
このコンテンツを視聴するには、JoVE 購読が必要です。 サインイン又は無料トライアルを申し込む。
Method Article
小オリゴマー形成膜タンパク質の沈降平衡:五量体安定性に対するヒスチジンプロトン化の影響
要約
Sedimentation equilibrium (SE) can be used to study protein-protein interactions in a physiological environment. This manuscript describes the use of this technique to determine the effect of pH on the stability of a homo-pentamer formed by the small hydrophobic (SH) protein encoded by the human syncytial respiratory virus (hRSV).
要約
分析用超遠心分離(AUC)は、相互作用の強さの広い範囲にわたって、生理的条件下において高分子間の可逆的な相互作用を研究するために使用することができる。これはAUC定量的に化学量論および生化学過程で一過性で可逆的でホモおよびヘテロ会合の熱力学を評価するための最適な方法になります。沈降平衡(SE)のモダリティでは、拡散と沈降の間のバランスは、特定の相関モデルに依存して半径方向距離の関数としてプロファイルを提供しています。本明細書において、詳細なSEプロトコルは、分析用超遠心を用いた小型膜タンパク質のオリゴマーのサイズ及びモノマー - モノマー関連エネルギーを決定するために記載されている。 AUC-ESは、物理的原理に基づいて、無標識であり、そして水溶性及び膜タンパク質の両方で使用することができる。例は後者の表示され、人間の呼吸器合胞体ウイルスの小さな疎水性(SH)タンパク質(はhRSV五量体イオンチャネルを形成する単一のαヘリックス膜貫通(TM)ドメインを持つ)、65アミノ酸のポリペプチド。 NMRベースの構造データは、SHタンパク質がチャネルの内腔に面して配向され、その膜貫通ドメイン二つのプロトン化のHis残基を有することを示している。 SE実験は、pHが会合定数及びSHタンパク質のオリゴマーサイズにどのように影響するかを決定するために設計されています。五量体形態はすべての場合に保存されたが、その結合定数が低いpHで減少した。これらのデータは、SHタンパク質内の2つのHis残基の内腔の向きと一致SHチャネル活性について観察同様のpH依存性と一致している。後者は、静電反発力が発生し、低pHでのオリゴマーの安定性を低減することができる。要約すると、この方法が適用されるたびに、生理学的条件の微妙なタンパク質 - タンパク質会合の変化に関する定量的情報を測定する必要がある。
概要
分析超遠心分離1-5は弱いと強い相互作用の両方にアクセス可能であること、生理的条件下での高分子の相互作用を研究するための最も重要な方法の一つである。この方法は、無標識であり、光吸収または干渉を利用し、さらに蛍光光学システムは、大きさが6の数桁にわたって濃度範囲にアクセスするために使用することができる。
ほとんどの生化学的過程が可逆的相互作用に依存するため、この方法は特に便利です。これらの相互作用の化学量論および強度を定量的生物学的プロセスを理解することを特徴とする必要があり、多数の方法が、この目的の7,8のために存在する。しかし、一時的な相互作用は、9を研究することは困難である。
巨大分子の相互作用を特徴付けるための方法の選択は、その静的または動的な性質に依存する。最初のケースでは、sedim entation速度(SV)は、半径方向の輸送速度が測定され、複合体が浮力質量と形状の違いに基づいて分画される場合、使用されている。
対照的に、実験の時間スケールで可逆的で動的な関連付けは、物理的に分離することができない。この場合には、自己または非共有結合性相互作用をもたらすヘテロ相互作用は、総タンパク質濃度に依存して平衡状態にある。これらの動的な相互作用は、沈降平衡(SE)及び沈降速度(SV)10の両方で研究することができる。しかし、第一の方法は、実行するために簡単であり、ここで説明されている。平衡拡散沈降の間に到達するようにSEでは、遠心分離が十分に低速で行われる。この時点で、半径方向距離の関数としての光信号(UV-VIS)の平衡プロファイルは、アソシエーション11用に事前設定された熱力学モデルを用いて分析することができる。
ve_content ">本研究では、沈降平衡試験があるため、その疎水性のイオンチャネルを形成し、ウイルス膜タンパク質の自己会合が示されているが、実験は、界面活性剤の存在下で実行され、この場合の密度のさ溶媒は、界面活性剤のそれに一致させる必要があるが、記載されるプロトコルは、水溶性タンパク質の場合には同一の場合と、無溶媒濃度マッチングを必要としないであろうことを除いて。使用されるタンパク質は、世界中のヒト呼吸器合胞体ウイルス(はhRSV)、乳児の下気道疾患の原因となるパラミクソウイルス科に包まニューモ、高齢者や免疫不全の集団12でエンコードされます。最大6400万はhRSV感染16万人の死亡の症例が毎年発生すると報告した。
はhRSVゲノム三膜タンパク質F、G、及び小さな疎水性(SH)を含む11のタンパク質を転写する。 SHタンパク質が関与しているRSV感染の病因に。 SH遺伝子(RSVΔSH)を欠くRSVは、生存可能であったシンシチウムの形成を引き起こし、野生型(WT)ウイルス13-16と同様に増殖した。しかし、RSVΔSHウイルスは上気道15、16にWTよりも低い効率で10倍に複製。また、RSVΔSHウイルスはin vivoでのマウスとチンパンジーモデル13、17で減衰されました。
SHタンパク質は、主にゴルジコン パートメント18の膜64に蓄積する(RSVサブグループA)または65(RSVサブグループB)アミノ酸長いII型内在性膜タンパク質である。 SHタンパク質は高度に保存されている20,21を単一の予測ヘリックス膜貫通(TM)ドメイン19を有している。 C-およびN-末端膜外ドメインは、それぞれ、内腔に/細胞外及び細胞質に配向されている。
両方の合成TMドメイン(残基18-43)及び全長SHタンパク質は、界面活性剤の様々なhomopentamersを形成することが示されている。ホモ五量体の形は、平面脂質二重層22,23におけるチャネル活性を担当しています。脂質二重層におけるTMモノマーの正しい配向は、第1のインターらせん配向に近い内腔であることのHis-22を示した部位特異的な赤外線二色23を使用して決定した。同一のTMドメイン配向は、ドデシルホスホコリンにおける全長タンパク質の五量ヘリックスバンドル(DPC)が22をミセルが再構成されたNMR研究により確認された。この「ミセル」モデルでは、単一のA-らせんTMドメインは、拡張B-ヘアピンによってC末端AヘリックスによってN末端が隣接し、そしてた。 SHタンパク質の2つのプロトン化可能な残基のHis-22およびHis-51、TMドメイン(内腔に配向)に配置され、膜外C末端βヘアピンの先端のそれぞれ、(遠チャンネル孔から)。バイセルエンバイロでnmentは、しかし、TMのαヘリックスは、彼の-51にまで延びており、両方のHis残基は、チャンネル24の内腔にアクセス可能である。チャネル構造は、(のCys-45へのSer-29)狭い地域漏斗のようなアーキテクチャ22は、22は疎水性側鎖(イル·32、イル= 36、イル= 40およびLeu-44)が並んで採用しており、イル·36は、チャンネル内腔に狭いポイントを定義します。のHis-51が最も小さい開口部の先端部に、一方のHis-22は、このファンネルの最大の開口部に配置されている。
本論文では、沈降平衡モードでの分析の遠心分離は、Hisプロトン化は、SHタンパク質五量の安定性に影響を与えるかどうかを決定するために使用されてきた。この場合、SHタンパク質は、そのSHタンパク質の形態五量体オリゴマー22を表示するために以前に使用されているC14-ベタイン界面活性剤で可溶化した。
プロトコル
このプロトコルは、詳細および特別な考慮事項3、25-28のために参照すべきである次のリソースに基づいています。
2 H 2 Oを有する界面活性剤ミセルの1密度マッチング
注:緩衝液の濃度は、界面活性剤ミセルの濃度に一致する必要がある。一般的な密度調節剤は、2 H 2 O、H 2 18 O、2 H 2 18 O、グリセロールおよびスクロース29を含む。 H 2 18 O 2 H 2 Oと同じ密度を有し、タンパク質中の交換可能なプロトンの重水素化が望まれていない場合に良い選択であり得る。この手順では、50mMトリスpH7.3の中で、3-(N、N-dimethylmyristylammonio)-propanesulfonate(C14SB)界面活性剤の濃度は、100mMのNaClを2 H 2 Oと一致するの最初の推測として、下記の濃度30、10、および50%v / vの2 H 2 Oが使用される。
1.1。試料調製
- 以下のストック溶液とフィルターは0.2μmのシリンジフィルターに通して滅菌の準備:50 mLの500 mMトリスpHは7.3と1MのNaCl(10×緩衝液); 1 mLの250 mMのC14SB(50X洗剤液)。
- 20μlの10×緩衝液、4μlの50×洗剤溶液、20μlの2 H 2 O(99.9%)、および156μlの脱イオンH 2 Oを混合することにより、試料溶液 200μlを準備20μlの10×緩衝液、20μlの2 H 2 O(99.9%)、および160μlの脱イオンH 2 Oを混合することにより、 参照溶液200μlを準備
- 適切に2 H 2 OとH 2 Oの量を調整する他の2 H 2 Oの濃度、すなわち、30%および50%を繰り返し、ステップ1.1.2。
1.2。 6チャンネルAの組立細胞へのUC細胞とサンプルのローディング。
注:サンプルローディング方法に応じAUCセルの2つのタイプがある。細胞を密封した後、細胞をロードすることができ、外部充填一方、外部フィルない細胞は、細胞を密封する前にロードされている。外部フィルAUC電池の組み立ては、先に3に記載されている。このプロトコルでは、 外部の充填せず、6チャンネルAUC電池の組み立てについて説明する。主な違いは、別々に締めする必要が両側のねじリングを有していることであり、それは(図1)のハウジングのプラグを必要としない。組み立て工程の違いは以下の強調表示されます。

図1.外部塗りなし6チャンネルAUCセルの分解図。この図は、ベックマン·コールターAN-50 TiおよびAN-60はTi Analytiから変更されているCALローター、細胞、およびカウンターバランスのユーザーマニュアル。
- サファイア窓の代わりに、石英窓(図2)との各AUCセル用の2窓アセンブリを準備します。窓ホルダーにウィンドウのガスケットを置きます。少し窓ライナーを曲げて、ギャップが窓ホルダーキー溝に対向して形成されるように、窓ホルダーにそれを置く。窓ホルダーキー溝にマークを合わせて、窓ライナーの内側にサファイア窓を配置します。
注:石英窓は、圧縮可能であり、したがって、高速28でより多くの光の屈折を作り出す30になるので30000 rpmで上記干渉測定のために、このよう密度マッチングの実験と同様に、サファイアウィンドウが使用されている。サファイア窓は、石英窓より重く、その側面上にエッチングさ "X"を有している。

図2.分解図OF窓アセンブリ。この図は、ベックマン·コールターAN-50 Tiと-60 Tiの分析ローター、細胞、およびカウンターバランスのユーザマニュアルから変更されている
- 部品番号逆さまにセルハウジングを置きます。住宅の鍵と整合キー溝を使用すると、ウィンドウが(図1、左)を下に向けて1窓アセンブリに続いて、下に斜めの側で6セクタ目玉、まずセルハウジングにスライドさせます。
- 軽くコートspinkoteスクリューリングスレッドとネジリングワッシャー。窓アセンブリの上部のネジリングワッシャを置きます。外に面した言葉「OUT」でウィンドウハウジングにネジリングを取り付けます。セル整列ツールを使用して、ネジリングを手で締め。
- トルクレンチを使用して、 唯一の60インチポンドにネジリングを締めます。
- 直立と12正午に位置部品番号でセルを配置します。 LOAD左行に120μlの参照、右の行に110μlのサンプル。各サンプルおよび参照が正しくペアリングされていることを確認してください。
注:正確なサンプル量が重要ではないが、参照サンプルのメニスカスが異なるであろうように、サンプル(5~10μl)を僅かに上回る容量を有する必要がある。 - 慎重にウィンドウが(図1、右)下に向けてセルハウジング1窓アセンブリにスライドさせます。過度に細胞を乱すと内容をこぼさないように注意してください。
- 手順を繰り返し1.2。3および1.2。4、120インチポンドへの第2ネジリングを締める。セルを反転し、120インチポンドに第1スクリューリングを再度締めます。
- ローターに細胞を読み込み、遠心分離機にローターをインストールし、製造元の指示28に従ってモノクロメータをインストールしてください。
注:このSTの詳細をepが、この特許文献3に見出すことができる。
1.3。干渉測定のセットアップ
- 簡単に次のステップに要約され、製造者の指示に応じて、AUC機器のためのユーザ·インタフェース·ソフトウェアを起動し、3000rpmで、各セル用のレーザセットアップおよび放射状リブレーションを行う。
- 1.3.1.1は、十分な真空た後、3000rpmで遠心分離を実行し、(<100ミクロン)に達しました。ユーザインタフェースソフトウェアで干渉パターンをプレビューし、最も高いコントラストを得るために、レーザパラメータを調整する。
- 「均衡」と「干渉」の測定を指定して|(新規ファイルのファイル)は、新しいセットアップファイルを作成します。沈降平衡法(「メソッド」ボタン)を20℃のラン温度で、45000 RPMまたはいずれか高い方のタンパク質試料のために予想される最高速度で実行し、1スキャンを収集するために15分ごとに設定します。 MonitorはHeteroAnalysisのデータファイルを開くと、少なくとも12時間後に「一致」機能を選択することによって、平衡の進行(約一晩、図3)。
注:同様の機能もSEDFIT(オプション|ロードオプション|均衡にテストアプローチ)で利用可能です。

HeteroAnalysisマッチ関数から図3の結果。マッチ関数は、連続スキャンと最終スキャンの間RMSDを比較することによって、平衡の進行状況を監視するために使用することができる。 X軸に漸近RMSD値で示されるように、この例では8時間後に平衡の達成を示している。
1.4。データ分析
- サンプルの各セットは、2 H 2 O濃度に対する動径分布プロファイルの傾きをプロット。
注:配布が非常に浅く、指数そのAPになりますproaches直線。 X軸切片が一致2 H 2 O濃度に対応する。 - より正確な結果を得るためには、いくつかの複製で実験を行う。別の方法として、2 H 2 O濃度の狭い範囲で実験を繰り返します。
C14SBミセルにおけるSHの2沈降平衡
2.1。実行パラメータ
- SEDNTERPを使用して、バッファ密度および粘度、タンパク質部分比体積及び遠心分離速度を計算する。 D 2 O濃度を含む、「バッファ·データの選択」の項でバッファ密度と粘度は、[計算を計算し、それに応じて緩衝液成分を入力します。
- 2.1.1.1「V-バー」セクションの蛋白部分比体積は、[計算を計算し、タンパク質のアミノ酸配列を入力します。 ":N =このモノマーからオリゴマーを作る」で最も高い期待オリゴマーのサイズを指定この場合フィールド、N = 5σ≈1までメインウィンドウのRPMフィールドに値を入力して速度を計算。これは、半径方向の分布形状25の良好な指数関数的な形状を確保するための経験則である。
注:ρ= 1.03839グラム/ mlで、η= 1.0267センチポイズ= 0.7569ミリリットル/ gの、ω1 = 16000 RPM:この実験のために計算された値は以下の通りであった。 - 1速での分布プロファイルとの間に十分な差が次の25を確実にするために従うように、後続の速度を計算します。
注:これは、考慮溶液カラム(充填容積)かかりSEDFITで「平衡ロータ速度関数を推定する」機能からもで行うことができる。
2.2。試料調製
- 100μlの10Xバッファーsolutioを混合することにより、密度マッチング実験(セクション1)から求めた5 mMのC14SBで1ml 基準溶液と32.3%2 H 2 Oを準備nは(ステップ1.1.1)、20μlの50X界面活性剤溶液(ステップ1.1.1)、323μlの2 H 2 O(99.9%)および527μlの脱イオンH 2 O.
- 例えばメタノール又は50%v / vのアセトニトリル水溶液のような適切な溶媒中で(先に31記載の発現および精製)凍結乾燥し、HPLCで精製したSHペプチドを溶解する。に希釈したときにマイクロリットルスケールのUV / Vis分光光度計、アリコート三つのサンプル用の280を得た、12ミリメートル = 0.3、0.5、0.8(280、10ミリメートル = 0.25、0.417、0.67)各溶解ペプチドのA280を測定する130μL。一晩試料を凍結乾燥し、再懸濁する130μlの標準液(ステップ2.2.1)で試料溶液を得た。
注:それがTrp及びTyr残基が含まれているため、SHタンパク質は、280nmにおけるUV /可視吸光度を検出することができる。芳香族残基なしのタンパク質は、トリプトファン含有変異体を使用して、適切な発色団でそれらをタグ付けすることによって、または妨害を用いて検出することができるNCEの測定値の代わりに、吸光度。 - 石英窓付きの 6チャンネルAUCセルを組み立てるためのセクション1.2の手順に従ってください。ロータ中心から最も遠いロータ中心に最も近いチャネルにおける最高濃度のサンプル(280、12ミリメートル = 0.8)と最低濃度のサンプル(280、12ミリメートル = 0.3)をロードします。
2.3。吸光度測定のセットアップ
- 「均衡」と「吸光度」の測定を指定して|(新規ファイルのファイル)は、新しいセットアップファイルを作成します。検出器波長として280nmのを指定します。
- ラジアルステップサイズで例えば、低解像度でのデータ収集を指定して、スキャンオプションで「最初のスキャンの前にラジアルキャリブレーション」をチェックして、3000rpmで放射状のキャリブレーションを実行= 0.01センチ、レプリケート= 3(低解像度、速い)、および単一のスキャンを実行する。スキャンが完了したら、オプションをオフにします。
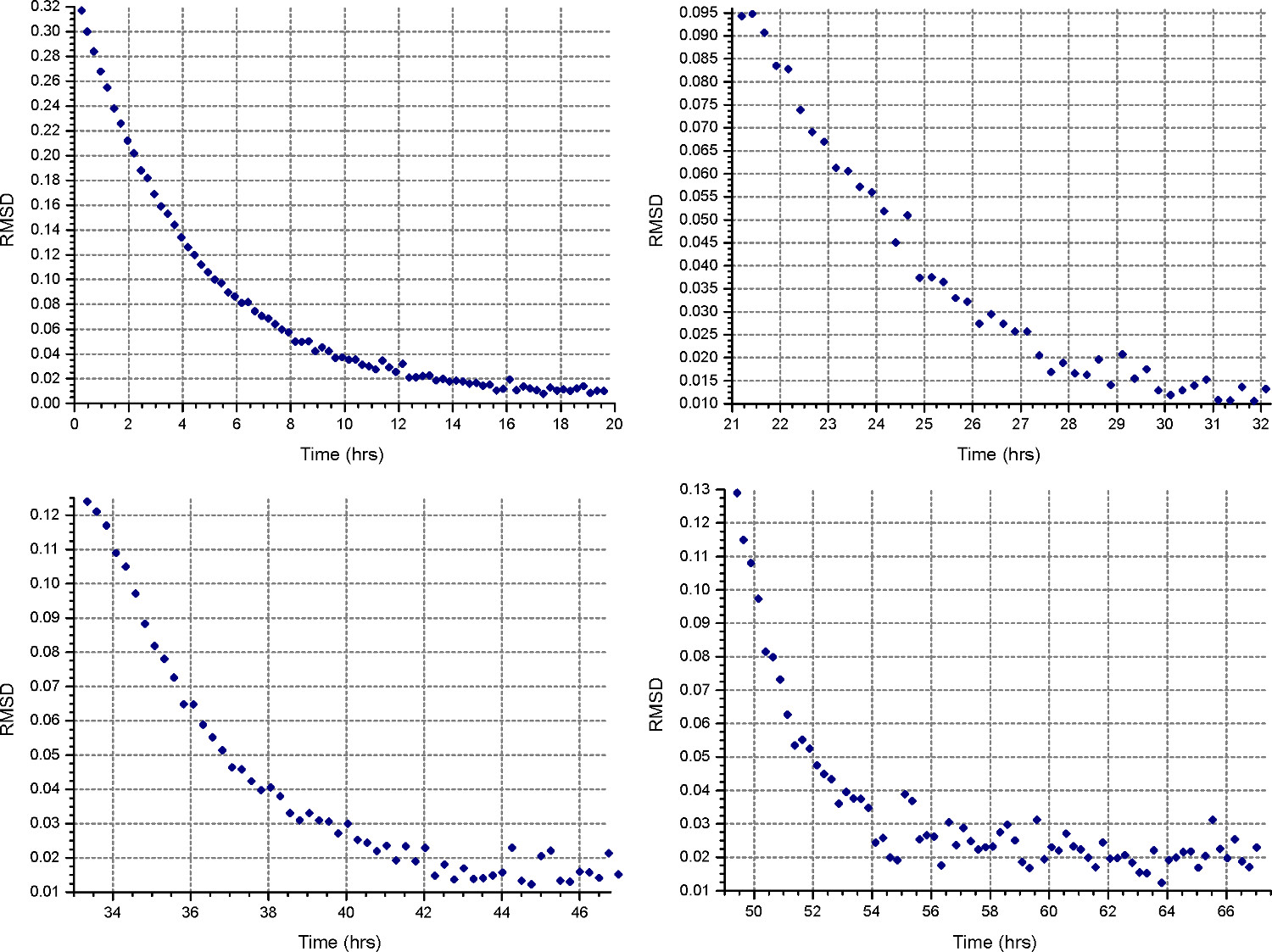
- 沈降平衡法(「メトを設定するD "ボタン)20℃で、ステップ2.1.3で計算された第1の速度で動作し、1スキャンごとに30分を収集する。各細胞の「詳細」には、ステップ2.3.2のように低解像度でのデータ収集を指定します。 HeteroAnalysisのデータファイルを開くと、少なくとも18時間後に「一致」機能を選択することによって、平衡の進行状況を監視(約一晩、図4)。
注:その後の速度はより少ない時間がかかるだろう、一方、均衡の達成は、第1速用の実質的に長い時間がかかることがあります。

HeteroAnalysis MATCH関数から図4.結果は、第 1、第2の速度は、(上部左右)が平衡に達したと思われるが、それは確かにいくつかのより多くの時間を待つことをお勧めします。比較して、第3および第4の速度が(左下および右)を明確に平衡に達している短い時間で。 この図の拡大版をご覧になるにはこちらをクリックしてください。
{kind=link}
- 平衡に達した後、ラジアルステップサイズ= 0.001 cmのレプリケート= 10(高分解能、遅い)と、高解像度、例えば、少なくとも一回の走査を収集する。
- スキャンが完了したら、次の速度のためのステップ2.3.3と2.3.4を繰り返します。
- 、沈降平衡法は、ステップ2.1.3において算出すべての速度を含むことをスキャンし、それぞれの平衡時間後に1スキャンを収集するように設定する必要に応じて、ときに、各速度の平衡に達するのに必要な時間は(計算されたまたは経験から)知られているスピード。この場合、各セルの「詳細」の高解像度データ収集を指定する。
2.4。 SEDFITとSEDPHATにおけるデータ解析
注:データ分析リーダーで詳細および考慮事項については、Tと呼ばれているwww.analyticalultracentrifugation.com:次のWebサイトO。
- SEDFITに高解像度スキャンを開きます(データ|ロード沈降平衡データ)と(これらは、各サンプルの異なる濃度に対応し、オプション|ロードオプション| 3サブセットで6チャンネル生データを保存します)3チャネルにデータを分割する。
- 再オープンSEDFITで同じサンプルと同じ濃度が、異なる速度に属しているデータ·ファイル。メニスカス(縦の赤線)、セルの底(縦の青線)とフィッティングの限界(縦の緑色の線)を調整し、SEDPHATで使用するためにデータをエクスポート(データ| SEDPHATするデータのエクスポート)。入力ステップ2.1.1で算出されたパラメータ、並びにロータ型と目玉タイプが要求されている。すべてのサンプルおよび濃度について、この手順を繰り返します。
- SEDPHATに同じサンプルからすべてのデータ(すべての濃度と速度)を開き、実験パラメータを入力。一例を図4に示す。 5。
D 2 Oがバッファに追加された場合、申:注意交換可能なプロトンのerationは著しく、特に水溶性のタンパク質について、タンパク質の分子量を変化させることができる。膜に埋め込まれた領域が交換から保護されているため、膜タンパク質、SHタンパク質のような、特に小さなものは、あまり影響を受けます。これを修正するには、入力「バッファDのモル分率」。
注:このステップではメニューデータを選択することで、個別に編集データセットを保存することをお勧めします|コピーすべてのデータと保存新しい設定として。 - モデルを選択して、そのモデルのグローバルパラメータを入力してください。
注:例として、「モノマー-nのメール自己会」モデル及びそのパラメータを図4に示す。 6。

図5.実験パラメータに記入する方法の例。
ファイル/ ftp_upload / 52404 / 52404fig6.jpg "/>
図6.モノマー-N-Merの自己会合モデルのグローバルパラメータに記入する方法の例。
- メニューフィットを選択することで、グローバルフィットを実行します|グローバルフィットとフィット収束するまで待ちます。書き留め(またはのスクリーンショットを撮る)フィッティング結果、特にグローバルな減少カイ二乗とKの値を記録します。熱力学情報を表示|そのようなフィット感データやメニューをコピーし、表示からフィッティング残差などの他のデータを抽出します。
- グローバルパラメータに戻り、(1)モノマー分子量に合わせてステップ2.4.5を繰り返してMをご確認ください。フィット分子量や地球減少カイ二乗を下に注意してください。
- 繰り返しステップ2.4.3は、各モデルをテストするための2.4.6及び減少した地球カイ二乗値ならびにフィッティング残差を比較することにより、各モデルの適合の品質を比較する。
注意:小とランダムフィッティング残差が一般的に良好なフィット感を示し、でしょう最高のフィットモデル最小のグローバルな減少カイ二乗を持っている。フィットモノマー分子量とそのカイ二乗値は固定(理論値)の分子量のものと実質的に異なるべきではありません。 - 第1の選択統計によって値で取得したログのKaの信頼区間を計算|重大なエラー表面突起のためのカイ二乗と希望信頼区間を入力する。 1次元のエラー表面突起を生成し、ログのKa用のカイ二乗値を得るためにグローバルパラメータダイアログでのKaを記録選択解除|次、統計に行く。
注:読者は、この方法33の方法32だけでなく、イラストの詳細については、以下のソースを(http://www.analyticalultracentrifugation.com/ sedphat / statistics.htm)に相談することをお勧めします。
結果
50mMのトリスC14SBの界面活性剤ミセルの径方向分布プロフィール、100mMのNaCl pH7.3の形の線形モデル( 図7A)に装着することができ、非常に浅い指数関数。この分布の傾きは逆にD 2 O濃度( 図7B)と相関している。傾きがゼロとなる点、 すなわち、マッチングD 2 O濃度は、32.3%であることが見出された。
以下の図7を
ディスカッション
本論文では、平衡沈降を使用して、洗剤の小さな膜タンパク質のオリゴマー化の試料調製および分析のための実験プロトコルを提供します。密度マッチングステップが必要とされないように記載されるプロトコルは、可溶性タンパク質に同等に有効で-and simpler-ある。実際に、システムは、界面活性剤とタンパク質との混合物で構成されている。それは、粒子浮選に寄与しないように、沈降研...
開示事項
The authors have nothing to disclose.
謝辞
This work has been funded by the National Research Foundation grant NRF-CRP4-2008-02 (J.T.) and Tier 1 grant RG 51/13.
資料
| Name | Company | Catalog Number | Comments |
| 3-(N,N-dimethylmyristylammonio)propanesulfonate | Sigma | T0807 | |
| Deuterium oxide 99.8% | Cambridge Isotope | DLM-4-99.8 | |
| An-50 Ti Rotor, Analytical, 8-Place | Beckman Coulter | 363782 | |
| An-60 Ti Rotor, Analytical, 4-Place | Beckman Coulter | 361964 | |
| Cell housing | Beckman Coulter | 334784 | |
| 12 mm six-channel centerpiece, epon charcoal-filled | Beckman Coulter | 331376 | |
| Window holder | Beckman Coulter | 305037 | |
| Window gasket | Beckman Coulter | 327021 | |
| Window liner | Beckman Coulter | 362329 | |
| Sapphire window | Beckman Coulter | 307177 | |
| Quartz window | Beckman Coulter | 301730 | |
| Screw-ring washer | Beckman Coulter | 362328 | |
| Screw ring | Beckman Coulter | 301922 | |
| Spinkote | Beckman Coulter | 306812 | |
| Torque stand assembly | Beckman Coulter | 361318 | |
| Counterbalance | Beckman Coulter | 360219 | |
| Cell alignment tool | Beckman Coulter | 362340 | |
| SEDNTERP | http://bitcwiki.sr.unh.edu/index.php/Main_Page | ||
| HeteroAnalysis | http://www.biotech.uconn.edu/auf/?i=aufftp | ||
| SEDFIT | http://www.analyticalultracentrifugation .com/sedfit.htm | ||
| SEDPHAT | http://www.analyticalultracentrifugation .com/sedphat/default.htm |
参考文献
- Laue, T. M., Stafford, W. F. Modern applications of analytical ultracentrifugation. Annu. Rev. Biophys. Biomol. Struct. 28, 75-100 (1999).
- Lebowitz, J., Lewis, M. S., Schuck, P. Modern analytical ultracentrifugation in protein science: A tutorial review. Protein Sci. 11, 2067-2079 (2002).
- Balbo, A., Zhao, H., Brown, P. H., Schuck, P. . Assembly, Loading, and Alignment of an Analytical Ultracentrifuge Sample Cell. , e1530 (2009).
- Rivas, G., Stafford, W., Minton, A. P. Characterization of heterologous protein-protein interactions using analytical ultracentrifugation. Methods-a Companion to Methods in Enzymology. 19, 194-212 (1999).
- Howlett, G. J., Minton, A. P., Rivas, G. Analytical ultracentrifugation for association and assembly the study of protein. Curr. Opin. Chem. Biol. 10, 430-436 (2006).
- MacGregor, I. K., Anderson, A. L., Laue, T. M. Fluorescence detection for the XLI analytical ultracentrifuge. Biophys. Chem. 108, 165-185 (2004).
- Phizicky, E. M., Fields, S. Protein-Protein Interactions - Methods for Detection and Analysis. Microbiol. Rev. 59, 94-123 (1995).
- Alexandrov, A. A facile method for high-throughput co-expression of protein pairs. Mol. Cell. Proteomics. 3, 934-938 (2004).
- Nooren, I. M. A., Thornton, J. M. Structural characterisation and functional significance of transient protein-protein interactions. J. Mol. Biol. 325, 991-1018 (2003).
- Ebel, C. Sedimentation velocity to characterize surfactants and solubilized membrane proteins. Methods. 54, 56-66 (2011).
- Minton, A. P. Quantitative characterization of reversible macromolecular associations via sedimentation equilibrium: an introduction. Exp. Mol. Med. 32, 1-5 (2000).
- Dowell, S. F. Respiratory syncytial virus is an important cause of community-acquired lower respiratory infection among hospitalized adults. J. Infect. Dis. 174, 456-462 (1996).
- Bukreyev, A., Whitehead, S. S., Murphy, B. R., Collins, P. L. Recombinant respiratory syncytial virus from which the entire SH gene has been deleted grows efficiently in cell culture and exhibits site-specific attenuation in the respiratory tract of the mouse. J. Virol. 71, 8973-8982 (1997).
- Fuentes, S., Tran, K. C., Luthra, P., Teng, M. N., He, B. Function of the respiratory syncytial virus small hydrophobic protein. J. Virol. 81, 8361-8366 (2007).
- Jin, H. Recombinant respiratory syncytial viruses with deletions in the NS1, NS2, SH, and M2-2 genes are attenuated in vitro and in vivo. Virology. 273, 210-218 (2000).
- Karron, R. A. Respiratory syncytial virus (RSV) SH and G proteins are not essential for viral replication in vitro: clinical evaluation and molecular characterization of a cold-passaged, attenuated RSV subgroup B. Proc. Nat. Acad. Sci. USA. 94, 13961-13966 (1997).
- Whitehead, S. S. Recombinant respiratory syncytial virus bearing a deletion of either the NS2 or SH gene is attenuated in chimpanzees. J. Virol. 73, 3438-3442 (1999).
- Rixon, H. W. The small hydrophobic (SH) protein accumulates within lipid-raft structures of the Golgi complex during respiratory syncytial virus infection. J. Gen. Virol. 85, 1153-1165 (2004).
- Collins, P. L., Mottet, G. Membrane orientation and oligomerization of the small hydrophobic protein of human respiratory syncytial virus. J. Gen. Virol. 74, 1445-1450 (1993).
- Collins, P. L., Olmsted, R. A., Johnson, P. R. The small hydrophobic protein of human respiratory syncytial virus: comparison between antigenic subgroups A and B. J. Gen. Virol. 71, 1571-1576 (1990).
- Chen, M. D., Vazquez, M., Buonocore, L., Kahn, J. S. Conservation of the respiratory syncytial virus SH gene. J. Infect. Dis. 182, 1228-1233 (2000).
- Gan, S. W. The small hydrophobic protein of the human respiratory syncytial virus forms pentameric ion channels. J. Biol. Chem. 287, 24671-24689 (2012).
- Gan, S. W., Ng, L., Xin, L., Gong, X., Torres, J. Structure and ion channel activity of the human respiratory syncytial virus (hRSV) small hydrophobic protein transmembrane domain. Protein Sci. 17, 813-820 (2008).
- Li, Y. Inhibition of the Human Respiratory Syncytial Virus Small Hydrophobic Protein and Structural variations in a bicelle environment. J. Virol. 88 (22), 11899-914 (2014).
- Burgess, N. K., Stanley, A. M., Fleming, K. G. Determination of membrane protein molecular weights and association equilibrium constants using sedimentation equilibrium and sedimentation velocity. Meth. Cell. Biol. 84, 181-211 (2008).
- Cole, J. L., Lary, J. W., Moody, T. P., Laue, T. M. Analytical Ultracentrifugation: Sedimentation Velocity and Sedimentation Equilibrium. Meth. Cell. Biol. 84, 143-179 (2008).
- Fleming, K. G. Determination of membrane protein molecular weight using sedimentation equilibrium analytical ultracentrifugation. Curr. Protoc. Prot. Sci. 53, 17.12.11-17.12.13 (2008).
- . . An-50 Ti and An-60 Ti Analytical Rotor, Cells, and Counterbalance. , (2005).
- Mayer, G. Studying membrane proteins in detergent solution by analytical ultracentrifugation: Different methods for density matching. Prog. Colloid Polym. Sci. 113, 176-181 (1999).
- Laue, T. Ch. 20.3. Current Protocols in Protein Science. 20, 20.23.21-20.23.13 (2001).
- Gan, S. W. The Small Hydrophobic Protein Of The Human Respiratory Syncytial Virus Forms Pentameric Ion Channels. J. Biol. Chem. 287, 24671-24689 (2012).
- Bevington, P. R., Robinson, D. K. . Data reduction and error analysis for the physical sciences. 336, (1969).
- Schuck, P., Radu, C. G., Ward, E. S. Sedimentation equilibrium analysis of recombinant mouse FcRn with murine IgG1. Molecular Immunology. 36, 1117-1125 (1999).
- Gan, S. W., Vararattanavech, A., Nordin, N., Eshaghi, S., Torres, J. A cost-effective method for simultaneous homo-oligomeric size determination and monodispersity conditions for membrane proteins. Anal. Biochem. 416, 100-106 (2011).
- Montserret, R. NMR structure and ion channel activity of the p7 protein from hepatitis C virus). J. Biol. Chem. 285, 31446-31461 (2010).
- Stouffer, A. L., DeGrado, W. F., Lear, J. D. Analytical Ultracentrifugation Studies of the Influenza M2 Homotetramerization Equilibrium in Detergent Solutions. Progr Colloid Polym Sci. 131, 108-115 (2006).
- Sorkin, A., von Zastrow, M. Signal transduction and endocytosis: Close encounters of many kinds. Nat. Rev. Mol. Cell Biol. 3, 600-614 (2002).
- Gan, S. W., Ng, L., Lin, X., Gong, X., Torres, J. Structure and ion channel activity of the human respiratory syncytial virus (hRSV) small hydrophobic protein transmembrane domain. Protein science : a publication of the Protein Society. 17, 813-820 (2008).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved