
Method Article
Расшифровка структурных эффектов активации соматических мутаций EGFR с помощью моделирования молекулярной динамики
В этой статье
Резюме
Цель этого протокола заключается в использовании молекулярной динамики моделирования для изучения динамических структурных изменений, которые происходят из-за активации мутаций белка киназы EGFR.
Аннотация
Многочисленные соматические мутации, происходящие в эпидермальных рецепторов фактора роста (EGFR) семьи (ErbB) рецепторов тирозин киназ (RTK) были зарегистрированы у больных раком, хотя относительно немногие из них были протестированы и показано, вызывают функциональные изменения в ErbBs. Рецепторы ErbB затемнены и активируются при связывании лиганда, а динамические конформационные изменения рецепторов присущи индукции сигнализации ниже по течению. Для двух мутаций, экспериментально измененных функцией EGFR, A702V имутацией удаленияELREA750, мы иллюстрировать в следующем протоколе, как молекулярная динамика (MD) моделирования могут зондировать (1) конформационную стабильность структуры мутант тирозинкиназы по сравнению с диким типом EGFR; (2) структурные последствия и конформации переходов и их связь с наблюдаемых функциональных изменений; (3) влияние мутаций на прочность связывающего АТФ, а также на связывание доменов киназы в активированном асимметричном димере; и (4) влияние мутаций на ключевые взаимодействия в месте связывания EGFR, связанное с активированным ферментом. Протокол обеспечивает подробную пошаговую процедуру, а также руководство, которое может быть более в целом полезно для исследования белковых структур с использованием моделирования MD в качестве средства для зондирования структурной динамики и отношения к биологической функции.
Введение
Семейство рецепторов эпидермального фактора роста человека (EGFR) (ErbB) рецепторов тирозинкиназ (RTKs) включает в себя четыре члена - EGFR/ErbB1/HER1, ErbB2/HER2, ErbB3/HER3 и ErbB4/HER4. Рецепторы ErbB регулируют фундаментальные клеточные процессы, такие как рост клеток и пролиферации,дифференциация, миграция и выживание 1,2, и, таким образом, мощные прото-онкогены. Анома дюрант активность рецепторов ErbB, особенно EGFR и ErbB2, часто ассоциируется с раком человека, что делает рецепторы ErbB ключевыми целями длялечения рака 2,3.
Несколько соматических изменений генов ERBB были зарегистрированы от злокачественных новообразованийчеловека 3,,4,,5. Лучшие характерные примеры включают периодические, активации точечных мутаций и короткие в кадре удаления в домене EGFR киназы при не-малоклеточного рака легких (NSCLC). Эти мутации EGFR представляют собой ключевые факторы роста рака, и предсказать чувствительность к EGFR ориентациирака наркотиков 6,,7,,8. Однако, в большинстве видов рака, соматические мутации в EGFR происходят за пределами этих повторяющихся "горячих точек" и распространяются по всему 1210-остаток диапазона рецепторов. Действительно, большинство остатков вдоль первичной последовательности EGFR были найдены, чтобы мутировать в рак человека9. Тем не менее, помимо нескольких горячих точек, функциональная значимость подавляющего большинства связанных с раком мутаций EGFR остается неизвестной.
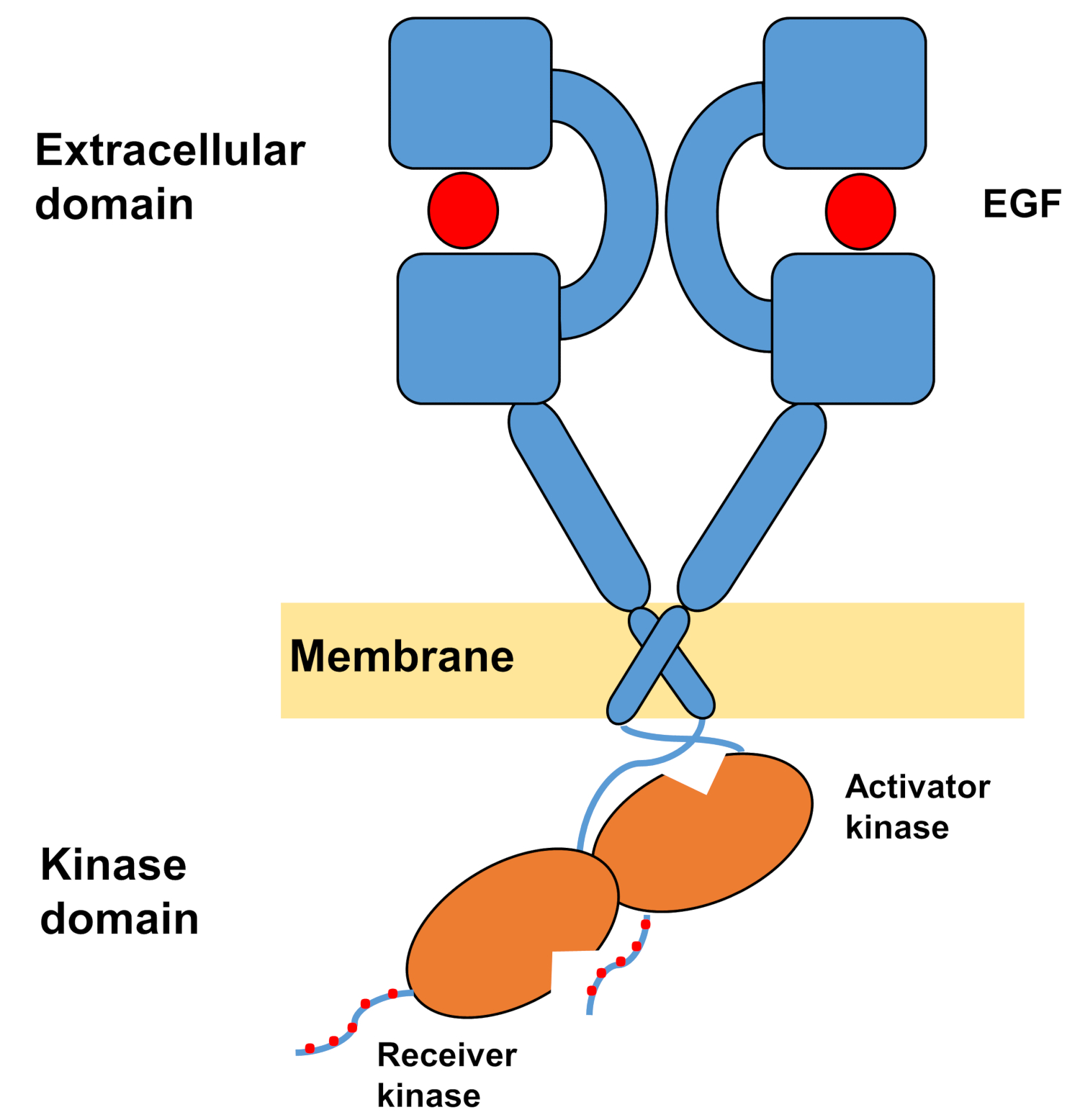
Мономерная структура ErbBs состоит из большого амино-терминала внеклеточного домена, за которым следует одна трансмембранная спираль, ведущая к внутриклеточному домену тирозинкиназы и области хвоста C-терминала, которая содержит стыковочный участки для внутриклеточных сигнальных белков. Лиганд связывания вызывает резкое конформации изменения во внеклеточной области, которая облегчает формирование рецепторов димеров, подвергая димеризации оружия, которые симметрично пересекают друг друга и взаимодействуют с их ароматических / гидрофобных поверхностей. При формировании рецептора димер тирозинкиназы вступают в контакт асимметрично(рисунок 1), в результате чего активация киназы, что фосфорилат C-терминал хвосты рецепторов мономеров, а затем в активации вниз потечению сигнализации 10,11.

Рисунок 1: Структура димера EGFR. EGFR dimerizes, когда внеклеточные домены связывают фактор роста (EGF, эпидермальный фактор роста). Затем домен приемника киназы активируется через асимметричное взаимодействие с доменом киназы активатора, а хвосты C-терминала самофосфорилируются на остатках тирозина (модифицированные из Тамирата идр. 12). Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}
Из-за динамических структурных перестановок, которые происходят во время мономерных переходов димера, наряду с активацией киназы, которая  связана с образованием асимметричного димера, мутации по всей длине структуры рецепторов потенциально могут иметь влияние на функцию рецепторов. Здесь мы описываем несколько примеров из наших предыдущих исследований, в которых моделирование мутации и визуализации было достаточно, чтобы объяснить последствия для функции.
связана с образованием асимметричного димера, мутации по всей длине структуры рецепторов потенциально могут иметь влияние на функцию рецепторов. Здесь мы описываем несколько примеров из наших предыдущих исследований, в которых моделирование мутации и визуализации было достаточно, чтобы объяснить последствия для функции.
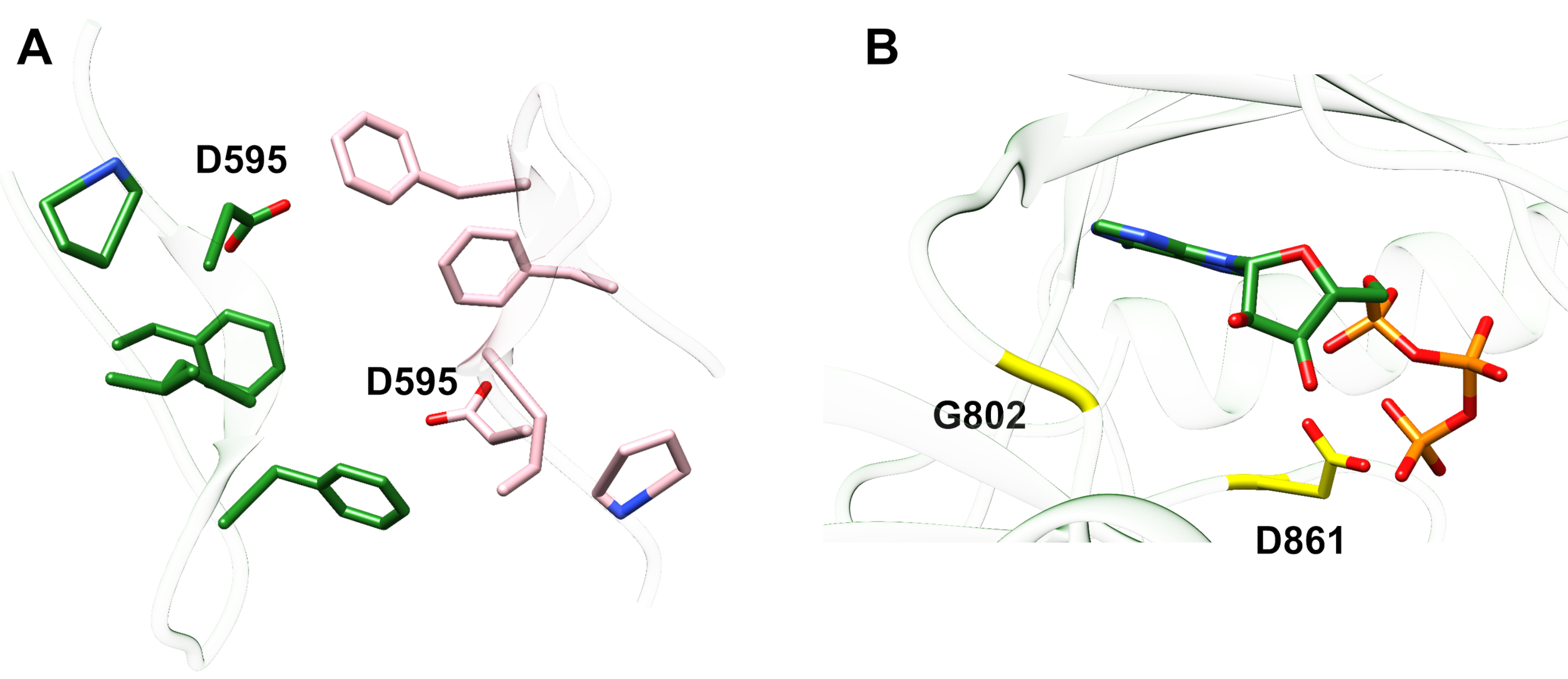
Пример 1: Один сообщил мутации, D595V в ErbB413, привело к увеличению ErbB4 димеризации ифосфорилирования 14. Визуализация местонахождения мутации была критическим фактором в понимании наблюдаемых функциональных эффектов: D595V произошел при симметричном кроссовере димерикических рук эктодомена(рисунок 2А). Руки в основном ароматические и гидрофобные, и замена полярной аспарагиновой кислоты валин, как ожидается, увеличит "липкие" гидрофобные взаимодействия, стабилизируют димер и, следовательно, увеличить продолжительность времени, когда фосфорилирование происходит14. Это было неожиданностью на первый найти аспарта в каждой руке, но в ретроспективе можно было бы думать о нем, как механизм времени для деятельности, где полярной кислоты боковых цепей уменьшить близость и срок службы нетронутыми димер и, следовательно, ограничить киназы опосредованного фосфорилирования и сигнализации. Замена валин затем удалить эту гарантию путем дальнейшей стабилизации ErbB4 димер.

Рисунок 2: Расположение активации мутации ErbB4 и мутаций, производящих киназы мертвых ErbB4. (A) D595 (активация мутации D595V) расположен на ароматических/гидрофобных димерикических руках эктодомаина модели ErbB4; оружие ассоциируется с обязательным фактором роста; (близлежащие остатки показаны как палочки). (B) В ErbB4, G802 (инактивация мутации G802dup) помогает сформировать связывающий карман вокруг аденинового кольца АТФ и каталитической D861 (инактивация мутации D861Y) связывает как Mg2 "(не показано) и γ-фосфатной группы АТФ. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}
Пример 2: Можно предвидеть, что соматические мутации, нацеленные на АТФ-связывающее место домена киназы, изменят или устранят энзиматические действия, ведущие к ослаблению или киназно-мертвому рецептору, неспособен к сигнализации. Из девяти зарегистрированных мутаций у пациентов с грудной, желудочной, колоректальной или NSCLC15,две из девяти мутаций при тестировании имели сильно уменьшеннуюфосфорилционную активность 16: G802dup (G → GG) и D861Y. Обе инактивирующие соматические мутации были найдены в месте связывания АТФструктуры домена тирозинкиназы (рисунок 2B):гибкий глицин, дублируется, изменит место ангенин кольцо и небольшой аспарагиновой кислоты заменен громоздкий тирозин вблизи терминала фосфатов будет физически предотвратить Mg2 "-ATPот связывания. Тем не менее, так как ErbB4 может образовывать гетенодимер с ErbB2 - ErbB2 не связывает фактор роста и зависит от ассоциации с ErbB, что делает для того, чтобы гетенодимеризировать - ErbB2 (активный)-ErbB4 (киназы мертвых) гетенодимер будет стимулировать пролиферацию клеток через Erk / Akt сигнализации путь еще клетки не будут дифференцировать из-за киназы мертвых ErbB4 и отсутствие активации путиSTAT5 16.
В более поздних исследованиях стало очевидно, что динамические движения эрббов имеют отношение к пониманию влияния некоторых мутантов на функцию ErbB, особенно мутаций, которые происходят в области тирозинкиназы. Область тирозинкиназы состоит из N-доли (в основном β листов) и C-доли (в основном альфа-хеликал), которые разделены каталитической сайт, где АТФ связывает. N-доля включает в себя спираль qC и P-петлю, в то время как активация (A-loop) и каталитические петли присутствуют в C-доле17,18,19. Кристаллические структуры домена тирозинкиназы выявили две неактивные конформации, большинство структур имеют Src-подобное неактивное состояние. В активной конформации каталитический аспартат A-петли указывает на сайт связывания АТФ, а спираль КЗ ориентирована на связующий карман АТФ («К-в» конформации), образуя сильное взаимодействие глутамата-лизина ион-пары.
Поскольку эрббы и область компонентов киназы являются высокодинамиными объектами, и особенно в тех случаях, когда влияние мутаций на функцию и биологическую активность, вероятно, тесно связано с конформационными состояниями Эрббов, важно оценить мутации в отношении диапазона динамических изменений, которые они будут испытывать. Рентгеновские кристаллические структуры Эрббса обеспечивают статические снимки 3D-структуры, которые могут иметь или не иметь отношения к пониманию динамических последствий мутации. Для зондирования диапазона динамических изменений, соответствующих "энергетическому ландшафту", доступен трехмерной (3D) структуре, молекулярной динамики (MD) моделирования широкоиспользуются 20. В случае мутаций, которые привели бы к локальным конформационным изменениям в домене тирозинкиназы или стабилизации комплекса, моделирование порядка 100 нс может быть достаточным. Однако более масштабные конформные изменения (например, переходы между активными и неактивными конформациями домена киназы) требуют более длительного времени моделирования - порядкамикросекунд 21.
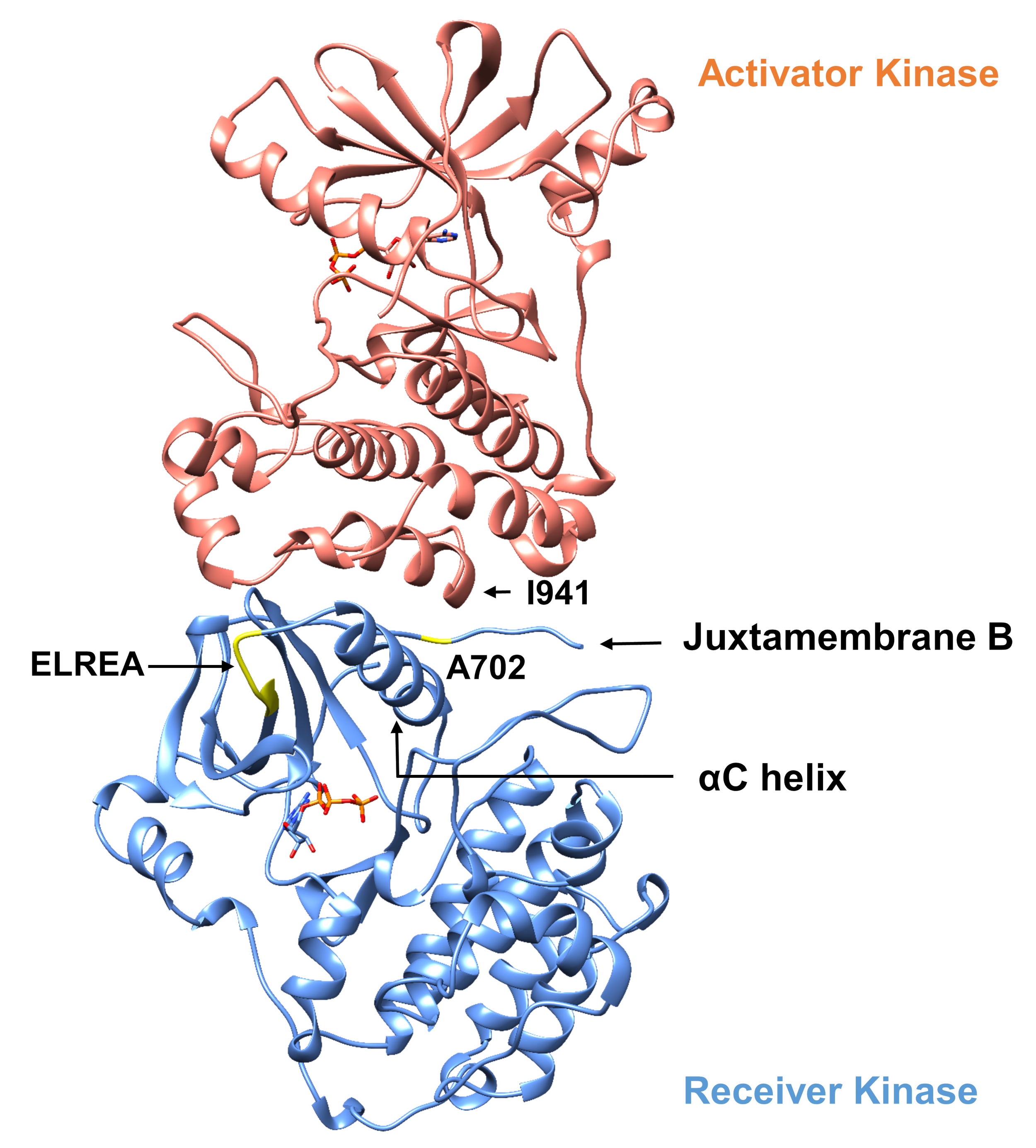
Что касается протокола, описанного ниже, мы рассматриваем две активации мутаций в домене тирозинкиназы(рисунок 3). Обе мутации расположены в домене киназы в местах, которые испытывают локальные конформационные изменения, которые диктуют, активна ли киназа или нет, и, таким образом, в обоих случаях применялись моделирования MD. В первом случае мы рассматриваем изменения, которые непосредственно влияют на сайт связывания АТФ и каталитический механизм домена киназы приемника EGFR, в частности, изучая последствия мутации удаления exon 19, которая широко замешана в NSCLC4,7. МутацияNO 746ELREA750, которая уменьшает длину цикла Q3-К, предшествующего спирали QC - спирали, которая движется к связывающему/активному месту активации киназы и участвует в формировании критического электростатического взаимодействия между E762 спирали и K745 путем позиционирования лизина для взаимодействия с АТФ - предрасполагает доменк активации 12. Во втором случае, мы рассматриваем A702V мутации EGFR, показано, что новый усиления функции активации мутации выявлены iScreamплатформы 9 и определены в NSCLC пациента22. Аланин-702 на домене приемника киназы расположен на juxtamembrane сегменте B на интерфейсе приемника и активатора киназы доменов, в которых этот асимметричный киназы димер комплекс и киназы конформации изменения необходимы дляактивации 9.

Рисунок 3: Асимметричный домен киназы димер EGFR. Мутация A702V будет расположена на критическом интерфейсе доменов активатора и приемника киназы, прилегающих к спирали КК и близко к изолеуцину 941 активатора киназы. Конформациальные изменения, вызванные образованием асимметричного димера, приводят к активации киназы. Цикл NO3-К, содержащий последовательность ELREA, непосредственно предшествует спирали КК; во время активации спираль QC движется внутрь к месту связывания АТФ. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}
протокол
ПРИМЕЧАНИЕ: Подробные шаги, предпринятые для изучения влияния мутации ЗЕЛЬРЕА и A702V на структуру EGFR с помощью моделирования MD, обсуждаются следующим образом:
1. Подготовка структуры
ПРИМЕЧАНИЕ: Для изучения структурных воздействий мутации ЗЕЛРЕА, формы апо-активных, активных и неактивных мономерных структур ЭГФР, связанных с АТФ, готовятся следующим образом.
- Откройте программу визуализации Chimera23 (https://www.cgl.ucsf.edu/chimera/)для подготовки активной структуры АПО дикого типа EGFR. В меню Файла нажмите на опцию Fetch по ID и выберите базу данных Protein Data Bank (PDB24)и укажите код PDB 2GS225 (разрешение 2.8). PDB является хранилищем для 3D-структур, решенных с помощью различных экспериментальных методов, включая рентгеновской кристаллографии, ядерной магнитно-резонансной спектроскопии, криоэлектронной микроскопии и нейтронной дифракции.
- Создайте недостающие структурные элементы 2GS2, взяв эти сегменты из структур EGFR PDB 1M1426 (2,6 евро) и 3W2S27 (1,9 евро). Для этого откройте 1M14 и 3W2S и наложить их на 2GS2, используя опцию MatchMaker в меню сравнения → Структуры инструментов.
- Урожай из сегментов, которые будут добавлены из 1M14 и 3W2S. Выберите терминальные атомы остатков перед зазорами в 2GS2 и атомы, которые будут добавлены из 1M14 и 3W2S (для получения подробной информации о добавленных сегментах, см. таблицу 1). На командной строке типа облигаций sel и хит введите. Эта структура является структурой шаблона.
- Используйте структуру шаблона от шага 1.2 для построения мутантной формы мутанта удаления киназы EGFR. Создать последовательность формата FASTA ЗЕЛЬРЕА EGFR, сохранив последовательность структуры шаблона(Любимая → Последовательность → Файл →сохранить как ), а затем удалив последовательность ELREA на остатках номера 746-750.
- Откройте последовательность ЗЕЛЬРЕА EGFR в Chimera и согласуйте ее с последовательностью структуры шаблонов 2GS2 с помощью меню Sequence. В окне выравнивания выберите параметр Structure → Modeller (гомология)28.
- В всплывающем окне укажите композитную структуру 2GS2 в качестве шаблона и последовательность мутантов в качестве запроса, который будет смоделирован. Затем нажмите OK. Выберите модель мутантов среди полученных моделей, основываясь на оценке zDOPE (обычно самый низкий балл) и визуальном осмотре.
- Для подготовки структуры киназы ЕГФР, связанной с АТФ, в качестве основной структуры используйте структуру PDB2ITX 29 (2,98 евро). Создавайте недостающие сегменты (см. таблицу 1)с использованием структур2GS6 25 (2,6 евро) и 3W2S, следующих за процедурой в шаге 1.2. Преобразование лиганд ANP в результирующую структуру в АТФ путем открытия файла PDB в текстовом редакторе и изменения атома азота N3B ANP на атом кислорода.
- Откройте структуру в шаге 1.4 в Chimera, как указано в шаге 1.1. Добавьте ион магния в эту структуру из структуры PDB2ITN 29 (2,47 евро), чтобы достичь аналогичного позиционирования для иона Mg2.
- Структурой, полученной в результате шага 1.5, моделируете форму мутанта ЗЕЛЕРЕА, следуя шагу 1.3.
| Апо активный EGFR | Апо неактивных EGFR | Активный EGFR, связанный с АТФ | |
| Основная структура | 2GS2 | 2GS7 | 2ITX |
| Структуры, используемые для построения отсутствующих петель | 1М14 (723-725) | 3W2S (958-984) | 2GS6 (862-865) |
| 3W2S (967-981) | 4HJO (848-850) | 3W2S (990-1001) |
Таблица 1: Структуры, используемые для построения композитных моделей активных, апо-неактивных и активных структур, связанных с АТФ. Недостающие области (аминокислотный диапазон в скобках) в основной структуре были построены из перечисленных структур.
- Чтобы подготовить неактивную структуру киназы апо дикого типа EGFR, откройте структуру PDB 2GS725 (2,6 евро), как в шаге 1.1, и удалите связанные лиганды и кристаллографические воды. Добавьте недостающие сегменты в 2GS7 (см. таблицу 1)из структур 3W2S и 4HJO30 (2,75 евро), используя процедуру в шаге 1.2. Основываясь на окончательной неактивной структуре EGFR, подготовьте модель мутантов, используя процедуру в шаге 1.3.
ПРИМЕЧАНИЕ: Для исследования мутации A702V изучается асимметричная структура димера EGFR, так как мутация находится в сегменте juxtamembrane B киназы, который составляет большую часть димерного интерфейса. Структуры мутантов дикого типа и A702V EGFR подготовлены следующим образом: - Асимметричная структура димера дикого типа построена из структуры PDB 2GS2, которая изначально отображается в мономерной форме. Для преобразования в биологическую сборку, которая содержит активатор и приемник киназ в асимметричном расположении, откройте 2GS2 в Химере, как в (1.1) и выполнить расчеты симметрии, нажав на меню Tools → Higher-Order Structure → Unit Cell. Выберите структуру 2GS2 и введите копии Make. Наконец, выберите и сохраните один асимметричный димер из нескольких копий димера в результате операций симметрии.
- Используя асимметричную структуру EGFR дикого типа из шага 1.8, создайте мутант A702V, заменив аланин 702 на валин с помощью инструмента → Structure, редактируя → Rotamers в Chimera.
ПРИМЕЧАНИЕ: В совокупности, шесть мономерных и две димерикические структуры EGFR подготовлены для исследований мутаций ЗЕЛЬРЕА и A702V, соответственно. Каждая структура впоследствии обрабатывается для моделирования с использованием мастера приготовления белка в программе Maestro31 и MD моделирования сделаны с янтаря программы32. - Откройте структуру в Maestro с помощью параметра структуры → файлов. Затем нажмите на кнопку мастера приготовления белка и выберите следующее: добавьте атомы водорода, постройте недостающие атомы боковой цепи, определите состояния протонирования ионизируемых остатков при рН 7.0 с помощью PROPKA, оптимизируйте ориентацию аспарагина, глутамина и гистидина остатков для склеивания водорода и, наконец, минимизировать структуру.
2. Настройка системы
- Откройте високосную программу, включенную в пакет программного обеспечения Amber. Импорт ff14SB силовоеполе 33 (источник leaprc.protein.ff14SB) и TIP3Pмолекулы воды 34 (источник leaprc.water.tip3p). Для систем, связанных с АТФ, также импортируют параметры дляАТФ 35 (loadamberparams frcmod.phosphate, loadamberprep ATP.prep). Затем загрузите структуру(mol и loadpdb structure.pdb).
- Solvate структуры в октагедоральной коробке с явным TIP3P молекулы воды, которая простирается на 10 евро во всех направлениях от поверхности атомов белка(solvateoct мол TIP3PBOX 10.0).
- Проверьте построенную систему( Проверить моль) и нейтрализовать его, добавив необходимыеионы (дополнения мол Наз 0). Чтобы достаточно моделировать биомолекулярные системы,добавьтедополнительные атомы Na q/Cl- в коробку для моделирования, чтобы довести концентрацию соли в системе до 0,15 М (дополненияmol Na' X, дополнения mol Cl- X), где X заменяется результатом: желаемой концентрации соли и количества молекул воды , объема на молекулу воды и номера Авогадро.
- Создание и сохранение топологии и координации файлов системы, которые служат в качестве входных данных для последующего моделирования производства(saveamberparm mol X.prmtop X.inpcrd).
3. Моделирование молекулярной динамики
- Используя Янтарь, первоначально подвергте систему моделирования 5000 циклов крутейшее спуск и конъюгировать минимизацию энергии градиента для того чтобы обойти неблагоприятные конфигурации. Проведение минимизации в несколько этапов, постепенно снижая ограничение, применяемое на растворимых атомов от 25ккал мол -1 -2 до 0 ккалмол -1 -2.
- В файле минимизации ввода, min.in, настроить переменную maxcyc для общего цикла минимизации (maxcyc 5000) и ncyc, чтобы узнать количество циклов для крутого алгоритма спуска. Используйте restraint_wt переменную для применения удерживающего силы на растворимые атомы, указанные параметром удерживающего устройства. Затем запустите минимизацию следующим образом:
$AMBERHOME/бин/сандер -O -i min.in -o min.out -p X.prmtop -c X.inpcrd-r min.rst -ref X.inpcrd
ПРИМЕЧАНИЕ: Используемая стратегия и фактические параметры могут варьироваться в зависимости от собственных предпочтений. Подробную информацию и рекомендации можно найти в руководстве и веб-сайтеЯнтаря (https://ambermd.org/index.php)
- В файле минимизации ввода, min.in, настроить переменную maxcyc для общего цикла минимизации (maxcyc 5000) и ncyc, чтобы узнать количество циклов для крутого алгоритма спуска. Используйте restraint_wt переменную для применения удерживающего силы на растворимые атомы, указанные параметром удерживающего устройства. Затем запустите минимизацию следующим образом:
- Нагрейте систему на 100 ps от 0 K до 300 K, установив 10 ккалмол -1 -2 удерживающих на растворимых атомов. Для этого установите tempi - 0,0, temp0 - 300.0, dt - 0,002 ps, nstlim - 50000 и restraint_wt - 10 в heat.in входном файле. Выполнить отопление со следующей командой:
$AMBERHOME/бин/сандер -O -i heat.in -o heat.out -p X.prmtop -c min.rst-r heat.rst -x heat.mdcrd -ref min.rst - Равноденствие системы для 900 ps под ансамблем ДНЯО; постоянное количество атомов, температура(температура 0 и 300,0) и давление(ntp No 1), контролируя его методом Берендсена(ntt No 1). Установите 9 -3 расстояния отсечения(разрез 9,0) для дальних электростатических взаимодействий. Постепенно опустите ограничение растворимого атома до 0,1 ккалмола -1 -2 (restraint_wt 0,1). Запустите файл ввода эквилибрации, equil.in описывает вышеперечисленные параметры следующим образом:
$AMBERHOME/бин/сандер -O -i equil.in -o equil.out -p X.prmtop -c heat.rst-r equil.rst -x equil.mdcrd -ref heat.rst - Завершите эквилибровку с безудержной симуляцией 5 ns (набор dt 0.002 ps, ntslim 2500000).
$AMBERHOME/бин/сандер -O -i equil_final.in -o equil_final.out -p X.prmtop -c equil.rst -r equil_final.rst -x equil_final.mdcrd -ref equil.rst - Убедитесь, что система имеет equilibrated путем изучения температуры, давления, плотности и энергетических значений.
$AMBERHOME/bin/process_mdout.perl heat.out equil.out equil_final.out
xmgrace резюме. ТЕМП/ПЛОТНОСТЬ/ЭТОТ/EPTOT/EKTOT - Проведение производственного моделирования для 100 нс (набор dt 0,002 ps, ntslim 50000000 в prod.in) и сохранить конформации каждые 10 ps (ntwx 5000).
$AMBERHOME/бин/сандер -O -i prod.in -o prod.out -p X.prmtop -c equil_final.rst-r prod.rst -x prod.mdrcd -ref equil_final.rst
4. Анализ
- Визуальный осмотр
- Визуализуйте конформации, отобранные во время моделирования киназы дикого типа и мутанта EGFR, открыв файлы топологии янтаря X.prmtop и соответствующие файлы траектории prod.mdcrd в VMD36. Используя удобные вторичные представления структуры, проанализируйте общую структурную динамику белков с записанной траектории. Просмотр конкретных взаимодействий между атомами/остатками интереса, такими как каталитически необходимый соляной мост K745 - E762.
- Кроме того, сохраните несколько конформаций, отобранных во время моделирования в формате PDB, и откройте их с помощью программы Chimera. Наложить структуры на начальную или медианную структуру с помощью опции MatchMaker. Отображение начальной/медианной структуры в твердом и остальных выровненных структурах в выцветшей белой. Такой подход позволяет визуализировать записанные структурные движения с большей ясностью.
ПРИМЕЧАНИЕ: Предложения по эффективному представлению и обработке конформных ансамблей от MDS можно найти в Melvin et al.37.
- Анализ RMSD и RMSF
- Вычислить расчеты корневого среднего квадратного отклонения (RMSD) и расчеты квадратных колебаний корневых колебаний (RMSF) с помощью программы Cpptraj 38 для анализа глобальной стабильности белков и изучения гибкости различных структурных единиц. В файлах rmsd.in и rmsf.in ввода указываются атомы позвоночника (для RMSD) и атомы C(для RMSF) исходной структуры в качестве эталона для установки RMS. В файлах rmsd/rmsf.in импортируются файлы топологии янтаря(parm X.promtop)и соответствующие файлытраектории (trajin prod.mdcrd). Затем запустите команду Cpptraj-i rmsd/rmsf.in. Участок выходных данных для анализа.
- Кроме того, выровнять конформативные ансамбли и цвет каждого остатка на основе атома RMSD. Для этого откройте конформации в Chimera и согласуйте их с опцией Matchmaker.
- Перейти к инструментам → изображение → Рендер по атрибуту. Выберите остатки конформации ансамбля и СЗ RMSD в качестве атрибутов и нажмите OK. Цепной след конформаций затем будет окрашен из синего → белого →, соответственно отражающего области высокой, средней и низкой структурной стабильности.
- Анализ водородных связей
- Проанализируйте взаимодействие водородных связей между АТФ и диким типом/ЭЛЕРЕА EGFRs. Подготовь сценарий Cpptraj, hbond.in,для выполнения этой задачи. Определите водородную связь с расстоянием донора-приемора менее 3,5 евро и угол связи больше или равен 135 градусов. Укажите анализ только для интермолекулярных водородных связей с переменной ноинтрамол, т.е. водородных связей между АТФ и EGFR(hbond All nointramol dist 3.5 out nhb.agr avgout avghb.dat). Запустите сценарий, как Cpptraj-i hbond.in.
- Используйте этот скрипт для оценки внутримолекулярных взаимодействий, например между остатками K745 и E762, которые являются ключевыми остатками активности EGFR киназы. Для этого укажите K745 в качестве донора водородных облигаций и E762 в качестве приемлемого водородного облигаций в hbond.in и запустите сценарий соответствующим образом.
- Мониторинг расстояния между атомами
- Измерьте расстояние между K745 и E762, открыв траектории диких типов и Apo EGFRs в VMD. Выберите C q Glu762 и Nz Lys745, нажав на → мыши → облигаций. Мониторинг расстояния во время моделирования путем построения графика с графикой и → на → связи → графике.
- Бесплатные расчеты энергии
- Для расчета расчетных связывающих свободных энергий между АТФ и ЭГФРА дикого типа/ЗЕЛЬРЕА, а также между активатором и приемником киназ дикого типа/A702V EGFRs используйте модуль молекулярной механики обобщенной поверхности Born surface area (MM-GBSA)39, доступный в пакете AMBER. Установите АТФ в качестве лиганда и EGFR в качестве рецептора в исследовании ЗЕЛЬРЕА. В исследовании A702V укажите приемник киназы как лиганд и киназу активатора в качестве рецептора.
- Сначала подготовьте лиганд, рецептор и лиганд-рецептор комплекса PDB файлы отдельно в високосной программе, установив значение PBRadii до mbondi2. Для файлов PDB сохраните топсологию янтаря газовой фазы (.prmtop) и координируйте (.inpcrd) файлы.
- Затем, в файле ввода mmgbsa, mmgbsa.in,установить igb No 2, saltcon 0.1. Выполняйте связывающие энергетические вычисления с использованием траекторий моделирования, подготовленных рецепторов/лигандовых янтарных файлов и параметров в mmgbsa.in с MMPBSA.py скриптом, доступным в Amber следующим образом:
$AMBERHOME/bin/MMPBSA.py-O -i mmgbsa.in -o mmgbsa.dat -sp X.prmtop-cp complex.prmtop-rp receptor.prmtop-lp ligand.prmtop -y prod.mdcrd-eo output.csv - Проанализируйте выходные данные, output.csv, путем построения графиков.
Результаты
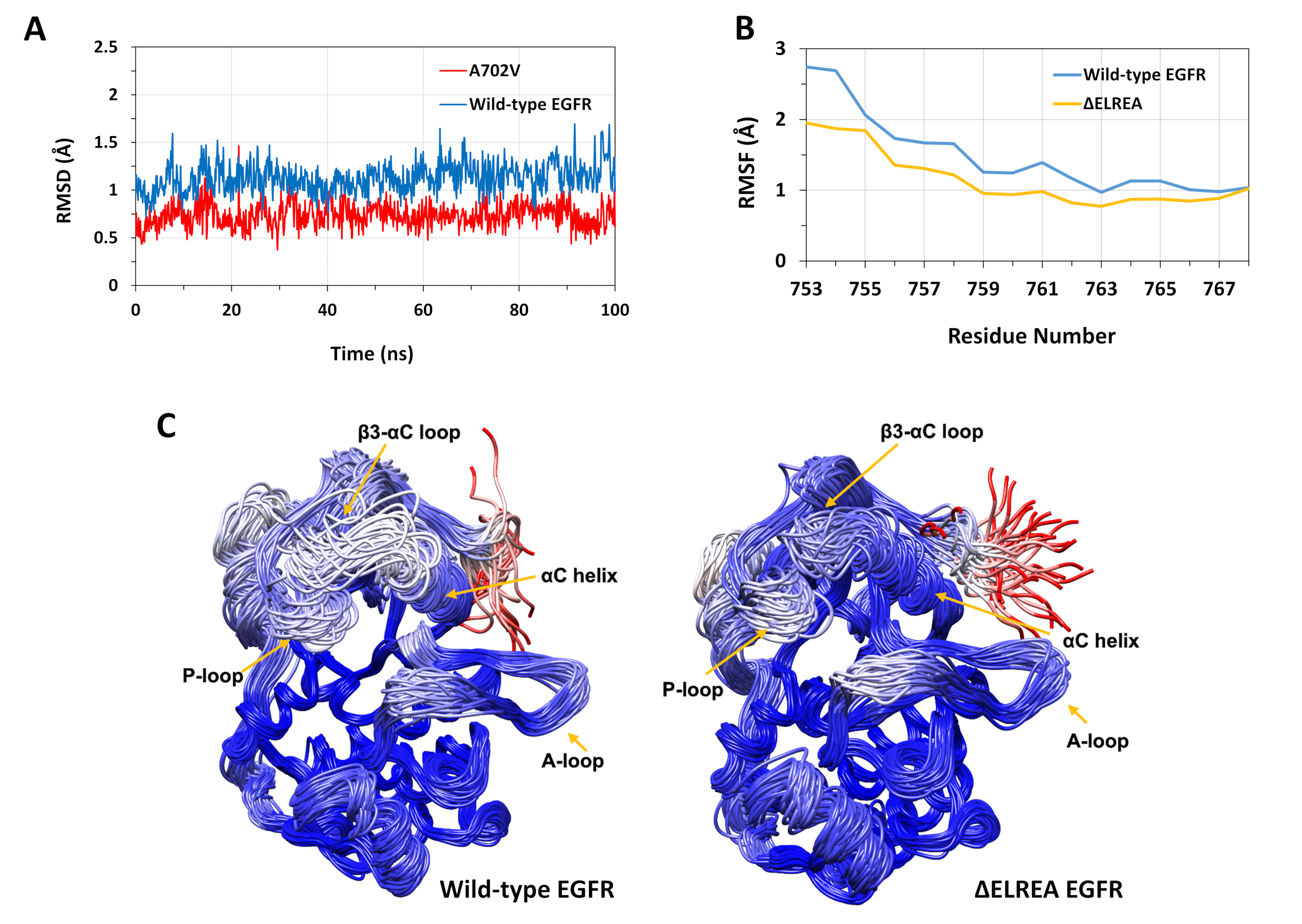
Описанный протокол был использован для изучения структурных эффектов мутаций ЗЕЛЕРЕА и A702V на структуру киназы EGFR. Одним из применений протокола было исследовать влияние мутаций на местную структурную/конформациюную стабильность путем вычисления значений RMSD и RMSF из моделирования MD. Поскольку мутация A702V находится в сегменте juxtamemembrane B, RMSD этого сегмента приемника киназы относительно стартовой структуры был рассчитан как для дикого типа, так и для ЭГФР A702V. Результат (Рисунок 4A) показал, что сегмент juxtramembrane B мутанта увеличил конформационную стабильность во время моделирования 100 ns (средний RMSD 0.7 - 95% интервал доверия (CI) 0.009) по сравнению с областью киназы дикого типа EGFR (средний RMSD 1.1 й - 95% CI 0.01). Это очень вероятно, в результате ужесточения гидрофобных взаимодействий на димер интерфейс из-за замены аланина 702 (метиловая группа боковой цепи) на громоздкие гидрофобные остатки, валин (изопропил группы боковой цепи), что приводит к увеличению гидрофобных взаимодействий V702 на приемник киназы домена с изолеуцином 941 активора киназы домена.
Мутация ЗЕЛЬРЕА расположена в петле No3-К, примыкающей к функционально критической спирали КК; конформация спирали КК является ключом к сдвигам между активными и неактивными состояниями киназы EGFR. Конформациальная стабильность спирали КЗ в активном состоянии была оценена путем изучения RMSF над атомами атомов C-атомов остатков в спирали во время моделирования MD(рисунок 4B): в целом, есть более низкие колебания в мутанте (средний RMSF 1.1 - 95% CI 0.4) по сравнению с диким типом (средний RMSF 1.5 - 95% CI 0.57); с наибольшей разницей в колебаниях, зарегистрированных для остатков N-терминала. Выборонные конформации, соответственно, налагаются на медианную структуру домена киназы дикого типа и домена киназы ЗЕЛРЕА также поддерживают эти результаты (рисунок4C):как области дикого типа, так и домены киназы QELREA имеют общую аналогичную стабильность для сверхпоставленных конформаций, за исключением цикла Q3-C и спирали QC, которые явно более стабильны в EGREA. Эти данные свидетельствуют о том, что удаление последовательности ELREA сдерживает движение активного состояния спирали КК, следовательно, сдерживая и тем самым стабилизируют активную конформацию. Кроме того, поскольку спираль КС является частью асимметричного димерного интерфейса, ограничения на спираль к КЗ у мутанта, скорее всего, стабилизируют асимметричный димер, продлевая продолжительность активированного состояния.
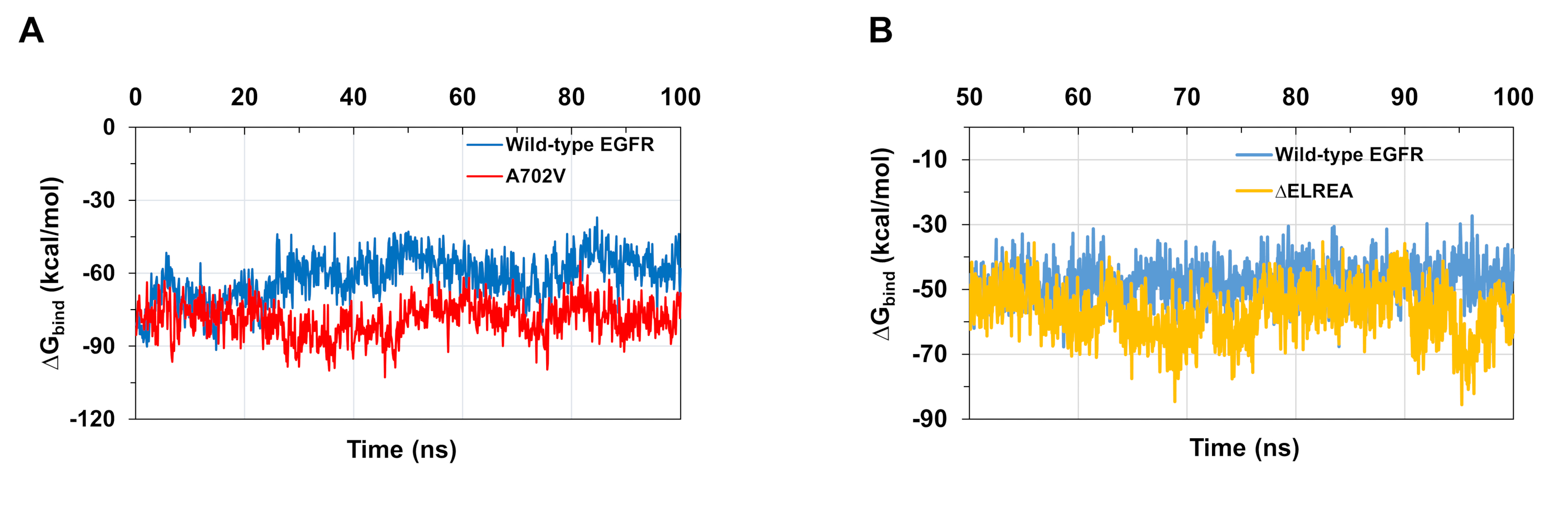
Еще одно применение протокола заключается в расследовании поведения ключевых внутри- и межмолекулярных взаимодействий, происходящих во время моделирования. Таким образом, взаимодействие между K745 и E762, которое имеет основополагающее значение для enzymatic деятельности EGFR, был проанализирован как для активной формы дикого типа, так и для киназы ЗЕЛЬРЕА EGFR путем измерения процентной заполняемости водородных связей, образоваваемых между боковыми полярными атомами двух остатков во время моделирования MD (рисунок5A):это ключевое электростатическое взаимодействие чаще формировалось в домене киназы ЗЕЛЕРЕА по сравнению с доменом киназы дикого типа, благодаря более стабильному домену E.C. Взаимодействия между Mg2 "-ATPи дикого типа и ELREA EGFR киназы доменов(рисунок 5B) в ходе моделирования были также оценены(рисунок 5C): количество водородных связей были больше для ЗЕЛЕРЕА (среднее значение 4,0 - 95% CI 0,03), чем для дикого типа EGFR (среднее значение 3,2 - 95% CI 0,04). Дальнейший анализ водородных связей показал, что K745 чаще взаимодействует с фосфатными группами АТФ в ЗЕЛЬРЕА EGFR, что связано с более стабильным взаимодействием K745-E762, отмеченным в моделировании домена киназы мутанта ЭГФРА.
Моделирование MD, описанное в протоколе, также полезно для оценки относительно свободной энергии связывания для белково-белковых и белково-лигандовых взаимодействий. Связывающие энергии между активатором и приемником киназы доменов дикого типа и A702V EGFR, а также между АТФ и дикого типа и qELREA мутант EGFR киназы доменов, были вычислены изbind молекулярной механики обобщенных Борн поверхности (MMGBSA) расчеты (Рисунок 6A): A702V мутант производится более низкое среднее значение связывания йG (средний йGсвязывать -76 ккал / мол - 95% CI 0.47), представляющих более благоприятные димерные взаимодействия, в отличие от домена EGFR дикого типа (средняяпривязка КГ -61 ккал/мол - 95% ДИ 0,61). Это наблюдение согласуется с более стабильным сегментом juxtamembrane B и более жестким интерфейсом димера из-за увеличения гидрофобных взаимодействий, наблюдаемых для домена киназы A702V EGFR. В случае привязки АТФ к доменам киназы дикого типа и ЗЕЛЬРЕА EGFR(рисунок 6B),расчеты MMGBSA предсказывают более сильную привязку АТФ с мутантом КЕЛЕРЕА (средняя привязка -57 ккал/мол - 95% CI 0,43) по сравнению с EGFR дикого типа (средняя привязка -48 ккал/мол - 95% CI 0,33).bind bind Этот результат соответствует большему числу водородных связей, зарегистрированных между АТФ и ЗЕЛЬРЕА EGFR(рисунок 5C) по сравнению с областью дикого типа.
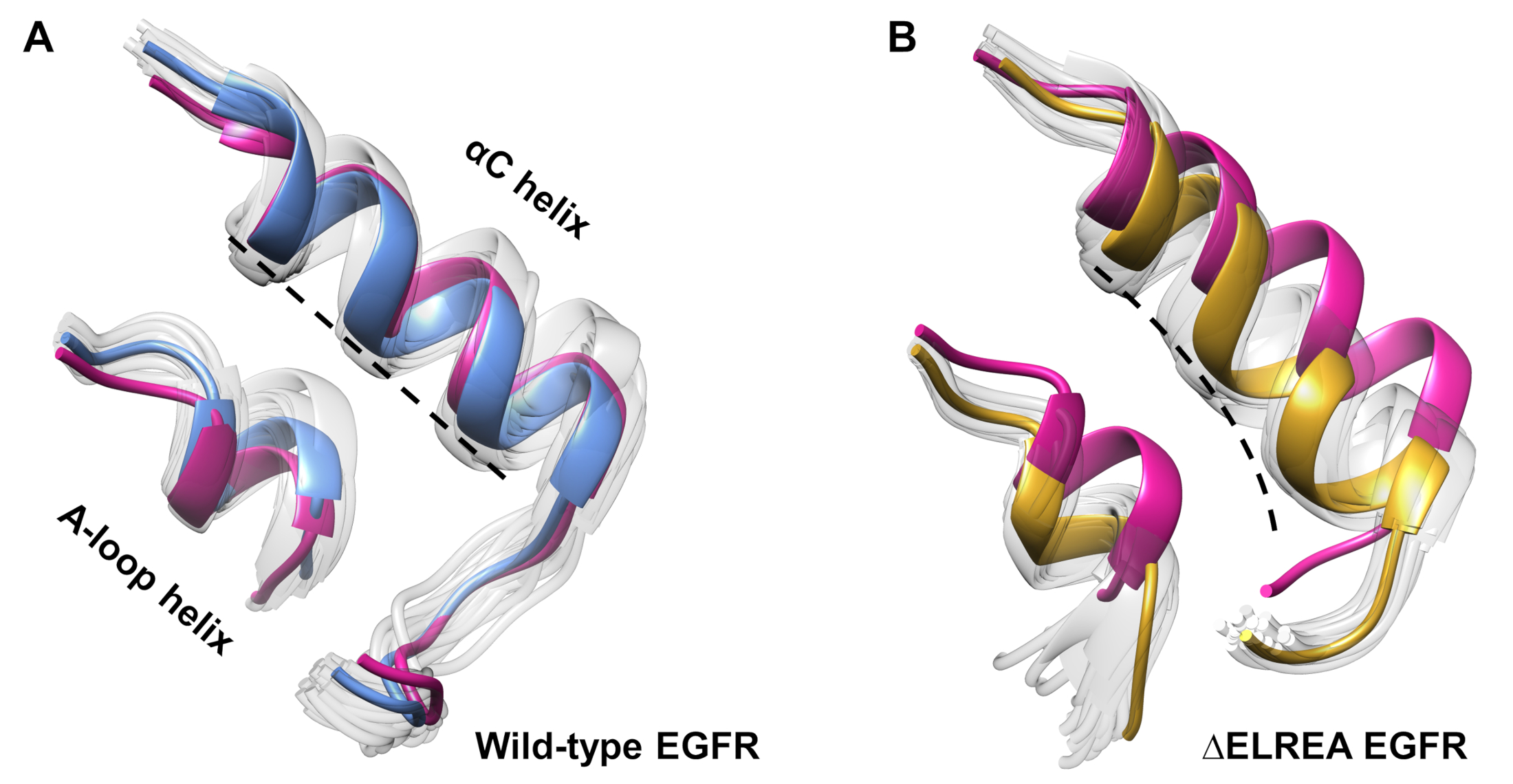
Протокол также может быть использован для исследования конформациальных изменений, наблюдаемых во время моделирования. В текущем исследовании влияние мутации ЗЕЛЕРЕА на неактивную конформацию EGFR изучалось визуальным осмотром и суперпозиционированием отобранных конформаций из моделирования. Анализ выявил внутреннее движение спирали КК в домене киназы ЗЕЛЬРЕА EGFR (рисунок7A),структурные изменения, ожидаемые при переходе в активное состояние. В отличие от этого, спираль дикого типа неактивных EGFR сохранила свою начальную конформацию(рисунок 7B). Таким образом, моделирование MD поддерживает предложение о том, что мутация удаления, показанная экспериментально дляувеличения активности киназы 40,41, способствует конформации переход от неактивной киназы к активному государству.

Рисунок 4: Дикий тип и мутант конформации стабильности активного домена EGFR киназы во время моделирования MD. (A) RMSD (атомы позвоночника) над сегментом juxtamembrane B дикого типа (синий) и A702V (красный) приемник киназы домена. (B) RMSF (атомы СЗ) над остатками спирали КК: дикого типа (синий) и ЗЕЛЕРЕА (золото). (C)Спроверенные образцы конформаций дикого типа (слева) и ЗЕЛЬРЕА (справа) домена EGFR киназы; цепные следы, окрашенные на основе RMSD (атомов C) каждого остатка по отношению к медианной структуре. Окраска варьируется от синего до белого до красного, представляющих регионы с высокой до низкой конформации стабильности. Обратите внимание, что «свободные» N-терминалы изолированных киназных доменов, окрашенных в красный цвет, не будут проявлять такой уровень мобильности в нетронутой структуре EGFR. Цифры, адаптированные из Chakroborty et al.9 (рисунок 4A воспроизводится с разрешения журнала биологической химии) и Тамират и др.12. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 5: Ключевые особенности, замеченные в активном приемнике киназы во время моделирования MD: соляной мост K745-E762, спираль к КЗ и взаимодействие с АТФ. (A)Процентная заполняемость взаимодействия K745-E762 во время моделирования доменов киназы дикого типа (синий) и ЗЕЛЬРЕА (золото). (B)Остатки дикого типа и мутанта ЗЕЛРЕА, взаимодействующих с АТФ (палками). Mg2 "(зеленый) координирует с АТФ и D855. (C) Количество водородных связей, образованных АТФ как с диким типом, так и с доменами киназы ЕГФРА во время моделирования MD. Рисунок из Тамират и др.12. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 6: Нижние относительные свободные энергии связывания наблюдаются для доменов мутантной киназы во время моделирования. (A)Связывание энергий, рассчитанных для взаимодействия между активатором и приемником киназы доменов дикого типа (синий) и A702V (красный) EGFRs. (B)привязкаАТФ к областям киназы дикого типа (синий) и ЗЕЛЬРЕА (золото). Цифры, адаптированные из Chakroborty et al.9 (Рисунок 6A воспроизводится с разрешения журнала биологической химии) и Тамират идр. 12. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 7: Суперпоставленные конформации от дикого типа и неактивного домена EGFR kinase. Конформация спирали ИК и спирали A-петли(A)дикого типа (средняя структура синим цветом) и(B)ЗЕЛЬРЕА EGFRs (золото). Другие образцы конформаций, выцветшие белые; первоначальные структуры до моделирования MD, розовый. Рисунок из Тамират и др.12. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}
Обсуждение
Протокол, описанный в этом исследовании, фокусируется на использовании моделирования молекулярной динамики для исследования локальных и глобальных структурных изменений, возникающих в результате активации соматических мутаций домена EGFR kinase. Хотя рентгеновские кристаллические структуры диких и мутантных EGFRs обеспечивают неоценимую структурную проницательность, они изображают одно или несколько статических представлений. Однако, присущие биологической функции ErbBs является необходимым переходов между энзиматично неактивных и активных тирозинкиназы, ссылаясь на динамические изменения как в структуре и внутримолекулярных взаимодействий между киназы мономеров. Таким образом, было проведено моделирование MD для зондирования динамического характера домена тирозинкиназы EGFR, включая структуру дикого типа, введенную мутацию удаления ЗЕЛЬРЕА и мутацию A702V. Эти моделирования были успешными в выяснении вероятной роли этих мутаций в структурах и как их влияние на конформацию домена тирозинкиназы приведет к экспериментально наблюдаемому увеличению активности EGFR киназы.
Важным шагом в этом протоколе является использование соответствующей структуры для оценки воздействия мутации. Один из способов выбрать соответствующую структуру ввода моделирования заключается в визуализации местоположения мутации в статической 3D-структуре и изучении ее возможного воздействия по отношению к соседним аминокислотам и структурным единицам. В этом исследовании, например, так как мутация A702V EGFR находится в сегменте juxtamembrane B, который формирует асимметричный интерфейс димера, использование структуры димера для моделирования, в отличие от мономера имеет решающее значение. Использование мономерной структуры подвергло бы juxtamembrane B сегмент киназы приемника растворителя, лишая его от стабилизирующими взаимодействиями, усиленными мутацией к более большим гидрофобным остаткам и взаимодействиям с изолеуцином 941 от остатков C-доли киназы активатора. Кроме того, следует отметить, что 3D-структура, представленная координатами в файле PDB, не обязательно соответствует биологически релевантной структуре, которая должна использоваться для изучения. Например, со структурой ErbB4, PDB-кодом 3BCE, координаты PDB соответствуют тримеру, но это связано с кристаллическими контактами (при визуализации этой структуры видны контакты между мономерами). Матрицы в файле PDB могут быть использованы (например, в Chimera) для реконструкции кристаллографически связанных структур, которые могут быть визуализированы для выявления цепей, которые соответствуют биологически релевантной 3D-структуры, как сообщается в первоначальнойпубликации 42. Другим важным шагом протокола является правильное подготовить структуру ввода моделирования, например, создание недостающих аминокислот в различных областях цикла, и особенно там, где находится в непосредственной близости от мутации. Хотя в PDB существуют многочисленные структуры EGFR дикого типа, доступно лишь ограниченное число мутантных структур EGFR. Следовательно, структуры мутантов также должны быть смоделированы; для одной остаточной мутации, такой как A702V, Химера использовалась для мутации остатков; в то время как для мутации удаления ЗЕЛЕРЕА использовался Моделлер.
Различные параметры, используемые в файлах ввода моделирования - например, количество циклов минимизации, нагревание системы до нужной температуры за один раз или вместо нагрева медленно через несколько промежуточных температур, период времени для равноденствия и для производственного моделирования - могут быть изменены на основе молекулы исследования, цели работы и собственных предпочтений. При проведении моделирования MD, он также является общим, чтобы встретить ошибки, которые могут возникнуть из входных файлов, вопросы, связанные с программным обеспечением моделирования в использовании или даже ошибка пользователя. Поэтому очень важно понимать источник ошибок, тщательно изучая любые сообщения об ошибках. Большинство программ моделирования имеют список рассылки, где пользователи могут задавать вопросы разработчикам программного обеспечения и другим пользователям, с помощью которых большинство проблем может быть решено. Кроме того, руководства пользователей оказывают существенную помощь в понимании деталей протокола моделирования, включая предположения и ограничения. Хотя моделирование MD является важным инструментом для изучения динамических свойств молекул, помните, что вычислительные результаты должны быть тщательно оценены в сочетании с другими источниками информации для оценки их достоверности. Всякий раз, когда это возможно, работа вместе с исследователями, которые являются экспертами по белкам в исследовании, особенно там, где соответствующие мокрой лаборатории экспериментальных исследований, которые служат для обеспечения результатов для структурной интерпретации, а также предложить эксперименты, которые могут быть сделаны на основе структурных наблюдений для проверки гипотез.
В этом исследовании протокол был эффективен в изучении динамических структурных воздействий мутаций ЗЕЛЕРЕА и A702V на структуры киназы EGFR. Моделирование показало, что ЗЕЛЕРЕА сдерживает функционально необходимую спираль КК и способствует конформации перехода от неактивной киназы к стабилизированной активной киназе. Результаты моделирования независимо поддерживаются данными о реакции на наркотики, которые продемонстрировали влияние ингибиторов тирозинкиназы на линии раковых клеток легких, имеющих мутацию удаления ЗЕЛЬРЕА и ЭГФР дикого типа, где большее ингибирование препаратами, распознавающих активную конформацию киназы, было сообщено для ЗЕЛЬРЕА, чем для EGFRдикого типа 12. С мутацией A702V, моделирование MD показывает, по сравнению с диким типом, повышенную стабилизацию интерфейса киназы активатора-приемника, а также более высокое сродство активатора и приемника киназы друг для друга, вместе поддерживая поддержание активированной конформации киназы EGFR. Мутация A702V, расположенная на сегменте juxtamembrane B киназы приемника, увеличит гидрофобные взаимодействия с активатором киназы, функционирующей для продления продолжительности активированного состояния. Мутация A702V поддерживает выживаемость клеток при отсутствии фактора роста и была выявлена в пробирке при скрининге на мутации EGFR9.
Раскрытие информации
Авторов нечего раскрывать.
Благодарности
Это исследование финансируется за счет грантов M.S.J от Академии Финляндии (308317, 320005), Сигрид Juselius Фонда и Tor, Джо и Пентти Борг мемориальный фонд, и К.Е. из Академии Финляндии (274728, 316796), Онкологический фонд Финляндии, и Турку университетской центральной больницы. М.З.Т. финансируется докторской сетью информационной и структурной биологии Збо Академи. Мы благодарим ИТ-центр CSC за вычислительные ресурсы и д-ра Jukka Lehtonen за ИТ-поддержку в рамках сети биоинформатики Biocenter Finland; и сеть инфраструктуры структурной биологии Biocenter Finland.
Материалы
| Name | Company | Catalog Number | Comments |
| Amber software | University of California, San Francisco | Version 2018 | Executable |
| Chimera program | Resource for Biocomputing, Visualization, and Informatics at the University of California, San Francisco | Version 1.13.1 | Executable |
| EGFR struture files | The Protein Data Bank | 3D coordinates of EGFR structures | |
| Maestro | Schrödinger LLC | Version 2018-3 | Executable |
| Modeller program | The Andrej Šali Lab, Departments of Biopharmaceutical Sciences and Pharmaceutical Chemistry, University of California San Francisco | Included in the Chimera program | |
| VMD software | Theoretical and Computational Biophysics Group, University of Illinois at Urbana-Champaign | Version 1.9.3 | Executable |
Ссылки
- Yarden, Y., Sliwkowski, M. X. Untangling the ErbB signalling network. Nature Reviews Molecular Cell Biology. 2, 127-137 (2001).
- Lemmon, M. A., Schlessinger, J., Ferguson, K. M. The EGFR family: not so prototypical receptor tyrosine kinases. Cold Spring Harbor Perspectives in Biology. 6, a020768 (2014).
- Arteaga, C. L., Engelman, J. A. ERBB receptors: From oncogene discovery to basic science to mechanism-based cancer therapeutics. Cancer Cell. 2, 282-303 (2014).
- Mishra, R., Hanker, A. B., Garrett, J. T. Genomic alterations of ERBB receptors in cancer: Clinical implications. Oncotarget. 8, 114371-114392 (2017).
- . cBioPortal for Cancer Genomics Available from: https://www.cbioportal.org (2020)
- Lynch, T. J., et al. Activating mutations in the epidermal growth factor receptor underlying responsiveness of non-small-cell lung cancer to gefitinib. New England Journal of Medicine. 350 (21), 2129-2139 (2004).
- Paez, J. G., et al. EGFR mutations in lung cancer: correlation with clinical response to gefitinib therapy. Science. 304 (5676), 1497-1500 (2004).
- Pao, W., et al. EGF receptor gene mutations are common in lung cancers from "never smokers" and are associated with sensitivity of tumors to gefitinib and erlotinib. Proceedings of the National Academy of Sciences U.S.A. 101 (36), 13306-13311 (2004).
- Chakroborty, D., et al. Unbiased in vitro screen for activating EGFR mutations. Journal of Biological Chemistry. 294 (24), 9377-9389 (2019).
- Leahy, D. J. Structure and Function of the Epidermal Growth Factor (EGF/ErbB) Family of Receptors. Advances in Protein Chemistry. 68, 1-27 (2004).
- Roskoski, R. ErbB/HER protein-tyrosine kinases: Structures and small molecule inhibitors. Pharmacological Research. 87, 42-59 (2014).
- Tamirat, M. Z., Koivu, M., Elenius, K., Johnson, M. S. Structural characterization of EGFR exon 19 deletion mutation using molecular dynamics simulation. PLoS ONE. 14 (9), e0222814 (2019).
- Ding, L., et al. Somatic mutations affect key pathways in lung adenocarcinoma. Nature. 455, 1069-1075 (2008).
- Kurppa, K. J., Denessiouk, K., Johnson, M. S., Elenius, K. Activating somatic ERBB4 mutations in non small-cell lung cancer. Oncogene. 35 (10), 1283-1291 (2016).
- Soung, Y. H., et al. Somatic mutations of the ERBB4 kinase domain in human cancers. International Journal of Cancer. 118, 1426-1429 (2006).
- Tvorogov, D., et al. Somatic mutations of ERBB4: selective loss-of-function phenotype affecting signal transduction pathways in cancer. Journal of Biological Chemistry. 284, 5582-5591 (2009).
- Hubbard, S. R., Till, J. H. Protein tyrosine kinase structure and function. Annual Review of Biochemistry. 69 (1), 373-398 (2000).
- Huse, M., Kuriyan, J. The conformational plasticity of protein kinases. Cell. 109 (3), 275-282 (2002).
- Jura, N., et al. Catalytic control in the EGF receptor and its connection to general kinase regulatory mechanisms. Molecular Cell. 42, 9-22 (2011).
- Karplus, M., Kuriyan, M., J, Molecular dynamics and protein function. Proceedings of the National Academy of Sciences U.S.A. 102 (19), 6679-6685 (2005).
- Shan, Y., Arkhipov, A., Kim, E. T., Pan, A. C., Shaw, D. E. Transitions to catalytically inactive conformations in EGFR kinase. Proceedings of the National Academy of Sciences U.S.A. 110 (18), 7270-7275 (2013).
- Reckamp, K. L., et al. A phase I trial to determine the optimal biological dose of celecoxib when combined with erlotinib in advanced non-small cell lung cancer. Clinical Cancer Research. 12 (11 Pt 1), 3381-3388 (2006).
- Pettersen, E. F., et al. UCSF Chimera-a visualization system for exploratory research and analysis. Journal of Computational Chemistry. 25 (13), 1605-1612 (2004).
- Berman, H. M., et al. The Protein Data Bank. Nucleic Acids Research. 28 (1), 235-242 (2000).
- Zhang, X., Gureasko, J., Shen, K., Cole, P. A., Kuriyan, J. An Allosteric Mechanism for Activation of the Kinase Domain of Epidermal Growth Factor Receptor. Cell. 125 (6), 1137-1149 (2006).
- Stamos, J., Sliwkowski, M. X., Eigenbrot, C. Structure of the epidermal growth factor receptor kinase domain alone and in complex with a 4-anilinoquinazoline inhibitor. Journal of Biological Chemistry. 277 (48), 46265-46272 (2002).
- Sogabe, S., et al. Structure-Based Approach for the Discovery of Pyrrolo[3,2-d]pyrimidine-Based EGFR T790M/L858R Mutant Inhibitors. ACS Medicinal Chemistry Letters. 4 (2), 201-205 (2013).
- Sali, A., Blundell, T. L. Comparative protein modelling by satisfaction of spatial restraints. Journal of Molecular Biology. 234 (3), 779-815 (1993).
- Yun, C. H., et al. Structures of lung cancer-derived EGFR mutants and inhibitor complexes: mechanism of activation and insights into differential inhibitor sensitivity. Cancer Cell. 11 (3), 217-227 (2007).
- Park, J. H., Liu, Y., Lemmon, M. A., Radhakrishnan, R. Erlotinib binds both inactive and active conformations of the EGFR tyrosine kinase domain. Biochemical Journal. 448 (3), 417-423 (2012).
- . Release 2018-3: Maestro Available from: https://www.schrodinger.com/maestro (2018)
- Case, D. A., et al. . AMBER 2018. , (2018).
- Maier, J. A., Martinez, C., Kasavajhala, K., Wickstrom, L., Hauser, K. E., Simmerling, C. ff14SB: Improving the Accuracy of Protein Side Chain and Backbone Parameters from ff99SB. Journal of Chemical Theory and Computation. 11 (8), 3696-3713 (2015).
- Jorgensen, W. L., Chandrasekhar, J., Madura, J. D., Impey, R. W., Klein, M. L. Comparison of simple potential functions for simulating liquid water. Journal of Chemical Physics. 79 (2), 926-935 (1983).
- Meagher, K. L., Redman, L. T., Carlson, H. A. Development of polyphosphate parameters for use with the AMBER force field. Journal of Computational Chemistry. 24 (9), 1016-1025 (2003).
- Humphrey, W., Dalke, A., Schulten, K. VMD: Visual molecular dynamics. Journal of Molecular Graphics. 14 (1), 33-38 (1996).
- Melvin, R. L., Salsbury, F. R. Visualizing ensembles in structural biology. Journal of Molecular Graphics and Modelling. 67, 44-53 (2016).
- Roe, D. R., Cheatham, T. E. PTRAJ and CPPTRAJ: Software for processing and analysis of molecular dynamics trajectory data. Journal of Chemical Theory and Computation. 9 (7), 3084-3095 (2013).
- Miller, B. R., et al. MMPBSA.py: An Efficient Program for End-State Free Energy Calculations. Journal of Chemical Theory and Computation. 8 (9), 3314-3321 (2012).
- Guha, U., et al. Comparisons of tyrosine phosphorylated proteins in cells expressing lung cancer-specific alleles of EGFR and KRAS. Proceedings of the National Academy of Sciences U.S.A. 105 (37), 14112-14117 (2008).
- Furuyama, K., et al. Sensitivity and kinase activity of epidermal growth factor receptor (EGFR) exon 19 and others to EGFR-tyrosine kinase inhibitors. Cancer Science. 104 (5), 584-589 (2013).
- Qiu, C., et al. Mechanism of Activation and Inhibition of the HER4/ErbB4 Kinase. Structure. 6 (3), 460-467 (2008).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеThis article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены