
Method Article
Антимикробные пептиды, производится путем селективного давления включения неканонических аминокислот
* Эти авторы внесли равный вклад
В этой статье
Резюме
Протокол представляет Escherichia coli-на основе селективного давления включения низина антимикробного пептида lactococcal неканонических аминокислот (ncAAs). Его свойства могут быть изменены во время рекомбинантных выражение через замену с желаемой ncAAs в определенных питательных сред. Обусловленные этим изменения в биологическую сопоставляются анализов ингибирование роста и микроскопии флуоресцирования.
Аннотация
Природа имеет множество возможностей для создания новых функций белков, изменяя последовательность отдельных аминокислот строительных блоков. Однако все варианты основаны на 20 канонические аминокислоты (cAAs). Как способ привнести дополнительные физико-химическим свойствам полипептидов включение неканонических аминокислот (ncAAs) все шире используется в машиностроении белка. Из-за их сравнительно короткой длины модификация ribosomally синтезированных и post-translationally модифицированных пептиды, ncAAs особенно привлекательным. Новые функциональные возможности и химические ручки может быть порождена конкретные изменения отдельных остатков. Селективного давления включения (SPI) метод использует ауксотрофных хост штаммов, которые лишены аминокислотой в химически определенных питательных сред. Несколько структурно и химически аналогичные аминокислот аналогов затем может быть активировано путем соответствующего аминоацил тРНК-синтетазы и предоставляют специфичные для остатков cAA(s) → ncAA(s) замен в последовательности целевого объекта пептид или белок. Хотя, в контексте метода SPI, ncAAs также включены в принимающей протеома во время фазы экспрессии рекомбинантных генов, большинство клеток ресурсов присваиваются экспрессии гена целевого объекта. Это позволяет эффективное включение остатков конкретных ncAAs часто сопровождается с высоким количеством изменение целевого. Представленная работа описывает в естественных условиях включения шести пролина аналогов в низина антимикробного пептида, lantibiotic, естественным образом вырабатывается Lactococcus lactis. Антимикробные свойства низина могут быть изменены и расширены в ходе ферментации и выражение в ауксотрофных Escherichia coli штаммов в определенных питательных сред. Таким образом последствия остатков конкретных замена Суга с ncAAs может доставить изменения в активности антимикробной и специфичность. Антимикробная активность анализов и микроскопии флуоресцирования используются для тестирования новых вариантов низина для ингибирования роста штамма индикатор грамположительных Lactococcus lactis . Масс-спектроскопии используется для подтверждения включения ncAA в вариантах биоактивные низина.
Введение
Открытие антибиотиков в двадцатом веке и параллельной разработки новых антимикробных соединений против патогенных микроорганизмов включен целевых лечения бактериальных инфекций. Однако из-за появления лекарственно устойчивой патогенов, таких как метициллин резистентный Staphylococcusaureus (MRSA), к ванкомицину энтерококков (VRE), MDR (с множественной лекарственной устойчивостью) Salmonella typhimurium Фаговые типа 10 (DT10 ), и Klebsiella pneumoniae, это срочно необходимо для создания новых противомикробных препаратов1. Антимикробных пептидов (AMPs) являются универсальным, часто весьма конкретных соединений, которые являются перспективными кандидатами для разработки новых лекарств благодаря их физико-химических свойств, гибкость, размер, гидрофобность и режим действий2. Усилители являются малые пептиды, обычно состоящая из 7-100 аминокислот. Часто они имеют структуру Катионный богаты положительно заряженных остатков аргинина и лизин, которые взаимодействуют с целевой микробной клеточной мембраны, которое противоположно взимается3. Определенной подгруппе AMPs, ribosomally синтезированных и posttranslationally модифицированных пептидов (RiPPs)4. Они производятся многие организмы от царства грибов и домен бактерий. Один из наиболее известных и широко используемым RiPPs это низина, естественным образом вырабатывается молочнокислые бактерии Lactococcus lactis (lactis л). Активен в отношении грамположительных бактерий группы, Низина был использован как Биоконсервант в пищевой промышленности для более чем 50 лет из-за его антимикробные свойства и отсутствие развивались сопротивления в целевых микробных штаммов5. Исследования показали, что низина дестабилизирует и генерирует поры в бактериальных клеточных мембран, ведущих к антимикробной активностью в отношении как грамположительных и грамотрицательных патогенов6. Связываясь с липидов II, синтез клеточной стенки бактерий является тормозится7. Низина кодируется nisA как линейной прекурсоров пептид, который состоит из лидера и ядро пептид региона (рис. 1). После рибосомных синтеза prenisin сначала изменяется путем дегидратазы НИСМ. Здесь серина и треонина остатков в регионе prepeptide ядро обезвожены dehydroalanine (ДГВ) и dehydrobutyrine (Dhb)8. Впоследствии, обезвоженный остатков соединены с цистеина в форме lanthionine кольца (отсюда название «lantibiotic» для lanthionine кольцо содержащих антибиотики) добавлением фермента катализированное Майкл. Этот Посттрансляционная модификация (ПТМ) катализируемой циклазы помощь. В л lactisмодифицированных prenisin затем транспортируется из клетки транспортер NisT, а лидер пептид расщепляется, протеиназы НИСП выпустить зрелой и активных низина форма9. Ответственный лидер пептидаза НИСП имеет высокий субстратная специфичность, поскольку он только процессы изменения низина эффективно10.
В общем активные RiPPs результатом действий PTM ферментов (например, NisBC), которые резко увеличить пространство химических короткие пептиды, например, через ацетилирования, гликозилирования, метилирование или фосфорилирования. Этот уровень сложности далее может быть расширена путем прямого включения ncAAs. Хотя часто возможно, химический синтез усилителей является проблемой для крупномасштабного производства вследствие их структурной сложности. Например общая химического синтеза lantibiotic lactosin S в 71 реакция шагов была достигнута с окончательный выход 10% и что низина с сырой доходность только 0,003%11,12. Таким образом биологического производства предлагает жизнеспособную альтернативу, благодаря генерации правильного stereocenters и продукции с высокой концентрации.
До сегодня более чем 150 ncAAs, например, имея функциональные группы, содержащие фтор или азиды, были включены в рекомбинантных белков, и несколько примеров ncAA модифицированные усилителей были сообщалось13,14, 15,16. С введением ncAAs Роман физико-химических свойств могут быть созданы по сравнению с обычными мутагенеза. Разнообразие существующих пептидов может быть увеличено, возможно, приведет к Роман антибиотики.
Один метод для включения ncAAs рекомбинантные пептиды — селективного давления включения (SPI) на основе использования ауксотрофных бактериальных штаммов17. Эти штаммы не способны синтезировать соответствующий УГА аналог ncAA. Методология использует часто наблюдаемых расслабленной субстратная специфичность, особенностью многих природных аминоацил тРНК-синтетаз (aaRSs)18. Помимо их природных субстратов УГА эти ферменты способны часто признавать и активировать нужный ncAA и поручить его на их родственных tRNA(s). Это приводит к рибосомальной включение ncAA в целевой продукт гена остатков конкретным образом (то есть, cAA → ncAA замещение). Это, конечно возможно только когда желаемый ncAA структурно и химически похож на канонической аминокислоты и попустительстве физиологии клетки, перевод машин и пептид или белок последовательности целевого объекта. В частности экспериментальной установки клетки ауксотрофных хозяина культивируемых в определенной среде, поставляется с предельной концентрации родной аминокислоты быть замещенным. Клеточного роста или обмен cAA свободной среде приводит к внутриклеточное истощение УГА. На следующем шаге добавляется ncAA и целевое выражение гена индуцируется. Неизбежно ncAAs теперь также включены в многих других белков в клетки-хозяина во время этой фазы целевого выражения гена. Тем не менее, токсичность SPI установки хранится на низком уровне, поскольку штамм Escherichia coli (E. coli) преобразуется с плазмида, перевозящих целевого гена под контролем промотора сильного (обычно очень конкурентоспособным промоутер T7 / Система РНК полимеразы)19. Сразу же после индукции (обычно когда исчерпаны УГА), принимающей клетки перестают расти и их цитоплазматических ферментативные механизмов используются главным образом для экспрессии гена на основе плазмида целевой. Сайт Направленный мутагенез может использоваться для определения сайта установки специфичные для остатков ncAA в целевого гена20.
Как пептид модель для включения ncAAs была выбрана низина AMP пентациклических A. Это 34 аминокислот длиной и имеет только один пролина остатков в ядро пептид последовательности (рис. 1). Как и Субтилизин, ericin A и S и epidermin также, как и низина Z и низина Q сохраняемой proline, по-видимому, существенно важное значение для деятельности9,21. УГА proline играет особенно важную роль в peptidyl проли Амида вращения и вторичная структура стабилизации. Ее боковой цепи кольцо конформации (exo / заправка Эндо) отвечают за термодинамических стабилизации амидных связей. Целевые химические модификации (например, hydroxylations, fluorinations, methylations) нитей проли часто критически влиять на складной стабильности, леска жесткостью и функций многих биологических структур22. Таким образом вполне вероятно, ожидать, что Pro→ пролина аналоговый замены будет наделять кольцо B, второе кольцо низина, Роман и необычными свойствами.
Здесь, пролина ауксотрофных штамм E. coli был использован для производства рекомбинантного низина. Это требует выражение prepeptide гена nisA , а также модификация ферментов генов nisBC. Генетически закодированный пептид продукта несет N-неизлечимо его меткой лидером для очистки через аффинной хроматографии. Для определения активности, L. lactis выражения и секреции NisPT используется для активации рекомбинантных низина варианты от lysates клетки Escherichia coli или образцов очищенной пептида (рис. 1). Зрелые AMP освобождается после расщепления лидера организован НИСП. В этом методе диффузии агар AMP образец диффундирует в твердого роста среднего и может подавлять рост грамположительных микроорганизмов. После инкубации это можно наблюдать визуально, ореолы ингибирование роста. В дополнение к л lactis как индикатор изменение низина вариантов показал Антимикробная активность против Enterococcus faecalis, Bacillus cereus, золотистый стафилококк и Lactobacillus johnsonii 21,23.
Альтернативных и экспериментально другой метод для включения ncAAs в RiPPs-стоп кодон подавления (SCS)24. Для этого, ортогональные tRNA / аминоацил тРНК синтетазы (Орсе) пара требуется для соответствующего ncAA. В идеале все эти три компонента являются bioorthogonal, т.е. они не взаимодействуют с эндогенного tRNAs и aaRSs. NcAA конкретных Орсе может быть порождена модификации фермента активного узла и скрининга генетической библиотек мутант синтетаз25. Кроме того введение ncAA требует кодоном который переназначается и которая не кодируются для УГА. Обычно Янтарный стоп-кодон — используется24,26.
Недавно SPI была создана для включения α-chloroacetamide содержащих и клик химия совместимые ncAAs в NisA27. К примеру Nε- alloc-лизин была включена в captistruin пептид Лассо с сайта (СКС) и остатка (SPI) включение методов и впоследствии в пробирке рутений катализированное метатеза28 . По сравнению с SPI метод SCS является более сложным с ортогональными tRNA / Орсе пары должен быть совместно выразили. На сегодняшний день, o пар для включения пролина были развитые29, но в меру наших знаний, поступили не пример пролина аналоговый инкорпорации.
Следует отметить, что не все ncAAs могут быть включены с помощью методологии SPI. Во-первых поглощение ncAAs в цитоплазму регулируется целым рядом транспортных белков, которые внедряются в цитоплазматической мембраны, которая является внутренней мембраны для грамотрицательных бактерий, таких как кишечная палочка. Как правило E. coli , способен перевозить широкий спектр аналогов аминокислот в клетку с боковых цепей структурно и химически похожие на канонической аминокислоты. Во-вторых, многие химически реактивным или нестабильной ncAAs может выступать в качестве ингибитора направлении клеточного роста, как они являются токсичными для метаболизма и физиологии принимающей ячейки30. Таким образом поглощение и токсичность ncAA для принимающей страны производства должны быть проверены заранее. Чтобы избежать инактивации PTM механизма в качестве побочного эффекта, установку строго контролируемых выражение генов, ответственных может использоваться для включения природных аминокислот в модификация ферментов (например, nisBC) и ncAA в целевых генов ( Например, Ниса). Это может быть достигнуто с использованием двух разных промоутеров и индукции целевых экспрессии генов, как показано в специально SPI протоколы31. Как указывалось выше, метод SPI опирается на расслабленной субстратная специфичность Орсе, который позволяет для активации ncAA и родственных tRNA зарядки. Впоследствии tRNA доставляется на рибосомы следуют Амида Бонд формирования и складывающиеся пептида целевой (поли). В этом процессе корректуры и редактирования механизмы могут стать соответствующим32. По этим причинам, это большое значение для целевой ncAA которая структурно и химически похож на УГА. Другие важные вопросы являются достаточной стабильности (оба в средствах массовой информации роста и подвергается клеточный метаболизм) и растворимости ncAA. Кроме того оно должно быть коммерчески доступных или легко химически синтезированных.
Здесь мы описываем протокол для SPI, позволяя остатков конкретного включения ncAAs в рекомбинантной RiPPs. Особенно, различными пролина аналогов включены в антимикробного пептида низина A с помощью кишечной палочки как организм хозяина. Масс-спектрометрия используется для проверки аминокислоты замены и пептидных продуктов анализируются на биологическую, используя анализы ингибирование роста и микроскопии флуоресцирования, с использованием штаммов микроорганизмов индикатор.
Основное требование для успешной рекомбинантной низина выражение с определенными ncAAs требует подходящей пролина ауксотрофных штамм E. coli . Для этого auxotrophy, proA должен быть неблагополучных, например достигнут геномной нокаутом. Клетки полностью лишен биосинтеза внутриклеточных Pro (т.е. удаление proABC) без возможности возврата стабильной auxotrophs. Широко используется ген нокаут методы являются Фаговые трансдукция или одного гена нокаут согласно Даценко и Ваннер33. Кроме того proA нокаут штаммы могут быть получены общественного репозиториях, таких как Addgene, CGSC или коллекции Кэйо. Поскольку выражение рекомбинантных nisABC , показанный здесь основывается на использовании T7 промоутеров, штамм узла выражение должен нести индуцибельной гена для T7 РНК-полимеразы. Это может быть достигнуто путем введения λDE3 Профаг в принимающей генома, например, с помощью коммерческих комплект. Кроме того штаммы такие BL21(DE3) можно выполнить ауксотрофных, как описано выше.
протокол
1. клонирование векторы выражения и преобразования штамма ауксотрофных производства
Здесь гены для биосинтеза низина, а именно nisABC, были взяты из L. lactis и переведены в векторы выражения на основе T7 плазмиды. Полной последовательности ДНК nisABC можно найти в GenBank запись X6830734. Ген прекурсоров пептида (nisA) был сделан на ПЭТ 3a вектор, который наделяет ампициллин сопротивления. Гены дегидратазы (НИСМ) и циклазы (помощь) были размещены на вектор pRSFDuet-1, как сообщалось ранее35, который совещается канамицин сопротивления.
Примечание: Для nisA, кодонов последних четырех аминокислот (АСПР) лидер последовательности были мутировал кодировать VSLR36 оказать остатков пролина в ядро пептид уникальный и для обеспечения надлежащей обработки организован НИСП prepeptide. В N-terminus гекса-гистидина окружении компоновщика остатков был добавлен для целей очистки (см. Рисунок 1).

Рисунок 1 . Схематическое представление биосинтез и ПТМ Ниса в E. coli и лидер расщепления по L. lactis индикатор напряжения в последующей деятельности пробирного. На первом шаге, кодируемых неактивных линейных prenisin (в составе лидера и ядро региона пептид, содержащий уникальный пролина (розовый) в позиции 9) Ниса ribosomally синтезированы. Далее prenisin posttranslationally изменяется путем обезвоживания серина и треонина остатков в dehydroalanine (ДГВ) и dehydrobutyrine (Dhb) как катализируемой НИСМ. Циклазы центр образует тиоэфиры мосты через Майкл добавлением цистеина сульфгидрильных групп с ДГВ или Dhb. Неактивные модифицированных prenisin очищенного от кишечной палочки и проверяются на антимикробной активности. Здесь он перевозится в ячейку грамположительных л lactis индикатор напряжения. Лидером является расщепляется протеазы НИСП (как указано стрелкой) выпустить полностью активные низина. Она также может быть удален в пробирке путем обращения с трипсина (*). Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
{kind=link}
- Используйте стандартные теплового шока протокол37 или электропорации38 для преобразования proline ауксотрофных штамм E. coli (см. выше) с плазмид ПЭТ 3a Ниса(VSLR) и pRSFDuet-1 nisBC.
- Пипетка 25-100 мкл суспензии трансформированных клеток на плиты агара, содержащий ампициллина, канамицин и 1% (w/v) глюкозы. Использование разбрасыватель пластины или стеклянные бусины распространить решение равномерно на тарелках.
- Инкубировать пластин на ночь при 37 ° C.
- После обеда, используйте единую колонию для инокуляции 10 мл LB носитель, содержащий ампициллина, канамицин и 1% (w/v) глюкозы в 50 мл флакон.
- Встряхните культуры на ночь (12-16 ч) при 37 ° C и 200 об/мин.
- 250 мкл стерильные 80% глицерина и 550 мкл культуры, смесь хорошо в 2-мл пробирку и хранить как замороженные клетки запас на 80 ° C.
2. новый минимальный средний (НММ) подготовка
Примечание: Этот протокол использует NMM20 как химически определенных жидкий бактериальный рост среднего. Кроме того рекомендуется строго следовать порядок подготовки. В противном случае может произойти осадки. Амино кислоты форм, отличающихся от тех, которые перечислены в таблице материалы (например, гидрохлоридов) Проверьте растворимость. NMM19 содержит 19 аминокислот Кроме УГА заменить (здесь, пролин), аналог ncAA. Приведена таблица 1 заключительный ингредиент концентрации. В зависимости от бактериального штамма, используемые для производства биотин и тиамина может быть необязательным.
- Подготовка смеси аминокислот
- Растворите 0,5 g Phe, ГТО и Tyr в 100 мл ddH2O с добавлением нескольких капель концентрированной HCl до роспуска.
- Весят, 0,5 г каждого из оставшихся 16 аминокислот. Смешать с 22 мл 1м х2PO4 и 48 мл 1 M K2HPO4. Добавьте ddH2O ~ 800 мл. Перемешайте до тех пор, пока решение становится ясно.
- Добавить растворенные Phe, ГТО и Tyr и отрегулировать громкость решения 1 Л с ddH2O.
- Стерилизация смеси аминокислот вакуумной фильтрации с единицей фильтрации верхнего бутылку.
- Складе решения для NMM19
- Во-первых, подготовить 1 М складе решения следующих компонентов: (NH4)2,4, KH2PO4, K2HPO4, MgSO4 и 5 М раствора NaCl акции. Стерилизация автоклавированием.
- Подготовка 50 мл запасов D-глюкоза (1 М), CaCl2 (1 г/Л), FeCl2 (1 г/Л), тиамин (10 г/Л), биотин (10 г/Л) и микроэлементов (CuSO4, ZnCl2, НКД2, (NH4)2MoO4; 1 мг/Л ). Стерилизуйте каждый путем фильтрации с помощью шприца фильтра.
- NMM19 подготовка
- Смешайте все биржевые решения для получения конечной концентрации 7,5 мм (NH4)2т4, 1,7 мм NaCl, 22 мм х2PO4, 50 мм K2HPO4, 1 мм MgSO4 и 20 мм D-глюкоза, амино кислоты 50 мг/Л Mix, 1 мкг/Л CaCl2, 1 мкг/Л FeCl2, 10 мкг/Л тиамина, биотин 10 мг/Л и прослеживающие элементы 0,01 мкг/мл.
3. выражение рекомбинантных низина с включения Proline аналогов по SPI
В этом разделе, рекомбинантных выражение prepeptide (здесь: Ниса) и ПТМ генов (здесь: nisBC) производится. Во-первых клетки выращивают в присутствии всех Суга, так как LB сложные среды используется. Глюкоза добавляется для подавления экспрессии генов цели на уровне фон, который в противном случае может привести к производству одичал тип пептида (здесь: Низина) из-за негерметичности промоутеров. Только после назначения УГА (здесь: пролина) является ncAA добавляется в исчерпаны, и целевое выражение гена индуцируется в химически определенной среде. Инкубации жидких культур должно выполняться в подходящих колбы с аэрацией (например, 500 мл в колбу Эрленмейера 2 Л на 200 об/мин).
- С помощью кончика стерильной пипеткой, начать свежие ночи культуры из замороженных клеток акции или свежие колонии (см. шаг 1). Использование 25 мл LB носитель, содержащий ампициллина, канамицин и 1% (w/v) глюкозы и инкубировать на ночь (12-16 ч) при 37 ° C и 200 об/мин.
- Прививок 1 Л стерильных свежей среды с 10 мл на ночь культуры (1% v/v) и инкубировать при 37 ° C и 200 об/мин до600 OD = 0.5.
- Центрифуга при температуре 4 ° C на 15 мин на 4500 x g.
- Слить супернатант и Ресуспензируйте лепешка с 20 мл NMM19 (подготовленных на шаге 2.3) содержащих антибиотики и 1% (w/v) глюкозы. Центрифуга при температуре 4 ° C для 10 мин на 4500 x g.
- Ресуспензируйте Пелле клеток в 500 мл же среды и Инкубируйте на 30 ° C и 200 rpm за 1 час.
Примечание: На этом шаге, происходит истощение УГА (здесь, пролин). - Делят на равные части (один для каждого ncAA) культуры. Побудить каждой культуры с 1 мм изопропиловый β-D-1-тиогалактопиранозид (IPTG) и поставок пролин аналогов 1 мм (4S/R- fluoroproline, 4S/R- гидроксипролина или 4S/R- methanoproline).
Примечание: Как управления, одна культура может поставляться с 1 мм пролина, что приводит к одичал тип пептида. - Инкубируйте на ночь (12-16 ч) на 28 ° C и 200 об/мин.
- Центрифуга ячейки культур в 50 мл трубки на 4 ° C на 20 мин в 5000 x g. слить супернатант и хранить окатышей на-80 ° C до очистки.
4. изоляция и очищение его меткой низина аналогов
Пептиды очищаются под денатурации условий с Гуанидин гидрохлорид (GuHCl)39, сильный денатурат.
Предупреждение: GuHCl вред при проглатывании или вдыхании и вызывает серьезное раздражение глаз и кожи. Надевайте защитные очки и перчатки.
- Подготовка 250 мл привязки буфер (5 M GuHCl, 300 мм NaCl, 25 трис, рН 7,4), мыть буфера (300 мм NaCl, 25 мм трис, 25 мм имидазола, рН 7,4) и Элюирующий буфер (300 мм NaCl, 25 мм трис, 250 мм имидазола, рН 7,4). Для этих переноса твердых частиц во флаконе 250 мл и заполнить до 200 мл с ddH2O. смесь хорошо и скорректировать рН 7,4 с 1 M NaOH или HCl. Затем заполните до 250 мл с ddH2O. фильтр всех буферных растворов с помощью фильтрации верхнего бутылку.
- Лизис клеток
Здесь используется sonicator (мощностью 200 Вт Максимальная высокой частоты (ВЧ)); Обратите внимание, что параметры питания, необходимых для разрушения клеток может отличаться для других инструментов. Все действия выполняются на льду. В качестве альтернативы можно использовать химические клеток лизис, жидкий гомогенизатор или французская пресса.- Добавьте 12 мл буфера привязки каждого пластиковых пробирок (шаг 3,8) и Ресуспензируйте vortexing.
- Погрузите кончик зонда sonicator в суспензии клеток. Установите sonicator на 40% амплитуды с импульса 1 s / 5 s от 15 мин.
Примечание: Чистый кончик sonicator между образцов, чтобы избежать уноса. Протрите sonicator зонд с 70% этиловом спирте. - Центрифуга суспензию лизированных клеток на 4 ° C для 40 мин на 15000 x g для пеллет ячейки мусор. Передача supernatants к новой пробке реакции.
- Хромотография сродства
Для иммобилизованных ион металла сродство хроматографии (ИАЦ)40Перистальтический насос или системы ПСОК может использоваться с 1 мл картридж (здесь заполнены с Ni-НТА смолы). Для приготовления буферов смотрите шаг 4.1.
Примечание: IMAC очистки осуществима поскольку производства рекомбинантного пептид носит N-неизлечимо его меткой лидера, который удаляется в шаге 6, лидер пептидаза НИСП, выпуская Зрелые низина. Выполнение очистки при комнатной температуре или при 4 ° C. Используйте скорость потока по 1 мл в минуту если применимо для IMAC картриджа.- Во-первых мыть патрон с 5 томов столбца (cv) ddH2O удалить буфер хранения.
- Сбалансировать с 10 cv привязки буфера.
- Процесс клеток lysate (шаг 4.2) с помощью шприца фильтр, чтобы удалить частицы, а затем применить к картриджа.
- Стирка с 15 cv мыть буфера для того, чтобы удалить неспецифических и несвязанные материал.
- Элюировать с 10 cv Элюирующий буфер и собирать 1 мл фракций в 1,5 мл пробирок. Хранение дробей в 4 ° C на короткий срок (до 3 дней) или при-20 ° C для более длительные сроки.
- Для хранения, мыть патрон с 10 cv ddH2O следуют 5 резюме 20% этанола.
5. LC-ЭСИ TOF масс-спектрометрических анализ низина аналогов
Примечание: Таблица Просмотреть материалы за пример инструментирования для жидкостной хроматографии в сочетании с электроспрей ионизации время полета масс-спектрометрия (LC-ЭСИ-TOF-MS).
- Выполните ВЭЖХ разделения раствора 15-20 мкл пептида (подготовленных на шаге 4.3) на столбец C5 с мобильных фазы воды (A) и ацетонитриле, (B) оба дополнена 0,1% муравьиной кислоты и градиент от 5-80% B более 20 мин. Для масс-спектрометрия (МС) используйте элюции через 5 мин.
Примечание: В зависимости от пептида содержание и близость к столбцу ВЭЖХ, образец томов и разделения может понадобиться оптимизации. - Используйте соответствующее программное обеспечение для deconvolute измеренной массы спектры и рассчитать заряда различных пептид государства41. Сравнение массы видов наблюдаемых пептид расчетная масса одичал типа, изменено путем замены ncAA → УГА. Принимать во внимание, что линейные prepeptide posttranslationally изменение восемь dehydrations (-8 H2O) и пять cyclizations (см. Рисунок 1).
Примечание: Использование буферов, содержащих натрия, MS анализ положительных режиме может показать что аддукты натрия. Они становятся видимыми как дополнительные пики с большей массой deconvoluted (для каждого натрия аддукт, наблюдаемых deconvoluted масса 22.99 да выше). Чтобы удалить эти аддукты, ВЭЖХ очистки42 или обширные диализа43 может быть выполнена.
6. антимикробной активности тест
- Подготовка GM17-агар пластин в стерильных условиях
- Подготовьте на ночь культуры индикатор напряжения NZ9000 L. lactis перевозящих плазмида pNG nisPT44 при 30 ° C в М17 бульон45 с 1% (w/v) глюкозы (= GM17) и 5 мкг/мл хлорамфеникола.
- Мера ОД600, прививать свежие средне-ОД600 = 0.1 и инкубировать до600 OD = 0,4-0,6. Затем поместите колбу на льду.
Примечание: Каждый ОД600 измерения будет потреблять тома культуры. Имейте в виду, что для каждого пробирного агар пластины, 1 мл бактериальной культуры потребуются. При необходимости наращивать объем жидкости культуры соответственно. - Для 1,5% агар весят, 4.5 g агар в стеклянной бутылке СМИ. Добавьте 300 мл ddH2O, смеси и автоклав.
- Готовить 2 x M17 бульон (двукратная концентрированного) в 300 мл ddH2O и автоклав.
- Смешайте 25 мл 2 x M17 бульон, содержащий 10 мкг/мл хлорамфеникола и 2% раствор глюкозы с 1 мл preculture lactis л (4% v/v).
- Добавьте 25 мл 1,5% расплавленный агар (свеже газобетона или подогревом в микроволновую печь).
Примечание: Перед этим, пусть бутылка прохладный на ощупь (около 50 ° C). Это необходимо, поскольку л lactis мезофильных организма, чувствительные к высоким температурам. - Залейте раствор в большой Петри. Сухие пластины для 10-15 мин.
- Стерилизация концы стеклянной пипетки Пастера пламени. Ждать для того охладить вниз, а затем использовать широкий конец для создания отверстия в затвердевшей GM17-агар.
- Подготовка образца
- 1 мл E. coli выражение культур (создана на шаге 3.7) маркировкой 1,5 мл трубки и центрифуги для 3 мин на 7000 x g. аспирата оставшиеся среднего и Ресуспензируйте Пелле клеток в 500 мкл Na-P (50 мм натрия фосфат буфера рН 7,4 из натрия дигидрогенфосфат 0,5 М и 0,5 М динатрия гидрогенфосфат).
- Sonicate образцы на льду (Сравните шаг 4.2.2). Погрузите кончик зонда sonicator в суспензии клеток. Установите sonicator на 30% амплитуды с импульса 1 s на и 5 s покинуть на 3 мин.
- Центрифуга ячейки lysate 10 мин на 13 000 x g для пеллет ячейки мусор. Передача супернатант к новой пробке реакции на льду.
- Разбавить и нормализовать supernatants клетки экстракт 1 мл ОД600 = 0,6, по отношению к плотности заготовленных клеток, с Na-P.
- Активности тест
- 40 мкл каждого нормализованных образца, в отверстие индикатор агар пластины (рис. 3). Используйте хлорамфеникола на 400 мкг/мл как антибактериальные управления соединения. Использование Элюирующий буфер как отрицательный контроль. Подождите, пока все образцы распространяются в агар. Инкубировать пластину на ночь на 30 ° C.
- Сфотографировать плиты агара, с помощью планшетного сканера или цифровой камеры. Рост ингибирование гало размеры могут быть измеренной вручную или с помощью ImageJ46.
7. Люминесцентная микроскопия
Для того, чтобы наблюдать эффект усилителей на бактериальных клеток, может использоваться свет и флуоресцентной микроскопии. Обратите внимание, что режим низина действий опирается на дестабилизацию и формирование поры в бактериальной мембраны6. Здесь Нил красный используется для пятно бактериальной клеточной мембраны, которая становится лизис клеток разбросаны и агрегированные по.
Примечание: Таблица см материалы например инструментария. Количество дополнительного усилителя решения может быть скорректирована в зависимости от концентрации пептида и биологическую.
- Подготовка клетки
- Подготовка 10 мм Нила красный раствор в диметилсульфоксида (ДМСО).
- Расти ОД600 л lactis индикатор напряжения = 1.0 как шаг 6.1.1-6.1.2.
- Центрифуга 1 мл культуры за 3 мин при 4 ° C и 5000 x g.
- Отменить супернатанта, Ресуспензируйте в 1 мл физиологического раствора фосфатного буфера (PBS)47.
- Центрифуги и Ресуспензируйте снова.
- Добавить 1 мкл Нила красный Стоковый раствор, осторожно перемешать.
- Приобретение микроскопических изображений
- 30 мкл подготовка клетки на обложке слайд во время захватывающей на 520 Нм.
- Установите приобретение время 0.2 s, кинетическая серии 0,1 Гц, длина 200 серии изображений.
- Добавить 0.3 - 1.5 мкл lysate клетки или IMAC образца (от шаг 6.2.4 или 4.3.5, соответственно). Для образцов IMAC Элюирующий буфер может использоваться как отрицательный контроль.
- Мониторинг и запись флуоресценции выбросов на λ ≥ 560 Нм.
- Анализ данных
- Микроскопия последовательности изображений хранятся как файлы фильмов (.avi). Отдельные изображения анализируются с ImageJ46.
Результаты
Этот протокол предназначен для включения производства низина ncAA модифицированные варианты включения специфичные для остатков пролина аналогов методом SPI. Для рекомбинантных производства полностью измененные одичал тип низина39были сообщалось ранее, осуществимость и урожайности 24 мг/л. С помощью метода SPI, целевой пептида/белка урожай часто хорошо и может достигать количествах недалеко от дикого типа производства48. Как первые эксперименты рекомбинантных одичал тип Рипп производства должны быть проверены в узле выбранной ауксотрофных. Здесь, пролина ауксотрофных E. coli MG1655 Δпроба:: frt ΔproC:: frt (DE3) был использован в качестве принимающей штамма. Для включения ncAAs, культивирования и индукции как сроков, так и среднего состава и температуры могут быть оптимизированы к максимальной пептид доходность.
Антимикробная активность пробирного
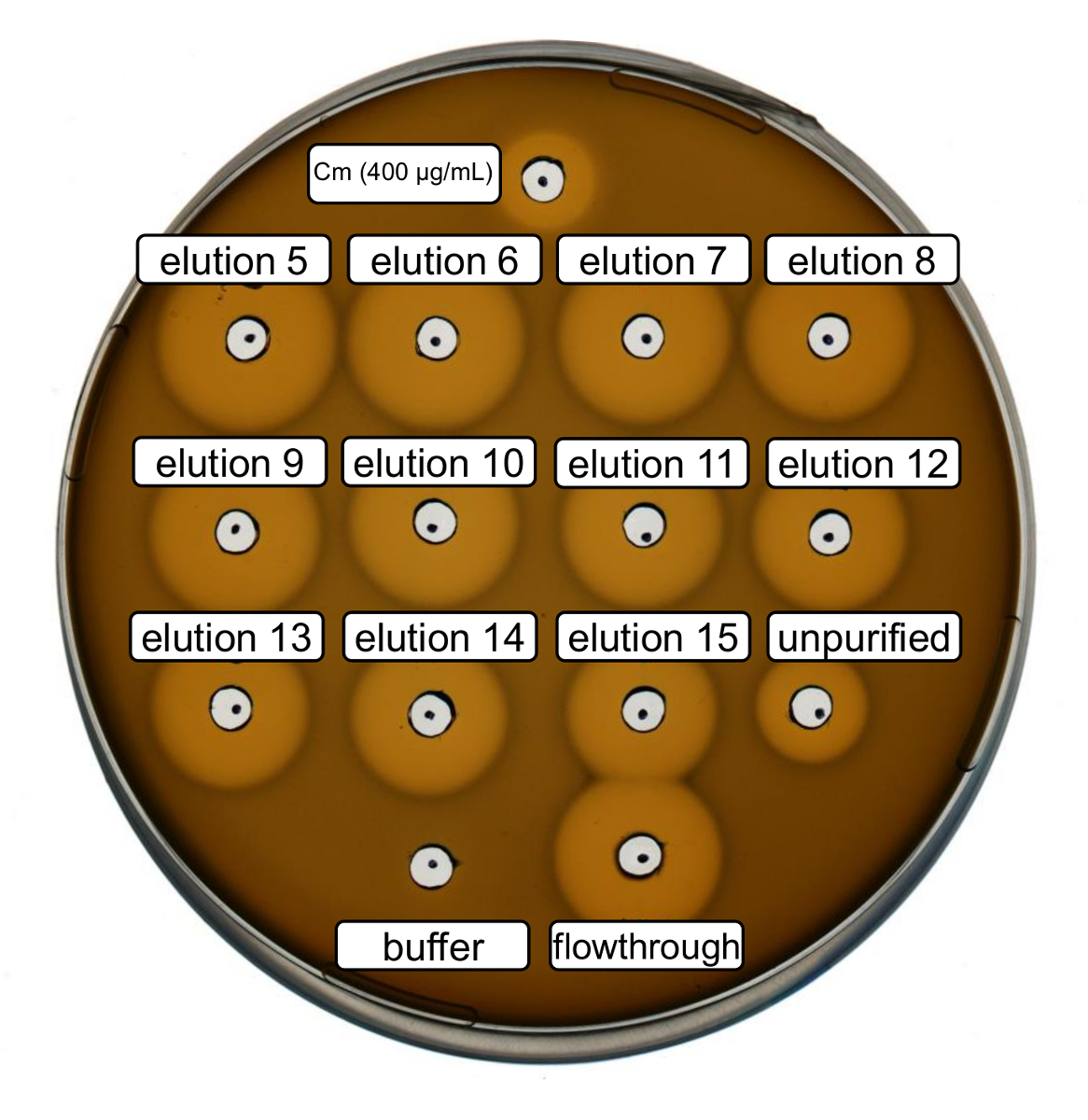
Рекомбинатные низина одичал тип производства и очистки были выполнены после выше протокол. В этом случае "ПроЛайн" был использован вместо его производные ncAA на шаге 3.6. Антимикробной активности был использован для проверки Рипп производства и сравнить антимикробной активности до и после очистки. Для анализа активности элюции дробями и потока через IMAC были непосредственно использованы и испытаны против грамположительных л lactis индикатор напряжения (рис. 2). Как этот штамм выражает НИСП, Низина варианты, содержащиеся в E. coli lysates клетки или образцов очищенной пептид, соответственно, стали активироваться протеолитического расщепления лидер пептида. Очевидно поток через показал рост ингибиторная активность. Это может объясняться Биоактивный материал не привязки к столбцу IMAC. Всех протестированных элюции фракций показал повышенная активность по сравнению с неочищенную образца, указывающее концентрация пептида его тегами, IMAC. Обратите внимание, что Элюирующий буфер (как отрицательный контроль) не влияют рост L. lactis в этот assay.

Рисунок 2 . Антимикробная активность теста после очистки IMAC recombinantly низина одичал типа. По сравнению с неочищенную lysate клетки (разбавленный для нормализации600 OD) против штамма индикатор л lactis были протестированы элюции фракций 5 до 15 и через поток очистки IMAC. Размер гало ингибирование роста показывает антимикробной активности. Хлорамфеникол при концентрации 400 мкг/мл был использован как положительный контроль и IMAC Элюирующий буфер как отрицательный контроль. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
{kind=link}
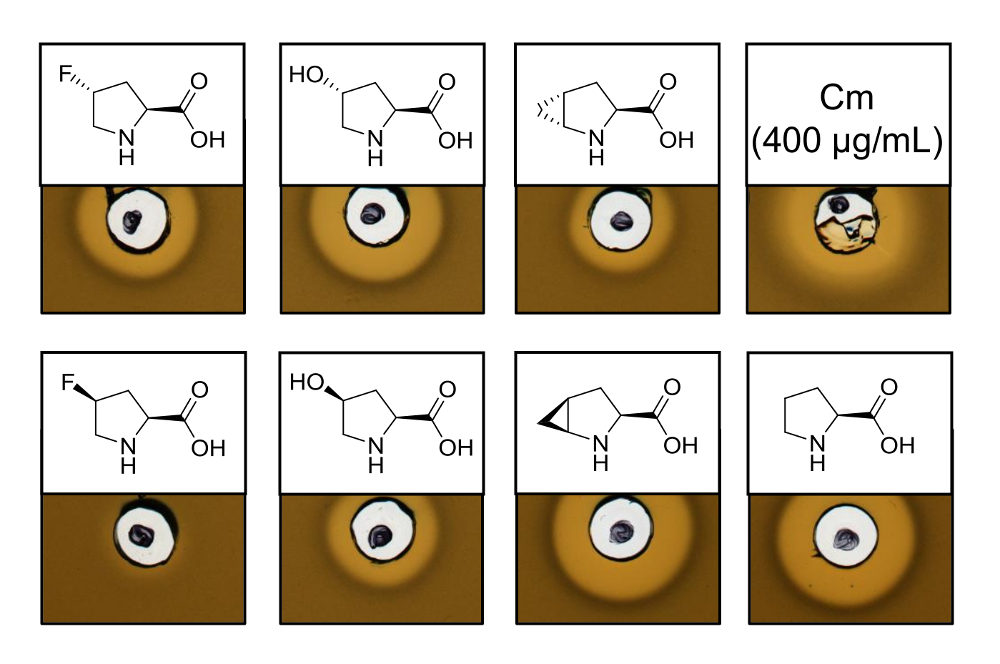
Чтобы доказать антимикробной активности recombinantly производства низина, варианты, содержащий шесть различных пролина аналогов, assay деятельность была выполнена для тестирования ингибирование л lactis индикатор напряжения. На рисунке 3 показано ингибирование роста для пяти из шести образцах, вырабатываемый пролина аналогов. Наилучшие результаты (как рассужено от размеров гало) наблюдались для включения экспериментов аналоговые (4R) - fluoroproline, (4R) - гидроксипролина и 4S-methanoproline. Сравнив размер гало ингибирование роста одичал тип низина изготовлены и испытаны параллельно, все три варианта низина показали аналогичные ингибирование. Однако, не может быть только размер гало используется для ослов конкретной деятельности, поскольку концентрация AMP не был определен. Таким образом анализов служат только для того, чтобы качественно проверить сохранились ли потерял антимикробной активности результате низина вариантов. Для определения удельной активности, концентрация низина вариантов должен быть количественных обязательств (см. обсуждение).

Рисунок 3 . Антимикробная активность пробирного lysates клетки содержащие варианты низина производится через SPI с аналогами пролина. Сравнение вариантов низина с рекомбинантным образцов дикого типа. Все образцы были ОД600-нормализуются после лизиса клеток по отношению к плотности культуры заготовленных клеток. Ореолы указывают отпорности в форме торможение роста индикатор напряжения. Первый ряд слева направо: (4R)-fluoroproline, (4R) - гидроксипролина, (4R) - methanoproline и хлорамфеникола (400 мкг/мл; антимикробной положительный контроль). Второй ряд: (4S)-fluoroproline, (4S) - гидроксипролина, (4S) - methanoproline и пролина (одичал тип управления). Обратите внимание, химической номенклатуры; например, (4R) - fluoroproline также называется транс-4 - fluoroproline. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
{kind=link}
LC-ЭСИ-TOF масс-спектрометрия
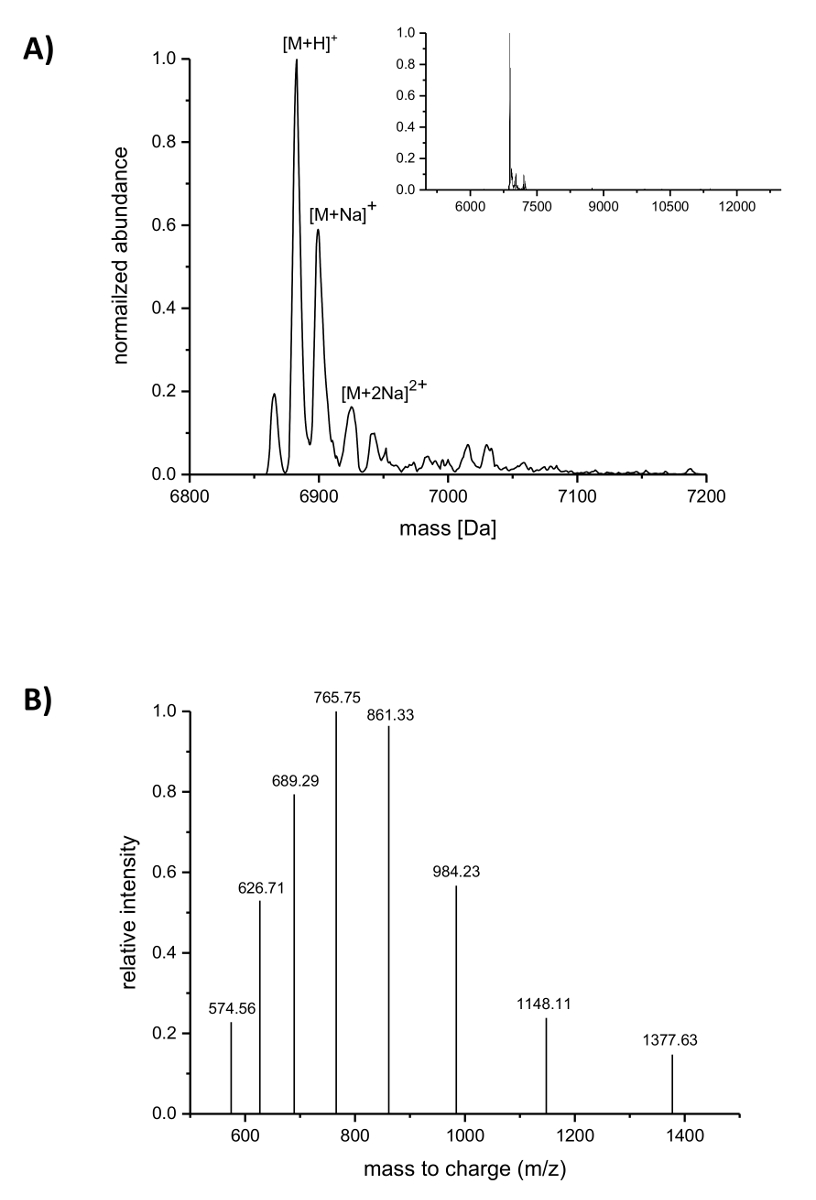
После очистки IMAC включение ncAAs в низина был проанализирован LC-ЭСИ-TOF масс-спектрометрии. Рисунок 4 показывает deconvoluted массы спектры низина variant, содержащее (4R)-fluoroproline. Этот вариант был IMAC очищенный как описано выше и впоследствии проанализированы LC-ЭСИ-TOF масс-спектрометрии, поэтому он по-прежнему осуществляется руководителем. Основной пик в Рисунок 4A соответствует изменение низина, содержащие (4R)-fluoroproline с deconvoluted массой 6883.18 Да (расчетная масса 6882.05 Да, расчетная масса соответствующих одичал тип пептид с пролина в положение 9 — 6864.06 да). Две вершины с нижней изобилия и выше массы соответствует натрия аддукты как указано. Рисунок 4 B показывает по-разному заряженных видов основных составных как найденные алгоритмом деконволюции. Например, пик на 1148.11 m/z соответствует шесть раз заряженных видов ([M + 6H]6 +).

Рисунок 4 . LC-ЭСИ-TOF масс-спектрометрия IMAC-очищенная рекомбинантных низина вариантов, содержащих (4R)-fluoroproline. (A) Deconvoluted Хроматограмма масс-спектрометрия (масштаб в вставкой) для варианта низина (по-прежнему носить лидер) с (4R)-fluoroproline (ожидается масс (Da): [M + H]+ = 6882.05, [M + Na]+ = 6904.03, [M + 2На]2 + = 6926.02). (B) сложные спектры для видов [M + H]+. Ожидается масс (Da): [M + 5H]5 + = 1377.41, [M + 6 H]6 + = 1148.01, [M + 7 H]7 + = 984.15, [M + 8 H]8 + = 861.26, [M + 9 H]9 + = 765.67, [M + 10 H]10 + = 689.21, [M + 11 H]11 + = 626.64, [M + 12 H] 12 + = 574.50. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
{kind=link}
Микроскопии флуоресцирования
Антимикробная активность рекомбинантного низина и его ncAA содержащих варианты могут отображаться также путем прямого наблюдения л lactis индикатор напряжения через микроскопии флуоресцирования. Нил красный, высокой гидрофобностью Люминесцентную краску, был использован для пятно бактериальной мембраны клетки. Рисунок 5 показывает качественного изменения агрегации состояния культуры клеток и морфология одну ячейку. Клетки были окрашенных с красным Нила и хранение на слайд крышка микроскопии. Верхняя строка показывает клетки непосредственно в начале когда буфер, рекомбинантных одичал тип низина, или низина содержащие (4R)-fluoroproline, или (4R)-гидроксипролина были добавлены. Нижней панели показывает соответствующие изображения после 20 минут инкубации.

Рисунок 5 . Микроскопии флуоресцирования рекомбинантных низина воздействия на грам-положительных клеток. Микроскопических изображений 30 мкл л lactis индикатор напряжения (OD600 = 1), отмеченные Нила, красный были приняты до (Верхняя панель) и после (Нижняя панель) 20 минут инкубации с 1 мкл буфера (A), 0,3 мкл рекомбинантных одичал тип низина (B) и 0,6 мкл низина вариантов, содержащих (4R)-fluoroproline (C) и (4R)-гидроксипролина (D). Синие круги Марк регионов с агрегированные или деформированных клетки, синие стрелки точки в регионы, где можно наблюдать диффузии флуоресцентные мембраны фрагментов. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
{kind=link}
Рисунок 5 A показывает, что общий внешний вид ячеек не изменился в течение 20 минут наблюдения. Только количество ячеек, которые сданы на хранение во время Роуз и, следовательно, большее количество клеток является видимым в пределах 80 мкм x 80 мкм наблюдается региона. Рисунок 5 B показывает, что грам-положительных клеток появился агрегированных и размыто (отмечены синими кругами) после 20 минут воздействия одичал тип низина, даже при низкой суммы (здесь, 0,3 мкл IMAC элюции) были добавлены. Кроме того легкий материал рассеянный из ячейки в буфер, указав, что фрагменты мембраны, отмеченные красным Нила были мобилизованы за период (отмечен синие стрелки). Эти выводы свидетельствуют о лизис клеток, как это было показано происходят после лечения с низина6,49. Аналогичные эффекты наблюдались после инкубации с низина вариант содержащие (4R) - fluoroproline (рис. 5C) и низина, содержащие (4R) - гидроксипролина (рис. 5D) оба показаны большие количество искаженные и агрегированных клеток после 20 минут, в отличие элемента управления образца (рис. 5A).
| Компонент | Концентрация |
| (NH4) 2 ТАК4 | 7.5 мм |
| NaCl | 8.5 мм |
| KH2PO4 | 22 мм |
| K2HPO4 | 50 мм |
| MgSO4 | 1 мм |
| D-глюкоза | 20 мм |
| Все канонические аминокислоты (за исключением одного, для замены) | 50 мг/Л |
| CA2 + | 1 мкг/мл |
| FE2 + | 1 мкг/мл |
| Прослеживающие элементы (Cu2 +, Zn2 +, Mn2 +, MoOH2 +) | 0,01 мкг/мл |
| Тиамин | 10 мкг/мл |
| Биотин | 10 мкг/мл |
Таблица 1. Состав NMM19 химически определяется средний рост бактерий после подготовки согласно шаг 2.
Обсуждение
С помощью SPI для вставки пролина аналогов, целевых низина структуры может быть существенно изменен, создавая Роман пептид варианты с уникальную последовательность комбинации и химические свойства. Таким образом можно обойти основные предел классической генной технологии, который ограничивается боковой цепи химия из 20 угас. В естественных условиях химические диверсификации низина, как свидетельствуют приведенные выше примеры демонстрирует общий подход для дополнения природных PTMs и резко повысить химического пространства RiPPs. Мы считаем, что расширение репертуара природных аминокислот имеет большие надежды, особенно для усилителей. В белках огромный спектр функций может быть реализована посредством определенного механизма 20 угас в трехмерных структур. С только 7-100 aa в длину3способы достижения таких структурных особенностей только через Суга будет ограничено для усилителей. Это, таким образом, не удивительно, что природные усилители в виде RiPPs обычно широко post-translationally изменение4. Таким же образом ncAAs как альтернативных строительных блоков держать большие перспективы для улучшения их фармакодинамические и фармакокинетические параметры (см. Baumann et al. 201735 и ссылки на них).
SPI методология, используемая в этой работе выгоды от относительно простой экспериментальной установки, простой исполнения и высокая воспроизводимость. Благодаря глобальной замены возможна также мультисайтовой ncAA включение в цели пептиды. С другой стороны этот метод не может быть достаточным для замены аминокислот, часто встречающихся в целевой продукт гена. В принципе нежелательные позиции может быть изменен сайт направленного мутагенеза, но эти дополнительные изменения могут также повлиять на несколько свойств AMP, включая структуру и биологическую. После того, как ауксотрофных штамм для производства доступен, несколько ncAAs может испытываться без обширные изменения на генетическом уровне. Кроме того, метод не ограничивается ауксотрофных E. coli штаммов, но также могут быть выполнены с использованием природных производства узла. К примеру, Чжоу et al. показал, что SPI для производства Роман RiPPs также работает естественно ГТО auxotroph L. lactis: с помощью определенных средств массовой информации, были включены три триптофан аналогов на четырех позициях в низина50.
Поскольку метод SPI приводит к глобальной замены выбранного УГА, ncAA, это правило применимо к широкий спектр целевых пептидов и белков. Диапазон ауксотрофных E. coli доступен штаммов (см. шаг 1), позволяя несколько угас каждый быть проверен ncAAs для замены. Встретил аналогов с помощью мета-дефицит штаммы (например, B834(DE3)) наиболее часто используются. Azidohomoalanine (Aha) и homopropargylglycine (Hpg), коммерчески доступных ncAAs, которые могут быть эффективно включены примеры isostructural встретил аналогов. Как ввести bioorthogonal ручки, которые позволяют прикрепления молекул, перевозящих совместимый алкины или азид функциональность, соответственно. К примеру, флуоресцентных красителей или постановление полиэтиленгликоля (PEG) могут быть прикреплены к меди (I)-катализированное азид алкины циклоприсоединения (CuAAC)51.
Хотя оба рекомбинантные методы включения ncAA (SPI и СКС) обычно достижения достаточного количества целевых, урожайность часто уменьшаются по сравнению с производством одичал тип соответствующего пептидов и белков. Как часто чистоты коррелируют с эффективностью производства, дополнительной очистки могут потребоваться действия, особенно для низкого обильные видов. В данном конкретном случае производство рекомбинантных низина последовательность его меткой лидера значительно упрощает Рипп очистки путем селективного обогащения от lysates клетки. Очистки, показано в этом протоколе улучшает чистоту и концентрация низина, но часто не дают достаточно для определения урожайности и удельной активности проб AMP. Помимо IMAC, часто используемые методы очистки AMP включают ВЭЖХ, ионообменной хроматографии (IEC) и осадков (например. с помощью ацетона или трихлоруксусной кислоты (TCA)) или их - комбинации, что приводит к схеме многоступенчатого очищения52 . Следует отметить, что их часто polycationic природа может облегчить IEC очистки. Паром для лиофильной сушки часто используется для хранения очищенной AMPs.
В идеале ncAAs для включения в AMPs должен быть коммерчески доступных по доступным ценам или легко подготовленных простых и воспроизводимые химического синтеза протоколов. Не менее важной предпосылкой для метода SPI является что ncAA признано, активируется и взимается на родственных tRNA эндогенного или совместно выразили Орсе. Это может быть протестированных в vitro аминокислоты активации или тРНК aminoacylation пробирного53. В качестве альтернативы легко SPI тест выражения модели белков, таких как Зеленый флуоресцентный белок (ГПУП) провела в присутствии и в отсутствие ncAA добавок могут быть выполнены. Кроме того растворимость в рост средних и клеток проницаемость, а также химическая стабильность являются важными факторами.
В этом примере антимикробной активности был показан с помощью штамм грамотрицательных индикатор. Как он выражает лидер пептидаза НИСП, последний шаг низина созревания катализировано. Удаление последовательности лидер (его тегами для целей очистки) также может быть выполненной в пробирке путем обращения с очищенной НИСП50 или трипсина54. Выходит за рамки данной работы патогенных организмов и множественной лекарственной устойчивостью штаммов может затем быть проверена для ингибирования бактериостатическое и бактерицидное ампер, с использованием аналогичной методологии. Клинически значимых видов включают в себя MRSA, МЛУ туберкулезом микобактерии, VRE, Acinetobacter baumannii, пневмококк, синегнойной палочки и Klebsiella pneumoniae. Помимо распространения плита агара пробирного представлены здесь, роста ингибирования также могут быть выполнены с использованием соответствующих жидких сред прививанным с бактериальными видами и дополнены AMP. С помощью методов разбавления отвара, минимальный тормозной концентрации (MIC) может быть определена с помощью Чистая пептидов55. Assay активности, представленные здесь может также использоваться для оценки биологическую и потенции AMP-содержащих решений относительно ведения соединений, например коммерчески доступных низина.
Рекомбинатные производство RiPPs часто является осуществимым39, который обычно включает в себя совместное экспрессии генов ПТМ. Как только штамм производство переносится в химически определяется или синтетических минимальный носитель, содержащий подходящие ncAA, замена остатков конкретных соответствующих Гага занимает место. Таким образом другие RiPPs могут быть изготовлены по той же методологии, условии, что их производство рекомбинантных осуществимым и условия могут быть найдены где ncAA включение и ПТМ дают достаточное количество целевого продукта. Обратите внимание, что помимо принимающей ячейки протеом, пептид PTM машины также может стать изменение ncAA во время SPI. Следовательно сроки целевого выражения индукции и длина следующих инкубационный период может требуют оптимизации. Так как ncAA включение в ферменты PTM может повлиять на их стабильность и деятельность, производство обработанных Рипп может в принципе быть затронуты. Достаточную эффективность фермента PTM обозначается формирования обработанные prepeptide, как определяется MS и отпорности анализов. Как выше, различные индуцибельной промоутеров могут быть использованы для того чтобы произвести PTM генов сначала (в отсутствии ncAA) следуют индукции гена пептид прекурсоров в присутствии ncAA. В целом производство cAA содержащих целевой пептид должны быть подавлены перед добавлением ncAA, именно поэтому жесткие репрессии гена прекурсоров не требуется. В рамках этого протокола это достигается путем катаболических репрессии путем добавления глюкозы до среднего роста. Тем более PTM ферментов, необходимых для prepeptide обработки возникают обычно от генетически далеких хоста, температуры и кодон использование выражений соответствующих генов может потребовать оптимизации если рекомбинантных производства до сих пор не создана. В принципе присутствие ncAAs в prepeptide может мешать распознавания и обработки PTM ферментов, в случае низина NisBC и НИСП. Для включения в ncAA в AMPs таким образом рекомендуется выполнять эксперименты мелких выражение сначала для выявления подходящего выражения условий и надежности деятельности пробирного AMP.
Раскрытие информации
Авторы заявляют, что они не имеют никаких финансовых интересов.
Благодарности
J.H.N., т.б. и м.б. признать финансирование по программе ЕС FW7 (SYNPEPTIDE). F.-и.с. и т.ф. признают финансирование от федерального министерства образования и науки (BMBF программа «HSP 2020», ту-WIMIplus проекта SynTUBio) и Немецкий исследовательский фонд (кластер передового опыта «Unifying концепции катализ»).
Материалы
| Name | Company | Catalog Number | Comments |
| sodium chloride | Carl Roth, Germany | P029 | |
| guanidine hydrochloride | Carl Roth, Germany | 0035.2 | |
| dipotassium hydrogen phosphate | Carl Roth, Germany | P749.3 | |
| potassium dihydrogen phosphate | Carl Roth, Germany | 3904.3 | |
| sodium dihydrogen phosphate monohydrate | Carl Roth, Germany | K300.2 | |
| disodium hydrogen phosphate | Carl Roth, Germany | P030.2 | |
| L-alanine | Carl Roth, Germany | 3076.2 | |
| L-arginine | Carl Roth, Germany | 3144.3 | |
| L-asparagine monohydrate | Carl Roth, Germany | HN23.1 | |
| L-aspartic acid | Carl Roth, Germany | T202.1 | |
| L-cysteine | Carl Roth, Germany | 3467.3 | |
| L-glutamine | Carl Roth, Germany | 3772.1 | |
| L-glutamic acid | Carl Roth, Germany | 3774.1 | |
| L-glycine | Carl Roth, Germany | 3187.3 | |
| L-histidine | Carl Roth, Germany | 3852.3 | |
| L-isoleucine | Carl Roth, Germany | 3922.3 | |
| L-leucine | Carl Roth, Germany | 3984.3 | |
| L-lysine monohydrate | Carl Roth, Germany | 4207.2 | |
| L-methionine | Carl Roth, Germany | 9359.4 | |
| L-phenylalanine | Carl Roth, Germany | 4491.2 | |
| L-proline | Carl Roth, Germany | T205.3 | |
| L-serine | Carl Roth, Germany | 4682.4 | |
| L-threonine | Carl Roth, Germany | T206.2 | |
| L-tryptophan | Carl Roth, Germany | 4858.2 | |
| L-tyrosine | Carl Roth, Germany | T207.2 | |
| L-valine | Carl Roth, Germany | 4879.4 | |
| ammonium sulfate | Carl Roth, Germany | 3746.3 | |
| magnesium sulfate | Carl Roth, Germany | 0261.2 | |
| D-glucose | Carl Roth, Germany | 6780 | prepare a 20% (w/v) solution for addition into molten agar |
| calcium chloride | Carl Roth, Germany | PN93.2 | |
| iron(II) chloride | Sigma-Aldrich, Germany | 372870 | |
| thiamine hydrochloride | Sigma-Aldrich, Germany | T1270 | |
| biotin | Carl Roth, Germany | 3822.2 | |
| copper(II) sulfate | Merck, Germany | 102792 | |
| manganese(II) chloride | Carl Roth, Germany | KK36.2 | |
| zinc chloride | Merck, Germany | 108816 | |
| immonium orthomolybdate | Sigma-Aldrich, Germany | 277908 | |
| glycerol | Carl Roth, Germany | 7533.3 | |
| isopropyl β-D-1-thiogalactopyranoside | Sigma-Aldrich, Germany | I6758 | |
| ampicillin sodium salt | Carl Roth, Germany | K029.5 | working concentration 100 µg/mL for E. coli, prepare 100 mg/mL stock in ddH2O |
| kanamycin sulfate | Carl Roth, Germany | T832.2 | working concentration 50 µg/mL for E. coli, prepare 50 mg/mL stock in ddH2O |
| chloramphenicol | Carl Roth, Germany | 3886.1 | working concentration 5 µg/mL for L. lactis, prepare 37 mg/mL stock in ethanol |
| (4S)-fluoroproline | Bachem, Switzerland | F-3970 | |
| (4R)-fluoroproline | Bachem, Switzerland | F-3975 | |
| (4S)-hydroxyproline | Bachem, Switzerland | F-1395 | |
| (4R)-hydroxyproline | Bachem, Switzerland | F-2980 | |
| (4S)-methanoproline | chemically synthesized | ||
| (4R)-methanoproline | chemically synthesized | ||
| hydrochloric acid (HCl) | Carl Roth, Germany | P074.4 | |
| ethanol | VWR, Germany | 20825.324 | |
| M17-broth | Sigmal-Aldrich, Germany | 56156 | commercial product, see Terzaghi BE & Sandine WE, Appl Microbiol., 1975, 29(6):807-13 for contents and preparation |
| agar-agar | Carl Roth, Germany | 5210.5 | |
| Nisin from Lactococcus lactis | Sigma-Aldrich, Germany | N5764 | commercial product, can be used as reference for bioactivity |
| dimethyl sulfoxide (DMSO) | Carl Roth, Germany | A994.1 | |
| imidazole | Carl Roth, Germany | 3899.3 | |
| 1.5 mL autosampler vial for LC-MS | Sigma-Aldrich, Germany | Supelco 854165 | |
| acetonitrile | VWR, Germany | HiPerSolv CHROMANORM ULTRA for LC-MS, 83642 | LC-MS grade required |
| formic acid | VWR, Germany | HiPerSolv CHROMANORM for LC-MS, 84865 | LC-MS grade required |
| 1 mL Ni-NTA IMAC column, e.g. HisTrap FF Crude | GE Healthcare, UK | 29-0486-31 | different manufacturers and resins available for IMAC |
| 0.45 µm bottle top filter unit | VWR, Germany | 10040-470 | sterile filtration of solutions using a vacuum pump |
| 0.45 µm syringe filter PVDF membrane | Carl Roth, Germany | CCY1.1 | sterile filtration of solutions using a syringe and to remove particles from cell lysates |
| luer-lock syringe, PP, 50 ml | Carl Roth, Germany | T552.2 | sterile filtration of solutions |
| 1.5 mL Eppendorf tubes | Eppendorf, Germany | 30120086 | |
| petri dishes (polystyrene, sterile) | Carl Roth, Germany | TA19 | |
| Nile red | Sigma-Aldrich, Germany | 72485 | |
| E. coli ΔproA strain | CGSC, Keio collection | JW0233-2 | proline-auxotrophic E. coli K-12 strain |
| E. coli B834(DE3) | Novagen (Merck), Germany | 69041 | methionine-auxotrophic E. coli B strain |
| λDE3 Lysogenization Kit | Novagen (Merck), Germany | 69734-3 | |
| Lactococcus lactis NZ9000 pNG nisPT | bacterial indicator strain, see Khusainov R & Kuipers OP, PLoS One, 8 (9), e74890 | ||
| benchtop centrifuge for 1.5 mL Eppendorf tubes | Eppendorf, Germany | 5427 R | |
| peristaltic pump | GE Healthcare, UK | P1 | |
| FPLC system | GE Healthcare, UK | Äkta Purifier 10 or the like | |
| inverted microscope | Nikon | TI Eclipse wide-field fluorescence microscope with 100x (N.A. 1.4) objective and Mercury Lamp | example setup for fluorescence microscopy |
| electron multiplying CCD (EMCCD) camera | Andor Technologies, UK | Andor Luca | example setup for fluorescence microscopy |
| fluorescence excitation filter | Thorlabs, USA | Dichroic cube (TLV-U-MF2-TRITC) | example setup for fluorescence microscopy |
| fluorescence emission filter | AHF Analysentechnik, Germany | T 560 LPXR | example setup for fluorescence microscopy |
| cover slip 24 x 60 mm | Carl Roth, Germany | LH26.1 | example setup for fluorescence microscopy |
| Immersion Oil | Carl Zeiss, Germany | Immersol 518 F | example setup for fluorescence microscopy |
| probe sonicator | Bandelin, Germany | Sonopuls HD3200 with sonotrode MS-72 | 200 W maximum HF output |
| C5 HPLC column (2.1x100 mm, 3 µm particle size) | Sigma-Aldrich, Germany | 567227-U | example setup for mass spectrometry |
| ESI-TOF coupled to HPLC system | Agilent, USA | Agilent 6530 Accurate Mass QTOF with 1260 HPLC | example setup for mass spectrometry |
Ссылки
- Ferri, M., Ranucci, E., Romagnoli, P., Giaccone, V. Antimicrobial resistance: A global emerging threat to public health systems. Crit Rev Food Sci Nutr. 57 (13), 2857-2876 (2017).
- Bahar, A. A., Ren, D. Antimicrobial peptides. Pharmaceuticals. 6 (12), 1543-1575 (2013).
- Ageitos, J. M., Sánchez-Pérez, A., Calo-Mata, P., Villa, T. G. Antimicrobial peptides (AMPs): Ancient compounds that represent novel weapons in the fight against bacteria. Biochem Pharmacol. 133, 117-138 (2017).
- Arnison, P. G., et al. Ribosomally synthesized and post-translationally modified peptide natural products: overview and recommendations for a universal nomenclature. Nat Prod Rep. 30 (1), 108-160 (2013).
- Lubelski, J., Rink, R., Khusainov, R., Moll, G. N., Kuipers, O. P. Biosynthesis, immunity, regulation, mode of action and engineering of the model lantibiotic nisin. Cell Mol Life Sci. 65 (3), 455-476 (2008).
- Shin, J. M., Gwak, J. W., Kamarajan, P., Fenno, J. C., Rickard, A. H., Kapila, Y. L. Biomedical applications of nisin. J Appl Microbiol. 120 (6), 1449-1465 (2016).
- Scherer, K. M., Spille, J. -H., Sahl, H. -G., Grein, F., Kubitscheck, U. The lantibiotic nisin induces lipid II aggregation, causing membrane instability and vesicle budding. Biophys J. 108 (5), 1114-1124 (2015).
- Jung, G. Lantibiotica - ribosomal synthetisierte Polypeptidwirkstoffe mit Sulfidbrücken und α,β-Didehydroaminosäuren. Angew Chemie. 103 (9), 1067-1084 (1991).
- Rink, R., et al. Lantibiotic structures as guidelines for the design of peptides that can be modified by lantibiotic enzymes. Biochemistry. 44 (24), 8873-8882 (2005).
- Lagedroste, M., Smits, S. H. J., Schmitt, L. Substrate Specificity of the Secreted Nisin Leader Peptidase NisP. Biochemistry. 56 (30), 4005-4014 (2017).
- Ross, A. C., Liu, H., Pattabiraman, V. R., Vederas, J. C. Synthesis of the lantibiotic lactocin S using peptide cyclizations on solid phase. J Am Chem Soc. 132 (2), 462-463 (2010).
- Fukase, K., et al. Synthetic Study on Peptide Antibiotic Nisin. V. Total Synthesis of Nisin. Bull Chem Soc Jpn. 65 (8), 2227-2240 (1992).
- Dumas, A., Lercher, L., Spicer, C. D., Davis, B. G. Designing logical codon reassignment - Expanding the chemistry in biology. Chem Sci. 6 (1), 50-69 (2015).
- Kuthning, A., Durkin, P., Oehm, S., Hoesl, M. G., Budisa, N., Süssmuth, R. D. Towards Biocontained Cell Factories: An Evolutionarily Adapted Escherichia coli Strain Produces a New-to-nature Bioactive Lantibiotic Containing Thienopyrrole-Alanine. Sci Rep. 6, 33447(2016).
- Piscotta, F. J., Tharp, J. M., Liu, W. R., Link, A. J. Expanding the chemical diversity of lasso peptide MccJ25 with genetically encoded noncanonical amino acids. Chem Commun (Camb). 51 (2), 409-412 (2015).
- Hartman, M. C. T., Josephson, K., Lin, C. -W., Szostak, J. W. An expanded set of amino acid analogs for the ribosomal translation of unnatural peptides. PLoS One. 2 (10), 972(2007).
- Johnson, J. A., Lu, Y. Y., Van Deventer, J. A., Tirrell, D. A. Residue-specific incorporation of non-canonical amino acids into proteins: recent developments and applications. Curr Opin Chem Biol. 14 (6), 774-780 (2010).
- Ibba, M., Söll, D. Aminoacyl-tRNAs: setting the limits of the genetic code. Genes Dev. 18 (7), 731-738 (2004).
- Studier, F. W., Moffatt, B. A. Use of bacteriophage T7 RNA polymerase to direct selective high-level expression of cloned genes. J Mol Biol. 189 (1), 113-130 (1986).
- Budisa, N., Steipe, B., Demange, P., Eckerskorn, C., Kellermann, J., Huber, R. High-level biosynthetic substitution of methionine in proteins by its analogs 2-aminohexanoic acid, selenomethionine, telluromethionine and ethionine in Escherichia coli. Eur J Biochem. 230 (2), 788-796 (1995).
- Rink, R., et al. Dissection and modulation of the four distinct activities of nisin by mutagenesis of rings A and B and by C-terminal truncation. Appl Environ Microbiol. 73 (18), 5809-5816 (2007).
- Kubyshkin, V., Durkin, P., Budisa, N. Energetic contribution to both acidity and conformational stability in peptide models. New J Chem. 40 (6), 5209-5220 (2016).
- Molloy, E. M., Field, D., O'Connor, P. M., Cotter, P. D., Hill, C., Ross, R. P. Saturation mutagenesis of lysine 12 leads to the identification of derivatives of nisin A with enhanced antimicrobial activity. PLoS One. 8 (3), 58530(2013).
- Wang, L., Brock, A., Herberich, B., Schultz, P. G. Expanding the genetic code of Escherichia coli. Science. 292 (5516), 498-500 (2001).
- Liu, C. C., Schultz, P. G. Adding new chemistries to the genetic code. Annu Rev Biochem. 79, 413-444 (2010).
- Anderson, J. C., Wu, N., Santoro, S. W., Lakshman, V., King, D. S., Schultz, P. G. An expanded genetic code with a functional quadruplet codon. Proc Natl Acad Sci U S A. 101 (20), 7566-7571 (2004).
- Zambaldo, C., Luo, X., Mehta, A. P., Schultz, P. G. Recombinant macrocyclic lanthipeptides incorporating non-canonical amino acids. J Am Chem Soc. 139 (34), 11646-11649 (2017).
- Al Toma, R. S., et al. Site-directed and global incorporation of orthogonal and isostructural noncanonical amino acids into the ribosomal lasso peptide capistruin. Chembiochem. 16 (3), 503-509 (2015).
- Chatterjee, A., Xiao, H., Schultz, P. G. Evolution of multiple, mutually orthogonal prolyl-tRNA synthetase/tRNA pairs for unnatural amino acid mutagenesis in Escherichia coli. Proc Natl Acad Sci U S A. 109 (37), 14841-14846 (2012).
- Budisa, N., Pal, P. P. Designing novel spectral classes of proteins with a tryptophan-expanded genetic code. Biol Chem. 385 (10), 893-904 (2004).
- Voller, J. -s, Thi To, T. M., Biava, H., Koksch, B., Budisa, N. Global substitution of hemeproteins with noncanonical amino acids in Escherichia coli with intact cofactor maturation machinery. Enzyme Microb Technol. 106, 55-59 (2017).
- Moghal, A., Hwang, L., Faull, K., Ibba, M. Multiple Quality Control Pathways Limit Non-protein Amino Acid Use by Yeast Cytoplasmic Phenylalanyl-tRNA Synthetase. J Biol Chem. 291 (30), 15796-15805 (2016).
- Datsenko, K. A., Wanner, B. L. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc Natl Acad Sci U S A. 97 (12), 6640-6645 (2000).
- Engelke, G., Gutowski-Eckel, Z., Hammelmann, M., Entian, K. D. Biosynthesis of the lantibiotic nisin: genomic organization and membrane localization of the NisB protein. Appl Environ Microbiol. 58 (11), 3730-3743 (1992).
- Baumann, T., Nickling, J. H., Bartholomae, M., Buivydas, A., Kuipers, O. P., Budisa, N. Prospects of In vivo Incorporation of Non-canonical Amino Acids for the Chemical Diversification of Antimicrobial Peptides. Front Microbiol. 8, 124(2017).
- Plat, A., Kluskens, L. D., Kuipers, A., Rink, R., Moll, G. N. Requirements of the engineered leader peptide of nisin for inducing modification, export, and cleavage. Appl Environ Microbiol. 77 (2), 604-611 (2011).
- JoVE Science Education Database. Basic Methods in Cellular and Molecular Biology. Bacterial Transformation: The Heat Shock Method. J Vis Exp. , (2017).
- JoVE Science Education Database. Basic Methods in Cellular and Molecular Biology. Bacterial Transformation: Electroporation. J Vis Exp. , (2017).
- Shi, Y., Yang, X., Garg, N., van der Donk, W. A. Production of lantipeptides in Escherichia coli. J Am Chem Soc. 133 (8), 2338-2341 (2011).
- Hochuli, E., Bannwarth, W., Döbeli, H., Gentz, R., Stüber, D. Genetic Approach to Facilitate Purification of Recombinant Proteins with a Novel Metal Chelate Adsorbent. Nat Biotechnol. 6 (11), 1321-1325 (1988).
- Zhang, Z., Marshall, A. G. A universal algorithm for fast and automated charge state deconvolution of electrospray mass-to-charge ratio spectra. J Am Soc Mass Spectrom. 9 (3), 225-233 (1998).
- JoVE Science Education Database. High-Performance Liquid Chromatography (HPLC). J Vis Exp. , (2017).
- JoVE Science Education Database. Dialysis: Diffusion Based Separation. J Vis Exp. , (2017).
- Khusainov, R., Kuipers, O. P. The presence of modifiable residues in the core peptide part of precursor nisin is not crucial for precursor nisin interactions with NisB- and NisC. PLoS One. 8 (9), 74890(2013).
- Terzaghi, B. E., Sandine, W. E. Improved medium for lactic streptococci and their bacteriophages. Appl Microbiol. 29 (6), 807-813 (1975).
- Schindelin, J., Rueden, C. T., Hiner, M. C., Eliceiri, K. W. The ImageJ ecosystem: An open platform for biomedical image analysis. Mol Reprod Dev. 82 (7-8), 518-529 (2015).
- Phosphate-buffered saline (PBS). Cold Spring Harb Protoc. 2006 (1), (2006).
- van Hest, J. C. M., Tirrell, D. A. Efficient introduction of alkene functionality into proteins in vivo. Febs Lett. 428 (1-2), 68-70 (1998).
- Prince, A., et al. Lipid-II Independent Antimicrobial Mechanism of Nisin Depends On Its Crowding And Degree Of Oligomerization. Sci Rep. 6 (1), 37908(2016).
- Zhou, L., et al. Incorporation of tryptophan analogues into the lantibiotic nisin. Amino Acids. 48 (5), 1309-1318 (2016).
- Presolski, S. I., Hong, V. P., Finn, M. G. Copper-Catalyzed Azide-Alkyne Click Chemistry for Bioconjugation. Curr Protoc Chem Biol. 3 (4), 153-162 (2011).
- Abts, A., et al. Easy and rapid purification of highly active nisin. Int J Pept. 2011, 175145(2011).
- Francklyn, C. S., First, E. A., Perona, J. J., Hou, Y. -M. Methods for kinetic and thermodynamic analysis of aminoacyl-tRNA synthetases. Methods. 44 (2), 100-118 (2008).
- van Heel, A. J., et al. Production and Modification of Five Novel Lantibiotics Using the Promiscuous Nisin Modification Machinery. ACS Synth Biol. 5 (10), 1146-1154 (2016).
- Wiegand, I., Hilpert, K., Hancock, R. E. W. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat Protoc. 3 (2), 163-175 (2008).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены