
Method Article
분열 효 모 Schizosaccharomyces pombe 에 깊은 시퀀싱 지원, 자발적인 억압 화면
요약
선물이 분열에 간단한 억압 화면 프로토콜 효 모. 이 방법은 효율적, 돌연 무료이 고 단일 게놈 소재 시에 자주 발생 하는 돌연변이 대 한 선택적입니다. 프로토콜은 진압 완화 돌연변이 또는 약물에 의해 발생 하는 액체 문화에서 성장 결함 격리 적합 합니다.
초록
Phenotypic 결함 돌연변이 의해 발생을 억제 하는 돌연변이 체 대립 유전자에 대 한 유전 스크린 밀접 하 게 관련 된 생화학 경로에 속하는 유전자를 식별 하는 강력한 접근 이다. 합성 유전자 배열 (SGA) 분석, 및 자외선 (UV) 또는 에틸 methanesulfonate (EMS) 또는 N-에틸-N-nitrosourea (ENU) 같은 화학 물질을 사용 하 여 임의의 mutagenesis 기법 등 이전 방법을 널리 사용 하지만 종종 비용이 많이 드는 고 힘 드는. 또한, 이러한 돌연 기반 심사 방법을 자주 분리는 진압의 복합성에 추가 하는 여러 돌연변이 유도 하는 유기 체에 심각한 부작용에 연관 됩니다. 여기, 우리 Schizosaccharomyces pombe성장 결함을 부여 하는 돌연변이에 억압 돌연변이 식별 하는 간단 하 고 효과적인 프로토콜을 제시. 연장된 기간 동안 자동된 96 잘 접시 리더를 사용 하 여 복구에 대 한 표준 풍부한 액체 미디어 또는 합성 액체 미디어 성장 결핍과 세포의 적합성을 모니터링할 수 있습니다. 일단 셀 문화에서 억압 돌연변이 획득, 하위 노르만족 부모의 셀의. 부모 셀의 경쟁력 있는 성장 이점을 복구 된 셀 다음 격리 하 고 부모의 셀과 backcrossed 수 있습니다. 억압 돌연변이 다음 전체 게놈 시퀀싱을 사용 하 여 식별 됩니다. 이 방법을 사용 하 여, 우리는 성공적으로 고립 된 Elf1는 AAA + 가족 ATPase 핵 mRNA 전송 및 게놈 안정성의 유지 보수에 중요 한 그의 손실에 의해 발생 하는 심각한 성장 결함을 완화 하는 여러 진압. 현재 400 유전자 이상에 있다 S. pombe 성장 결함을 부여 하는 돌연변이 함께. 이 유전자의 많은 특징이 되지 않습니다, 우리는 우리의 방법은이 사용자 친화적인, 높은 처리량 방법 소설 기능 상호 작용의 id를 서두르는 것입니다 제안 합니다.
서문
유전자 간의 기능 링크 이해의 기초는 분자 따라 장치를 마더보드에는 복잡 한 유전 특성 생산 다양 한 고기1분기를 식별 하는 기능에 의존 합니다. 분열 효 모, Schizosaccharomyces pombe (S. pombe)에서 단백질 코딩 유전자의 대부분은 생존2불가결. 이 결과이 유전자의 unimportance 하지만 이러한 유전자가 속한 생화학 경로 기본 복잡 한 보상 메커니즘에 오히려 말을 하지 않습니다. 이러한 보상 메커니즘을 해 부 epistasis 지도, 포괄적인 유전 상호 작용을 발견 하 고 확대 기능 생화학 경로3,4의 우리의 이해는 생성 했다.
높은 처리량 방법 (예를 들어, 합성 유전자 배열 분석, 또는 SGA) 신진 효 모, 게놈 전체 유전자 상호 작용을 식별 하 여 분열 효 모5,6에 사용 하기 위해 확장 되어 개발 되었습니다. 이러한 방법을 자주 모든 가능한 단일 단백질 코딩 유전자 삭제를 포함 하는 긴장의 라이브러리에 의존 (약 3300 단일 삭제 돌연변이 취재 분열 효 모 게놈의 92% 이상), 사이 유전 십자가 수행 하기 위해 로봇 팔 필요는 관심과 library6에서 모든 가능한 긴장 긴장 또한, SGA 기술을 적절 하 고 효율적인 짝짓기를 라이브러리 긴장의 능력에 따라, 현재 444로 비정상 이다 표현 형 특징 S. pombe2유전자.
유전 상호 작용의 복잡성에도 불구 하 고 각 유전자의 개별 돌연변이 나르는 2 개의 긴장의 표현 형에 2 개의 유전자에 돌연변이 들고 긴장의 표현 형을 비교 하나를 가질 수의 2 개의 주목할 만한 결과: 1) 이중 돌연변이 표현 형이 가장 극단적인 경우, 치 사 율, 또는 질병의 형태로 예상된 곱셈 부모의 고기 보다 더. 이 부정적인 유전자 상호 작용 이라고 그리고 일반적으로 두 유전자 병렬 생물학 통로에 행동 하는 서명입니다. 2) 이중 돌연변이 표현 형 부모의 고기로 알려진 또한 긍정적인 유전자 상호 작용의 예상된 조합 보다 낫다. 때문에이 유전자 동일한 프로세스에서 작동 나타냅니다 긍정적인 유전자 상호 작용 특히 흥미롭습니다. 2 개의 긍정적인 상호 작용 유전자는 3 개의 잠재적인 관계: 돌연변이 유전자 수 최대-규제는 병렬 통로에 다른 유전자의 표현, 두 유전자는 서로 동일한 통로 하류 내 콘서트에서 일할 수 있습니다 또는 두 유전자 인코딩 직접 상호 작용 하는 단백질. 따라서, 긍정적인 유전자 상호 작용 유전자 규정 노드를 지도 하 고 생화학 경로7,8에 새롭거나 유전자 분류를 사용할 수 있습니다.
억압은 돌연변이 다른 유전자의 돌연변이의 질병 표현 형을 완화 수 있는 일반적으로 대표 하는 두 개의 유전자9,10사이의 긍정적인 유전 상호 작용. 그들은 억제 하는 돌연변이의 다른 장소에 억압 돌연변이 extragenic 진압으로 알려져 있습니다. 그들은 합성으로 치명적인 형 (나사로 효과 라고도 함)11를 구출 하 여 비-가능한 유전자 변이 공부에 특히 귀중 한. 그들은 또한 유전 질병12,13치료에 잠재적인 치료 응용 프로그램 있다.
모든 이러한 이유로, 다양 한 모델 생물에서 억압 돌연변이의 식별은 다양 한 생화학 경로14,,1516의 우리의 이해를 촉진 하기 위하여 널리 이용 되어. 진압에 대 한 심사에 돌연변이의 표현 형에 일반적으로 기반으로 하며 돌연변이 표현 형을 완화 것을 임의의 mutagenesis 실시. 거의 모든 모형 유기 체는 무작위 mutagenesis 방법을 설립 했다. 예를 들어 N-에틸-N-nitrosourea (ENU) 및 ethylmethanesulfonate (EMS), 두 mutagens 포인트 DNA에 돌연변이 유도 할 수 있다, 쥐17,,1819 에 박테리아에서 다양 한 모델에서 널리 채택 된다 . 또한, 염화 망간 오래 사용 되었습니다 효 모에 망간 양이온의 능력을 억제 하는 DNA 수 선 통로20에 대 한. 또 다른 일반적인 접근은 UV 유도 된 mutagenesis 생성 게놈 넓은 돌연변이 pyrimidine 이합체21,22.
비록 억압 돌연변이 식별 하기 위해 화학 mutagenesis의 활용은 인기가, 메서드는 위험한 화학 물질, 높은 변수 성공률, 그리고 여분의 혼란 변수의 도입의 사용을 포함 하 여 많은 단점 여러 세포 프로세스23,24에 mutagen의 부정적인 부작용을 제시한. 또한, 화학 mutagenesis는 종종 추가 유전 사용 하 여 복잡 하 고 시퀀싱 기술을 억압 표현 형 유기 체25에서 수 여 하는 정확한 돌연변이 식별 하는 게놈에서 여러 돌연변이 유도 합니다.
현재 mutagenesis 방식의 단점을 해결 하기 위해 우리는 어떤 mutagens 또는 삭제 라이브러리에 의존 하지 않는 분열 효 모에 자발적인 억압 돌연변이 대 한 화면을 방법 제시. 메서드를 사용 하면 격리 긍정적인 선택 분석 결과 통해 진압 합니다. 이 방법의 원리는 자동된 플레이트 리더에 의해 모니터링 될 수 이다 액체 문화에서 돌연변이 억압 부분 모집단의 성장 우위를 기반으로 합니다. 짝짓기와 감수 분열 한 유전 배경 정리 또는 전체 게놈 시퀀싱 전에 진압의 monogenic 대립 유전자의 존재를 확인 하려는 경우에 사용 됩니다. 억제 형 단일 돌연변이 의해 발생 하는 경우 억압 형과 부모의 종자 backcrossing 후 2: 2를 분리 합니다. 억압 돌연변이 다음 전체 게놈 시퀀싱을 사용 하 여 확인할 수 있습니다. 이 메서드는 액체 문화에서 큰 인구를 성장할 수 있는 모든 미생물에 진압 심사 적용 제안 한다.
프로토콜
1. 스트레인 건설 및 준비
- 돌연변이 또는 (yfm, 좋아하는 돌연변이) 유전자의 삭제26위에서 설명한 대로 표준 사이트 감독 mutagenesis (SDM)를 사용 하 여 생성 합니다.
- 시작 하기 전에 화면, (최적의) 유전 배경 청소 부모의 긴장으로 신선한 태어난 돌연변이 세포를 생성 하는 야생-타입 긴장과 돌연변이 체 긴장 backcross. 연속 부모의 스트레인 개별 식민지 표준 리치 미디어 판에. 무작위로 8 16 독립적인 식민지 (생물 복제) 플레이트 리더 분석 결과 대 한 원하는 돌연변이와 선택 (3.1 참조).
참고:이 프로토콜 부모의 긴장 있다 액체 미디어 (최소 또는 풍부 하 고, 마약, 없이 또는 성장 결함을 일으키는 원인이 되는 온도 교대)에 결함 성장 하는 경우에 효과적입니다. 모든 부모의 긴장 단일 따라서 유전자 보완 짝짓기 형식과 다른 단일 긴장 교차 될 수 있게 해야 합니다.
2. 플레이트 리더 분석 결과
- 메 마른 주걱 단계 1.1 (정확한 양을 접종 시작 하는 데 필요한 문화)에서 준비 하는 식민지의 각각의 작은 금액을 소요와 96 잘 폴리스 티 렌 microplate에. (부자 또는 최소한의 또는 약 없이) 적절 한 액체 미디어의 200 µ L에서 식민지의 각각을 일시 중단 합니다. 동일한 미디어 (셀)의 200 µ L을 포함 하는 접시에 모든 행에 대해 빈 잘 포함 됩니다.
- 플레이트 리더 탐지 소프트웨어에는 다음 프로토콜 자동된 microplate 리더에 연결 된 실행: 연속 빠른 궤도 흔들어 (425 cpm, 3 m m 진폭)와 30 ° C에서 24 시간 및 온도 대 한 운동 프로그램을 설정. 광학 밀도, 600nm의 파장에서 빛 분산을 측정 하는 광학 읽기 설정 하 고 2 분 (잘 당 24 h 동안 721 총 읽습니다)의 독서 주파수에서 접시까지 읽기 빛을 설정 합니다.
- 24 h, 마지막 숨겨진된 광학 밀도 판독 (숨겨진된 OD600)를 기록 하 고 다음 수식을 사용 하 여 각 마약까지 샘플을 희석 하는 데 필요한 볼륨을 확인 후 = 0.1:

참고: 플레이트 리더 소프트웨어에서 데이터를 내보내기 하 고 스프레드시트 소프트웨어를 사용 하 여 일괄 처리 하는 함수에서 각 실험 잘 사용할 수 희석 볼륨 처리로 위의 수식을 삽입할. - 모든 24 시간 하루 0 마약으로 같은 미디어를 사용 하 여 샘플의 각각 희석 0.1 (약 1.5 x 106 셀/mL) = 2.3 단계에 표시 된 수식을 사용 하 여. 모든 성장 곡선 매일 생성 되 고 동일한 유전적 배경 가진 동료의 나머지 부분 보다 훨씬 높은 최종 마약 또는 유사의 성장 곡선에 의해 판단 하는 증가 증가 비율을 보여 주는 어떤 개별 식민지를 저장 야생-타입의 식민지
참고:이 분석 결과 일반적으로 약 7-14 일 걸립니다. 무 균 조건 하에서 모든 단계를 수행 합니다.
3. 억압 식민지 형의 확인의 선택.
- 플레이트 리더 분석 결과 (2.4 단계)의 마지막 날에서 액체 문화를 눈에 띄게 복구 된 성장 율, 아마도 부모의 돌연변이의 표현 형을 완화 수 있는 억압 돌연변이 확보 하 여 저장 합니다. 전송 및 250 µ L 액체 문화 50% 글리세롤의 250 µ L을 포함 하는 cryotube 혼합. 플래시 액체 질소에는 세포를 동결 하 고 무기한-80oC에서 긴장을 저장 합니다.
- 억압 돌연변이 유전자 상속 요소 인지를 확인 하려면 메서드 사용 하 여 표준 유전 교차점 yfm P를 (대 한 플레이트 리더 분석 결과의 시작 부분에 사용 되는 부모, 긴장) yfm S (대 한 억압, 부담 끝에 저장 된 플레이트 리더 분석 결과의). 억압 돌연변이 유전자 상속 요소 이면 실제로 yfm P × yfm S까지 있는 두 식민지 부모의 스트레인의 질병 표현 형 있고 두 식민지 억압의 복구 된 성장 율을 산출 해야 한다 스트레인입니다.
- 3.2 단계의 십자가에서 억압 형 (S 긴장)와 3 개의 식민지를 선택 하 고 동일한 유전자에서 부모의 표현 형 (P 스트레인)와 3 개의 식민지 (각 3 생물 복제), 교차 하 고 게놈 DNA 추출 진행 및 시퀀싱 단계 아래입니다.
참고: 단계 3.2 및 3.3이 좋습니다 있지만 필요 하지 않습니다. 또는, 하나 하나의 식민지로 풍부한 매체에 3.1에서 수집 하는 복구 된 액체 문화 확산 다음 무작위로 추가 유전자 확인 없이 전체 게놈 시퀀싱에 대 한 생물 학적 triplicates로 3 개의 식민지를 선택 수 있습니다. 이 경우에, 부모의 긴장의 3 개의 생물 학적 triplicates 게놈 시퀀싱 비교를 위해 사용 되어야 한다.
4. 게놈 DNA 추출, 도서관 생산 및 연속.
- DNA 추출, 라이브러리 준비 및 시퀀싱, 무작위로 yfm P 스트레인, 당 3 개의 생물 복제 선택 하 고 3 생물 각 개별적으로 일어 나 yfm S 긴장 또는 유전 십자가 (3.2 단계)에서 복제는 접시 S 스트레인 (단계 3.3 참고)의 단일 식민지를 확산.
- 리치 미디어에 10 mL 문화에서 긴장 중간 로그 단계로 성장 (마약 = 0.5-0.8, 0.75-1.2 x 107 셀/mL에 대 한), 떨고 인큐베이터를 사용 하 여 지속적으로 250 rpm 동요 30 ° C에 액체 문화를 성장 하 고. 1000 x g에 5 분 동안 4 ° C에서 원심 분리 하 여 세포를 수집 합니다.
- 수송과 세포 DNA 추출 버퍼의 400 µ L 일시 중단 (2% 트라이 톤 X-100, 1 %SDS, 100 mM NaCl, 10 mM Tris-Cl (pH 8.0), 1 m m 나2-EDTA), 유리 구슬의 400 µ L 및 400 µ L 25:24:1 페 놀: 클로 프롬: isoamyl 알콜의 추가. 4 ° c.에 2 분 동안 적극적으로 소용돌이
- 여러 번을 반전 하 여 DNA 추출 버퍼와 섞어 추가 200 µ L를 추가 합니다. 20000 x g에서 4 ° C에서 5 분 동안 원심 분리기.
- 깨끗 한 관을 수성 단계 전송의 RNase A/T1, 20 µ g을 추가 하 고 15 분 동안 37 ° C에서 품 어.
- 25:24:1 페 놀: 클로 프롬: isoamyl 알콜, 20000 x g에 4 ° C에서 5 분 동안 회전 급강하의 동등한 볼륨 추가 다음 깨끗 한 관을 수성 단계를 전송 합니다.
- 클로 프롬의 동일한 볼륨을 추가, 여러 번을 반전 하 여 혼합 다음 20000 x g에 4 ° C에서 5 분 동안 회전 깨끗 한 관을 수성 단계를 전송 합니다.
- 100% 에탄올의 두 개의 볼륨 플러스 적어도 2 시간 동안-20 ° C에서 3 M NaOAC (pH 4.3)의 10% 볼륨 촉진 DNA는 다음 20000 x g 와 수집 펠 릿에서 4 ° C에서 5 분에 대 한 스핀.
- 펠 릿 (침전 된 DNA) 냉장된 70% 에탄올 (20000 x g, 5 분, 4 ° C에서 원심 분리기)로 두 번 세척 하 고 50 µ L 10 mm Tris 버퍼 (pH 7.4)에 펠 릿을 일시 중단.
- 사용 라이브러리 준비 키트 ( 재료의 표참조) 제조 업체의 권장 사항을 전체 게놈 시퀀싱 라이브러리를 준비 하 당.
참고: 오류 돌연변이 PCR 증폭 하는 동안 생성을 최소화 하는 PCR 증폭 없이 게놈 라이브러리의 건설을 허용 하기 때문에 재료의 테이블에 에서 나열 된 키트는 것이 좋습니다. 또한, 게놈 라이브러리 준비 하는 동안 허용 하지 않습니다에 구슬 비드 건조 시간 1-2 분을 단축 하 여 완전히 건조. - 라이브러리 준비 하는 동안 매개 변수를 전단에 대 한 집중된 sonicator 사용 ( 재료의 표참조)와 20%, 버스트, 당 200 사이클 175 W, 피크 전력 듀티 계수를 설정 하 고 5.5oC ~ 6 ° C 45에서 주파수 연소 모드 또는 s., 사용을 DNA와 chomatin 시스템 전단 ( 재료의 표참조) 다음 설정을 가진: 펄스 모드, 15 4 ° C에서 50% 진폭 s에와 15-10 분, 20 분의 총 처리 시간에 대 한 시작.
- 주의이 단계에서 사용 되는 유해 물질을 처리 하기 위해 필수적 이다. NaOAC, 에탄올, 25:24:1 페 놀: 클로 프롬: isoamyl 알콜 및 클로 프롬을 처리 하기 위한 적절 한 재료 안전 데이터 시트와 기관의 환경 건강 및 안전 사무실을 참조 하십시오.
- 결과 게놈 라이브러리를 시퀀스. 전체 시퀀싱 읽기 단일 뉴클레오티드 범위에서 해상도 전체 게놈의 적어도 3 시간을 포함 해야 한다. 짝-엔드 시퀀싱 (또는 최신 기술) 것이 좋습니다.
5. 생물 정보학 분석 억압 돌연변이의 id에 대 한
- 보호자 사이 일관 되 게 식별 되는 게놈 변화에 초점을 생물 정보학 분석을 수행 하 고 모든 생물에서 억제 yfm 변종 복제 합니다.
참고: 완전 한 파이프라인 프로세스는 아래에 설명 된, 하지만, 또한, 두 개의 일반 텍스트 BASH 스크립트 파일, fastq_to_vcf.sh 및 vcfprocess.sh, VCF variant 읽기의 처리에서 워크플로의 예를 보여을 보충 자료로 포함 되어 있습니다. 파일 처리 및 교차로의 VCF 파일, 각각. - 트림 짧은-읽기 전단 (https://github.com/jbpease/shear) 다음 명령줄 (모든 다른 옵션 기본값)를 사용 하 여:
shear.py-fq1 $FASTQ1-fq2 $FASTQ2-out1 $OUTFQ1-out2 $OUTFQ2 \
-barcodes1 $BARCODE-플랫폼 TruSeq-trimqual 20시 20분 \
trimpolyat-0-trimambig-filterlength 50-filterunpaired - 지도는 PomBase (ftp://ftp.ebi.ac.uk/pub/databases/pombase/pombe/Chromosome_Dumps/fasta/)에서 얻은 S. pombe 참조 게놈 v2.30 BWA v0.7.1527을 사용 하 여 읽습니다. 다음 명령줄 (모든 다른 옵션 기본값)를 사용 하 여:
bwa mem-8 $GENOME $OUTFQ1 $OUTFQ2 t > $SAM1 - GATK를 통해 맞춤 SAM 파일을 넣어 유용한28 변형 전화 GATK v 3.629, PicardTools v2.5.0 (http://broadinstitute.github.io/picard), 그리고 SAMtools v1.3.130을 사용 하 여 파이프라인. 사용 하 여 다음 명령줄과 매개 변수 (모든 다른 옵션 기본값).
자바-Xmx30g-picard.jar AddOrReplaceReadGroups 입력 항아리 $SAM1 = \
출력 $BAMMARKED RGID = 1 RGLB = lib01 RGPL = illumina = \
RGPU $BARCODE RGSM = $SAMPLENUMBER =
samtools fixmate-O bam $BAMMARKED $BAMFIXED
samtools 정렬-O bam-o $BAMSORTED-T /home/peasejb/tmp $BAMFIXED
samtools 인덱스 $BAMSORTED
자바-Xmx30g-GenomeAnalysisTK.jar-T 자 HaplotypeCaller \
-R $GENOME-$BAMSORTED-genotyping_mode 발견 나 \
-stand_emit_conf 10-stand_call_conf 30-o $VCFRAW - Tabix를 사용 하 여 압축 및 인덱스 VCF 파일:
bgzip $VCFRAW.vcf
tabix $VCFRAW.vcf.gz - 보호자 중 VCF 파일을 비교 및 억압 변종 BCFtools v1.3.127을 사용 하 여 연속 복제. 사용 하 여 다음 명령줄과 매개 변수 (모든 다른 옵션 왼쪽된 기본값).
bcftools isec-n + 1 $VCFPARENTAL1.gz $VCFPARENTAL2.gz $VCFPARENTAL3.gz \
$VCFMUTANT1.gz $VCFMUTANT2.gz $VCFMUTANT3.gz > common_variants.list
참고:이 명령은 나왔고 시퀀스 변종만 첫 번째 돌연변이에 나타나는 것이 어디 "000100", "000010", "000111,"로 모든 3 명의 돌연변이 서만 두 번째 돌연변이로 이진 인코딩된 이진 패턴을 사용 하 여 인코딩된 파일 등. 이러한 파일은 보호자의 각 집합에 대해 생성 된 그리고 VCF 파일을 복제 하는 돌연변이. - UNIX grep 명령을 사용 하 여 각 라인에 추가 하는 파일 이름과 함께 변형 교차로 목록 파일을 컴파일하십시오.
grep "." *.list > all.list - 사용자 지정 Python 스크립트 (variant_characterize.py)를 사용 하 여 현재 GFF3 주석 파일 (ftp://ftp.ebi.ac.uk/pub/databases/pombase/pombe/Chromosome_Dumps/gff3/schizosacchar omyces_pombe.chr.gff3)와 함께 완전 한 변형 목록을 상호 참조 에 단백질 코딩 영역 (동의어 및 비 동의어), 5 '과 3 ' Utr, 및 ncRNA에서 일관 된 SNP 사이트를 식별 합니다.
python3 variant_characterize.py-목록 common_variants.list \
-gff schizosaccharomyces_pombe.chr.gff3 \
-fasta Schizosaccharomyces_pombe입니다. ASM294v2.30.dna.genome.fa \
-패턴 000100-all.list.filter.000100 개
이 스크립트 수정 반복--패턴 출력 파일의 접미사 (-밖으로) 이진 파일을 사용 하 여
패턴: 000010, 000001, 000110, 000011, 000101, 및 000111 - 스프레드시트도 볼 수 있도록 탭으로 구분 된 파일에서 이러한 모든 스크립트 실행의 출력을 결합 합니다. 이체의 주석된 테이블 배경 기준 중 하나 또는 둘 다 돌연변이 strain(s)에 나타나는 포함 합니다. 이진 플래그 필드 나타냅니다 (000100, 000010, 000001), 단일 돌연변이 스트레인에 어느 모양 두 돌연변이 체 긴장 (000011 000101, 000110), 또는 모든 3 개의 돌연변이 체 긴장 (000111).
- 변종 부모의 샘플에서 찾을 수 없습니다 하지만 1, 2, 또는 모든 3 개의 돌연변이 샘플에서 발견의 출력 주석 목록을 분석. 주석을 genomic 위치 및 variant (동의어/비-동의어 코딩 영역에서 3/5' UTR, 비 코딩, 등.)의 클래스를 나타냅니다. 후보 돌연변이의이 목록에서 강하게 관련 후보자의 예는 모든 3 개의 긴장에 일관 되 게 나타나는 비 동의어 코딩 변종 수 있습니다. 또 다른 유형의 강한 후보 가깝게 또는 동일한 유전자에서 나타나는 돌연변이 체 긴장에 다양 한 비 동의어 또는 상 상속 규정 돌연변이의 축적 될 것 이다.
결과
느린-성장 돌연변이 액체 문화에서 phenotypic 회복을 보여
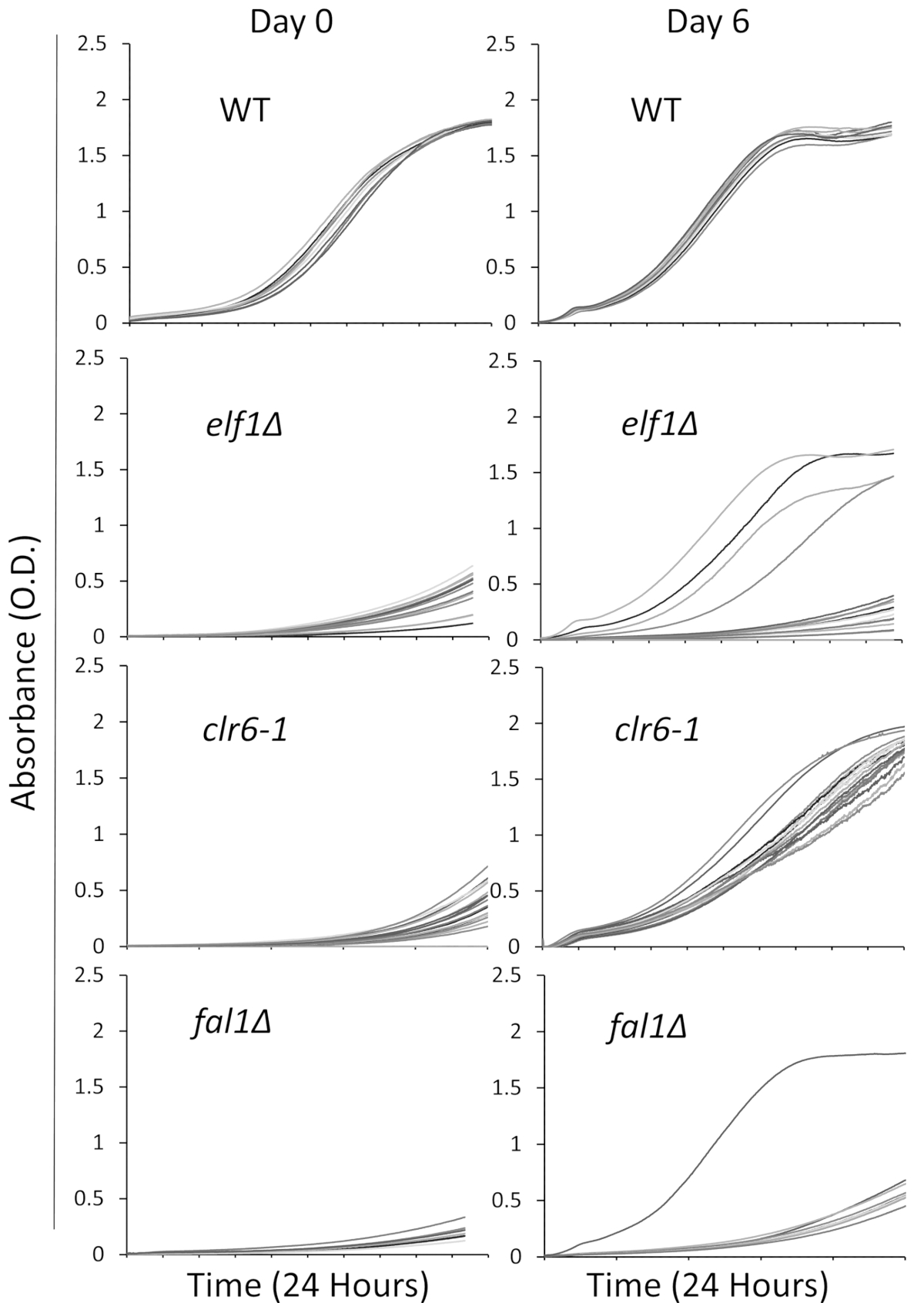
우리 다양 한 아픈, 느린 성장 형으로 생물 학적 경로에 관련 된 3 명의 돌연변이 선택: AAA 가족 ATPase Elf1, 히스톤 Deacetylase Clr6, 그리고 엑손 접합 복잡 한 구성 요소 Fal1. 야생-타입 긴장과 긴장 야생-타입 종자와 backcrossed 했다이 3 개의 유전자의 돌연변이 들고 개별 식민지, 줄무늬 했다 그리고 16 단일 식민지 했다 무작위로 선택 교양으로 96 잘 접시를 사용 하 여 풍부한 액체 미디어에 위에서 설명한. 개별 식민지의 성장 곡선 연속 모니터링 플레이트 리더를 사용 하 여 초기 시간 시점 (0 일)과 6 일 동안 기록 되었다. 예상 했던 대로, 야생-타입 식민지 실험31 (그림 1)를 통해 그들의 성장 곡선에 더 눈에 띄는 변화를 보여줍니다. 특히, elf1∆ 배경 4 식민지와 하나의 fal1∆ 식민지 보여 극적인 변화 성장의 몇 가지 다양 한 수준으로 느린 성장에서 성장에 유사 하거나 야생-타입 식민지의 가까이. 극적으로, 모든 clr6-1 돌연변이 분석 결과31 (그림 1)의 끝으로 빠른 속도로 성장 하 고 일관 된 phenotypic 회복을 보여. 다른 고기 특성, 우리는 느린-성장 하 고 "P 긴장" (또는 보호자 긴장)는 원래 긴장 하 고 긴장 phenotypic 복구 "S 긴장" (또는 억제 된 변종)로 표시를 참조 하십시오. 그림 1 심사 실험의 한 라운드의 예 이며, 총 비 보완 진압 식별 및 다음 대표 결과 순차화를 대표 하지 않습니다를 note 하시기 바랍니다.
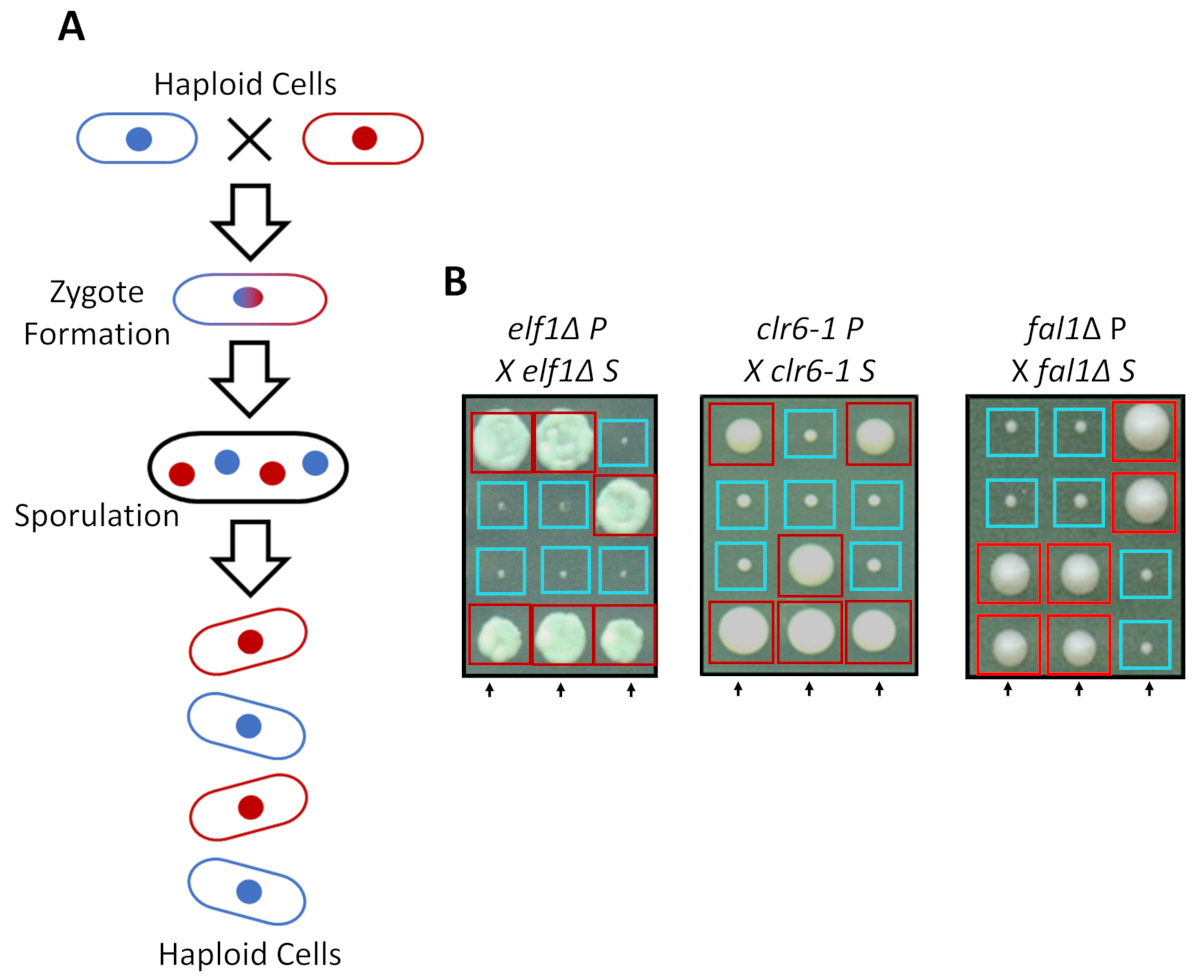
Phenotypic 복구 상속 특성에 기인
S. pombe 리치 미디어에서 단일 하지만 질소 기아에서 보완 짝짓기 종류 친구와 두 개의 단일 긴장으로 성장할 수 있다. 분열 효 모 감수 분열 세포 분열의 2 라운드 뒤 중복의 라운드를 수반 한다. 성적 사이클 2:2 (그림 2A) 클래식 Mendelian 유전학의 규칙에 따라 유전 형질의 분리와 부모의 스트레인의 유전 물질을 운반 하는 4 개의 단일 포자의 형성 결과입니다. 동일한 시간 동안 동일한 접시에 성장, 우리 확인 2:2 분리 때 다시 건너 모든 억압 긴장 (S 변종)과 작은 2 (성장 결함) 및 대형 2 (억압 형)는 그들의 부모의 종자 (P 긴장) 식민지입니다. 억제 된 elf1∆, clr6-1 및 fal1∆ 세포에 대 한 개별 예제는 그림 2B에 표시 됩니다. 우리는 모든 격리 된 S 긴장 그들의 P 긴장 (데이터 표시 되지 않음)의 느린 성장 형 억제 monogenic 유전적 요소를가지고 확인 했습니다.
전체 게놈 시퀀싱 성공적으로 억압 돌연변이 식별
예를 들어, 우리는 elf1∆ S 긴장에서 phenotypic 복구에 대 한 책임 유전 요소를 식별 하 쌍 간 전체 게놈 시퀀싱을 사용. 데이터 분석의 자세한 설명 사용할 수 온라인31이다. 짧게, 우리 두 독립적으로 생성 된 elf1∆ P 긴장의 생물 triplicates와 elf1∆ S 긴장, 각각 포함 된 다른 진압의 5 비 보완 그룹의 생물 학적 중복 고용. 후 우리 생물 정보학 분석 (6.1-10)에서 주석 변종의 목록 우리는 우리의 분석에 관련 된 이체의 특정 클래스 우선. 우리 모든 개별 elf1∆ S 긴장 그들의 부모의 elf1∆ P 긴장 (그림 3 및 보조 테이블 1-4에 비해의 생물 복제의 동일 했다 일관 된 게놈 변경 식별에 초점을 맞춘 ). 우리는 모두 5 가지 elf1∆ S 변종이 포함 rli1 +, SPBPJ4664.02, cue2 + rpl2702 +에서 CD 영역에서 5 nonsynonymous 변경 확인. S-a 1과 S-a 2 다른 아미노산에는 돌연변이 발생 하지만 돌연변이 SPBPJ4664.02를 포함 되어 있습니다. SPBPJ4664.02 긴 유전자 (11,916 뉴클레오티드)의 반복 이므로 돌연변이 시퀀싱 뒤 PCR을 수행 하 여 확인 될 수 없었습니다. S-a 3 삭제 돌연변이 생물 모두 중복에 일치 하는 rli1 에 포함 되어 있습니다. 그러나, 돌연변이 공동 elf1∆ 배경에서 S 형으로 분리를 하지 않았다. 우리는 아미노산으로 S 1에서 cue2 돌연변이 (cue2-1) 396-400 누락 확인. S-b 2 Aspartate31위치 45에서 아미노산 글리신에서 변경 되는 rpl2702 돌연변이 (rpl2702-1)에 포함 되어 있습니다. 아래와 같이 cue2-1 및 rpl2702-1 elf1∆ 진압으로 확인 되었다.
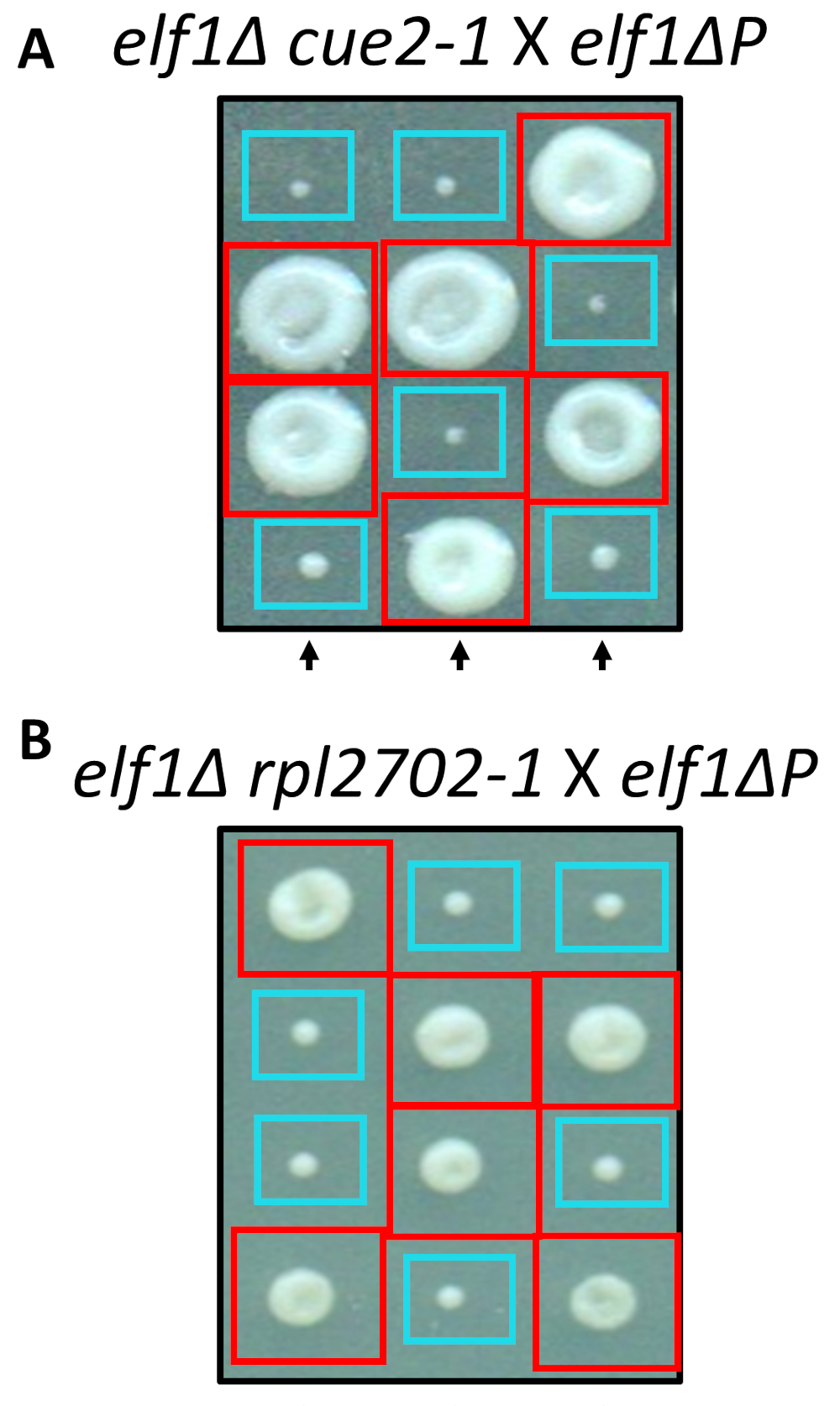
복구 형의 상속 가능성을 확인 하는 확인 된 억압 돌연변이의 유전자 확인
확인 된 nonsynonymous 변경, cue2-1 rpl2702-1, 2 사이트 지시 된 mutagenesis에 대 한 표준 프로토콜을 사용 하 여 실험실에서 재건 되었다. 이중 돌연변이 변종 cue2-1 elf1∆ P 와 rpl2702-1 elf1∆ P 무료 elf1∆ P 스트레인31 (그림 4) 교차 했다. 이 화면을 통해 확인 된 nonsynonymous 돌연변이, elf1∆ P를 억제 하기에 충분 했다, 다음 결과 표시 됩니다 2:2 큰 비율에 작은 식민지에 각 tetrad 4 포자에서 결과에. 실제로, 유전자 교차 확인 된 억압 돌연변이 elf1∆ P 의 느린 성장 형 억제에 성공 하는 및 상속는 보여주었다.

그림 1: Phenotypic 복구 플레이트 리더에서 성장 곡선을 기록 하 여 모니터링할 수 있습니다. 야생-타입 (WT), elf1∆, clr6 1, fal1∆ 의 16 단일 식민지 96 잘 접시에 배치 했다. 성장 곡선 24 h 동안 기록 되었다 그리고 식민지 리치 미디어에 (서) 매일 다시 희석 했다. 성장 결함 0 하루에 24 시간 기간의 끝에 낮은 흡 광도 (마약)에 의해 분명 하다. Phenotypically 복구 된 종자는 비슷한, 또는의 가까이, 성장 곡선을 표시 하는 주 6에서 24 h 동안 야생-타입. Elf1∆, fal1∆의 한 식민지 및 clr6-1 의 모든 식민지의 4 식민지 6 일 후 phenotypic 복구의 다양 한 수준으로 나타났다. 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.
{kind=link}

그림 2: 유전 교차점 phenotypic 복구 단일 상속 대립 유전자에 따라 결정 됩니다 확인할 수 있습니다. (A) 때 분열 효 모 세포 질소 기아, 보완 짝짓기 형식과 두 개의 단일 셀을 복종 된다 zygote tetrad 4 포자의 생성 하기 위해 sporulates를 생성할 수 있습니다. 부모의 유전 재료는 Mendelian 유전학의 규칙에 따라 감수 분열 동안 분리 합니다. (B) Phenotypically 식민지 회복 (레이블된 S 표시 되지 대 한)로 나타난 부모의 genotypes 했다 그들의 무료 부모 식민지로 다시 넘어 (아무 phenotypic 복구, P에 대 한 분류를 보여 주는 부모의). 유전 십자가 2:2 작은 표시 (불 쌍 한 피트 니스) 대형 (복구 된 피트 니스) 식민지 phenotypic 복구 상속 이며 단일 유전 요소에 표시 될 수 있습니다 보여줍니다. 빨간 상자는 식민지 억압 대립 유전자를 운반 하 고 파란색 상자는 부모의 대립 유전자를 운반 하는 식민지. 이 그림에서 Marayati 그 외 여러분, 201831수정 되었습니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.
{kind=link}

그림 3: 유전 요소 phenotypic 복구의 책임을 식별 하기 위해 전체 게놈 시퀀싱 데이터의 분석. 3 생물 두 부모의 "P" 긴장의 복제 (P A와 P B), 그리고 phenotypically 복구 5의 두 생물 학적 복제 전환 "S" 긴장 (S-a 1, a S 2와 S-a 3에서에서 발견 한 P-A; S-B1 및 P b에서 S-B2), 시퀀싱 했다 변이 (예: P-A 대 S-A1, 등)에서 파생 된 부모의 긴장 게놈에 비교 하 여 복구 된 스트레인에 각 변이의 목록으로 구성 했다. 이러한 모든 인덱스도 비교의 전체 게놈에서 감지 된 변이의 총 수는 660 이었다. 총 44 돌연변이 같은 "S" 스트레인의 두 생물 학적 복제에서 발생 하는 유일한 돌연변이 선정 됐다 때 확인 되었다. 44 돌연변이에서 12 돌연변이 삽입/삭제 (INDEL) 또는 비-동의어 변이 했다. 12에서 INDEL 또는 비 동의어 돌연변이, 5 단백질 코딩 순서에서에서 발생 했습니다. 5 변이 잠재적으로 연관 단일 유전자 요소 phenotypically 복구 된 긴장에 대 한 책임: SPBPJ4664.02 에 비 동의어 돌연변이 발견 S a 1과 S-a 2에서 rli1 에 INDEL INDEL cue2에 a S 3에 S 1와 S b 2에서 발견 되는 rpl2702 에 비 동의어 돌연변이에서 발견. 변이 필터링 된 배경에 대 한 자세한 시퀀스 정보 보충 표 1-4에 포함 됩니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.
{kind=link}

그림 4: 전체 게놈 시퀀싱을 통해 식별 하는 진압의 확인. 전체 게놈 시퀀싱 결과 독립적으로 돌연변이 생성 하 고 수행에 의해 확인 되었다 유전 십자가 elf1∆ P 스트레인, 및 elf1∆ elf1∆ cue2-1 변형 건너 phenotypic 복구를 확인 하 rpl2702-1 elf1∆ P 긴장. 3 대표적인 수직까지 표시 됩니다. 빨간 상자는 이중 돌연변이 식민지 (elf1 cue2-1또는 elf1 rpl2702-1); 파란색 상자는 elf1∆ 식민지. 이 그림에서 Marayati 그 외 여러분, 201831수정 되었습니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.
{kind=link}
보충 표 1. 이 테이블을 다운로드 하려면 여기를 클릭 하십시오.
보조 테이블 2 . 이 테이블을 다운로드 하려면 여기를 클릭 하십시오.
추가 표 3 . 이 테이블을 다운로드 하려면 여기를 클릭 하십시오.
추가 표 4 . 이 테이블을 다운로드 하려면 여기를 클릭 하십시오.
코딩 파일 추가 . 파일을 다운로드 하려면 여기를 클릭 하십시오.
토론
여기에 설명 된 프로토콜 나타냅니다 소설과 자발적인 억압 돌연변이 돌연변이 분열 효 모, S. pombe에 400 개 이상의 유전자의 표현 형에 있는 느린 성장 부여의 phenotypic 복구 통해 감지에 대 한 간단한 화면에 함수는 많은의 알 수 없는2,32남아 있습니다. 이전 방법 억압 돌연변이 미생물, 온도 민감한 돌연변이 배경33mutagens21의 사용 또는 온도 변화에의 응용 프로그램을 포함 하 여에 대 한 화면에 다른 접근을 하고있다. 이 프로토콜 phenotypic 복구 추가 환경/화학 간섭 없이 발생 하 고 결국 액체에 사용할 수 있는 리소스를 인계 억제 돌연변이 피트 니스 이용 하이라이트를 표시 하는 반면, 문화입니다. 그것은 elf1∆ 또는 fal1∆ 같은 두 손실의 기능 돌연변이 및 clr6-1, 만큼 돌연변이 점 돌연변이 대 한 효과가 있기 때문에이 화면 우회 진압 또는 상호 작용 진압의 격리를 허용 액체 문화에서 피트 니스 결함을 보여 줍니다.
지금까지 우리가 조사 하는 복구 된 S 긴장 전부 phenotypic 복구의 다양 한 각도 증명 하고있다. 유전자 교차를 통해 감지, 복구 된 표현 형 단일 유전 요소에 기인 이며 상속 (예제 그림2). 이 종종 여러 개의 게놈 loci를 대상 화학 물질 또는 UV 기반 억압 화면에 비해이 방법의 가장 중요 한 장점 중 하나입니다. 16 식민지/스트레인 (약 10%)에서 복구 하는 하나 또는 두 개의 식민지를 관찰 하는 것이 일반적입니다. 내 주. 그러나, 우리는 Rrp6, 핵 관련 exosome 소 단위 적 거의 야생-타입 성장 속도를 복구의 기능의 손실 등 특정 돌연변이 elf1Δ 셀31에서 관찰을 주의 했다. 그것은 가능성이 그 Rrp6의 기능 수만 부분적으로 보상 진압, 달리 다른 돌연변이 테스트, fal1∆, 규제에 그것의 중요 한 기능을 통해 심각한 meiotic 결함을 표시 했다는 등의 함수에 의해 34접합 우리는 yfg 세포 성장에 고유, 비 교체 역할 때 그 대체 억압 심사 방법 같은 문제가 대상이 될 것 이라고 믿습니다.
게놈 시퀀싱을 수행 하기 전에 다시-크로스 플레이트 리더, 유전 배경 선택을 취소 하 고를 가져오는 생물 복제 부모의 긴장에서에서 식별 phenotypically 복구 된 식민지에 최적입니다. 또한, 깊은 전체 게놈 시퀀싱 단일 뉴클레오티드 변화, 대부분의 심사에 대 한 작은 관심의 생물 복제 사이 동일 하지 않은의 수백을 식별 합니다. 예를 들어 우리는 모든 3 개의 염색체 두 elf1Δ P와 5 다른 S 긴장 (그림 3) 사이 걸쳐 660 게놈 변경의 총 발견. 우리 않았다 일반적으로 관찰 하지 각 스트레인의 복제 시퀀스 생물 사이 동일한 돌연변이 새로운 돌연변이 게놈 라이브러리 건설 하기 전에 elf1Δ 세포의 경작 하는 동안 발생할 수 있습니다 또는 임의의 오류가 있을 수 있습니다 제안 라이브러리 건설 및 시퀀싱 하는 동안 도입. 그러므로, 생물 복제에서 일치 하는 돌연변이 고립 시키는 전체 게놈 시퀀싱을 사용 하 여 진압의 성공적인 식별에 중요 한 측면 이다.
우리는 확인 하 고 5 시퀀스 S 긴장에서 CD 영역에 두 개의 진압을 확인. SPBPJ4664.02 에 돌연변이 S a 1과 S-a 2 긴장에서 발견 되었습니다, 하지만 그것은 S-a 1과 S-a 2에서 포함 되지 않은 동일한 유전자에 억압은 서로 (보완 하기 때문에 SPBPJ4664.02 는 유효한 억압 데이터에 표시 되지 않음)입니다. 우리는 또한 rli1 S-A3, 공동 elf1Δ와 backcrossed 때 S 형으로 분리 하지 않았다에 확인 하지 않았다. 또는, 우리는 S-a 1, S-a 2와 S a 3에서 비 코딩 영역에 특정 돌연변이 발견. 이러한 변경 된 비 코딩 게놈 영역 우리의 미래 연구에 해결 될 것입니다 elf1Δ 표현 형을 완화 가능 하다. 연계 분석 결과, 유전자 변이 매핑할 수 년이 걸릴 수 있습니다, 같은 전통적인 방법에 비해 monogenic 요소 S 표현 형 발생을 확인 한 후 2 개월 이내 두 진압 파악. 전체 게놈 시퀀싱 기술의 빠른 발전으로 우리는 낙관적이 방법을 더 효율적 장래에 일관 된 유전자 변이 식별 된다는 것입니다.
요약 하자면,이 프로토콜 성공적으로 액체 문화에서 느린 성장 결함 관심의 모든 유전자에 대 한 억압 돌연변이 식별 하는 단계별 지침을 제공 합니다. 이 분석 결과의 단순 작은 실습 교육의 여러 유전 배경의 대규모 심사에 대 한 수 있습니다. 추가 매일 희석을 수행 하는 액체 처리 로봇을 사용 하 여 프로세스를 자동화 하는 공간이 있다. 미생물의 실험실 조작 성장 액체 문화, 헬스, 본질적으로 선택적 프로세스를 필연적으로 필요 하므로이 프로토콜 박테리아와 같은 다른 큰 인구 모형 유기 체에 광범위 하 게 적용 될 수는 제안 하 고 다른 효 모 종입니다.
공개
저자는이 방법과 아무 경쟁 금융 관심사에 사용 된 계기의 제조 업체에 의해 아무 보증을 선언 합니다.
감사의 말
이 작품에 의해 국립 연구소의 일반 의료 과학, 보조금 1R15GM119105-01 K.Z. 지원 했다 우리는 모든 검토자를 대 한 통찰력 있는 의견 감사합니다. 우리 또한이 원고에 대 한 의견 및 토론에 대 한 제임스 터 커, 앨 리 앤더슨, 엘리자베스 블랙과 글렌 Marrs을 감사합니다.
자료
| Name | Company | Catalog Number | Comments |
| Adenine, Powder | Acros Organics | 147441000 | Use at 75 mg/L to make liquid and solid rich media (YEA) |
| Bacteriology Petri Dish | Corning, Falcon | C351029 | 100 ×15 mm, use to grow strains to single colonies on solid rich media |
| D-Glucose Anhydrous, Powder | Fisher Chemical | D16-1 | Use at 30 g/L to make liquid and solid rich media (YEA) |
| Difco Agar, Granuated | Becton, Dickinson and Co. | 214530 | Use at 20 g/L to make solid rich media (YEA) |
| DNA extraction buffer | 2% Triton X-100, 1% SDS, 100 mM NaCl, 10 mM Tris-Cl (pH 8.0), 1mM Na2-EDTA | ||
| Focused-ultrasonicator | Covaris Inc. | S220 | Alternatively, use QSonica Q800R sonicator/DNA and chromatin shearing system |
| Gen5 Data Collection and Analysis Software | Biotek, Inc. | GEN5SECURE | Or equivalent, must be compatible with the micro-plate reader, use to export data readings from the micro-plate reader |
| Hydrochloric Acid 1N, Liquid | Fisher Chemical | SA48-4 | Use to adjust pH to 5.5 in liquid and solid rich media |
| Liquid Rich Media (liquid YEA) | 30 g/L D-Glucose, 5 g/L Yeast Extract, 75 mg/L Adenine, pH adjusted to 5.5 with 1 M HCl | ||
| Microplate Reader, Synergy H1 Hybrid Multi-Mode Reader | Biotek, Inc. | BTH1MG | Or equivalent, must read visible light at 600 nm wavelength range |
| Rich Media agar plates (YEA plates) | 30 g/L D-Glucose, 5 g/L Yeast Extract, 75 mg/L Adenine, 20 g/L Agar, pH adjusted to 5.5 with 1 M HCl. | ||
| RNase A/T1 mix | Thermo Fisher Scientific | EN0551 | Use according to manufacturer recommendation |
| Sterile Polystyrene Inoculating Loop | Corning, Inc. | OS101 | Or equivalent, use to transfer colonies from agar plates to 96-well plate |
| Sterile workspace and burners | |||
| Tissue Culture Plate, 96-well Optical Flat Bottom with Low Evaporation Lid | Corning, Falcon | C353072 | Or equivalent, must have optical flat bottom for micro-plate ready |
| TruSeq DNA PCR-Free LT/HT Library Prep Kit | Illumina, Inc. | 20015962 | Use to prepare the whole-genome sequencing library |
| Yeast Extract, Powder | Fisher Chemical | BP1422-500 | Use at 5 g/L to make liquid and solid rich media (YEA) |
참고문헌
- McKay, J. K., Latta, R. G. Adaptive population divergence: Markers, QTL and traits. Trends in Ecology and Evolution. 17 (6), 285-291 (2002).
- Wood, V., Harris, M. A., et al. PomBase: A comprehensive online resource for fission yeast. Nucleic Acids Research. 40 (D1), (2012).
- de Visser, J. A. G. M., Cooper, T. F., Elena, S. F. The causes of epistasis. Proceedings of the Royal Society B: Biological Sciences. 278 (1725), 3617-3624 (2011).
- Sailer, Z. R., Harms, M. J. Detecting high-order epistasis in nonlinear genotype-phenotype maps. Genetics. 205 (3), 107911088(2017).
- Kuzmin, E., Costanzo, M., Andrews, B., Boone, C. Synthetic genetic arrays: Automation of yeast genetics. Cold Spring Harbor Protocols. 2016 (4), 326-332 (2016).
- Tong, A. H. Y., Boone, C. Synthetic genetic array analysis in Saccharomyces cerevisiae. Methods in Molecular Biology. 313 (1), 171-192 (2006).
- Dixon, S. J., Costanzo, M., Baryshnikova, A., Andrews, B., Boone, C. Systematic Mapping of Genetic Interaction Networks. Annual Review of Genetics. 43 (1), 601-625 (2009).
- Boone, C., Bussey, H., Andrews, B. J. Exploring genetic interactions and networks with yeast. Nature Reviews Genetics. 8 (6), 437-449 (2007).
- Bai, X., Yang, Z., Jiang, H., Lin, S., Zon, L. I. Genetic suppressor screens in haploids. Methods in Cell Biology. , 129-136 (2011).
- Manson, M. D. Allele-specific suppression as a tool to study protein-protein interactions in bacteria. Methods. 20 (1), 18-34 (2000).
- Motter, A. E., Gulbahce, N., Almaas, E., Barabási, A. L. Predicting synthetic rescues in metabolic networks. Molecular Systems Biology. 4, 168(2008).
- Peterson, R. T., Shaw, S. Y., et al. Chemical suppression of a genetic mutation in a zebrafish model of aortic coarctation. Nature Biotechnology. 22 (5), 595-599 (2004).
- Giorgini, F., Guidetti, P., Nguyen, Q., Bennett, S. C., Muchowski, P. J. A genomic screen in yeast implicates kynurenine 3-monooxygenase as a therapeutic target for Huntington disease. Nature Genetics. 37 (5), 526-531 (2005).
- Forsburg, S. L., Patton, E., et al. The art and design of genetic screens. Nature reviews. Genetics. 2 (9), 659-668 (2001).
- Johnston, D. S. The art and design of genetic screens. Genetics. 3 (March), 176-188 (2002).
- Jorgensen, E. M., Mango, S. E. The art and design of genetic screens: Caenorhabditis elegans. Nature Reviews Genetics. 3 (5), 356-369 (2002).
- Gocke, E., Müller, L. In vivo studies in the mouse to define a threshold for the genotoxicity of EMS and ENU. Mutation Research - Genetic Toxicology and Environmental Mutagenesis. 678 (2), 101-107 (2009).
- Suzuki, T., Hayashi, M., et al. A comparison of the genotoxicity of ethylnitrosourea and ethyl methanesulfonate in lacZ transgenic mice (Muta(TM)Mouse). Mutation Research - Genetic Toxicology and Environmental Mutagenesis. 395 (1), 75-82 (1997).
- Uttam, J., Alberico, C., De Stasio, E. ENU Mutagenesis. International C. elegans Meeting. , (1995).
- Putrament, A., Baranowska, H., Ejchart, A., Prazmo, W. Manganese Mutagenesis in Yeast. Methods in Cell Biology. 20, 25-34 (1978).
- Bose, J. L. Chemical and UV mutagenesis. Methods in Molecular Biology. 1373, 111-115 (2016).
- Ikehata, H., Ono, T. The Mechanisms of UV Mutagenesis. Journal of Radiation Research. 52 (2), 115-125 (2011).
- Shrivastav, N., Li, D., Essigmann, J. M. Chemical biology of mutagenesis and DNA repair: cellular responses to DNA alkylation. Carcinogenesis. 31 (1), 59-70 (2010).
- De Stasio, E. A., Dorman, S. Optimization of ENU mutagenesis of Caenorhabditis elegans. Mutation Research - Genetic Toxicology and Environmental Mutagenesis. 495 (1-2), 81-88 (2001).
- Probst, F. J., Justice, M. J. Mouse mutagenesis with the chemical supermutagen ENU. Methods in Enzymology. 477 (C), 297-312 (2010).
- Bähler, J., Wu, J. Q., et al. Heterologous modules for efficient and versatile PCR-based gene targeting in Schizosaccharomyces pombe. Yeast. 14 (10), 943-951 (1998).
- Li, H., Durbin, R. Fast and accurate short read alignment with Burrows – Wheeler transform. Bioinformatics. 25 (14), 1754-1760 (2009).
- Van der Auwera, G. A., Carneiro, M. O., et al. From fastQ data to high-confidence variant calls: The genome analysis toolkit best practices pipeline. Current Protocols in Bioinformatics. 43, 11.10.1-11.10.33 (2013).
- Mckenna, A., Hanna, M., et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Research. 20, 1297-1303 (2010).
- Li, H., Handsaker, B., et al. The Sequence Alignment/Map format and SAMtools. Bioinformatics. 25 (16), 2078-2079 (2009).
- Marayati, B. F., Drayton, A. L., et al. Loss of Elongation-Like Factor 1 Spontaneously Induces Diverse, RNase H-Related Suppressor Mutations in Schizosaccharomyces pombe. Genetics. 209 (4), 967-981 (2018).
- Harris, M. A., Lock, A., Bähler, J., Oliver, S. G., Wood, V. FYPO: The fission yeast phenotype ontology. Bioinformatics. 29 (13), 1671-1678 (2013).
- Xu, X., Wang, L., Yanagida, M. Whole-Genome Sequencing of Suppressor DNA Mixtures Identifies Pathways That Compensate for Chromosome Segregation Defects in Schizosaccharomyces pombe. G3: Genes|Genomes|Genetics. 8 (3), 1031-1038 (2018).
- Marayati, B. F., Hoskins, V., et al. The fission yeast MTREC and EJC orthologs ensure the maturation of meiotic transcripts during meiosis. RNA. 22 (9), 1349-1359 (2016).
재인쇄 및 허가
JoVE'article의 텍스트 или 그림을 다시 사용하시려면 허가 살펴보기
허가 살펴보기더 많은 기사 탐색
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. 판권 소유