
Method Article
Оптогенетические манипуляции нейронной активности модулировать поведение в свободно движущихся мышей
В этой статье
Резюме
С оптогенетической манипуляции конкретных популяций нейронов или областей мозга, поведение может быть изменено с высоким временным и пространственным разрешением в свободно движущихся животных. С помощью различных оптогенетических инструментов в сочетании с хронически имплантированных оптических волокон, различные нейрональные модуляции и поведенческие испытания могут быть выполнены.
Аннотация
Оптогенетическая модуляция нейронных цепей у свободно движущихся мышей влияет на острое и долгосрочное поведение. Этот метод способен выполнять манипуляции отдельных нейронов и региона конкретных передатчик релиз, до целых нейронных схем в центральной нервной системе, и позволяет прямое измерение поведенческих результатов. Нейроны выражают оптогенетические инструменты через инъекцию вирусных векторов, несущих ДНК выбора, таких как Channelrhodopsin2 (ChR2). Свет попадает в определенные области мозга с помощью хронических оптических имплантатов, которые прекращаются непосредственно над целевой областью. После двух недель восстановления и правильного инструмента-выражения, мыши могут быть неоднократно использованы для поведенческих тестов с оптогенетической стимуляции нейронов, представляющих интерес.
Оптогенетическая модуляция имеет высокое временное и пространственное разрешение, которое может быть достигнуто с высокой специфичностью клеток, по сравнению с широко используемыми методами, такими как химическая или электрическая стимуляция. Свет не вредит нейронной ткани и поэтому может быть использован для долгосрочных экспериментов, а также для нескольких поведенческих экспериментов в одной мыши. Возможности оптогенетических инструментов практически безграничны и позволяют активации или замалчивания целых нейронов, или даже манипуляции конкретного типа рецепторов светом.
Результаты таких поведенческих экспериментов с интегрированной оптогенетической стимуляцией непосредственно визуализируют изменения в поведении, вызванные манипуляцией. Поведение одного и того же животного без стимуляции света в качестве базового является хорошим контролем для индуцированных изменений. Это позволяет детальное обзор типов нейронов или нейромедиатора систем, участвующих в конкретных поведениях, таких как тревога. Пластичность нейронных сетей также может быть исследована в очень подробно через долгосрочную стимуляцию или поведенческие наблюдения после оптической стимуляции. Оптогенетика поможет просветить нейронную сигнализацию при нескольких видах неврологических заболеваний.
Введение
Модуляция нейронных цепей в центральной нервной системе и их поведенческие результаты важны для понимания того, как работает мозг, особенно при психических заболеваниях и когнитивных задачах, таких как обучение и память. С оптогенетикой, одиночные клетки или населенности клетки до целых схем можно модулировать светом. Общие оптогенетические инструменты, такие как Channelrhodopsin2 (ChR2) или Archaerhodopsin (Арка) способны активировать или заставить замолчать нейроны, или увеличить или ингибировать высвобождение передатчика на аксоновыхтерминалах,проецирующих на различные области мозга1,2,3,4. Тем не менее, Арка должна быть использована тщательно, как было показано, что его активация на пресинаптических терминалах увеличивает спонтанный релиз передатчика5. Арка является внешним исправление протонного насоса, который изменяет значение рН внутри клетки. Эта щелочная среда индуцирует приток кальция и усиливает высвобождение передатчика5. Специально модулировать внутриклеточные сигнальные пути, рецепторные химеры, состоящие из легкого активируемого оптогенетического инструмента, такого как родопсин или конусный опсин, в сочетании с адекватным G-белковым рецептором,могут быть созданы 6,,7,8. Количество и вариации оптогенетических инструментов, доступных значительно увеличилось за последнее десятилетие9.
Целью оптогенетики является манипулирование нейронными схемами во время поведения. Оптогенетика позволяет, например, измерять острые поведенческие изменения, такие как изменения в поведении тревоги. Оптогенетические инструменты доставляются в целевые области мозга с помощью вирусных векторов. С помощью специальных промоутеров и усилителей, или Cre-loxP системы, специфика типа клетки может быть обеспечена для выражения оптогенетических инструментов (рисунок 1A). Есть несколько генетически модифицированных линий мыши, выражают фермент Cre-Recombinase только в конкретных типах клеток. Например, мышей Nex-Cre выразить Cre-Recombinase в пирамидальных нейронов в коре головного мозга и гиппокампа под контролем Nex-продвигателя10. Этот фермент способен инвертировать ДНК-последовательности, которые окружены локсP сторон11. Следовательно, ДНК-последовательность двойной флоксированный оптогенетический инструмент, который инвертирован и в окружении loxP сторон, может быть транскрибирована только нейронов, которые обладают Cre-Recombinase, но не другие нейрональныетипы 12,13. В случае мышей Nex-Cre, оптогенетический инструмент будет выражен исключительно в пирамидальных нейронов. Световая стимуляция некоторых областей мозга достигается путем хронической имплантации оптических волокон непосредственно над областью интереса. Затем животные могут быть соединены с подходящим источником света и свободно вести себя почти во всех видах поведенческих тестов.

Рисунок 1: Инъекция и имплантация. A) Cre-loxP система для ChR2-YFP. Двойной floxed оптогенетический инструмент упакован в адено связанных вирусов (AAV) для инъекций в ткани мозга. B) Стрельцое представление о впрыске вируса и имплантации оптического нейронального интерфейса в/над областью IL mPFC. Инъекции и имплантация были сделаны сверху. Показаны все регионы, представляющие интерес, IL, BLA и DRN. C)Детальный обзор имплантированного оптического волокна, рукава и источника света. D)Распространение стимуляции синего и красного лазерного света в тканях мозга серого вещества из 200 мкм светового волокна (Yizhar et al. 2011). Синий свет распространяется, максимум, 0,5 мм в ткани, красный свет около 1 мм. Цветовое кодирование: красный 50%, желтый 10%, зеленый 5%, синий 1%, если свет достигает этой области. E) Корональная точка зрения односторонней имплантации непосредственно над левым IL с оптическим волокном 200 мкм. Область IL имеет ширину 0,25 мм в каждом полушарии и глубину 0,2 мм. Синие и красные лампочки являются границы 5% свет распространяется и передаются из Yizhar и др. в нужном размере. LoxP: локус X-over P1; ChR2: Channelrhodopsin; YFP: желтый флуоресцентный белок; dflox: двойная флокса; IL: инфралибическая кора; BLA: базолатеральная миндалина; DRN: спинные ядра рафэ; PrL: прелимбийский регион. Эта цифра была изменена с Berg 201948. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
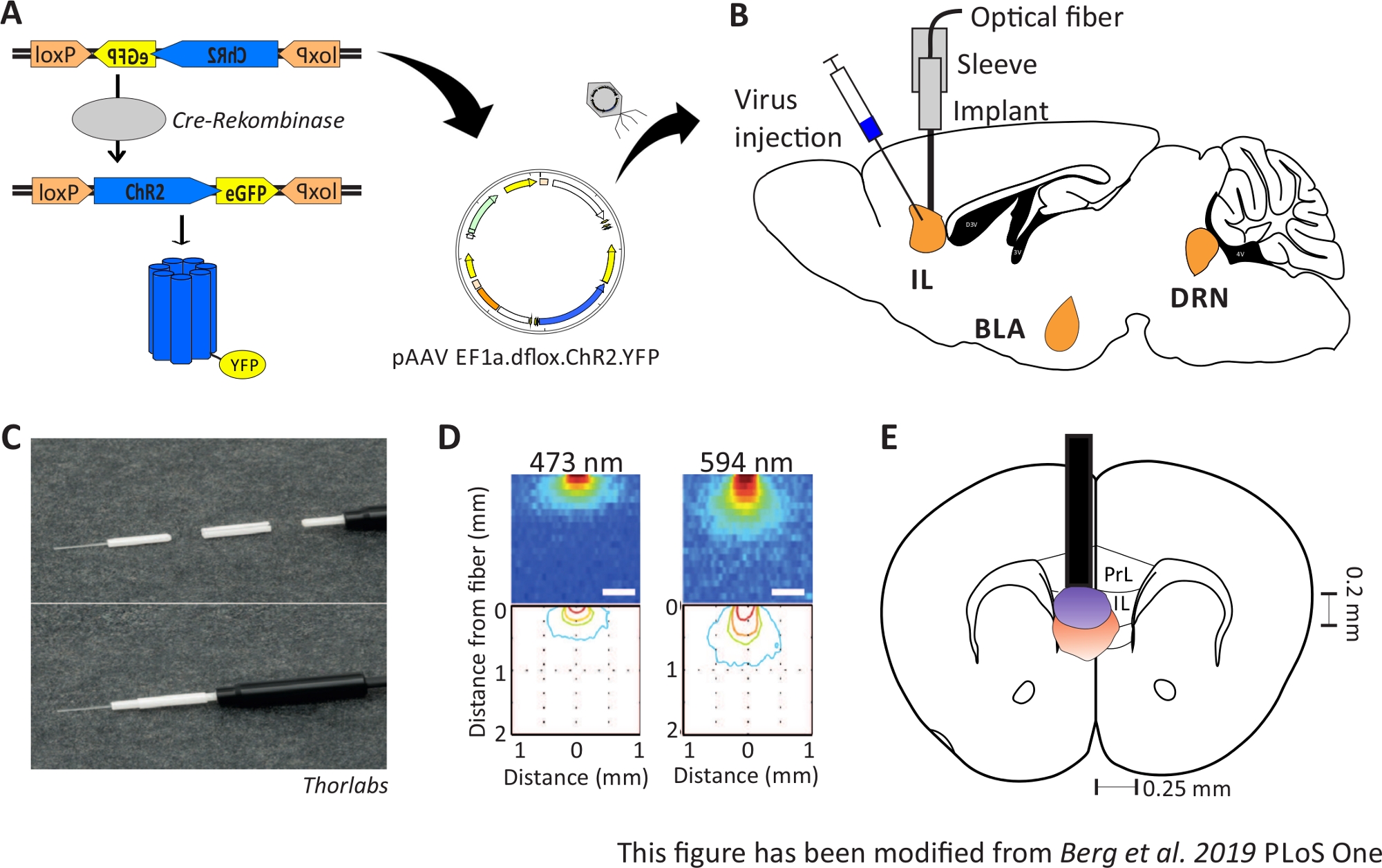{kind=link}
Используются оптогенетические подходы, позволяющие как высокое временное, так и пространственноеразрешение 14 и специфическую модуляцию типа клеток. Кроме того, можно повторно использовать имплантированное устройство без дальнейшего лечения. После стереотаксической операции, где проводится инъекция адено-ассоциированного вируса, несущего оптогенетический инструмент, и имплантация оптического волокна, мыши могут восстановиться в течение двух недель. Мы выбрали время восстановления только 2 недели, потому что это достаточно времени, чтобы оправиться от операции и для вируса, чтобы выразить. По мере того как поведенческие эксперименты после этого иммуногистохимия, мы должны обеспечить что мыши не получают слишком старыми во время эксперимента; в противном случае качество тканей снижается. Они не показывают очевидных поведенческих нарушений от имплантата и участвовать в типичном поведении клетки. Конечно, имплантация сопровождается значительным хирургическим поражением; поэтому мышей интенсивно контролируют. После операции, мыши должны быть одной размещены, как группа размещены мыши, как правило, травмируют друг друга свежие раны и имплантаты. Тем не менее, жилищные условия имеют большое влияние на уровень тревоги мышей мужского пола, как одиночные размещены мыши показывают болеенизкие уровни тревоги 15 и в целом менее депрессивные симптомы16.
Химические или электрические манипуляции мозговых схем не имеют высокой клеточной специфики оптогенетики и имеют более низкое временное и пространственноеразрешение14,17,,18. В зависимости от экспериментального вопроса, электрическая или химическая стимуляция может иметь различные преимущества. При прохождении волоконных терминалов в определенной области также необходимо стимулировать, электрическая стимуляция является лучшим методом. Химическая стимуляция является хорошим выбором, когда передатчик конкретных рецепторов в целом регионе должны быть активированы агонистов. Еще одним большим преимуществом оптогенетики по сравнению с химической или электрической стимуляции является то, что эндогенно, нейроны не чувствительны к свету, что позволяет избежать возникновения побочныхэффектов 19. Действительно, высокая итонимность света может вызватьэффекты нагрева 8,20, но из-за надлежащих контрольных групп, поведенческие эффекты из-за оптогенетических манипуляций могут быть устранены.
Исследование поведения грызунов, особенно в отношении психических заболеваний, значительно улучшилось с оптогенетики в свободно движущихся животных, так как это позволяет прямой модуляции отдельных рецепторов до конкретныхпопуляций клеток 21 исхемы 22. Возможность измерения острого воздействия таких модуляций, а также долгосрочные поведенческие эффекты после определенноговремени 23 или после хроническойстимуляции 24, обеспечивает широкую гибкость экспериментальных конструкций и обеспечивает очень подробное понимание мозговых схем. Световая стимуляция может быть использована для модуляции нейронов, расположенных в месте инъекции оптогенетического инструмента. Когда и инъекции и имплантации адрес той же области мозга, клеточных тел и спины проецирования аксонов принципа нейронов и интернейронов в этойобласти могут быть направлены 3,6,8. Тем не менее, световое волокно также может быть имплантировано в области, отличается от вводили один. В этом случае световая стимуляция может модулировать высвобождение передатчика на аксон-терминалах в проекционных зонахинъекционнойобласти 25,26,,27.
В исследовании здесь, оптогенетика используется в сочетании с экспериментами для анализа тревоги, связанных с поведением. Тревожные психические заболевания затрагивают более трети населения мира28,29,,30 ивызывают высокое экономическоебремя 31. Пострадавшие страдают от чувства возбуждения, напряжения и беспокойства, за которым следует поведениеизбегания 32,,33. Эти хронически происходящие негативные эмоции, которые в основном ориентированы набудущие события 34,сильно вмешиваются в повседневную жизнь пациентов. Общие методы лечения, такие как бензодиазепины или селективные ингибиторы обратного захвата серотонина (SSRIs) являются успешными только у некоторых пациентов. Большое количество людей не реагируют на лечение вообще35, показывая, что механизм, лежащий в основе таких заболеваний еще не до конца понял. Медиальная префронтальная кора (mPFC), как известно, играет важную роль вразвитии и проявлении тревоги 21,,25,,27,,36,,37,,38. В частности, переактивация региона инфралибной коры головного мозга (ИЛ) в MPFC может быть частью тревожных расстройств39,40. Пример эксперимента, описанного здесь, может помочь понять, как модуляции в области IL mPFC влияют на тревожное поведение. Кроме того, потенциально может быть поддержана разработка новых терапевтических стратегий в области психических заболеваний, связанных с тревогой.
2-6 месяцев самец Nex-Cre мышей используются для выражения ChR2 конкретно в пирамидальных нейронов в пределах области IL mPFC41. Nex-Cre мышей C57Bl/6 фона и выразить фермент Cre-рекомбиназы конкретно в пирамидальных нейронов. Во время стереотаксической операции двойной флоксированная chR2-DNA вводится в область IL с помощью адено связанных вирусных векторов. Оптический имплантат помещается непосредственно над областью интереса(рисунок 1B)и имплантат фиксируется зубным цементом. Контрольные животные получают инъекцию двойного флокса tdTomato-ДНК в той же области, чтобы имитировать клеточное выражение.
До дня операции животные размещаются в группе, а затем размещаются в одиночных домах, чтобы избежать травм от других мышей. Мыши размещаются в отдельных вентилируемых клетках (IVC) стеллажи в TypI-L клетки для одиноких мышей. Светло-темный цикл следует ритму 12:12 ч, фаза света начинается в 10 утра. Все поведенческие эксперименты проводятся в темной фазе, напоминающей активную фазу грызунов. Вода и стандартные пищевые гранулы доступны ad libitum. После двух недель восстановления, которое обеспечивает достаточное выражение ChR2 в пирамидальных нейронов, мыши используются для поведенческих экспериментов.
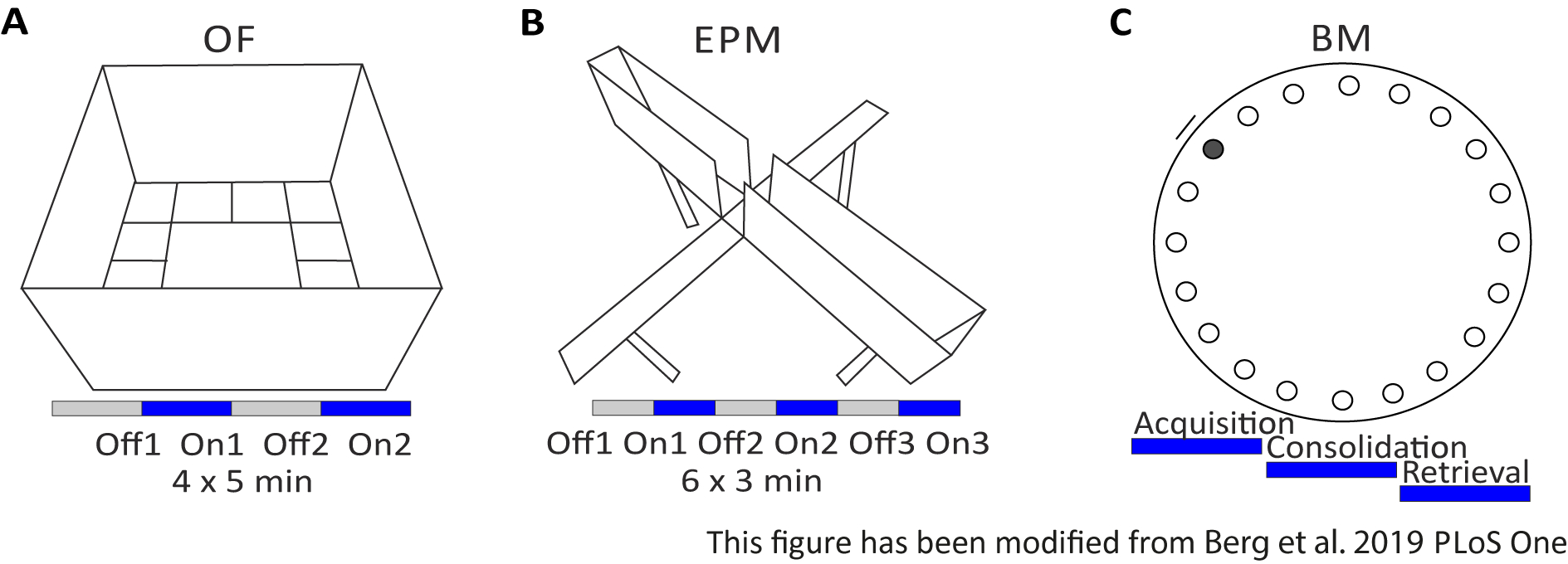
Открытое поле (OF) является 50 см х 50 см в квадрате лабиринт с пескоструйной 40 см высокие стены. Земля разделена на 16 квадратов, где внутренняя 4 представляют центр. Измеренное поведение: 1) время, проведенное в центре, 2) количество входов в центр и 3) общее расстояние перемещено. В ходе этого эксперимента, Есть 4 испытания на общую сумму 20 минут. В испытаниях 1 и 3, не происходит стимуляции света, и в испытаниях 2 и 4, стимуляция 20 Гц с 5 мс светового импульса и 1 мВт интенсивность света 473 нм выполняется (Рисунок 2A). В более поздних испытаниях, привыкание к испытательному полигону было принято во внимание, но использование фиктивных инъекционных контрольных животных показывает, как выражается привыкание.
Лабиринт Барнса - это эксперимент по обучению и памяти. Представляет собой круглую платформу диаметром 92 см и содержит 20 равноудающих отверстий вокруг окружности. 19 отверстий закрыты и под одним отверстием представлена коробка для побега. В течение 4 дней подряд, мыши имеют 4 учебных испытаний, чтобы узнать расположение коробки побега. На5-й день, побег окно удаляется, и мыши проверяются на сколько времени им нужно, чтобы найти правильное отверстие. Измеренное поведение: 1) Время до тех пор, пока окно побега/правильное отверстие не будет найдено, 2) Количество целевых посещений и ошибок, и 3) Расстояние, перемещенное до тех пор, пока в поле побега. Свет стимуляции в различных группах делается либо во время приобретения или консолидации, которые происходят в учебные дни 1-4, или во время поиска в день тестирования, который день 5 (Рисунок 2D).

Рисунок 2: Поведенческие эксперименты с оптогенетическими протоколами. A)Схематический рисунок эксперимента «Открытое поле» с соответствующим протоколом стимуляции света. C)Схематический рисунок эксперимента Elevated-Plus Maze с соответствующим протоколом стимуляции света. D)Схематический рисунок эксперимента Barnes Maze с соответствующим протоколом стимуляции света. EPM: Повышенный плюс лабиринт; OF: Открытое поле; БМ: Тест Барнса в лабиринте. Эта цифра была изменена с Berg 201948. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}
Для оптогенетической стимуляции интенсивность и частота света должны быть адаптированы к оптогенетического инструмента и нейронального типа, который находится под следствием. Самая низкая возможная интенсивность света должна быть использована для того, чтобы избежать повреждения тканей, так как несколько исследований показали, что есть возможные эффекты нагрева из-засильной интенсивности света 8,,20. Для ChR2, 20 Гц стимуляции с 5 мс световой импульс обычно используется2. Поскольку ChR2 достаточно светочувствительный, достаточно интенсивности света 1 мВт. Протокол стимуляции света чередуется между выключением света и испытаниями для непосредственного измерения поведенческих изменений. Внешние комнатные условия для поведенческих экспериментов должны оставаться стабильными для всей группы животных. Важными условиями для рассмотрения являются шум (имейте в виду, что сами устройства могут шуметь), запах (всегда очистить поведенческие установки с этанолом), интенсивность света, и экспериментатор. Экспериментатор всегда должен быть один и тот же человек. Кроме того, время суток экспериментов должно быть одинаковым для всех животных в одной группе, через несколько часов после начала темной фазы на объекте предпочтительнее.
Целью этого эксперимента является увеличение коэффициента возбуждения/ингибирования (E/I) в области IL за счет сильной активации возбуждающей пирамидальной нейроны. Повышенное соотношение E / I в этой специальной области коры головного мозга, как известно, увеличение уровнятревоги у мышей 40,42,43,44.
протокол
Процедуры с участием животных были одобрены институциональным научно-исследовательским центром для животных и "Сенаторин фюр Виссеншафт, Gesundheit und Verbraucherschutz" в Университете Бремена (#146)
1. Подготовка оптического имплантата9 (Рисунок 1C)
- Поместите керамическую феррульную плоскую сторону вверх в тись на скамейке.
- Полоса пальто 200 мкм диаметром стеклянное волокно с волокном зачистки инструмент и сократить 2-3 см в длину штук с керамическим писцом волокна.
- Поместите кусок стеклянного волокна в керамический феррул с ровный свес с обеих сторон.
- Поместите каплю суперклея на плоской стороне керамического феррула с инъекцией канулы.
ПРИМЕЧАНИЕ: Протокол можно приостановить здесь. - Возьмите предварительный имплантат из тиза скамейки и на круглой стороне керамической феррулы, вырезать стекло волокна как можно короче с керамическим писцом волокна.
- Поместите предимплантуатуру на шайбу для полировки феррула и отполировать круглую сторону на 4 различных полировальных бумагах, нарисовав восемь 20 раз на бумагу (30 мкм песка, 6 мкм песка, 1 мкм песка, и, наконец, 0,02 мкм песка).
- Возьмите предимплантный из феррулы полировки шайбу и вырезать стеклянное волокно на плоской стороне керамического феррула до длины, необходимой для имплантации. Начните измерять длину за выступающим суперклеем.
- Для равномерной резки поверхности просто поцарапать стекло волокна 2-3 раза, а затем разорвать его.
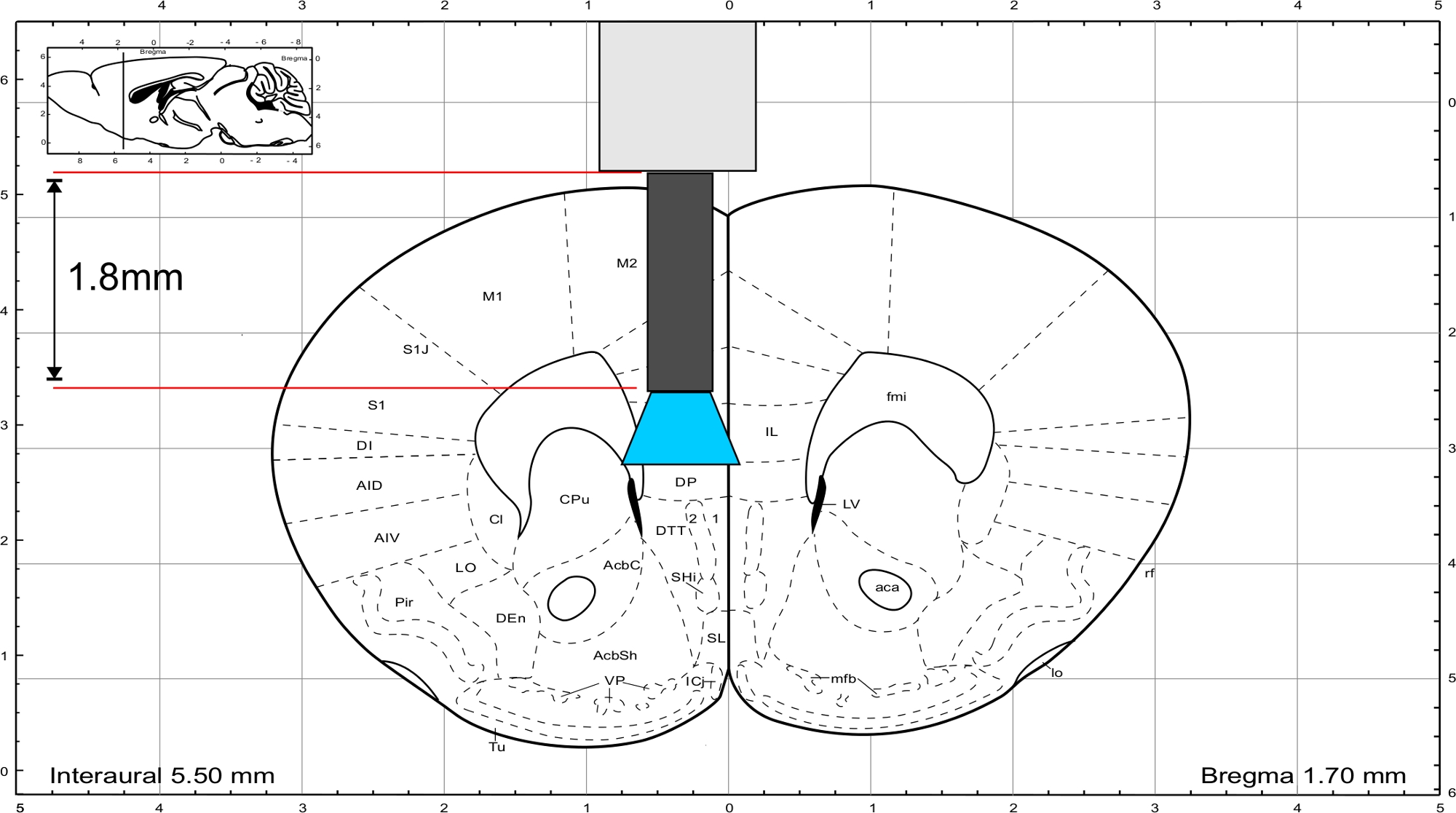
ПРИМЕЧАНИЕ: Используйте атлас мозга мыши от Paxinos и Франклин45 для того чтобы высчитать длину имплантата. Имплантат должен закончиться непосредственно над областью интереса и толщина черепа должна быть включена в расчет длины. Чтобы стимулировать область IL, стекловолокно имеет длину 1,8 мм(рисунок 3).
- Для равномерной резки поверхности просто поцарапать стекло волокна 2-3 раза, а затем разорвать его.

Рисунок 3: Атлас мозга мыши (Paxinos и Франклин) с репрезентативной длиной имплантата для достижения области IL. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}
- Дезинфицировать готовый имплантат в течение 10 минут в этаноле и дайте ему высохнуть перед имплантацией.
2. Инъекция и имплантация
- Транспорт одной мыши в хирургическую комнату и взвесить его. Нанесите анестезию с внутриперитонеальной (i.p.) инъекцией кетамина/ксилазина (кетамин 0,12 мг/г, ксилазин 0,01 мг/г).
- Исправить мышь левой рукой и повернуть его на спину с головой низко.
- Нацелиться на левый нижний квадрант живота шприцем и ввести инъекцию канулы 1 см под кожу.
- Вводите анестезию в медленном и постоянном движении в брюшную полость.
- Поместите мышь обратно в клетку и ждать, пока она достигнет глубокого состояния анестезии.
ПРИМЕЧАНИЕ: Глубина анестезии может быть определена отсутствием мигающих и между ног рефлексов.
- Поместите мышь на нагревательной пластине и зафиксните голову в стереотаксической раме. Зафиксить нос и зубы спереди, и уши с обеих сторон.
ПРИМЕЧАНИЕ: Голова должна быть прямо на лево-правой и рострал-каудальной оси, чтобы обеспечить правильные стереотаксических координат. - Нанесите анальгезию с 2 мг/кг карпрофена подкожно в задней части мыши и применить непрозрачные мази глаза на обоих глазах, чтобы защитить их от высыхания.
- Смочить волосы на кожу головы с мокрым бумажным полотенцем, а затем отрезать его с помощью ножниц. Убедитесь в том, чтобы удалить все свободные волосы с мокрым бумажным полотенцем. Для дезинфекции кожи головы используйте ватную палочку и возьмите 0,5 мл настойки, содержащей йод (Betaisodona 100 мг/мл Povidon йода и 11 мг/мл йода) и дайте ему высохнуть.
ПРИМЕЧАНИЕ: Вместо ножниц, также электрический клипер может быть использован для надлежащего удаления волос. - Поднимите кожу головы над областью интереса с помощью пинцета и вырезать 1 см вдоль средней линии. Используйте два пинцета, чтобы отодвинуть кожу в сторону, чтобы разоблачить череп. Убедитесь в том, чтобы также удалить тонкую кожу над черепом и пусть подвергаются черепа высохнуть.
- Грубить череп для более поздней имплантации.
- Нанесите 2 мм х 2 мм капли фосфорной кислоты (37%) из клея комплекта (например, Optibond) на черепе, распределить его кончиком шприца и дайте ему в силу в течение 15 с.
- Удалить все кислоты с ватной палочкой и промыть череп с 1 мл 0,9% NaCl в два раза.
- Высушите череп хлопчатобумажной палочкой и сжатым воздухом.
Внимание: Фосфорная кислота опасна и должна быть полностью удалена, чтобы избежать повреждения тканей.
- Рассчитайте F-фактор для отдельных координат.
- Поместите стеклянную канулу в стереотаксическую раму и найдите ее прямо над брегмой.
- Нулевой системы координат и переместить стеклянную канулу в lambda.
- Рассчитайте F-фактор46 со следующей формулой:

- Умножьте F-фактор с координатами из атласа мозга мыши, чтобы настроить их на индивидуальную мышь.
- Просверлите отверстие в черепе для инъекций.
- Используйте скорректированные координаты, чтобы найти расположение на черепе непосредственно над структурой интереса и пометить его с помощью кончика инъекционной канулы, поцарапав его над поверхностью кости.
- Используйте инъекционную канулу, чтобы просверлить отверстие в черепе в отмеченной локации, вращая канулу на месте. Если кровь вытекает из заусенцев отверстие, промыть 1 мл 0,9% NaCl и высушить череп потом.
- Возьмите раствор вируса в стеклянную канулу.
- Поместите каплю 100 л 0,9% NaCl на череп и кусок парафильма (1 см х 1 см) сверху, стерильную сторону вверх.
- Поместите 1-2 Л вирусного раствора на парафильм и опустите кончик стеклянной канулы в него.
- Подключите стеклянную канулу к шприцу, нанесите минимальное отрицательное давление и подождите, пока раствор вируса не будет взят канюлей (в течение нескольких секунд).
ПРИМЕЧАНИЕ: Важно, чтобы остановить применение отрицательного давления, прежде чем воздух взял в канулу. Таким образом, всегда будет небольшой остаток вирусного раствора.
- Ввимить вирусное решение в область, представляющий интерес.
- Поместите вирус заполнены стеклянные канулы над заусенцев отверстие.
- Медленно опустите канулу в заусенцевое отверстие и обнуляйте z-координацию, когда кончик канулы находится на уровне черепа.
- Тщательно опустите канулу в самое низкое положение места инъекции.
- Сосредоточьте бинокль на мениска раствора вируса в канюле.
- Нанесите небольшое количество положительного давления со шприцем, пока мениск не будет снижен незначительно.
- Пусть вирус распространяется в течение 2-3 минут, прежде чем двигать стеклянную канулу вверх в следующее положение.
- Применяем вирусное решение каждые 200-300 мкм по всему региону, представляющим интерес.
- Удалите стеклянную канулу очень медленно и отбросьте ее после окончательной инъекции.
- Подготовь череп к имплантации с помощью адгезионерного комплекта (например, OptibondTMFL).
- Высушите череп сжатым воздухом.
- Нанесите 5 Л грунтовки (например, Оптибонд, 1-30% (этанол, силиховая кислота, глицеринфосфатиметакрилат, 2-(2-(метакрилоилокси)этиоксикарбонил),бензоэзаур, 2-Гидроксиэтилметхакрилат)) с соответствующей палкой и дайте ей высохнуть.
- Применить 5 МКЛ связи (например, Optibond, 15-20% 2-Гидроксиэтилметакрилат 1-2% Alkalihexafluorosilikat (Na)) с той же палкой и вылечить его на 20 с ультрафиолетовым светом (420-480 нм).
ПРИМЕЧАНИЕ: Важно, чтобы череп был сухим и чтобы грунтовка и связь применялись в очень тонком слое.
Внимание: Не смотрите непосредственно в ультрафиолетовый свет, так как ультрафиолетовый свет может нанести вред глазам.
- Распоить имплантат непосредственно над областью интереса.
- Зафиксировать имплантат в соответствующем держателе.
- Высушите череп сжатым воздухом.
- Распоить кончик стеклянного волокна прямо над отверстием заусенцев и осторожно опустить его.
- Прекратите опускать имплантат, когда оставшаяся лампочка суперклея коснется черепа. Не давите на череп!
ПРИМЕЧАНИЕ: Если инъекции и имплантации делается в различных регионах (например, спинной раф игиппокамп ), просверлить все необходимые отверстия после применения фосфорной кислоты, но до 2-компонентной адгезии, а затем следовать инструкциям, как описано ранее (шаг 2,8-2,14).
- Исправьте имплантат.
- Проверьте, полностью ли высох череп.
- Нанесите жидкий зубной цемент (например, Gradia direct flo) вокруг имплантата и в окрестностях и излечив на 20 с ультрафиолетовым светом (420-480 нм).
ПРИМЕЧАНИЕ: Количество зубного цемента зависит от свободной области черепа. Весь череп должен быть покрыт зубным цементом. - Нанесите еще два слоя цемента и полностью заполните свободную и высушенную область черепа. Лечить каждый слой ультрафиолетовым светом (420-480 нм).
- Закончите операцию.
- Нанесите на всю рану 0,5 г йодной мази (бетаисодона 100 мг/мл йода и 11 мг/мл йода).
- Введение 0,1 мл глюкозы растворяется в 0,9% NaCl подкожно в шею для быстрого восстановления.
- Отпустите фиксацию носа и уха, принесите мышь в свежую клетку и поместите ее под нагревательной лампой, чтобы избежать потери тепла тела.
- Когда мышь проснется, верните ее в помещение.
- Проверяйте состояние его здоровья не реже одного раза в день. Примите соответствующие меры, если мыши отображают какие-либо плохие конституции (например, обеспечить послеоперационную анальгезию с карпрофеном до 3 дней, если мыши проявляют какие-либо признаки боли).
ПРИМЕЧАНИЕ: После двух недель восстановления, мыши могут быть использованы для поведенческих экспериментов.
3. Создание нового эксперимента (Пример стимуляции ChR2 и открытого поля)
- Pulser
- Программа импульсного импульса (например, Prizmatix) для стимуляции света.
- Откройте программное обеспечение и выберите порт USB COM, в который подключен источник света.
- Выберите режим работы Select (3) Выполните последовательности импульсов после запуска HIGH, а затем остановить, когда LOW, чтобы внешнее программное обеспечение для управления источником света.
- Запрограммировать световой протокол. Для стимуляции 20 Гц с световым импульсом 5 мс: выберите TI 23 мс, P1D и 5 мс, P1I 22 мс и P2D 0 мс.
- Последовательность запуска прессы. Этот статус будет сохраняться до завершения экспериментов.
ПРИМЕЧАНИЕ: Программное обеспечение импульсного импульса (Prizmatix Pulser) должно быть запущено до программного обеспечения для отслеживания видео; в противном случае программное обеспечение для отслеживания видео не сможет распознать устройство.
- Программное обеспечение для отслеживания видео (например, Ethovision XT)
- Создайте новый эксперимент из заранее определенного шаблона.
- Откройте программное обеспечение, перейдите в файл, выбрать новый из шаблона. Выберите Применить заранее определенный шаблон.
- Выберите отслеживание в реальном времени и выберите камеру, нажав на Источник и подтвердите подключенный Basler GenICam.
ПРИМЕЧАНИЕ: Живое изображение камеры теперь будет отображаться в окне в правом верхнем. - Нажмите Далее и выберите животное, которое должно быть записано( Грызуны, Мышь).
- Нажмите Далее и выберите шаблон арены Открытое поле, квадрат. Выберите шаблон зоны Центр, Граница, Углы и подтвердите с Next.
- Подтвердите 1 тему, которую следует отслеживать с помощью Next.
- Выберите центр-точка, нос-точка и хвост-база и подтвердить цвет животного по сравнению с фоном, как темнее с Next.
- Подтвердите рекомендуемую выборку 12,5 с Next и закончите шаг.
- Назовите эксперимент подходящим и выберите место для сохранения.
- Определите экспериментальные настройки.
- Перейти к настройке и экспериментальным настройкам. Выберите центральную точку, точку носа и обнаружение хвостовой базы в качестве отслеживаемых функций.
- Выберите использование оборудования пробного управления и перейдите в настройки.
- Выберите коробку Noldus USB-IO и подтвердите с Ok.
- Выберите пользовательское оборудование в качестве типа устройства в порту TTL, который был подключен к устройству импульсивного устройства, и подтвердите с Ok.
- Определите настройки арены.
- Перейти к Арена Настройки и выберите Арена Настройки 1.
ПРИМЕЧАНИЕ: Теперь камера автоматически откроет фоновое изображение. - Подтвердите изображение с Grab.
- Адаптировать заранее определенные зоны к реальной арене, изменив их размер. Используйте стрелку и два символа справа от нее. Если некоторые зоны не нужны, удалите их.
- Нажмите 1. Нарисуйте шкалу, чтобы откалибровать и вытащить линию из одного угла лабиринта в другой. Введите длину реального расстояния в см.
- Повторите это для другой оси.
- Перейти к Арена Настройки и выберите Арена Настройки 1.
- Проверьте, работает ли световая стимуляция.
- Перейти к Арене - Оборудование Картирование и выбрать тест на серой бар.
- Выберите командный выход 1 High и нажмите тест.
ПРИМЕЧАНИЕ: Там должен быть свет, излучающий из конца оптического волокна. При выборе выход 1 Низкий и тест, стимуляция должна прекратиться.
- Определите параметры пробного управления в течение 20 минут эксперимента. Установите испытания Off1, On1, Off2 и On2, чтобы каждый из них был 5 минут.
- Перейдите на настройку пробного контроля и выберите продолжительность трека 30 минут.
- Подготовьтесь к основному правилу, регулируя условие: время до 20 минут, выбрав настройки и измените от 30 до 20 минут. Подтвердите с Ok.
ПРИМЕЧАНИЕ: Условие для начала трека должно быть, когда объект находится на арене в течение 2 секунд. Таким образом, система автоматически начнет отслеживать, когда мышь находится на арене. - Создайте под-правило для стимуляции света: Перейти к структурам, больше и выберите Под-правило.
- Дайте ему имя, такое как протокол стимуляции света.
- Поместите его ниже основного правила и распределить две коробки, выбрав синюю область с мышью courser.
- Перейти к условиям, время и дать ему имя, как свет на 1.
- Состояние регулировки выполнено через 5 минут. Подтвердите с Ok.
- Поместите коробку прямо за окном Правило Начало подуряда, потянув его к черной линии.
- Перейти к действию (ru) Пользовательское оборудование и назовите его: свет на 1.
- Выберите действие для выполнения в качестве выхода 1 High и подтвердите с Ok.
- Поместите коробку прямо за коробку Условия.
ПРИМЕЧАНИЕ: Теперь, после 5 минут эксперимента, должна начаться световая стимуляция. - Повторите шаги, чтобы определить состояние времени После 5 минут и действия Выход 1 Низкий, чтобы остановить стимуляцию света еще через 5 минут.
- Повторите шаги снова, чтобы запрограммировать другой свет Off и свет на суде.
- Перейти к структурам Подправие ссылки и проверить, что ссылка принадлежит к правильному подправе.
- Выберите условия запуска, как без задержек и стоп-условия, как выполнить один раз в состоянии запуска. Подтвердите с Ok.
- Поместите справочную коробку между экшн-боксом 1 и коробкой условий 2 основного правила и нарисуйте линию от Action - начните отслеживать ссылку.
ПРИМЕЧАНИЕ: Теперь основное правило непосредственно запускает подуряд после запуска трека.
- Определите параметры обнаружения, чтобы показать системе, что она должна отслеживать.
- Перейдите к настройкам обнаружения и выберите настройки обнаружения 1.
- Поместите тестовую мышь на арену и выберите автоматизированную настройку.
- Выберите грызунов в качестве животного типа и использовать курсер мыши, чтобы нарисовать коробку вокруг мыши на арене. Подтвердите результаты OK? вопрос с Да.
- Определите список испытаний для всех экспериментальных животных, которые должны быть отслеированы.
- Перейти к Пробный список и планировать все животные для записи сегодня: Выберите добавить испытаний и выбрать номер.
- Выберите все условия, определенные ранее для каждой мыши.
- Назовите Animal-ID и лечение правильно, чтобы позже упростить анализ.
ПРИМЕЧАНИЕ: Animal-ID не имеет значения для системы и имеет важное значение только для более поздних анализов данных экспериментатором. Группа по лечению и контролю важна для системы, чтобы знать, как группы и как сравнить все треки в более позднем анализе шагов.
- Перейти к приобретению и начать с эксперимента.
- Создайте новый эксперимент из заранее определенного шаблона.
4. Открытый полевой эксперимент (тревога)
- Принесите экспериментальную мышь в поведенческую комнату прямо перед экспериментом, чтобы обеспечить надлежащий уровень тревоги.
ПРИМЕЧАНИЕ: Поведенческие эксперименты должны быть выполнены во время темной фазы, когда мыши бодрствуют, и всегда в тот же временной интервал для обеспечения сопоставимости. - Пара мыши через рукав к источнику света, нажав его осторожно на сетку клетки.
- Поместите его в клетку ожидания со свежим мусором в течение 10 минут, чтобы акклиматизироваться к световому кабелю.
- Начните приобретение, нажав кнопку «Пуск» в программном обеспечении для отслеживания видео (например, Ethovision XT).
- Перенесите мышь из клетки ожидания в левый верхний угол открытого поля. Удалите руку в течение 2 секунд, чтобы избежать отслеживания руку вместо мыши.
- Оставьте поле зрения мыши во время эксперимента и сохраняйте спокойствие.
- Через 20 минут, когда эксперимент будет закончен, удалите мышь из лабиринта, отключите световой кабель и поместите его обратно в домашнюю клетку.
- Верните мышь обратно в помещение.
5. Барнс лабиринт (обучение)
- Принесите всех экспериментальных мышей в поведенческую комнату примерно за 1 час до эксперимента.
- Подготовьтесь к лабиринту Барнса, закрыв все отверстия, кроме одного, под которым помещается коробка для побега. Поместите стену из коробки в середине платформы, которая является отправной точкой для мыши.
- Подключите одну мышь к источнику света (рукав на световом кабеле) на обоих имплантатах.
- Поместите мышь прямо в середину лабиринта Барнса в стену коробки, которая предотвращает мышь от бегать до начала эксперимента.
- Нажмите Начните с программного обеспечения для отслеживания видео (например, Ethovision XT) и удалите коробки.
ПРИМЕЧАНИЕ: Программное обеспечение отслеживает мышь до тех пор, пока правильное отверстие не будет достигнуто, но будьте готовы остановить испытание вручную только в случае, если программное обеспечение не распознает переход отверстия. - Вынюхи мышь из лабиринта и удалите соединение с световым кабелем.
ПРИМЕЧАНИЕ: Если это тренировочный день с несколькими испытаниями на мышь, оставьте мышь в зале ожидания рядом с поведенческой комнате до начала следующей тренировки. Если это был день тестирования только с одной пробной версией тестирования на мышь, принесите мышь обратно на объект.
6. Анализ данных (пример открытых полевых данных с 4 различимыми испытаниями)
- Программное обеспечение для отслеживания видео (например, Ethovision XT)
- Определите экспериментальные группы и исследования в профиле данных.
- Перейдите к профилям данных слева и выберите Treated vs. Control.
- Перейдите в Nesting в новом окне в левом середине и выберите состояние пробного управления.
- Выберите интервал состояния из действия элемента: начните отслеживать действие элемента: свет идет на 1.
- Поместите коробку Nesting между обработкой коробки фильтра и соответствующей коробкой результатов.
ПРИМЕЧАНИЕ: Этот определенный интервал Off1, который описывает первые 2,5 минуты эксперимента, где нет стимуляции света присутствует. - Повторите шаги для интервалов On1 (от элемента Действий: свет переходит на 1 к элементу Действие: свет выходит 1), Off2 (от элемента Действий: свет выходит 1 к элементу свет идет на 2) и On2 (от элемента Действий: свет идет на 2 к элементу Действий: стоп-трек).
- Повторите 4 интервала для группы фильтра управления.
ПРИМЕЧАНИЕ: Каждая коробка гнездования нуждается в своей собственной коробке результатов с именами Off1, On1, Off2, On2. Теперь и группа обработки и управления разделена в 4 по-разному испытания стимуляции света которые проанализированы отдельно.
- Определите параметры для анализа в профиле анализа.
- Перейти к анализу профилей слева и выберите В зонах.
- Выберите зависимую переменную в зоне и выберите центр в качестве зоны.
- Дважды нажмите на In center и выберите любую из выбранных точек и выберите только в центре.
- Перед выходом из окна перейдите на пробную статистику и выберите частоту, совокупную продолжительность и задержку к первому.
- Добавьте перемещенное зависимое переменное расстояние.
ПРИМЕЧАНИЕ: В групповой статистике,выбрать, следует ли использовать стандартную ошибку или стандартное отклонение в качестве ошибки. С помощью этого профиля доступны данные о времени, проведенном в центре, записи Центра и Общее расстояние.
- Извлекайте данные
- Перейти к результатам и выбрать статистика и диаграммы.
- Нажмите Рассчитайте, чтобы увидеть проанализированные данные.
ПРИМЕЧАНИЕ: Судебная статистика дает информацию о каждой мыши и группы статистика анализирует среднее и ошибка для обеих групп, разделенных на 4 испытания с соответствующим графиком бар. - Нажмите данные экспорта и выберите статистику проб и местоположение для сохранения.
ПРИМЕЧАНИЕ: Экспортируемые данные сохраняются как файл Excel и с индивидуальными значениями для каждой мыши. В этом файле Excel Animal-ID помогает идентифицировать мышей. - Перейти к визуализации Heatmap и нажмите Участок Heatmaps.
- Выберите испытания справа, чтобы увидеть отдельные тепловые карты для каждой мыши и суда.
- Сделайте правильный щелчок на мышь и экспортировать тепловые карты в качестве изображений.
- Определите экспериментальные группы и исследования в профиле данных.
- Печати
- Откройте файл электронной таблицы на компьютере и вычислите средства и стандартные ошибки (SEM) для всех 4 испытаний в каждом измеренном состоянии и группе.
- Создание графиков в статистической программе (например, Sigma Plot).
- Копировать средства и SEM в правильный порядок из файла электронной таблицы в Sigma Участок. Строки должны содержать данные для Off1, On1 и т.д., а столбцы содержат пробную версию, среднее и SEM в качестве головок.
- Выберите все три столбца и перейдите к созданию графика.
- Выберите поле бара и выберите негрупповые бары с ошибкой (верхний ряд, третья коробка).
- Подтвердите с отделкой, чтобы открыть новую страницу графика.
- Этикетка весь график, а затем перейти к дому, выберите граф поле слева и нажмите Экспорт. Выберите папку назначения и выберите MetaFile (К.wmf) в качестве формата.
ПРИМЕЧАНИЕ: Формат .wmf может быть обработан позже в графическом программном обеспечении, как CorelDraw.
- Рассчитайте статистику полученных данных.
- Копирование необработанных данных из электронной таблицы (Off1, On1 и т.д.) в отдельные столбцы Sigma Plot.
- Отметите столбцы, чтобы сравнить и перейти к анализу, выбрать t-тест и нажмите Run.
- Подтвердите формат данных Raw with Next и запустите тест с Finish.
Результаты
Целью данного протокола является измерение изменений в поведении генетически модифицированных мышей во время оптогенетического эксперимента. Оптогенетические манипуляции делается путем инъекций адено связанных вирусного вектора. Световая стимуляция у свободно движущихся мышей возможна путем имплантации легкого волокна непосредственно над областью интереса.
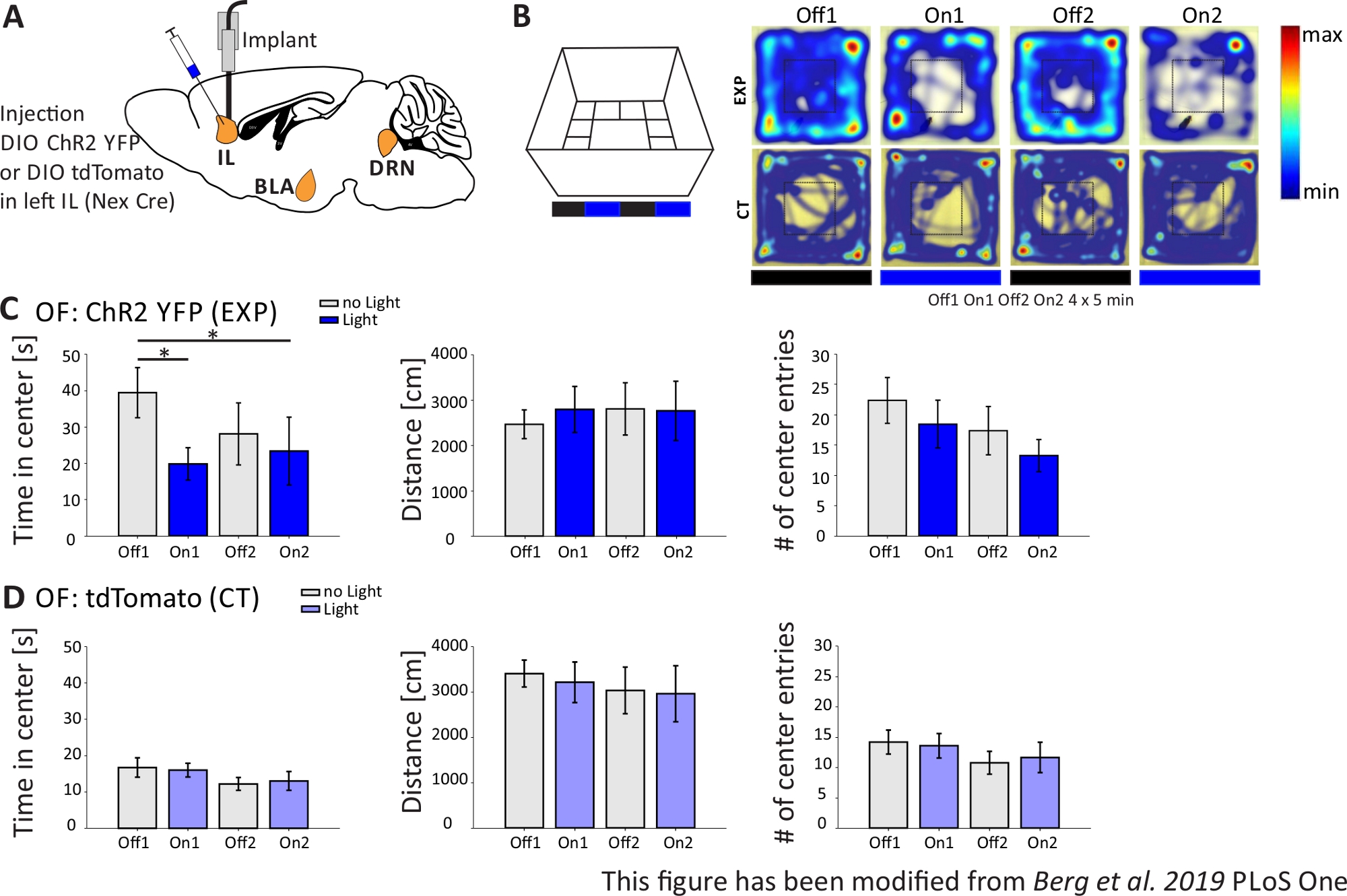
На рисунке 4представлены результаты оптогенетического эксперимента. Сильная активация возбуждающей пирамидальных нейронов в области IL через ChR2 повышенное тревожное поведение в Открытом поле. ChR2 был введен в области IL mPFC в Nex-Cre мышей для выражения в пирамидальных нейронов (Рисунок 4A). Во время двух тестов тревоги, Открытоеполе (рисунок 4B,C) и Новизна-Подавленный тест кормления(рисунок 4F,G), ChR2 стимулируется с синим светом и активирует пирамидальные нейроны. В качестве контроля другая группа мышей получила инъекцию фторфора tdTomato вместо ChR2(рисунок 4D,G). В таком эксперименте тревога определяется как избегание более яркой центральной области. Мыши показывают внутреннее избегание открытых площадок, потому что они беспокоятся о хищниках.
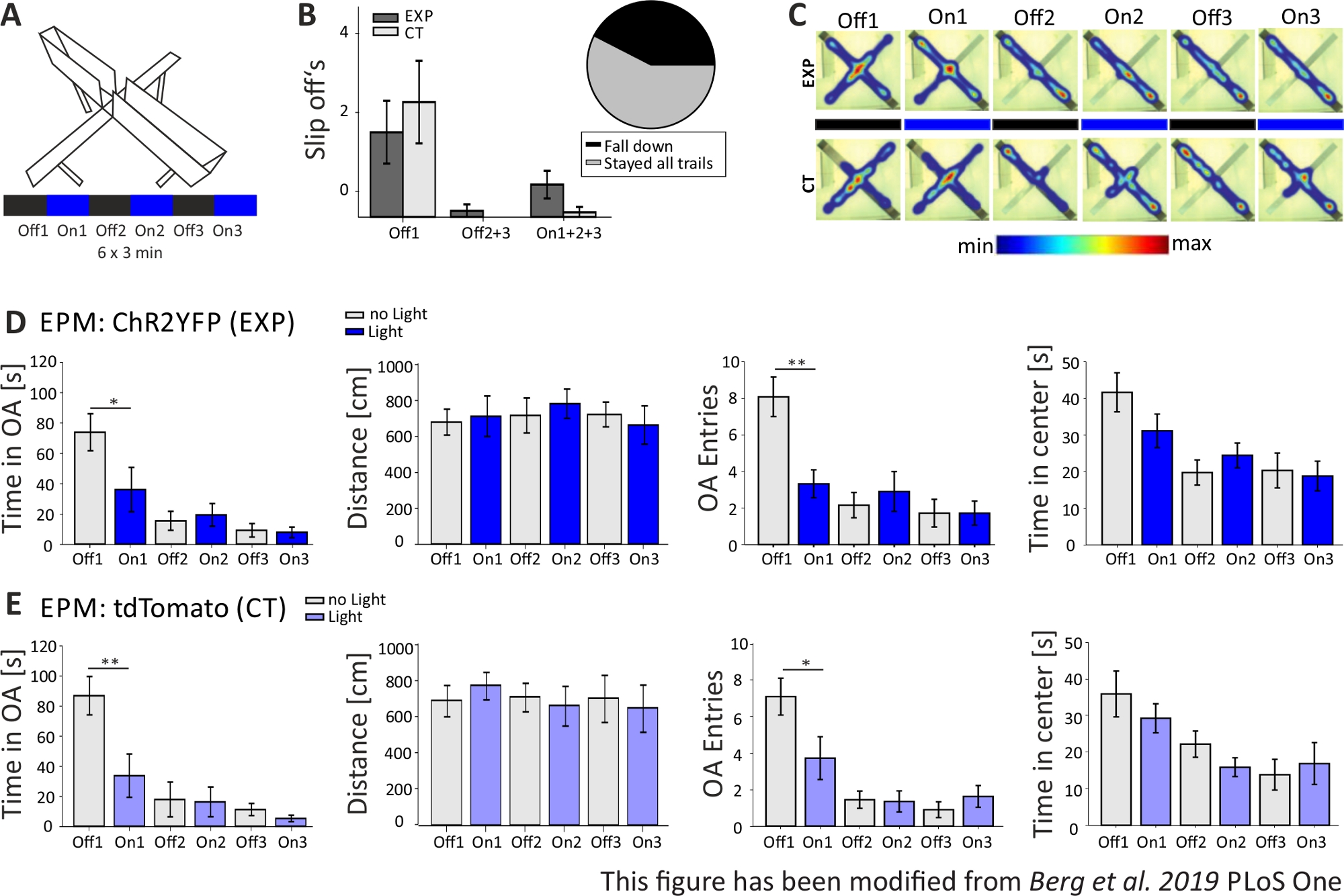
В эксперименте Open Field, показанный на рисунке 4B,мыши выполнили 4 испытания по 5 минут каждый. В испытаниях 1 и 3 не происходило стимуляции света (Off1,2), а в испытаниях 2 и 4 была проведена стимуляция синего света с частотой 20 Гц (5 мс светового импульса) и интенсивностью 1 мВт (On1,2). Тепловые карты показывают, что в экспериментальной группе продолжительность центра отличалась между испытаниями Off и On. Во время стимуляции света мыши преимущественно остаются в пограничной зоне. Контроль животных также предпочитают границы, но не меняют свое поведение на свет стимуляции. На рисунке 4Cдля экспериментальной группы показаны основные поведенческие измерения во время эксперимента «Открытое поле». Если данные прошли тест Шапиро-Вилка на нормальность, статистика была сделана с помощью независимого двуххвостого т-теста. Если тест на нормальность провалился, тест Манн-Уитни-Ранк Сум использовался в качестве немарметрической альтернативы. Для такого рода экспериментов, в группе сравнения был выбран для расследования, если свет стимуляции может непосредственно изменить поведение тревоги с течением времени, независимо от базовой тревоги экспериментальных и контроля животных. Продолжительность центра значительно уменьшилась во время обоих испытаний стимуляции света, что указывает на повышенный уровень тревоги. Общее пройденное расстояние не было изменено, что показывает, что поведение локомотива не было затронуто. Количество входов в центр было увеличено, хотя и не значительно. На рисунке 4Dотображаются данные контрольной группы. Контроль животных не отображает каких-либо поведенческих изменений между Off и On испытаний в любом из проанализированных параметров, показывая, что стимуляция света или имплантации не вызывает наблюдаемых эффектов. В целом, этот тест показывает повышенную тревожность во время легкой стимуляции ИЛ пирамидальных нейронов через ChR2.
На рисунке 5показаны данные неудачного оптогенетического эксперимента для лабиринта Elevated-Plus. Во время эксперимента Elevated-Plus Maze, который представлен на рисунке 5A,мыши завершили 6 испытаний по 3 минуты каждый. В испытаниях 1, 3 и 5 не проводилась стимуляция света (Off1, Off2, Off3), а в испытаниях 2, 4 и 6 была проведена стимуляция синего света с частотой 20 Гц (5 мс светового импульса) и интенсивностью 1 мВт (On1, On2, On3). В этих примерных результатах длина оптогенетического протокола и конструкция самого лабиринта не подходили для трансгенной линии мыши. На рисунке 5Bвидно, что несколько мышей соскользнули с лабиринта с задних лап или даже упали. Когда это произошло, мыши получили второй шанс выполнить EPM один день спустя. Если они упали снова, они были исключены из анализа. Когда мыши соскальзывания несколько раз, но удалось остаться в лабиринте, данные были проанализированы нормально. Тем не менее, данные должны быть интерпретированы очень тщательно и контроля животных получить большее значение. У мышей Nex-Cre были моторные трудности, чтобы остаться на узких сеяных объятиях. Чтобы избежать этого, маленькие стены, высотой 1 см, помогли бы для надежного удержания задних лап на руках лабиринта. И тепловые карты, и графики показывают, что экспериментальные, а также контроль мышей, начали избегать сеяных рук от суда 2 (On1) на (Рисунок 5C-E). Время на сеяных объятиях значительно сократилось для обеих групп, как и открытые руки записей. Анализируя экспериментальную группу, получены только данные, подразумевающие большой аксиогенный эффект стимуляции света, так как время на открытой руке и открытых руках значительно снижается во время испытания On1. Однако при сравнении этих данных с контрольной группой, которые показывают одинаковое поведение, становится ясно, что наблюдаемое поведение опосредовано не оптогенетической стимуляцией, а избеганием рас раскройки оружия в целом из-за привыкания к лабиринту. Эти данные подчеркивают важность надлежащей контрольной группы для проведения различия между поведенческими эффектами, опосредованными оптогенетической стимуляцией, и возможной поведенческой адаптацией. Кроме того, эти данные проливают свет на важность правильной адаптации экспериментальной установки в соответствии с конкретной линией мусса и экспериментальным вопросом.
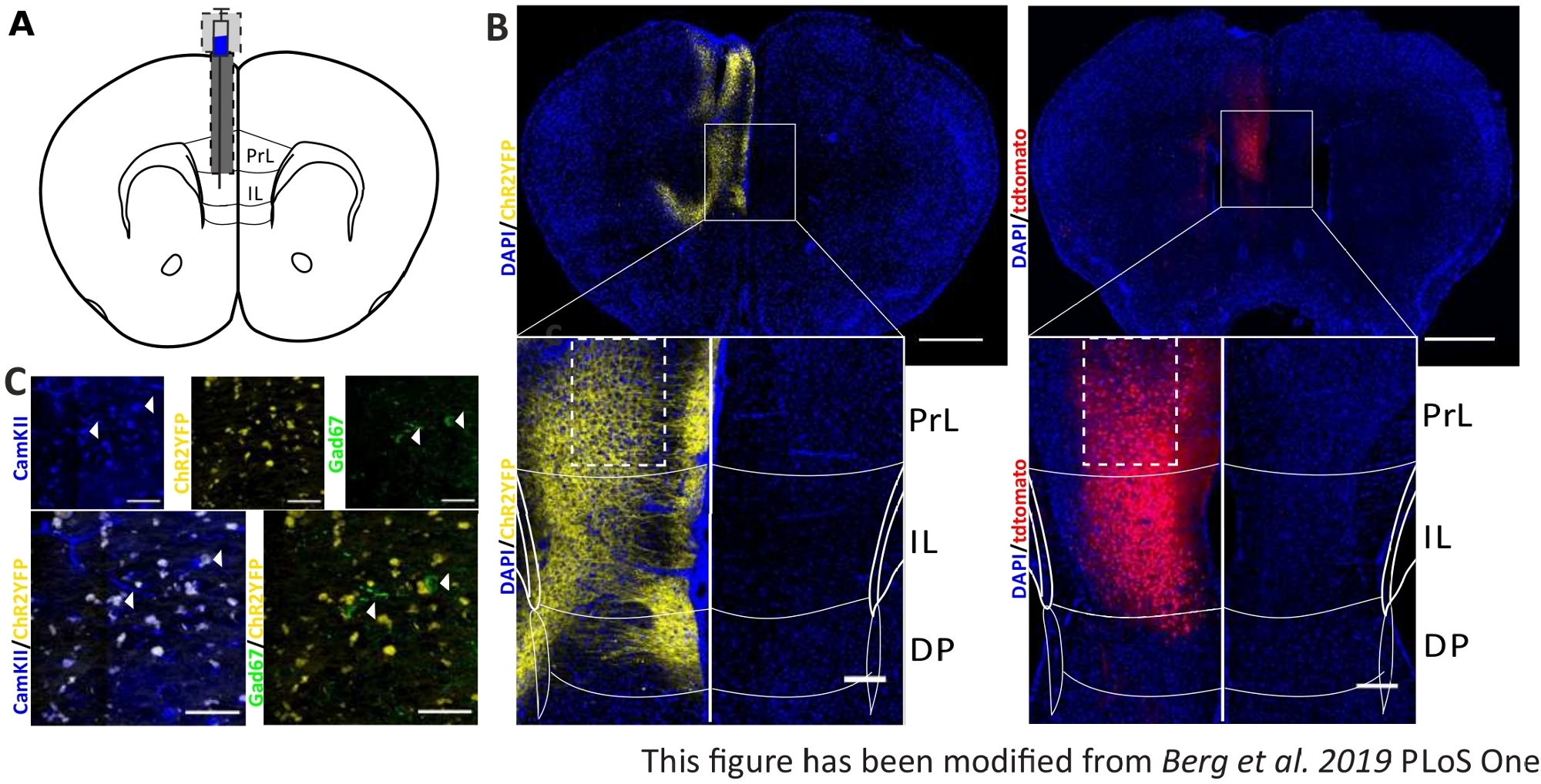
Для проверки и укрепления собранных поведенческих данных мозг мышей удаляется после последнего эксперимента по контролю за правильной инъекцией и имплантацией(рисунок 6). Мозги фиксируются в 4% параформальдегиде и удаляются из черепа. Мозг обезвоживается в 30% сахарозы в течение 1-2 дней и криослоцирован впоследствии. 40 мкм толщиной корональных ломтиков мозга мыть и монтируется на сверххолодных объективных слайдов с монтажной среды, содержащей DAPI, который пятна ядра клеток. Это позволяет идентифицировать целевые области в корональных срезах. Флуоресценция YFP-тега или tdTomato сама указывает на расположение инъекций вируса. На рисунке 6B представлены образцовые места инъекций ChR2-YFP слева (желтый) и tdTomato справа (красный). С помощью шаблона, адаптированного из атласа мозга мыши Paxinos и Franklin45, область IL может быть идентифицирована. В обоих слайдах оптогенетический инструмент выражается не только в области IL, но и в соседних областях мозга. Для правильной интерпретации, распространение синего света в тканях мозга консультируется8 (Рисунок 1D,E). Видно, что синий свет достигнет области DP ниже IL только с менее чем 5% от первоначальной интенсивности света 1 мВт на кончике волокна (Голубая линия на рисунке 1D)8. Кроме того, небольшое количество света может пойти вверх в регион PrL из-за обратного рассеяния47. Следовательно, можно сказать, что область IL освещена наиболее сильно, однако соседние регионы, такие как DP и PrL региона также могут быть слегка стимулировали. Таким образом, ИЛ-клеточная специфическая стимуляция не гарантируется, и иммуногистохимический анализ прилегающих регионов должен быть выполнен, чтобы увидеть, если активность клеток PrL и DP модулируется через свет. На рисунке 6Cпоказан еще один важный элемент управления: специфика линии мыши Nex-Cre. Через антитела окрашивания против двух типов клеток в регионе IL, глутаматергический принцип нейронов и ГАМК-интернейронов, можно увидеть, что выражение ChR2-YFP происходит только в глутаматергических нейронов, а не с ГАМК.
В целом, наши эксперименты показывают, что при оптогенетических манипуляциях во время поведенческого тестирования можно наблюдать изменения в поведении, связанном с тревогой. Используя несколько тестов для одного и того же поведения, можно сделать надежный вывод. Кроме того, иммуногистохимический анализ подтверждает полученные данные. Наши эксперименты показывают, что специфическая активация пирамидальных нейронов в инфралибной коре увеличила тревожное поведение в определенных анализах.

Рисунок 4: Оптогенетическая активация пирамидальных нейронов IL увеличивает тревожное поведение. Стимуляция света во время экспериментов: 473 нм, 1 мВт, 20 Гц стимуляции. A)Схематический рисунок места инъекций и имплантации для ChR2-YFP или tdTomato в IL. В ходе эксперимента пирамидальные нейроны в области IL mPFC активируются ChR2. Saggital ломтики мозга адаптированы из Paxinos и Франклин мыши атлас мозга, saggital: боковой o,6. B) Открытое поле лабиринт с протоколом стимуляции света (20 мин с 4x5 мин чередующихся Off и On испытаний; слева) и тепловые карты образцовых ChR2-инъекций (EXP) и tdTomato-инъекций (КТ) мышей во всех 4 испытаниях эксперимента (справа). EXP животные проводят меньше времени в центре OF, когда стимулировали с синим лазерным светом. Для КТ животных время, проведенное в центре, не отличается между светом Off и On испытаний. C)Данные группы для животных EXP в OF, n'11. Мыши тратят значительно меньше времени в центре OF, когда стимулируется с синим светом (Off1 39,49±6,9 с, На 1 19.87±4.47 s, Off2 28.13±8.55 s, On2 23.42±9.32 s, Off1:On1, t-test, p 0.033, Off1:On2, MWRS, стр.0,049, . Расстояние перемещено не влияет (Off1 2703.09±292.65 cm, On1 3113.4±491.15 cm, Off2 3331.86 ±482.62 cm, On2 3082.17±658.61 см). - центральные записи уменьшаются со временем, но не показывают существенных различий (с 1 22.36±3.78, On1 18.45±3.95, Off2 17.36±1.99, On2 13.27±2.64). D) Данные группы для животных CT в OF, n'15. Время мышей проводят в центре OF, пройденное расстояние, q входов центра не меняет между светом дальше и с проб (Время в центре Off116.73±2.65 s, On1 16.02±1.89 s, Off2 12.02±1.76 s, On2 13.04±2.58 s; Расстояние От 1 3399.69±296.77 cm, On1 3210.6±446.9 cm, Off2 3030.28±513.83 cm, On2 2955±617.7 cm; - центральных записей Off1 14.2±1.98, On1 13.6±2.02, Off2 10.8±1.88, On2 11.67±2.5). КТ мышей показывают значительно более высокую базовую тревожность (Off1 EXP: CT, MWRS, p'0.005, q). Значения средние±S.E.M. - указывают на значительные различия (p≤0.05), - указывают на значительные различия (p≤0.01). t-test всегда двуххвостый, MWRS: тест Манн-Уитни Rank Sum; IL: инфралибическая кора; BLA: базолатеральная миндалина; DRN: спинные ядра рафэ; OF: Открытое поле; КТ: контроль животных; EXP: экспериментальное животное; Л: легкий. Эта цифра была изменена с Berg et al. 2019, PLoS One43 и с Berg 201948. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 5: Эксперимент EPM не показал поведенческих эффектов у мышей Nex-Cre. Стимуляция света во время экспериментов: 473 нм, 1 мВт, 20 Гц стимуляции. A)Повышенный-Плюс лабиринт с протоколом стимуляции света (18 мин, 6x3 мин, чередуя Off и On испытаний). B) Данные группы для мышей, которые "соскользнули" включены в данные, всего No 23. Мышей Nex-Cre имели тенденцию соскальзывания с открытой руки с их задние лапы, независимо от экспериментальной группы (слева). Только мыши, которые остались в лабиринте для всех 6 испытаний были рассмотрены в более поздних анализов. Slip Off в первом этапе Off1 являются причиной для более позднего избежания сеяных рук (Off1 EXP 1.63±0.6, КТ 2.2±0.79, Off2'3 EXP 0.125±0.125, CT 0±0, On1'2'3 EXP 0.625±0.26, CT 0.1±0.1). Pie диаграммы (справа) показывает мышей, падающих из лабиринта в течение 18 минут с 42,42%. Только 57,57% закончили эксперимент. C)Тепловые карты образцовых EXP и КТ мышей во всех 6 испытаниях эксперимента. Обе группы показывают уменьшение продолжительности открытой руки после суда Off1. D) Данные группы для животных EXP в EPM, n'12. Время, проведенное в сеяных руках значительно сократилось в течение первых двух испытаний и постоянно потом (Off1 73,91±12,22 с, On1 36,15±14,65 с, Off2 15.61±6.23 s, On2 19.49±7.51 s, Off3 9.36±4.44 s, On3 7.96±3.47 s. Off1:On1, t-test, p'0,041, q). Пройденное расстояние не влияет (Off1 679.96±71.63 cm, On1 712.24±112.82 cm, Off2 717,49±97,39 см, On2 782,51±81,11 см, Off3 722,11±68,60 см, On3 663,90±106,57 см). Количество открытых записей руки значительно уменьшается от Off1 до On1, а затем остается постоянным (Off1 8.08±1.08, На 1 3.33±0.76, Off2 2.16±0.69, на 2.91±1.09, Off3 1.73±0.75, на3 1.73±0.66. Off1:On1, t-test, стр.0.002, q). Время, проведенное в центре EPM уменьшается вдоль испытаний, но не показывает существенной разницы от Off to On trial (Off1 41.71±5.34 s, На 1 31,2±4,59 с, Off2 19,8±3,44 с, На 2 24,49±3,38 с, Off3 20,37±4,77 с, На3 18,85±4,07 с). E) Данные группы для КТ животных в EPM, n-11. Данные КТ показывают такое же значительное снижение, как и данные EXP, что указывает на то, что эксперимент не сработал должным образом (время в сеяных объятиях Off1 86.92±12.74 s, On1 33.78±14.38 s, Off2 18.01±11.61 s, On2 16.41±9.61 s, Off3 11.36±4.01 s, On3 5.43±2.07 s. Off1:On1, MWRS, p'0.009, Расстояние от 1 705,11±88,36 см, On1 789,45±77,53 см, Off2 724.74±80.49 см, On2 676.57±111.99 см, Off3 716.99±132.47 cm, On3 663.03±132.46 см; Открытые записи руку Off1 7.09±1, On1 3.72±1.17, Off2 1.45±0.47, On2 1.36±0.58, Off3 0.91±0.43, Off3 1.64±0.9. Off1:On1, MWRS, стр. 0,01, Время проводит в центре Off1 35.89 s, На 1 29.25±3.96 s, Off2 22.17±3.58 s, On2 15.9±2.57 s, Off3 13.86±4.2 s, На3 16.89±5.75 s). Значения являются средними ± S.E.M. - указывают на значительные различия (p≤0.05), - указывают на значительные различия (p≤0.01). t-test всегда двуххвостый, MWRS: тест Манн-Уитни Rank Sum; EPM: Повышенный плюс лабиринт; КТ: контроль животных; EXP: экспериментальное животное; О.А.: с открытыми объятиями. Эта цифра была изменена с Berg et al. 2019, PLoS One43 и с Berg 201948. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 6: Инъекционная сторона ChR2 и tdTomato в специфичности IL и Nex-Cre. A)Схематический рисунок места имплантации на короналовых срезах мозга на AP 1,66 мм, мл 0,3 мм, DV -1,8 мм, с односторонней инъекцией и имплантацией (адаптирован из атласа мозга мыши, Paxinos и Франклин, Bregma 1,54 мм). B) Образцовые места инъекций ChR2-YFP (слева, желтый) и tdTomato (справа, красный) слились с ядрами окрашенных клеток DAPI (синий) у мышей Nex-Cre. Масштабная планка 1 мм. Insets показывают высокое увеличение il области. Шкала бар 150 мкм. Белые ящики указывают расположение вставки. C) Верхний ряд: конфокальные изображения левой области IL мыши Nex-Cre, окрашенной CamKII в качестве маркера для глутаматергических нейронов (синий), и ChR2-YFP (желтый) или Gad67 в качестве маркера для ГАМК-нейронов (зеленый), Nex-Cre мыши. Нижний ряд: колокализация ChR2-YFP (желтый) с CamKII (слева, синий), но не с Gad67 (справа, зеленый), показывая специфику Nex-Cre мышей для глутаматергических нейронов. Шкала бар 50 мкм. PrL: прелимбинальная кора; IL: инфралибическая кора; ДП: спинная педункулярная кора. Эта цифра была изменена с Berg et al. 2019, PLoS One43 и с Berg 201948. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}
Обсуждение
Использование света для манипулирования нейрональной сигнализации был метод выбора в течение почти одного десятилетия. С 2005 года количество опубликованных статей о разработкеновых оптогенетических инструментов 4,,6,,8,,14,,49,,50,51 и исследованиях,где такие инструменты используютсядля исследования мозговых цепей 21,,23,,40,,43,,52,значительно увеличилось. С одной стороны, с огромным разнообразием инъекционных оптогенетических инструментов, вариантов имплантации, трансгенных линий мыши и поведенческих экспериментов, возможность экспериментов многообразна и неограничена. С другой стороны, возможность ошибок при выборе экспериментальных условий очень высока, а эксперименты настолько специфичны, что зачастую сопоставимость с другими исследованиями затруднена.
Критические шаги
Одним из важных важных этапов этого протокола является надлежащее планирование. Выбор оптогенетического инструмента должен соответствовать научному вопросу. Нужно ли только манипулировать общей активностью нейрона или синапса? Тогда коммерчески предоставленные инструменты, такиекак ChR2 21,25,27 и Arch37 являются хорошим выбором. Но кроме этого, если один специальный нейромедиатор системы или даже одного рецептора должны манипулировать, индивидуальный рецептор химера частолучший выбор 3,6. Несколько рецепторов химер с GPCRs, так называемые Opto-XRs, и руководящие принципы для их производства ужедоступны 4,50. Помимо выбора оптогенетических инструментов, линия мыши в сочетании с поведенческим экспериментом также имеет решающее значение. Различные фоновые штаммы, такие как, например, C57Bl/6 и BALB/cByJ, отображают различные поведенческие фенотипыв некоторых отношениях 53,54. C57Bl/6 мышей имеют низкую базовую тревожность и могут быть использованы для аксиогенных манипуляций, в то время как BALB / cByJ показать более высокий уровень тревоги и, следовательно, более чувствительны к анксиолитических препаратов. Кроме того, трансгенные варианты этих фоновых штаммов также могут варьироваться в их фенотипе48. При правильном сочетании конкретных промоутеров в сочетании с оптогенетическим инструментом и трансгенной линией мыши, почти каждая желаемая популяция клеток может быть мишенью.
Важным шагом во время операции является таргетинг правильного местоположения. С помощью атласа мозга мыши, правильные координаты передней-задней оси, и медиальной боковой оси, и глубина структуры могут бытьустановлены 45. На самом деле, каждый череп имеет несколько иную форму и размер. Таким образом, F-фактор46 для регулировки стереотаксических координат является весьма важным, как и правильное фиксация носа и уха во время стереотаксической хирургии. Если головка мыши наклонена, инъекционная канула не сможет ориентироваться на нужную область интереса.
Кроме того, диаметр инъекционной канулы также имеет решающее значение. Если он слишком мал, ни один вирус не может быть выпущен в ткань, если он слишком широк, канула будет утечка вирусного раствора на своем пути в область интереса. Если имплантированное оптическое волокно прекращается непосредственно над целевой областью, экспрессия вируса в вышеуказанных областях коры не имеет значения. Но если имплантат находится над другими регионами для стимулирования аксон-терминалов, аксоны верхних областей коры также будут активированы светом и фальсифицировать полученные данные. В качестве примера: Область IL и прелимбийской (PrL) области как проект базальной миндалины55,56, но имеют совершенно разные функции и роли в модуляции тревоги26,57., Если имплантат находится над миндалиной для активации аксонных терминалов из региона IL, а во время инъекции раствор вируса также помещается в PrL из-за неправильной инъекционной канулы, риск также активации аксоновых терминалов из PrL очень высок.
Во время подготовки черепа к фиксации имплантата, редкое использование грунтовка и связь имеет решающее значение для надежной и прочной фиксации. Если 2-компонентная система адгезии не применяется тонко, зубной цемент может отделиться от черепа через пару дней или недель. Кроме того, череп также должен быть полностью высушен перед фиксацией имплантата, так как в противном случае цемент не будет прикрепляться должным образом к черепу.
Критические шаги также существуют в поведенческой части этого протокола. Во-первых, строительство лабиринта очень важно. В каждой поведенческой установке в литературе существует несколько вариантов относительно размера и формы, а также длясамой процедуры 58,,59,,60. Важно выбрать вариант, который делает данные сопоставимыми и воспроизводимыми. Также следует учитывать специальные требования к использованным линиям мыши43,,48. В репрезентативных данных для EPM видно, что несколько мышей Nex-Cre упали из лабиринта или соскользнули несколько раз(рисунок 2b). Для этих мышей лабиринт с небольшой стеной вокруг сеяных рук был бы лучшей альтернативой.
Во-вторых, очень важно сохранить все внешние условия комнатыпостоянными 61, в противном случае различные группы мышей не были бы сопоставимы вообще. В связи с этим очень важно выбрать время эксперимента как место, где экспериментальная установка вакантна, а экспериментатор всегда присутствует. Кроме того, для того, чтобы избежать вмешательства в полученные данные, следует учитывать такие события в здании, как строительные работы, тестирование любых систем (пожарная сигнализация) или день очистки мышей.
Наконец, обработка и жилищные условия имеют решающее значение для поведенческих экспериментов. Когда имплантация выполняется, мышей должны быть одной размещены из-за риска получения травмы от других мышей. Чтобы обеспечить хорошую сопоставимость между группами и низкую ошибку в пределах одной группы, каждая мышь должна иметь одинаковый размер клетки и обогащение. Для тревожных экспериментов, одно жилье имеет некоторые преимущества, как синге размещены мышей показать более низкий базовый уровень тревоги, меньше изменений в их уровень тревоги, и менеедепрессивные, как симптомы 15,16. Группа размещены мышей мужского пола может сильно отличаться по уровню тревоги из-за иерархии среди мышей. Помимо жилья, постоянное и равное обращение со всеми мышами и группами также имеет важное значение. Захват мыши для того, чтобы соединить световое волокно на имплантате очень напряженный. Таким образом, эта процедура должна быть одинаковой для каждой мыши, то есть той же техники и одного и того же экспериментатора. Кроме того, время привыкания в клетке ожидания, которая предназначена для успокоения мыши от стрессовой процедуры подключения, также должна иметь равные условия по продолжительности, помету и положению в лабиринте. Обработка в механизме мыши также имеет решающее значение для более поздних поведенческих характеристик. Экспериментальные и контрольные животные не должны быть очищены в разные дни или разными людьми, так как это также стресс для мышей. Кроме того, день уборки не должен быть экспериментальным днем, чтобы избежать различий в поведении.
Устранение неполадок
Есть несколько проблем, которые могут возникнуть во время протокола. Например, бурение целого в черепе во время стереотаксической операции может привести к повреждению кровеносных сосудов. Как правило, сильное кровотечение происходит, особенно выше брегмы и лямбды. Если это произойдет, не пытайтесь остановить кровотечение с ватными палочками, поскольку они, как правило, распространяются еще больше кровотечения из сосуда из-за их абсорбции, вместо этого, непосредственно промыть NaCl.
Также может случиться так, что инъекция давления вирусного раствора не работает. В этом случае, это может быть то, что парафильм, парши из отверстия заусенцев или ткани мозга, засоряет кончик канулы. В этом случае, удалить канулу медленно из мозга, не изменяя х- или у оси и использовать пинцет, чтобы удалить 1-2 мм передней части кончика канулы. Прежде чем снова снизить канулу, проверьте функциональность, применяя небольшое количество давления, чтобы увидеть, если вирус выходит из кончика канулы. Чтобы избежать запоров, опустите канулу с постоянной скоростью и не останавливайте движение до тех пор, пока не будет достигнута самая глубокая глубина стороны инъекции. Если слишком много кончика канулы удаляется и диаметр слишком велик, канула повредит ткани и риск применения вируса все сразу будет увеличена. Таким образом, убедитесь, что только забитая часть наконечника тщательно удаляется.
Во время поведенческого эксперимента, установка эксперимента в программное обеспечение для отслеживания видео (например, Ethovision XT) может вызвать проблемы. Если, например, световой выход не работает должным образом, это может быть связано с несколькими причинами. Pulser должен быть открыт, запрограммирован и запущен до открытия Ethovision XT. Оборудование должно быть выбрано правильно в "Экспериментальной установке" (шаг 3.2.2.4). Если выбран неправильный IO-Box или что-либо, кроме "Costume Hardware", устройство Pulser не может управляться Ethovision. Если проверка светового вывода будет успешной, но запрограммированный протокол света в "Настройках пробного контроля" не работает во время приобретения, ссылка на подуряд или подуряд может быть расположена неправильно или условия и действия неясны. Например: относится ли ссылка к правильному подправе? Правильно ли запрограммирована ссылка (например, как часто выполняется подуряд)?
Кроме того, может случиться так, что во время "обнаружения настройки" животное надлежащим образом отслеживается, но во время приобретения Есть образцы, где субъект не найден. В этом случае проверьте, была ли изменена подсветка в экспериментальной комнате, или же что-либо произвело нежелательные тени в лабиринте. Вся нижняя часть лабиринта должна иметь тот же цвет, так как настройка будет работать только для одной конкретной комбинации. Если по каким-либо причинам не избежать различных нижних цветов или теней, определите настройки обнаружения в самой темной части лабиринта.
Чтобы изменить настройки после приобретения первых животных, не применяй эти изменения в уже используемых настройках. Дублируйте их, чтобы настроить их. Это также означает, что уже записанное испытание больше не действует для анализа данных. В таком случае зафиксируйте всех животных для этой экспериментальной группы с исходными настройками и создайте новый эксперимент, в котором записанные видео анализируются вместо живого отслеживания. В этом эксперименте «из видео» можно использовать несколько параметров для анализа, не теряя сопоставимости между животными или даже данными.
Ограничения и будущие приложения
Этот метод манипулирования поведением с помощью оптогенетики у свободно движущихся животных также включает в себя ограничения. Во время операции близость двух имплантатов ограничена. Для двойной имплантации расстояние между двумя имплантатами должно быть минимально шириной аппарата для удержания имплантата. Аппарату необходимо опустить второй имплантат в отверстие заусенца, в то время как первые имплантаты уже зафиксированы. Решением для этого может быть угловой имплантации, где кончики стеклянного волокна может быть очень близко в то время как керамические ферулы надчерепом имеют большее расстояние 23,55,56,57,62,63. Недостатком угловой имплантации является распространение света. Когда кончик волокна наклонен вместо прямо выше, стимулируемая область отличается. В случае двух целевых областей, расположенных в непосредственной близости, необходимо учитывать измененную позицию стимуляции света.
Во время поведенческого эксперимента строительство лабиринта может помешать оптическому кабелю, подключенного к животному. Некоторые поведенческие тесты, такие как светло-темная коробка, содержат крытуюобласть 64,65, и другие лабиринты содержат отсеки, которые мышь должна войти. Такие эксперименты не могут быть выполнены с этой установкой. Кроме того, беспроводная система может быть вариант22,26,66. Но, к счастью, некоторые лабиринты, такие как лабиринт Барнса, могут быть расположены таким образом, что мыши могут войти в соответствующиеотсеки 67.
Помимо тех, с закрытыми зонами, лабиринты, которые являются слишком широкими может вызвать также проблемы. Чем больше площадь лабиринта, тем длиннее кабель должен быть, чтобы позволить животному идти в каждую позицию в лабиринте. Необходимо позаботиться о том, чтобы животное не могла наступить на кабель или схватить его и укусить. Решением для этого может быть конструкция, которая свертывает избыточный кабель. Недостатком является то, что перетащить развернуть кабель трудно для мышей. Это решение лучше подходит для крыс. Другим возможным вариантом может быть сделать стимуляцию света заранее, а не во время эксперимента, конечно, это возможно только в том случае, если долгосрочный эффект из-за стимуляции светапроисходит 23.
Сравнение с существующими/альтернативными методами
Альтернативные методы будут химической или электрической стимуляции вовремя поведения 8,,18. Химические агонисты или антагонисты способны активировать или заставить замолчать нейроны через конкретные рецепторы, а также могут манипулировать одной нейромедиаторасистем 38,68. С одной стороны, рецептор-специфичность довольно высока для химических веществ, потому что специфический агонист или антагонист активирует только определенныерецепторы 39. С другой стороны, специфичность рецепторных подтипов одной и той же группы нейромедиаторов часто недостаточна. Большинство химических веществ связываются по крайней мере с двумя подтипами с разной вероятностью69. Кроме того, химические вещества не могут различать типы нейронных клеток до тех пор, пока они обладают теми же типами рецепторов. Помимо этого, временное и пространственное разрешение плохо для химических манипуляций по сравнению с оптогенетикой. Агонисты или антагонисты часто вводят устно35 или с помощью системных инъекций57,70. Если вливание химического вещества делается непосредственно в ткани мозга, эффекты появляются быстрее, чем при пероральном применении, но все же на более медленном сроке, чем при стимуляции света. Поскольку вводимые химические вещества диффузные в головном мозге и не являются специфическими для нейрональных типов или областей мозга, манипуляции конкретных схем мозга не возможно.
Электрическая стимуляция имеет более высокое временное разрешение, чемхимическая стимуляция 9,,14. Распространение внутри нейронной ткани меньше, чем при химической стимуляции, и пространственное разрешение лучше, чем при химической стимуляции. Тем не менее, электрическая стимуляция не имеет возможности конкретно решать различные типы нейронных клеток или типов рецепторов, так как каждый нейрон в непосредственной близости от электрода будет реагировать на электрическую стимуляцию.
Альтернативные методы поведения у свободно движущихся мышей, например, электрофизиологические записи в срезах мозга, где одиночные нейроны или аксоны могут модулироваться с помощью оптогенетики и вызвали эффекты могут быть измерены с помощью записиэлектродов 6,71. Эксперименты in vitro предлагают возможность исследовать молекулярную и клетчатую основу оптогенетической стимуляции, но имеют ограничение, что внутренняя связь и вход из других областей мозга отсутствует. Другим вариантом является использование оптогенетики в сочетании с мультифотоннойвизуализацией 1,,72. В этом случае, мыши имеют их голову фиксированной и может быть анестезировано или бодрствовать, чтобы решить простые задачи.
Для успешного оптогенетического эксперимента в настоящее время доступен широкий спектр инструментов и приложений. Выбор оптогенетических инструментов и поведенческой настройки имеет решающее значение для ответа на конкретные вопросы исследования. При выборе правильного сочетания инструментов и экспериментов оптогенетика позволяет провести беспрецедентное, углубленное исследование нейронных схем с высоким временным и пространственным разрешением. Это поможет понять и разработать новые терапевтические стратегии для психических заболеваний и познания.
Раскрытие информации
Авторов нечего раскрывать.
Благодарности
Большое спасибо профессору Клаусу-Армину Нарве и доктору Сандре Геббельс (Max-Plank-Institute of Experimental Medicine, Goettingen, Германия) за любезное предоставление мышей Nex-Cre. Кроме того, мы благодарим нашу видео-команду Yunus Dikici и Рубена Визнера за запись и обработку видео JoVE для этой статьи. Кроме того, большое спасибо Кристин Клауссен за ее голос за кадром и Кимберли Энн Go для корректирования рукописи.
Представленные результаты были получены в Рур-университете в Бохуме, и видео было записано в Бременской университете.
Эта работа финансировалась Deutsche Forschungsgemeinschaft (DFG, Немецкий исследовательский фонд) - Projektnummer 122679504 - SFB 874 и DFG MA 4692/3-2.
Материалы
| Name | Company | Catalog Number | Comments |
| Ketamin | Sigma-Aldrich | K2753-64 | Anestasia |
| 20 % Glucose | AlleMan Pharma | Injection s.c. for fast recovery | |
| Behavioral mazes | Costum made | Measure anxiety | |
| Bepanthen | Bayer | Ophthalmic oinment | |
| Betaisodona | Monodipharma | Sterilant containing iodine | |
| Betaisodona | Monodipharma | Iodine oinment | |
| Binocular | Olympus | SZ52, 110AL0.62x WD160 | Surgery |
| Ceramic ferrules | Thorlabs | CFLC230-10 | Implant |
| Ceramic Fiber Scribe | Thorlabs | CSW12.5 | Cutting of the glass fiber |
| Channelrhodopsin2-YFP virus | Penn Vector Core | Addgene 20298 | Optogenetic tool |
| Compressed air | Kontakt Chemie | Druckluft 67 | Drying of the skull |
| Coordinate system | Stoelting | Stereotactic coordinates for the surgery | |
| Correl Draw | Graphical software version 13 | ||
| Cryoslicer | MICROM | HM500OM | Production of brain slices for staining |
| Ethovision XT 14 | Noldus | Software for behavioral tracking | |
| Exel | Statistical Software | ||
| Ferrule Polishing Puck | Thorlabs | D50-F | Polishing implants round side |
| Fiber Patch Cord dual | Prizmatix | Optogenetics-Fiber 500, 1,20 m, Ferrule core 1,25 mm | Cables, which are connected with the two implants of a bilateral implantation |
| Fiber Patch Cord single | Prizmatix | Optogenetics-Fiber 500, 1,20 m, Ferrule core 1,25 mm | Cable, which is connected with the implant via a sleeve |
| Fiber Stripping Tool | Thorlabs | T06S13 | Stripping glass fiber for implant |
| Filter paper | VWR European | 516-0300 | Cut into pieces for the Novelty-Suppressed Feeding test |
| Food pellets | Mühle Levers | Höveler Nagerfutter | Nutrition for the mice |
| Glass pipettes | Harvard Apparatus | GC150-10 | Injection pipettes |
| Gradia direct-Flo | Henry Schein | 103322 | Fluid dental cementum |
| Heating lamp | efbe-Schott/Phillips | R95E | Prevent the mice from cooling after the surgery |
| Heating plate | Stoelting | Integrated into coordinate system | |
| Injection canula | Braun | 100 Sterican, 0,4 x 20 mm, Gr. 20 | All injections and to bore hole into the skull |
| Litter | T 1350 | Grounding for the Novelty-Supressed Feeding test | |
| Mouse cages | Zoonlab | 405 cm^2 | Single housing for experiments |
| Optibond FL | Kerr | 26684E | Preparation of the skull for implantation |
| Optical glass fiber | Thorlabs | FT200EMT | Light fiber for implant |
| Optogenetics-LED.STSI | Prizmatix | Optogenetic toolbox for light stimulation during behavioral experiments | |
| Paraformaldehyde | Sigma-Aldrich | 16005-1KG-R | Perfusion of mice to remove the brains |
| Polishing sheet 0.02 µm grit | Thorlabs | LFCF | Polishing implants round side |
| Polishing sheet 1 µm grit | Thorlabs | LF1D | Polishing implants round side |
| Polishing sheet 30 µm grit | Thorlabs | LF30D | Polishing implants round side |
| Polishing sheet 6 µm grit | Thorlabs | LF6D | Polishing implants round side |
| Pulser Software | Prizmatix | Software for light device control | |
| Rimadyl-Carprofen | Zoetis | Analgesia | |
| Sigma Plot | Software for statistics | ||
| Sleeve | Thorlabs | FT200EMT | Connection of implant and light cable |
| SodiumCloride (NaCl) | Braun | 3570410 | Rinsing of the skull |
| Superglue | Pattex Henkel | To Fix the glass fiber in the ferrule | |
| td-Tomato virus | Penn Vector Core | Addgene 51503 | Optogenetic tool |
| UV light | KoQGHJ | wireless, 1200 mW/cm^2 | Polymeration lamp for dental cementum |
| Xylavet-Xylazin | cp pharma | Anesthesia |
Ссылки
- Chow, B. Y., et al. High-performance genetically targetable optical neural silencing by light-driven proton pumps. Nature Letters. 463, 98-102 (2010).
- Zhao, S., et al. Cell type-specific channelrhodopsin-2 transgenic mice for optogenetic dissection of neural circuitry function. Nature Methods. 8, 745-752 (2011).
- Spoida, K., Masseck, O. A., Deneris, E. S., Herlitze, S. Gq/5-HT2c receptor signals activate a local GABAergic inhibitory feedback circuit to modulate serotonergic firing and anxiety in mice. Proceedings of the National Academy of Science of the United States of America. 111, 6479-6484 (2014).
- Kleinlogel, S. Optogenetic user's guide to Opto-GPCRs modified GPCRs. Frontiers in Bioscience. 21, 794-805 (2016).
- Mahn, M., Prigge, M., Ron, S., Levy, R., Yizhar, O. Biophysical constraints of optogenetic inhibition at presynaptic terminals. Nature Neuroscience. 19, 554-556 (2016).
- Masseck, O. A., et al. Vertebrate Cone Opsins Enable Sustained and Highly Sensitive Rapid Control of Gi/o Signaling in Anxiety Circuitry. Neuron. 81, 1263-1273 (2014).
- Oh, E., Maejima, T., Liu, C., Deneris, E., Herlitze, S. Substitution of 5-HT 1A Receptor signaling by a light-activated G protein-coupled receptor. Journal of Biological Chemistry. 285, 30825-30836 (2010).
- Yizhar, O., Fenno, L. E., Davidson, T. J., Mogri, M., Deisseroth, K. Optogenetics in Neural Systems. Neuron Primer. 71, 9-34 (2011).
- Masseck, O. A., Manahan-Vaughan, D. A Guide to Optogenetic Applications, With special Focus on Behavioral and In Vivo Electrophysiological Experiments. HandboOk of In Vivo Neural Plasticity Techniques - A Systems Neuroscheince Approach to the Neural Basis of Memory and Cognition. , 557 (2019).
- Goebbels, S., et al. Genetic Targeting of Principal Neurons in Neocortex and Hippocampus of NEX-Cre Mice. Genesis. , 611-621 (2006).
- Yang, Y. S., Hughes, T. E. Cre Stoplight: A red/green fluorescent reporter of Cre recombinase expression in living cells. Biotechniques. 31, 1036-1041 (2001).
- Schnütgen, F., et al. A directional strategy for monitoring Cre-mediated recombination at the cellular level in the mouse. Nature Biotechnology. 21, 562-565 (2003).
- Taniguchi, H., et al. A Resource of Cre Driver Lines for Genetic Targeting of GABAergic Neurons in Cerebral Cortex. Neuron. 71, 995-1013 (2011).
- Deisseroth, K. Optogenetics. Nature Methods. 8, 268-273 (2011).
- Palanza, P., Gioiosa, L., Parmigiani, S. Social stress in mice: Gender differences and effects of estrous cycle and social dominance. Physiology and Behavior. 73, 411-420 (2001).
- Karolewicz, B., Paul, I. A. Group housing of mice increases immobility and antidepressant sensitivity in the forced swim and tail suspension tests. European Journal of Pharmacology. 415, 197-201 (2001).
- Masseck, O. A., Rubelowski, J. M., Spoida, K., Herlitze, S. Light- and drug-activated G-protein-coupled receptors to control intracellular signalling. Experimental Physiology. 96, 51-56 (2011).
- Aravanis, A. M., et al. An optical neural interface: in vivo control of rodent motor cortex with integrated fiberoptic and optogenetic technology. Journal of Neural Engineering. 4, (2007).
- Zhang, F., et al. Multimodal fast optical interrogation of neural circuitry. Nature Article. 446, 633-639 (2007).
- Owen, S. F., Liu, M. H., Kreitzer, A. C. Thermal constraints on in vivo optogenetic manipulations. Nature Neuroscience. 22, 1061-1065 (2019).
- Hare, B. D., et al. Optogenetic stimulation of medial prefrontal cortex Drd1 neurons produces rapid and long-lasting antidepressant effects. Nature Communication. 10, 1-12 (2019).
- Allsop, S. A., Vander Weele, C. M., Wichmann, R., Tye, K. M. Optogenetic insights on the relationship between anxiety-related behaviors and social deficits. Frontiers in Behavioral Neuroscience. 8, 1-14 (2014).
- Fuchikami, M., et al. Optogenetic stimulation of infralimbic PFC reproduces ketamine's rapid and sustained antidepressant actions. Proceedings of the National Academy of Science of the United States of America. 112, 8106-8111 (2015).
- Correia, P. A., et al. Transient inhibition and long-term facilitation of locomotion by phasic optogenetic activation of serotonin neurons. Elife. 6, 1-26 (2017).
- Felix-Ortiz, A. C., Burgos-Robles, A., Bhagat, N. D., Leppla, C. A., Tye, K. M. Bidirectional modulation of anxiety-related and social behaviors by amygdala projections to the medial prefrontal cortex. Neuroscience. 321, 197-209 (2016).
- Marek, R., Xu, L., Sullivan, R. K. P., Sah, P. Excitatory connections between the prelimbic and infralimbic medial prefrontal cortex show a role for the prelimbic cortex in fear extinction. Nature Brief Communication. , (2018).
- Parfitt, G. M., et al. Bidirectional Control of Anxiety-Related Behaviors in Mice: Role of Inputs Arising from the Ventral Hippocampus to the Lateral Septum and Medial Prefrontal Cortex. Neuropsychopharmacology. 42, 1715-1728 (2017).
- Bandelow, B., Michaelis, S. Epidemiology of anxiety disorders in the 21st century. Dialogues in Clinical Neuroscience. 17, 327-335 (2015).
- Kessler, R. C., et al. Lifetime Prevalence and Age-of-Onset Distributions of DSM-IV Disorders in the National Comorbidity Survey Replication. Archives of General Psychiatry. 62, 593-602 (2005).
- Kessler, R. C., Petukhova, M., Sampson, N. A., Zaslavsky, A. M., Wittchen, H. U. Twelve-month and lifetime prevalence and lifetime morbid risk of anxiety and mood disorders in the United States. International Journal of Methods Psychiatric Research. 21, 169-184 (2014).
- Andlin-Sobocki, P., Wittchen, H. U. Cost of anxiety disorders in Europe. European Journal of Neurology. 12, 39-44 (2005).
- Forster, G. L., Novick, A. M., Scholl, J. L., Wall, M. J. The Role of the Amygdala in Anxiety Disorders. Intech. , 61-102 (2012).
- Liberzon, I. Neural circuits in anxiety and stress disorders a focused review. Therapeutics and Clinical Risk Management. 11, 115-126 (2015).
- Sylvers, P., Lilienfeld, S. O., LaPrairie, J. L. Differences between trait fear and trait anxiety: Implications for psychopathology. Clinical Psychology Review. 31, 122-137 (2011).
- Daws, L. C., Koek, W., Mitchell, N. C. Revisiting Serotonin Reuptake Inhibitors and the Therapeutic Potential of 'Uptake-2' in Psychiatric Disorders. ACS Chemical Neuroscience. 4, 16-21 (2013).
- Felix-Ortiz, A. C., et al. BLA to vHPC inputs modulate anxiety-related behaviors. Neuron Report. 79, 658-664 (2013).
- Padilla-Coreano, N., et al. Direct Ventral Hippocampal-Prefrontal Input Is Required for Anxiety-Related Neural Activity and Behavior. Neuron Article. 89, 857-866 (2016).
- Lisboa, S. F., Stecchini, M. F., Corrêa, F. M. A., Guimarães, F. S., Resstel, L. B. M. Different role of the ventral medial prefrontal cortex on modulation of innate and associative learned fear. Neuroscience. 171, 760-768 (2010).
- Bi, L. L., et al. Enhanced excitability in the infralimbic cortex produces anxiety-like behaviors. Neuropharmacology. 72, 148-156 (2013).
- Yizhar, O., et al. Neocortical excitation/inhibition balance in information processing and social dysfunction. Nature Article. 477, 171-178 (2011).
- Goebbels, S., et al. Genetic Targeting of Principal Neurons in Neocortex and Hippocampus of NEX-Cre Mice. Genesis. 44, 611-621 (2006).
- Rubenstein, J. L. R., Merzenich, M. M. Model of autism: increased ratio of excitation/ inhibition in key neural systems. Genes, Brain and Behavior. 2, 255-267 (2003).
- Berg, L., Eckardt, J., A, M. O. Enhanced activity of pyramidal neurons in the infralimbic cortex drives anxiety behavior. PLoS One. 14, 1-19 (2019).
- Meunier, C. N. J., Amar, M., Lanfumey, L., Hamon, M., Fossier, P. 5-HT1A receptors direct the orientation of plasticity in layer 5 pyramidal neurons of the mouse prefrontal cortex. Neuropharmacology. 71, 37-45 (2013).
- Paxinos, G., Franklin, K. B. J., Paxinos, G., Franklin, K. B. J., Paxinos, G., Franklin, K. B. J. . Mouse Brain in Stereotaxic Coordinates. 2, (2004).
- Gore, B. B., Soden, M. E., Zweifel, L. S. Manipulating gene expression in projection-specific neuronal populations using combinatorial viral approaches. Current Protocols in Neuroscience. 435, 1-6 (2014).
- Stujenske, J. M., Spellman, T., Gordon, J. A. Modeling the Spatiotemporal Dynamics of Light and Heat Propagation for InVivo Optogenetics. Cell Report. 12, 525-534 (2015).
- Berg, L. Imbalance of excitation and inhibition within the prefrontal cortex supports anxiety behavior. Ruhr-University Bochum. , (2019).
- Boyden, E. S. A history of optogenetics: The development of tools for controlling brain circuits with light. F1000 Biology Reports. 3, 1-12 (2011).
- Airan, R. D., Thompson, K. R., Fenno, L. E., Bernstein, H., Deisseroth, K. Temporally precise in vivo control of intracellular signalling. Nature. 458, 1025-1029 (2009).
- Sparta, D. R., et al. Construction of implantable optical fibers for long-term optogenetic manipulation of neural circuits. Nature Protocol. 7, 12-23 (2012).
- Covington, H. E., et al. Antidepressant Effect of Optogenetic Stimulation of the Medial Prefrontal Cortex. Journal of Neuroscience. 30, 16082-16090 (2010).
- Lepicard, E. M., Joubert, C., Hagneau, I., Perez-Diaz, F., Chapouthier, G. Differences in anxiety-related behavior and response to diazepam in BALB/cByJ and C57BL/6J strains of mice. Pharmacology, Biochemistry and Behavior. 67, 739-748 (2000).
- Schmidt, M. V., Müller, M. B. Animal models of anxiety. Elsevier. 3, 369-374 (2006).
- Cho, J. H., Deisseroth, K., Bolshakov, V. Y. Synaptic Encoding of Fear Extinction in mPFC-amygdala Circuits. Neuron Article. 80, 1491-1507 (2013).
- Adhikari, A., et al. Basomedial amygdala mediates top-down control of anxiety and fear. Nature. 527, 179-185 (2015).
- Suzuki, S., et al. The infralimbic and prelimbic medial prefrontal cortices have differential functions in the expression of anxiety-like behaviors in mice. Behavioural Brain Research. 304, 120-124 (2016).
- Carola, V., D'Olimpio, F., Brunamonti, E., Mangia, F., Renzi, P. Evaluation of the elevated plus-maze and open-field tests for the assessment of anxiety-related behaviour in inbred mice. Behavioural Brain Research. 134, 49-57 (2002).
- Prut, L., Belzung, C. The open field as a paradigm to measure the effects of drugs on anxiety-like behaviors: A review. European Journal of Pharmacology. 463, 3-33 (2003).
- Tye, K. M., et al. Amygdala circuitry mediating reversible and bidirectional control of anxiety. Nature Letter. 471, 358-362 (2011).
- Bouwknecht, J. A., et al. Differential effects of exposure to low-light or high-light open-field on anxiety-related behaviors: Relationship to c-Fos expression in serotonergic and non-serotonergic neurons in the dorsal raphe nucleus. Brain Research Bulletin. 72, 32-43 (2007).
- Overstreet, D. H., Knapp, D. J., Angel, R. A., Navarro, M., Breese, G. R. Reduction in repeated ethanol-withdrawal-induced anxiety-like behavior by site-selective injections of 5-HT1A and 5-HT2C ligands. Psychopharmacology. 187, 1-12 (2006).
- Takahashi, A., et al. Glutamate Input in the Dorsal Raphe Nucleus As a Determinant of Escalated Aggression in Male Mice. Journal of Neuroscience. 35, 6452-6463 (2015).
- Klemenhagen, K. C., Gordon, J. A., David, D. J., Hen, R., Gross, C. T. Increased Fear Response to Contextual Cues in Mice Lacking the 5-HT1A Receptor. Neuropsychopharmacology. 31, 101-111 (2006).
- Ramos, A. Animal models of anxiety: do I need multiple tests. Trends in Pharmacological Science. 29, 493-498 (2008).
- Isosaka, T., et al. Htr2a-Expressing Cells in the Central Amygdala Control the Hierarchy between Innate and Learned Fear. Cell. 163, 1153-1164 (2015).
- Regev, L., Goshen, I. Employing Optogenetics in Memory Research. Optogenetics: A Roadmap. , 219-256 (2017).
- Shah, A. A., Sjovold, T., Treit, D. Inactivation of the medial prefrontal cortex with the GABA A receptor agonist muscimol increases open-arm activity in the elevated plus-maze and attenuates shock-probe burying in rats. Brain Research. 1028, 112-115 (2004).
- Knight, A. R., et al. Pharmacological characterisation of the agonist radioligand binding site of 5-HT2A, 5-HT2B and 5-HT2C receptors. Naunyn-Schmiedebergs Archiv of Pharmacology. 370, 114-123 (2004).
- Graeff, F. G., Viana, M. B., Mora, P. O. Dual Role of 5-HT in Defense and Anxiety. Neuroscience and Biobehavioral Reviews. 21, 791-799 (1997).
- Cheriyan, J., Sheets, P. L. Altered Excitability and Local Connectivity of mPFC-PAG Neurons in a Mouse Model of Neuropathic Pain. Journal of Neuroscience. 38, 4829-4839 (2018).
- Boyden, E. S., Zhang, F., Bamberg, E., Nagel, G., Deisseroth, K. Millisecond-timescale, genetically targeted optical control of neural activity. Nature Neuroscience. 8, 1263-1268 (2005).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены