
Method Article
올리고뉴클레오티드의 화학적 트리포스포릴화
요약
올리고뉴클레오티드 5'-트리포스페이트는 필수 생물학적 경로에서 유비쿼터스 성분이며 생명 공학 응용에 점점 더 많이 사용되고 있습니다. 여기에서, 우리는 표준 자동화 합성 기술에 의해 제조된 올리고뉴클레오티드로부터 출발하여 올리고뉴클레오티드 5'-트리포스페이트의 일상적인 합성 및 정제를 위한 기술을 기술한다.
초록
5'-트리포스페이트는 모든 생애에서 발견되는 필수적인 핵산 변형이며, 생명공학 및 합성 생물학에서 올리고뉴클레오티드의 기능적 변형으로 점점 더 많이 사용되고 있다. 올리고뉴클레오티드 5'-트리포스페이트는 역사적으로 효소적 방법에 의해 시험관내에서 제조되어 왔다. 그러나, 이들 방법은 천연 RNA 올리고뉴클레오티드로 제한되고, 강한 서열 선호도를 가지며, 이종 생성물을 생산하는 경향이 있다. 화학적 트리포스포릴화의 새로운 방법은 포스포르아미다이트 화학에 의한 자동화된 올리고뉴클레오티드 합성의 비용 절감과 현재 이용 가능한 다양한 뉴클레오티드 변형을 보완한다. 따라서, 임의의 서열 및 길이의 올리고뉴클레오티드 트리포스페이트의 합성, 및 선택적으로 다양한 비천연 변형을 함유하는, 이제 접근이 가능하다.
이 논문은 살리실 포스포로클로리다이트 및 피로포스페이트를 사용하는 올리고뉴클레오티드의 화학적 트리포스포릴화를 위한 적절한 방법 및 기술을 제시한다. 이 방법은 상업적으로 이용가능한 시약을 사용하고, 표준 고상 합성 방법에 의해 제조된 대부분의 올리고뉴클레오티드와 상용성이며, 탈보호 및 정제 전에 올리고뉴클레오티드 합성에 이어 2시간 내에 완료될 수 있다. 촉매 RNA 효소를 위한 기질로서 화학적으로 트리포스포릴화된 올리고뉴클레오티드의 두 가지 용도가 입증되며, 비생물학적 L-RNA 트리포스페이트로부터의 해머헤드 리보자임의 미러 이미지 버전의 합성을 포함한다.
서문
RNA의 5'-트리포스포릴화된 형태는 생명의 모든 도메인에서의 RNA 전사와 많은 RNA 바이러스의 수명주기 동안 RNA 복제에 의해 생성되기 때문에 생물학에서 유비쿼터스이다. 이들 트리포스페이트는 진핵생물에서 7-메틸구아닐레이트-캡핑된 mRNA의 형성을 위한 기질로서 작용하며, 따라서, 단백질 발현1에 필수적인 역할을 한다. 대조적으로, 삼인산염은 박테리아와 바이러스에 보유된다; 따라서, RNA 5'-트리포스페이트는 진핵생물 2,3,4,5,6,7에서 선천적 면역 반응 조절자에 의해 인식된다. 생물학 밖에서, RNA 리가아제 리보자임의 숙주는 시험관내8에서 5'-트리포스페이트를 사용하도록 진화되었고, 진단 검정 9,10,11,12,13,14,15에 사용하기 위해 변형되었다. 하나의 그러한 리보자임은 작은 L-RNA 올리고뉴클레오티드 5'-트리포스페이트16,17,18로부터의 천연 D-RNA의 비생물학적 "미러-이미지" 거울상이성질체인 L-RNA의 주형-의존성 합성에 사용될 수 있다. 다양한 서열 및 백본 조성의 트리포스포릴화된 올리고뉴클레오티드의 일상적인 준비는 이들 시스템을 조사하는 데 필수적이다.
실험실에서 RNA 5'-트리포스페이트를 제조하는 가장 일반적이고 접근하기 쉬운 방법은 시험관내 전사에 의한 것이다. 그러나, 이러한 방법에 의해 생산된 RNA는 RNA 중합효소의 프로모터 및 기질 요건에 의해 서열 및 크기가 제한된다. T7 RNA 중합효소 및 특수 유도체는 이러한 목적을 위해 사용되는 가장 일반적인 중합효소(19,20,21,22)이다. 이들 효소로 제조된 시험관내 전사된 RNA는 5'-말단 퓨린으로 시작되어야 하며, 처음 10개 뉴클레오티드23,24에서 퓨린에 강하게 편향된다. 더욱이, 염기 또는 골격-변형된 뉴클레오티드의 효소적 혼입은 기껏해야 비효율적이며 천연 중합효소로는 더 자주 불가능하며, 천연 D-RNA 이외의 것으로 구성된 올리고뉴클레오티드 5'-트리포스페이트를 생산할 기회를 제한한다. 또 다른 제한 인자는 시험관내 전사에 의해 생성된 RNA가 실질적인 5'- 및 3'- 이질성을 함유할 수 있고, 20 nt 23,24,25,26,27보다 짧을 때 매우 이질적인 생성물로서 생성된다는 것이다.
대조적으로, 고상 포스포르아미다이트 합성에 의해 제조된 올리고뉴클레오티드의 화학적 트리포스포릴화28,29,30,31,32,33,34,35는 임의의 서열의 3-50 nt 길이의 올리고뉴클레오티드 5'-트리포스페이트를 제조하는데 사용될 수 있다. 추가적으로, 포스포르아미다이트 합성에 접근할 수 있는 방대한 핵산 변형은 5'-트리포스포릴화 14,15,16,17,18,29,36 이전에 올리고뉴클레오티드에 첨가될 수 있다. 이들 방법 중 다수는 모노뉴클레오시드37의 용액상 트리포스포릴화를 위해 Ludwig 및 Eckstein에 의해 개발된 포스피틸화 시약 살리실 포스포로클로리다이트를 사용한다. 이 시약을 사용한 올리고뉴클레오티드의 트리포스포릴화는 올리고뉴클레오티드 5'-히드록실의 포스피틸화, 피로포스페이트와의 반응 및 산화에 의한 트리포스페이트로의 전환, 이어서 고체 지지체로부터 올리고뉴클레오티드의 절단, 탈보호 및 정제를 위한 표준 절차에 의해 고체상 상에서 달성된다(도 1)28.

도 1: 합성 올리고뉴클레오티드의 트리포스포릴화를 위한 반응식. 첫 번째 단계에서, 올리고뉴클레오티드 5ʹ-히드록실은 SalPCl로 포스피틸화된다. 다음 단계에서, 5ʹ-살리실 포스파이트는 TBAP와 반응하여 고리형 메타포스파이트를 형성한 다음, 세 번째 단계에서 산화되어 DNA/RNA 합성기 산화 용액(0.1 M 요오드/피리딘/H2O/THF)에서 순환 5ʹ-트리메타포스페이트를 생성하고, 이는 신속하게 가수분해되어 동일한 용액28,33에서 선형 5ʹ-트리포스페이트를 생성하며, 37. 고체 CPG 지지체로부터의 후속 알칼리성 절단 및 수성 MeNH2/암모니아 중의 올리고뉴클레오티드의 탈보호는 임의의 잔류 고리형 트리메타포스페이트를 선형 형태로 가수분해할 것이다. 약어: SalPCl = 살리실 포스포로클로리다이트; TBAP = 트리부틸암모늄 피로포스페이트; THF = 테트라히드로푸란; CPG = 조절된 기공 유리; MeNH2 = 메틸아민. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
이 방법을 사용하는 초기 발표 된 보고서는 종종 불량한 수율과 바람직하지 않은 부제품 28,37,38로 고통 받았지만 무수 조건을주의 깊게 유지하는 것이 일상적으로 높은 수율을 얻는 데 필요한 모든 것입니다. 이는 시약을 신중하게 준비하고 표준 플라스틱 성분으로 조립 된 간단한 반응 장치를 사용하여 달성 할 수 있습니다. 여기에서, 우리는 시약의 제조, 반응 챔버의 조립, 트리포스포릴화 반응, 및 트리포스포릴화된 올리고뉴클레오티드의 후속 탈보호 및 정제를 포함하는 올리고뉴클레오티드의 화학적 트리포스포릴화를 위한 적절한 단계를 입증한다. 또한 천연 D-RNA 및 비생물적 L-RNA 골격을 갖는 더 큰 핵산 생성물의 합성을 위한 리가아제 리보자임(ligase ribozymes)을 위한 기질로서 5'-트리포스포릴화된 올리고뉴클레오티드의 대표적인 용도가 포함된다.
프로토콜
1. 고체 지지체 상의 5'-히드록실 올리고뉴클레오티드의 자동화된 고체상 합성
- 표적 올리고뉴클레오티드 조성 및 기기 지침에 따라 시약 및 포스포르아미다이트로 자동화된 DNA/RNA 합성기를 준비하십시오.
- 고체-지지체를 포함하는 합성 컬럼을 합성기 상에 로딩하고 합성기 기기 프로토콜에 따라 올리고뉴클레오티드를 합성한다.
참고: 트리포스포릴화 절차는 1 μmole 규모로 제조된 올리고뉴클레오티드에 대해 최적화되었다. - 5'-디메톡시트리틸 보호기를 제거하여 이전 단계에서 합성된 올리고뉴클레오티드의 일부로서 고체 담지 올리고뉴클레오티드 5'-히드록실을 수득하거나, 합성기 기기 프로토콜에 따라 말단 탈취 단계를 수행함으로써 수득한다.
- 고체 지지체 상의 5'-히드록실 올리고뉴클레오티드를 함유하는 컬럼을 합성기로부터 제거하고, 잔류 용매를 제거하기 위해 10분 동안 집진공 하에 건조시키고, 트리포스포릴화를 위한 물질이 제조되면 트리포스포릴화(섹션 3 및 4)를 진행한다(섹션 2).
참고: 즉시 사용하지 않을 경우, 건조된 컬럼은 -20°C에서 건조제가 있는 밀봉된 플라스틱 용기에 정상적인 분위기 하에서 저장될 수 있다. 이 단계에서 컬럼은 섹션 3에서 트리포스포릴화 전에 완전히 건조되기 때문에 더 이상의 건조가 필요하지 않다.
2. 삼인산화를 위한 재료 준비
- 압력 조절 가능한 건조 아르곤 소스를 적어도 두 줄의 가스 매니폴드에 부착하고 버블러에 연결하십시오. 반응 장치와의 연결을 용이하게 하기 위해 1 mL 플라스틱 주사기에서 라인이 종결되도록 하십시오.
- 1mL 플라스틱 주사기, 3방향 폴리프로필렌 스토콕, 비코링 니들, 1.5mL 폴리프로필렌 튜브 및 소형 금속 주걱을 포함하여 트리포스포릴화 중에 사용할 장비를 수집합니다. 밀봉 된 용기 또는 사용 전에 적어도 1 일 동안 실온에서 건조제가있는 데시케이터에 보관하십시오.
- 건조 트랩(밀봉된 멤브레인 패킷에 4 Å 분자체 체)으로 건조된 30 mL 유리 병에 무수 1,4-디옥산, 3:1 디옥산:피리딘, N,N-디메틸포름아미드(DMF) 및 아세토니트릴(ACN)을 각각 30 mL씩 준비하십시오. 고무 셉타로 밀봉하고 건조제가있는 데시케이터에 보관하십시오.
- 고체 2-클로로-4H-1,3,2-벤조디옥사포스포린-4-온(살리실 포스포로클로리다이트, SalPCl)을 4°C에서 건조제가 있는 밀봉된 용기 안에 원래의 용기에 보관하십시오. 항상 사용 사이에 아르곤으로 컨테이너를 플러시하십시오.
- 트리포스포릴화 반응의 적어도 5일 전에 트리부틸암모늄 피로포스페이트(TBAP) 용액을 준비한다:
- 건조된 30-mL 유리 병에 고체 TBAP 1-5 g을 칭량하고, TBAP의 g 당 1 mL의 DMF 및 0.5 mL의 트리부틸아민에 용해시킨다.
- 세 개의 건조 트랩을 추가하고 병을 아르곤 아래의 고무 격막으로 밀봉하고 아르곤으로 30 분 동안 거품을 일으켜 탈기하십시오.
- 분자체가 모든 미량의 물을 흡수할 수 있도록 4°C에서 5일 동안 건조제로 밀봉된 항아리 안에 보관하십시오. 항아리를 -20°C에 보관하고 6개월 후에 신선하게 준비한다.
3. 조립 및 삼인산화 장치 사용
- 합성 컬럼이 -20°C에서 보관으로부터 회수되는 경우 실온으로 가온되도록 허용한다.
- 그림 2에 표시된 반응 챔버를 조립합니다.
- 전실을 준비하십시오 : 건조한 1 mL 주사기에서 플런저를 제거하고, 가위 또는 면도날을 사용하여 주사기의 상단을 잘라 내고, 주사기를 합성 컬럼에 부착하십시오. 세 방향 스톱콕을 주사기 상단에 부착하고 스톱콕의 측면 입구를 버블러가있는 건조 아르곤 소스에 부착하여 스톱콕의 상단 입구가 시약 주입 포트가되도록하십시오.
- 이 장치를 클램프가있는 스탠드에 고정하고 왁스 씰링 필름으로 모든 업스트림 조인트를 밀봉하십시오. 주입구가 닫히고 장치가 아르곤 소스에 개방되도록 스톱콕을 조정하십시오. 버블러를 닫고, 아르곤을 저압 (<10 psi)에서 5분 동안 반응 챔버를 통해 스트림하도록 허용한다.
참고: 다중 반응 챔버는 트리포스포릴레이트 2-4 올리고뉴클레오티드와 병렬로 설정될 수 있다. 그러나 매니폴드의 한 줄은 시약 병에 아르곤을 공급하기 위해 예약해야합니다. - 버블러를 다시 열고 주사기를 폐기물 주사기가 될 합성 컬럼의 바닥에 부착하십시오. 상기 폐기물 주사기를 이용하여 컬럼을 통해 아르곤을 반복적으로 당기는 단계; 그런 다음 플런저를 완전히 밀어 넣은 상태에서 주사기를 다시 부착하십시오.
참고: 시약을 적재하지 않는 한, 도 2A에 도시된 바와 같이, 주입구가 닫히고 장치가 아르곤 소스에 개방되도록 스톱콕을 설정해야 한다. 유사하게, 폐주사기는 시약을 활발히 제거하지 않는 한 왁스 밀봉 필름으로 밀봉된 합성 컬럼에 부착되고 접합되어야 한다.
- 시약 또는 용매를 추가하려면,
- 마른 아르곤 소스에 바늘을 부착하고 시약 또는 용매 병 중격에 넣고 바늘을 병 내용물에 담그지 않도록주의하십시오.
- 바늘로 건조한 주사기를 조립하고 병 내용물에 담그지 않고 시약 또는 용매 병 중격에 삽입하십시오. 주사기를 아르곤으로 채우고 중격에서 바늘을 꺼내 아르곤을 배출하십시오. 주사기를 아르곤으로 채우고 다시 추방하십시오. 그런 다음 아르곤 압력 하에서 필요한 부피의 용매 또는 시약으로 주사기를 채 웁니다.
- 아르곤 소스가 닫히고 주입 포트가 열리도록 장치의 스톱콕을 조정합니다(그림 2B). 채워진 주사기와 바늘을 소스 병에서 신속하게 제거하고 바늘의 측면 또는 끝에 붙어있는 용매를 닦아 낸 다음 바늘을 주사 포트에 삽입하십시오. 시약을 장치의 전실로 배출하고, 바늘을 제거하고, 주입 포트를 닫고, 장치를 아르곤 소스로 다시 엽니 다.
- 폐기물 주사기를 사용하여 합성 컬럼을 통해 전실로부터 액체를 부드럽게 끌어당겨 모든 액체가 이제 폐주사기 내에 유지되도록 한다. 이제 천천히 용액을 합성 컬럼으로 밀어 넣어 컬럼에 가스 버블이 없는지 확인하십시오. 혼합하거나 교반하려면 폐주사기로 용액을 컬럼 위로 부드럽게 위아래로 당깁니다.
참고: 항상 액체를 반응 챔버를 통해 천천히 부드럽게 움직여 씰이 깨지지 않도록 하여 공기가 장치에 들어갈 수 있도록 합니다.
- 컬럼으로부터 시약 또는 용매를 제거하려면:
- 용액을 천천히 폐주사기로 당깁니다. 용액의 벌크가 폐주사기 내로 통과된 후, 아르곤을 통해 당겨 컬럼으로부터 잔여 용매를 플러시한다.
- 폐기물 주사기 조인트 주변의 왁스 밀봉 필름을 제거한 다음 주사기를 제거하고 폐기물 용액을 폐기하십시오. 폐기물 주사기를 새로운 건식 주사기로 교체하고 왁스 밀봉 필름으로 조인트를 다시 밀봉하십시오.

도 2: 트리포스포릴화 장치. 혼합 또는 반응 동안, 장치(A)는 아르곤 공급원(i)을 개방하고, 삼방향 스톱콕(ii)을 조정함으로써 공기에 폐쇄된다. 시약은 폐주사기(v)에 의해 전실(iii)로부터 합성 컬럼(iv)으로 끌어당겨진다. 시약은 모든 액체를 폐주사기 (v)로 끌어당기고 폐기함으로써 제거된다. 시약(B)을 로딩할 때, 삼방향 스톱콕(ii)은 대기로 개방되고, 시약은 주사기 및 바늘(vi)에 의해 전실(iii) 내로 로딩된다. (c) 시약 혼합 및 반응을 위한 (A)에서와 같이 조립된 장치 세트의 사진. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
4. 합성 5'-히드록실 올리고뉴클레오티드의 컬럼 상의 트리포스포릴화
- -20°C의 저장소에서 SalPCl 및 TBAP 용액을 제거하고 사용 전에 실온으로 가온하십시오.
- 단계 3.3.1-3.3.3에 따라 200μL의 피리딘/다이옥산을 전실에 첨가한다. 그러나, 단계 4.4까지 합성 컬럼 상에 용매를 로딩하지 않는다.
- 건조 금속 주걱을 사용하여 6-12 mg의 SalPCl을 무겁게 1.5 mL 마이크로원심분리 튜브에 넣고, 마이크로원심분리 튜브 내에서 용매를 상하로 부드럽게 시닝하여 다이옥산 100 μL에 용해시킨다.
- 용해된 SalPCl을 전실에 첨가하고, 이를 합성 컬럼 상에 로딩하고, 이어서 단계 3.3. 15 분 동안 반응시키고 5 분마다 용액을 교반하십시오. 3.4단계에 따라 SalPCl 솔루션을 제거하고 버립니다.
참고 : SalPCl은 과량으로 과량 첨가되며 준비 및 반응 챔버로 로딩하는 동안 흡수 된 물을 제거합니다. 그러나, 단계 4.5 및 4.6 동안 임의의 수분의 도입은 최종 올리고뉴클레오티드 5'-트리포스페이트 수율을 손상시킬 것이다. - 250 μL의 TBAP 용액을 전실에 첨가하고, 이를 단계 3.3에 이어 합성 컬럼 상에 로딩한다. 20 분 동안 반응하게하고 5 분마다 동요시킵니다. 3.4단계에 따라 TBAP 솔루션을 제거하고 폐기합니다.
- 컬럼을 0.5 mL의 DMF로 세척한 다음, 0.5 mL의 ACN으로 세척하고, 단계 3.3 및 3.4에 따라 각각 첨가한 후에 용매를 제거하였다.
- 250 μL의 산화제 용액 (테트라 하이드로 퓨란 (THF) / 피리딘 / 물 중 0.1 M 요오드, 88 : 10 : 2)을 전실에 넣고 3.3 단계에 따라 합성 컬럼에 로딩하십시오. 30 분 동안 반응하게하고 10 분마다 동요시킵니다. 단계 3.4에 따라 산화제 용액을 제거하고 폐기한다.
- 컬럼을 0.5 mL의 ACN으로 세척하고, 단계 3.3 및 3.4에 따라 제거한다.
- 반응 장치를 분해한다. 합성 컬럼을 5 mL의 ACN으로 세척하고 컬럼을 건조시킨다.
5. 고체 지지대, 탈보호 및 정제로부터의 절단
- 건조된 고체 지지체 수지를 합성 컬럼으로부터 제거하고, 이를 실리콘 o-링이 있는 1.5 mL 폴리프로필렌 스크류 캡 밀봉가능한 튜브로 옮긴다.
- 수지를 28%-30% 암모니아수 및 40% 메틸아민(AMA) 수용액의 1:1 혼합물 중 1 mL에 현탁시키고 튜브를 단단히 밀봉한다. 65°C에서 10분 동안 온화한 역전(39)에 의한 간헐적 혼합으로 인큐베이션한다. 40 nt보다 긴 올리고뉴클레오티드에 대해 실온에서 2 h 동안 더 온화한 처리를 사용하십시오.
주의: AMA 용액을 가열하면 튜브가 고압 상태로 유지됩니다. 튜브가 단단히 밀봉되지 않았거나 암모니아 호환 가능 (실리콘) O 링을 사용하지 않는 경우, 튜브는 가스 또는 누출 용매를 배출하여 안전 또는 최종 제품 수율을 손상시킬 수 있습니다. 뜨거운 AMA 용액이 가스를 격렬하게 진화시킬 수 있으므로 주위 온도 이상에서는 튜브를 열지 마십시오. - 튜브를 얼음 위에 식히고 간단히 원심분리하십시오 (10 초 동안 6,000-12,000 × g ). 수지로부터 상층액을 제거하고, 0.2 μm 필터가 장착된 주사기를 통해 여과하고, 새로운 멸균 폴리프로필렌 튜브로 옮긴다. 암모니아 중화 화학 트랩이 장착된 진공 원심분리기를 사용하여 용액을 증발시켜 건조시킨다.
참고: 합성 올리고뉴클레오티드가 2'-실릴 보호기를 갖는 RNA 뉴클레오티드를 포함하지 않는 경우, 단계 5.8로 건너뜁니다. - 건조된 물질을 THF에 1 mL의 1 M 테트라부틸암모늄플루오라이드(TBAF)에 용해시키고, 55°C로 가열하고, 필요에 따라 진탕하여 올리고뉴클레오티드를 완전히 용해시키고, 실온에서 16-24시간 동안40,41시간 동안 인큐베이션함으로써 실릴 보호기를 제거한다.
- TBAF 용액을 1 mL의 1 M 트리스 완충액, pH 7.5로 켄칭하고, 진공 원심분리기를 사용하여 THF를 제거하였다.
- 제조업체의 지침에 따라 일회용 크기 배제 컬럼을 사용하여 TBAF 염을 제거하십시오. 15 nt보다 짧은 올리고뉴클레오티드의 경우, 분획에서 용출액을 수집하여 생성물의 용출을 확인하고, UV-Vis 분광광도계 상에서 260 nm에서의 흡광도로 주요 생성물 분획을 확인한다.
- 탈보호된 올리고뉴클레오티드를 15 nt 미만일 경우 동결건조 또는 진공 원심분리에 의해, 또는 15 nt를 초과하는 경우 에탄올 침전에 의해 농축한다.
- 올리고뉴클레오티드 크기, 겔 플레이트 및 스탠드에 적합한 프로토콜에 따라 19:1 모노:비스 아크릴아미드 스톡을 사용하여 10%-20% 폴리아크릴아미드/8 M 우레아/1x TBE 겔을 준비하십시오. 저장고에 1x TBE로 겔 스탠드에서 겔 플레이트를 조립하고 적어도 30분 동안 35W(또는 겔 플레이트 포맷에 적합함)에서 미리 실행한다.
- 고체 올리고뉴클레오티드를 우레아 겔 로딩 완충액 (8 M 우레아, 10% 수크로오스, 50 mM 트리스, pH 8, 1 mM EDTA와 브로모페놀 및 크실렌 시아놀 실행 염료)에 용해시키고 80°C로 가열한다. 폴리아크릴아미드 겔에 하중을 가하고 25-35 W에서 1-2 h 동안 실행한다 (또는 겔 플레이트 포맷 및 올리고뉴클레오티드 크기에 적합함).
참고: 브로모페놀 블루 또는 자일렌 시아놀은 에탄올 침전 없이 용출된 올리고뉴클레오티드로부터 이들 염료를 제거하기 어렵기 때문에 생성물과 함께 공동 이동되는 경우 15 nt보다 짧은 올리고뉴클레오티드에 대한 겔 로딩 완충액에서 제외되어야 한다. - 겔 전기영동이 완료되면, 겔 스탠드로부터 겔 플레이트를 제거하고, 겔 플레이트를 분해하고, 겔을 폴리비닐클로라이드 필름으로 감싸라. 254nm 자외선으로 백섀도잉하여 제품 밴드를 식별하고 면도날을 사용하여 주요 제품 밴드를 소비합니다.
- 분쇄 및 담그는 방법42에 의해 적출된 겔로부터 올리고뉴클레오티드를 추출한다.
- 적출된 겔을 플라스틱 주사기를 통해 또는 기계적으로 압출하여 분쇄한다.
- 15 nt보다 긴 올리고뉴클레오티드의 경우:
- 3x 부피의 분쇄 및 완충액 (300 mM NaCl; 10 mM 트리스, pH 8; 1 mM EDTA)을 교반 또는 진탕으로 적어도 12 시간 동안 담그십시오.
- 용액을 0.2 μm 필터가 장착된 주사기를 통해 통과시켜 고체 겔 조각을 제거하고 에탄올 침전에 의해 올리고뉴클레오티드를 농축시킨다.
- 15 nt보다 짧은 올리고뉴클레오티드의 경우:
- 3x 부피의 뉴클레아제가없는 물을 교반 또는 흔들면서 적어도 12 시간 동안 묽으십시오.
- 용액을 0.2 μm 필터가 장착된 주사기를 통해 통과시켜 고체 겔 조각을 제거하고 동결건조에 의해 농축한다.
- 단계 5.6에서와 같이 일회용 크기 배제 컬럼을 사용하여 잔류 염 및 용질을 제거한다. 동결 건조에 의해 집중하십시오.
- 정제된 트리포스포릴화 올리고뉴클레오티드를 -20°C에서 TE 완충액 (10 mM 트리스, pH 8; 1 mM EDTA) 또는 유사한 올리고뉴클레오티드 저장 완충액에 저장한다.
- UV-Vis 분광광도계를 이용하여 260 nm에서 흡광도를 측정하여 올리고뉴클레오티드의 농도를 결정한다.
참고: 올리고뉴클레오티드의 추정된 흡광 계수는 그것의 5'-인산화 상태에 의해 영향을 받아서는 안 되며, 올리고뉴클레오티드 흡광 계수 계산기를 사용하여 그의 서열로부터 계산될 수 있다. - 질량 분광법에 의한 삼인산화를 검증한다. 5'-히드록실 올리고뉴클레오티드의 질량에 비해 +239.94 Da의 예상 질량을 찾아본다.
참고: 매트릭스 보조 레이저 탈착/이온화 또는 전기 분무 이온화 질량 분광법(각각 MALDI-MS 또는 ESI-MS)은 핵산에 최적화된 프로토콜을 사용할 때 올리고뉴클레오티드의 삼인산화 상태를 식별하는 데 적합합니다. 그러나 ESI-MS는 이온 단편화 속도가 낮기 때문에보다 일관된 결과를 제공합니다. 대표적인 상업 서비스는 자료 표에 나와 있습니다.
- UV-Vis 분광광도계를 이용하여 260 nm에서 흡광도를 측정하여 올리고뉴클레오티드의 농도를 결정한다.
6. 리보자임 자기복제를 위한 기질로서의 트리포스포릴화 올리고뉴클레오티드
주의: 32P는 방사성 동위원소이며 다음 단계는 실험실에서 방사성 물질 작업을 위한 표준 안전 프로토콜을 사용하고 관련 환경 보건 및 안전 부서에서 방사성 물질 사용을 인증받은 연구원이 수행해야 합니다. 대안으로서, 자기-복제 리보자임 기질 A는 단계 7.9에서와 같이 5'-플루오레세인 라벨(14 )을 사용하여 합성적으로 제조되고 형광으로 이미징될 수 있다.
- 용액 A를 자가복제체 리보자임 E 및 5'-32P-표지된 RNA 기질 A의 혼합물로서14 내지 0.30 μM 및 30 μM의 농도로 각각 준비한다. 용액 B를 15 μM 트리포스포릴화 RNA 기질 B로 전사 및 B-합성하고, 75 mM EPPS 완충액, pH 8.5 및 37.5MgCl2에서 각각 위와 같이 시험관내 전사14 또는 화학적 트리인산화에 의해 제조한다. 두 용액을 모두 42 °C로 가져 오십시오.
참고: 모든 RNA 성분은 물질 표에 열거되어 있습니다. - 자기 복제를 개시하기 위해, 5 μL의 용액 A를 10 μL의 용액 B와 신속하게 혼합하고, 0.1 μM E, 10 μM A, 10 μM B, 25 mMMgCl2 및 50 mM EPPS 완충액, pH 8.5의 최종 농도로 전사 또는 B-합성하였다. 반응 혼합물을 42°C에서 인큐베이션한다.
- 일정한 간격으로 0.5 μL 분취량을 취하여 9.5 μL의 포름아미드 겔 로딩 완충액 (95% 포름아미드; 10 mM EDTA, pH 8)에서 켄칭한다.
- 겔 플레이트 및 스탠드에 대한 적절한 프로토콜에 따라 분석 폴리아크릴아미드/8 M 우레아/1x TBE 겔을 준비합니다. 캐스트 젤과 플레이트를 겔 스탠드에 조립하고, 저장소를 1x TBE로 채우고, 40W(또는 다른 겔 플레이트 및 스탠드에 적합한)에서 30분 동안 프리런합니다.
- 켄칭된 반응 샘플을 80°C로 가열하고, 각 웰에 샘플 5 μL를 로딩하고, 대략 40분 동안 40 W에서 겔을 실행한다(또는 상이한 겔 플레이트 및 스탠드에 적합함).
- 겔 스탠드에서 겔 플레이트를 제거하고, 분해하고, 겔을 폴리염화비닐 필름으로 감싸십시오. 겔을 형광체 스크린으로 덮고 1 h 동안 노출시킨다 (또는 32Pcpm에 적합하게); 형광/인광 젤 스캐너를 사용하여 화면을 스캔합니다.
- 겔 이미지 정량 소프트웨어를 사용하여 반응 수율을 정량화하십시오.
- 분석 도구 상자를 사용하여 젤 이미지를 열고 분석을 선택하| 모양 정의, 사각형 모양| 영역을 선택하고 각 시간 및 두 반응 모두에 대해 미반응 A 및 생성물 E에 해당하는 밴드 주위에 동일한 크기의 사각형을 그립니다.
- 분석 | 선택 배경 빼기, 사각형 모양| 영역을 선택하고 젤 이미지의 빈 부분에 같은 크기의 사각형을 그립니다. 모든 사각형에 대한 배경 빼기 방법을 이미지 사각형으로 변경합니다.
- 2 영역 창| 창을 선택한 다음 편집을 선택합| Excel로 내보내 정량화된 밴드 픽셀 볼륨을 스프레드시트 파일로 내보냅니다.
- 제품 E의 농도 대 시간을 플로팅하고 통계 데이터 피팅 소프트웨어를 사용하여 데이터를 물류 성장 Eq (1)에 맞춥니다.
[E] = (1)
(1)
여기서 a는 반응 최대 범위이고, b는 시그모이디시티의 정도이고, c는 지수 성장률이다.- 수출 된 스프레드 시트에서 제품 E 부피를 기질 A와 생성물 E 부피의 합으로 나누어 각 시간과 두 반응 모두에 대한 제품의 분수 수율을 결정합니다. 기질 A의 초기 농도 (10 μM)를 곱하여 시간의 함수로서 생성물 E의 수율을 결정한다.
- 통계 데이터 피팅 소프트웨어에서 파일 |을 선택하십시오. 새로운 | 새 프로젝트 파일, 새 테이블 및 그래프에서 XY를 선택하고 만들기를 클릭합니다. 인접한 컬럼에서 두 반응 모두에 대한 반응 시간 및 생성물 E 수율을 입력하고, 그에 상응하게 컬럼을 라벨링한다(예를 들어, "시간", "전사된 B", "합성 B").
- | 삽입을 선택하십시오. 새 분석에서 XY 분석을 선택하| 비선형 회귀(커브 맞춤)를 클릭한 다음 확인을 클릭합니다. 성장 곡선 | 선택 물류 성장 및 클릭 확인; 다른 매개 변수를 조정하지 마십시오. 결과에서 적합 매개변수와 신뢰 구간을 관찰하고 그래프 아래의 데이터 점과 적합 곡선의 플롯을 관찰합니다.
7. L-RNA의 교차 키랄 복사
- 50 mM 트리스, pH 8.3의 10 μL에서 20 μM D-RNA 27.3t 교차 키랄 중합효소, 2 μM 5'-플루오레세인 표지된 L-RNA 프라이머, 4 μM 비오티닐화 L-RNA 해머헤드 주형 및 20 μM L-RNA pppCUG, pppAUG, pppAGG 및 pppCGC를 각각 함유하는 RNA 용액 10 μL를 제조하였다. RNA를 1분 동안 90°C로 가열하고 PCR 열사이클러에서 0.2°C/s에서 23°C로 냉각하여 어닐링한다. 중합효소, 프라이머 및 주형에 대한 자세한 내용은 재료 표를 참조하십시오.
- RNA 용액을 17°C에서 인큐베이션하고, 10 μL의 2x 시작 완충액 (400 mMMgCl2, 500 mM NaCl, 및 50 mM 트리스, pH 8.3)을 첨가하여 반응을 시작한다. 반응의 모든 성분의 최종 농도가 10 μM 중합효소, 1 μM 프라이머, 2 μM 주형, 10 μM 각 트리뉴클레오티드 5'-트리포스페이트, 200 mMMgCl2, 250 mM NaCl, 및 50 mM 트리스, pH 8.3인지 확인한다.
- 반응이 진행됨에 따라, 10 μL 분취량을 취하고, 5 μL의 0.5 M EDTA, pH 8로 켄칭한다.
- 각각의 켄칭된 반응 샘플에, 0.05% 중성 세제와 함께 TE 완충액 중의 1 M NaCl 중 10 μL에 현탁된 스트렙타비딘 코팅된 자성 비드 (20 pmol 비오틴-올리고뉴클레오티드 결합 능력)의 0.1 mg을 첨가하고, 비오티닐화 주형을 통해 삼량체-연장된 프라이머를 포획하고, 진탕하면서 실온에서 30분 동안 인큐베이션한다.
- 비드-포획 자석 상에 비드를 포획하고, 액체를 제거 및 버리고, 50 μL의 세척 용액(0.05% 중성 세제를 사용한 TE 중의 250 mM NaCl)을 첨가한다. 구슬을 섞어서 다시 잡은 다음 세척 용액을 제거하십시오. 한 번 더 반복하십시오.
- 비드로부터 연장된 프라이머를 용출시키기 위해, 50 μL의 용출 용액 (0.05% 중성 세제를 사용한 25 mM NaOH)을 첨가하고, 비드를 혼합한다. 비드를 다시 포획하고, 용출 용액을 제거하고, 100 mM 트리스 (pH 7.5)로 켄칭하고, 에탄올로 침전시킨다.
- 겔 플레이트 및 스탠드에 적합한 프로토콜에 따라 분석 폴리아크릴아미드/8 M 우레아/1x TBE 겔을 준비합니다. 캐스트 젤과 플레이트를 겔 스탠드에 조립하고, 저장소를 1x TBE로 채우고, 40W(또는 다른 겔 플레이트 및 스탠드에 적합한)에서 30분 동안 프리런합니다.
- 침전된 RNA를 10 μL의 포름아미드 겔 로딩 완충액에 용해시키고, 0.5 μM의 미반응 최종 표지된 프라이머를 포름아미드 겔 로딩 완충액에 제조하였다. 샘플을 80°C로 가열하고, 샘플을 각 웰에 5 μL씩 로딩하고, 대략 40분 동안 40 W에서 겔을 실행한다(또는 상이한 겔 플레이트 및 스탠드에 적합함).
- 스탠드에서 겔 플레이트를 분리하고 형광/인광 겔 스캐너를 사용하여 스캔하여 교차 키랄 L-RNA 확장 제품을 시각화합니다.
결과
올리고뉴클레오티드는 포스포르아미다이트와 자동화된 DNA/RNA 합성기에 적합한 표준 프로토콜을 사용하여 합성되어야 하며, 생성물 올리고뉴클레오티드는 원래의 플라스틱 합성 컬럼의 고체 지지체로부터 분리되고, 5ʹ-말단 디메톡시트리틸 그룹을 제거하여 자유 5ʹ-히드록실을 생성한다(섹션 1). 이 데모에 사용된 모든 올리고뉴클레오티드는 고체 지지체로서 1,000 Å 조절된 공극 유리(CPG) 수지를 사용하여 제조되고 0.2 또는 1 μmole 규모로 진행되었다. 합성기 컬럼, 수지, 시약 및 포스포르아미다이트의 대표적인 예가 물질의 표에 제공된다. 더 큰 규모의 반응의 경우, 후속 단계에서 사용되는 부피와 시간을 조정해야 할 수도 있습니다.
상기 트리포스포릴화 반응은 표 of Materials에 열거된 표준적이고 상업적으로 이용가능한 성분들을 사용하여 맞춤형 반응 챔버(도 2, 섹션 3)에서 컬럼 상에서 수행되며, 도 1(섹션 4)28에 예시된 반응식을 따른다. 삼인산화 동안 조건을 엄격하게 무수로 유지하고, 모든 용매 및 시약을 미리 분자 체를 통해 준비하고 사용 전에 완전히 건조시키는 것이 중요합니다 (섹션 2). 트리포스포릴화는 전형적으로 일어나기까지 2시간이 걸리고, 그 후, 세척 및 건조된 컬럼은 표준 올리고뉴클레오티드 탈보호 및 정제 절차에 따라 처리될 수 있다(섹션 5).
탈보호 후, 올리고뉴클레오티드 트리포스페이트는 폴리아크릴아미드 겔 전기영동(PAGE)을 변성시켜 정제하여, 겔로부터 적출 및 용출될 수 있는 UV 백섀도잉에 의해 단일 주요 생성물 밴드를 나타낸다. 도 3A,B의 DNA 트리뉴클레오타이드 5ʹ-트리포스페이트, pppAAA 및 pppCCC, 및 L-RNA 트리뉴클레오타이드 5ʹ-트리포스페이트 pppGAA에 대해 나타낸 바와 같이 5'-트리포스페이트 생성물은 짧은 올리고뉴클레오티드에 대한 반응 측면 생성물로부터 쉽게 분리된다. AAA 및 CCC DNA 삼량체에 대한 5'-히드록실 및 5'-트리포스페이트 생성물 둘 다를 질량 분광법에 의해 적출 및 동정하고 도 3A에 상응하게 표지하였다. AAA DNA 삼량체에 대해 보이는 바와 같이 추가의 밴드는 일반적으로 회수 및 식별하기에 충분한 물질을 함유하지 않는다. 그러나 이들 밴드의 존재는 정제되지 않은 반응 생성물 내의 추가적인 생성물 질량과 상관관계가 있으며(도 3C), 전형적으로 아래에 논의된 바와 같이 5'-디포스페이트, 모노포스페이트 및 H-포스포네이트 부산물을 나타낸다.
PAGE 정제 후, 더 큰 올리고뉴클레오티드는 분쇄 및 담금질 방법(42 ) 및 후속 에탄올 침전을 사용하여 용출될 수 있다. 그러나, 15 nt 미만의 올리고뉴클레오티드는 에탄올이 효율적으로 침전될 수 없으며, 따라서, 겔 용출을 위한 변형된 절차가 필요하다(단계 5.11.3). 물질 표 에 열거된 일회용 크기 배제 컬럼은 10 nt보다 긴 올리고뉴클레오티드와 함께 사용하는 것에 대해서만 평가된다. 그러나 우리는 삼량체만큼 짧은 올리고뉴클레오티드가 제조업체의 권장 프로토콜을 사용하여 효과적으로 탈염 될 수 있음을 발견했습니다. 그럼에도 불구하고, 짧은 올리고뉴클레오티드를 탈염할 때(단계 5.6 및 5.11.3에서와 같이) 컬럼 용출액은 분획으로 수집하고, 생성물 분획은 UV-Vis 분광광도계를 사용하여 260nm에서의 흡광도로 확인하는 것이 좋다. 더 짧은 올리고뉴클레오티드에 대해 최적화된 크기 배제 컬럼이 대안적인 선택으로서 물질의 표 에 제공된다. 정제 후 합성된 올리고뉴클레오타이드 1 μmole 스케일로부터의 최종 수율은 50-300 nmol이다.
트리포스포릴화는 질량 분광법에 의해 확인될 수 있는데, 여기서 트리인산화된 생성물은 5'-히드록실 올리고뉴클레오티드보다 +239.94 Da 큰 질량을 갖지만, 5'-디- 및 모노포스페이트(각각 +159.96 및 +79.98 Da)에 상응하는 물질의 존재는 종종 관찰된다. 5'-OH 질량으로부터 +63.98 Da의 질량을 갖는 5'-H-포스포네이트 측 생성물이 또한 관찰될 수 있고, 이 생성물의 높은 수준은 트리포스포릴화 동안의 조건이 충분히 무수적이지 않았음을 나타낸다. 정제 전에, 탈보호된 올리고뉴클레오티드는 전형적으로 이러한 모든 생성물을 보여주지만(도 3C), 정제된 물질은 5'-디- 및 모노포스페이트와 함께 5'-트리포스페이트 생성물에 상응하는 피크를 보여줄 것이다(도 3D,E).
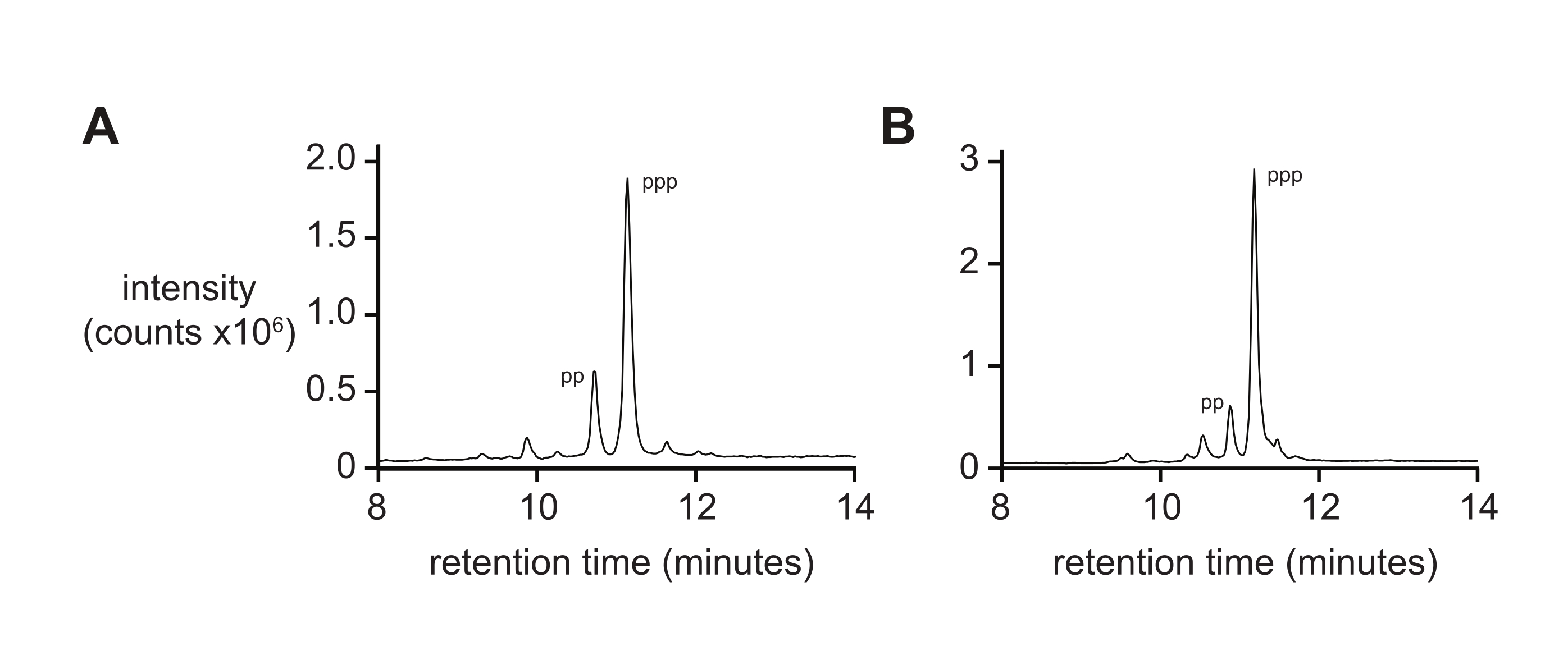
질량 분광법만으로는 일반적으로 이온화 동안 삼인산염의 차등 속도 및 단편화로 인해 5'-트리포스페이트 순도의 엄격한 측정을 제공하지 않습니다. 최종 생성물 순도를 측정하기 위해, 역상 액체 크로마토그래피 및 탠덤 ESI-MS(RP-LC/ESI-MS)가 권장되며, 특히 더 긴 올리고뉴클레오티드의 경우 더욱 그렇습니다. RP-LC/ESI-MS에 의한 D-RNA 5ʹ-트리포스페이트 pppACGAGG 및 pppGAGACCGCAACUUA의 분석(각각 도 4A, B)은 20% 5ʹ-디포스페이트를 포함하는 전형적인 최종 생성물 순도를 보여주는데, 이는 이 두 종들이 더 긴 올리고뉴클레오티드에 존재할 때 분리하기 어렵기 때문이다.
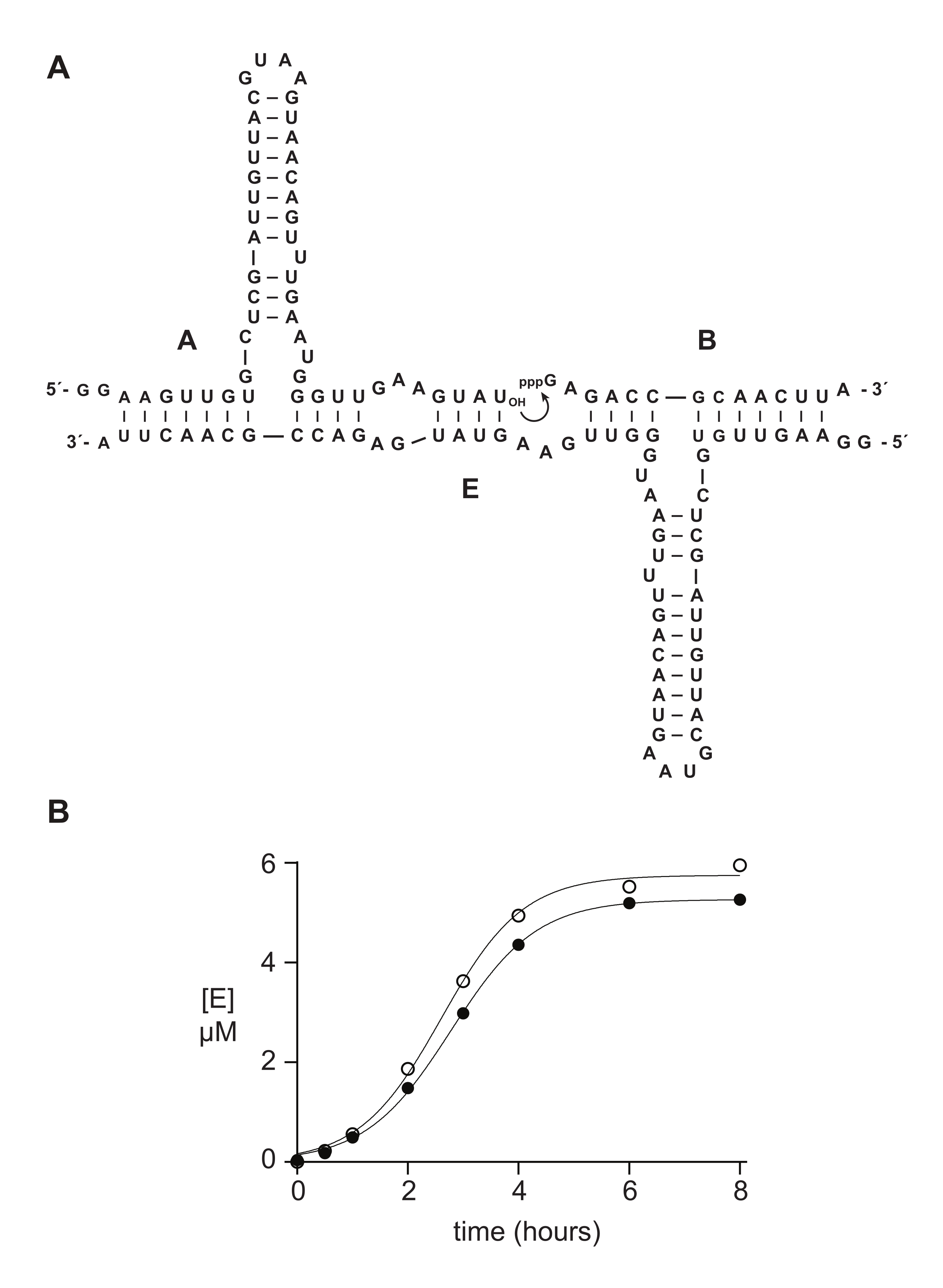
합성 5'-트리포스페이트 올리고뉴클레오티드는 전형적으로 생화학적 연구에서 효소적으로 제조된 물질보다 잘 또는 더 잘 기능한다. 섹션 6에서, 일례로, 합성적으로 또는 시험관내 전사에 의해 제조된 5'-트리포스페이트 14 nt RNA 기질을 RNA-촉매화된 자가-복제 반응 14,15,43,44,45에서 비교하였다. 리보자임 E는 기질 A와 B의 결합을 촉매하여 기하급수적으로 성장할 수 있는 자가촉매 반응에서 E의 새로운 카피를 생성한다(도 5A). E 및 32P표지된 A 성분은 시험관내 전사에 의해 제조되었고, 트리포스포릴화된 기질 B는 상기 기재된 바와 같이 합성적으로, 또는 시험관내 전사14에 의해 제조되었다. 자기 복제 반응 진행은 PAGE를 변성시켜 분석하고 형광/인광 겔 스캐너를 통해 정량화한 주기적인 샘플을 취하여 모니터링하였다. 물류 성장 기능에 적합한 결과 데이터는 전사 된 기질 또는 합성 B 기질이 기하 급수적 인 성장을 지원하지만 합성 B는 약간 더 많은 양의 제품을 제공한다는 것을 보여주었습니다 (그림 5B). 이러한 결과는 시험관내 전사23,24에 의해 제조된 RNA의 5'-말단에서의 조성적 이질성을 반영할 수 있다.
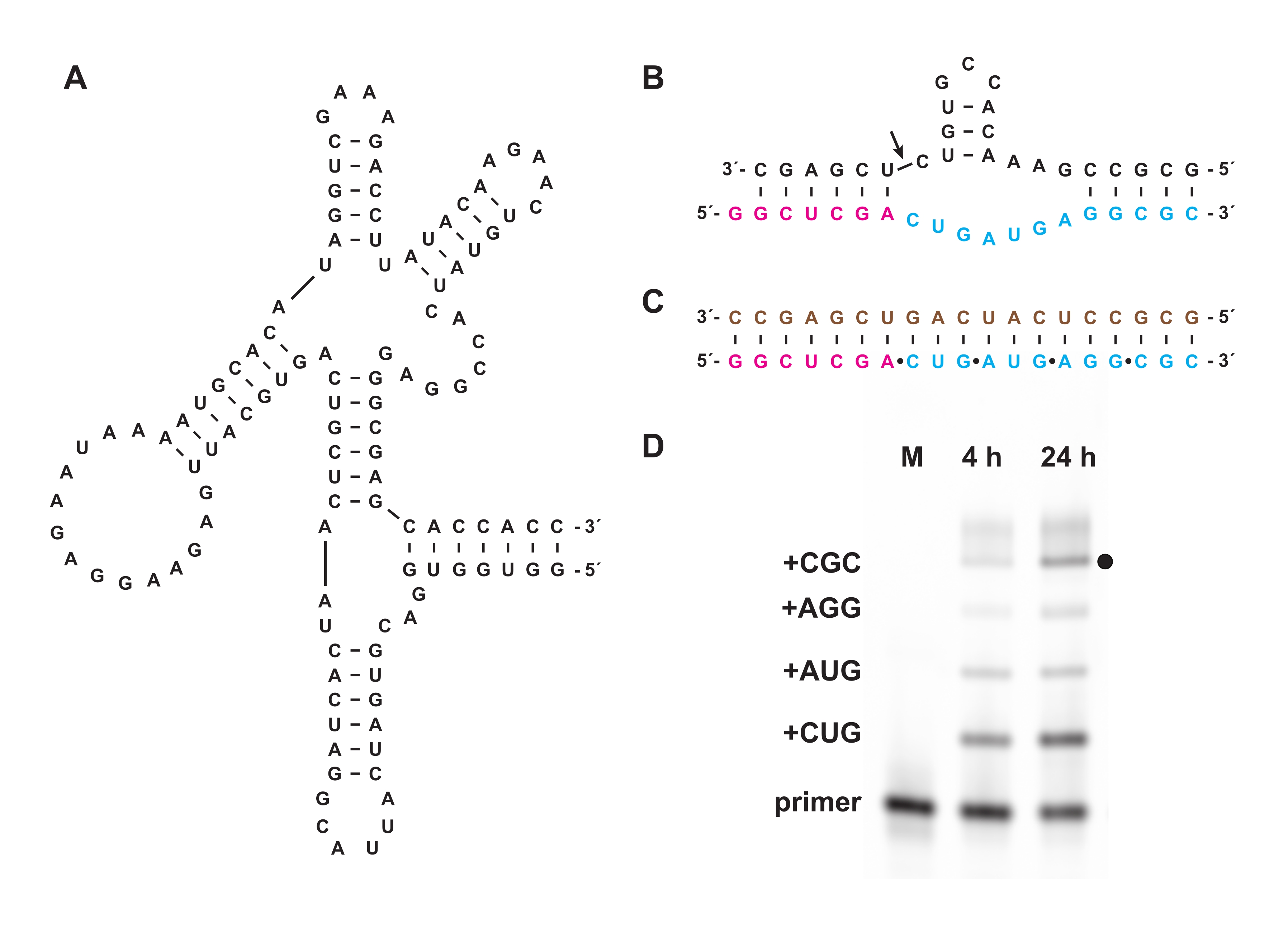
화학적 트리포스포릴화는 또한 시험관내 또는 세포에서 생물학적으로 제조될 수 없는 올리고뉴클레오티드 트리포스페이트의 합성을 가능하게 한다. 섹션 7에서, 섹션 1-5에서와 같이 제조된 천연 D-RNA의 거울상이성질체인 L-RNA로 구성된 비생물학적 트리포스페이트 올리고뉴클레오티드를 D-RNA "교차-키랄" 중합효소 리보자임 27.3t(도 6A)의 기질로 사용하였고, 이는 짧은 L-RNA 올리고뉴클레오티드 5'-트리포스페이트로부터의 더 긴 L-RNA 생성물의 주형 지향 중합을 서열-일반적인 방식으로 촉매한다. 일례로서, 리보자임은 해머헤드 자가절단 모티프의 L-RNA 버전을 합성할 수 있다(도 6B)18. 정제된 L-RNA 트리뉴클레오티드 트리포스페이트를 플루오레세인 표지된 L-RNA 프라이머 및 L-RNA 주형(도 6C)과 결합시키고, 교차 키랄 리가아제와 반응시켰다. 반응 과정에 걸친 샘플을 PAGE로 분석하고 형광/인광 겔 스캐너를 사용하여 이미징하여 주형에 의해 코딩되는 해머헤드 리보자임의 L-RNA 버전의 합성을 입증하였다(도 6D).

그림 3: 트리뉴클레오타이드 5ʹ-트리포스페이트의 정제. (A) DNA 트리뉴클레오티드의 트리포스포릴화의 PAGE 분석(UV-back-shadowing에 의해 시각화됨)은 트리데옥시아데노신(AAA, blue) 및 트리데옥시시티딘(CCC, 적색)이며, 사소한 측면 생성물을 시각화하기 위해 의도적으로 과부하되었다. 5ʹ-트리포스페이트 생성물(ppp) 및 5ʹ-히드록실(OH) 출발 물질을 모두 적출하고 MALDI-MS에 의해 확인하였다. (B) 주요 생성물 밴드를 적출하고 ESI-MS에 의해 5ʹ-트리포스페이트(ppp)로 확인된 L-RNA 트리뉴클레오티드 GAA의 트리포스포릴화의 준비 PAGE. (C) 탈보호 후의 조 반응 생성물의 MALDI-MS 및 (D) (A)로부터의 정제된 생성물. 5ʹ-트리포스페이트 (ppp; pppAAA 예상 1,119 Da, 관찰 1,118 Da; pppCCC 예상 1,047 Da, 관찰 1,046); 5ʹ-디포스페이트(pp), 5ʹ-모노포스페이트(p), 5ʹ-하이드록실(OH) 및 5ʹ-H-포스포네이트(Hp)라는 라벨이 붙어 있다. (E) (B)로부터 분리된 5ʹ-트리포스페이트 생성물의 직접 주입 ESI-MS로부터의 디컨볼루션된 질량 스펙트럼, 확인된 피크가 표지된 (예상 1,181.6 Da, 관찰된 1,181.0 Da). 5ʹ-디포스페이트 (pp) 생성물도 관찰되며, 트리포스페이트 및 디포스페이트 생성물 (+22 Da) 모두에 대한 나트륨 이온 피크도 관찰된다. 일반적인 오염 물질 피크에는 별표가 표시되어 있습니다. 비교의 용이성을 위해, 질량 스펙트럼은 각 스펙트럼에서 측정된 가장 높은 강도로 정규화되었고, 그 값에 상대적인 백분율로 보고된다. 약어: PAGE = 폴리아크릴아미드 겔 전기영동; MALDI-MS = 매트릭스 보조 레이저 탈착/이온화; ESI-MS = 전기 분무 이온화 질량 분광법. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

도 4: 6 nt 및 14 nt D-RNA 올리고뉴클레오티드 트리포스페이트의 분석 RP-LC . (A) 5ʹ-pppACGAGG-3ʹ 및 (B) 5ʹ-pppGAGACCGCAACUUA-3ʹ. Tandem ESI-MS는 둘 다의 주요 피크 (~ 70 %)를 5ʹ-triphosphate (ppp)로 확인했으며, 5ʹ-diphosphate (pp)의 양은 적습니다. 약어: RP-LC = 역상 액체 크로마토그래피; nt = 뉴클레오티드; ESI-MS = 전기 분무 이온화 질량 분광법. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

그림 5: 화학적 합성 또는 시험관내 전사에 의해 제조된 올리고뉴클레오티드 5ʹ-트리포스페이트 기질의 비교. (A) 자가복제 리보자임 E는 RNA A와 5'-트리 포스포릴화 RNA B를 리게이트한다. (B) 합성(개방 원) 또는 시험관내 전사(채워진 원)인 10μM A 및 10μM B를 사용한 자가 복제 반응의 비교. (B) 데이터는 로지스틱 성장 방정식에 적합하였다: [E] = a/(1+b e-ct), 여기서 a는 최종 수율이고, b는 시그모이디시티의 정도이고, c는 지수 성장률이다. 두 반응의 성장률은 1.14 h-1에서 동일했지만 최종 범위는 합성 B와의 반응에 대해 10 % 더 높았습니다.이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

도 6: 리보자임과의 교차-키랄 L-RNA 중합. (A) L-RNA의 주형-의존성 라이게이션을 촉매하는 D-RNA 27.3t 중합효소 리보자임. (b) 27.3t에 의해 합성된 L-RNA 생성물은 해머헤드 엔도뉴클레아제 모티프의 일부를 형성한다. (c) 비오티닐화된 L-RNA 주형(갈색), 말단 표지된 L-RNA 프라이머(마젠타), 및 네 개의 L-RNA 트리뉴클레오티드 트리포스페이트(시안)를 사용하여 27.3t에 의해 촉매된 L-RNA 중합, 합성적으로 제조됨. (d) 4 h 및 24 h에서 (B)의 연장 산물의 PAGE 분석은, 전장 생성물 (검은 점)까지의 각각의 트리뉴클레오티드 혼입을 보여준다. 미반응 L-RNA 프라이머는 참조 마커로서 포함된다. 약어: PAGE = 폴리아크릴아미드 겔 전기영동; M = 참조 마커. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
토론
여기에 설명된 트리포스포릴화 절차는 표준 포스포르아미다이트 화학을 이용한 올리고뉴클레오티드 합성과 광범위하게 양립가능하다. 뉴클레오시드 포스포르아미다이트는 포스파이트 상의 표준 β-시아노에틸, 및 핵염기의 외환형 아민 상의 이소부티릴, 디메틸포름아미딜, 아세틸, 페녹시아세틸 또는 4-이소프로필페녹시아세틸기를 포함하는 AMA39에서 신속한 탈보호와 양립가능한 염기-불안정한 보호기를 가져야 한다. 리보스 2'-히드록실기는 실릴 보호기, 즉 t-부틸디메틸실릴(TBDMS) 또는 트리이소프로필실릴옥시메틸(TOM)40,41에 의해 보호되어야 한다. 염기-불안정성 피발로일옥시메틸(PivOM) 그룹은 또한 화학적 트리포스포릴화30과 상용성인 것으로 보고되었다.
합성 올리고뉴클레오티드28,29,30,31,32,33,34,35의 화학적 트리인산화에 대해 다수의 방법이 기술되었다. 우리는 Ludwig-Eckstein 시약37을 사용한 트리포스포릴화가 가장 접근하기 쉬운 것 중 하나라는 것을 발견했으며, 시약의 특수 합성과 특수 장비가 필요하지 않습니다. 이 방법에 의해 제조된 올리고뉴클레오티드 5'-트리포스페이트는 RNA 리보자임(RNABE ribozymes)의 기질로서 일상적으로 사용되어 왔으며, 효소적으로 접근이 불가능한 L-RNA 올리고뉴클레오티드 트리포스페이트를 사용하여 주형-의존적 합성 및 이 "미러-이미지" 핵산의 복제를 달성하는것을 포함하여 14,16,17,18 . 이 방법은 또한 척추동물 6,7에서 선천적 면역 반응의 강력한 활성화제인 작은 5'-트리포스포릴화된 줄기-루프 RNAs의 제조에 적합하다.
Ludwig-Eckstein 시약인 살리실 포스포로클로리다이트37은 물에 매우 반응성이 높으며, 올리고뉴클레오티드 컬럼에 로딩하기 전에 시약을 용해시킬 때 도입된 모든 물을 효과적으로 제거한다. 그러나, 이 시점 이후, 5'-포스피실화된 올리고뉴클레오티드는 피로포스페이트 위에 시스템에 도입된 임의의 물과 우선적으로 반응하여, 워크업 28,37,38 후에 5'-H-포스포네이트 부생성물을 형성할 것이다. 트리인산화 시약 및 트리인산화 반응 챔버를 조심스럽게 준비하면 이러한 부산물이 형성되지 않도록 한다. 용제 건조를 위해, 유형 4 Å 분자체는 대부분의 올리고뉴클레오티드 합성 시약 회사에 의해 다양한 상표명으로 대부분의 유기 용매와 호환되는 테프론 백에 미리 포장되어 판매됩니다. 무수 분위기 하에서 글로브 박스에서 트리포스포릴화를 수행하는 것과 같은 추가적인 예방조치는 일반적으로 필요하지 않다.
TBAP와 5'-포스피실화된 올리고뉴클레오티드의 반응은 고리형 5'-트리메타포스파이트 중간체를 형성하고, 이는 올리고뉴클레오티드 합성 산화제 용액(물/피리딘/THF의 요오드)을 사용하여 고리형 5'-트리메타포스페이트로 산화된다. 상업용 산화제 용액은 다양한 양의 요오드를 사용하며 트리포스페이트에 대한 완전한 산화를 보장하기 위해 높은 0.1M 요오드 농도를 사용하는 것이 필수적이라는 점에 유의해야합니다. 고리형 생성물은 동일한 용액(37)에서 최종 선형 5'-트리포스페이트로 가수분해되고, 물 이외의 친핵체와의 선형화가 필요한 경우 대안적인 무수 산화제 용액을 사용해야 한다(용도는 아래 참조)33. 그러나, 임의의 잔류 고리형 트리메타포스페이트는 올리고뉴클레오티드의 후속 알칼리성 탈보호 동안 선형화될 것이다. 순환 5'-트리메타포스페이트의 가수분해는 분지형 트리포스페이트37,46이 아닌 선형만을 산출한다.
올리고뉴클레오티드 탈보호는 전형적으로 5'-트리포스포릴화를 수용하기 위해 변형될 필요가 없지만, 몇 가지 예방책이 취해져야 한다. 삼인산염은 알칼리성 조건에 대한 짧은 노출로 비교적 안정하지만, 필요한 것보다 더 오래 AMA에 삼인산염을 노출시키지 않도록 주의해야 한다. 65°C에서 10분 이상 암모니아 또는 AMA에서 보다 장기간의 처리를 필요로 하는 보호기는 피해야 한다. 실온에서 암모니아에서 2h와 같은 보다 온화한 처리는 다른 포스포르아미다이트 보호기와 양립할 때 허용가능하다. 실릴-보호된 합성 RNA 올리고뉴클레오티드에 대한 일반적인, 빠른 탈보호 방법은 트리에틸아민 트리히드로플루오라이드 및 고온47을 이용한다; 그러나, RNA 5'-트리포스페이트를 제조할 때는 장기간 산성 조건이 트리포스페이트 가수분해31,32를 가속화하는 것으로 밝혀짐에 따라 이것은 피해야 한다.
제조 PAGE는 5'-트리포스포릴화된 올리고뉴클레오티드의 탈보호 후 정제를 위한 가장 간단하고 가장 신뢰할 수 있는 방법임이 입증되었다(도 3 및 도 4). 그러나, 제조용 역상 HPLC는 또한 트리인산화된 생성물을 정제하는데 사용될 수 있다. 5'-디포스페이트 및 5'-모노포스페이트 생성물의 존재는 질량 분광법에 의한 삼인산화를 검증할 때 일상적으로 관찰된다. 우리는 화학 합성 또는 전사에 의해 제조 된 고순도 물질로부터 질량 분광법 중에 5'-트리 포스페이트 단편화를 관찰했으며, 특히 장비가 올리고 뉴클레오티드 분석에 최적화되지 않은 경우 특히 그렇습니다. 그럼에도 불구하고, RP-LC 분석은 종종 5'-디포스페이트 부산물의 10%-20%가 더 긴 5'-트리포스포릴화 올리고뉴클레오티드에 존재한다는 것을 보여준다(도 4). 트리부틸암모늄 피로포스페이트의 상업적 제제는 20% 모노포스페이트로 오염될 수 있으며, 이는 트리포스포릴화 동안 부생성물로서 5'-디포스페이트를 산출할 것이다30,31. 이 시약을 사내에서 조심스럽게 준비하면 훨씬 더 순수한 TBAP 주식(31)을 얻을 수 있다. 그러나, 우리는 TBAP의 상업적 공급원을 사용하여 트리포스포릴화된 올리고뉴클레오티드가 시험관내 전사에 의해 제조된 물질에 비해 효소 반응에서 기질로서 사용될 때 여전히 필적할만한 또는 더 큰 반응성을 나타낸다는 것을 발견하였다 (도 5B).
루드비히-에크슈타인 시약을 사용한 트리포스포릴화 올리고뉴클레오티드의 한 가지 주목할만한 추가의 사용은 고리형 트리메타포스파이트 중간체33을 이용한다. 후속 산화 단계가 무수 조건 하에서 올리고뉴클레오티드 산화에 자주 사용되는 헥산 중의 1 M t-부틸 퍼옥사이드로 진행되면, 포스파이트의 산화는 고리 개구 가수분해 없이 발생하여, 고리형 트리메타포스페이트를 산출한다. 이 중간체는 이어서 일차 아민 또는 알콜 친핵체와 반응하여 γ-포스페이트에서 변형된 5'-트리포스페이트를 수득할 수 있다. 이러한 변형은 포스포르아미데이트 결합에 의해 연결된 친유성 태그의 첨가를 포함하며, 이는 RP-LC에 의한 신속한 트리포스페이트 특이적 정제를 용이하게 하고, 이어서 트리포스페이트33으로부터 태그의 산성 가수분해를 수반한다. γ-포스페이트 위치에서의 형광 변형은 또한 리보자임-촉매 라이게이션 반응(15,33)을 위한 실시간 형광 리포터로서 사용하기 위해 도입될 수 있다.
공개
저자들은 경쟁적인 재정적 이익이 없다고 선언합니다.
감사의 말
저자는 Greg Springsteen, Natasha Paul, Charles Olea, Jr., Jonathan Sczepanski 및 Katrina Tjhung에게 화학 삼인산화 반응에 대한 모범 사례에 대한 유용한 토론과 Gerald Joyce에게 유용한 의견을 주신 것에 감사드립니다. 이 연구는 국립 과학 재단 (National Science Foundation)의 보조금 MCB 2114588에 의해 지원되었습니다.
자료
| Name | Company | Catalog Number | Comments |
| 0.22 µm polyethersulfone syringe filter | MilliporeSigma | SLMP025SS | Syringe filter for removing crushed polyacrylamide gel particles (Section 5) |
| 0.22 µm PTFE syringe filter | MilliporeSigma | SLLG013SL | Syringe filter for removing CPG resin (Section 5) |
| 1 mL plastic syringes | ThermoFisher Scientific | 14-823-434 (BD 309659) | Components of triphosphorylation apparatus (sections 2–4) |
| 1,4-Dioxane, anhydrous | MilliporeSigma | 296309 | Triphosphorylation solvent (sections 2–4) |
| 2-Chloro-4H-1,3,2-benzodioxaphosphorin-4-one, Salicyl Phosphorochloridite (SalPCl) | MilliporeSigma | 324124 | Triphosphorylation reagent (sections 2–4) |
| 30 mL glass bottles | MilliporeSigma | 23232 | Bottles for preparing triphosphorylation solvents and TBAP solution (section 2) |
| 3-way Stopcock, polycarbonate/polypropylene | Bio-Rad Laboratories | 7328103 | Component of triphosphorylation apparatus (sections 2–4) |
| 40% acrylamide/bis-acrylamide solution, 19:1 | Bio-Rad Laboratories | 1610144 | For PAGE (sections 5–7) |
| Acetonitrile, anhydrous, 100 mL | Glen Research | 40-4050-50 | Triphosphorylation solvent (sections 2–4) |
| Ammonia-neutralizing Trap | ThermoFisher Scientific | ANT100 and ANS121 | For use with Speedvac DNA130 (section 5) |
| Ammonium persulfate (APS) | Bio-Rad Laboratories | 1610700 | For PAGE, catalyst for acrylamide polymerization (sections 5–7) |
| Aqueous ammonia, 28% | MilliporeSigma | 338818 | For preparing AMA deprotection reagent (section 5) |
| Aqueous methylamine, 40% | TCI America | TCI-M0137 | For preparing AMA deprotection reagent (section 5) |
| Automated DNA/RNA oligonucleotide synthesizer | PerSeptive Biosystems | Expedite 8909 DNA/RNA Synthesizer | any column-based synthesizer is acceptable (section 1) |
| Bead-capture magnet | ThermoFisher Scientific | 12320D | For streptavidin bead capture (section 7) |
| Bromophenol blue | Bio-Rad Laboratories | 1610404 | For PAGE urea loading buffer (section 5) |
| Deep vacuum oil pump | ThermoFisher Scientific | VLP200-115 | For use with lyophilizer (section 5) |
| Drierite dessicant, 10-20 mesh | MilliporeSigma | 737828 | Desiccant for storing triphosphorylation chemicals and equipment (sections 1–2) |
| D-RNA 27.3t cross-chiral polymerase | prepared in house18 | 5′-GGUGGUGGAC GUGAUCAUUA CGGAUCACUA ACUCGUCAGU GCAUUGAGAA GGAGAAUAAA AUGCACAUAG GUCGAAAGAC CUUAUACAAG AACUGUAUCA CCGGAGGGCG AGCACCACC-3′ | For cross-chiral ribozyme reactions (section 7) |
| D-RNA CPG solid supports, 1,000Å, prepackaged 1 µmole synthesis columns | Glen Research | 20-3404-41E, 20-3415-41E, 20-3424-41E, 20-3430-41E | representative, for D-RNA oligonucleotide synthesis (section 1) |
| D-RNA TOM-protected phosphoramidites | ChemGenes | ANP-3201, 3202, 3203, 3205 | representative, for D-RNA oligonucleotide synthesis (section 1) |
| Empty Expedite Synthesis Columns, 1µm | Glen Research | 20-0021-01 | Synthesis columns for use with Expedite DNA/RNA synthesizer (section 1) |
| EPPS, N-(2-Hydroxyethyl)piperazine-N′-(3-propanesulfonic acid), solid | MilliporeSigma | E1894 | Ribozyme reaction buffer component (section 6) |
| Ethylenediaminetetraacetic acid (EDTA), solid | MilliporeSigma | EDS | Divalent metal ion chelator for use in various buffers (sections 5–7) |
| Filters for Expedite synthesis columns | Glen Research | 20-0021-0F | Expedite-style synthesis column filters, for use with empty synthesis columns (section 1) |
| Fluorescent/phosphorescent gel scanner | Cytiva | Amersham Typhoon RGB, 29187193 | For visualizing analytical PAGE (sections 6–7) |
| Formamide, deionized | VWR Life Science | 97062 | For PAGE formamide gel loading buffer (sections 6–7) |
| Gel image quantitation software | Cytiva | ImageQuant TL | For quantifying scanned gel images (section 6) |
| Glass desiccator | MilliporeSigma | CLS3121150 | Triphosphorylation solvent storage (section 2) |
| L-RNA CPG solid supports, 1,000Å, bulk | ChemGenes | N-4691-10, N-4692-10, N-4693-10, N-4694-10 | L-RNA oligonucleotide synthesis (section 1) |
| L-RNA hammerhead template | prepared in house18 | 5′-GCGCCUCAUC AGUCGAGCC-3′ | For cross-chiral ribozyme reactions (section 7) |
| L-RNA primer | prepared in house18 | 5′-fluorescein-GGCUCGA-3′ | For cross-chiral ribozyme reactions (section 7) |
| L-RNA TOM-protected phosphoramidites | ChemGenes | OP ANP-5201, 5202, 5203, 5205 | L-RNA oligonucleotide synthesis (section 1) |
| Lyophilizer/Freeze Dryer | VirTis | Benchtop K | For concentrating oligonucleotides (section 5) |
| Magnesium Chloride Hexahydrate, solid | MilliporeSigma | M2670 | For ribozyme reactions (sections 6–7) |
| N,N-Dimethylformamide, anhydrous | MilliporeSigma | 227056 | Triphosphorylation solvent (section 2) |
| NAP-25 Desalting column (Sephadex G-25 resin) | ThermoFisher Scientific | 45000150 | Disposable gravity-flow size exclusion chromatography columns containing Sephadex G-25 resin (section 5) |
| Non-coring stainless steel needle, 20 G | ThermoFisher Scientific | 14-815-410 | Needles for piercing rubber septa (sections 2–4) |
| Oligonucleotide extinction coefficient calculator | Integrated DNA Technologies | OligoAnalyzer Tool | Nearest-Neighbor Model Short Oligonucleotide 260nm extinction coefficient calculator (section 5) |
| Oxidizer solution, 0.1 M Iodine in THF/pyridine/water | ChemGenes | RN-1456 | Triphosphorylation reagent (section 4) |
| PAGE plates | Timberrock/CBS | NGP-250-BO and NO | For PAGE (sections 5–7) |
| PAGE power supply | Bio-Rad Laboratories | PowerPac HV 1645056 | For PAGE (sections 5–7) |
| PAGE spacers and combs (analytical) | Timberrock/CBS | VGS-0725 and VGC-0714 | For PAGE (sections 6–7) |
| PAGE spacers and combs (preparative) | Timberrock/CBS | VGS-3025R and VGC-3001 | For PAGE (section 5) |
| PAGE stand | Timberrock/CBS | ASG-250 | For PAGE (sections 5–7) |
| Parafilm M | ThermoFisher Scientific | 13-374-12 (Bemis PM999) | Wax sealing film for triphosphorylation apparatus (sections 2–4) |
| PCR thermocycler | Bio-Rad Laboratories | C1000 Touch Thermalcycler | For cross-chiral ribozyme reactions (section 7) |
| PD 10 Desalting column (Sephadex G-10 resin) | MilliporeSigma | GE17-0010-01 | Disposable gravity-flow size exclusion chromatography columns containing Sephadex G-10 resin, for oligonucleotides < 15 nt (section 5) |
| Phosphor screens | Cytiva | 28956480 | For visualizing 32P-labeled RNA (section 6) |
| Phosphoramidite synthesis reagents | Glen Research | 30-3142-52, 40-4050-53, 40-4012-52, 40-4122-52, 40-4132-52, 40-4060-62 | representative, for standard RNA/DNA synthesis (section 1) |
| Polypropylene screw-cap sealable tube | MilliporeSigma | BR780752 | 1.5 mL microcentrifuge tubes with screw-cap and silicone O-ring, for safe AMA deprotection (section 5) |
| Pyridine, anhydrous | MilliporeSigma | 270970 | Triphosphorylation solvent (section 2) |
| Reverse-phase liquid chromatography/electrospray ionization mass spectrometry (RP-LC/ESI-MS) | Novatia | n/a | Commercial service for LC/MS specializing in oligonucleotides (section 5) |
| Rubber Septa (ID x OD 7.9 mm x 14 mm), white | MilliporeSigma | Z564702 | Septa for preparing triphosphorylation solvents and TBAP (section 2) |
| Self-replicator ribozyme E | prepared in house14 | 5′-GGAAGUUGUG CUCGAUUGUU ACGUAAGUAA CAGUUUGAAU GGUUGAAGUA UGAGACCGCA ACUUA-3′ | For self-replicator ribozyme reactions (section 6) |
| Self-replicator substrate A | prepared in house14 | 5′-32P-GGAAGUUGUG CUCGAUUGUU ACGUAAGUAA CAGUUUGAAU GGUUGAAGUA U-3′-OH | For self-replicator ribozyme reactions (section 6) |
| Self-replicator substrate B, transcribed | prepared in house14 | 5′-pppGAGACCGCAA CUUA-3′ | For self-replicator ribozyme reactions (section 6) |
| Small Drying Traps, 4 Å molecular sieves | ChemGenes | DMT-1975 | Drying traps for DNA/RNA synthesizer phosphoramidites and triphosphorylation reagents (sections 1–2) |
| Sodium Chloride (NaCl), solid | MilliporeSigma | S7653 | Salt for use in various buffers (sections 5–7) |
| Sodium Hydroxide (NaOH), solid | MilliporeSigma | S8045 | Salt for use in various buffers (sections 5–7) |
| Statistical data-fitting software | GraphPad | Prism | For fitting data from analytical PAGE to kinetic models (section 6) |
| Streptavidin-coated magnetic beads | ThermoFisher Scientific | 65002 | For capturing biotin-labeled RNA in cross-chiral ribozyme reactions (section 7) |
| Sucrose | MilliporeSigma | 84097 | For PAGE urea loading buffer (section 5) |
| TBE running buffer, 10x | ThermoFisher Scientific | AAJ62788K3 | For PAGE (sections 5–7) |
| Tetrabutylammonium Fluoride, 1.0 M solution in Tetrahydrofuran | Aldrich | 216143 | For removing 2′-silyl protecting groups (section 5) |
| Tetramethylethylenediamine (TEMED) | Bio-Rad Laboratories | 1610801 | For polymerizing acrylamide for PAGE (sections 5–7) |
| Tributylamine | MilliporeSigma | 90781 | Triphosphorylation reagent (section 2) |
| Tributylammonium pyrophosphate (TBAP) | MilliporeSigma | P8533 | Triphosphorylation reagent (section 2) |
| Tris base | MilliporeSigma | T6666 | Buffering agent for use in various buffers (sections 5–7) |
| TWEEN20 polysorbate detergent | MilliporeSigma | P7949 | Neutral detergent for use with magnetic beads (Section 7) |
| Urea | MilliporeSigma | U5378 | For PAGE and gel loading buffer (sections 5–7) |
| UV-Vis spectrophotometer | ThermoFisher Scientific | NanoDrop 2000, ND2000 | For measuring oligonucleotide concentrations (section 5) |
| Vacuum centrifuge | ThermoFisher Scientific | Savant Speedvac DNA130-115 Vacuum Concentrator | For removing AMA and THF (section 5) |
| Xylene cyanol | Bio-Rad Laboratories | 1610423 | For PAGE urea loading buffer (section 5) |
참고문헌
- Shuman, S. What messenger RNA capping tells us about eukaryotic evolution. Nature Reviews. Molecular Cell Biology. 3 (8), 619-625 (2002).
- Pichlmair, A., et al. RIG-I-mediated antiviral responses single-stranded RNA bearing 5'-phosphates. Science. 314 (5801), 997-1001 (2006).
- Hornung, V., et al. 5'-Triphosphate RNA is the ligand for RIG-I. Science. 314 (5801), 994-997 (2006).
- Myong, S., et al. Cytosolic viral sensor RIG-I is a 5'-triphosphate-dependent translocase on double-stranded RNA. Science. 323 (5917), 1070-1074 (2009).
- Takeuchi, O., Akira, S. Pattern recognition receptors and inflammation. Cell. 140, 805-820 (2010).
- Wang, Y., et al. Structural and functional insights into 5'-ppp RNA pattern recognition by the innate immune receptor RIG-I. Nature Structural & Molecular Biology. 17 (7), 781-787 (2010).
- Goubau, D., et al. Antiviral immunity via RIG-I-mediated recognition of RNA bearing 5'-diphosphates. Nature. 514 (7522), 372-375 (2014).
- Joyce, G. F. Forty years of in vitro evolution. Angewandte Chemie. 46 (34), 6420-6436 (2007).
- Robertson, M. P., Ellington, A. D. In vitro selection of an allosteric ribozyme that transduces analytes to amplicons. Nature Biotechnology. 17 (1), 62-66 (1999).
- Robertson, M. P., Hesselberth, J. R., Ellington, A. D. Optimization and optimality of a short ribozyme ligase that joins non-Watson-Crick base pairings. RNA. 7 (4), 513-523 (2001).
- Hesselberth, J. R., Robertson, M. P., Knudsen, S. M., Ellington, A. D. Simultaneous detection of diverse analytes with an aptazyme ligase array. Analytical Biochemistry. 312 (2), 106-112 (2003).
- Lam, B. J., Joyce, G. F. Autocatalytic aptazymes enable ligand-dependent exponential amplification of RNA. Nature Biotechnology. 27 (3), 288-292 (2009).
- Lam, B. J., Joyce, G. F. An isothermal system that couples ligand-dependent catalysis to ligand-independent exponential amplification. Journal of the American Chemical Society. 133 (9), 3191-3197 (2011).
- Olea, C., Horning, D. P., Joyce, G. F. Ligand-dependent exponential amplification of a self-replicating L-RNA enzyme. Journal of the American Chemical Society. 134 (19), 8050-8053 (2012).
- Olea, C., Joyce, G. F. Real-Time Detection of a Self-Replicating RNA Enzyme. Molecules. 21 (10), (2016).
- Sczepanski, J. T., Joyce, G. F. A cross-chiral RNA polymerase ribozyme. Nature. 515 (7527), 440-442 (2014).
- Tjhung, K. F., Sczepanski, J. T., Murtfeldt, E. R., Joyce, G. F. RNA-catalyzed cross-chiral polymerization of RNA. Journal of the American Chemical Society. 142 (36), 15331-15339 (2020).
- Bare, G. A. L., Joyce, G. F. Cross-chiral, RNA-catalyzed exponential amplification of RNA. Journal of the American Chemical Society. 143 (45), 19160-19166 (2021).
- Milligan, J. F., Groebe, D. R., Witherell, G. W., Uhlenbeck, O. C. Oligoribonucleotide synthesis using T7 RNA polymerase and synthetic DNA templates. Nucleic Acids Research. 15 (21), 8783-8798 (1987).
- Chelliserrykattil, J., Ellington, A. D. Evolution of a T7 RNA polymerase variant that transcribes 2'-O-methyl RNA. Nature Biotechnology. 22 (9), 1155-1160 (2004).
- Ibach, J., et al. Identification of a T7 RNA polymerase variant that permits the enzymatic synthesis of fully 2′-O-methyl-modified RNA. Journal of Biotechnology. 167 (3), 287-295 (2013).
- Esvelt, K. M., Carlson, J. C., Liu, D. R. A system for the continuous directed evolution of biomolecules. Nature. 472 (7344), 499-503 (2011).
- Pleiss, J. A., Derrick, M. L., Uhlenbeck, O. C. T7 RNA polymerase produces 5' end heterogeneity during in vitro transcription from certain templates. RNA. 4 (10), 1313-1317 (1998).
- Schenborn, E. T., Mierendorf, R. C. A novel transcription property of SP6 and T7 RNA polymerases: dependence on template structure. Nucleic Acids Research. 13 (17), 6223-6236 (1985).
- Martin, C. T., Muller, D. K., Coleman, J. E. Processivity in early stages of transcription by T7 RNA polymerase. Biochemistry. 27 (11), 3966-3974 (1988).
- Gholamalipour, Y., Karunanayake Mudiyanselage, A., Martin, C. T. 3' end additions by T7 RNA polymerase are RNA self-templated, distributive and diverse in character-RNA-Seq analyses. Nucleic Acids Research. 46 (18), 9253-9263 (2018).
- Vasilyev, N., Serganov, A. Preparation of short 5′-triphosphorylated oligoribonucleotides for crystallographic and biochemical studies. Nucleic Acid Crystallography: Methods and Protocols. , 11-20 (2016).
- Lebedev, A. V., Koukhareva, I. I., Beck, T., Vaghefi, M. M. Preparation of oligodeoxynucleotide 5'-triphosphates using solid support approach. Nucleosides, Nucleotides & Nucleic Acids. 20 (4-7), 1403-1409 (2001).
- Paul, N., Springsteen, G., Joyce, G. F. Conversion of a ribozyme to a deoxyribozyme through in vitro evolution. Chemistry & Biology. 13 (3), 329-338 (2006).
- Zlatev, I., et al. Efficient solid-phase chemical synthesis of 5'-triphosphates of DNA, RNA, and their analogues. Organic Letters. 12 (10), 2190-2193 (2010).
- Zlatev, I., Manoharan, M., Vasseur, J. -. J., Morvan, F. Solid-phase chemical synthesis of 5'-triphosphate DNA, RNA, and chemically modified oligonucleotides. Current Protocols in Nucleic Acid Chemistry. , (2012).
- Zlatev, I., et al. Automated parallel synthesis of 5'-triphosphate oligonucleotides and preparation of chemically modified 5'-triphosphate small interfering RNA. Bioorganic & Medicinal Chemistry. 21 (3), 722-732 (2013).
- Goldeck, M., Tuschl, T., Hartmann, G., Ludwig, J. Efficient solid-phase synthesis of pppRNA by using product-specific labeling. Angewandte Chemie. 53 (18), 4694-4698 (2014).
- Sarac, I., Meier, C. Efficient automated solid-phase synthesis of DNA and RNA 5′-triphosphates. Chemistry-A European Journal. 21 (46), 16421-16426 (2015).
- Sarac, I., Meier, C. Solid-phase synthesis of DNA and RNA 5'-O-triphosphates using cycloSal chemistry. Current Protocols in Nucleic Acid Chemistry. 64 (1), 4-67 (2016).
- Perez, J. T., et al. Influenza A virus-generated small RNAs regulate the switch from transcription to replication. Proceedings of the National Academy of Sciences of the United States of America. 107 (25), 11525-11530 (2010).
- Ludwig, J., Eckstein, F. Rapid and efficient synthesis of nucleoside 5'-0-(1-thiotriphosphates), 5'-triphosphates and 2',3'-cyclophosphorothioates using 2-chloro-4H-1,3,2-benzodioxaphosphorin-4-one. The Journal of Organic Chemistry. 54 (3), 631-635 (1989).
- Gaur, R. K., Sproat, B. S., Krupp, G. Novel solid phase synthesis of 2'-o-methylribonucleoside 5'-triphosphates and their α-thio analogues. Tetrahedron Letters. 33 (23), 3301-3304 (1992).
- Reddy, M. P., Hanna, N. B., Farooqui, F. Fast cleavage and deprotection of oligonucleotides. Tetrahedron Letters. 35 (25), 4311-4314 (1994).
- Hogrefe, R. I., McCaffrey, A. P., Borozdina, L. U., McCampbell, E. S., Vaghefi, M. M. Effect of excess water on the desilylation of oligoribonucleotides using tetrabutylammonium fluoride. Nucleic Acids Research. 21 (20), 4739-4741 (1993).
- Pitsch, S., Weiss, P. A., Jenny, L., Stutz, A., Wu, X. Reliable chemical synthesis of oligoribonucleotides (RNA) with 2′-O-[(Triisopropylsilyl)oxy]methyl(2′-O-tom)-protected phosphoramidites. Helvetica Chimica Acta. 84 (12), 3773-3795 (2001).
- Green, M. R., Sambrook, J. Isolation of DNA fragments from polyacrylamide gels by the crush and soak method. Cold Spring Harbor Protocols. 2019 (2), (2019).
- Paul, N., Joyce, G. F. A self-replicating ligase ribozyme. Proceedings of the National Academy of Sciences of the United States of America. 99 (20), 12733-12740 (2002).
- Lincoln, T. A., Joyce, G. F. Self-sustained replication of an RNA enzyme. Science. 323 (5918), 1229-1232 (2009).
- Robertson, M. P., Joyce, G. F. Highly efficient self-replicating RNA enzymes. Chemistry & Biology. 21 (2), 238-245 (2014).
- Singh, J., et al. Synthesis of modified nucleoside oligophosphates simplified: fast, pure, and protecting group free. Journal of the American Chemical Society. 141 (38), 15013-15017 (2019).
- Bellon, L. Oligoribonucleotides with 2'-O-(tert-butyldimethylsilyl) groups. Current Protocols in Nucleic Acid Chemistry. , (2001).
재인쇄 및 허가
JoVE'article의 텍스트 или 그림을 다시 사용하시려면 허가 살펴보기
허가 살펴보기더 많은 기사 탐색
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. 판권 소유