100胚/バッチ)、初期テストとして100 pgの)胚バッチ(1-4細胞期に。
- 適切な段階では(上記の5.3を参照)、表現型の採点を行い、非注入対照と比較して死んでいる、および/または影響を受けた胚の統計的に有意な数が存在しないとなるで最高用量を決定する。これは "アッセイ"用量である。
- 変異ヒトmRNAのアッセイ量(; 50〜100胚/バッチ1-4細胞期)を注入する。表現型胚およびWT人間のmRNAの注射またはアッセイMO濃度の測定線量から得点に結果を比較します。成果については、図1を参照してください。
- 結果がMOと区別できない場合には、WT mRNAと人間の突然変異体のmRNAを滴と人間のWTと変異mRNAの注射と比較します。変異プラスWT mRNAを注入バッチと表現型の改善はドミナントネガティブを示します。改善は関数のゲインがないことを示します。
7。 in vivoでのテスト結果に再現
- in vivo試験で三回の最小値を繰り返します。
8。エビデンスの他のラインを用いたin vivo病原性データにゼブラフィッシュを統合
- in vitroでの研究では遺伝的血統内のデータ(該当する場合)、対照集団周波数データ、(細胞ベースのタンパク質の安定性のアッセイ、細胞内局在、シグナル出力、または酵素活性)にゼブラフィッシュの実験から得られた病原性のデータを比較してください。
結果
劣性とPseudorecessive障害
一次繊毛は、増殖、極性、分化、組織の維持を含め、複数の細胞運命の細胞シグナルの役割を果たしている脊椎動物のボディプランに近いユビキタス構造です。これらのオルガネラの5機能障害は、人間の遺伝性疾患の広い範囲につながると総称ciliopathies。6,7つのそのような臨床実体はバーデット-ビードル症候群(BBS)、網膜変性、肥満、性腺機能低下症、多指症、および腎機能障害によって特徴づけ多臓器小児疾患である。7アレル病原性のin vivoアッセイの開発が必要であったBBSので)、それは、少なくとも17の遺伝子7-10で発生主として民間非同義変化によって引き起こされる遺伝的異質性疾患であり、BBSファミリーの> 25%のB)oligogenic継承、第二のヘテロ接合体の変化の存在を特徴とBBS遺伝子(劣性プライマリー因果変異に加えて)臨床浸透度と表現力を調節することができる。通常、このような第三の対立は、このようにタンパク質の機能に及ぼす生物学的効果の正確な解釈を必要とする遺伝的観点、不明確病原電位からの非同義ヘテロ接合変化、である。11-13
アルマ - BBSにおける突然変異負荷に貢献する変異の病原性の可能性を調べるために、我々は、最初はBBS1で識別されるすべてのミスセンスの変更をテストした。我々と他の人の両方が基礎体温タンパク質の損失は平面内細胞極性の調節不全(PCP、非カノニカルWntシグナル伝達)を生じさせることが示されている。半ば体節ゼブラフィッシュ胚における収束拡張欠陥として顕在14この生理学的に関連する表現型の読み出しを使用して、我BBS遺伝子の抑制を短縮体軸、広く、薄く体節、広帯域、よじれnotochordsをもたらすことを見出した。15 <にSUP />(図2)MO-による抑制は、原腸形成の欠陥を生産、およびin vivoの方法で別の 3で得点に人間のmRNAと同時注射(と再現)著しくこれらの表現型を救出した。まず、胚は質的表現型基準(ノーマル、クラスIおよびクラスII、表現型のクラスの詳細な定義については、15を参照)によると、ライブスコア化した。次に、遊走細胞を可視化するフルオレセイントレーサーを用いることにより、被包中の細胞遊走を(卵黄セル16の上にセル層の薄層化と拡散することを特徴と早期発達段階)定量。最後に、我々は平らな形態素解析のためにマウントされたkrox20、PAX2とMyoDのリボプローブのカクテル、とハイブリダイズin situで九体節胚において体節トランクの長さを測定した。
この方法は、毛様体変異空間における対立遺伝子の過剰500でテストするために使用されている。いずれかで一人で勉強し、> 130対立遺伝子のin vivo試験では表現型のスコアの範囲を生産し、完全な、良性に分類された(図1)我々のプロトコルによって救出示さ(WT救助大きく異なっていない)、部分的には、(大幅に改善hypomorphsとして分類された救助)WT救助よりMOからではなく、より深刻な、救助に失敗は、機能ヌル(MO大きく異なっていない)、そして支配的なネガに分類されたMOに比べて単独の変異体mRNAによって誘発表現型として分類された。
また、ゼブラフィッシュにおけるin vivo相補アッセイの感度および特異性を評価した。特異性は、共注入共通のSNP(健康な対照集団における> 5%のマイナーアレル頻度)により確認したが、これらを14/17(> 82%)を試験に良性の表現型を与えることが見出され、示されるように、感度は98%であることが示された生体データと遺伝引数十分T 中の間に一致によりoはBBS血統で因果として対立遺伝子を属性17に加えて、表現型効果がイムノと細胞内局在研究を用いたin vivoの措置(ライブスコア、被包トラッキング、ISH形態計測)in vitroで検証された3つを用いて観察。これらの結果の解釈は病気の病因の少なくとも一つのメカニズム予備知識を必要としますが、この例では、我々のプロトコルの有用性と堅牢性を立証するための証拠を提供しています。我々は公平な突然変異スクリーニングとTTC21B、逆行性鞭毛内輸送タンパク質の機能解析研究における実験的証拠の複数の他のラインとの生体内のスコアを裏付け以来持っている。18
支配的な障害
肢帯筋ジストロフィー(LGMD)は腰と肩が遅い進行筋力低下を引き起こし、筋ジストロフィーの常染色体クラスである。この遺伝的にと私chanistically疾患の異質グループは、複数の、筋細胞膜サルコメア、細胞質、核タンパク質全体で優性と劣性変異の両方によって引き起こされる。臨床表現型と磁気共鳴画像から筋肉の関与の証拠の提示に基づいて、我々は、フィンランド、米国、イタリアの家族19で見つかった支配LGMDの原因を調査した。マッピングされた遺伝子座内の位置候補の配列がDNAJB6の変異を明らかにし、HSP70ファミリーの共同シャペロンをコードする遺伝子は、ヒトの少なくとも二つのスプライスアイソフォーム(核および細胞質)として表現。 DNAJB6機能とLGMDとの関連性にさらなる洞察を得るために、我々は、ゼブラフィッシュの筋肉の整合性におけるその役割を検討した。ゼブラフィッシュオルソログ(dnajb6b)のRT-PCRは、スプライスブロッキングモルホリノと胚の注入が続いた五体節期、早けれ式を検出しました。 48時間後の受精では、マスクされたスコアは、低速の剥離が認められた彼らの挿入ポイントからの繊維。この表現の特異性は、その後、第二の非重複MOでテストされ、WT人間DNAJB6 mRNAとその後救助された。
DNAJB6機能の喪失が筋肉の整合性の欠陥につながるどのように照会するには、我々は両方のアイソフォームのヒト転写に患者に見られるミスセンス変異を導入し、ゼブラフィッシュ胚にそれらを注入した。 WT人間のmRNAの注射は目立った表現型を生成しませんが、これらの変化は、細胞質に設計MOの機能効果の損失をphenocopiedが、核ではないアイソフォーム。これは、支配的な影響を示唆して、筋肉の表現型の強化された重大度を示し、変異体およびWT mRNAを、等モル量の同時注入が続いた。この概念をテストするために、さらなる注射は変異体およびWT mRNAのモル比を変えることにより行なった。予測と一致して、変異体mRNAの過剰は、胚におけるWT誘導し致死率と比較しながらWTのn過剰は次第に増加して救助を生産。これは、oligimerization特性および可能なタンパク質相互作用を決定するインビトロ実験を行った。 LOF BAG3での小児科のフォームを引き起こすので、これらは、変異が細胞質DNAJB6のantiaggregation活動を損なうともLGMDの病理学的機序に関連する変異体およびWT両方の売上高だけでなく、他の分子と相互作用し、BAG3、に干渉することを示した筋ジストロフィー20。我々はその後BAG3がゼブラフィッシュにおける変異DNAJB6bによって誘発表現型を調節することができるかどうか尋ねました。 WT BAG3の注入だけではなく表現型を生成しませんが、DNAJB6変異との混注を大幅BAG3は、このような変異体19の病原を媒介する役割を果たしていることを示唆し、表現型の重症度を増加させた。
| 過渡ゼブラフィッシュモデル | 変異ゼブラフィッシュライン | トランスジェニックマウスライン | |
| 調査中の人間の表現型の発症年齢 |
|
|
|
| 実現可能な表現型 |
|
|
|
| 使用するまでの時間 (誕生から) | 1-7日 | > 3ヶ月 | > 6ヶ月 |
| スループット | 中高 | 低い | 低い |
| メリット |
|
|
|
表1。 in vivoモデルの比較 。

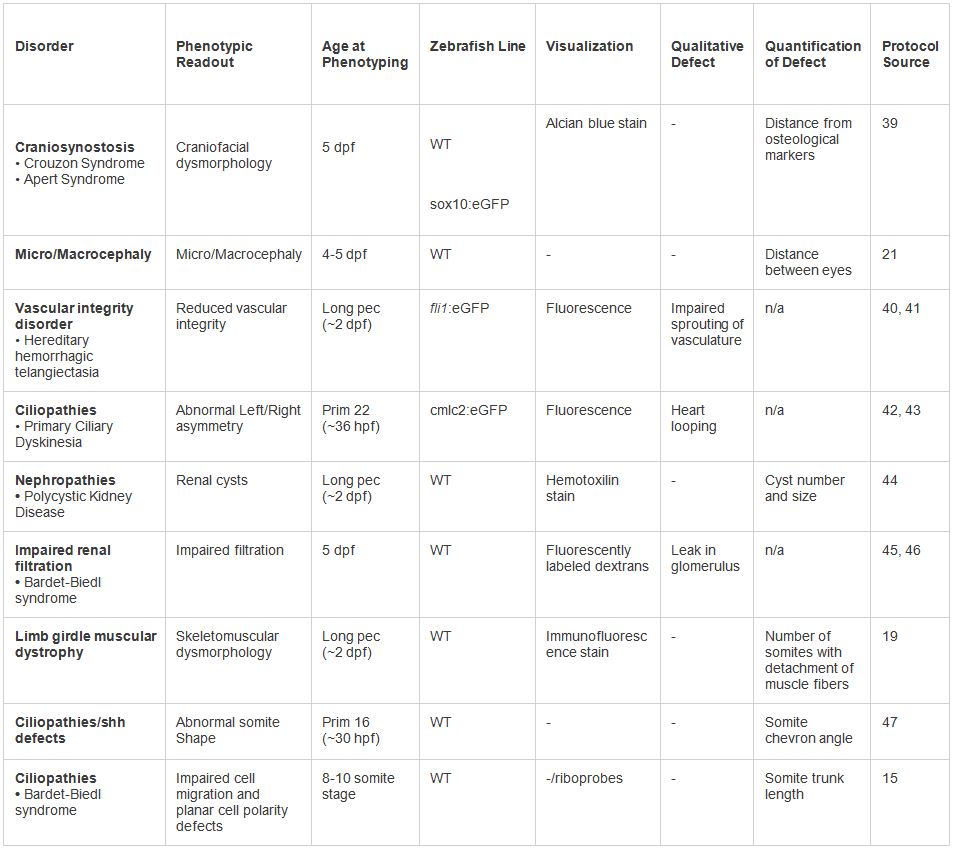
表2。人間dysmorphologiesのin vivoモデルでの例。様々な表現型は、提示されたプロトコルの下でテスト。表現型の読み出しと可視化技術の範囲は、障害の種類に基づいて採用することができる。 大きいテーブルを表示するには、ここをクリックしてください。
{kind=link}

図1非同義変異型のin vivo機能テストで 。すべての機能テストへの体系的なアプローチ不明または仮説意義eles。モルホリノマイクロインジェクション経由遺伝子ノックダウンは、WTおよび変異ヒトmRNAの両方の(共)注射のシリーズが続いている。表現型の結果の統計的分析は、対立遺伝子の病原性と分子機能に通知します。簡単に言えば、 機能検査の損失:ノックダウン変異体およびWTのmRNAによって同等に救出することができた表現型を生成する場合、対立遺伝子は(緑のボックス)おそらく良性です。ノックダウンの表現型の変異救助ノックダウン表現型と区別がつかない場合は、対立遺伝子は、おそらく機能ヌル(黄色の箱)です。ノックダウンの表現型の変異救助MOよりも統計的に良いですが、WTよりも悪い場合は、対立遺伝子は、おそらく低次形態(緑のボックス)が支配的な試験のために:。変異mRNAの注入は、野生型mRNAのと同等である場合、対立遺伝子は、良性または機能の喪失のいずれであってもよい、またはアッセイは(緑のボックス)が失敗した可能性があります。変異体mRNAの注入はindisである場合 MOノックダウンからtinguishable、遺伝子産物の機能は、おそらく何らかの方法で変更される。機能の変化を識別する、野生型mRNAと変異型のmRNAを滴定する。この滴定の結果は、単独で、野生型mRNAに区別できない場合には、変異タンパク質産物は、それによってドミナントネガティブの形質を(青箱)を示す、基質として野生型タンパク質を使用する。この滴定の結果は単独変異体mRNAに区別できない場合には、変異タンパク質産物は、もはや、野生型と同じ機能を持っていないので、おそらく機能(青箱)の利得である。まだ表現型はMOノックダウンから提示ないが、WT mRNAを、本行い 、さらなる実験が発生することができれば、WTヒトmRNAは、表現型を最小化するために滴定されるべきであり、また、新たな設定点として使用することができる。 WTおよび変異ヒトmRNAの更なる同時注入は変異(ピンク箱)の救出能力に基づいて評価することができます。ら= "_blank">より大きい数字を表示するには、ここをクリックしてください。

図2。ヒトで検出さMKS1変異の定量的および定性的評価。mks1 morphant胚の発達の欠陥。重大度に基づいて、表現型は3つのグループに分類した。各クラスの例としては、()が示されており、その胚集団内での有病率(nは= 100-160胚)(図示せず)を集計した。クラスとMO注入胚は私の表現型は肉眼正常な形態を持っていたが、同じ体節年齢(8-9体節)で注入胚を対照と比較して卵黄に過度の胚組織で、短かった。クラスII morphantsは、薄く短く、不十分頭と尾の構造を開発し、さらに、体節の定義を欠いていたと対称性。クラスIII胚は厳しく、未発達と不格好な体節で遅れnotochordsを波状、通常は10体節の段階を過ぎて生存しなかった。人間MKS1 mRNAの同時注射はmks1抑制に表現型の特異性を実証し、これらの欠陥のそれぞれを救出。krox20、PAX2とMyoDのリボプローブで染色し、11体節段階(±1体節)での胚のin situハイブリダイゼーション(A、B、C )。表現型は、Cで定量最初から各胚の最後のかなりの体節(矢印)に測定することによって定量した。図15から許可を得て適応した。

図3。人間の異形学のin vivoモデルでの例。 (a)は顔面異形学。 mRNAを注入された胚(左)と5 DPFでアルシアンブルーで染色した変異体、注入胚(右)を制御します。変異mRNAを注入胚は鰓弓と欠落しているか、不正な形式の構造をスプレイ含む軟骨頭蓋顔面骨格の一般的な解体と、特に小さいと不格好な頭を表示します(B)マイクロ/大頭。制御は5 DPFで胚(左)とkctd13 MO注入胚(右)を非注入。 Morphants目の間のスペースで見られるように、頭の拡大表示します21(c)は、血管の完全性を減少させた。コントロールは、胚を非注入(上)とENG MO注入胚(下)FLI1 2 DPFで蛍光顕微鏡を用いて画像化:のeGFPトランスジェニックレポーターライン。節間血管や他の血管構造の出芽障害Morphantsが表示。41(d)に改変された心臓のループを。ユニのin situハイブリダイゼーションccdc39 morphant胚(図示せず)が二国間の示した(右)または、ほとんどの場合、検出不可能なSPAW式ながらnjected野生型胚(左)は、左側の側板中胚葉におけるSPAWの発現を示す図43(e)の腎嚢胞が。非注入WT胚(上)とift80 morphant(下)。 Morphantsは大きな腎嚢胞(矢印)、心膜水腫(矢じり)とカールテールを表示しました。44(f)に削減糸球体ろ過。制御注入胚(上)とift80 morphant(下)心にローダミンデキストランの注入後24時間の蛍光可視化。血管系全体に蛍光分散し、コントロールの蛍光の完全な欠如が見られるようにほぼ完全に腎臓によって排出される。 morphantは減少、糸球体濾過を示唆している永続的な蛍光デキストランが表示されます46(g)の筋ジストロフィー。 WT DNAJB6はmRNA注入された胚は(上)抗遅いミオシン抗体を用いた免疫染色によって決定されるように隣接myosepta間で正常に体節にまたがる正常遅い筋線維を示す。ミュータントDNAJBb(下)は、1つまたは複数の体節にmyoseptaから筋繊維の剥離を完了する部分を示した図19(h)に体節角度。コントロール非注入(上)または30 HPFで撮像kif7 morphants(下)のライブ横ビューを拡大。 Morphantsは、ゼブラフィッシュの筋節内シグナリング異所ヘッジホッグに帰属異常形の体節を、表示します47
すべての数字は、許可を得て適応。
ディスカッション
方法は、ヒト遺伝病の表現型( 表2、図3)の様に関連付けられた非同義変化の測定に適用できる一般的なプロトコルを表し、ここで説明。我々のアプローチは、疾患の表現型上の変化の潜在的影響を評価するために有用であることが証明されており、病気のメカニズム(例えばバーデット-ビードル症候群へのドミナントネガティブ変異の寄与、主に常染色体劣性疾患17など)を詳細に分析を支援する。現在までに、提示された決定木の開発を通じて、我々は1,000対立遺伝子の過剰に、因果遺伝性疾患に関連付けられている200の遺伝子の過剰に合理的なコストと時間でモデル化しました。
ここでは詳細に説明されていないが、我々はまた、これらの方法には、例えばコピー数変異体(CNVsの)並びにと遺伝的相互作用などの遺伝的病変の他のタイプをモデル化することが適切であることが示されている。そのような事象の分析は、の範囲を超えてい本発明の方法の説明、彼らは根本的に臨床的に適切な表現型の誘発または増悪を決定するために、候補遺伝子の系統的なテスト(同時に注入された遺伝子のペアを含む)と同じ原理に依存しているものの。例えば、16p11.2 CNVにおける29の遺伝子のどれを解明することは注入し、頭部たセグメント内の29の遺伝子の各々に対応する観測660キロバイトゲノムセグメントの重複を持つ患者で観察された小頭症、mRNAに関連するかもしれないサイズ測定は、単一の転写、KCTD13の主要な貢献を明らかにする、2 DPFと5 DPFで行った。さらに21、我々はバーデット-ビードル症候群とヒルシュスプルング病の両方を持つ患者におけるゲノム病変のアッセイ遺伝的相互作用には、このモデルを使用しています22別々にかつ同時に2つの臨床アイデンティティの原因遺伝子の抑制MOの比較によって、我々は、バイとして得られた表現型を識別することができたngの相乗的相互作用ではなく、単に添加剤の重大。
ciliopathies 17に貢献変種用の高感度(98%)および特異性(> 82%)を設立したことにもかかわらず、我々はまだこれらがゼブラフィッシュモデル内のすべての表現型の読み出しに一般化されているかどうかを判断するための十分なデータがありません。そのためには、対立遺伝子の多くは、それぞれの表現型のカテゴリ内でテストする必要があり、遺伝的に良性または病原性のどちらかになると予測。これはVUSsの機能的解釈は偽陽性と偽陰性の堅牢な理解は医師と患者へのそのような結果の配信を伴うことができる場合にのみ診断と管理に知らせることができ、ここで、臨床の現場でこのようなアッセイの実施のために特に重要になります。それにもかかわらず、これらの方法は、ヒト遺伝病の風景のより良い理解に向けて大きく貢献することができる。私たちは、これらのモデルは唯一の財団としての役割を果たすことはありませんと予想臨床遺伝情報の改善された解釈のためdationなく、また治療の画面を実施する有用なモデルとして採用されます。vivoでのデータではまた、PolyPhen 23などのソースからインシリコ計算予測に比較することができる、24 SIFT、のSNP&GO 25、または26 MutPred一致を表示します。予測データベースのSNP&GOとMutPred以前の研究では、それぞれ、精度だけで0.82と0.81に達すると、最も正確であることが判明したことに注意してください。27
我々は小児解剖学的欠陥( 表2、図3)のサブセットのこれらの方法のロバスト性を概説してきたが、特定の表現型は、これらの方法による少なく扱いである。いくつかの例外にもかかわらず、我々のプロトコルに従順ではない疾患の3つの主なクラスがあります。成人発症性疾患(例えば、パーキンソン病など)胚システムのモデルへの挑戦を表す。スローPROGression変性表現型(例えば、前頭側頭型認知症のような)表現型を生成するためにMO活動の7 DPFウィンドウよりも多くの時間を必要とするかもしれません。 RNAiのおよびsiRNAなどの他の遺伝子ノックダウン技術は妨害または遺伝子標的を分解することが可能であるが、未ひいては期間を制限する、MOの28のように、特定の安定した、非毒性、又は長期持続しないことが示されている表現型解析のために。第三に、このような哺乳類の肺のようないくつかの脊椎動物の構造は、ゼブラフィッシュ胚において十分オーソログ構造を持っていません。我々は、これは珍しい、望ましくない状況で注意にもかかわらず我々はまた、WT人間のmRNA注入が表現型につながるでこれらの事件の捜査のための提案されたコンティンジェンシー·プランを提供してきた。
特定の病気の表現型は、その後抽象と代理出産の大きい程度を必要とするかもしれません。これは、遺伝子の機能が十分にモデルとTRの間に表現型の類似性を弱めるために分岐している可能性がありますUEの表現型、またはそのゼブラフィッシュ生理は本来誘発疾患の影響を複雑にします。このような場合には、事前に生成さ却下表現型のさらなる切開を示唆している。私たちは、このアッセイのために問題のある表現型はゼブラフィッシュ胚でモデル化されている、いくつかの成功例を生み出してきた。例えば、TCF8の突然変異、フックス角膜ジストロフィー(FCD)に関連付けられている遺伝子は、開発の初期段階では、この転写産物の既知の役割に基づいて、代理の表現型読み出しとして原腸形成の欠陥を利用して我々のプロトコルを使用して測定した。他の例では29、そのようなDNAJB6で突然変異によって引き起こさ成人発症型筋ジストロフィーとして、私たちは、人間が生活の最初の三から四十年の間にかなりの筋肉の病理を失っているという事実にもかかわらず、5dpf胚における筋線維表現型を生成することができました。19
ここで紹介する過渡変異モデルに加えて、他の人もアドバンタをとっている身体の様々なシステムにヒトの疾患をモデル化するために、この一時的なシステムのGE。一例では、網膜色素変性症、網膜細胞死をもたらす、遺伝子RP2のノックダウンにより、ゼブラフィッシュでモデル化および網膜ラミネーションを減少させた。 5変異mRNAのうち4人は救出に失敗しながら、ヒト野生型mRNAを使っての救済は、網膜のラミネーションのすべての3つの層の発展をもたらした。30人間の感覚障害のこのモデルは、形態学的表現型に基づいていますが、それはまたすることが可能ですこのような音響驚愕やプレパルス抑制などの刺激に対するアッセイ応答47
最近、ゼブラフィッシュモデルは、アミロイド前駆体タンパク質を介してアルツハイマー病の病因を研究するために使用された。31著者らは、遺伝子ノックダウンは、ヒトmRNAと救出することができる運動ニューロンの軸索伸長を損なう原因と示した。このモデルでは、マウスモデルだけの微妙なphenotを表示するように、特に有益であることが証明されているYPES(シングルノックダウン)、または出生後致死(ダブルノックダウン)。開発を通じてインビボでゼブラフィッシュ胚を評価する能力だけでなく、タンパク質が適切に機能するために細胞外および細胞内ドメインの両方を必要とすることの直接的な証拠を提供して、減圧アミロイド前駆体タンパク質の病原効果を識別する助けた。その他の注目すべきモデルが含まれ、追加の筋ジストロフィー32、ダイヤモンドブラックファン貧血33、アクセンフェルト-レイガー症候群(眼及び顔面開発)34、炎症性腸疾患35(抗菌活性)、パーキンソン病36(ニューロンと運動の損失)、および発作37のこと(水頭症と多動)。
より一般的には、ヒトの疾患の表現型を要約することも示されている変異体ゼブラフィッシュラインである。 1,38で検討、モデルは白血病、メラノーマ、拡張型心筋症、デュシェンヌ型筋ジストロフィーを含むおよび多くの他。
開示事項
著者は、彼らが競合する経済的利益を持っていないことを宣言します。
謝辞
私たちは、デューク大学ディーンのサマーリサーチフェローシップ()、米国心臓協会(AHA)の交わり11POST7160006(CG)、国立衛生研究所(NIH)から国立眼研究所(EED)、R01HD04260からの補助金R01-EY021872から支援を認める国立成育医療協会(NK)、糖尿病消化器腎疾患研究所(NK)からR01DK072301とR01DK075972、および欧州連合(GA NR下EU 第 7 回 FPによる資金援助を受けて241955、プロジェクトSYSCILIA; EED、 NK)NKは識別ジャンとジョージ·W·Brumley教授。
資料
| Name | Company | Catalog Number | Comments |
| Reagent | |||
| Phusion High-Fidelity DNA Polymerase | NEB | M0530S, M0530L | |
| DpnI restriction endonuclease | NEB | R0176L, R0176S | |
| Max Efficiency DH5α competent cells | Invitrogen | 18258-012 | |
| Big Dye Terminator | Applied Biosystems | 4337455 | |
| mMESSAGE mMACHINE Kit | Invitrogen | AM1340, AM1344, AM1348 | |
| Morpholino | Gene-Tools | n/a | |
| 1-phenyl-2-thiourea (PTU) | Sigma Aldrich | P7629 | Prepare as 0.003% PTU in embryo media |
| Paraformaldehyde (PFA) | Sigma Aldrich | P6148 | For embryos that must be fixed prior to phenotyping, prepare as 4% |
| Tricaine methane sulfonate | Western Chemical | N/A | For anesthetization and euthanasia |
| Equipment | |||
| PTC-225 Tetrad Thermal Cycler | BioRad | Any equivalent thermal cycler | |
| Nano Drop 2000 spectrophotometer | Thermo Scientific | ||
| SMZ 745T Stereomicroscope | Nikon | ||
| AZ100 Stereomicroscope | Nikon | ||
| DS Fi1 Digital Camera | Nikon | For color/fluorescent imaging | |
| DS QiMC Digital Camera | Nikon | For black/white imaging | |
| Advanced Resarch 3.2 Imaging Software | NIS- Elements | ||
参考文献
- Lieschke, G. J., Currie, P. D. Animal models of human disease: zebrafish swim into view. Nat. Rev. Genet. 8, 353-367 (2007).
- Nolan, T., Hands, R. E., Bustin, S. A. Quantification of mRNA using real-time RT-PCR. Nat. Protoc. 1, 1559-1582 (2006).
- Thisse, C., Thisse, B. High-resolution in situ hybridization to whole-mount zebrafish embryos. Nat. Protoc. 3, 59-69 (2008).
- Detrich, H. W., Westerfield, M. 3rd, Zon, L. I. Overview of the Zebrafish system. Methods Cell Biol. 59, 3-10 (1999).
- Gerdes, J. M., Davis, E. E., Katsanis, N. The vertebrate primary cilium in development, homeostasis, and disease. Cell. 137, 32-45 (2009).
- Hildebrandt, F., Benzing, T., Katsanis, N. Ciliopathies. N. Engl. J. Med. 364, 1533-1543 (2011).
- Zaghloul, N. A., Katsanis, N. Mechanistic insights into Bardet-Biedl syndrome, a model ciliopathy. J. Clin. Invest. 119, 428-437 (2009).
- Marion, V. Exome sequencing identifies mutations in LZTFL1, a BBSome and smoothened trafficking regulator, in a family with Bardet-Biedl syndrome with situs inversus and insertional polydactyly. J. Med. Genet. 49, 317-321 (2012).
- Otto, E. A. Candidate exome capture identifies mutation of SDCCAG8 as the cause of a retinal-renal ciliopathy. Nat. Genet. 42, 840-850 (2010).
- Schaefer, E. Molecular diagnosis reveals genetic heterogeneity for the overlapping MKKS and BBS phenotypes. Eur. J. Med. Genet. 54, 157-160 (2011).
- Katsanis, N. The oligogenic properties of Bardet-Biedl syndrome. Hum. Mol. Genet. 13 Spec No 1, R65-R71 (2004).
- Badano, J. L. Dissection of epistasis in oligogenic Bardet-Biedl syndrome. Nature. 439, 326-330 (1038).
- Badano, J. L. Heterozygous mutations in BBS1, BBS2 and BBS6 have a potential epistatic effect on Bardet-Biedl patients with two mutations at a second BBS locus. Hum. Mol. Genet. 12, 1651-1659 (2003).
- Gerdes, J. M. Disruption of the basal body compromises proteasomal function and perturbs intracellular Wnt response. Nat. Genet. 39, 1350-1360 (2007).
- Leitch, C. C. Hypomorphic mutations in syndromic encephalocele genes are associated with Bardet-Biedl syndrome. Nat. Genet. 40, 443-448 (2008).
- Kimmel, C. B., Ballard, W. W., Kimmel, S. R., Ullmann, B., Schilling, T. F. Stages of embryonic development of the zebrafish. Developmental dynamics : an official publication of the American Association of Anatomists. 203, 253-310 (1995).
- Zaghloul, N. A., et al. Functional analyses of variants reveal a significant role for dominant negative and common alleles in oligogenic Bardet-Biedl syndrome. Proc. Natl. Acad. Sci. U.S.A. 107, 10602-10607 (2010).
- Davis, E. E. TTC21B contributes both causal and modifying alleles across the ciliopathy spectrum. Nat. Genet. 43, 189-196 (2011).
- Sarparanta, J., et al. Mutations affecting the cytoplasmic functions of the co-chaperone DNAJB6 cause limb-girdle muscular dystrophy. Nat. Genet. 44, 450-455 (2012).
- Selcen, D. Mutation in BAG3 causes severe dominant childhood muscular dystrophy. Annals of neurology. 65, 83-89 (2009).
- Golzio, C., et al. KCTD13 is a major driver of mirrored neuroanatomical phenotypes of the 16p11.2 copy number variant. Nature. 485, 363-367 (2012).
- de Pontual, L., et al. Epistasis between RET and BBS mutations modulates enteric innervation and causes syndromic Hirschsprung disease. Proc. Natl. Acad. Sci. U.S.A. 106, 13921-13926 (2009).
- Adzhubei, I. A., et al. A method and server for predicting damaging missense mutations. Nature Methods. 7, 248-249 (2010).
- Kumar, P., Henikoff, S., Ng, P. C. Predicting the effects of coding non-synonymous variants on protein function using the SIFT algorithm. Nat. Protoc. 4, 1073-1081 (2009).
- Calabrese, R., Capriotti, E., Fariselli, P., Martelli, P. L., Casadio, R. Functional annotations improve the predictive score of human disease-related mutations in proteins. Hum. Mutat. 30, 1237-1244 (2009).
- Li, B., et al. Automated inference of molecular mechanisms of disease from amino acid substitutions. Bioinformatics. 25, 2744-2750 (2009).
- Thusberg, J., Olatubosun, A., Vihinen, M. Performance of mutation pathogenicity prediction methods on missense variants. Hum. Mutat. 32, 358-368 (2011).
- Summerton, J. E. Morpholino, siRNA, and S-DNA compared: impact of structure and mechanism of action on off-target effects and sequence specificity. Curr. Top. Med. Chem. 7, 651-660 (2007).
- Riazuddin, S. A., et al. Missense mutations in TCF8 cause late-onset Fuchs corneal dystrophy and interact with FCD4 on chromosome 9p. Am. J. Hum. Genet. 86, 45-53 (2010).
- Shu, X., et al. Knockdown of the zebrafish ortholog of the retinitis pigmentosa 2 (RP2) gene results in retinal degeneration. Investigative Ophthalmology & Visual Science. 52, 2960-2966 (2011).
- Song, P., Pimplikar, S. W. Knockdown of amyloid precursor protein in zebrafish causes defects in motor axon outgrowth. PloS one. 7, e34209(2012).
- Kawahara, G., Guyon, J. R., Nakamura, Y., Kunkel, L. M. Zebrafish models for human FKRP muscular dystrophies. Hum. Mol. Genet. 19, 623-633 (2010).
- Danilova, N., Sakamoto, K. M., Lin, S. Ribosomal protein S19 deficiency in zebrafish leads to developmental abnormalities and defective erythropoiesis through activation of p53 protein family. Blood. 112, 5228-5237 (2008).
- Bohnsack, B. L., Kasprick, D. S., Kish, P. E., Goldman, D., Kahana, A. A zebrafish model of axenfeld-rieger syndrome reveals that pitx2 regulation by retinoic acid is essential for ocular and craniofacial development. Investigative Ophthalmology & Visual Science. 53, 7-22 (2012).
- Oehlers, S. H., et al. The inflammatory bowel disease (IBD) susceptibility genes NOD1 and NOD2 have conserved anti-bacterial roles in zebrafish. Disease Models & Mechanisms. 4, 832-841 (2011).
- Sheng, D., et al. Deletion of the WD40 domain of LRRK2 in Zebrafish causes Parkinsonism-like loss of neurons and locomotive defect. PLoS genetics. 6, e1000914(2010).
- Teng, Y., et al. Loss of zebrafish lgi1b leads to hydrocephalus and sensitization to pentylenetetrazol induced seizure-like behavior. PloS one. 6, e24596(2011).
- Santoriello, C., Zon, L. I. Hooked! Modeling human disease in zebrafish. J. Clin. Invest. 122, 2337-2343 (2012).
- Javidan, Y., Schilling, T. F. Development of cartilage and bone. Methods Cell Biol. 76, 415-436 (2004).
- Lawson, N. D., Weinstein, B. M. In vivo imaging of embryonic vascular development using transgenic zebrafish. Dev. Biol. 248, 307-318 (2002).
- Lee, N. Y. Endoglin regulates PI3-kinase/Akt trafficking and signaling to alter endothelial capillary stability during angiogenesis. Molecular Biology of the Cell. 23, 2412-2423 (2012).
- Huang, C. J., Tu, C. T., Hsiao, C. D., Hsieh, F. J., Tsai, H. J. Germ-line transmission of a myocardium-specific GFP transgene reveals critical regulatory elements in the cardiac myosin light chain 2 promoter of zebrafish. Developmental dynamics : an official publication of the American Association of Anatomists. 228, 30-40 (2003).
- Merveille, A. C., et al. CCDC39 is required for assembly of inner dynein arms and the dynein regulatory complex and for normal ciliary motility in humans and dogs. Nat. Genet. 43, 72-78 (2011).
- Beales, P. L. IFT80, which encodes a conserved intraflagellar transport protein, is mutated in Jeune asphyxiating thoracic dystrophy. Nat. Genet. 39, 727-729 (2007).
- Drummond, I. A., Davidson, A. J. Zebrafish kidney development. Methods Cell Biol. 100, 233-260 (2010).
- Tobin, J. L., Beales, P. L. Restoration of renal function in zebrafish models of ciliopathies. Pediatr. Nephrol. 23, 2095-2099 (2008).
- Putoux, A. KIF7 mutations cause fetal hydrolethalus and acrocallosal syndromes. Nat. Genet. 43, 601-606 (2011).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved