Method Article
A Cultura 3D de Organoides de Criptas Intestinais Murinas e uma Única Célula-Tronco para Pesquisa de Organoides
Neste Artigo
Resumo
Descrevemos um protocolo para isolar criptas murinas do intestino delgado e cultura de organoides intestinais 3D das criptas. Adicionalmente, descrevemos um método para gerar organoides a partir de uma única célula-tronco intestinal na ausência de um nicho celular subepitelial.
Resumo
Atualmente, a cultura de organoides representa uma importante ferramenta para estudos in vitro de diferentes aspectos biológicos e doenças em diferentes órgãos. As criptas murinas do intestino delgado podem formar organoides que mimetizam o epitélio intestinal quando cultivadas em uma matriz extracelular 3D. Os organoides são compostos por todos os tipos celulares que cumprem várias funções homeostáticas intestinais. Estes incluem células de Paneth, células enteroendócrinas, enterócitos, células caliciformes e células de tufo. Moléculas bem caracterizadas são adicionadas ao meio de cultura para enriquecer as células-tronco intestinais (ISCs) marcadas com repetições ricas em leucina contendo o receptor acoplado à proteína G 5 e são usadas para conduzir a diferenciação para linhagens específicas; essas moléculas incluem fator de crescimento epidérmico, Noggin (uma proteína morfogenética óssea) e R-espondina 1. Além disso, um protocolo para gerar organoides a partir de um único ISC positivo para receptor hepatocelular B2 (EphB2) produtor de eritropoetina também é detalhado. Neste artigo de métodos são descritas técnicas para isolar criptas do intestino delgado e um único ISC dos tecidos e garantir o estabelecimento eficiente de organoides.
Introdução
Os organoides intestinais, que foram estabelecidos pela primeira vez em 2009, emergiram como uma poderosa ferramenta in vitro para o estudo da biologia intestinal, dada a sua semelhança morfológica e funcional com tecidos maduros. Recentemente, avanços tecnológicos em cultura de organoides derivados de células-tronco de tecidos adultos têm permitido o cultivo a longo prazo de células-tronco intestinais (ISCs) com potencial de autorrenovação e diferenciação. Esses organoides têm sido amplamente utilizados para estudos de pesquisa básica e translacional sobre fisiologia e fisiopatologia gastrintestinal1,2,3,4,5,6. Os organoides 3D desenvolvidos pelo grupo Clevers fornecem uma poderosa ferramenta para o estudo do epitélio intestinal com melhora da relevânciafisiológica7. Como os organoides intestinais são derivados de células-tronco teciduais e são compostos por múltiplos tipos celulares, eles recapitulam a funcionalidade do epitélio intestinal. É importante ressaltar que uma célula-tronco acoplada ao receptor 5-positivo (Lgr5+) para leucina contendo repetições ricas em leucina também pode gerar organoides 3D sem células de Paneth ou um nicho ISC, como o nicho epitelial ou o nicho estromal7. No entanto, a capacidade de formação de organoides de células Lgr5+ de classificação simples é baixa em comparação com as de células criptas e células ISC-Paneth8.
Um número crescente de estudos tem demonstrado que os métodos de incubação do ácido etilenodiaminotetracético (EDTA) ou dissociação da colagenase causam soltura no epitélio e liberação de criptas. Como a dissociação enzimática pode ter um efeito sobre o estado celular das criptas, um método de isolamento mecânico é geralmente usado para dissociar o tecido. Embora a digestão mecânica seja uma técnica rápida, esse método pode estar associado a rendimentos inconsistentes de criptas ou má viabilidade celular9. Portanto, o tratamento com EDTA e a dissociação mecânica podem ser combinados para gerar melhores rendimentos de criptas. Uma característica da metodologia apresentada neste artigo é o uso de agitação vigorosa dos fragmentos de tecido após quelação com EDTA10. A agitação vigorosa permite o isolamento eficiente de criptas de complexos cripta-vilosidades no intestino delgado. O grau de agitação manual determina a separação. Assim, a obtenção de criptas a partir de complexos é importante para os experimentadores neste campo. Além disso, a habilidade adequada pode reduzir a contaminação por vilosidades ao mínimo e aumentar o número de criptas.
Assim, este protocolo experimental, que emprega organoides do intestino delgado derivados da murina, pode isolar melhor criptas com força física após o tratamento com EDTA para dissociação. Sabe-se que o padrão de expressão do receptor hepatocelular B2 produtor de eritropoetina (EphB2) reflete, em parte, o ambiente das criptas. Por exemplo, as células positivas para EphB2 são organizadas de baixo para cima11. A classificação celular ativada por fluorescência (FACS) foi realizada com base na expressão de EphB2, e as células obtidas foram divididas em quatro grupos: EphB2alta, EphB2med, EphB2baixa e EphB2neg. Em seguida, o crescimento organoidede células altas EphB2 de classificação única em camundongos selvagens (WT) foi demonstrado.
Protocolo
Todos os experimentos com camundongos foram aprovados pelo Suntory Animal Ethics Committee (APRV000561), e todos os animais foram mantidos de acordo com as diretrizes do comitê para o cuidado e uso de animais de laboratório. Uma linhagem padrão WT de Mus musculus (C57BL6/J) foi usada. Foram utilizados camundongos machos e fêmeas de 10 semanas a 20 semanas de idade. Os camundongos foram eutanasiados por asfixia por CO2 .
1. Isolamento do intestino delgado
- Excisar o intestino delgado, incluindo o duodeno e a metade proximal do jejuno, com tesoura de laboratório.
- Transfira o tecido para uma placa de Petri e lave o intestino delgado com 5 mL de PBS-ABx frio (PBS + penicilina-estreptomicina [1%] + gentamicina [0,5%]) em uma seringa de 5 mL para limpar o conteúdo luminal.
- Corte o tecido aberto longitudinalmente com uma tesoura de laboratório e lave manualmente com PBS-ABx frio enquanto agita.
NOTA: Ao raspar as vilosidades com uma lâmina, a contaminação das vilosidades pode ser reduzida12. - Coletar pedaços de aproximadamente 5 mm x 5 mm do segmento intestinal com tesoura de laboratório. Transferir os fragmentos para um tubo de 50 mL com pinça e adicionar 25 mL de PBS-ABx frio.
- Lavar os fragmentos agitando para frente e para trás 10x com 25 mL de PBS-ABx frio para remover o conteúdo intestinal no tubo de 50 mL.
2. Isolamento da cripta
- Incubar as peças em PBS-ABx contendo EDTA 2 mM por 30 min em gelo sem agitação.
- Para facilitar a solidificação da matriz extracelular (MEC), incubar previamente uma placa de 24 poços em uma incubadora de cultura de tecidos a 37 °C.
- Aspirar a solução de EDTA do sistema de cultura celular com uma bomba de vácuo, adicionar 25 mL de PBS-ABx fresco e frio e, em seguida, agitar as peças para cima e para baixo vigorosamente com a mão 30x-40x para liberar os complexos cripta-vilosidades.
NOTA: As criptas e vilosidades separadas podem ser verificadas pela observação microscópica de uma gota de 25 μL da suspensão em aumento de 4x. - Em seguida, filtre a suspensão através de um filtro de 70 μm uma vez.
- Centrifugar a suspensão a 390 × g durante 3 min a 4 °C.
- Ressuspender o pellet da cripta em 20 mL de DMEM de sorbitol (DMEM avançado/F12 + penicilina-estreptomicina [1%] + gentamicina [0,5%] + soro fetal bovino [1%] + sorbitol [2%]) com pipetagem e transferir a suspensão da cripta para dois novos tubos de 15 mL para divisão em duas soluções de 10 mL para centrifugação em baixa velocidade.
NOTA: A massa celular grande e as células/detritos podem ser separadas usando centrifugação de baixa velocidade. A grande massa celular está na pelota, e as células/detritos estão no sobrenadante. - Centrifugar as duas suspensões de criptas a 80 × g durante 3 min a 4 °C e, em seguida, aspirar suavemente o sobrenadante.
OBS: Como a formação do pellet é fraca, não aspirar muito. Deixar 2 mL de sobrenadante em cada tubo. - Adicionar novamente 10 mL de sorbitol DMEM a cada tubo. Centrifugar a suspensão a 80 × g durante 3 min a 4 °C.
- Após a aspiração do sobrenadante, deixando 2 mL de sobrenadante em cada tubo, adicionar 10 mL de sorbitol DMEM para ressussão e centrifugar a suspensão da cripta a 80 × g por 3 min finais a 4 °C.
- Após a aspiração do sobrenadante, deixando 2 mL de sobrenadante em cada tubo, adicionar 10 mL de DMEM completo (DMEM avançado/F12 + penicilina-estreptomicina [1%] + gentamicina [0,5%] + soro fetal bovino [1%]) para ressuspensão do pellet por pipetagem para cima e para baixo, e deixar por 1 min.
NOTA: Aguarde 1 min para obter as criptas flutuantes de forma eficiente. - Após 1 min, coletar cada suspensão de 10 mL para um total de 20 mL e filtrar uma vez com um filtro de células de 70 μm para purificar as criptas.
- Antes de semear criptas essencialmente puras, conte o número de criptas no DMEM completo filtrado e, em seguida, centrifugue a 290 × g por 3 min a 4 °C.
- Gotículas de 25 μL em um prato de 6 cm em três pontos. Conte o número de criptas ao microscópio com aumento de 4x e calcule a concentração de criptas por gota de 25 μL.
- Suspender 100 criptas com 40 μL de ECM por poço. Pipetar para cima e para baixo 5x-10x para obter uma suspensão homogênea de criptas na MEC e, em seguida, semear em uma placa de 24 poços pré-aquecida a 37 °C.
OBS: Mantenha sempre o ECM no gelo para evitar a polimerização. Pipetar cuidadosamente para evitar a formação de bolhas de ar na MEC. - Incubar a placa de 24 poços por 15 min em uma incubadora de 37 °C e 5% de CO2 para a polimerização da MEC.
- Finalmente, cobrir a MEC com 500 μL de meio de cultura contendo fator de crescimento epidérmico (EGF) de camundongo, R-spondin 1 de camundongo recombinante e Noggin de camundongo recombinante à temperatura ambiente. A concentração final de materiais por poço é a seguinte: penicilina-estreptomicina (1%), 50 U/mL cada; gentamicina (0,5%), 25 μg/mL; EGF, 20 ng/mL; Noggin, 100 ng/mL; R-espondina 1, 500 ng/mL; L-glutamina, 2 mM.
- Inicie a cultura da cripta a 37 °C em uma incubadora de 5% de CO2 .
NOTA: Para o meio de cultura para organoides em uma placa de 24 poços, ver Tabela 1. - Realizar imagens ao vivo de longo prazo para observar a morfogênese organoide com um microscópio de imagem de lapso de tempo de gravação equipado com uma objetiva de 20x a cada 3 h por até 7 dias. Obtenha imagens seriais z-stacked em z-steps de 1 μm (1 μm x cinco passos).
- Troque o meio a cada dois dias.
3. Classificação celular ativada por fluorescência (FACS)
- Isole as criptas dos ratinhos (ver secção 2).
- Tratar as criptas isoladas com 2 mL de tripsina por 30 min a 37 °C.
- Pare a reação com 10 mL de PBS e, em seguida, passe por um filtro de células de 20 μm.
- Centrifugar a solução a 390 × g durante 3 min a 4 °C e voltar a suspender com 100 μL de DMEM completo.
- Adicionar o anticorpo anti-EphB2 APC-conjugado (1/50) e incubar por 30 min no gelo.
- Lave as células 3x com PBS e, finalmente, adicione 7-amino-actinomicina D (7-AAD) (1/100).
- Classifique as células coradas via FACS.
- Ajuste o fator de dimensionamento de área e classifique de acordo com o tamanho da célula (dispersão direta, FSC-A) versus granularidade (dispersão lateral, SSC-A).
- Classificar as células negativas e positivas de 7-AAD para viabilidade com o laser ajustado em um comprimento de onda de 488 nm e potência de 50 mV.
- Demarque as portas para classificar as células EphB2-alta (EphB2alta), EphB2-média (EphB2med), EphB2-baixa (EphB2baixa) e EphB2-negativa (EphB2neg) com o laser ajustado em um comprimento de onda de 640 nm e potência de 100 mV.
- Iniciar a cultura decélulas altas de EphB2 a 37 °C em uma incubadora de 5% de CO2 .
4. Organoides cultivados em células únicas
- Realizar o método de isolamento celular de acordo com os níveis de superfície EphB2 graduados11 e, em seguida, obter quatro populações distintas (alta, média, baixa e negativa).
- Coletar pastilha com centrifugação a 390 × g por 3 min a 4 °C e incorporar as célulasaltas EphB2 de triagem simples na MEC por pipetagem, seguida de semeadura em uma placa de 24 poços (100 singlets/40 μL de ECM/poço).
- Como na etapa 2.14, permitir que a MEC se polimerize e cobrir a MEC com um meio de cultura contendo um inibidor de quinase associada a Rho (ROCK) (10 μM) nos primeiros 2 dias para manter as célulasaltas de EphB2.
NOTA: O inibidor de ROCK é eficaz contra anoikis. - Inspecionar manualmente as células usando um microscópio invertido em aumento de 40x e observar organoides viáveis com formação esferoide e protrusão de criptas.
Resultados
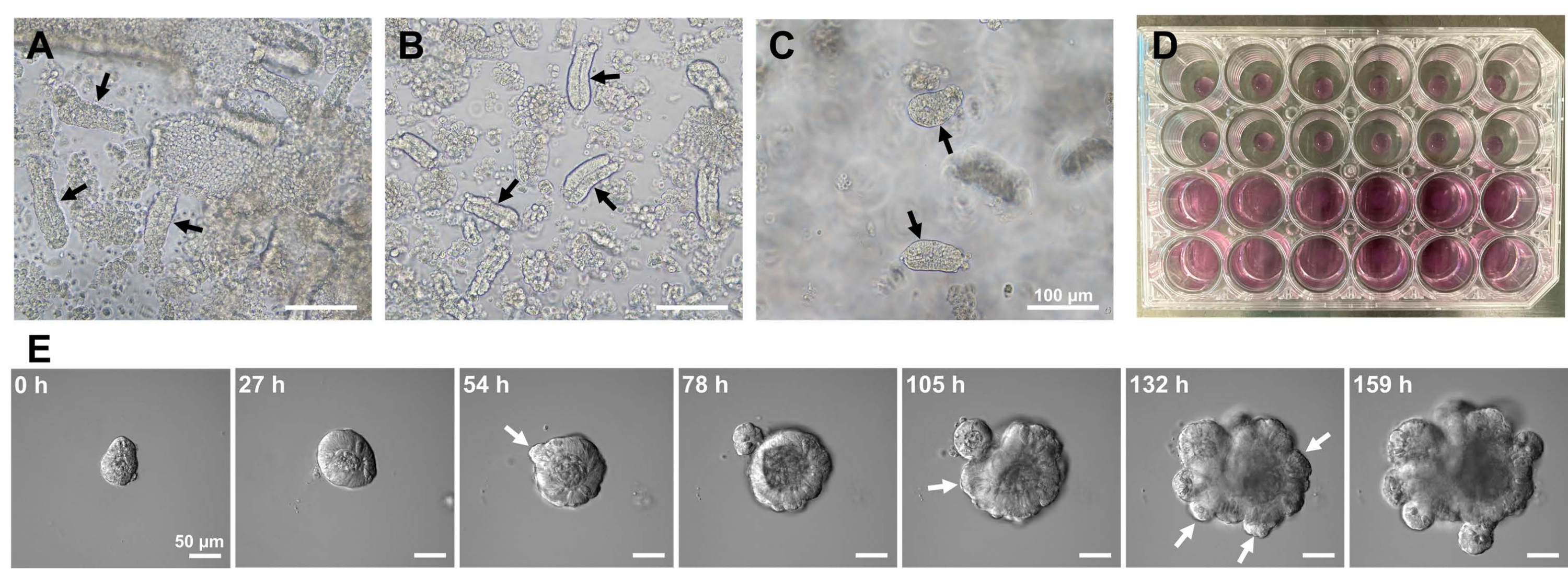
Para gerar organoides do intestino delgado de camundongos, uma combinação de tratamento com EDTA e um método de isolamento mecânico pode ser usada para isolar criptas de forma eficiente10,13. Os resultados deste estudo mostraram que quase todas as criptas isoladas foram imediatamente seladas e apareceram em forma de cone após serem espremidas para fora dos nichos epiteliais (Figura 1A). Para minimizar a contaminação das vilosidades, a suspensão resultante foi passada através de um filtro de células de 70 μm e, em seguida, o filtrado foi centrifugado. Como algumas criptas são interrompidas durante a filtragem e suspensão, essas etapas devem ser realizadas com cuidado. Os resultados mostraram que quase todas as criptas da fração final estavam integradas e adequadas para uso em cultura (Figura 1B). Para visualizar individualmente todas as criptas plaqueadas foram plaqueadas 100 criptas por poço (Figura 1C). Após a adição do meio de cultura de cripta específico (Figura 1D), o desenvolvimento dos organoides foi monitorado com um microscópio diariamente. Além disso, o crescimento organoide das criptas foi observado por imagens de lapso de tempo para monitorar seu desenvolvimento (Figura 1E e Vídeo Suplementar S1). As criptas cultas comportavam-se de forma estereotipada. O lúmen interno do organoide foi preenchido por uma massa de células apoptóticas. A proliferação ativa e diferenciação das ISCs ocorreram na região das criptas com brotamento (Figura 1E e Vídeo Suplementar S1). O brotamento foi acoplado à migração e proliferação de ISC e diferenciação de células Paneth. As células de Paneth diferenciadas estavam sempre localizadas no sítio de brotamento (Figura Suplementar S1). Como os organoides foram confirmados estáveis em cultura usando um microscópio invertido em aumento de 10x, a técnica pôde ser usada para examinar a formação de criptas no intestino delgado em desenvolvimento e determinar a capacidade de regeneração tecidual e a sobrevida a longo prazo do ISC para a produção de novas células epiteliais intestinais14,15,16.
Lgr5 é definido como um marcador ISC, e células Lgr5+ murinas formam organoides 3D7. No entanto, como a abundância da superfície celular da proteína LGR5 é baixa e há uma falta de anticorpos anti-LGR5 de alta afinidade, é um desafio isolar eficientemente ISCs murinas por FACS. A EphB2 foi previamente identificada como um marcador de superfície para a purificação de ISCs murinas e humanas de tecidos intestinais17,18. O padrão de expressão de EphB2 aumenta a complexidade envolvida nos marcadores ISC. As células EphB2-positivas organizam-se ao longo do compartimento proliferativo, com pico na parte inferior das criptas, enquanto diminuem em gradiente em direção ao topo das criptas11. Células de paneth e células progenitoras também estão localizadas na cripta. As células de Paneth expressam principalmente EphB3, que é necessário para seu posicionamento, e as células progenitoras acima delas na cripta expressam principalmente EphB2. Assim, a contaminação de ambos os tipos celulares pode ocorrer durante o curso da purificação do ISC usando o anticorpo anti-EphB2. Nesse sentido, sua expressão gênica marcadora e a capacidade formadora de organoides de células isoladas usando EphB2 por FACS devem ser avaliadas.
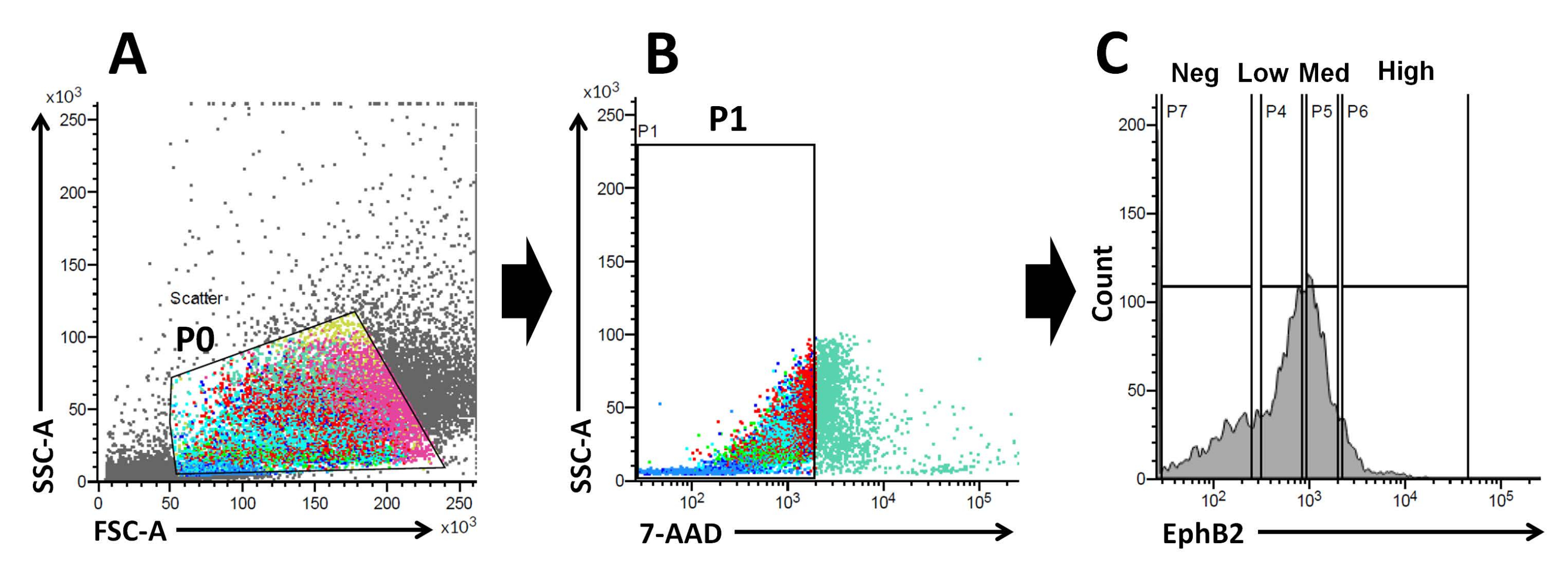
Com base nesses fatos, usando a análise FACS, células marcadas superficialmente EphB2 podem ser isoladas de criptas WT19. Foi investigado se a expressão de EphB2 pode distinguir entre quatro grupos com a expressão de marcadores específicos, tais como genes marcadores ISC-específicos (Lgr5, Ascl2 e Olfm4) e genes marcadores específicos de células progenitoras (Ki67, Myc e FoxM1). Este experimento demonstrou que as célulasaltas de EphB2 eram predominantemente ISCs, ao contrário das célulasMed EphB220,21. Finalmente, com base no método de isolamento celular, as células obtidas foram divididas em quatro grupos (células EphB2alta, EphB2med, EphB2baixa e EphB2neg) (Figura 2). Em seguida, células únicas expressando altos níveis de EphB2 classificadas por FACS foram cultivadas para crescimento organoide. Uma única célulaalta de EphB2 pode ser aplicada independentemente para tratamento localizado e recriar estruturas cript-vilosas auto-organizáveis que lembram o intestino delgado normal (Figura 3). Entretanto, as células derivadas de outros grupos (EphB2med, EphB2low e EphB2neg) não geram organoides20.
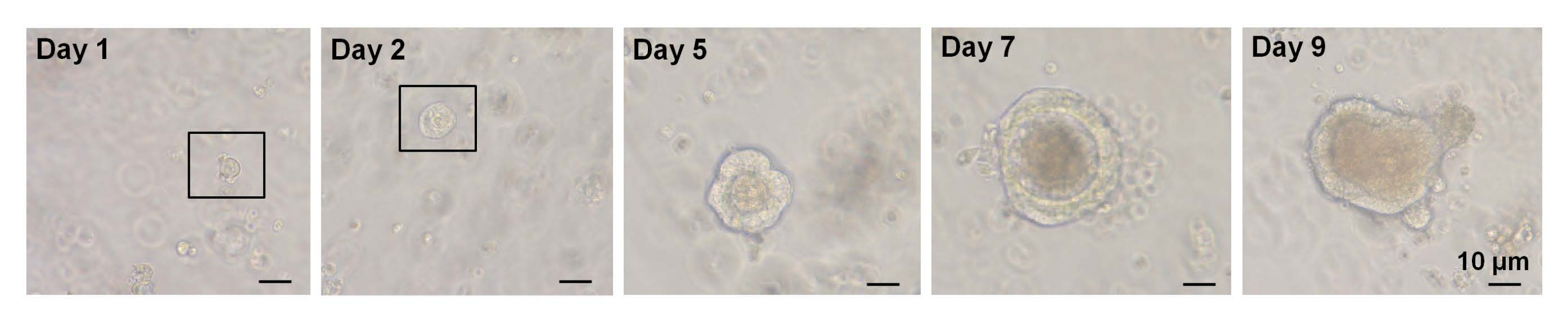
Em um estudo anterior, ~6% das células Lgr5-GFPhi de classificação única foram capazes de iniciar organoides cript-vilosos7. No entanto, as células remanescentes foram incapazes de gerar organoides e morreram nas primeiras 12 h7. Os autores presumiram que isso fosse resultado do estresse físico e/ou biológico inerente ao procedimento de isolamento7. Menos de 6% de crescimento organoide também foi obtido a partir de célulasaltas de EphB2 de triagem única em camundongos WT. No 5º dia de cultura, formaram-se estruturas esferoides (Figura 3). Do 7º ao 9º dia, ocorreu a evaginação das manchas para formação de criptas (Figura 3). É importante ressaltar que a aplicação de um inibidor de ROCK selecionado às célulasaltas EphB2 de classificação única diminuiu a apoptose induzida pela dissociação e aumentou a eficiência do crescimento organoide.

Figura 1: Geração de organoides do intestino delgado de camundongos. (A) Criptas preparadas por uma combinação de quelação de EDTA e dissociação mecânica. (B) Criptas purificadas resultantes. (C) Cripas embutidas na matriz extracelular. (A-C) As setas pretas indicam criptas. (D) Cultura tridimensional de criptas e organoides. (E) Imagens representativas de um organoide em crescimento derivado de uma cripta. As setas brancas indicam brotamento de cripta. Barras de escala = (A-C) 100 μm e (E) 50 μm. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 2: Estratégia de acoplamento por citometria de fluxo para obtenção de uma população de células EphB2-positivas (EphB2+) em camundongos selvagens. (A) Gráficos de dispersão frontal e lateral são usados para separar as células de acordo com seu tamanho e granularidade, respectivamente. (B) O espalhamento de fluorescência é usado para separar células viáveis de acordo com a intensidade de fluorescência 7-AAD (PerCP) das células. A porta para a população celular 7-AAD-negativa foi escolhida. (C) Foram escolhidas as portas para as populações de células EphB2-alta (EphB2alta), EphB2-média (EphB2med), EphB2-baixa (EphB2baixa) e EphB2-negativa (EphB2neg). Abreviações: FSC-A = área do pico de dispersão anterior; SSC-A = área do pico de dispersão lateral; 7-AAD = 7-amino-actinomicina D. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 3: Curso temporal do crescimento organoide decélulas altas de EphB2 em camundongos selvagens. Clique aqui para ver uma versão maior desta figura.
{kind=link}
Tabela 1: Meio de cultura para placa de 24 poços. Clique aqui para baixar esta tabela.
Vídeo Suplementar S1: Imagens em time-lapse de um organoide em crescimento. Barra de escala = 50 μm. Clique aqui para baixar este arquivo.
Figura suplementar S1: Imagem representativa da coloração de anticorpos antilisozima em um organoide. As setas brancas indicam células Paneth. Abreviação: DIC = microscópio de contraste de interferência diferencial. Barra de escala = 10 μm. Clique aqui para baixar este arquivo.
Discussão
Este protocolo descreve um método para isolar consistentemente criptas do intestino delgado e a subsequente cultura de organoides 3D. Para melhorar a taxa de liberação de criptas, um método de isolamento mecânico envolvendo agitação vigorosa após o tratamento com EDTA foi estabelecido. A composição do meio é diferente do protocolo original de Sato et al.7. O meio original é relativamente caro. Assim, um meio de cultura e meios personalizados para organoides murinos do intestino delgado contendo inibidores farmacológicos, fatores de crescimento recombinantes e/ou meios condicionados são mostrados na Tabela 1. Wnt3A e N-acetilcisteína não estão incluídos no meio de cultura neste protocolo. Como as células Paneth expressam Wnt3, as células produzem Wnt3 e suportam a manutenção ISC. Além disso, durante o curso do isolamento da cripta, o meio condicionado não é usado. O modelo organoide é dinâmico e apresenta heterogeneidade celular e estrutural (células de Paneth, enterócitos, células caliciformes, células enteroendócrinas, células tufo e ISCs). Assim, esses organoides podem ser usados em escala para estudar questões fundamentais da biologia organoide.
O gradiente EphB2 mantém a haste e proliferação do ISC ao longo do eixo cripta-vilosidade no intestino delgado adulto18. A vantagem de fazer organoides a partir de uma única célula EphB2 em comparação com criptas isoladas está relacionada à compreensão da biologia das ISCs murinas, já que as ISCs desempenham papéis-chave em vários distúrbios intestinais humanos. ISCs únicos dealta expressão de EphB2 podem ser cultivados para formar organoides de maneira semelhante ao desenvolvimento de organoides a partir de ISCs que expressam Lgr5 único. O passo mais importante é dividir precisamente as células em quatro grupos (EphB2alto, EphB2med, EphB2baixo e EphB2neg) de acordo com a expressão de EphB2 nas criptas usando FACS. Gráficos de dispersão direta versus lateral (FSC vs. SSC) são comumente usados para identificar células de interesse com base em seu tamanho e granularidade. FSC indica o tamanho da célula e SSC relaciona-se com a complexidade ou granularidade da célula na porta P0 (Figura 2A). Neste trabalho, as células que se encontravam dentro do portão definido (P0) foram posteriormente analisadas quanto à viabilidade. Em seguida, sua viabilidade foi determinada de acordo com as populações negativas e positivas de sinais de fluorescência 7-AAD. A fronteira entre as células 7-AAD-negativas e -positivas foi estritamente decidida para ganhar as negativas com mínima contaminação celular positiva. As portas EphB2 foram aproximadamente definidas com base na expressão graduada EphB2.
Para confirmar que os quatro grupos foram precisamente divididos, a expressão de RNAm de genes selecionados foi analisada. Os níveis de RNAm dos marcadores ISC são elevados em célulasaltas de EphB2220. Além disso, os níveis de RNAm de marcadores específicos de células progenitoras são relativamente altos em célulasEphB2 med 20. No entanto, a excompressão de EphB2 em células EphB2low e EphB2neg é baixa ou negativa em comparação com a EphB2high e EphB2med cells20. As medidas precedentes devem ser tomadas para garantir o enriquecimento daalta população celular de EphB2 antes do plaqueamento. No entanto, o crescimento organoide de menos de 6% de célulasaltas de EphB2 pode ser devido à morte de células-tronco durante o processo de cultura, e não ao tremor vigoroso durante o isolamento das criptas. Demonstrou-se que a aplicação de um inibidor seletivo da quinase associada à Rho (ROCK) em células-tronco embrionárias humanas diminui acentuadamente a apoptose induzida pela dissociação22. Assim, como uma mudança técnica, vale a pena tentar adicionar o inibidor de ROCK em uma concentração maior e com uma incubação mais longa para melhorar a viabilidade.
As células Paneth secretoras de Wnt3A ao lado das ISCs fornecem suporte essencial às ISCs8. De fato, os duplos de células ISC-Paneth exibem uma capacidade de formação de organoides fortemente aumentada em comparação com ISCs individuais8. Além disso, a adição de Wnt3A na concentração de 100 ng/mL nos primeiros 3 dias de cultura demonstrou aumentara capacidade de formação de organoides8. Assim, como outra mudança técnica, a adição de Wnt3A exógeno poderia melhorar a capacidade de formação de organoides de ISCs EphB2de alta expressão.
Comparados a abordagens in vivo, os organoides podem ser facilmente utilizados para manipulação genética, análise de fenótipos de malignidade e triagem de drogas20,23. Uma combinação de quelação de EDTA e um método de isolamento mecânico é eficaz, reprodutível e eficiente em termos de tempo para criar organoides do intestino delgado a partir de criptas e pode ser facilmente seguida pela equipe de laboratório sem qualquer experiência avançada. Assim, a adição do isolamento mecânico com agitação vigorosa após o tratamento com EDTA pode estabelecer eficientemente organoides murinos do intestino delgado ex vivo e fornecer uma ferramenta potencial para o cultivo de organoides e modelagem de doenças de outros tecidos epiteliais adultos.
As células epiteliais intestinais são polarizadas e orientadas com o lado apical direcionado para a luz. No entanto, o lado apical voltado para o lúmen dos organoides 3D está em seu interior. Assim, essa organização impede o acesso ao lado apical, o que é um problema quando se estudam os efeitos de componentes luminais, como nutrientes, micróbios e metabólitos sobre as células epiteliais. Para contornar essa desvantagem, uma cultura de células organoides como monocamadas2D foi desenvolvida 24. Em termos de aplicações futuras, a cultura de monocamadas de células organoides será utilizada, pois representa o sistema mais eficiente e tratável.
Divulgações
Os autores declaram não haver conflitos de interesse.
Agradecimentos
Este trabalho foi apoiado por Grants-in-Aid for Scientific Research (C) to T.T. (grant numbers JP17K07495 e JP20K06751). Agradecemos ao Prof. Mineko Kengaku pelo uso de equipamentos para a obtenção de imagens time-lapse de longa duração (LCV100; Olimpo).
Materiais
| Name | Company | Catalog Number | Comments |
| 1.5 mL Eppendorf tube | Eppendorf | 0030 125.215 | |
| 5 mL syringe | TERUMO | SS-05SZ | |
| 15 mL Falcon tube | Iwaki | 2325-015 | |
| 20 μm cell strainer | Sysmex | 04-004-2325 | |
| 24-well plate | Iwaki | 3820-024 | |
| 50 mL Falcon tube | Iwaki | 2345-050 | |
| 60 mm tissue culture dish | FALCON | 353002 | |
| 70 μm cell strainer | Falcon | 352350 | |
| 100 mm Petri dish | Iwaki | 3020-100 | |
| 7-AAD | BD Biosciences | 559925 | |
| Advanced DMEM/F12 | Gibco | 12634-010 | |
| Alexa Fluor 568 Goat Anti-Mouse IgG (H+L) | Invitrogen | A-11004 | |
| Anti-EphB2 APC-conjugated antibody | BD Biosciences | 564699 | |
| C57BL6/J mice | Japan SLC, Inc. | ||
| Clean bench | HITACHI | CCV-1306E | |
| Confocal laser scanning microscope | Olympus | FV3000 | |
| EDTA (0.5 mol/L) | Nacalai Tesque | 06894-14 | 2 mM |
| FACSMelody | BD Life Sciences-Biosciences | 661762 | |
| Fetal bovine serum | Sigma | 173012 | 1% (v/v) |
| Fiji (software) | https://fiji.sc/ | ||
| Gentamicin (10 mg/mL) | Nacalai Tesque | 16672-04 | 25 μg/mL |
| Hammacher laboratory scissor | SANSYO | 91-1538 | |
| Incubator | Panasonic | MCO-170-PJ | |
| Laboratory tweezer | AS-ONE | 7-164-04 | |
| L-Glutamine 200 mM | Gibco | 25030081 | 2 mM |
| Matrigel | BD Biosciences | 354230 | ECM for 3D organoids |
| Mouse Anti-Human Lysozyme | LSBio | LS-B8704-100 | |
| Murine EGF (20 μg/mL stock solution) | PeproTech | 315-09 | 20 ng/mL |
| PBS 1x | Gibco | 10010-023 | |
| Penicillin-Streptomycin (10,000 U/mL) | Gibco | 15140-122 | 50 U/mL |
| Pipetman (10 μL, 20 μL, 200 μL, and 1,000 μL) | GILSON | 1-6855-12, -13, -15, and -16 | |
| Recombinant murine Noggin (20 μg/mL stock solution | R&D Systems | 1967-NG-025 | 100 ng/mL |
| Recombinant murine R-Spondin 1 (250 μg/mL stock solution) | R&D Systems | 3474-RS-050 | 500 ng/mL |
| Sorbitol | Nacalai Tesque | 32021-95 | 2% (w/v) |
| TE2000-S (inverted microscope) | Nikon | 24131 | |
| Time-lapse image microscope | Olympus | LCV100 | |
| TrypLE Express 1x | Gibco | 12605-010 | |
| ULVAC | ULVAC KIKO Inc. | 100073 | |
| Y-27632 | Fujifilm | 331752-47-7 | 10 μM |
Referências
- Clevers, H. Modeling development and disease with organoids. Cell. 165 (7), 1586-1597 (2016).
- Seidlitz, T., et al. Human gastric cancer modelling using organoids. Gut. 68 (2), 207-217 (2019).
- Nikolaev, M., et al. Homeostatic mini-intestines through scaffold-guided organoid morphogenesis. Nature. 585 (7826), 574-578 (2020).
- Artegiani, B., Clevers, H. Use and application of 3D-organoid technology. Human Molecular Genetics. 27, R99-R107 (2018).
- Lancaster, M. A., Knoblich, J. A. Organogenesis in a dish: modeling development and disease using organoid technologies. Science. 345 (6194), 1247125 (2014).
- Dedhia, P. H., Bertaux-Skeirik, N., Zavros, Y., Spence, J. R. Organoid models of human gastrointestinal development and disease. Gastroenterology. 150 (5), 1098-1112 (2016).
- Sato, T., et al. Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature. 459 (7244), 262-265 (2009).
- Sato, T., et al. Paneth cells constitute the niche for Lgr5 stem cells in intestinal crypts. Nature. 469 (7330), 415-418 (2011).
- Aronowitz, J. A., Lockhart, R. A., Hakakian, C. S. Mechanical versus enzymatic isolation of stromal vascular fraction cells from adipose tissue. Springerplus. 4 (1), 713 (2015).
- Takahashi, T. New trends and perspectives in the function of non-neuronal acetylcholine in crypt-villus organoids in mice. Methods in Molecular Biology. 1576, 145-155 (2019).
- Batlle, E., et al. β-catenin and TCF mediate cell positioning in the intestinal epithelium by controlling the expression of EphB/ephrinB. Cell. 111 (2), 251-263 (2002).
- Baghdadi, M. B., Kim, T. -. H. Analysis of mouse intestinal organoid culture with conditioned media isolated from mucosal enteric glial cells. STAR Protocols. 3 (2), 101351 (2022).
- Takahashi, T., et al. Non-neuronal acetylcholine as an endogenous regulator of proliferation and differentiation of Lgr5-positive stem cells in mice. FEBS Journal. 281 (20), 4672-4690 (2014).
- Barker, N. Adult intestinal stem cells: Critical drivers of epithelial homeostasis and regeneration. Nature Reviews Molecular Cell Biology. 15 (1), 19-33 (2014).
- Fordham, R. P., et al. Transplantation of expanded fetal intestinal progenitors contributes to colon regeneration after injury. Cell Stem Cell. 13 (6), 734-744 (2013).
- Miyoshi, H., et al. Wnt5a potentiates TGF-β signaling to promote colonic crypt regeneration after tissue injury. Science. 338 (6103), 108-113 (2012).
- Jung, P., et al. Isolation and in vitro expansion of human colonic stem cells. NatureMedicine. 17 (10), 1225-1227 (2011).
- Merlos-Suárez, A., et al. The intestinal stem cell signature identifies colorectal cancer stem cells and predicts disease relapse. Cell Stem Cell. 8 (5), 511-524 (2011).
- Mao, W., et al. EphB2 as a therapeutic antibody drug target for the treatment of colorectal cancer. Cancer Research. 64 (3), 781-788 (2004).
- Takahashi, T., et al. Muscarinic receptor M3 contributes to intestinal stem cell maintenance via EphB/ephrin-B signaling. Life Science Alliance. 4 (9), e202000962 (2021).
- Jung, P., et al. Isolation of human colon stem cells using surface expression of PTK7. Stem Cell Reports. 5 (6), 979-987 (2015).
- Watanabe, K., et al. A ROCK inhibitor permits survival of dissociated human embryonic stem cells. Nature Biotechnology. 25 (6), 681-686 (2007).
- Schulte, L., Hohwieler, M., Müller, M., Klaus, J. Intestinal organoids as a novel complementary model to dissect inflammatory bowel disease. Stem Cells International. 2019, 8010645 (2019).
- Puzan, M., Hosic, S., Ghio, C., Koppes, A. Enteric nervous system regulation of intestinal stem cell differentiation and epithelial monolayer function. Scientific Reports. 8 (1), 6313 (2018).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados