
Method Article
В естественных условиях переноса гена к шванновских клеток в нерве Грызун седалищного путем электропорации
В этой статье
Резюме
Here, we present an in vivo technique for gene transfer to Schwann cells (SCs) in the rodent sciatic nerve. This simple technique is useful for investigating signaling mechanisms involved in the development and maintenance of myelinating SCs.
Аннотация
The formation of the myelin sheath by Schwann cells (SCs) is essential for rapid conduction of nerve impulses along axons in the peripheral nervous system. SC-selective genetic manipulation in living animals is a powerful technique for studying the molecular and cellular mechanisms of SC myelination and demyelination in vivo. While knockout/knockin and transgenic mice are powerful tools for studying SC biology, these methods are costly and time consuming. Viral vector-mediated transgene introduction into the sciatic nerve is a simpler and less laborious method. However, viral methods have limitations, such as toxicity, transgene size constraints, and infectivity restricted to certain developmental stages. Here, we describe a new method that allows selective transfection of myelinating SCs in the rodent sciatic nerve using electroporation. By applying electric pulses to the sciatic nerve at the site of plasmid DNA injection, genes of interest can be easily silenced or overexpressed in SCs in both neonatal and more mature animals. Furthermore, this in vivo electroporation method allows for highly efficient simultaneous expression of multiple transgenes. Our novel technique should enable researchers to efficiently manipulate SC gene expression, and facilitate studies on SC development and function.
Введение
The rapid transmission of sensory and motor information in the peripheral nervous system is permitted by the myelin sheath, which is formed by myelinating Schwann cells (SCs)1. Insulation of axons by the myelin sheath enables saltatory conduction, which increases the speed of nerve impulses. In disorders in which the development or maintenance of the myelin sheath is impaired, nerve conduction speed is reduced. This results in neuropathy involving motor and sensory dysfunction. Although there are many studies on the molecular mechanisms of myelination and demyelination in the peripheral nervous system, the roles of the numerous proteins involved in these processes remain unclear.
To study the molecular mechanisms of SC myelination/demyelination in vivo, genetic approaches have been used to modify gene expression in animals. A powerful approach is the use of knockout/knockin or transgenic animals. However, the generation of these animals is expensive and time consuming. For SC-specific gene manipulation, crossing floxed strains with Cre mice or other conditional gene expression methods are necessary. This again is laborious and time intensive. In recent years, a cutting-edge genetic technology, the CRISPR-Cas9 system, has made the generation of genetically modified mice much quicker (about 4 weeks)2,3, but this method is hindered by target sequence limitations, and suffers from off-target effects. As an alternative method, viral vector-mediated gene transfer is a faster and easier method of achieving gene transfer into SCs in vivo4-6. Indeed, the generation of viral vectors is less expensive, and takes a shorter time (within a few weeks), and gene manipulation of SCs can be achieved by simply injecting engineered viral vectors, such as adenoviral vectors, adeno-associated viral (AAV) vectors, and lentiviral vectors, into the sciatic nerve. Because these viral vectors have different characteristics, users have to choose the one best suited for their purpose. Adenoviral vectors infect axons and SCs in both young and mature sciatic nerves. In particular, adenoviral vectors have higher selectively for non-myelinating SCs than myelinating SCs. Adenoviruses can cause immune responses, and accordingly, immunodeficient strains should be used5. AAV vectors are currently the most widely used viral vectors, and allow in vivo gene transfer with lower toxicity7. AAV can transduce both axons and SCs by direct injection into the nerve fibers8,9. However, AAV-mediated protein expression usually requires 3 weeks or longer to reach maximum levels7,9. Therefore, it is difficult to analyze myelination, which actively progresses during the two week postnatal period. Lentiviral vectors have higher selectively for myelinating SCs than non-myelinating SCs, and do not have toxic effects on sciatic nerves. However, lentiviral vectors do not infect SCs in more mature nerves5, and therefore are unsuitable for analyzing events such as the demyelination process.
Electroporation is another faster and easier approach to achieve in vivo gene transfer. It has been reported that in vivo transfection of SCs can be achieved when electroporation is applied to transected rat sciatic nerves10. However, because this method requires nerve transection for gene delivery, the application is limited to the analysis of the damaged nerves. Here, we describe an alternative method that allows the delivery of transgenes into myelinating SCs in intact rat sciatic nerves using electroporation11. This method requires plasmid construction, which can usually be completed within a week. Then, by simply delivering electric pulses to the site on the sciatic nerve where the plasmid DNA was injected, highly selective transfection of myelinating SCs can be achieved in neonatal as well as in more mature animals. By electroporating multiple plasmids, simultaneous expression of a variety of genes can be easily achieved. The ability to simultaneous express multiple molecules, such as signaling proteins, short-hairpin RNAs (shRNAs) and functional probes, is crucial for investigating complex processes such as myelination and demyelination. The novel in vivo electroporation method described in this paper will be a powerful tool, allowing researchers to analyze the function of a multitude of molecules and their interactions in myelinating SCs.
протокол
Использование крыс для исследования было сделано в соответствии с правилами, установленными Комитетом по охране животных из Университета Токио.
1.Preparation плазмидной ДНК
- Генерирование ДНК плазмид для электропорации в естественных условиях путем субклонирования кДНК или последовательность shRNA в плазмиде экспрессии для клеток млекопитающих 12. Используйте цитомегаловируса ранние усиливающие и куриные бета-актина фьюжн (CAG) промотор-приводом плазмиды 13 , поскольку она позволяет сильной и стабильной экспрессии. Для экспрессии shRNAs под контролем промотора CAG, используют mir30 на основе системы кассетного shRNA дл субклонировани в shRNA 14.
- Очищают плазмидной ДНК с помощью макси-Prep Kit в соответствии с инструкциями производителя, и ресуспендируют ДНК с HEPES-буферном солевом растворе (140 мМ NaCl, 0,75 мМ Na 2 HPO 4, 25 мМ HEPES, рН 7,40). Отрегулируйте концентрацию ДНК ≥ 4 мкг / мкл.
- Приготовьте раствор плазмидной ДНК в концентрации 4 мкг / мкл, и добавить минимальное количество быстрого зеленого красителя (конечная концентрация 0,01%), чтобы маркировать место инъекции. Когда требуется одновременное электропорация нескольких плазмид, регулировать общую концентрацию раствора плазмидной ДНК до 4 мкг / мкл.
Примечание: оптимальный состав плазмидных ДНК должна быть определена в соответствии с эффективности трансфекции каждой плазмиды.
2. Стерилизация хирургических инструментов и физиологический раствор
- Автоклавы хирургические инструменты и 0,9% раствор NaCl.
3. Подготовка стекла микропипетки
- Потянуть стеклянные пипетки с помощью пипетки съемник. Обрежьте кончик пипетки диаметром 30-50 мкм. Используйте следующие параметры: Тепло, 600; Скорость, 50; Время, 75.
4. Хирургическое лечение животных, инъекции ДНК и электропорация
Примечание: за кадромвид этого шага описана на рисунке 1. Хотя процедура для крысят описаны здесь, способ также применим к более зрелых животных с использованием той же процедуры.
- Обезболить крыс с изофлуран в индукционной коробке, пока животное не будет неподвижен путем регулировки потока кислорода до 0,4 л / мин и изофлуран концентрации до 4% (об / об). Выполните палец щипать, чтобы подтвердить правильность обезболивание.
- Поместите крысу на подогретый теплее под бинокулярным микроскопом, и поддержания анестезии путем непрерывного введения изофлуран через маску. Отрегулировать поток кислорода до 0,2 л / мин и изофлуран концентрации до 2% (об / об). Используйте глазные капли, чтобы предотвратить сухость глаз, если глаза животного открыты.
- Зафиксируйте ноги с хирургической лентой.
- Очистите кожу на задней бедра с повидон-йодом, и сделать надрез скальпелем.
Примечание: Бритье хирургические области, если хирургические участки покрыты чвоздух. - Выставляют седалищный нерв, создавая отверстие между четырехглавой мышцы бедра и двуглавая мышца бедра с швейными иглами.
- Влажные нерва с 0,9% раствором NaCl. Поглощенный избыток воды с безворсовой бумаги.
- Вставьте основание стеклянной микропипетки на гибкой трубки, и заполнить достаточное количество раствора ДНК (по крайней мере, один микролитр) в микропипетки, осторожно аспирационных.
- Поднимите обнаженный нерв, осторожно потянув дистальной стороны нерва с помощью иглы.
Примечание: Не прикладывайте напряжение к нерву, чтобы минимизировать механическое напряжение. - Вставьте стекло микропипетки в дистальный участок на нерв, и впрыскивают раствор ДНК, применяя давление (например , путем продувки в открытый конец гибкой трубки). Вводят раствор ДНК до тех пор, пока не появится нерв зеленым (максимум 1 мкл). Поскольку частое введение микропипетки может повредить нерв, не вставляйте микропипетка более чем в два раза.
- Поместите TWEэцер типа платинового электрода около 1-2 мм друг от друга от нерва. Заполните зазор между электродом и нервом с 0,9% -ным раствором NaCl.
Примечание: Не держите нерва с электродом, чтобы избежать механических напряжений на нерв. - Применение электрических импульсов на месте инъекции с использованием электропоратора с электродом. После первого набора импульсов, инвертировать электрода и применить другой набор импульсов. Используйте следующие параметры: напряжение, 50 В; длительность импульса, 5 мс; интервал между импульсами, 100 мс; число импульсов, в 4 раза.
- Очистите сайт электропорации с 0,9% раствором NaCl.
- Повторите шаги 4.4-4.11 на контралатеральной седалищного нерва.
5. После электропорации
- Закройте надрезы с цианакриловым клеем.
- После высыхания клея, очистить рану с повидон-йодом.
- Отпустите детеныша от лицевой маски. Подогреть на детеныша теплее по крайней мере, в течение часа, с тем чтобы позволить ему полностью восстановиться от анестезии. Dо не оставляйте без присмотра щенка, пока он не пришел в сознание достаточное.
- После выхода из наркоза, вернуть щенка к матери крысы. Не возвращать щенка, пока полностью не выздоровел.
6. Послеоперационный
- Дом в крысят в клетке до проведения опытов 11 (смотри примеры на рисунке 3). Администрирование карпрофен (5 мг / кг; бр), нестероидный противовоспалительный препарат, или бупренорфин (0,1 мг / кг, подкожно), опиоидного анальгетика, в случае необходимости.
Примечание: Если щенок крысы не растут хорошо или воспаление наблюдается вокруг места операции, исключить животное из экспериментов.
Результаты
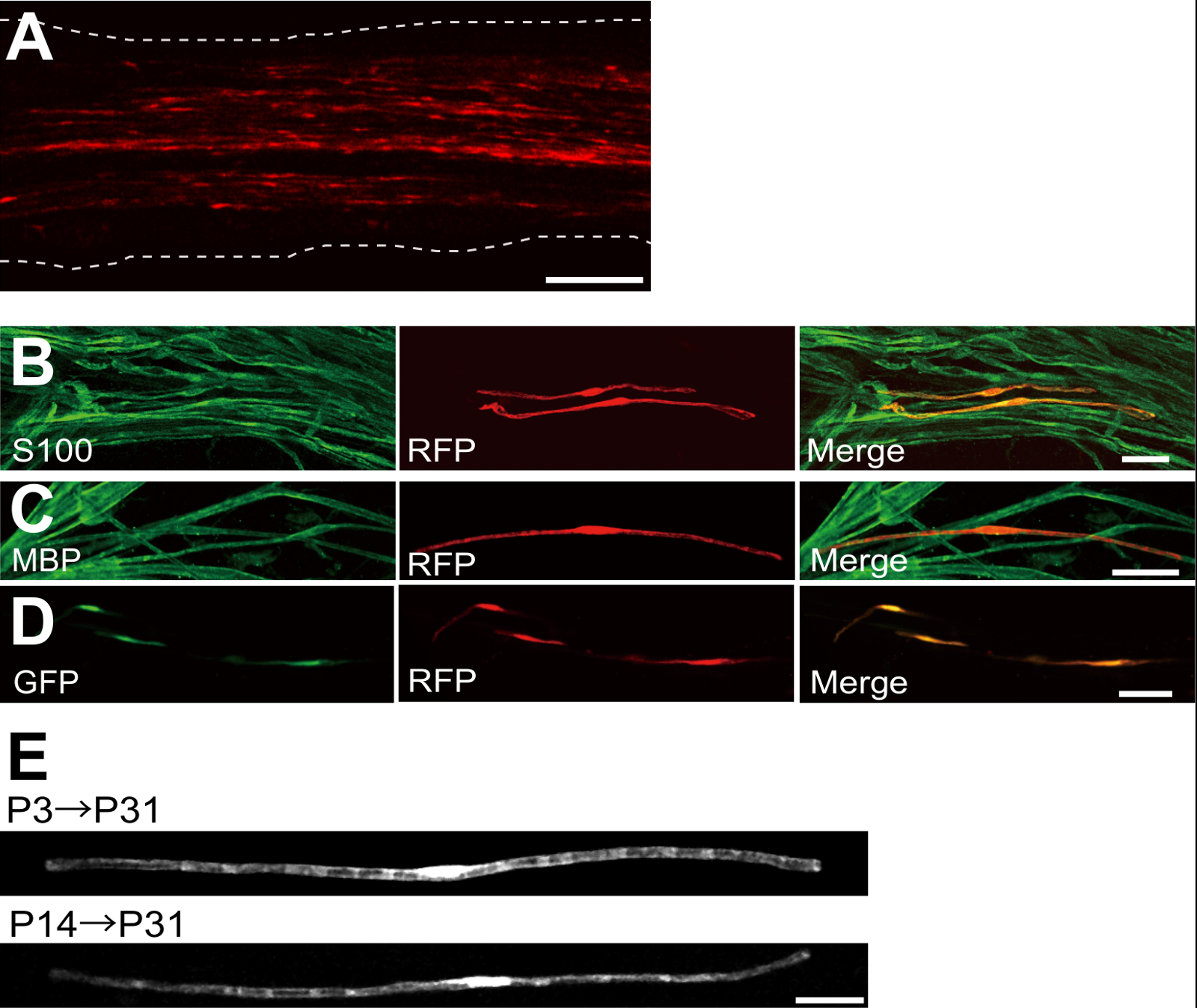
Примером седалищного нерва, трансфицированных красный флуоресцентный белок (RFP) -expressing плазмиды показана на фигуре 2А. Клетки, демонстрирующие биполярного морфологии, характеристика ГКС, были слабо трансфицировали RFP. Ни один ЗП флуоресценции не был обнаружен в аксонах. Как правило, мы находим ~ 100 трансфецированных в каждом SCs нерва. Эта эффективность трансфекции кажется похож на SC эффективность инфекции в естественных условиях с использованием лентивирусов векторов 4.
Иммунным окрашиванием эксперименты показали , что большинство (~ 96%) RFP-положительных клеток при Р7 со-меченого для S100, КА маркера (Фигура 2В), и 91% от RFP-положительных клеток при Р14 со-меченого для МВР, в myelinating SC маркера (фиг.2С), предполагая , что перенос генов с помощью электропорации является высоко селективным для myelinating ГКС.
Введение нескольких генов в SCs в естественных условиях будет чрезвычайно полезен для изучения механизмов миелинизации / демиелинизации. Основным преимуществом естественных условиях методом электропорации в описанном здесь является способность передавать несколько генов с помощью простой процедуры. На рисунке 2D показан репрезентативный изображение седалищного нерва , трансфицированных смесью GFP и RFP-экспрессирующих плазмид с использованием в естественных условиях электропорации. Около 97% ГКС были GFP и RFP двойной положительный, предполагая, что высокую эффективность доставки множества генов может быть достигнуто путем простого электропорации смеси нескольких плазмид.
У грызунов, миелинизации инициирует вокруг рождения, резко возрастает в течение первых двух недель после рождения, а затем постепенно уменьшается. Таким образом, с помощью методов генной манипуляции во время этих SCs развития временных окон, механизмы, лежащие в основе этих различных стадий миелинизации могут быть уточнены. Lentiviral векторы являются хорошим инструментом дляnalyzing миелинизации, в частности , так как они имеют минимальную токсичность, но лентивирусов инфицируют только неонатальной седалищного нервов 5,6. Для сравнения, электропорации-опосредованный перенос генов хорошо работает , когда трансфекция проводится на P3 (рис 2Е, сверху) или на Р14 (рис 2Е, внизу).
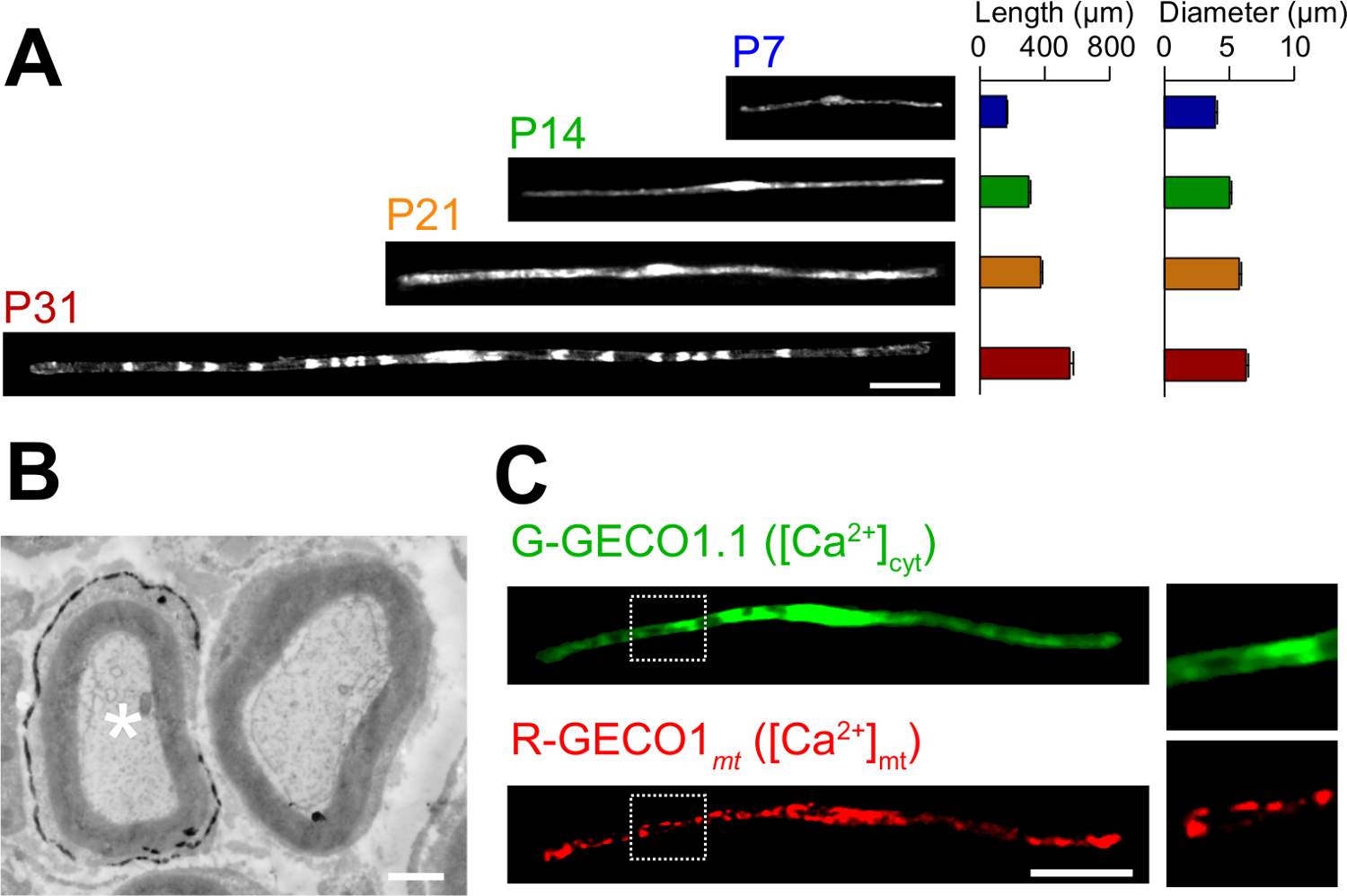
Применения романа в естественных условиях методом электропорации описаны здесь. На рисунке 3А показаны легкие микроскопические изображения GFP-экспрессирующие myelinating SCs на различных стадиях развития (P7, P14, P21 и P31). При легкой микроскопического анализа, изменения морфологических параметров, таких как длина и диаметр, можно оценить. Обратите внимание , что эти параметры имеют близкие значения по сравнению с неповрежденной крыс периферических нервов 15,16, предполагая , что электропорации нервы развиваются без значительных вредных эффектов. На рис 3B показана электронная микроскопическое изображение LacZ-expressing myelinating SCS. В этом случае, LacZ, был использован в качестве маркера экспрессии. β-галактозидазы окрашивание с использованием Bluo-гал, этанол нерастворимый субстрат, дает анализ структуры миелина трансфицированных SCs с помощью электронной микроскопии 11,17. В этих экспериментах роль сигнальных молекул могут быть исследованы с помощью глушителей или увеличении их выражение, таким образом, позволяя анализировать с потерей функции или усиления из-функции эффектов. В дополнение к анализу фиксированной ткани, в естественных условиях электропорации-опосредованный перенос генов может быть также применен жить эксперименты визуализации. Например, на рисунке 3C показывает myelinating SC коэкспрессирующей G-GECO1.1 18, зеленый флуоресцентный индикатор цитозольного Ca 2+ и R-19, GECO1mt красный флуоресцентный митохондриальный индикатор Ca 2+. Выражая эти показатели, мы определили сигнальный путь , который управляет цитозольных и митохондриальных Ca 2+ концентрации в myelinating SCs , Таким образом, настоящий способ может быть использован для изучения различных сигнальных механизмов, в частности, когда генетически кодируемые флуоресцентные зонды доступны для обнаружения сигналов, представляющих интерес.

Рисунок 1:. Схема I N естественных условиях электропорации первый метод, седалищного нерва у анестезированных крыс подвергается. Во-вторых, плазмидная ДНК вводят в седалищный нерв. В-третьих, электрические импульсы поступают на месте инъекции через щипцов-образный электрод. Наконец, рана закрыта с клеем. Эта процедура может быть повторена на контралатеральной нерва. Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой фигуры.
{kind=link}

Рисунок 2: Представитель Результаты по трансфицированных седалищного нервах (А) представитель образ трансфицированных седалищного нерва.. Нерв трансфицировали RFP-экспрессирующих плазмид в P3, и фиксируется на Р7. (Б) представитель образ RFP-трансформированных клеток при Р7 показывая колокализацию с S100, в SC маркером. (C) представитель образ RFP-трансфицировали седалищного нерва при P14 , показывая колокализацию с МВР, в myelinating SC маркера. (D) представитель образ седалищного нерва котрансфицируют с GFP и RFP , экспрессирующих плазмид. Трансфицированных Стволовые одновременно выражали GFP и RFP. (E) Изображение myelinating SCs на P31 , трансфицированные в P3, когда начинается миелинизации (вверху), и образ myelinating SCs на P31 трансфицировали в P14, когда большинство крупных аксонов становятся миелинизированы (внизу), предполагая , что Transfпрогиб из myelinating SCs может быть достигнуто не только в неонатальной нервов, но и в более зрелых нервов. Шкала баров = 200 мкм (А); 50 мкм (BE). Эта цифра была изменена с нашей предыдущей публикации 11. Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой фигуры.
{kind=link}

Рисунок 3: Применение в естественных условиях электропорации (а) световой микроскопический анализ развития myelinating ГКС.. Седалищный нервы были электропорации с GFP-экспрессирующие плазмиды в P3, и были зафиксированы на разных стадиях развития (P7, P14, P21 и P31). Типичные изображения GFP-положительных SCs показаны на левой стороне. Средняя длина и диаметр суммированы в виде среднего значения ± SEM (n = 30 - 47 из 3-х нервов) справа. Длина и диаметр myelinating SCS увеличивается по мере развития. (В) Электронно - микроскопический образ седалищного нерва , трансфицированных плазмидой , кодирующей LacZ. Трансфицированной SC (белая звездочка, слева) тонко метят выделениями продукта реакции β-галактозидазы. (C) Изображение SC котрансфицируют с G-GECO1.1, зеленый флуоресцентный цитозольного индикатора Са 2+ и R-GECO1mt, красный флуоресцентный митохондриального индикатора Са 2+. Области внутри белых пунктирными прямоугольниками показаны в увеличенном виде панелей справа. Масштабные полоски = 50 мкм (А и С); 1 мкм (Б). Эта цифра была изменена с нашей предыдущей публикации 11. Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой фигуры.
{kind=link}
Обсуждение
In this paper, we describe a simple and efficient method that allows in vivo gene transfer to myelinating SCs in the rat sciatic nerve using electroporation. This method allows highly selective gene expression in myelinating SCs by simply applying electric pulses to the plasmid DNA-injected sciatic nerve. Because the molecular mechanisms of myelination and demyelination in the peripheral nervous system remain unclear, the present in vivo electroporation method will be a powerful tool to clarify the roles of multiple genes of interest in living animals.
A critical requirement of this method is to keep damage to the nerve during surgery to a minimal level. Should surgical damage cause excessive inflammation, the sciatic nerve may degenerate. To avoid this, one must conduct surgery with extreme care, so as to not damage the blood vessels around the nerve. Mechanical stress to the nerve during the surgery can also be a cause of nerve damage. To minimize mechanical stress, lifting the exposed nerve should be done as gently as possible, and the tweezer-type electrode should be placed close to the nerve without contact. Furthermore, electrical pulses that are too strong can cause undesirable large leg movement, which leads to mechanical stress, or can burn the nerve. If significant damages are observed in the nerves, we recommend reducing the electrical pulse intensities or placing the electrode further away from the nerve.
In our present protocol, CAG promoter-driven plasmids were used as expression vectors. CAG promoter-driven plasmids allow high levels of gene expression in myelinating SCs in vivo. We also have tried a CMV promoter, another widely used universal promoter for mammalian gene expression, but expression of the gene product was very weak. This is consistent with previous results, in which electroporation-mediated transfection was conducted in the embryonic brain20. Therefore, we recommend using CAG promoter-driven plasmids for the in vivo electroporation method.
Because axonal signaling is a key factor in myelination/demyelination21, gene modification in neurons is also important. However, delivery of transgenes using our in vivo electroporation method is limited to SCs. It has been reported that gene delivery into sciatic nerve axons can be achieved when in vivo electroporation is applied to dorsal root ganglion (DRG) neurons in adult rats22. This suggests that delivery of plasmid DNA to the cell body is likely to be critical for in vivo transfection of peripheral axons. Thus, to examine the involvement of axonal molecules in myelination/demyelination, researchers should use neuron-specific genetic methods such as genetically modified animals, neuron-specific viral vectors, or in vivo electroporation to DRG neurons.
Compared with current methods, such as the generation of genetically modified animal lines23 and delivery of transgenes by viral vectors4-6, gene modification of SCs by in vivo electroporation is simpler. This method only requires several days for plasmid DNA construction and one day for electroporation surgery. Plasmid DNA construction does not require a biohazard room that is usually essential for viral vector handling. In addition, one of the advantages of the electroporation method is the capacity for simultaneous expression of multiple gene products using a simple protocol. Our novel technique will be useful for analyzing the interaction of a variety of signaling molecules involved in myelination and demyelination. In particular, by permitting the cotransfection of a number of different intracellular fluorescent probes, our method should be a powerful tool for investigating intracellular signaling dynamics in SCs using live imaging experiments.
Раскрытие информации
The authors declare that they have no competing financial interests.
Благодарности
This work was supported by grants from the Ministry of Education, Culture, Sports, Science and Technology to M.I. (21229004 and 25221304).
Материалы
| Name | Company | Catalog Number | Comments |
| Genopure Plasmid Maxi Kit | Roche | 03 143 422 001 | Plasmid DNA purification kit |
| Fast Green CFC | WAKO | 069-00032 | Dye for DNA injection |
| GC 150T-10 | HARVARD APPARATUS | 30-0062 | Glass capillary |
| Suction tubing | Drummond | 05-2000-00 | Suction tubing for micro injection |
| MODEL P-97 | SUTTER INSTRUMENT CO. | Micropipette puller | |
| CUY21 Single Cell | BEX | Electroporator CUY21 Single Cell | Pulse generator |
| Electric warmer | KODEN | CAH-6A | Warmer during the surgery |
| Isofluolane | Mylan | 1119701G1076 | Anesthetic |
Ссылки
- Nave, K. A., Werner, H. B. Myelination of the nervous system: mechanisms and functions. Annu Rev Cell Dev Biol. 30, 503-533 (2014).
- Wang, H., et al. One-step generation of mice carrying mutations in multiple genes by CRISPR/Cas-mediated genome engineering. Cell. 153, 910-918 (2013).
- Yang, H., Wang, H., Jaenisch, R. Generating genetically modified mice using CRISPR/Cas-mediated genome engineering. Nat Protoc. 9, 1956-1968 (2014).
- Cotter, L., et al. Dlg1-PTEN interaction regulates myelin thickness to prevent damaging peripheral nerve overmyelination. Science. 328, 1415-1418 (2010).
- Gonzalez, S., Fernando, R. N., Perrin-Tricaud, C., Tricaud, N. In vivo introduction of transgenes into mouse sciatic nerve cells in situ using viral vectors. Nat Protoc. 9, 1160-1169 (2014).
- Ozcelik, M., et al. Pals1 is a major regulator of the epithelial-like polarization and the extension of the myelin sheath in peripheral nerves. J Neurosci. 30, 4120-4131 (2010).
- Daya, S., Berns, K. I. Gene therapy using adeno-associated virus vectors. Clin Microbiol Rev. 21, 583-593 (2008).
- Glatzel, M., et al. Adenoviral and adeno-associated viral transfer of genes to the peripheral nervous system. Proc Natl Acad Sci U S A. 97, 442-447 (2000).
- Homs, J., et al. Schwann cell targeting via intrasciatic injection of AAV8 as gene therapy strategy for peripheral nerve regeneration. Gene Ther. 18, 622-630 (2011).
- Aspalter, M., et al. Modification of Schwann cell gene expression by electroporation in vivo. J Neurosci Methods. 176, 96-103 (2009).
- Ino, D., et al. Neuronal Regulation of Schwann Cell Mitochondrial Ca(2+) Signaling during Myelination. Cell Rep. 12, 1951-1959 (2015).
- Struhl, K. Chapter 3; Subcloning of DNA fragments. Curr Protoc Mol Biol. , Unit3 16 (2001).
- Niwa, H., Yamamura, K., Miyazaki, J. Efficient selection for high-expression transfectants with a novel eukaryotic vector. Gene. 108, 193-199 (1991).
- Chang, K., Elledge, S. J., Hannon, G. J. Lessons from Nature: microRNA-based shRNA libraries. Nat Methods. 3, 707-714 (2006).
- Schlaepfer, W. W., Myers, F. K. Relationship of myelin internode elongation and growth in the rat sural nerve. J Comp Neurol. 147, 255-266 (1973).
- Webster, H. D. The geometry of peripheral myelin sheaths during their formation and growth in rat sciatic nerves. J Cell Biol. 48, 348-367 (1971).
- Weis, J., Fine, S. M., David, C., Savarirayan, S., Sanes, J. R. Integration site-dependent expression of a transgene reveals specialized features of cells associated with neuromuscular junctions. J Cell Biol. 113, 1385-1397 (1991).
- Zhao, Y., et al. An expanded palette of genetically encoded Ca(2)(+) indicators. Science. 333 (2), 1888-1891 (2011).
- Suzuki, J., et al. Imaging intraorganellar Ca2+ at subcellular resolution using CEPIA. Nat Commun. 5, 4153 (2014).
- Tabata, H., Nakajima, K. Labeling embryonic mouse central nervous system cells by in utero electroporation. Dev Growth Differ. 50, 507-511 (2008).
- Taveggia, C., Feltri, M. L., Wrabetz, L. Signals to promote myelin formation and repair. Nat Rev Neurol. 6, 276-287 (2010).
- Saijilafu, E. M., Hur, F. Q., Zhou, Genetic dissection of axon regeneration via in vivo electroporation of adult mouse sensory neurons. Nat Commun. 2, 543 (2011).
- Tanaka, Y., Hirokawa, N. Mouse models of Charcot-Marie-Tooth disease. Trends Genet. 18, S39-S44 (2002).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены