
Method Article
Многоклеточная альвеолярная модель человека, состоящая из эпителиальных клеток и первичных иммунных клеток для оценки опасности
* Эти авторы внесли равный вклад
В этой статье
Резюме
Здесь представлен протокол первичной изоляции моноцитов крови человека, а также их дифференциации на макрофаги и дендритные клетки и сборки с эпителиальными клетками в многоклеточную модель легких человека. Биологические реакции кокультур, состоящих из иммунных клеток, дифференцированных либо от свежеиспеченные или оттаянные моноциты, при воздействии провоспалительных стимулов, сравниваются.
Аннотация
Модель совместной культуры альвеолярных клеток человека описана здесь для моделирования альвеолярного эпителиального барьера тканей, состоящего из альвеолярных эпителиальных клеток типа II и двух типов иммунных клеток (т.е. макрофагов, полученных из моноцитов человека ,MDM) и дендритных клеток (MDDCs). Предусмотрен протокол для сборки многоклеточной модели. Альвеолярные эпителиальные клетки (линия клеток A549) выращиваются и дифференцированы в условиях погружения на проницаемые вставки в двухкамерных скважинах, а затем в сочетании с дифференцированными MDM и MDDCs. Наконец, клетки подвергаются воздействию воздушно-жидкого интерфейса в течение нескольких дней. Поскольку первичные иммунные клетки человека должны быть изолированы от человека баффи пальто, иммунные клетки отличаются от свежих или оттаяли моноцитов сравниваются для того, чтобы адаптировать метод, основанный на экспериментальных потребностей. Трехмерные модели, состоящие из альвеолярных клеток со только что изолированными или оттаянными моноцитными иммунными клетками, показывают статистически значимое увеличение цитокинов (интерлекинов 6 и 8) при воздействии провоспалительных стимулов (липополисахарида и фактора некроза опухоли) по сравнению с необработанными клетками. С другой стороны, нет статистически значимой разницы между высвобождением цитокинов, наблюдаемым в кокультурах. Это показывает, что представленная модель реагирует на провоспалительные стимулы в присутствии МДМ и MDDCs отличается от свежих или оттаяющих периферических моноцитов крови (PBMs). Таким образом, он является мощным инструментом для исследования острой биологической реакции на различные вещества, включая аэрозольные препараты или наноматериалы.
Введение
Культуры клеток легких in vitro предлагают экономически эффективные, надежные и хорошо контролируемые платформы для оценки опасностей аэрозолей1. В качестве модели клеточной системы для человека альвеолярных пневмоцитов, эпителиальной линии клеток A549 изолированы от легочной аденокарциномычасто используется 2. Эти клетки представляют собой плоскоклеточный тип II эпителиальных клетокиз альвеолярной области 3 и широко используются линии клеток легкихдля оценки опасности итоксичности 1, 4,,5,,6,,7,,8,,9,10., Клеточная линия A549 обладает соответствующими особенностями альвеолярных эпителиальных клеток типа II, такими как наличие характерных ламеллярных тел, содержащих плотно упакованные фосфолипиды3.
Было показано, что, когда клетки культуруются на воздухо-жидком интерфейсе (ALI), сурфактант высвобождается на апической стороне открытых воздухом эпителиальных клеток, уменьшая поверхностноенатяжение 11,,12,,13. Эта особенность особенно важна при исследованиях наноматериальной респираторной опасности и токсичности. После вдыхания наноматериалов / токсикантов откладываются в альвеолярной области, они сначала взаимодействуют с легочным сурфактантом и вытесняются смачивающими силами в аквеозную гипофаз, где взаимодействие с легочнымиклетками происходит 14,15. Несмотря на то, что A549 клетки образуют монослой (который может разрастаться в многослойных на более поздних точках времени при выращивании в ALI) и производить сурфактант, недостатком является их недостаточное плотное образование соединения, в результате чего низкие значения трансепителиальной электрической устойчивости, но все еще представляет функциональный барьер против межклеточных (нано) частицтранслокации 16,17,18.
В легких существует множество популяций иммунных клеток, включая фагоцитные и профессиональные антиген-представления клеток (т.е. макрофагов и дендритных клеток), которые непосредственно общаются через контакт клеток или межклеточный сигнал для контроля и поддержания гомеостаза. Макрофаги и дендритные клетки являются критическими врожденными иммунными эффекторами и инициаторами адаптивного иммунного ответа19. Дендритные клетки, проживающие внутри или под эпителием, могут образовывать выступы через эпителий к люмену, чтобы поймать антигены. Альвеолярные макрофаги расположены на апической поверхности эпителия и действуют как дозорные клетки, представляющие собой первую клеточную защиту от инордного материала, а также бактериальные, вирусные и грибковые инфекции. Их фенотипическая пластичность позволяет быстро индукции провоспалительных реакций в ответ на такие раздражители, а также переход в запуск противовоспалительных (т.е. ингибирующие)реакции 20.
Для имитации альвеолярного эпителиального барьера ткани человека, мы создали тройную модель совместной культуры с клетками A549, дополненными макрофагами, полученными из моноцитов человека (MDM) и дендритными клетками (MDDCs) на апиальной и базальной сторонах,соответственно 17. Культивирование этой модели на ALI ранеесообщалось 16, даже до 72 ч после экспозиции21. Острые иммунные реакции на воздействие углеродных нанотрубок были значительно усилены в клеточной культуре, подвергаемой воздействию АЛИ, по сравнению с подводнымиусловиями 22. Модель кокультуры, культурные и подвергаются воздействию различных материалов на ALI, ранее был использован для исследования цитотоксичности, окислительного стресса и воспалительных реакций при воздействииоксида цинка, 23 графена связанных материалов24, золотые наночастицы25,26, углеродные нанотрубки21, и вулканического пепла и дизельныхвыхлопных частиц 27.
Кроме того, была подтверждена важная роль макрофагов и дендритных клеток в качестве иммунных клеток-эффекторов в модели легких человека in vitro. В частности, повышенная провоспламеняемая реакция в модели наблюдалась только при наличии иммунных клеток по сравнению с монокультурнымисистемами 7. Потенциальными недостатками использования первичных моноцитов, полученных из иммунных клеток, являются ограниченная доступность ПБМ, а также вариации от донора к донору. В качестве решения этих потенциальных недостатков, представленный здесь протокол введения криоконсервации свежеиспеченногоPBMs 28 для сборки модели культуры клеток. Цель этого исследования состоит в том, чтобы продемонстрировать 3D человека альвеолярной эпителиальной ткани модели сборки, в том числе изоляции ПБМ от человека баффи пальто. Реагирование на провоспалительные стимулы сравнивается с моделью, состоящей из МДМ и МДДК, дифференцированной от свежих ПБМ или дифференцированной от замороженных/оттаяющих ПБМ.
Работа с непроверенными образцами крови человека включает в себя конкретную помощь для предотвращения потенциальной передачи инфекционных заболеваний, таких как ВИЧ (вирус иммунодефицита человека), гепатит В и гепатит С. Таким образом, использование мер индивидуальной защиты, таких как перчатки, платья, маски и защита глаз имеют решающее значение и должны быть в соответствии с принципами хорошей лабораторной практики. Эти средства защиты снижают риск обиболивания кожи или слизистых оболочек с потенциально инфекционными жидкостями. Кроме того, для тех, кто участвует в обработке баффи пальто и ПБМ, вакцинация против вируса гепатита В является обязательным, и уровень титры крови антител против гепатита В должны быть выше 100 МЕ / Л (страна конкретных законодательных требований должны быть рассмотрены). Кроме того, вся работа должна проводиться в лабораториях уровня биобезопасности 2 (необходимо решать законодательные требования, предъявляемые к конкретным стране). При проведении всего протокола должны быть приняты стандартные меры предосторожности для здоровья и безопасности, связанные с работой в лабораторных условиях и обработкой культуры клеток млекопитающих, включая обработку отходов.
протокол
Работа с участием первичных моноцитов, изолированных от человеческой крови, была одобрена комитетом Федерального управления общественного здравоохранения Швейцарии (справочный номер: 611-1, Meldung A110635/2) для Института Адольфа Меркл.
1. Изоляция периферических моноцитов крови (PBMs) от человека баффи пальто
ПРИМЕЧАНИЕ: В следующем разделе описывается изоляция иммунных клеток от одного 50 мл мешок баффи пальто, приобретенные в Швейцарском центре переливания крови в Берне, Швейцария.
- Приготовление реагентов
- Подготовьте 100 мл буфера магнитного разделения на баффи пальто: 0,5% «w/v» бычьего альбумина сыворотки (BSA; в фосфатно-буферном солевом растворе «PBS») с этилендиаминтетраатетической кислотой (EDTA) и приспособиться к рН 7,2, стерильному фильтру с размером 0,22 дюйма). Держите на уровне 4 градусов по Цельсию на протяжении всей процедуры.
- Подготовьте клеточную культуру среды (CCM): RPMI 1640 с 10% фетальной сыворотки крупного рогатого скота (FBS), 1% L-глатамин (здесь, 2 мММ Л-глутамин) и 1% пенициллин-стрептомицин (здесь 100 единиц/мл пенициллина и 100 мкг/мл стрептомицина).
ПРИМЕЧАНИЕ: Необходимое количество каждого реагента зависит от количества клеток, которые будут посеяны в следующих шагах.
- Изоляция ПБМ
ПРИМЕЧАНИЕ: Все стекло и пластмассовые изделия должны быть стерилизованы перед использованием. По соображениям безопасности, использование пластичных средств рекомендуется при обработке образцов крови человека, чтобы уменьшить риск получения травмы с помощью стеклянной посуды.- Используйте ножницы, чтобы разрезать шланг конце мешок, содержащий баффи пальто.
- Распределите баффи пальто, наливая содержимое сумки через проток сумки непосредственно в две конические центрифуги 50 мл труб (по 25 мл каждый).
- Аккуратно налейте или пипетки PBS в трубы, чтобы достичь 50 мл томов. Смешайте содержимое, повернув трубку слегка вверх дном 3x.
- Разделите смесь баффи пальто-PBS на четыре новых 50 мл конической центрифуги труб, трубя 25 мл смеси в каждой свежей трубке.
- Медленно положите 13 мл градиентной среды плотности под баффи пальто-PBS смеси с помощью 10 мл серологической пипетки. Отсоедините заполненную пипету от держателя пипетки и немедленно подключите верхнее отверстие пипетки большим пальцем, чтобы предотвратить любую дополнительную утечку градиентной среды плотности в баффи пальто-PBS смеси.
ПРИМЕЧАНИЕ: Удерживая верхнее отверстие большим пальцем, поместите заполненные пипетки в нижней части конической центрифуги трубки так, что плотность градиента среды медленно течет под баффи пальто-PBS смеси, в результате чего примерно 1 мл плотности градиента среды внутри пипетки. - Повторите шаг 1.2.5 с другими тремя трубками, содержащими баффи пальто-PBS смеси.
- Центрифуга всех четырех трубок, содержащих смеси в течение 20 минут при 1000 х г и 25 градусов по Цельсию в режиме медленного торможения. Используйте держатели с защитными крышками для центрифугации.
- Откройте крышку каждой трубки, удалите верхний слой, содержащий плазму и тромбоциты с помощью серологического пипетки, и утилизировать в контейнере для биологических жидких отходов.
- Используйте серологическую пипетку для сбора периферического слоя моноядерных клеток крови, который выглядит как беловатый мутный небольшой фракции (2-3 мм в толщину) между плазмой и плотностью градиент средних слоев. Гранулы содержат красные кровяные тельца в нижней части. Избегайте переноса эритроцитов, которые образуют самый нижний слой. Повторите это для всех четырех трубок.
ПРИМЕЧАНИЕ: Периферические моноядерные клетки крови состоят из ПБМ и лимфоцитов. PBMs будут отделены от лимфоцитов позже во время магнитного разделения CD14 ". - Бассейн периферических моноядерных клеток крови из четырех трубок в две трубки 50 мл.
- Заполните две трубки с PBS до 50 мл и накройте крышкой.
- Отбросьте остатки эритроцитов и плазмы из оригинальных четырех трубок в контейнере с жидкими отходами.
- Центрифуга две трубки в течение 8 мин при 500 х г и 18-20 градусов по Цельсию на регулярной скорости центрифуги.
- После центрифугации удалите супернатант серологическим пипеткой и выбросьте его в контейнер для биологических жидких отходов.
- Повторное использование клеток с 5 мл PBS с помощью серологического пипетки.
- Бассейн ячейки суспензии в один 50 мл конической центрифуги трубки и заполнить до 50 мл с PBS.
- Используйте 5 МКЛ клеточной подвески для подсчета ячеек с счетчиком ячеек с помощью метода исключения трипан-голубого (45 МКЛ).
- Pipette 10 йл из trypan сине-PBMs решение в камеру счетчика клеток и подсчитать количество ячеек в стандартном протоколе подсчета. Используйте Equation 1 для расчета общего числа ячеев, CT.

- После подсчета клеток, центрифуга 50 мл трубки, как это делается в шаге 1.2.13.
- CD14 положительный выбор
- Аккуратно откройте крышку каждой трубки, затем удалите и отбросьте супернатант с помощью серологического пипетки, не нарушая гранулы.
- Добавьте рассчитанную сумму (Equation 2) буфера магнитного разделения (здесь 80 МКЛ буфера на 1 х 107 общих ячеек) и повторно найдите гранулы ячейки, засовыв раствор вверх и вниз.

- Рассчитайте с помощью Equation 3 (здесь 10 йл на 1 х10 7 общих ячеек) соответствующий объем магнитных бусин CD14 и пипетки соответствующего объема.

- Хорошо перемешать, трубя вверх и вниз, закрыть крышку, и инкубировать раствор при 4 градусов по Цельсию в течение 15 мин.
- После инкубации заполните трубку до 50 мл магнитным буфером разделения.
- Центрифуга, как это делается в шаге 1.2.13.
- Аспирировать и отбрасывать супернатант с помощью серологического пипетки, не нарушая гранулы клетки.
- Pipette соответствующее количество магнитного буфера разделения (Equation 4; здесь, 500 йл буфера на 1 х 108 ячеек) и осторожно смешивать путем трубопроводов вверх и вниз 3x.

- Дезинфицировать магнитное отделение станции путем распыления и вытирая его с стерилизованным агентом. Поместите в капот ламинарного потока вместе с колонкой для магнитного разделения.
- Поместите магнитное разделительное столбец в магнитное поле и поместите пустую коническую центрифугу 50 мл трубки прямо под колонку для сбора промывки и неоцеливых клеток (т.е. отходов).
- Промыть магнитное разделение столбец путем трубопроводов 3 мл магнитного буфера разделения в столбец. Не позволяйте столбец высохнуть на протяжении всей процедуры.
- Подготовь 15 мл конической центрифуги трубки и пипетки 1 мл магнитного буфера разделения.
- Нанесите подвеску ячейки (подготовленную в шаге 1.3.8) к магнитному разделительному столбу. В 50 мл конической центрифуги трубки под фильтром, собирать неоцеличные клетки, которые прошли через.
ПРИМЕЧАНИЕ: Не превышать 2 х 109 ячеек на столбец, чтобы избежать блокировки столбца. - Как только резервуар столбца пуст (т.е. когда клетки прошли через столбец), нанесите 3 мл буфера магнитного разделения с помощью серологического пипетки и дайте ему пройти через столбец. Повторите это 3x.
- Удалите магнитный разделительный столб из магнитного сепаратора, аккуратно потянув руками, затем поместите его в трубку 15 мл, содержащую предварительно трубчатый 1 мл буфера магнитного разделения (подготовленный в шаге 1.3.12).
- Добавьте 5 мл буфера магнитного разделения к столбецу и смойте магнитно помеченные клетки, твердо толкая поршень в столбец.
- Подготовка реагентов к дифференциации МДМ и MDDC
- Подсчитайте ячейки с счетчиком ячеек, используя метод trypan blue exclusion, как это делается в шаге 1.2.17.
- Рассчитайте необходимые объемы СКК или FBS для дальнейших шагов следующим образом: либо объем СКК, соответствующий плотности ячейки 1 х 106 ячеек/мл (шаг 1.5.1), либо объем FBS, соответствующий плотности ячейки 6 х10 6 ячеек на 0,9 мл FBS (шаг 1.6.3).

- Закройте крышку, поместите трубку в центрифугу и центрифугу, как это делается в шаге 1.2.13. Удалите и отбросьте супернатант, не нарушая гранулы клетки. Приступайте к шагу 1.5 для посева клеток или шаг 1.6 для замораживания клеток.
- Посев и дифференциация ПБМ на МДМ и ЦРТ
- Повторное использование клеточной гранулы в расчетном объеме СКК, рассчитанном в шаге 1.4.2 (здесь, окончательная концентрация 1 x 106 ячеек/мл) путем трубопроводов вверх и вниз в 3 раза.
- Pipette количество клеток, предназначенных для дифференцирования в MDMs и MDDCs в отдельных конических трубок центрифуги с помощью серологического пипетки.
- Pipette дифференциации факторов СКК с ПБМ и хорошо перемешать, трубопроводов вверх и вниз. Дифференцировать факторы применяются следующим образом:
- Для MDDCs: окончательная концентрация 10 нг/мл интерлеукина-4 (IL-4) и 10 нг/мл гранулоцитов-макрофагов колоний-стимулирующих факторов (GM-CSF).
- Для МДМ: окончательная концентрация макрофагового фактора 10 нг/мл (M-CSF).
- Пипетты клеточной подвески в СКК с добавленными дифференцируя факторами в 6 пластин хорошо, распределяя 3 мл подвески на колодец (соответствует 3 х 106 ячеек / хорошо, т.е., 1 х 106 клеток / мл).
- Поместите 6 пластин хорошо в инкубатор культуры клеток (37 КК, 5% CO 2 ) и дайте имдифференцироватьв течение 6 дней без обновления СКК.
ПРИМЕЧАНИЕ: Дифференциация колеблется от 5 до 8 дней в зависимости от локальной доступности баффи пальто и экспериментальной установки, при условии, что эффективность дифференциации определяется с использованием соответствующих методов (ссылка на раздел обсуждения).
- Замораживание PBM
- Перепрофилировать клеточные гранулы в криопротекторной среде (здесь, FBS и диметилсуфсид »DMSO; цитотоксический») в соотношении 9:1 (v/v) путем трубчатого объема предварительной FBS. Это соответствует окончательной концентрации клеток 6 х 106 клеток/мл, учитывая дальнейшее добавление 10% DMSO (v/v).
- Отметь нужное количество криовилов в капоте ламинарного потока (т.е. завечете дату, код изоляции и количество ячеек).
- Pipette 0.9 mL клеточной подвески в чистом FBS (здесь, 6 x 106 клеток в 0.9 мл FBS) к каждому криовиалю. Впоследствии медленно пипетка 0,1 мл DMSO и смешать подвеску хорошо, повернув криовиалы вверх и вниз 3x.
- Перенесите криовиалы в контейнер для замораживания клеток и немедленно установите его до -80 градусов по Цельсию в течение 24 ч.
- После 24 ч снимите криовиалы из морозильной камеры и контейнера -80 градусов по Цельсию и поместите их в резервуар с жидким азотом, пригодный для хранения клеток.
- Таяние и дифференциация ПБМ на МДМ и MDDCs
- Разогреть все необходимые реагенты до 37 градусов по Цельсию на водяной бане (20-30 мин).
- Подготовь соответствующее количество из 6 пластин, соответствующих количеству оттаялых клеток (здесь одна пластина на 1,8 х 107 клеток, т.е. 3 криовиала). Pipette 2 мл СКК для каждого хорошо в асептических условиях. Поместите пластины в инкубатор (5 % CO2, 37 градусов по Цельсию) в течение 15 минут, чтобы рН эквивалентно.
- Возьмите необходимое количество криовилов с замороженными клетками из резервуара с жидким азотом и аккуратно закружить их в водяной бане 37 градусов по Цельсию (1-2 мин), чтобы обеспечить равномерное оттаивание клеточной подвески.
- Снимите криовиал с водяной бани и обеззараживайте стерилизатором, гарантируя, что агент не взаимодействует с крышкой и O-кольцом.
ПРИМЕЧАНИЕ: Таким образом, все шаги должны быть завершены в асептических условиях. - Подготовь соответствующее количество конических центрифуг 15 мл, соответствующее количеству криовилов, которые будут разморожены (здесь 6 х10 6 клеток/труб). Pipette 9 мл довоенной СКК в каждой трубке.
- Пипетт медленно (капля за каплей) содержимое криовиальной в трубку, содержащую СКК. Закройте крышку, повторите для каждой трубки, и центрифуга на 200 х г в течение 5 минут на регулярной скорости центрифуги.
- Откажитесь от супернатанта, не нарушая гранулы.
- Resuspend гранулы из каждой трубки (которая содержит клетки из одного криовиального) в 2 мл довоенной СКК путем трубопроводов вверх и вниз с помощью серологической пипетки (плотность клеток, соответствующих 3 х 106 клеток / мл).
- Из каждой трубки пипетка перерасходует клетки в две скважины (1 мл на скважину) из 6 скважин, содержащих 2 мл ранее подготовленного СКК, чтобы достичь плотности клеток 3 х 106 клеток/колодец (соответствующих окончательной концентрации 1 х 106 клеток/мл). Повторите это для всех других труб.
- Продолжить дифференциацию, описанную в разделе 1.5.3.
- Поместите 6 пластин хорошо в инкубатор культуры клеток (37 КК, 5% CO 2 ) и дайте имдифференцироватьв течение 6 дней без обновления СКК.
2. Тройная клеточная модель совместной культуры альвеолярной эпителиальной ткани человека
ПРИМЕЧАНИЕ: В этом разделе содержатся инструкции по томам и номерам ячеей, соответствующим 12 вставокпластинок. На рисунке 1 кратко излагается предлагаемая шкала времени сборки моделей.
- Эпителиальная клетка (линия клеток A549) посев
- Культура эпителиальных клеток в соответствии с рекомендациями, предоставленными поставщиком (ATTC). Короче говоря, субкультуры клеток в СКК на 80% клеточного слияния (примерно, 2x-3x в неделю).
ПРИМЕЧАНИЕ: Субкультура A549, по крайней мере четыре прохода до композиции модели кокультуры, используя A549 клеток в диапазоне прохода 5-25. - Пипетка 1,5 мл довоенного СКК в 12 скважин (количество скважин соответствует желаемому количеству моделей).
- Поместите отдельные 12 хорошо клеточной культуры вставки в колодцы 12 хорошо пластины с использованием стерилизованных пинцетом.
- Отсоедините клетки от колбы в соответствии с протоколом субкультивации (т.е. с помощью отсоединения агента удалите агент по центрифуге, как это делается в шаге 1.2.13). Ставки в соответствующем объеме СКК, соответствующие конечной концентрации клеток A549 (здесь, 50 х 104 клеток/мл; 0,5 мл клеточной подвески на вставку, т.е. 25 х 104 ячейки/вставки, что соответствует плотности посева 27,8 х 104 ячейки/см 2).
- Пипетка 0,5 мл клеточной подвески (т.е. 25 х 104 ячеек/вставки) в апиалятную сторону вставки с помощью 1 мл пипетки.
- Обложка пластины с крышками и поместить их в инкубатор культуры клеток (37 градусов по Цельсию, 5 % CO2) в течение 4 дней.
ПРИМЕЧАНИЕ: Регулярно проверяйте слияние клеток A549 под фазо-контрастным микроскопом.
- Культура эпителиальных клеток в соответствии с рекомендациями, предоставленными поставщиком (ATTC). Короче говоря, субкультуры клеток в СКК на 80% клеточного слияния (примерно, 2x-3x в неделю).
- Посев MDDC
- Аспират СКК с незакрепленными клетками в 6 пластинах, содержащих MDDCs.
- Добавьте 1 мл свежего довоенированного СКК к каждому колодец.
- Используйте клеточный скребок, отсоедините (отсоедините) адепта MDDCs от каждой скважины, аккуратно промыть колодцы с существующим 1 мл CCM 3x, и объединить их в одну коническую трубку центрифуги.
- Подсчитайте ячейки с счетчиком ячеек с помощью метода trypan blue exclusion с использованием 10 МКЛ клеточной подвески и 10 МКЛ трипан синего раствора.
- Центрифуга подвески клетки, как это делается в шаге 1.2.13.
- Рассчитайте объем СКК, необходимый для повторного успения (Equation 5):
Требуемая плотность MDDC 42 x 104 ячейки/мл; каждая вставка требует 6,3 х10 4 клеток, что соответствует плотности семенной клетки 7 х10 4 ячейки/см 2 (здесь, 150 йл добавил на 0,9см 2 в шаге 2.2.10.).
Объем СКК для повторного деления клеток (Vm; Уравнение 5):
- Аккуратно аспирировать и выбросить СКК из верхней камеры 12 хорошо пластин с ростом A549 на вставки.
- Поместите вставки с клетками A549 в перевернутом положении в стерильную чашку Петри с помощью стерилизованных пинцетов. Подготовь коническую центрифугу трубку (50 мл) с ПОМОЩЬЮ PBS и предварительно увлажнить клеточный скребок.
- Очистите клетки A549 от базальной поверхности вставки (т.е. верхней части в перевернутом положении), которая должна расти через поры вставок.
ПРИМЕЧАНИЕ: Промыть скребок с PBS (подготовлен в трубке) между соскоб отдельных образцов и держать его мокрым на протяжении всей процедуры. - После центрифугации (шаг 2.2.5), аспирировать и отказаться от супернатанта, а затем redisperse MDDC гранулы в расчетном количестве СКК (шаг 2,2,6) и пипетки вверх и вниз 3x.
- Пипетка 150 МКЛ клеточной подвески поверх каждой вставки так, чтобы вся базальная поверхность вставки поровну покрывалась жидкостью и не содержала пузырьков.
- Накройте блюдо крышкой и поместите в инкубатор клеточной культуры на 70 минут. Аспирировать СКК из пластин клеточной культуры (где вставки первоначально были помещены), выбросить его в биоопасных жидких отходов, и пипетки 1,5 мл свежих СКК в каждой хорошо. Накройте пластину крышкой и поместите в клеточный инкубатор (37 градусов по Цельсию, 5% CO2).
ПРИМЕЧАНИЕ: Не превышать период времени, упомянутый выше, чтобы избежать высыхания клеток. - После инкубации тщательно держите каждую вставку с стерилизованным пинцетом и поместите их на пластины, содержащие СКК, в обычном положении. Обложка пластины с крышкой и вернуть его в клеточный инкубатор (37 градусов по Цельсию, 5% CO2).
- Посев макрофагов (МДМ)
- Возьмите 6 хорошо пластин, содержащих предварительно дидифференцированных MDMs. Поместите их из клеточного инкубатора в ламинарный капюшон потока.
- Аспирировать и отбрасывать СКК с неприсоединения MDMs, выращенных в 6 пластин хорошо, и пипетка 1 мл свежих довоенных СКК в каждой хорошо.
- Используя сотовый скребок, аккуратно удалите адептов MDM из отдельных скважин (как это делается в шаге 2.2.3 для MDDCs).
- Pipette 10 йл трипан синий в колодец или трубку и добавить 10 МКЛ подвески МДМ для достижения окончательного разбавления 1:1 (v/v). Подсчитайте количество MDM с помощью соответствующего протокола подсчета.
- Центрифуга подвески клетки, как это делается в шаге 1.2.13.
- Рассчитайте необходимый объем (Equation 6):
Требуемая плотность МДМ 2,5х 10 4 ячейки/мл в СКК (здесь каждая вставка требует 1,25 х10 4 ячейки в 0,5 мл СКК, что соответствует плотности семенной клетки 1,4 х 104 ячейки/см2). (6)
(6) - После центрифугации, аспирата и отбрасывания супернатанта, переделайте гранулы МДМ в расчетном количестве СКК (шаг 2.3.6), и пипетку вверх и вниз в 3 раза.
- Тщательно пипетка 0,5 мл подвески МДМ (подготовлена в шаге 2.3.7) на стене клеточной культуры вставляется с A549 и MDDCs (не непосредственно на эпителиальных клетках) с помощью 1 мл пипетки. Обложка пластин с крышками и место в инкубаторе культуры клеток (37 градусов по Цельсию, 5 % CO2) для 24 ч.
- Перевод модели совместной культуры в воздушно-жидкий интерфейс (ALI)
- По окончании инкубационного периода в 24 ч (2 ч) собранной модели в инкубаторе клеточной культуры, аспират и отбрасывание СКК как из апических, так и из базальных частей клеточной культуры вставки и из колодцев.
- Используя стерилизованные пинцеты, поднимите отдельные вставки из колодцев и пипетку 0,6 мл свежей предварительной СКК к каждой скважине с помощью 1 мл пипетки. Не добавляйте СКК в апическую сторону вставки.
- Обложка пластин с крышками и место в инкубаторе клеточной культуры (37 КК, 5 % CO2) для 24 ч до дальнейшего использования.
3. Воздействие отдельных положительных элементов управления (известные стимулы для индуцирования провоспалительных реакций)
ПРИМЕЧАНИЕ: Воздействие моделей кокультуры на известный провоспалительный стимул эндотоксина липополисахарид (LPS)7 и провоспалительный фактор некроза опухоли цитокинов No (TNF-Q)7 используется для иллюстрации отзывчивости модели. Кроме того, воздействие моющего средства (Triton X-100) используется для подтверждения чувствительности лактат дегидрогеназы (LDH) анализа.
- Подготовье положительных контрольных решений: запасы LPS (1 мг/мл в дистиллированной воде), запасы ТНФ-З (100 мкг/мл в дистиллированной воде) и Тритон-Х 100 (2% в ПБС).
- При инкубации модели coculture на 24 ч в условиях ALI, аспирировать и выбросить супернатант из базального отсека. Используя стерилизованные пинцеты, поднимите отдельные вставки из колодцев и пипетку 0,6 мл свежего довоенного СКК в каждую скважину.
- Подготовка индивидуальных положительных решений контроля рабочих путем разбавления запасов в СКК в конической центрифуге труб следующим образом: 1 мкг / мл LPS, 1 мкг / мл TNF-З, и 0,2% Тритон-X 100. Объемы соответствуют количеству проверенных вставок (здесь 100 йл/вставки). Смешайте растворы хорошо, трубя вверх и вниз 3x.
- Нанесите 100 мкл каждого положительного раствора управления, медленно пипетки на стену вставки культуры клеток. Обложка хорошо пластины с крышкой и место в инкубаторе культуры клеток (37 градусов по Цельсию, 5% CO2) для 24 ч. После инкубации, аспирировать и отказаться от жидкости на апической стороне вставки, удерживая отдельные вставки с помощью пинцета.
- Соберите СКК в базальных отсеках и храните при 1) 4 кк для дальнейшего анализа LDH, обозначая разрыв клеточной мембраны опосредованную цитотоксичность и/или 2) хранить при -80 градусов по Цельсию для дальнейшего анализа высвобождения белка с помощью ферментно-связанного иммуносорбентного анализа (ELISA). Вы запустите анализы в соответствии с рекомендациями поставщика комплекта.
- После удаления СКК, мыть вставки с PBS 3x и исправить клетки на клеточной культуры вставки в 4% "W / V" параформальдегид (в PBS, 15 мин при комнатной температуре), обеспечивая обе стороны вставки хорошо покрыты раствором PFA. Впоследствии мыть 3x с PBS, чтобы удалить PFA. Храните образцы, погруженные в PBS при 4 кк для дальнейшего иммуностимления (пример этого метода был описан ранее17).
Результаты
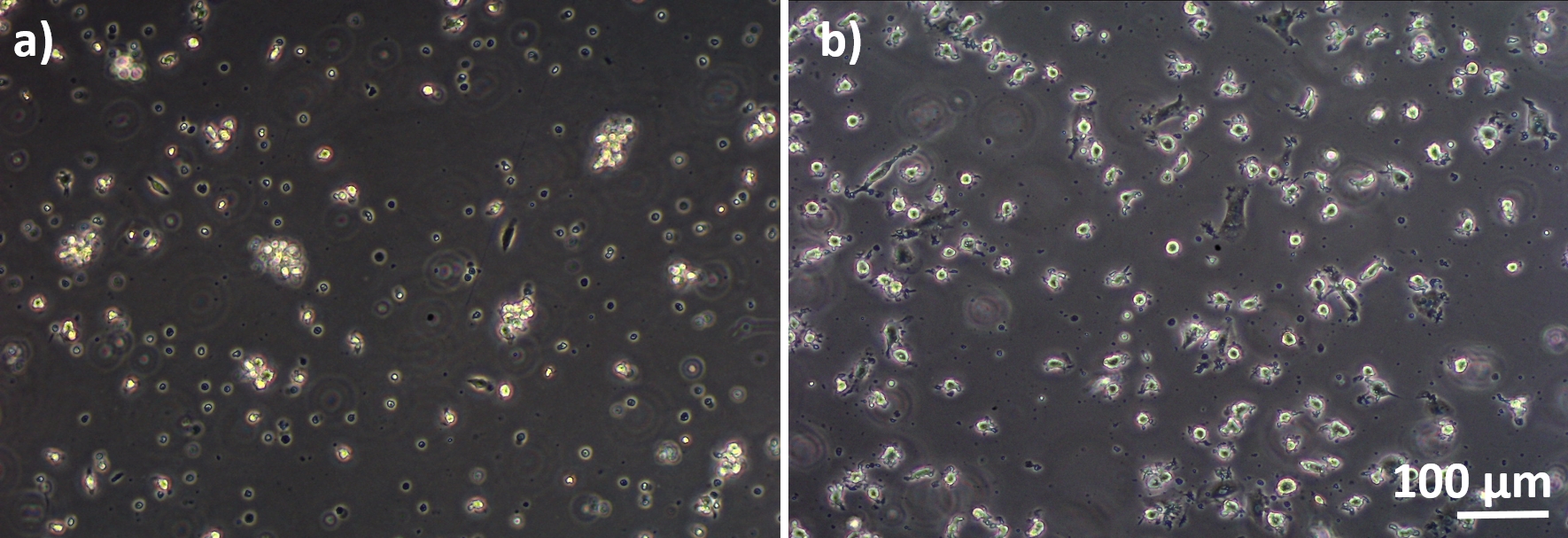
Модели кокультуры легких человека, состоящие из альвеолярных эпителиальных клеток и иммунных клеток, были собраны либо из свежих или замороженных MDDCs и MDMs прародителей (здесь, человека периферических кроветворных моноцитов). Как по представлено на рисунке 1, A549 клетки были посеяны через 3 дня после первого раздела с участием моноцитов изоляции / оттаивания. После 6 дней дифференциации, дифференцированные MDMs появились круглой формы, в то время как MDDCs сформировали более удлиненную форму с наблюдаемыми выступами. Они также появились в качестве агломератов, особенно когда отличаются от свежих моноцитов(рисунок 2, рисунок 3). Эпителиальные клетки образуют плотный клеточный слой клеток после 3 дней роста на мембранныхвставок (рисунок 4), когда кокультуры были собраны. После 24 ч сборки и еще 24 ч, которые подверглись условиям АЛИ, кокультуры были подготовлены для экспозиции.
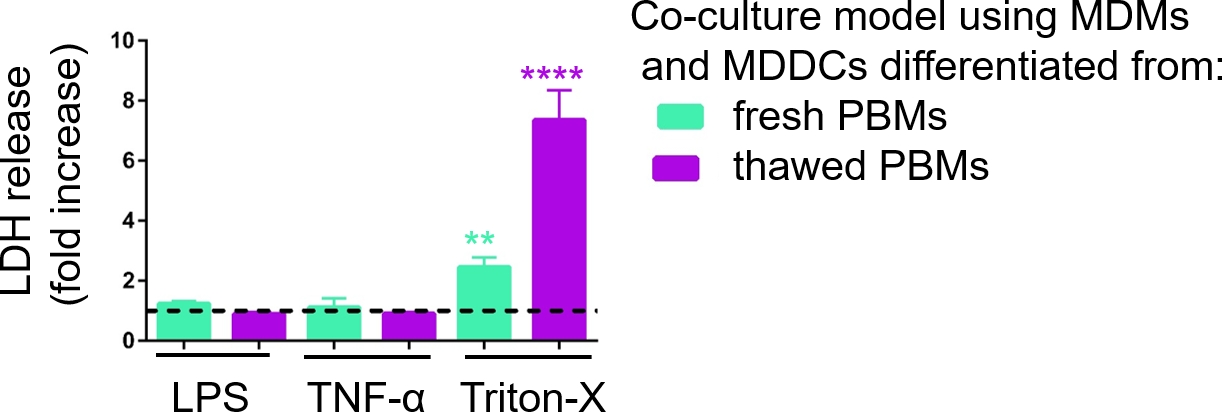
Ответственность моделей культуры 3D клеток была исследована при воздействии известных провоспалительных стимулов с использованием псевдо-ALI подход, как описаноранее 29. Провоспламеняемые стимулы, LPS и TNF-q, были добавлены в низких объемах (100 л) на апкическую поверхность модели клеток, подвергшихся воздействию воздуха. Параллельно с анализом LDH оценивалось отсутствие разрыва мембраны в качестве меры цитотоксии. Значительное увеличение выброса LDH в СКК базального отсека наблюдалось при воздействии положительного контроля при разрыве мембраны моющего средства Triton-X 100(рисунок 5). Эти результаты доказали отзывчивость модели к цитотоксическому веществу, в то время как никакого увеличения выброса LDH не наблюдалось при апической стимуляции с помощью TNF-q или LPS.
Возможная причина различных измеренных значений ЛДГ в образцах, собранных со свежими или ранее замороженными ПБМ, может быть отнесена к хранению образцов. Образцы из свежих ПБМ хранились в течение более длительного времени при -80 градусов по Цельсию; поэтому активность фермента LDH может снизиться. Примечательно, что LDH стабилен только до 4 дней в СКК; таким образом, рекомендуется выполнить анализ последних 2 дней после сбора супернатантов. Кроме того, можно заморозить супернатанты сразу после сбора. Тем не менее, важно учитывать, что замораживание может уменьшить энзиматической активности LDH.
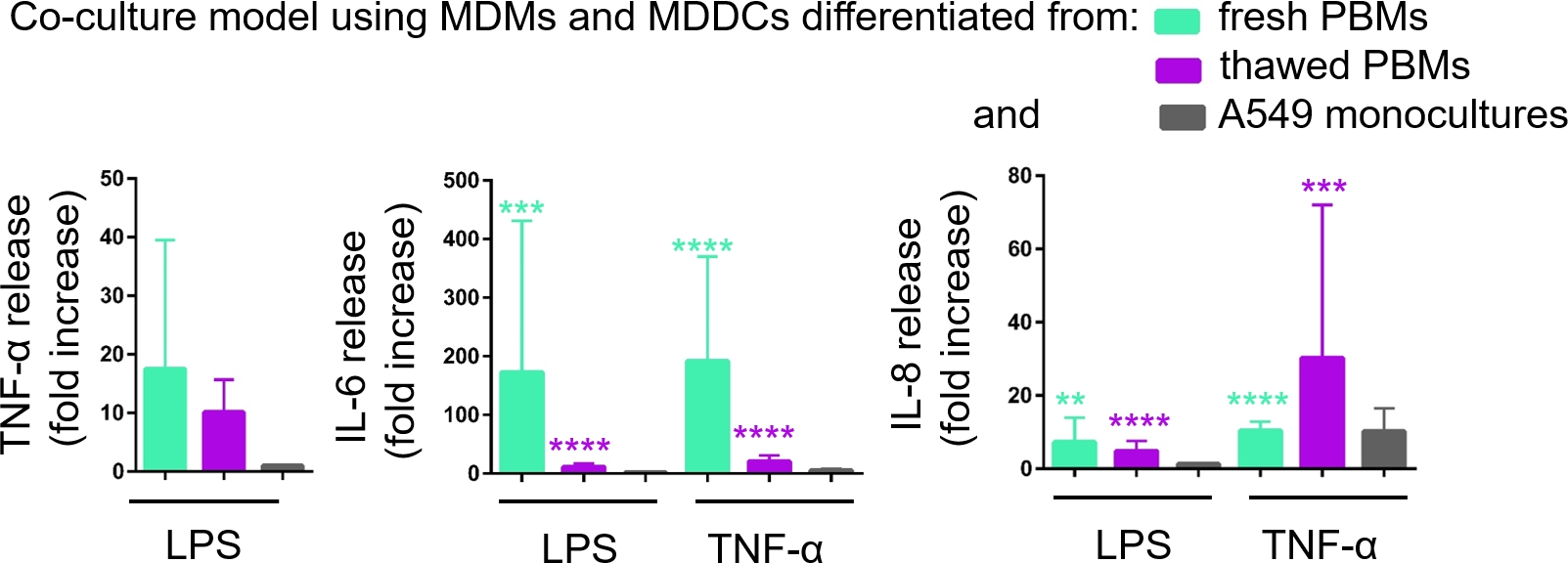
Через ELISA была количественно определена секреция провоспламеняющихся посредников (здесь ТНФ-я и интерлейкины 6 и 8 Ил-8) в базальный СКК. Статистически значимое (p lt; 0.05, в одну сторону ANOVA) увеличение выпуска IL-6 и IL-8 наблюдалось как в LPS- и TNF-- обработанных образцах по сравнению с соответствующими необработанными клетками, так и в моделях клеточной культуры, собранных из любого источника PBMs(рисунок 6). Хотя концентрации (пг/мл) всех проверенных цитокинов в базальных СКК были выше в кокультурах, состоящих из свежих ПБМ, различия между двумя кокультурами и монокультурами не были статистически значимыми (стр. 0,05) (Рисунок 6). Чтобы подтвердить добавленную стоимость моделей совместной культуры в отношении 2D эпителиальной клеточной культуры, монокультуры A549 также подверглись воздействию LPS или TNF-Q. Как и ожидалось, выпуск всех исследуемых посредников из монокультур A549 был ниже по сравнению с обеими моделями совместной культуры; хотя, разница между ними не была статистически значимой (стр. 0,05, в одну сторону ANOVA).
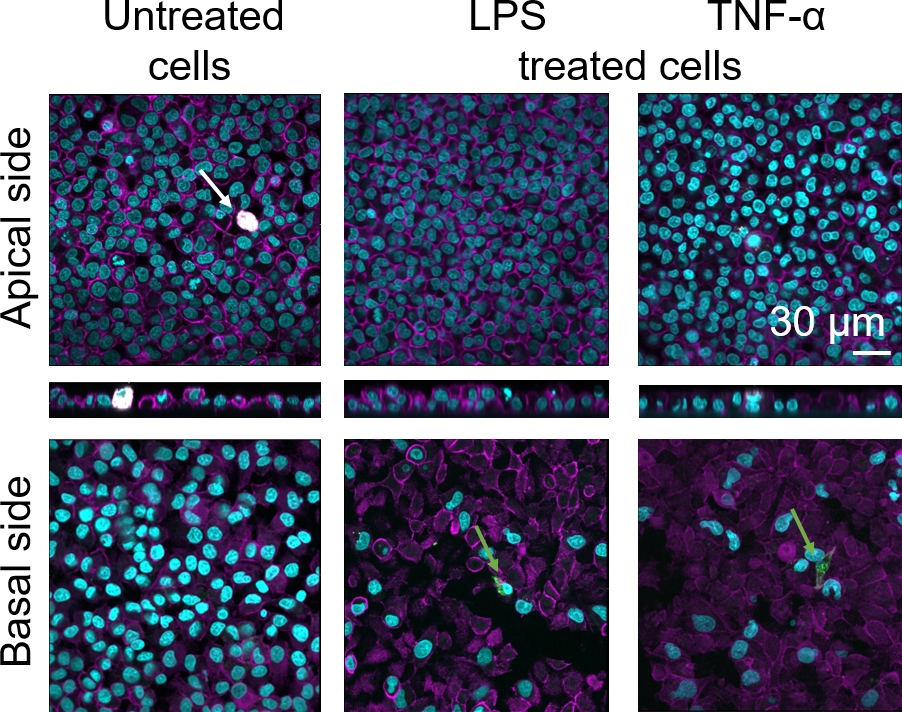
Клеточная морфология 3D альвеолярного эпителиального барьера ткани человека оценивалась с помощью конфокальной лазерной сканирующей микроскопии (LSM). Чтобы визуализировать состав каждой модели, макрофаги в рамках моделей совместной культуры (MDM) были окрашены зрелым маркером макрофага 25F9. MDDCs были окрашены CD83, который является важным маркером для активированных дендритных клеток30. Что касается клеточной морфологии, то никакой разницы между моделями совместной культуры с использованием МДМ и MDDCs от свежих ПБМ по сравнению с теми, которые используют оттаявые ПБМ, не наблюдалось. В lpS- и TNF--открытых кокультурах, как состоящих из свежих и замороженных иммунных клеток, был замечен нарушенный эпителиальный слой в изображениях LSM, чего не было в необработанных клетках(рисунок 7, Рисунок 8).

Рисунок 1: Схематическая хронология протокола. Презентация подготовки, сборки и применения модели 3D-кокультуры (воздействие испытанного вещества). АЛИ - воздушно-жидкий интерфейс, MDDCs - моноциты, производные дендритные клетки, МДМ - моноциты, макрофаги, ПБМ - периферические моноциты крови. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 2: ЦРТ и MDDCs отличаются от свежих ПБМ. Фазово-контрастное микроскопия изображения дифференцированных(A) MDMs и(B) MDDCs от свежих PBMs (6 дней после изоляции клеток). ЦРТ имеют круглую форму, в то время как MDDCs часто наблюдаются как агломераты. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 3: MDMs и MDDCs дифференцируются от замороженных ПБМ. Фазово-контрастное микроскопия изображения дифференцированных(A) MDM и(B)MDDCs от оттаялих ПБМ (6 дней после оттаивания). MDMs круглой формы, но некоторые удлиненные клетки могут наблюдаться. MDDCs также появляются круглой формы с выступами. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 4: Эпителиальные клетки роста на мембранных вставок. Фазово-контрастное микроскопия изображения стеченного A549, растущего на мембранной вставке через 4 дня после посева, образует плотный слой клеток. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 5: Результаты цитотоксичности исследованы с помощью анализа разрыва мембраны (LDH). Данные представлены как увеличение створки по сравнению с необработанными клетками (средний й SD, n No 3, звездочка обозначает статистически значимое увеличение по сравнению с необработанными клетками, q p lt; 0.01, йоут; 0.0001). В зеленых моделях представлены MDM и MDDCs из свежих ПБМ, а в фиолетовых моделях представлены собранные из размороженных ПБМ. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 6: Провоспламеняемые реакции в кокультурах и монокультурах. Провоспламеняемые посредники (ТНФ-З, Ил-6 и Ил-8) выпускают в кокультурах на 24 ч вызов с LPS или TNF-З. Данные представлены по отношению к необработанным ячейкам (средний - SD, n No 3, хт; 0,01, йоут; 0,001, йоут; 0,0001). В зеленых моделях представлены MDM и MDDCs из свежих ПБМ, а в фиолетовых моделях представлены собранные из размороженных ПБМ. Серый цвет представляет монокультуры A549. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 7: Морфология кокультур, состоящих из свежих иммунных клеток. LSM изображения апических и базальных сторон модели кокультуры с xz проекции апических сторон модели с использованием MDMs и MDDCs из свежих PBMs. Cyan представляет ядра (DAPI), пурпурный представляет цитоскелет (родамин-фаллоидин), белый представляет MDMs (25F9), а зеленый представляет MDDCs (CD 83). Белая стрелка обозначает МДМ, в то время как зеленая стрелка обозначает MDDC. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 8: Морфология кокультур, состоящих из замороженных иммунных клеток. LSM изображения апической стороны модели кокультуры с соответствующими проекциями xz, и базальная сторона модели с использованием MDMs и MDDCs от оттаялого PBMs. Cyan представляет ядра (DAPI), пурпурный представляет цитоскелет (родамин-фаллоидин), белый представляет MDMs (25F9), а зеленый представляет MDDCs (CD 83). Белая стрелка обозначает МДМ, в то время как зеленая стрелка обозначает MDDC. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}
Обсуждение
Новое производство новых материалов, включая химические вещества и наркотики, постепенно увеличивает потребность в прогностический in vitro моделей. В соответствии с тремя принципами замены, сокращения иуточнения тестирования на животных 32, в пробирке модели клеток стали мощными инструментами в отношении замены и сокращения аспект для выяснения механизмов действия препарата или материала8,9,10,11. Здесь представлен подробный протокол сборки многоклеточной модели с использованием иммунных клеток, которые либо недавно изолированы, либо оттаяли от ранее замороженных моноцитов. Также описано культивирование модели в ALI. Наконец, протокол иллюстрирует пример воздействия провоспалительных стимулов и сравнивает реакцию двух моделей, содержащих либо свежие, либо замороженные моноциты.
Различные исследования были проведены, чтобы подтвердить и оправдать добавленную стоимость повышенной сложности моделей, выращенных и подвергшихся воздействию в условиях АЛИ по сравнению с обычными погруженныхвоздействия 7,22,31. Наблюдение за более высокой провоспламеняемой реакцией в кокультурах по сравнению с монокультурами эпителиальных клеток подтверждает предыдущее исследование. В исследовании использовалась представленная модель кокультуры (стимулируемая LPS) и показана более высокая реакция на уровни экспрессии генов TNF и IL1B по сравнению с монокультурной эквивалентной моделью A5497. С другой стороны, обе модели показали более высокие различия в измеренных значениях выброса провоспламеняемого посредника по сравнению с монокультурами A549. Это можно объяснить использованием иммунных клеток от различных доноров (баффи пальто) в биологических повторений (т.е. одно повторение, один донор), как ранеепоказано 7. При желании различия между репликациями могут быть преодолены на 1) с помощью разморожевания ПБМ от одного и того же донора или 2) объединения ПБМ от различных доноров до замораживания клеток, а затем последующего использования одного и того же пула в каждом повторении. В том числе более биологических повторений также рекомендуется.
Техника замораживания клеток может рассматриваться как критический шаг; однако, это обычная лабораторная процедура для сохранения клеток для фенотипического и функционального анализа. Различные исследования показали, что качество замороженных ПБМ имеет жизненно важное значение для их выживания, и соответствующий метод замораживания является ключом к успеху последующих анализов с темиже клетками 28,32. Модификация протокола может быть выполнена путем замораживания ПБМ, что обеспечивает гибкость в экспериментальной установке, так как доступность баффи пальто, как правило, ограничена. Еще одним преимуществом использования замороженных ПБМ (в нескольких флаконах) по сравнению со свежеисплиенными является то, что они могут быть использованы в последующих экспериментах даже после 1 года. Это уменьшает потенциальную проблему изменчивости от донора к донору, если это желаемый или необходимый параметр в экспериментальном эксперименте.
Результаты межлабораторного сравнения, выполненного после 13 месяцев, показывают, что ПБМ, при правильном хране в резервуаре с жидким азотом, могут использоваться в течение длительного периода без какого-либо влияния на жизнеспособность клеток или восстановлениеклеток 33. Более длительное время хранения (более 1 года) может быть возможным при тщательной проверке жизнеспособности клеток и реакции клеток перед выполнением эксперимента. Кроме того, температура в резервуаре с жидким азотом должна оставаться стабильной в все времена. Основным фактором, влияющим на жизнеспособность криоконсервированных ПБМ, была обнаружена концентрация ДМСО с оптимальной концентрацией 10%-20% (v/v)28. Чтобы свести к минимуму потенциально вредные последствия замораживания, различные источники белков, FBS или BSA (с широким диапазоном концентрации от 40% до 100 %34) часто добавляются в замораживание среды в качестве естественных защитных компонентов, которые могут увеличить выживаемость клеток.
В связи с высоким цитотоксическим потенциалом ДМСО рекомендуется сначала разогнать ПБМ в FBS, а затем добавить DMSO в PBMs, уже рассеянные в FBS. Примечательно, что, несмотря на более высокие концентрации FBS (nogt;40%) не показывают никакого улучшения жизнеспособности клеток, в то же время, они не причиняют вреда клеткам28. Тем не менее, замораживание моноцитов является возможным подходом к преодолению проблем ограниченной доступности баффи пальто. Однако, если использование MDDCs и MDMs от свежих PBMs желательно, иммунные клетки могут быть дифференцированы и использованы 5-8дней после изоляции 7,,16,,17,,35,,36,37. Если экспериментальное планирование позволяет, рекомендуется по крайней мере 6 дней дифференциации как в MDDCs, так и в MDM. Тем не менее, согласованность между различными повторениями в одном и том же эксперименте, наряду с рутинными проверками их конкретных выражений маркера поверхности, имеют решающее значение. Реагирование на провоспламеняющие стимулы, такие как LPS, после дифференциации время также должны быть регулярно проверены.
Многие исследования с использованием линии клеток A549 были проведены в ALI, либо в качестве монокультуры или в сочетании с другими типами клеток (макрофаги, дендритные клетки, или фибробласты) в 3D-моделькокультуры 22,24,29,38. Используя эту 3D модель кокультуры, цитотоксичность, окислительный стресс, или провоспламеняющий эффект (нано-) материалы были исследованы до 72 ч1,17,21,24,29. Сходство модели с тканью in vivo ранее исследовалось на основе конфокального лазерного сканирования изображения модели16. При сборке модели важно учитывать как пролиферацию клеток (которая может повлиять на A549 в представленной здесь модели), так и производительность первичных (не размножающихся) иммунных клеток (здесь MDDCs и MDMs). Важно также учитывать, что не все положительные моноциты CD14 дифференцируются на MDDCs и MDM, и что клетки могут присутствовать как в прикрепленных, так и в подвешенных формах. Основываясь на природе сборки кокультуры (здесь оба типа клеток должны прикрепляться к существующему эпителиальный слой), рекомендуется использовать только приверженцев под популяций обоих типов иммунных клеток. Кроме того, рутинный анализ моноцитов, монокультуры MDDC и MDM, реагирующих на LPS, и выражение конкретных поверхностных маркеров (CD14, CD163, CD86, CD93 или CD206, данные не показаны) показали, что 6 и 7 дней дифференциации являются оптимальными точками времени.
Несмотря на то, что реальное количество альвеолярных эпителиальных клеток в легких человека соответствует 160 000клеток/см 2,количество клеток A549, подсчитанных вмодели,составляет 1 000 000 клеток/см2 после 9 дней, культурных навставке 16,18. Таким образом, необходимо учитывать ограничения этой модели in vitro. Во-первых, плотность эпителиальных клеток была установлена на основе их способности образовывать стечение слоя на растущей мембране. Важно также отметить, что A549 представляет собой эпителианные клетки типа II с кубоидной формой, в отличие от эпителиальных клеток типа I, которые являются плоскими и перепрочитанными. С другой стороны, необходимое количество иммунных клеток было установлено на основе литературы и представлено в этом протоколе как номерклетки/площадь поверхности 39,,40,,41. Плотность клеток MDDCs в диапазоне 400 клеток/мм2 (4 клетки/см2)16 сопоставима с плотностью клеток стабильного состояния 500-750 клеток/мм2 (5- 7 клеток/см2),о чем сообщается в исследованиях in vivo39. Плотность MDMs в этой модели находится в пределах того же диапазона in vivo ситуации в области альвеолярных человека40.
Зрелое окрашивание макрофагового маркера (25F9) наблюдалось как в апической стороне (где присутствуют МДМ), так и в базальной стороне (т.е. в месте дендритных клеток). Транслокация иммунных клеток через мембрану вставляет поры возможно, а также наблюдается спомощью этой модели 16, которые могут объяснить наблюдаемые различия в интенсивности окрашивания. Тем не менее, другое возможное объяснение заключается в том, что зрелый маркер макрофагов также может быть выражен на дендритных клетках, но выражение является весьма доноромконкретных 42. Кроме того, интенсивность выражения 25F9 гораздо выше в MDMs(рисунок 7, рисунок 8). Оба провоспалительных стимула (LPS и TNF-я) повлияли на целостность легочного эпителиального барьера в обеих кокультурах(рисунок 7, рисунок 8). Это было ожидаемо на основе предыдущихпубликаций 43,44, показывающих, что провоспалительные цитокины и бактериальные продукты нарушают целостность эпителиальных барьеров.,
3D многоклеточная модель альвеолярного эпителия человека, установленная ихарактерная ранее 17, служила мощным и полезным инструментом для оценки биологических реакций (т.е. острых провоспламеняющих реакций, окислительной реакции стресса, распределения частиц и сотовой связи) впробирке21,24,,25,,45. Результаты подтверждают ответственность моделей совместной культуры за провоспалительные стимулы (здесь, LPS и TNF-я). Реакция была несколько увеличена при использовании иммунных клеток от свежих ПБМ; однако статистически значимой разницы между кооперативами, использующими свежие и размороженные ПБМ, не было. Кроме того, провоспламеняемые реакции обеих моделей кокультуры были выше, чем реакции монокультур эпителиальных клеток, культивируемых в одинаковых (АЛИ) условиях. Таким образом, протокол описывает сборку 3D человека альвеолярной эпителиальной ткани совместной модели с использованием либо свежие или оттаяли ПБМ для дифференциации в MDMs и MDDCs. Показано, что обе модели в значительной степени реагируют на провоспалительные стимулы; поэтому они могут служить мощными инструментами для оценки потенциальной опасности и токсичности.
Раскрытие информации
Авторов нечего раскрывать.
Благодарности
Авторы хотели бы поблагодарить д-ра Мигеля Спуха-Гольвара за схему совместного ства на рисунке 3 и д-ра Бедию Бегум Каракочек за критическое чтение. Это исследование было поддержано проектом PATROLS, Программой исследований и инноваций Европейского союза Horizon 2020 в рамках грантового соглашения No 760813 и Фондом Адольфа Меркле. B.D. благодарит фонд Питера и Траудла Энгельхорна за финансовую поддержку.
Материалы
| Name | Company | Catalog Number | Comments |
| Benchmark microplate reader | does not have to be specific, for example BioRad, Cressier, Switzerland | ||
| Cell culture Incubator | does not have to be specific | ||
| Cell freezing container (for example Mr. Frosty) | does not have to be specific | ||
| Centrifuge | does not have to be specific | ||
| Confocal laser scanning microscope | does not have to be specific, for example | Zeiss LSM 710 meta | |
| Heamatocytometer, or automatic cell counter | does not have to be specific | ||
| Laminar bio-safety hood class II | does not have to be specific | ||
| MultiStand Macs (Macs Cell Separator) | Miltenyi, Germany | 130-042-303 | |
| pH meter | does not have to be specific | ||
| Phase contrast inverted light microscope | does not have to be specific | ||
| Pipette boy, pipettors (different volumes) | do not have to be specific | ||
| Scissors | do not have to be specific | ||
| Vacuum pump | does not have to be specific | ||
| Water bath | does not have to be specific | ||
| Disposable small equipment/glassware | Catalogue Number | ||
| 15 mL and 50 mL conical centrifuge tubes | does not have to be specific | ||
| 6- and 12-well cell culture plates, flat bottom, low evaporation lid, sterile | Falcon, Switzerland | 353046 and 353043 | |
| Cell culture inserts, transparent PET membrane, 12-well, 3 μm pore size | Falcon, Switzerland | 353181 | |
| Cell scrapper | does not have to be specific, for example VWR, Switzerland | 353085 | |
| Cryovials | do not have to be specific | ||
| Glass autoclaved Petri Dishes | do not have to be specific | ||
| LS Columns | Miltenyi, Germany | 130-042-401 | |
| Sterile filtration cup for vacuum filtration, 0.2 μm pore size | does not have to be specific, for example VWR, Switzerland | 10040-446 | |
| Sterile Lab Bottle compatible with Filtration cup (min. 100 mL) | does not have to be specific | ||
| Sterile pipettes | do not have to be specific | ||
| Chemicals | |||
| Bovine serum albumine (BSA) | Sigma-Aldrich, Switzerland | A7030-100g | |
| CD14+ MicroBeads human - magnetic beads | Miltenyi, Germany | 130-097-052 | |
| Deattachnig agent Trypsin-EDTA, 0.05%, phenol red | Gibco, Switzerland | 25300054 | |
| Density gradient medium Lymphoprep | Alere Technologies AS, Norway | 1114547 | |
| Dimethyl Sulfoxide (DMSO) | Sigma Aldrich, Switzerland | D2438 | |
| Ethylenediaminetetraacetic acid (EDTA) | Sigma-Aldrich, Switzerland | E6758-100g | |
| Fetal bovine serum (heat inactivated) | Gibco, Switzerland | 10270-106 | |
| Human granulocyte-macrophage colony-stimulating factor (GM-CSF), premium grade | Miltenyi, Germany | 130-093-864 | |
| Human Interleukin 4 (IL-4), premium grade | Miltenyi, Germany | 130-095-373 | |
| Human macrophage colony-stimulating factor (M-CSF), premium grade | Miltenyi, Germany | 130-096-485 | |
| L-glutamine | Gibco, Switzerland | 25030-024 | |
| Lipopolysaccharid (LPS) from Escherichia coli | Sigma-Aldrich, Switzerland | 4524-5mg | |
| Paraformaldehyde (PFA) | Sigma-Aldrich, Switzerland | 158127 | |
| Penicilin-Streptomycin | Gibco, Switzerland | 15140-122 | |
| Phosphate Buffer Saline (PBS) | Gibco, Switzerland | 10010-015 | |
| Roswell Park Memorial Institute-1640 Medium (RPMI) | Gibco, Switzerland | 42401-018 | |
| Triton X-100 | Sigma-Aldrich, Switzerland | T8787 | |
| Trypan blue solution (0.4%) | Sigma Aldrich, Switzerland | ||
| Tumor necrosis factor alpha (TNF-α) | Immunotools | 11343015 | |
| Assays used for cytotoxicity, (pro-)inflammatory response | |||
| Cytotoxicity Detection Kit (LDH) | Roche, Switzerland | 11644793001 | |
| Human IL-6 DuoSet ELISA | R&D, Biotechne, Switzerland | DY206 | |
| Human IL-8/CXCL8 DuoSet ELISA | R&D, Biotechne, Switzerland | DY208 | |
| Immunostaining | |||
| 4′,6-diamidino-2-phenylindole (DAPI), concentration 2 μg/mL | Sigma-Aldrich, Switzerland | 10236276001 | |
| Goat anti-mouse IgG (H+L) Alexa Fluor 647 conjugated, concentration 20 μg/mL | Abcam, UK | ab150115 | |
| Goat anti-rabbit IgG antibody (H+L) Dylight 488 conjugated, concentration 10 μg/mL | Agrisera, Sweden | AS09 633 | |
| Mature Macrophage Marker Monoclonal Antibody, concentration 50 μg/mL | eBioScience, Thermo Fischer, Switzerland | 14-0115-82 | |
| Phalloidin rhodamine, concentration 0.264 µM | Molecular Probes, Life Technologies, Switzerland | R415 | |
| Recombinant Anti-CD83 antibody, 1:50 dillution | Abcam, UK | ab244204 |
Ссылки
- Rothen-Rutishauser, B., Blank, F., Mühlfeld, C., Gehr, P. In vitro models of the human epithelial airway barrier to study the toxic potential of particulate matter. Expert Opinion on Drug Metabolism and Toxicology. 4 (8), 1075-1089 (2008).
- Giard, D., et al. In vitro cultivation of human tumors: establishment of cell lines derived from a series of solid tumors. Journal of National Cancer Institute. 51 (5), 1417-1423 (1973).
- Ochs, M., Weibel, E. R., Grippi, M. A. Ch. 2: Functional Design of the Human Lung for Gas Exchange . Fishman's Pulmonary Diseases and Disorders, 5e. , (2008).
- Foster, K. A., Oster, C. G., Mayer, M. M., Avery, M. L., Audus, K. L. Characterization of the A549 Cell Line as a Type II Pulmonary Epithelial Cell Model for Drug Metabolism. Experimental Cell Research. 243 (2), 359-366 (1998).
- Guo, X. Y., Lu, M., Chen, X. Q., He, F. D., Li, A. Correlation study of biological characteristics of non-small cell lung cancer A549 cells after transfecting plasmid by microbubble ultrasound contrast agent. Asian Pacific Journal of Tropical Medicine. 9 (6), 582-586 (2016).
- Cooper, J. R., et al. Long Term Culture of the A549 Cancer Cell Line Promotes Multilamellar Body Formation and Differentiation towards an Alveolar Type II Pneumocyte Phenotype. PLoS ONE. 11 (10), 0164438 (2016).
- Bisig, C., Voss, C., Petri-Fink, A., Rothen-Rutishauser, B. The crux of positive controls - Proinflammatory responses in lung cell models. Toxicology In Vitro. 54, 189-193 (2019).
- Rothen-Rutishauser, B., et al. A newly developed in vitro model of the human epithelial airway barrier to study the toxic potential of nanoparticles. ALTEX. 25, (2008).
- Braakhuis, H. M., et al. Progress and future of in vitro models to study translocation of nanoparticles. Archives of Toxicology. 89 (9), 1469-1495 (2015).
- Thai, P., Chen, Y., Dolganov, G., Wu, R. Differential regulation of MUC5AC/Muc5ac and hCLCA-1/mGob-5 expression in airway epithelium. American Journal of Respiratory Cell and Molecular Biology. 33 (6), 523-530 (2005).
- Wu, J., et al. Characterization of air-liquid interface culture of A549 alveolar epithelial cells. Brazilian Journal of Medical and Biological Research. 51 (2), 6950 (2017).
- Shapiro, D. I., Nardone, L. L., Rooney, S. A., Motoyama, E. K., Munoz, J. L. Phospholipid biosynthesis and secretion by a cell line (A549) which resembles type II aleveolar epithelial cells. Biochimica and Biophysica Acta. 530 (2), 197-207 (1978).
- Balis, J., Bumgarner, S. D., Paciga, J. E., Paterson, J. F., Shelley, S. A. Synthesis of lung surfactant-associated glycoproteins by A549 cells: description of an in vitro model for human type II cell dysfunction. Experimental Lung Research. 6 (3-4), 197-213 (1984).
- Schurch, S., Gehr, P., Im Hof, V., Geiser, M., Green, F. Surfactant displaces particles toward the epithelium in airways and alveoli. Respiration Physiology. 80 (1), 17-32 (1990).
- Gehr, P., Schurch, S., Berthiaume, Y., Hof, V. I., Geiser, M. Particle Retention in Airways by Surfactant. Journal of Aerosol Medicine. 3 (1), 27-43 (2009).
- Blank, F., Rothen-Rutishauser, B., Gehr, P. Dendritic Cells and Macrophages Form a Transepithelial Network against Foreign Particulate Antigens. American Journal of Respiratory Cell and Molecular Biology. 36 (6), (2007).
- Rothen-Rutishauser, B. M., Kiama, S. G., Gehr, P. A three-dimensional cellular model of the human respiratory tract to study the interaction with particles. American Journal of Respiratory Cell and Molecular Biology. 32, (2005).
- Blank, F., Rothen-Rutishauser, B. M., Schurch, S., Gehr, P. An optimized in vitro model of the respiratory tract wall to study particle cell interactions. Journal of Aerosol Medicine. 19, (2006).
- Jardine, L., et al. Lipopolysaccharide inhalation recruits monocytes and dendritic cell subsets to the alveolar airspace. Nature Communications. 10 (1), 1999 (2019).
- Kopf, M., Schneider, C., Nobs, S. P. The development and function of lung-resident macrophages and dendritic cells. Nature Immunology. 16 (1), 36-44 (2015).
- Chortarea, S., et al. Repeated exposure to carbon nanotube-based aerosols does not affect the functional properties of a 3D human epithelial airway model. Nanotoxicology. 9 (8), 983-993 (2015).
- Hilton, G., Barosova, H., Petri-Fink, A., Rothen-Rutishauser, B., Bereman, M. Leveraging proteomics to compare submerged versus air-liquid interface carbon nanotube exposure to a 3D lung cell model. Toxicology In Vitro. 54, 58-66 (2019).
- Brandenberger, C., et al. Effects and uptake of gold nanoparticles deposited at the air-liquid interface of a human epithelial airway model. Toxicology and Applied Pharmacology. 242, (2010).
- Drasler, B., et al. Single exposure to aerosolized graphene oxide and graphene nanoplatelets did not initiate an acute biological response in a 3D human lung model. Carbon. 137, 125-135 (2018).
- Durantie, E., et al. Carbon nanodots: Opportunities and limitations to study their biodistribution at the human lung epithelial tissue barrier. Biointerphases. 13, (2018).
- Brandenberger, C., et al. Quantitative evaluation of cellular uptake and trafficking of plain and polyethylene glycol-coated gold nanoparticles. Small. 6 (15), 1669-1678 (2010).
- Tomašek, I., et al. Combined exposure of diesel exhaust particles and respirable Soufrière Hills volcanic ash causes a (pro-)inflammatory response in an in vitro multicellular epithelial tissue barrier model. Particle and Fibre Toxicology. 13 (1), 67 (2016).
- Nazarpour, R., et al. Optimization of Human Peripheral Blood Mononuclear Cells (PBMCs) Cryopreservation. International Journal of Molecular and Cellular Medicine. 1 (2), 88-93 (2012).
- Endes, C., et al. An in vitro testing strategy towards mimicking the inhalation of high aspect ratio nanoparticles. Particle and Fibre Toxicology. 11 (1), (2014).
- Ju, X., et al. The Analysis of CD83 Expression on Human Immune Cells Identifies a Unique CD83+-Activated T Cell Population. Journal of Immunology. 197 (12), 4613-4625 (2016).
- Lenz, A. G., et al. Inflammatory and Oxidative Stress Responses of an Alveolar Epithelial Cell Line to Airborne Zinc Oxide Nanoparticles at the Air-Liquid Interface: A Comparison with Conventional, Submerged Cell-Culture Conditions. BioMed Research International. , 12 (2013).
- Germann, A., Schulz, J. C., Kemp-Kamke, B., Zimmermann, H., von Briesen, H. Standardized serum-free cryomedia maintain peripheral blood mononuclear cell viability, recovery, and antigen-specific T-cell response compared to fetal calf serum-based medium. Biopreservation and Biobanking. 9 (3), 229-236 (2011).
- Weinberg, A., et al. Optimization and Limitations of Use of Cryopreserved Peripheral Blood Mononuclear Cells for Functional and Phenotypic T-Cell Characterization. Clinical and Vaccine Immunology. 16 (8), 1176 (2009).
- Freshney, R. I., Freshney, R. I. . Culture of animal cells: a manual of basic technique. , 321-334 (2005).
- Lehmann, A. B. C., Blank, F., Gehr, P., Rothen-Rutishauser, B., Yarmush, M. L., Langer, R. S. . Alternatives to animal testing. , 239-260 (2010).
- Steiner, S., et al. Reduction in (pro-)inflammatory responses of lung cells exposed in to diesel exhaust treated with a non-catalyzed diesel particle filter. Atmospheric Environment. 81, 117-124 (2013).
- Martinez, F. O., Gordon, S., Locati, M., Mantovani, A. Transcriptional Profiling of the Human Monocyte-to-Macrophage Differentiation and Polarization: New Molecules and Patterns of Gene Expression. The Journal of Immunology. 177 (10), 7303 (2006).
- Chortarea, S., et al. Profibrotic activity of multi-walled carbon nanotubes upon prolonged exposures in different human lung cell types. Applied In Vitro Toxicology. 5 (1), (2019).
- Holt, P. G. Pulmonary Dendritic Cells in Local Immunity to Inert and Pathogenic Antigens in the Respiratory Tract. Proceedings of the American Thoracic Society. 2 (2), 116-120 (2005).
- Pinkerton, K. E., Gehr, P., Castañeda, A., Crapo, J. D., Parent, R. A. . Comparative Biology of the Normal Lung (Second Edition). , 105-117 (2015).
- Crapo, J., Barry, B., Gehr, P., Bachofen, M., Weibel, E. R. Cell number and cell characteristics of the normal human lung. American Review of Respiratory Disease. 126 (2), 332-337 (1982).
- Maniecki, M. B., Møller, H. J., Moestrup, S. K., Møller, B. K. CD163 positive subsets of blood dendritic cells: The scavenging macrophage receptors CD163 and CD91 are coexpressed on human dendritic cells and monocytes. Immunobiology. 211 (6), 407-417 (2006).
- Chignard, M., Balloy, V. Neutrophil recruitment and increased permeability during acute lung injury induced by lipopolysaccharide. American Journal of Physiology-Lung Cellular and Molecular Physiology. 279 (6), 1083-1090 (2000).
- Coyne, C. B., et al. Regulation of Airway Tight Junctions by Proinflammatory Cytokines. Molecular Biology of the Cell. 13 (9), 3218-3234 (2002).
- Durantie, E., et al. Biodistribution of single and aggregated gold nanoparticles exposed to the human lung epithelial tissue barrier at the air-liquid interface. Particle and Fibre Toxicology. 14 (49), (2017).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеThis article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены