
Для просмотра этого контента требуется подписка на Jove Войдите в систему или начните бесплатную пробную версию.
Method Article
Функционализация атомной силы Микроскоп Кантилеверс с одно-T-клеток или одночастицы для иммунологической одноклеточной силы спектроскопии
В этой статье
Резюме
Мы представляем протокол для функционализации атомной силы микроскопа (AFM) кантилеверс с одной Т-клетки и частицы биса для иммунологических исследований. Показаны процедуры зондирования однопартной Т-дендритной связывания клеток AFM и мониторинга клеточной реакции макрофагов в реальном времени на одну твердую частицу с помощью флуоресценции.
Аннотация
Атомная силовая микроскопия на основе одноклеточной силовой спектроскопии (AFM-SCFS) является мощным инструментом для изучения биофизических свойств живых клеток. Этот метод позволяет зондировать прочности взаимодействия и динамики на мембране клетки в реальном маштабе времени, включая те между клетками, приемными устройствами и ligands, и наряду со многими другими изменениями. Он также работает как механизм для доставки физического или биохимического стимула на одиночных клетках в пространственно контролируемой манере, что позволяет конкретной активации клеток и последующих клеточных событий, которые будут контролироваться в режиме реального времени в сочетании с живой клеткой флуоресценция. Ключевым шагом в этих измерениях AFM-SCFS является функционализация AFM-cantilever, или, другими словами, присоединение предмета, интересующего к кантилевер. Здесь мы представляем методы модификации AFM кантилеверов с одной Т-клеток и одной бустирой полистирола соответственно для иммунологических исследований. Первый включает в себя биосовместимый клей, который пары одного Т-клеток на кончике плоского кантилевера в растворе, в то время как последний опирается на эпоксидный клей для одной саженки биса в воздушной среде. Предусмотрены также два иммунологических применения, связанных с каждой модификацией кантилевера. Описанные здесь методы могут быть легко адаптированы к различным типам клеток и твердым частицам.
Введение
Атомная микроскопия силы (AFM), универсальный инструмент, нашелмного применений в исследованиях клеточной биологии 1,2,3,4. Помимо своей высокой возможности изображения разрешения, родная функция зондирования силы позволяет биофизические свойства живых клеток, которые будут исследованы непосредственно на месте на одноклеточном уровне6,7. К ним относятся жесткости субклеточныхструктур или даже целые клетки 8,9,10,11,12, конкретные лиганд / рецептор связывания сильные силы на одномолекулярный уровень на поверхности клетки13,и силы стыковки междуоднопартными парами твердых частиц и клеток или между двумя клетками 1,2,14,15. Последние два часто классифицируются как одноклеточная спектроскопия силы (SCFS)16. Благодаря легкодоступным кантилеверам с различными весенними константами диапазон сил, доступный для AFM, довольно широк от нескольких пиконевтонов (pN) до микроньютонов (кН), который адекватно покрывает весь спектр клеточных событий, включающих силы из нескольких десятков pN, таких как рецептор на основе одномолекулярных связывания, на nN, таких как фагоцитные клеточные события15. Этот большой динамический диапазон сил делает AFM выгодным по сравнению с другими методами зондирования силы, такими как оптические/магнитные пинцеты и биомембранный зонд силы, так как они больше подходят для измерений слабой силы, с силой, как правило, менее 200 pN17 , 18. Кроме того, AFM может функционировать как высокоточный манипулятор для доставки различных стимулов на одиночные клетки в пространственно определенным образом4,19. Это желательно для проведения одноклеточных исследований в режиме реального времени. В сочетании с живой клеткой флуоресценции изображения, последующая клеточная реакция на конкретные стимулы могут контролироваться одновременно, что делает AFM основе SCFS чрезвычайно надежным, как оптические изображения, обеспечивая практический инструмент для зондирования клеточной сигнализации. Например, AFM был использован для определения штаммов, необходимых для получения преходящей кальция в остеобластах20. В этой работе, переходные кальция были отслежены флуоресцентно через кальций коэффициентметрической визуализации после применения локализованных сил на культивированных остеобластов с aFM отзыв. Недавно AFM был использован для растяжения колагенных фибрилков, на которых были выращены печеночные клетки стеллата (HSC), и эта механо-трансиндуцированная активация HSC в режиме реального времени контролировалась флуоресцентным биосенсором Src, фосфорилирование которого представлено Интенсивность флуоресценции биосенсора коррелирует с активацией HSC3.
В экспериментах SCFS на основе AFM правильная функционализация кантилеверов AFM является ключевым шагом на пути к успешным измерениям. Так как наш исследовательский интерес фокусируется на активации иммунных клеток, мы регулярно функционализируем кантилеверы с твердыми частицами, такими как отдельные твердые частицы, которые могут вызвать фагоцитоз и/или сильные иммунные реакции4,14 , 15 и одиночные Т-клетки, которые могут образовывать иммунный синапс с антигеном, представляя клетки, такие как активированные дендритные клетки (DC)2. Одиночные твердые частицы нормально соединены к cantilever через эпоксидный клей в окружающей среде воздуха, тогда как одиночные клетки T, из-за их non-клейкого природы, функционализированы к cantilever через биосовместимый клей в растворе. Здесь мы описываем методы выполнения этих двух типов модификации кантилевера и даем два связанных приложения. Первое приложение заключается в зондировании взаимодействий Т-клеток/DC с AFM-SCFS, чтобы понять подавляющий механизм регуляторных Т-клеток с точки зрения механики клеток. Второй включает в себя сочетание AFM с живой клеткой флуоресценции изображения для мониторинга клеточной реакции макрофага на твердую частицу в режиме реального времени, чтобы выявить молекулярный механизм рецептор-независимый фосфатидинозинол 4,5-бисфосфат (PIP2)- Моезин опосредованный фагоцитоз. Цель этого протокола заключается в том, чтобы обеспечить ориентировочную основу для заинтересованных исследователей для разработки и реализации своих собственных экспериментальных настроек с AFM на основе одноклеточного анализа для иммунологических исследований.
протокол
Протокол эксперимента мыши следует за директивами по внимательности животных университета Tsinghua
1. Функционализация кантилеев с одними Т-клетками
- Подготовка клеток селезенки мыши
- Пожертвуйте мышью (8-16 недель (либо пол); например, штамм C57BL/6 с использованием двуокиси углерода, а затем вывихшей шейки матки.
- Очистите мышь с 75% этанола и сделать разрез кожи средней линии следуют спленэктомии.
- Гомогенизировать селезенку в 4 мл PBS, содержащего 2% сыворотки крупного рогатого скота плода (FBS) с помощью стеклянных слайдов и удалить агрегаты и мусор, пройдя клеточной подвески через 70 мкм сетки нейлоновый ситечко.
- Центрифуги яточной подвески на 500 х г в течение 5 минут, отбросить супернатант и resuspend клетки в 2 мл крови люцинированного буфера (сбалансированный при комнатной температуре) в течение 5 мин. Прекратите реакцию лисиса, добавив 8 мл раствора PBS.
- Центрифуги яточной подвески на 500 х г в течение 5 минут и повторной концентрации клеток при плотности 1 х 108 ячеек/мл в PBS, содержащий 2% FBS и 1 мМ EDTA (маркированные как решение А), как правило, 0,25-2 мл в зависимости от плотности клеток. Перенесите переложенные ячейки на 5 мл (12 х 75 мм) полистирола круглой нижней трубки.
- Мышь CD4 "Т-клетки подготовки
- Добавьте в образец клетки, полученный с шагом 1.1.5, 50 крысоловных сывороток (см. таблицу материалов)и 50 коктейлей изоляции клеток (см. таблицу материалов). Смешайте и инкубировать в течение 10 минут при комнатной температуре.
- Vortex запас стрептавидин покрытием магнитной частицы решения (см. Таблица материалов) в течение 30 с или до тех пор, пока частицы появляются равномерно рассеяны.
- Добавьте в образец клетки 75 магнитных частиц с стрептавидином покрытием, на который внедряется 75 л/мл. Смешайте и инкубировать в течение 2,5 мин при комнатной температуре.
- Добавьте решение А, чтобы пополнить образец клетки до 2,5 мл и перемешайте, аккуратно понижая вверх и вниз в течение 2-3 раз.
- Поместите образец трубки (без крышки) в магнит (см. Таблицаматериалов) и инкубировать в течение 5 минут при комнатной температуре. Тщательно вылейте обогащенную суспензию клетки в новую 5 мл полистирола кругло-дно трубки.
- Центрифугия клеточной подвески на 500 х г в течение 5 мин. Отбросьте супернатант и resuspend обогащенных Т-клеток в 500 Л л раствора А.
ПРИМЕЧАНИЕ: Обогащенные клетки CD4'T содержат как обычные, так и регуляторные Т-клетки.
- Регулятивное отделение Т-клеток от обычных Т-клеток
- Добавьте 25 зл блокировщика FcR (см. ТаблицаМатериалов) к обогащенному образцу Т-клеток, полученному со ступени 1.2.6. Смешайте и инкубировать в течение 5 минут при комнатной температуре.
- Добавьте 25 Зл регулятивных T-клеток положительный отбор коктейль (см. Таблица материалов) в образец Т-клеток. Смешайте и инкубировать в течение 10 минут при комнатной температуре.
- Добавьте в образец Т-клеток 10 кЛ коктейля pe selection (см. таблицуматериалов). Смешайте и инкубировать в течение 5 минут при комнатной температуре.
- Vortex штепсить dextran-покрынный решение магнитной частицы (см. Таблица материалов) для 30 s или до тех пор пока частицы не будут появиться ровно рассеяно.
- Добавьте 10 зл магнитных частиц с деквентным покрытием в образец Т-клеток. Смешайте и инкубировать в течение 5 минут при комнатной температуре.
- Добавьте решение А, чтобы пополнить образец Т-клеток до 2,5 мл и перемешайте, аккуратно понижая вверх и вниз в течение 2-3 раз.
- Поместите Т-образную трубку (без крышки) в магнит и инкубировать в течение 5 минут при комнатной температуре. Аккуратно вылейте супернатант в новую трубку.
ПРИМЕЧАНИЕ: Супернатант содержит обогащенные обычные клетки CD4'T. - Centrifuge обогащенных обычных клеток CD4'T на 500 х г в течение 5 мин. Отбросьте супернатант и resuspend клетки в 4 мл RPMI1640, содержащий 10% FBS, 0,05 мм мм -Mercaptoethanol, 0,01 M HEPES и 1% пенициллин / стрептомицин (помечены как средний B).
- Удалите трубку, в которой регуляторные Т-клетки обогащаются от магнита. Добавьте 2,5 мл раствора А в трубку и перемешайте, аккуратно потянив вверх и вниз в течение 2-3 раз. Положите трубку обратно в магнит, инкубировать в течение 5 минут, а затем осторожно вылить и отбросить супернатант. Повторите этот шаг еще три раза.
- Приостановить обогащенные регуляторные Т-клетки в 2 мл среднего B.
- Инкубировать как очищенные обычные Т-клетки и регуляторные Т-клетки с 100 U/mL hIL-2 ночь или по крайней мере 4 ч при 37 градусах По Цельсию в увлажненный инкубатор с 5% CO2 перед использованием для функционализации кантилевера.
- Препарат дендритных клеток
- Подготовка раствора пираньи, смесь 30% H2O2 (30%) и 70% H2SO4 (conc) (v/v). Медленно налейте 3 мл H2O2 в 7 мл H2SO SO4 при постоянном перемешивании и охлаждении.
ВНИМАНИЕ: Пиранья раствор очень коррозионный, и он может сжигать и разрушать ткани тела. Поэтому безопаснее использовать раствор пираньи под капотом и носить соответствующее оборудование безопасности, так как смесь будет плескаться вокруг стакана. Нейтрализовать раствор с NaOH рН 7 после использования. - Погрузите стеклянный покрывало диаметром 24 мм в раствор пираньи в течение 30 минут и тщательно промыть с помощью стерильной ультрачистой воды.
- Опустите пару остроконечных пинцетов в 75% этанола в течение 30 минут для холодной дезинфекции.
- Введите очищенные стеклянные крышки в 6-колодую культурную тарелку пинцетом.
- Наклоните 6-сантиметровое блюдо культуры 6 см, в котором DC2.4 клетки были предварительно культивированы с 4 мл среднего B и аспирировать все среды. Добавьте 2 мл PBS в блюдо культуры, чтобы промыть клетки DC2.4 и отказаться от PBS. Повторите этот шаг промывки еще два раза.
- Добавьте 1 мл 0,25% трипсина EDTA в культурное блюдо в течение 2 мин. Добавьте 1 мл среднего B к этому блюду, чтобы положить конец реакции пищеварения фермента. Перенесите переваренные клеточные суспензии в трубку 15 мл.
- Центрифуги яточной подвески на 500 х г в течение 5 мин и resuspend DC2.4 клеток при плотности 2 х 105 клеток / мл в среднем B.
- Семена DC2.4 клетки на стеклянных крышках, приготовленные на шаг 1.4.4 и инкубировать клетки на ночь в увлажненной камере при 37 градусах Цельсия с 5% CO2.
ПРИМЕЧАНИЕ: Для измерения взаимодействующих сил между двумя одиночными ячейками, относительно низкая концентрация клеток DC2.4 (т.е. lt;10% confluency) необходима для того, чтобы иметь надлежащее расстояние между клетками.
- Подготовка раствора пираньи, смесь 30% H2O2 (30%) и 70% H2SO4 (conc) (v/v). Медленно налейте 3 мл H2O2 в 7 мл H2SO SO4 при постоянном перемешивании и охлаждении.
- AFM кантилевер подготовки
ПРИМЕЧАНИЕ: Кантилеверы, которые подходят для экспериментов с спектроскопией одноклеточной силы, являются те, с низкими константами пружины, как правило, в диапазоне 0,01-0,06 Н/м. Здесь мягкие кантилеверы без наконечников предпочтительнее для одиночных клеток и однозначных твердых частиц, функционализации.- Очистите кантилеверы путем обработки Piranha или плазмы или УФ-озона очистки.
- Установите очищенный кантилевер к голове сканирования AFM.
- Подготовьте чистую образец камеры, наполненной чистой водой, и откалибруйте кантилевер в водном растворе, сначала запустив кривую силы на стеклянном субстрате, чтобы получить чувствительность (наклон линейной пригонки над отталкивающей частью приближающейся кривой), а затем запись теплового шума спектра для извлечения весны постоянной в соответствии с инструкцией руководства AFM.
- Удалите aFM сканирующую голову из раствора, вымойте установленный кантилевер несколькими каплями чистого этанола и держите кантилевер сухим на сканирующей голове.
- Прикрепление одиночных Т-клеток к кантилеверу
- Разогреть корпус среды обитания живых клеток с 5% CO2 при температуре 37 градусов по Цельсию.
- Установите стеклянный покрывало с клетками DC2.4, выращенными на нем от шага 1.4.8 до сборки камер ы образца, добавьте 600 qL среднего B в камеру немедленно, а затем поместите сборку на этап образца AFM.
- Добавьте в камеру образца инкубированные клетки CD4-T (обычные или регулятивные Т-клетки) hIL-2.
ПРИМЕЧАНИЕ: Общий объем выборки не должен превышать 1 мл. - Подождите, пока добавленные ячейки CD4'T полностью установятся на дне крышки.
ПРИМЕЧАНИЕ: Воздушные пузыри вызовут большое помехи для эксперимента, поэтому желательно избегать пузырьков воздуха в step1.6.2 и 1.6.3. - Добавьте каплю 2 юл биосовместимого клея на конец установленного кантилевера с пипеткой, как показано на рисунке 1, а затем поместите сканирующую голову на этап образца быстро, тем самым позволяя кантилеверу, покрытому биосовместимым клеем, погрузиться в Решение.
ВНИМАНИЕ: Не прикасайтесь к стеклянному блоку или кантилеверу кончиком пипетки. Поскольку биосовместимый клей, используемый здесь, склонен к окислению в воздухе, этот шаг следует сделать как можно быстрее. - Найдите здоровую Т-клетку под кончиком кантилевера грубо под микроскопом, переместив этап образца, а затем мелко отрегулируйте позиционирование, переместив сканирующую головку.
ПРИМЕЧАНИЕ: Здоровая ячейка CD4'T, как правило, имеет относительно большой размер, гладкие края и оптически трансмиссивную в ярко-поле изображения. - Опустите кантилевер вручную с размерами шага, начиная с 50 мкм, затем до 10, 5, 2 и 0,5 мкм постепенно, контролируя двигатели степпера. Удерживайте положение двигателей степпера и отрегулируйте позиционирование сканирующей головки для лучшего выравнивания между кончиком кантилевера и клеткой, как только кантилевер вступает в твердый контакт с целевой Т-клеткой, о чем свидетельствует небольшое смещение лазерного луча положение в фотодетекторе, соответствующее типичному диапазону силы 0,5-1,5 нн.
ПРИМЕЧАНИЕ: Этот шаг также может быть сделано путем запуска одного измерения силы, в котором набор-точка (сила применяется к ячейке) и время контакта может быть четко определена в программном обеспечении. Однако, из-за неклейкого характера Т-клеток, ручной подход обеспечивает большую гибкость в управлении прицеливанием, позиционированием и временем контакта, чем автоматическое приближение, и он надежно работает для сцепления Т-клеток. Будущие экспериментаторы должны попробовать как ручной и автоматический подход, чтобы выяснить, какие работает лучше для их систем интересов. - Убирать кантилевер после 30 с контакта.
ПРИМЕЧАНИЕ: Если ячейка движется с кантилевером, вложение успешно. Если нет, повторите шаг 1.6.6, но на другой Т-клетке. Биосовместимый клей легко окисляется. Шаг 1.6.5-1.6.7 должен быть завершен в течение 5 минут. Кроме того, если один и тот же кантилевер трижды выходит из строя для вложения Т-клеток, следует использовать новый кантилевер, а процедура крепления должна начинаться с шага 1.5.2 снова.

Рисунок 1: Схематическое представление добавления небольшой капли биосовместимого клея на установленный кантилевер. Кантилевер устанавливается через зажимную пружину на держателе стеклянного блока, который устанавливается на сканирующую головку AFM (не нарисованную здесь). Когда сканирующая головка стоит на выровневой поверхности, кантилевер вертикально ориентирован, как показано на рисунке. Около 2 биосовместимых клея л можно добавить к кончику кантилевера с микро-пипеткой. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
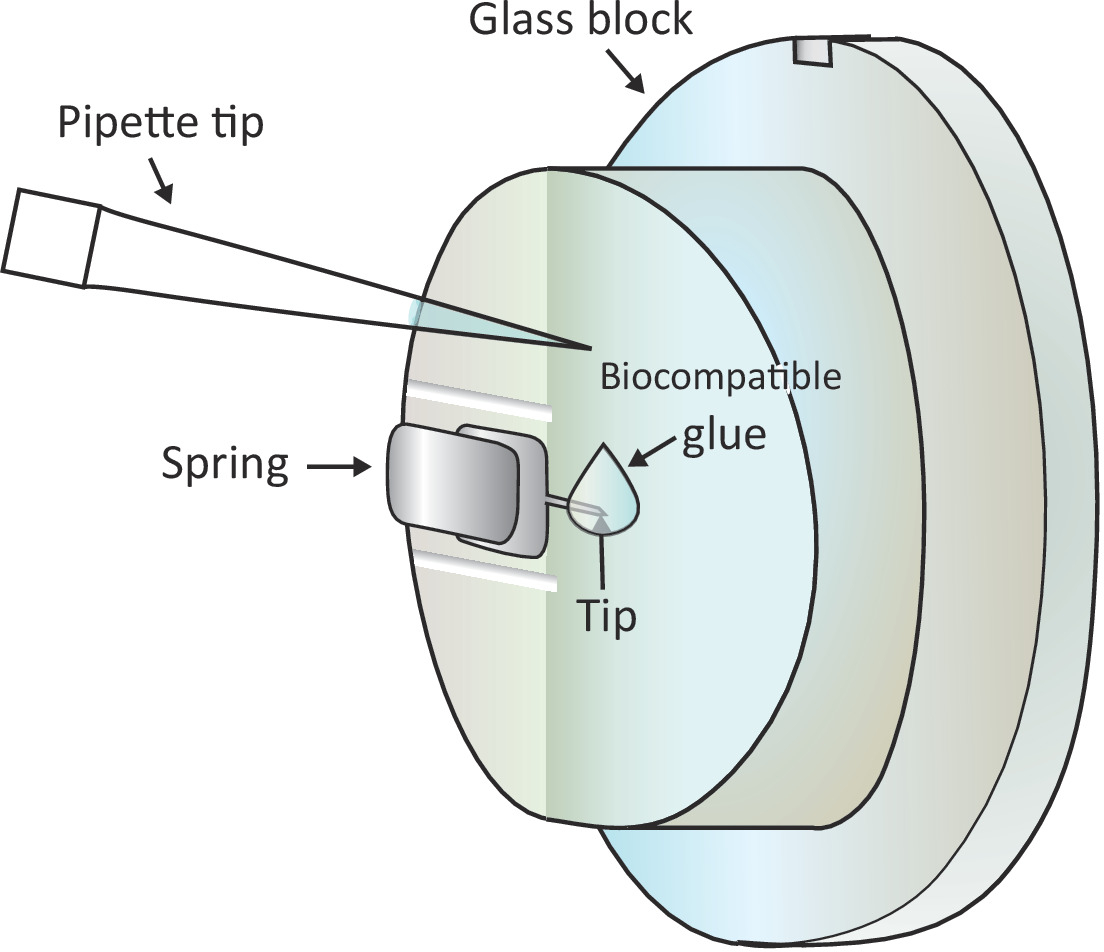{kind=link}
- Силовая спектроскопия однопартного Т-клеточного/дендритного взаимодействия клеток
ПРИМЕЧАНИЕ: Для зондирования взаимодействия клеток/клеток требуется AFM с диапазоном, превышающее обычные 10-15 мкм, чтобы полностью отделить две клетки. AFM, используемый здесь, имеет диапазон в 100 мкм, что является достаточным для отделения Т-клеток от дендритной клетки после контакта клетки/клетки.- Расположите прикрепленную Т-клетку над отдельной ячейкой DC2.4, переместив этап образца и/или сканирующую головку (см. рисунок 2).
- Установите правильные параметры и запустите силовую спектроскопию.
ПРИМЕЧАНИЕ: Обычно используются следующие ключевые настройки: Setpoint 0.5 nN, Тяговая длина 50 мкм, постоянная скорость движения, скорость расширения 5 мкм/с, Время контакта 10 с, Постоянная сила режима задержки. Для каждой пары T-DC собрано 20 повторов кривых силы и не менее 14 кривых силы. - Установите новый очищенный кантилевер, откалибруйте его в чистой воде, как в шаге 1.5.3, и вернитесь к тому же образцу Т-DC-клеток, чтобы повторить шаг 1.6 и 1.7 для другой пары T-DC. Зонд по крайней мере 5 пар для каждого состояния.

Рисунок 2: Экспериментальная конфигурация силового зондирования между одной Т-ячейкой и DC. (A) Схематический рисунок экспериментальной конфигурации, в которой Т-клетка, прикрепленная к кантилеверу, привозима в DC, выращенный на субстрате для силового зондирования. (B) Ярко-поле изображение Т-клеток функционализованной кантилевер и DC. Шкала бар, 20 мкм. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
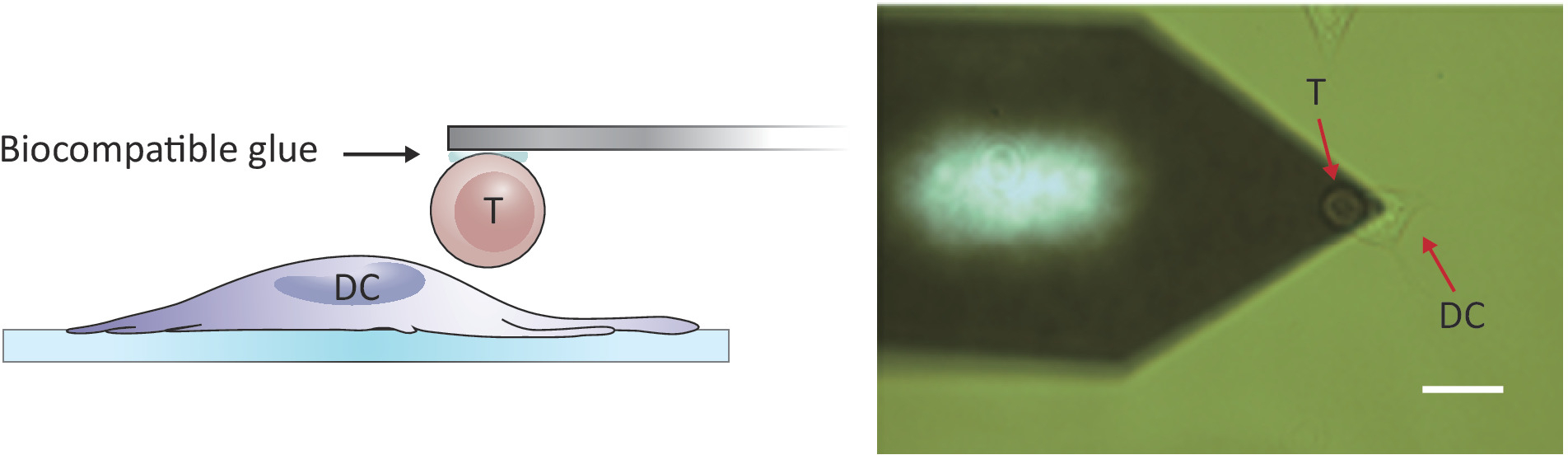{kind=link}
2. Функционализация кантилевера с одной полистироловых шариков
- Подготовка однобиеных шариков
- Разбавить запас нойся подвеской 6 мкм полистирола в 100% этанола.
ПРИМЕЧАНИЕ: Концентрация разбавленного раствора бисера должна быть достаточно низкой, чтобы при добавлении к стеклянной поверхности крышки, отдельные бусы хорошо разделены без значительного кластеризации после испарения растворителя. - Очистите стеклянный покрывало диаметром 24 мм с помощью этанола и удалите любую пыль воздушным потоком N 2.
- Установите очищенное стекло крышкой к сборке камеры образца и положите сборку на микроскоп.
- Положите каплю разбавленного раствора бисера на левую сторону, но близко к центру крышки (см. Рисунок3) и проверьте расстояние между бисером после испарения растворителя в ярком поле под микроскопом с целью 20x. Переходке к следующему шагу, если отдельные бусинки хорошо разделены.
- Опустите кончик микропайпета или зубочистку в хорошо смешанный эпоксидный клей, а затем перенесите небольшое количество такого клея в три отдельных места с последовательными нежными касаниями на правой стороне, но близко к центру крышки.
ПРИМЕЧАНИЕ: Три клея пятна должны быть вертикально выровнены (см. Рисунок 3). Последнее место с наименьшим количеством клея будет использовано позже.
- Разбавить запас нойся подвеской 6 мкм полистирола в 100% этанола.

Рисунок 3: Схематическое представление рабочего потока для функционализации одношариков на кантилевере. Хорошо разделенные микрон размера бусы готовятся на левой стороне субстрата и небольшое количество эпоксидного клея передается на правую сторону субстрата через 3 последовательных нежных прикосновений, в результате чего 3 клея пятна. Только последнее место с наименьшим количеством клея (указано кругом) используется, чтобы покрыть самый конец кантилевера. Подход ккильтер в клей слева, а затем переместить кантилевер назад, как только он погружается в клей, чтобы ограничить клей в самом конце кантилевера. Принесите целевой бис под кантилевер и выровнять их должным образом, прежде чем сделать твердый контакт (обычно 2-5 nN) для стегесы биса. Когда бисуспешно успешно функционализирован на кантилевере, новый кантилевер может быть установлен, чтобы начать новый цикл функционализации. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
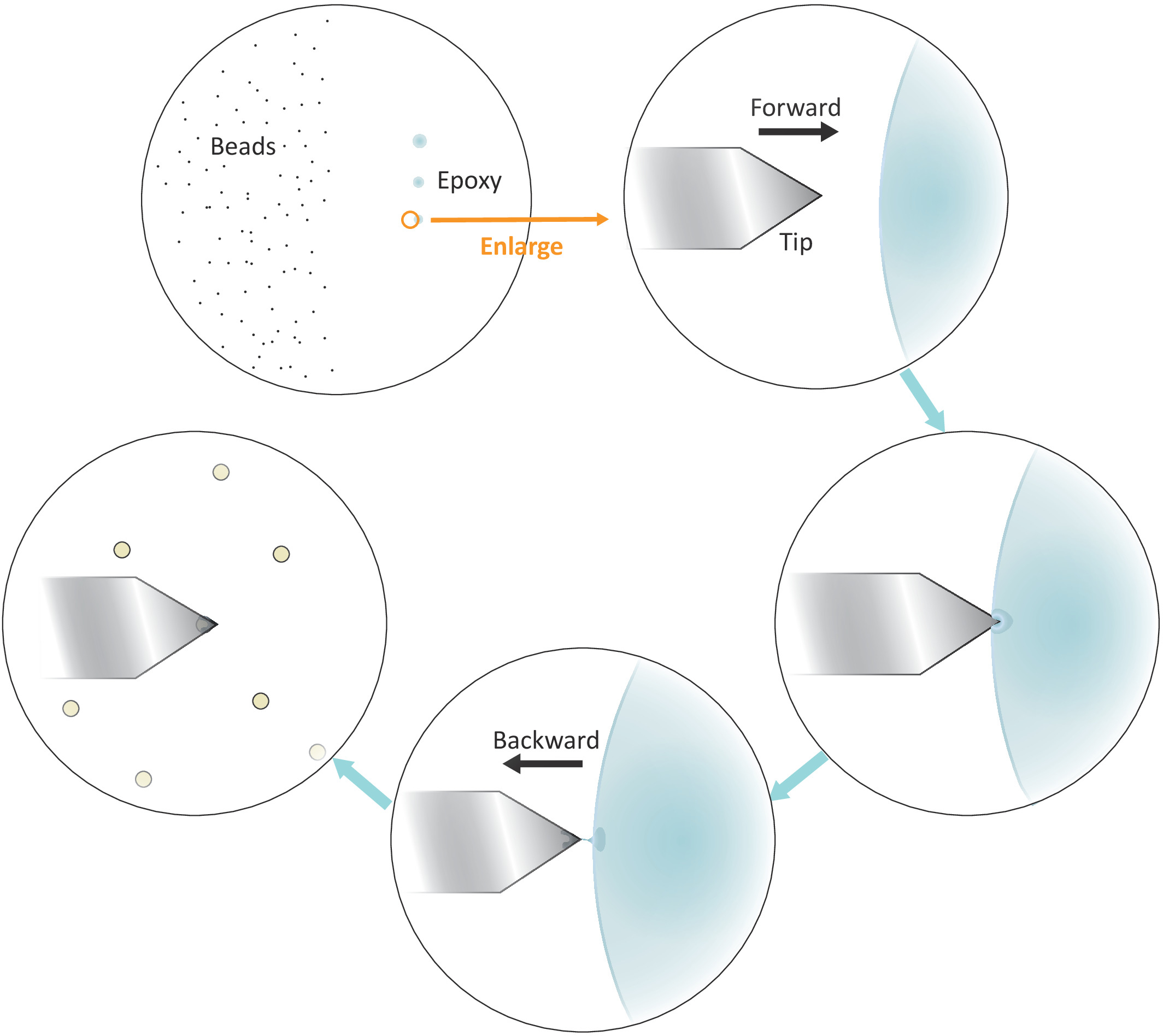{kind=link}
- AFM кантилевер подготовки
- Установите очищенный наконечник менее кантилевер на aFM сканирования головы.
- Откалибровать этот кантилевер в воздухе с чистой поверхностью, чтобы получить пружинную константу.
- Прикрепление одиночных шариков к кантилеверу
- Расположите кончик кантилевера над левой границей последнего пятна эпоксидного клея, как показано на рисунке 3.
- Принесите кантилевер близко к клею медленно, опустив ступенчатые двигатели с небольшими размерами шага.
- Вытяните кантилевер от быстро от клея боковой путем перемещения AFM сканирования голову назад (влево) вручную, как только кончик погружается в клей.
ПРИМЕЧАНИЕ: Убедитесь, что только небольшое количество клея придерживается в самом конце кончика. Если есть чрезмерный клей на кончике, можно уменьшить количество клея, касаясь следуют скольжения кончика на пустой поверхности. - Переместите кончик кантилевера поверх хорошо изолированной одной бусы.
- Подойдите к кантилеверу к одной бисерной басоле медленно и сделать прочный контакт с бисером (как указано смещение положения лазерного луча в фотодетекторе, соответствующий типичному диапазону силы 2-5 nN) около 10 с, в течение которых тонкая регулировка т IP позиционирование боковой поможет лучше найти бисовву в самом конце кончика. Уречку наконечника в конце контакта.
ПРИМЕЧАНИЕ: Исчезновение самого биса из исходного фокусного плана указывает на успешное событие адепта. - Сделать бисиню модифицированный кантилевер тщательно и хранить его в кантилевер поле на ночь для полного затвердевания клея.
- Флуоресценция визуализации клеточной реакции макрофага на одну бисо, доставленную AFM.
ПРИМЕЧАНИЕ: Флуоресценция была выполнена на самодельных объективного типа общего внутреннего отражения флуоресценции микроскоп на основе коммерческого микроскопа стенда. Эта система визуализации оснащена 4 лазерными источниками (405 нм, 488 нм, 561 нм, 647 нм), сплиттером для двухцветного обнаружения и электронным умножающим зарядом в паре с устройством (EMCCD) для широкоугольной визуализации.- Выращивайте RAW264.7 клетки на стеклянном покрывале при 37 градусах по Цельсию в 5% CO2 увлажненной камере.
- Transfect Moesin-EGFP и ПЛКЗ-PH-mCherry к RAW264.7 клеткам с помощью трансфекционного комплекта (см. Таблица материалов) в рамках протокола производителя, чтобы флуоресцентно визуализировать моэсин и фосфатидинозитол 4,5-бисфосфат (PIP2) молекулы Соответственно.
ПРИМЕЧАНИЕ: Moesin имеет мотив ITAM, который может активировать Сик, ключевой игрок в фагоцитоз. ИЗВЕСТНО, что PIP2 завербует Moesin в клеточную мембрану. - Положите стеклянный покрывало с клетками на сборку камер образца и установите сборку на этап образца AFM.
- Установите бис модифицированного кантилевера к голове сканирования AFM.
- Запустите кривую силы в пустой области и откалибруйте силу с чувствительностью из этой кривой и константой пружины, измеренной в шаге 2.2.2.
- Найдите хорошо изолированную ячейку с надлежащей интенсивностью флуоресценции как в зеленом (Moesin-EGFP), так и в красных (ПЛКЗ-PH-mCherry) каналах с 488/561 нм возбуждения.
- Доставка голый 6 мкм полистирола с AFM на поверхность клетки с 1 nN постоянной силы и 500 с контактным временем.
- Запись флуоресценции изображения серии клетки в контакте с бисодля для анализа (обычно 10 кадров / с).
ПРИМЕЧАНИЕ: Чтобы уменьшить фотоотбеление флюорофоров, относительно низкая сила возбуждения должна быть использована для поиска клеток, представляющих интерес. Кроме того, для продления времени флуоресценции можно использовать схему прерывистой возбуждения, если динамика реакций клеток находится в медленном временном масштабе.
Результаты
На рисунке 4A показаны типичные кривые силового расстояния от связывающего взаимодействия между одно-Т-клеточной и однодневкой в одном цикле подхода-оттягивания. Светло-красная кривая — кривая расширения, а темно-красная — кривая опровержения. Поскол...
Обсуждение
Спектроскопия одноклеточной силы на основе AFM превратилась в мощный инструмент для решения биофизических свойств живых клеток. Для этих приложений, кантилевер должен быть функционировал должным образом для того, чтобы зондировать конкретные взаимодействия или свойства на клетках, пр...
Раскрытие информации
Авторам нечего раскрывать.
Благодарности
Эта работа поддерживается Национальным фондом естественных наук Китая Общей программы (31370878), Государственной ключевой программы (31630023) и Инновационной исследовательской группы программы (81621002).
Материалы
| Name | Company | Catalog Number | Comments |
| Material | |||
| 10 μl pipette tip | Thermo Fisher | 104-Q | |
| 15 ml tube | Corning | 430791 | |
| 6 cm diameter culture dish | NALGENE nunc | 150462 | |
| 6-well culture plate | JET | TCP011006 | |
| AFM Cantilever | NanoWorld | Arrow-TL1-50 | tipless cantilever |
| β-Mercaptoethanol | Sigma | 7604 | |
| Biocompatible glue | BD Cell-Tak | 354240 | |
| CD4+ T cell isolation Cocktail | STEMCELL | 19852C.1 | |
| DC2.4 cell line | A gift from K. Rock (University of Massachusetts Medical School, Worcester, MA) | ||
| Dextran-coated magnetic particles | STEMCELL | SV30010 | |
| EDTA | GENEray | Generay-E1101-500 ml | |
| Epoxy | ERGO | 7100 | |
| Ethanol | twbio | 00019 | |
| FBS | Ex Cell Bio | FSP500 | |
| FcR blocker | STEMCELL | 18731 | |
| Glass coverslip | local vender (Hai Men Lian Sheng) | HX-E37 | 24mm diameter, 0.17mm thinckness |
| Glass slides | JinTong department of laboratory and equipment management, Haimen | N/A | customized |
| H2O2 (30%) | Sino pharm | 10011218 | |
| H2SO4 | Sino pharm | 80120892 | |
| HEPES | Sigma | 51558 | |
| Magnet | STEMCELL | 18000 | |
| Mesh nylon strainer | BD Falcon | REF 352350 | |
| Moesin-EGFP | N/A | cloned in laboratory | |
| Mouse CD25 Treg cell positive isolation kit | STEMCELL | 18782 | Component: FcR Blocker,Regulatory T cell Positive Selection Cocktail, PE Selection Cocktail, Dextran RapidSpheres, |
| Mouse CD4+ Tcell isolation kit | STEMCELL | 19852 | Component:CD4+T cell isolation Cocktail, Streptavidin RapidSpheres, Rat Serum |
| NaOH | Lanyi chemical products co., LTD? Beijing | 1310-73-2 | |
| PBS | Solarbio | P1022-500 | |
| PE selection cocktail | STEMCELL | 18151 | |
| Penicillin-Streptomycin | Hyclone | SV30010 | |
| PLCδ-PH-mCherry | Addgene | 36075 | |
| Polystyrene microspheres 6.0μm | Polysciences | 07312-5 | |
| polystyrene round bottom tube | BD Falcon | 352054 | |
| Rat serum | STEMCELL | 13551 | |
| RAW264.7 | ATCC | ||
| Recombinant Human Interleukin-2 | Peprotech | Peprotech, 200-02-1000 | |
| Red blood cell lysis buffer | Beyotime | C3702 | |
| Regulatory T cell positive selection cocktail | STEMCELL | 18782C | |
| RPMI 1640 | Life | C11875500BT | |
| Sample chamber | Home made | ||
| Streptavidin-coated magnetic particles | STEMCELL | 50001 | |
| Transfection kit | Clontech | 631318 | |
| Trypsin 0.25% EDTA | Life | 25200114 | |
| Tweezers | JD | N/A | |
| Name | Company | Catalog Number | Comments |
| Equipment | |||
| 20x objective NA 0.8 | Zeiss | 420650-9901 | Plan-Apochromat |
| Atomic force microscope | JPK | cellHesion200 | |
| Centrifuge | Beckman coulter | Allegra X-12R | |
| Fluorescence imaging | home-made objective-type total internal reflection fluorescence microscop based on a Zeiss microscope stand | ||
| Humidified CO2 incubator | Thermo Fisher | HERACELL 150i | |
| Inverted light microscope | Zeiss | Observer A1 manual |
Ссылки
- Benoit, M., Gabriel, D., Gerisch, G., Gaub, H. E. Discrete interactions in cell adhesion measured by single-molecule force spectroscopy. Nature Cell Biology. 2 (6), 313-317 (2000).
- Chen, J., et al. Strong adhesion by regulatory T cells induces dendritic cell cytoskeletal polarization and contact-dependent lethargy. Journal of Experimental Medicine. 214 (2), 327-338 (2017).
- Liu, L., et al. Mechanotransduction-modulated fibrotic microniches reveal the contribution of angiogenesis in liver fibrosis. Nature Materials. 16 (12), 1252-1261 (2017).
- Mu, L. B., et al. A phosphatidylinositol 4,5-bisphosphate redistribution-based sensing mechanism initiates a phagocytosis programing. Nature Communications. 9, (2018).
- Qi, C., et al. Pathology-targeted cell delivery via injectable micro-scaffold capsule mediated by endogenous TGase. Biomaterials. 126, 1-9 (2017).
- Muller, D. J., Helenius, J., Alsteens, D., Dufrene, Y. F. Force probing surfaces of living cells to molecular resolution. Nature Chemical Biology. 5 (6), 383-390 (2009).
- Muller, D. J., Dufrene, Y. F. Atomic force microscopy: a nanoscopic window on the cell surface. Trends in Cell Biology. 21 (8), 461-469 (2011).
- Radotic, K., et al. Atomic force microscopy stiffness tomography on living Arabidopsis thaliana cells reveals the mechanical properties of surface and deep cell-wall layers during growth. Biophysics Journal. 103 (3), 386-394 (2012).
- Kuznetsova, T. G., Starodubtseva, M. N., Yegorenkov, N. I., Chizhik, S. A., Zhdanov, R. I. Atomic force microscopy probing of cell elasticity. Micron. 38 (8), 824-833 (2007).
- Scheuring, S., Dufrene, Y. F. Atomic force microscopy: probing the spatial organization, interactions and elasticity of microbial cell envelopes at molecular resolution. Molecular Microbiology. 75 (6), 1327-1336 (2010).
- Berdyyeva, T. K., Woodworth, C. D., Sokolov, I. Human epithelial cells increase their rigidity with ageing in vitro: direct measurements. Physics in Medicine and Biology. 50 (1), 81-92 (2005).
- Sokolov, I., Dokukin, M. E., Guz, N. V. Method for quantitative measurements of the elastic modulus of biological cells in AFM indentation experiments. Methods. 60 (2), 202-213 (2013).
- Bozna, B. L., et al. Binding strength and dynamics of invariant natural killer cell T cell receptor/CD1d-glycosphingolipid interaction on living cells by single molecule force spectroscopy. Journal of Biological Chemistry. 286 (18), 15973-15979 (2011).
- Flach, T. L., et al. Alum interaction with dendritic cell membrane lipids is essential for its adjuvanticity. Nature Medicine. 17 (4), 479-487 (2011).
- Ng, G., et al. Receptor-independent, direct membrane binding leads to cell-surface lipid sorting and Syk kinase activation in dendritic cells. Immunity. 29 (5), 807-818 (2008).
- Helenius, J., Heisenberg, C. P., Gaub, H. E., Muller, D. J. Single-cell force spectroscopy. Journal of Cell Science. 121 (11), 1785-1791 (2008).
- Litvinov, R. I., Shuman, H., Bennett, J. S., Weisel, J. W. Binding strength and activation state of single fibrinogen-integrin pairs on living cells. Proceedings of the National Academy of Sciences of the United States of America. 99 (11), 7426-7431 (2002).
- Evans, E., Ritchie, K., Merkel, R. Sensitive Force Technique to Probe Molecular Adhesion and Structural Linkages at Biological Interfaces. Biophysics Journal. 68 (6), 2580-2587 (1995).
- Lamprecht, C., Hinterdorfer, P., Ebner, A. Applications of biosensing atomic force microscopy in monitoring drug and nanoparticle delivery. Expert Opinion on Drug Delivery. 11 (8), 1237-1253 (2014).
- Charras, G. T., Horton, M. A. Single cell mechanotransduction and its modulation analyzed by atomic force microscope indentation. Biophysics Journal. 82 (6), 2970-2981 (2002).
- Sun, M. Z., et al. Multiple membrane tethers probed by atomic force microscopy. Biophysics Journal. 89 (6), 4320-4329 (2005).
- Yan, J. C., Liu, B., Shi, Y., Qi, H. Class II MHC-independent suppressive adhesion of dendritic cells by regulatory T cells in vivo. Journal of Experimental Medicine. 214 (2), 319-326 (2017).
- Hao, J. J., et al. Phospholipase C-mediated hydrolysis of PIP2 releases ERM proteins from lymphocyte membrane. Journal of Cell Biology. 184 (3), 451-462 (2009).
- Rodriguez, R. M., et al. Lymphocyte-T Adhesion to Fibronectin (Fn) - a Possible Mechanism for T-Cell Accumulation in the Rheumatoid Joint. Clinical and Experimental Immunology. 89 (3), 439-445 (1992).
- Kimura, A., Ersson, B. Activation of Lymphocytes-T by Lectins and Carbohydrate-Oxidizing Reagents Viewed as an Immunological Recognition of Cell-Surface Modifications Seen in the Context of Self Major Histocompatibility Complex Antigens. European Journal of Immunology. 11 (6), 475-483 (1981).
- Miller, K. The Stimulation of Human Lymphocyte-B and Lymphocyte-T by Various Lectins. Immunobiology. 165 (2), 132-146 (1983).
- Vitte, J., Pierres, A., Benoliel, A. M., Bongrand, P. Direct quantification of the modulation of interaction between cell- or surface-bound LFA-1 and ICAM-1. Journal of Leukocyte Biology. 76 (3), 594-602 (2004).
- Beaussart, A., et al. Quantifying the forces guiding microbial cell adhesion using single-cell force spectroscopy. Nature Protocols. 9 (5), 1049-1055 (2014).
- Shu, F., et al. Cholesterol Crystal-Mediated Inflammation Is Driven by Plasma Membrane Destabilization. Frontiers in Immunology. 9, (2018).
- Hosseini, B. H., et al. Immune synapse formation determines interaction forces between T cells and antigen-presenting cells measured by atomic force microscopy. Proceedings of the National Academy of Sciences of the United States of America. 106 (42), 17852-17857 (2009).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены