
Method Article
Количественный анализ визуализации in vitro для фагоцитоза мертвых клеток нейробластомы iPSC-макрофагами
В этой статье
Резюме
Нейродегенеративные заболевания связаны с дисрегулируемыми функциями микроглии. В данной статье описан анализ in vitro фагоцитоза клеток нейробластомы iPSC-макрофагами. Количественные показания микроскопии описаны как для покадровой визуализации живых клеток, так и для визуализации с высоким содержанием в фиксированных клетках.
Аннотация
Микроглия организует нейроиммунные реакции при нескольких нейродегенеративных заболеваниях, включая болезнь Паркинсона и болезнь Альцгеймера. Микроглия очищает мертвые и умирающие нейроны через процесс эффероцитоза, специализированной формы фагоцитоза. Функция фагоцитоза может быть нарушена экологическими или генетическими факторами риска, которые влияют на микроглию. В данной работе представлен быстрый и простой протокол микроскопии in vitro для изучения эффероцитоза микроглии в модели микроглии индуцированных плюрипотентных стволовых клеток (iPSC) с использованием клеточной линии нейробластомы человека (SH-SY5Y), помеченной pH-чувствительным красителем для фагоцитарного груза. Процедура приводит к высокому выходу мертвых клеток нейробластомы, которые отображают поверхностный фосфатидилсерин, распознаваемый фагоцитами как сигнал «съесть меня». 96-скважинный пластинчатый анализ подходит для покадровой визуализации живых клеток, или пластина может быть успешно зафиксирована перед дальнейшей обработкой и количественно определена с помощью микроскопии с высоким содержанием. Микроскопия с высоким содержанием фиксированных клеток позволяет масштабировать анализ для скрининга ингибиторов малых молекул или оценки фагоцитарной функции генетических вариантов линий iPSC. Хотя этот анализ был разработан для изучения фагоцитоза целых мертвых клеток нейробластомы iPSC-макрофагами, анализ может быть легко адаптирован для других грузов, имеющих отношение к нейродегенеративным заболеваниям, таких как синаптосомы и миелин, а также другие типы фагоцитарных клеток.
Введение
Микроглия является макрофагами- резидентами ткани мозга, и их функции включают иммунный надзор, координацию воспалительных реакций на травму / инфекцию, синаптическое ремоделирование и фагоцитоз мертвых клеток, миелина, белковых агрегатов и патогенов. Фагоцитоз — это процесс, с помощью которого микроглия распознает груз с поверхностными рецепторами и реорганизует их цитоскелет, чтобы поглотить объект в фагосому, которая затем сливается с лизосомами для деградации груза. Здоровая микроглия фагоцитозы апоптотических клеток мозга удаляет их до того, как они станут некротическими1. Фагоцитоз апоптотических клеток также известен как эффероцитоз и требует отображения сигнала фосфатидилсерина «съесть меня» умирающей клеткой2. Многочисленные рецепторы микроглии напрямую связываются с фосфатидилсерином, включая TIM-4, BAI1, Stabilin-2 и TREM2. Микроглиальные TAM-рецепторы (например, MERTK) и интегрины косвенно связываются с фосфатидилсерином, используя вспомогательные белки GAS6 или MFG-E8 соответственно. Другие сигналы «съесть меня» могут быть необходимы для распознавания умирающих клеток, к ним относятся изменения гликозилирования или заряда поверхностных белков; экспрессия внутриклеточных белков ICAM3, калретикулина, аннексина-I на поверхности клетки; окисленные ЛПНП; или покрытие апоптотической клетки микроглией, продуцируемым комплементом C1q1,2.
Нейродегенеративные заболевания, включая болезнь Паркинсона, болезнь Альцгеймера, лобно-височную деменцию и боковой амиотрофический склероз, были связаны с нарушением функции микроглии, включая накопление продуктов жизнедеятельности мозга, таких как мертвые клетки, фрагменты миелина и белковые агрегаты, и преувеличенные воспалительные реакции на эти стимулы3. Фагоцитоз может быть нарушен при нейродегенеративных заболеваниях и способствовать патологии, обусловленной сочетанием старения, воспаления или специфических вариантов генетического риска4,5. С другой стороны, есть также данные из животных моделей нейродегенеративных заболеваний, что микроглия может ненадлежащим образом фагоцитозировать жизнеспособные нейроны или синапсы6,7,8. Механизм, вероятно, будет спровоцирован фосфатидилсериновой дисплеем поврежденных нейритов, который непосредственно ощущается рецепторами фагоцитоза микроглии TREM2 или GPR56 или косвенно ощущается растворимым комплементом C1q, покрывающий обогащенную фосфатидилсерином мембрану, что приводит к CR3-опосредованный фагоцитоз9,10,11.
In vitro анализы функции фагоцитоза, например, для оценки фенотипического воздействия варианта генетического риска в микроглии, часто выполняются с использованием нефизиологических грузов, таких как латексные шарики4. Также используются флуоресцентно меченые бактерии и зимозан, которые являются физиологическими, но не имеют отношения к нейродегенеративным заболеваниям. Нефизиологические фагоцитарные грузы могут быть использованы для обнаружения дефектов в основном механизме фагоцитарного поглощения, но не могут точно смоделировать первый шаг «распознавания» в фагоцитозе апоптотических нейронов. Размер, форма, жесткость и тип груза также диктуют внутриклеточные сигнальные пути, которые активируются, что приводит к различным результатам состояния активации микроглии. Например, бактерии E.coli маленькие и жесткие, в отличие от клеток человека, а липополисахариды на их поверхности распознаются Toll-подобным рецептором 4 (TLR4), который активирует фагоцитоз и провоспалительные сигнальные пути2,12.
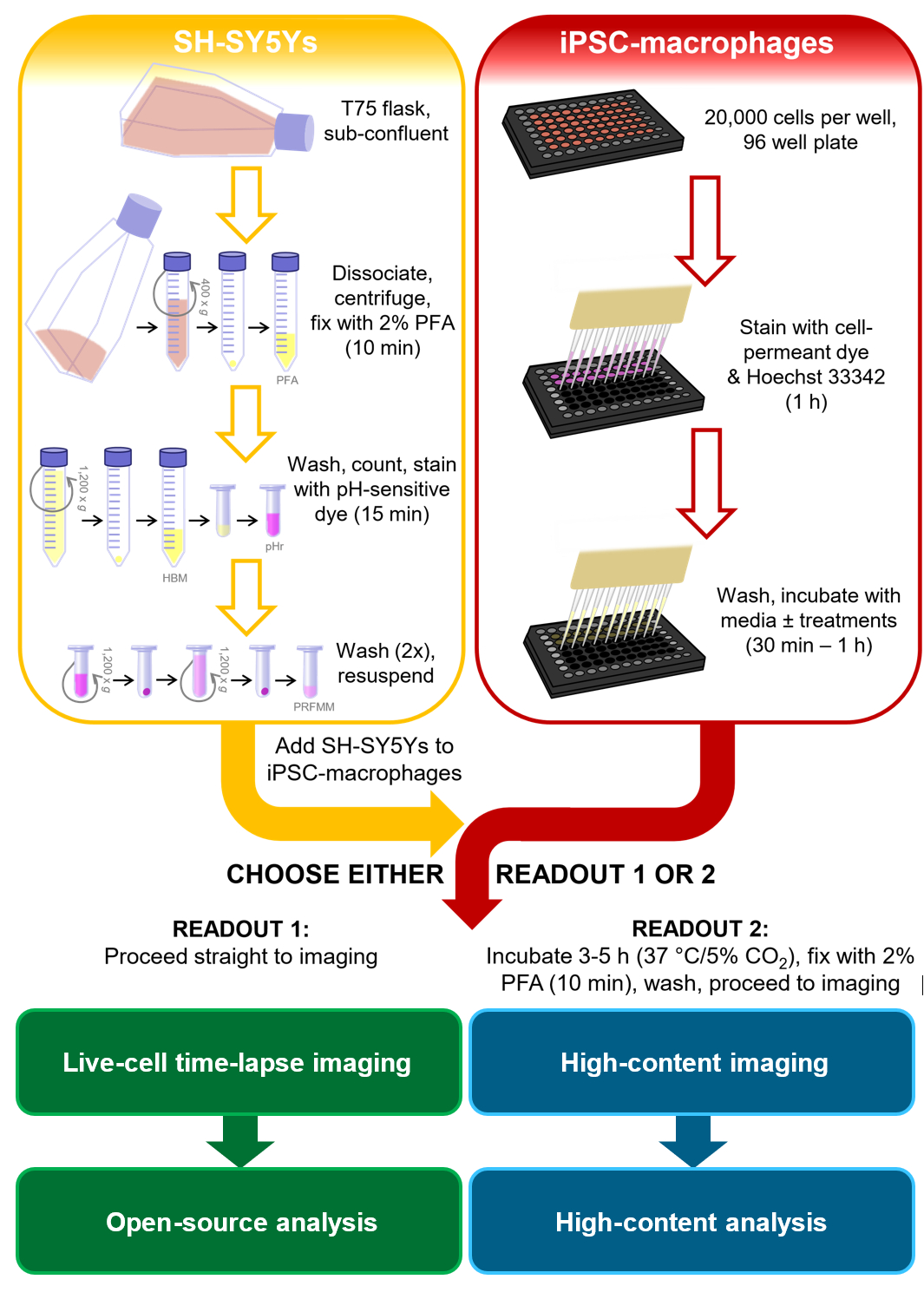
В контексте исследований нейродегенеративных заболеваний более релевантный фагоцитарный груз будет иметь фосфатидилсерин на плазматических мембранах млекопитающих и в идеале будет человеческим и нейронным, включая сигналы, с которыми может столкнуться микроглия. Для этого протокола фагоцитоза клеточная линия нейробластомы человека SH-SY5Y была выбрана в качестве модели нейрона, которую легко культивировать. Постоянный поверхностный дисплей фосфатидилсерина был искусственно индуцирован параформальдегидом, который, как было ранее показано, вызывает фосфатидилсериновый дисплей тромбоцитов13. Для модели клеток микроглии человека использовались iPSC-макрофаги, которые имитируют онтогенез и транскрипционный профиль микроглии человека и являются фагоцитически компетентными14,15,16,17. iPSC-макрофаги не являются самой аутентичной моделью микроглии, например, они не имитируют морфологию микроглии; однако при желании его можно заменить более аутентичной монокультурной iPSC-моделью микроглии, такой как Haenseler et al.15. Модели iPSC человека предпочтительнее первичных микроглий грызунов для изучения нейродегенерации из-за опасений по поводу ограниченного перекрытия транскрипционных модулей микроглии, наблюдаемых в тканях нейродегенеративных заболеваний человека имыши18. Мертвые SH-SY5Y окрашиваются кислотно-чувствительным красителем, который слабо флуоресцирует при нейтральном рН и сильнее внутри фаголизосом iPSC-макрофагов после фагоцитоза. Использование кислоточувствительного красителя повышает точность обнаружения фагоцитарных событий, с универсальностью для различных считывания живых и фиксированных макрофагов19. Этот протокол описывает как покадровую визуализацию фагоцитоза живыми клетками, так и фиксированный анализ изображений с высоким содержанием для фагоцитоза с теми же этапами подготовки клеток до считывания(рисунок 1).

Рисунок 1:Принципиальная схема методологии. Контур анализа фагоцитоза, где параллельно проводится подготовка SH-SY5Ys и окрашивание iPSC-макрофагов, а затем SH-SY5Ys пипетируются на iPSC-макрофаги. Либо покадровая визуализация живых клеток выполняется немедленно, либо клетки инкубируют при 37 °C / 5% CO2 в течение требуемой продолжительности и фиксируют перед выполнением микроскопии с высоким содержанием. PFA: параформальдегид, HBM: фенольные красные буферизованные среды HEPES, pHr: pH-чувствительный красный флуоресцентный раствор красителя STP Ester, PRFMM: фенольные среды макрофагов без красного цвета. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
протокол
Протокол следует руководящим принципам использования человеческих iPS-клеточных линий, полученных в Оксфордском университете, Оксфордском центре болезни Паркинсона (Комитет по этике: Национальная служба здравоохранения, Управление исследований в области здравоохранения, Комитет NRES South Central, Беркшир, Великобритания (REC 10/H0505/71)). Человеческие ИПСК должны обрабатываться в шкафу безопасности класса II, чтобы защитить работника от возможных адвентитивных агентов. Должны соблюдаться местные, национальные и ес правила охраны труда и техники безопасности. Композиции сред клеточных культур подробно описаны в таблице 1,а все материалы перечислены в дополнительной Таблице материалов.
1. Клеточная культура перед экспериментом
- Культивируемые иПСК в средах иПСК(таблица 1)в 6-скважинных пластинах, предварительно покрытых матрицей базальной мембраны, сертифицированной hESC, субслеженной и с малым проходным числом.
- Дифференцировать человеческие ИПСК к предшественникам iPSC-макрофагов: посеять четыре миллиона iPSC в микролунку с низкой адгезией 24-скважинной пластины с 2 мл эмбриоидной среды тела(Таблица 1)для стимулирования формирования эмбрионального тела и выполнения 75% изменений среды ежедневно в течение 5-6 дней. Перенос эмбриоидных тел в колбы T175, приблизительно 150 эмбриоидных тел на колбу, содержащую 20 мл заводских сред(таблица 1). Кормят еженедельно добавлением 10-20 мл заводских носителей.
ПРИМЕЧАНИЕ: Предшественники iPSC-макрофагов появляются в надводном веществе примерно через 2-3 недели и производятся непрерывно в течение нескольких месяцев. Для этого эксперимента предпочтительно использовать клетки примерно через 6 недель после установки фабрик дифференцировки. Ранее собранные iPSC-макрофаги могут сохранять некоторую пролиферативную способность и менее адгезивны, препятствуя даже посеву при низкой плотности клеток. Верхний возрастной предел способности фагоцитоза не определен. - Дифференцировать предшественники iPSC-макрофагов к iPSC-макрофагам: собирать прекурсоры путем удаления необходимого объема надпомножателя; пропустить его через клеточный сетчатый фильтр 40 мкм для удаления комков; центрифуга по 400 х г в течение 5 мин к грануляционным клеткам и повторное суспендирование в макрофаговы(таблица 1). Семена iPSC-макрофагов по 20 000-30 000 клеток на скважину в 96-скважинной микропластине, обработанной культурой тканей (TC), с черными стенками скважины и оптически чистым дном, в 100 мкл среды макрофагов на скважину. Избегайте краевых колодцев и заполняйте их PBS; это важно для уменьшения влияния испарения на анализ. Дифференцировать в течение 6-10 дней путем инкубации при 37 °C/5% CO2.
ПРИМЕЧАНИЕ: Для этого анализа использовалась линия iPSC BIONi010-C (ECACC ID: 66540023); однако может быть заменена другая линия iPSC. - Поддерживают SH-SY5Ys до субсовпадения в колбах T75 с 20 мл среды SH-SY5Y(таблица 1),проходя каждые 3-4 дня.
| Имя | Базовые носители | Добавка, конечная концентрация |
| Носители iPSC | мТеСР1 | - |
| Эмбриоидные среды тела | мТеСР1 | БМП4, 50 нг/мл |
| VEGF, 50 нг/мл | ||
| СКФ, 20 нг/мл | ||
| Заводские носители | XVIVO15 | ГлутаМАКС, 2 мМ |
| Пенициллин, 100 ед/мл | ||
| Стрептомицин, 100 мкг/мл | ||
| 2-меркаптоэтанол, 50 мкМ | ||
| Ил-3, 25 нг/мл | ||
| М-ликвор, 100 нг/мл | ||
| Макрофаговы | XVIVO15 | ГлутаМАКС, 2 мМ |
| Пенициллин, 100 ед/мл | ||
| Стрептомицин, 100 мкг/мл | ||
| М-ликвор, 100 нг/мл | ||
| SH-SY5Y носитель | ДМЕМ/Ф12 | Фетальная бытовая сыворотка, 10% |
| Пенициллин, 100 ед/мл | ||
| Стрептомицин, 100 мкг/мл |
Таблица 1: Медиа-рецепты.
Составляющие клеточных культурных сред, используемых в протоколе. Более подробную информацию о компонентах носителей можно найти в Таблице материалов.
2. Приготовление мертвых SH-SY5Ys
- В шкафу биологической безопасности класса II диссоциируют SH-SY5Ys путем добавления 4 мл буфера диссоциации клеток, содержащего рекомбинантные трипсиноподобные ферменты и 1,1 мМ ЭДТА (см. Таблицу материалов),которые должны быть немедленно удалены таким образом, чтобы менее 1 мл оставалось в виде тонкой пленки, покрывающей клетки. Инкубировать в течение 2-3 мин при 37 °C / 5% CO2.
- Добавьте 10 мл HBSS в колбу T75 для промывки и переложите SH-SY5Ys в коническую центрифужную трубку размером 15 мл. Центрифуга при 400 х г в течение 5 мин. Аспирировать супернатант и повторно суспендировать клетки в 2 мл фенольной безкрасной HEPES-буферной среды (см. Таблицу материалов). Убедитесь, что гранулы аккуратно повторно суспендированы, пипеткой 100-1000 мкл, чтобы разбить комки перед фиксацией.
- Фиксируйте клетки, добавляя в трубку 2 мл 4% параформальдегида (конечная концентрация 2%). Инкубировать в течение 10 мин при комнатной температуре с периодическим мягким перемешиванием тюбика.
- Добавьте 10 мл HBSS в пробирку. Центрифуга при 1,200 х г в течение 7 мин и повторное суспендировать в 2 мл фенольного красного цвета HEPES-буферной среды.
ПРИМЕЧАНИЕ: После этапа 2.4 препарат с фиксированным SH-SY5Y может контролироваться по качеству путем окрашивания приложением V-FITC для получения доступного фосфатидилсерина и йодида пропидия для измерения проницаемости клеток с помощью показаний проточной цитометрии. Сравните фиксированный препарат с живым SH-SY5Ys, полученным из шага 2.2. См. раздел 7 и дополнительный рисунок S1. Хранение фиксированных SH-SY5Ys после шага 2.4 не рекомендуется, так как это не было оценено.
3. Маркировка мертвых SH-SY5Ys pH-чувствительным красным флуоресцентным красителем
- После шага 2.4 подсчитайте клетки и удалите общее количество необходимых клеток в 2 мл низкобелк-связывающей трубки. На каждый 1 миллион SH-SY5Ys составьте общий объем в пробирке 2 мл до 300-500 мкл с фенольными красными HEPES-буферными средами. Ненадолго нагрейте трубку на водяной бане с 37 °C.
- Воссоздайте pH-чувствительный красный флуоресцентный краситель STP эфир (см. Таблицу материалов)и добавьте 12,5 мкг красителя на миллион SH-SY5Y в теплую 2 мл пробирку клеток. Аккуратно перемешайте путем пипетки. Инкубировать трубку при комнатной температуре в течение 30 мин, защищенных от света.
ПРИМЕЧАНИЕ: Эфир STP-чувствительных красителей вступает в реакцию с первичными аминами, и поэтому буфер маркировки не должен содержать свободных аминов. Из-за потенциально ограниченной растворимости в водных буферах добавляйте растворенный в ДМСО краситель только в теплый водный буфер, немедленно перемешайте и исследуйте на наличие признаков осадка (темных частиц под световым микроскопом). - Добавьте 1 мл HBSS и центрифугу при 1200 х г в течение 7 мин при 4 °C. Выбросьте супернатант и промойте 2 мл HBSS. Центрифугирование повторить.
- Выбросьте супернатант и повторно суспендировать клетки в фенольной среде макрофагов без красных (см. Таблицу материалов)до концентрации 200 000-1,2 миллиона клеток/мл, так что 50 мкл составляет 10 000-60 000 клеток (т.е. в 0,5x-3 раза больше SH-SY5Ys, чем iPSC-макрофагов).
ПРИМЕЧАНИЕ: Фенол красный в средах увеличивает фоновую флуоресценцию, и поэтому для визуализации живых клеток следует использовать фенольный ред-жид. Хранить окрашенные SH-SY5Ys более нескольких часов не рекомендуется, так как это не было оценено. Храните окрашенные SH-SY5Ys на льду и защищайте от света.
4. Окрашивание iPSC-макрофагов
- В шкафу биологической безопасности готовят раствор в макрофаговой среде темно-красного флуоресцентного, клеточно-проницаемого, сукцинимидилового эфиро-реакционноспособного красителя (см. Таблицу материалов). Добавить Hoechst 33342 (см. Таблицу материалов). Прогрейте рабочий раствор до 37 °C на водяной бане.
- Аспирировать iPSC-макрофаговую среду осторожно, пипетируя клеточный супернатант многоканальной пипеткой в стерильный резервуар. Добавьте 70 мкл/целую часть раствора красителя, приготовленного на этапе 4.1, к iPSC-макрофагам, используя многоканальную пипетку. Инкубировать в течение 1 ч при 37 °C/5% CO2.
- Подготовьте экспериментальные методы лечения в фенольных средах макрофагов без красного цвета. Включите 10 мкМ цитохалазина D в качестве отрицательного контрольного лечения. После инкубации аспирировать iPSC-макрофагальную среду очень осторожно с помощью многоканальной пипетки и добавить 100 мкл / цель буферного солевого раствора Хэнка (HBSS) для промывки. Немедленно удалите HBSS путем бережного пипетирования, затем добавьте 100 мкл среды ± соединений. Инкубировать в течение 10 мин-1 ч при 37 °C/ 5% CO2.
ПРИМЕЧАНИЕ: Цитохалазин D является мощным ингибитором актина и блокирует фагоцитоз. Для любых экспериментальных методов лечения, требующих более длительной инкубации, например, 24-72 ч, проводят экспериментальную обработку до этапа 4.1, используя 100 мкл/велл обработки в полной среде макрофагов. Следуйте шагам 4.1-4.3 в соответствии с протоколом, чтобы было выполнено окрашивание клеток, а затем лечение повторно применено в фенольных красных макрофагов для остальной части анализа фагоцитоза.
5. Визуализация фагоцитоза
Ниже приведены два различных метода считывания фагоцитоза, выберите подраздел 5.1 или 5.2.
- Покадровая визуализация в живых клетках
- Перед фагоцитозом включите микроскоп покадровой визуализации живых клеток (см. Таблицу материалов),компьютер, камеру окружающей среды и газ CO2. Открытое программное обеспечение для захвата изображений. Убедитесь, что в микроскоп установлены световые кубики DAPI, RFP и CY5. Нажмите на | Time Lapse Инкубационная | Включите камеру окружающей среды и выберите нагреватель до 37 ° C с газом CO2, а также убедитесь, что влажность не выбрана. Дайте 30 минут, чтобы микроскоп нагрел до 37 °C.
- Во время инкубации соединения на этапе 4.3 загрузите пластину iPSC-макрофага в микроскоп.
- Нажмите на | изображений Захват | Эксперт по судам. Выберите Плиту скважины и выберите тип плиты на 96 скважин.
- На вкладке Изображение включите фазовый канал и настройте грубую и тонкую фокусировку с помощью вертикальных ползунков, чтобы ячейки были в фокусе. Отрегулируйте уровень освещения с помощью горизонтального ползунка. Нажмите на каналы DAPI, RFP и CY5 и отрегулируйте уровни освещения для каждого канала.
- На вкладке «Система» нажмите «Калибровка выравнивания сосуда» и следуйте инструкциям на экране.
- Нажмите на | Time Lapse Процедуры | Создайте новую процедуру. На первом экране мастера замедленной съемкиназовите процедуру. Нажмите далее. На втором экране выберите цель 20x, выберите Монохромный захват и выберите каналы DAPI, RFP, CY5 и Phase. Не выбирайте следующие параметры: Авто найти образец, Авто тонкая фокусировка, Z-Стек, Автоматическое освещение. Нажмите далее.
- На следующем экране установите маяк в центре каждой скважины, который позволит микроскопу возвращаться к тем же координатам с одинаковыми настройками освещения для каждой точки времени. Настройки фокусировки и освещения для каждого маяка независимы. Чтобы установить маяк: перетащите синий круг в место на карте пластины, используйте вертикальный ползунок грубой и тонкой фокусировки, а когда вы удовлетворены, нажмите «Добавить маяк». Настройки маяка можно обновить позже с помощью кнопки Обновить выбранное.
- Когда вы будете готовы начать анализ фагоцитоза, снимите пробирную пластину и поместите ее в шкаф биологической безопасности. Используйте многоканальную пипетку, чтобы добавить 50 мкл SH-SY5Ys на скважину, добавляя в сторону каждой скважины на краю жидкости.
- Загрузите пластину в микроскоп и подождите примерно 30 минут, пока тепловой сдвиг не уравновесятся.
ПРИМЕЧАНИЕ: В течение первых 30 минут, когда пластина находится в микроскопе, изменение температуры пробирной пластины приведет к смещению фокуса. Если пластине не позволено уравновешиваться, захваченные изображения будут смещаться из фокуса в течение таймлапса. - Нажмите на каждый маяк и обновите настройку фокуса. Нажмите далее. На следующем экране мастера замедленной съемкивыберите формат файла TIFF,включите параметр Сохранять отдельные каналыи включите параметр Создать видео для каждого маяка,а также разрешите параметры в разделе Включить следующую информацию в качестве водяного знака. Нажмите далее.
- Установите для числа сцен значение 1. Нажмите далее. Установите продолжительность и интервалы интервалов, например, 3 ч и визуализацию каждые 5 мин. Не выбирайте «Захват только одного кадра». Нажмите далее.
- Включите камеру окружающей среды с температурой 37 °C и CO2 (влажность необязательна для коротких экспериментов). Нажмите далее дважды. Выберите путь для сохранения данных. Нажмите далее. Нажмите кнопку Пуск, чтобы начать таймлапс.
- Визуализация с высоким содержанием фиксированных клеток
- Используйте многоканальную пипетку, чтобы добавить 50 мкл меченого SH-SY5Ys на скважину, добавляя на сторону каждой скважины на краю жидкости. Инкубировать при 37 °C/ 5% CO2 в течение 3-5 ч.
- После инкубации фагоцитоза осторожно аспирировать клеточные супернатанты путем пипетки многоканальной пипеткой и выбросить. Вымойте один раз со 100 мкл PBS.
- Зафиксируйте пластину добавлением 100 мкл 2% параформальдегида, инкубируйте в течение 15 мин при комнатной температуре.
- Аспирировать скважины и добавить 100 мкл PBS. Крышка с пластинчатым герметителем и фольгой; хранить при 4 °C до тех пор, пока это потребуется.
ПРИМЕЧАНИЕ: Пробирная пластина может храниться таким образом в течение не менее недели без значительного ухудшения сигнала; более длительное хранение не было протестировано. - Включите микроскоп с высоким содержанием изображений (см. Таблицу материалов)и откройте программное обеспечение для захвата изображений. Загрузите пробирную пластину в микроскоп, нажав на значок «Загрузить» в верхней части экрана.
- Выберите вкладку Настройка. В выпадающих меню левого верхнего поля: выберите соответствующий тип пластины, выберите опцию автофокусировки Два пика (по умолчанию),выберите цель 40x Water, NA1.1, выберите Конфокальный режим и выберите биннинг 1.
- Промывайте 40-кратный объект воды перед использованием через меню «Настройки».
- В поле Выбор канала используйте значок +, чтобы добавить каналы DAPI, Alexa 647 и Alexa 568. Установите их для измерения в одной плоскости 1 мкм. Оптимизируйте настройки времени и мощности для повышения эффективности окрашивания пробирной пластины.
ПРИМЕЧАНИЕ: В качестве ориентира установите DAPI на 200 мс экспозиции и 100% мощности, Alexa 647 на 1500 мс экспозиции и 100% мощности, а Alexa 568 на 100 мс экспозиции и 40% мощности. - Убедитесь, что каналы не измеряются одновременно, щелкнув Последовательность каналов, чтобы разделить каналы.
- В разделе Навигационные | Определите макет,выберите скважины для измерения и выберите 9-12 месторождений на скважину.
- Во время настройки нажмите на репрезентативное поле на карте пластины и проверьте каждый измерительный канал по очереди, чтобы убедиться, что окрашивание присутствует и что изображения сфокусированы, регулируя смещение канала.
- Чтобы загрузить данные на сервер для удаленного анализа, нажмите на поле Online Jobs (Онлайн-задания) и соответствующее имя экрана; это позволит автоматически загружать данные на сервер после создания образа.
- Сохраните протокол анализа, нажав на кнопку Сохранить.
- Нажмите на вкладку «Выполнить эксперимент» в верхней части и назовите пластину эксперимента, затем нажмите «Пуск».
6. Анализ данных
Ниже приведены два различных метода анализа данных: выберите подраздел 6.1, если был соблюдена подраздел 5.1, или подраздел 6.2, если был выполнен подраздел 5.2.
- Анализ изображений фагоцитоза, полученных с помощью живоклеточного покадрового микроскопа
- Загрузите и установите рекомендуемое программное обеспечение с открытым исходным кодом (см. Таблицу материалов). Откройте программное обеспечение.
- В поле Модули ввода выберите Изображения.
- В проводнике Windowsоткройте папку данных, содержащую подпапки с именами Beacon-1, Beacon-2 и т.д. Выберите и перетащите все папки маяков в список Файл.
- В поле Модули ввода выберите Метаданные. Для параметра Извлечь метаданные?выберите Да. В раскрывающемся меню рядом с пунктом Метод извлечения метаданныхвыберите «Извлечь из имен файлов/папок». В поле Источник метаданныхвыберите Имя папки. Нажмите на увеличительное стекло справа от регулярного выражения и введите ".*[\.*](? P.*)$в текстовое поле Regex (исключая кавычки). Нажмите кнопку Отправить. В параметре «Извлечь метаданные из»выберите «Все изображения». Нажмите «Обновить» в нижней части экрана. Изображения теперь будут сгруппированы по маяку.
- В поле Модули ввода выберите NamesAndTypes. Следующий процесс позволит назначить изображения для каждой временной точки правильному каналу флуоресценции. Присвойте имя Правилам сопоставления изображений (раскрывающееся меню). Выберите критерий правила, соответствующий всем (раскрывающееся меню) следующих правил. Файл (раскрывающееся меню), Делает (раскрывающееся меню), Contain (раскрывающееся меню), DAPI (текстовое поле). Имя, присваиваемые этим изображениям DAPI (текстовое поле). Выберите тип изображения Оттенки серого Image (выпадающее меню). Задайте диапазон интенсивности в основе метаданных изображения (раскрывающееся меню).
- В нижней части экрана нажмите «Добавить еще одно изображение»и повторите шаг 6.1.5. Замените DAPI на RFP,чтобы изображения RFP были сгруппированы.
- Повторите шаг 6.1.6 для изображений канала CY5.
- Нажмите «Обновить» в нижней части экрана, файлы изображений теперь будут перечислены в трех столбцах с меткой DAPI, RFP и CY5.
- В поле Модули ввода выберите Группы. Для параметра Вы хотите сгруппировать изображения?выберите Да. В раскрывающемся меню Категория метаданныхвыберите Маяк.
- В поле Модули анализа щелкните правой кнопкой мыши пустое пространство, чтобы вызвать список всех модулей.
- Нажмите на Добавить | | обработки изображений EnhanceOrSuppressФунтерии. Выберите DAPI в первом раскрывающемся раскрывающемся списка в качестве входного изображения. Назовите выходное изображение "DAPIspeckles". Выберите тип операции Enhance и тип объекта Speckles, с размером объекта 20 пикселей. Выберите скорость и точность опции Fast/Hexagonal.
- Создайте новый модуль. Добавить | | обработки объектов ИдентификацияПервичных Объектов. Выберите DAPIspeckles из первого раскрывающегося списка в качестве входного изображения. Назовите первичные объекты «Ядрами». Введите типичный диаметр объектов в единицах от 10 до 35 пикселей; этот параметр можно оптимизировать. Выберите пороговую стратегию Global,метод пороговых значений RidlerCalvard,метод сглаживания Automaticи дайте пороговый поправочный коэффициент как 12 с нижней и верхней границами 0-1. Измените метод, чтобы различать слипнутые объекты на Shape, но оставьте другие параметры в настройках по умолчанию.
ПРИМЕЧАНИЕ: Ядра iPSC-макрофагов были примерно сегментированы на шаге 6.1.12 после этапа обработки изображений, который уменьшает диаметр и увеличивает контрастность ядер. Важно, чтобы отбирались только самые яркие ядра, поскольку SH-SY5Ys будут отображаться как более слабые ядра и будут ошибочно приняты за iPSC-макрофаги. Для корректировки доли ядра, которые выбраны, увеличивают или уменьшают пороговый поправочный коэффициент. На этапе тестирования сравните полученный выбор ядер с фазовым изображением маяка, где легко отличить iPSC-макрофаг от SH-SY5Y с помощью клеточной морфологии. - Создайте новый модуль. Добавить | | обработки изображений CorrectIlluminationСчитай . Выберите CY5 в первом раскрывающемся списка в качестве входного изображения. Назовите выходное изображение "IllumCY5". В разделе «Выбрать способ освещения»выберите «Фон» в раскрывающемся меню. Оставьте остальные параметры в настройках по умолчанию.
- Создайте новый модуль. Нажмите на Добавить | | обработки изображений Правильноимощное приложение. Выберите CY5 в первом раскрывающемся списка в качестве входного изображения. Назовите выходное изображение "CorrCY5". В разделе «Выбрать подсветку»выберите IllumCY5 в раскрывающемся меню. В разделе «Выбрать способ освещения»выберите «Разделить» в раскрывающемся меню.
ПРИМЕЧАНИЕ: Цель этапов 6.1.13-6.1.14 заключается в исправлении изменений в фоновом освещении изображений CY5, которые в противном случае препятствовали бы правильной сегментации ячеек. - Создайте новый модуль. Нажмите на Добавить | | обработки объектов ИдентификацияСекундарные Объекты. Выберите CorrCY5 в первом раскрывающемся списка в качестве входного изображения. Выберите Ядра в качестве входных объектов. Назовите вторичные объекты "Mac". Выберите способ идентификации как Расстояние - B. Выберите пороговую стратегию Global,пороговый метод RidlerCalvard,метод сглаживания No smoothing,и дайте пороговый поправочный коэффициент как 1 с нижней и верхней границами 0-1. Оставьте другие параметры в настройках по умолчанию.
ПРИМЕЧАНИЕ: Этот этап сегментации ячеек может потребовать оптимизации путем корректировки порогового поправого коэффициента для роста или сжатия границ ячеек. Эффективность сегментации также может быть улучшена за счет увеличения силы окрашивания iPSC-макрофагов или освещения светового куба CY5 во время визуализации. - Создайте новый модуль. Нажмите на Добавить | | обработки изображений EnhanceOrSuppressФунтерии. Выберите RFP из первого раскрывающегося списка в качестве входного изображения. Назовите выходное изображение "FilteredRFP". Выберите тип операции Enhance и тип объекта Speckles, с размером объекта 15 пикселей. Размер функции можно оптимизировать. Выберите скорость и точность опции Fast/Hexagonal.
- Создайте новый модуль. Нажмите на Добавить | | обработки объектов ИдентификацияПервичных Объектов. Выберите FilteredRFP в первом раскрывающемся раскрывающемся списка в качестве входного изображения. Назовите первичные объекты "pHr". Введите типичный диаметр объектов в единицах 5-20 пикселей. Выберите стратегиюпороговых значений Manual и введите пороговое значение вручную, например, 0,005. Измените метод, чтобы различать слипнутые объекты на Shape, но оставьте остальные параметры в настройках по умолчанию.
ПРИМЕЧАНИЕ: SH-SY5Ys были сегментированы на этапе 6.1.17 после этапа обработки изображения, который уменьшает диаметр и увеличивает контрастность пункты. Крайне важно выполнить ручное пороговое значение, так как интенсивность pH-чувствительного красителя увеличивается со временем в фагоцитированных частицах, а другие стратегии порогового значения будут искусственно завышать количество pH-чувствительных красителей пункты в ранние временные точки. Ручное пороговое значение должно быть скорректировано для каждого последующего экспериментального повтора с использованием тестового режима. - Создайте новый модуль. Нажмите на Добавить | | обработки объектов Связанные объекты. Выберите входные дочерние объекты pHr в раскрывающемся меню. Выберите входные родительские объекты Mac из раскрывающегося меню. Для параметра Рассчитать средние значения для каждого родителя для всех дочерних измерений?выберите Да. Не вычисляйте расстояния между дочерними и родительскими (Нет).
ПРИМЕЧАНИЕ: Шаг 6.1.18 связывает pH-чувствительный сигнал красителя с iPSC-макрофагами, что позволяет измерять среднее количество фагоцитированных объектов на iPSC-макрофаг. - Создайте новый модуль. Нажмите на Добавить | | обработки файлов ExportToSpreadsheet. Выберите разделитель столбца tab и добавьте префикс для имен файлов, чтобы указать номер маяка. Выберите конкретные измерения для экспорта, как указано ниже (этапы 6.1.19.1 - 6.1.19.4); оставляя другие параметры в настройках по умолчанию.
- | изображений Количество | Выберите pHr и Mac
- | изображений Имя файла
- | изображений Группа
- | Для Mac Детские | пЧ
- В поле Вывод нажмите Просмотреть настройки вывода. Создайте новую папку на рабочем столе для этого эксперимента и установите ее в качестве выходной папки по умолчанию.
- Сохранение | файлов конвейера СохранитьПроект как....
- Протестируйте и оптимизируйте конвейер на репрезентативном изображении, щелкнув Запустить тестовый режим в левом нижнем углу. Программа автоматически выбирает первое изображение для тестирования и каждый шаг конвейера можно просмотреть, нажав на символы глаза, что делает вывод видимым, а затем нажав на Run. Чтобы изменить маяк, используемый для тестирования, в верхней строке меню нажмите на Test | Выберите Группа изображений. Чтобы изменить изображение (точку времени) в маяке, в верхней строке меню нажмите на Test | Выберите Набор изображений. Параметры, которые необходимо оптимизировать, указаны в предыдущих шагах.
- Когда вы удовлетворены конвейером, нажмите на Exit Test Mode и нажмите на открытые символы глаз, чтобы закрыть их. Сохраните конвейер. Нажмите «Анализировать изображения», чтобы начать полный анализ изображений.
- Текстовые файлы, которые генерируются, могут быть открыты в виде электронной таблицы с соответствующим программным обеспечением для электронных таблиц, а файл с надписью «Изображение» будет содержать строку для каждой точки времени изображения со столбцами, представляющими параметры.
ПРИМЕЧАНИЕ: Count_Mac и Count_pHr представляют собой количество iPSC-макрофагов и количество идентифицированных pH-чувствительных объектов на изображении. Не используйте Count_pHr данные, так как количество включает тусклые флуоресцентные SH-SY5Y, которые не были фагоцитозированы. Столбец Mean_Mac_Children_pHr_Count принимает среднее количество фагоцитированных объектов pHr на Mac (шаг 6.1.18 RelateObjects) для отдельного изображения, т.е. отдельной временной точки маяка. - Расположите данные таким образом, чтобы каждый маяк был отдельным столбцом в электронной таблице, изображения располагались в виде строк хронологического порядка, с различными параметрами, занимающими разные листы книги электронной таблицы.
- Умножьте Mean_Mac_Children_pHr_Count измерений на Count_Mac, чтобы сгенерировать параметр Количество пятен на изображении. Рассчитайте средний Count_Mac для каждого маяка. Разделите число пятен на изображение на среднее Count_Mac для этого маяка, сгенерив параметр Количество пятен на ячейку.
ПРИМЕЧАНИЕ: Шаг 6.1.26 исправляет любые ошибочные флуктуации, которые могут возникнуть в количестве iPSC-макрофагов (Count_Mac), путем нормализации данных до среднего количества iPSC-макрофагов во всех временных точках маяка. - Назначьте время с начала фагоцитоза (в мин) каждой строке изображения.
- Генерировать средние и стандартные отклонения для реплицированных скважин/маяков. График количества пятен на клетку (ось Y) по отношению ко времени (ось X) для визуализации скорости фагоцитоза.

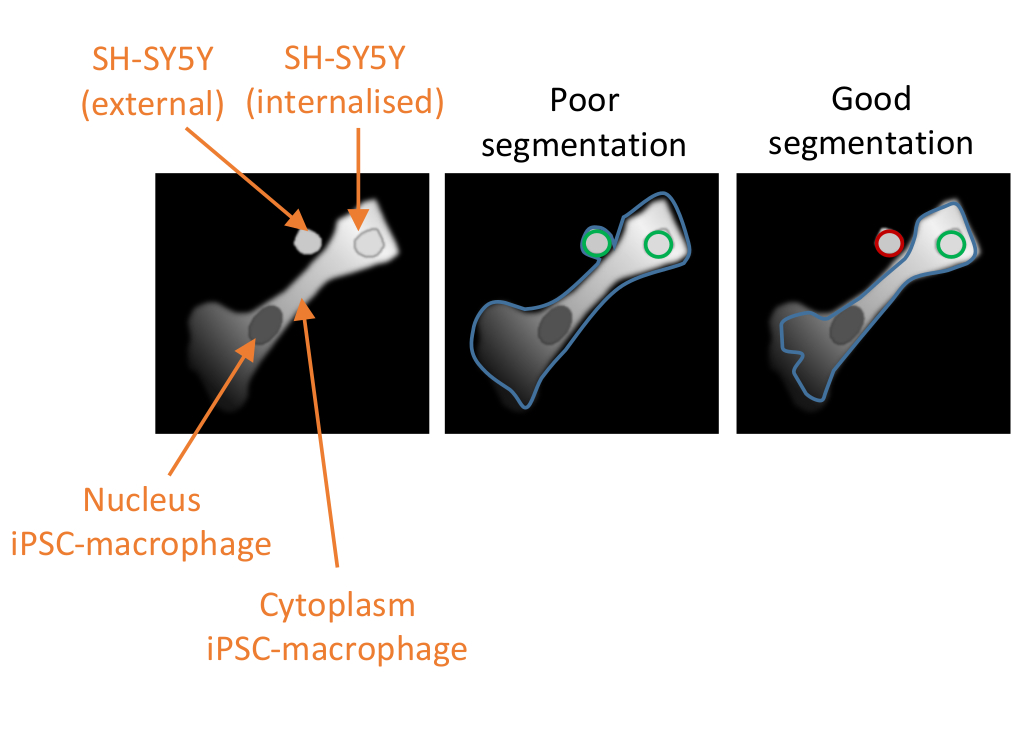
Рисунок 2:Сегментация клеток в анализе фагоцитоза с высоким содержанием. Иллюстрация, демонстрируя хорошую и плохую сегментацию iPSC-макрофага в непосредственной близости от нефагоцитозного SH-SY5Y, со вторым SH-SY5Y, полностью фагоцитозным. С обоими типами клеток, показанными серым цветом, граница клеток iPSC-макрофага, очерченная компьютерным анализом, очерчена (синий). SH-SY5Y, которые учитываются как события фагоцитоза, очерчены зеленым или красным цветом, если исключены из анализа. Изображение посередине показывает плохую сегментацию; iPSC-макрофаг имеет неоптимальное очертания, которое включает нефагоцитозный SH-SY5Y в пределах клеточной границы, что будет учитываться как событие фагоцитоза. Изображение справа показывает хорошую сегментацию из-за более строгих параметров, определяющих границу клеток iPSC-макрофага, что привело к тому, что нефагоцитозный SH-SY5Y был правильно исключен из анализа. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
- Анализ изображений фагоцитоза, полученных с помощью высокосодержимого микроскопа
- Войдите в рекомендуемое программное обеспечение для обработки изображений (см. Таблицу материалов).
- Выберите папку с именем экрана и вложенную папку запуска образа в меню слева. Нажмите на значок Image Analysis (экран с увеличительным стеклом). Выберите репрезентативную скважину на макете пластины для настройки аналитического трубопровода.
- Первым строительным блоком анализа является входное изображение. Оставьте настройки по умолчанию для обработки стека(Отдельные плоскости)и коррекции плоского поля(Нет). Нажмите на знак + в правом верхнем углу блока, чтобы добавить следующий строительный блок, и выберите Найти ядра.
- В Find Nucleiзадайте канал как DAPI,популяцию ROI как None,метод сегментации как C. Поле метода содержит раскрывающееся меню, которое позволяет оптимизировать параметр, с настройками общего порога (т.е. 0,40) и площади (т.е. >30 мкм2). Назовите выходную популяцию "Ядра". Добавьте следующий строительный блок, щелкнув символ + и выбрав «Найти цитоплазму».
- В Find Cytoplasmзадайте канал как Alexa 647 и метод как B. Поле метода содержит раскрывающееся меню, которое позволяет оптимизировать параметр с настройками общего порога (т.е. 0,45) и индивидуального порога (т.е. 0,20). Добавьте следующий строительный блок, щелкнув символ + и выбрав Выбрать население.
ПРИМЕЧАНИЕ: Крайне важно правильно оптимизировать сегментацию цитоплазмы таким образом, чтобы она исключала любые соседние SH-SY5Y, которые не были фагоцитозированы, но не исключала фагоцитозированный груз (см. Рисунок 2). - В разделе Выбрать популяциюсохраните параметры по умолчанию, которые будут заключаться в популяции Nuclei, методе Common Filters,галочкой Remove Border Objectsи выходной популяцией с именем "Nuclei Selected". Добавьте следующий строительный блок, щелкнув символ + и выбрав Рассчитать свойства морфологии.
- В разделе Вычисление свойств морфологиизадайте для совокупности значение Выбранные ядра,для области — Ячейка,для метода — значение Стандартный. В раскрывающемся меню убедитесь, что выбраны площадь и округляемость (мкм2). Назовите выходную популяцию «Морфологическая клетка». Добавьте следующий строительный блок, щелкнув символ + и выбрав Выбрать население.
- В разделе Выбор популяции (2)выберите выбранные ядра популяциии метод Фильтровать по свойствам. В раскрывающемся списках Фильтр F1выберите Область ячейки морфологии [мкм2]. Выберите > в раскрывающемся списка справа и введите 160 в поле справа от него. Назовите выходную популяцию "Ядра, выбранные 2". Добавьте следующий строительный блок, щелкнув символ + и выбрав Найти пятна.
ПРИМЕЧАНИЕ: Этот шаг исключает любые неправильно сегментированные клетки и любые мертвые клетки из дальнейшего анализа. Может возникнуть необходимость в оптимизации путем увеличения или уменьшения размера отсечки. - В разделе Найти пятнавыберите канал Alexa 568,ROI популяции Nuclei Selected 2,область ROI Cell,метод Bи назовите выходную популяцию «Spots». Метод может быть оптимизирован, при необходимости, с помощью выпадающего меню, с настройками чувствительности обнаружения (т.е. 0,20) и чувствительности разделения (т.е. 0,400). Добавьте следующий строительный блок, щелкнув символ + и выбрав Рассчитать свойства морфологии.
- В разделе Вычисление свойств морфологии (2)выберите точкипопуляции, область Пятнои метод Стандарт. В раскрывающемся меню убедитесь, что выбраны площадь и округляемость (мкм2). Назовите выходные свойства "Morphology Spot". Добавьте следующий строительный блок, щелкнув символ + и выбрав Выбрать население.
- В разделе Выбор популяции (3)выберите Точки популяции и метод Фильтровать по свойствам. В раскрывающемся списках в разделе Фильтр F1выберите Область пятна [px2], >, 20. В раскрывающихся списках в разделе Фильтр F2выберите Область пятна [px2], <, 2500. В раскрывающемся списках в разделе Фильтр F3выберите Округлость пятна морфологии, >, 0,6. В раскрывающихся списках в разделе Фильтр F4выберите Интенсивность по точкам региона, >, 2.5. Назовите выходное население "Выбранные пятна". Добавьте следующий строительный блок, щелкнув символ + и выбрав Выбрать население.
ПРИМЕЧАНИЕ: Автоматизированный выбор пятно будет сегментировать множество крошечных флуоресцентных пятнышков, которые являются результатом автофлуоресцентных тел в iPSC-макрофагах. Этот шаг направлен на фильтрацию автофлуоресцентных тел путем применения строгих отсечек к площади, округлости и интенсивности пятен и может потребовать некоторой оптимизации. - В разделе Выбор популяции (4)выберите ядра популяции Selected 2 и метод Фильтровать по свойствам. В раскрывающихся списках Фильтр F1выберите Количество мест, >, 0,5. Назовите выходную популяцию «Точечные положительные клетки». Добавьте следующий строительный блок, щелкнув символ + и выбрав Определить результаты.
- В разделе Определение результатоввыберите первый метод в качестве Списка выходных данных. По умолчанию используется количество объектов, вычисляемых для каждой совокупности. Щелкните раскрывающееся меню Население: Ядра выбраны 2 и убедитесь, что установлен флажок Количество объектов, а в раскрывающемся меню Применить ко всем выберите ВСЕ. Для популяции Spot Positive Cellубедитесь, что отмечено количество объектов. Для других популяций нет необходимости сообщать какие-либо параметры. Выберите второй метод в качестве Formula Outputи введите формулу (a/b)*100. Выберите как переменную A Spot Positive Cell- Количество объектов,а в качестве переменной B выберите Nuclei Selected 2- Number of Objects. Назовите выходные данные "Точечные положительные ячейки (%)".
- Сохраните конвейер: нажмите на иконку Сохранить анализ на диск (дискета со стрелкой вниз).
- Нажмите на иконку Пакетный анализ (символ воронки и винтиков в верхней части экрана). Из экспериментальных папок слева выберите файл необработанных данных, который должен обновить количество выбранных измерений до 1. В области Параметры анализа щелкните раскрывающееся меню Методи выберите Существующий анализ. Нажмите на ... рядом с полем Файл сценария и найдите сохраненный файл анализа (с суффиксом .aas). Затем нажмите на зеленую стрелку рядом с пунктом Начать анализ. Ход анализа можно отслеживать, нажав на Job Status (в правом верхнем углу экрана).
- После завершения анализа перейдите на вкладку Экспорт,выберите папку эксперимента и выберите папку назначения. Оставьте настройки по умолчанию, которые экспортируют данные, но не изображения TIFF, и начните экспорт.
- Откройте загруженный файл как электронную таблицу в соответствующем программном обеспечении для работы с электронными таблицами. Скважины расположены рядами, а параметры в столбцах. Выберите данные в столбцах с метками Точечные положительные ячейки (%), Выбранные ядра 2 - Количество пятен - Среднее на скважину и Выбранные ядра 2 - Общая площадь пятна - Среднее на скважину и скопируйте их в свежие электронные таблицы для каждого параметра. Рассчитайте среднее значение параметра для реплицированных скважин каждого условия и график при необходимости.
7. Контроль качества анализа однородности фиксированных SH-SY5Ys
- Собрать аликвоту живых SH-SY5Ys на этапе 2.2 и повторно суспендовать в буфере связывания аннексина из набора для окрашивания В-ФИТЦ приложения (см. Таблицу материалов)в концентрации приблизительно 200 000 клеток на мл.
- Собрать аликвоту фиксированных SH-SY5Ys со этапа 2.4 и повторно суспендировать в буфер связывания аннексина в концентрации приблизительно 200 000 клеток на мл.
- Подготовьте две пробирки с 5 мкл приложения V-FITC и 5 мкл йодида пропидия (см. Таблицу материалов). Добавьте 500 мкл живого SH-SY5Ys в одну трубку и 500 мкл фиксированных SH-SY5Ys в другую.
- Подготовьте три контрольные трубки: одну с 5 мкл приложения V-FITC, одну с 5 мкл йодида пропидия и одну трубку пустую. Смешайте соотношение 1:1 живых и фиксированных SH-SY5Y и добавьте 500 мкл этого в каждую контрольную трубку.
- Аккуратно перемешайте трубы путем пипетки. Инкубировать при комнатной температуре в течение 10 мин, защищенных от света.
- Немедленное измерение на проточном цитометре(Ex = 488 нм; Em = 530 нм) с использованием детектора сигналов FITC (обычно FL1) для приложения V-FITC и детектора сигналов излучения фикоэритрина (обычно FL2) для йодида пропидия.
- Используйте любое программное обеспечение для анализа проточной цитометрии для отображения точечных графиков сигнала FITC против PI и используйте прямоугольный инструмент для выбора двойной отрицательной популяции. В двойной отрицательной популяции отобразите FSC против SSC и используйте полигональный инструмент для создания исключающей вентили вокруг популяции с очень низким FSC и SSC, которая классифицируется как мусор и, следовательно, исключается из дальнейшего анализа. Отобразите остальные события в виде сигнала FITC vs PI и используйте одноокрашенные и незапятнанные элементы управления для установки квадрантного затвора для событий FITC-/PI-, FITC+/PI-, FITC -/PI+ и FITC+/PI+.
ПРИМЕЧАНИЕ: Избегайте грубого обращения, вихрей или длительных инкубаций с живым SH-SY5Ys, которые могут искусственно индуцировать отображение фосфатидилсерина. Приступайте к проточной цитометрии без задержек. Желательным результатом является то, что доля событий FITC-/PI- составляет <5% в фиксированных SH-SY5Ys. Репрезентативные результаты показаны на дополнительном рисунке S1.
Результаты
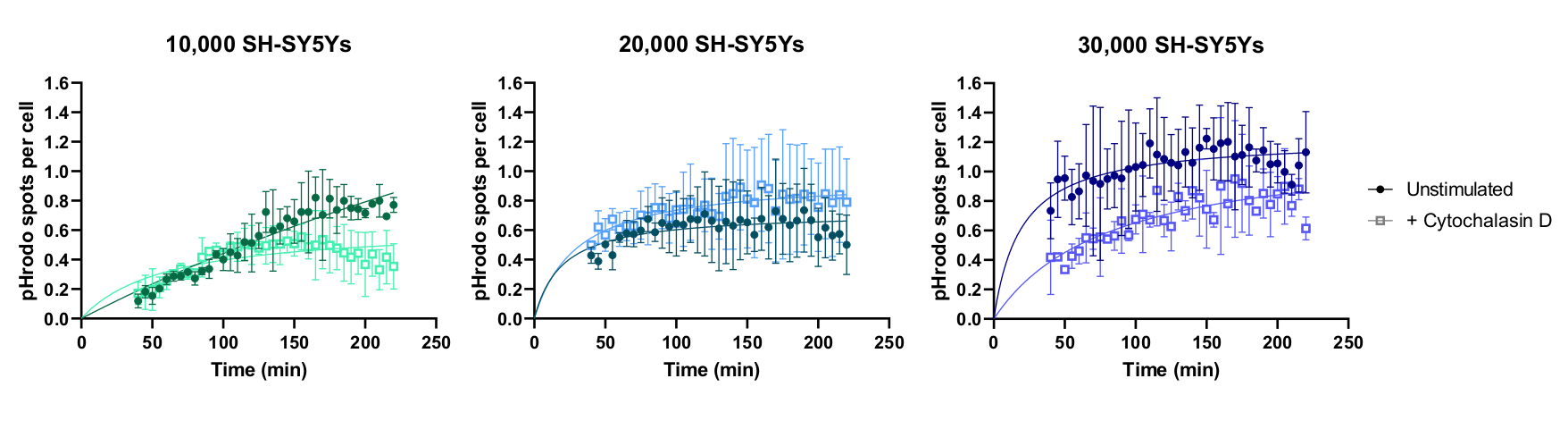
Покадровая визуализация живых клеток была выполнена с использованием ранее описанного протокола, с iPSC-макрофагами дикого типа, посеянными на 20 000 клеток на скважину. Применяли различные количества SH-SY5Ys (10 000-30 000 на скважину, по оценкам по количеству клеток на этапе 3.1), и ингибитор фагоцитоза цитохалазин D был предварительно инкубирован (1 ч) с некоторыми скважинами, действуя как контроль для ингибирования фагоцитоза для каждого количества SH-SY5Ys. Визуализация началась через 40 минут после добавления SH-SY5Ys, и изображения были сделаны с интервалом в 5 минут в течение следующих 3 ч (данные включают начальную задержку 40 минут). Репрезентативное покадровое видео включено в Дополнительные данные,а проанализированные количественные данные показаны на рисунке 3. При количестве 10 000 SH-SY5Ys на скважину количество фагоцитированных частиц (пятен) на клетку увеличивалось линейно со временем и ингибировалось примерно на 50% цитохалазином D. Ингибирование цитохалазином D было слабее, чем ожидалось, скорее всего, вызвано недостаточными техническими или биологическими репликациями, поскольку только одна скважина на состояние была изображена с тремя полями изображения. При более высоких количествах SH-SY5Ys на скважину (20 000 и 30 000) фагоцитоз проявлял плохую линейность, вероятно, из-за плохой сегментации iPSC-макрофагов и SH-SY5Ys в более переполненном поле зрения.
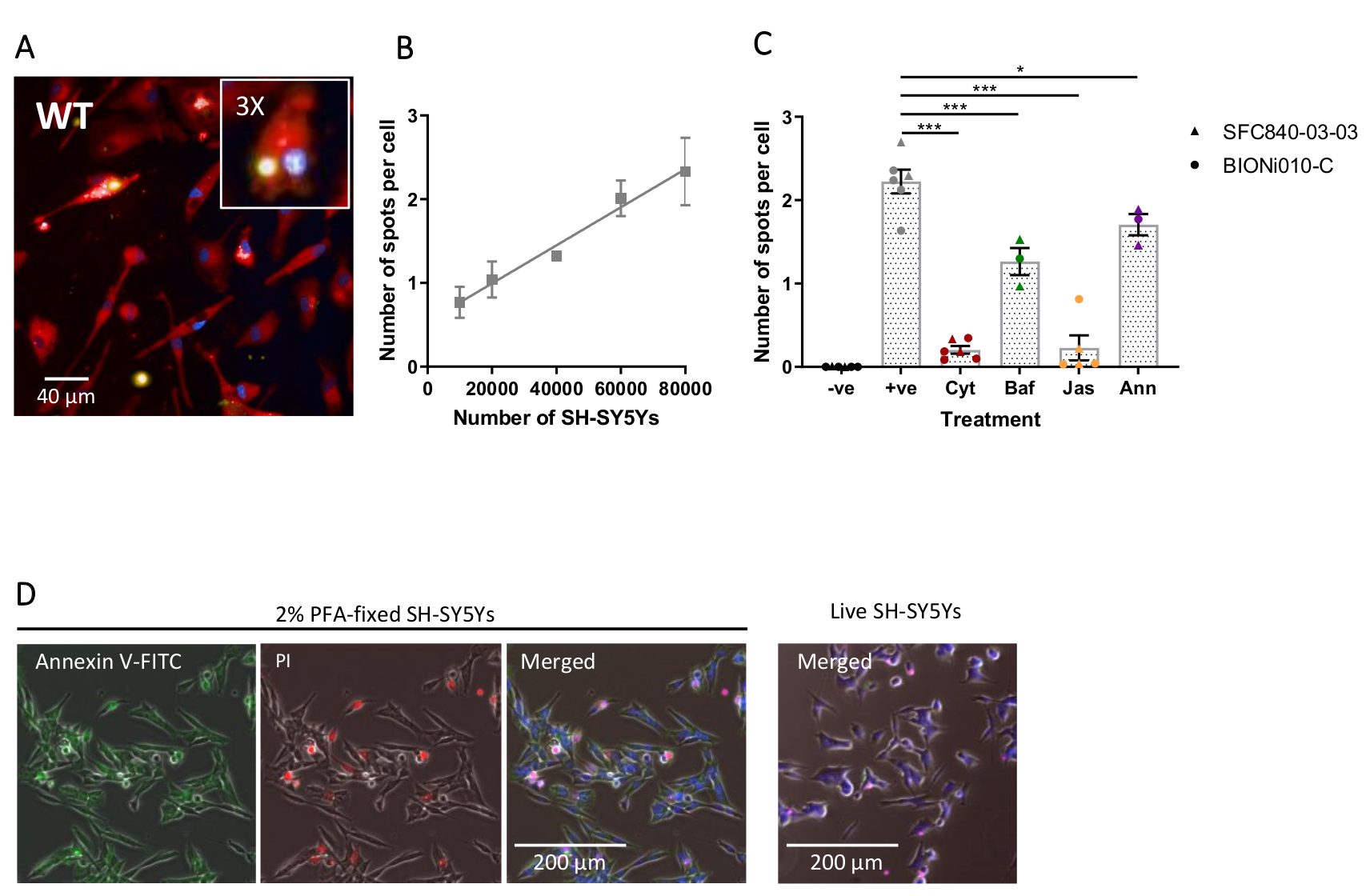
Визуализация с высоким содержанием фиксированных клеток была выполнена с использованием ранее описанного протокола, с iPSC-макрофагами дикого типа при 20 000 клеток на скважину, несколькими различными количествами SH-SY5Ys (10 000-80 000 на скважину), и анализная пластина была зафиксирована и визуалирована через 5 ч. Репрезентативное изображение фагоцитоза представлено на фиг.4А,а проанализированные данные показаны на фиг.4В17. Увеличение количества SH-SY5Ys приводило к увеличению количества фагоцитированных частиц (пятен) на клетку; однако удвоение количества SH-SY5Y приводит только к увеличению числа пятен на ячейку в 1,5 раза. Это указывает на то, что тестируемые количества не ограничивают скорость фагоцитоза. Впоследствии высокосоставленный визуализационный анализ фагоцитоза был подтвержден с использованием нескольких ингибиторов фагоцитоза(Рисунок 4C)17. Ингибиторы полимеризации актина цитохалазин D и джасплакинолид достоверно ингибировали фагоцитоз на 91% и 90% соответственно при предварительной инкубации в течение 1 ч до фагоцитоза. Надежный Z' анализа, когда цитохалазин D или джасплакинолид используются в качестве отрицательных контрольных элементов, рассчитывается как 0,7 и 0,8 соответственно20. Ингибитор подкисления лизосом бафиломицинА А1 достоверно уменьшал фагоцитоз на 31%, при инкубации за 1 ч до фагоцитоза. Более слабый эффект ингибитора подкисления лизосом по сравнению с ингибиторами актина предполагает, что обнаружение интернализованного груза может не требовать полного подкисления фагосомы. Рекомбинантный аннексин V использовался в качестве контроля для специфический блокирования фосфатидилсерина, подвергающегося воздействию на поверхность SH-SY5Ys, предотвращая доступ фагоцитарных рецепторов к лиганду, важному сигналу «съесть меня». Добавление рекомбинантного аннексина V значительно уменьшало фагоцитоз на 30%, при добавлении в колодцы непосредственно перед добавлением SH-SY5Y. Было подтверждено, что фиксированные SH-SY5Y подвергают воздействию фосфатидилсерина с использованием флуоресцентного зонда присоединения V, тогда как живые SH-SY5Y были отрицательными для окрашивания аннексина V(рисунок 4D).
Ранее было показано, что рецептор фагоцитоза микроглии TREM2 важен для фагоцитоза апоптотических нейронов21. Мутация R47H TREM2 является геном риска позднего начала болезни Альцгеймера и, как предполагается, уменьшает связывание лигандов TREM223. С целью оценки фагоцитарной функции R47H TREM2 и TREM2 KO проводили анализ высококонтактного фагоцитоза с фиксированными клетками с использованием изогенных линий iPSC-макрофагов с WT/R47H/KO TREM217. Было протестировано несколько длин фагоцитоза продолжительностью от 1 до 5 ч с использованием поэтапного добавления фагоцитарного груза (40 000 SH-SY5Ys). Полученный сигнал линейно увеличивается до 4 ч, слегка выравниваясь через 5 ч(рисунок 5)17. Снижение скорости и емкости фагоцитоза (% пятноположительных клеток) было очевидно при TREM2 KO по сравнению с WT, тогда как мутант R47H TREM2 не показал измененного фагоцитоза. Фагоцитарный дефект в клетках TREM2 KO не является фенокопируемым мутацией R47H TREM2, по-видимому, потому, что функция TREM2 достаточна для поддержки нормального фагоцитоза.

Рисунок 3:Пример данных для анализа фагоцитоза с временными клетками. Поглощение мертвого SH-SY5Ys дикими iPSC-макрофагами BIONi010-C (ECACC ID: 66540023), сфотографируемым с интервалом 5 мин в течение 3 ч. Время, показанное на графике, от начала фагоцитоза, включая первые 40 минут без измерения. По графику построена средняя площадь пятен на ячейку из трех реплицированных скважин. Фагоцитоз 10 000 SH-SY5Ys ингибируется 10 мкМ цитохалазина D за 1 ч до лечения, тогда как более высокие количества SH-SY5Ys (20 000 и 30 000) имеют субоптимальную количественную оценку фагоцитоза. Среднее ± стандартного отклонения (SD), N = 1 эксперимент. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 4:Оптимизация и валидация анализа фагоцитоза с высоким содержанием фиксированных клеток. (A) Репрезентативное микроскопическое изображение с высоким содержанием фагоцитоза SH-SY5Ys, сфагозированного iPSC-макрофагами дикого типа BIONi010-C (ECACC ID: 66540023). Показана 3-часовая точка времени с 40 000 SH-SY5Ys. Флуоресцентные каналы сливаются, при этом окрашивание iPSC-макрофагов показано красным цветом, ядра — синим, а SH-SY5Ys — желтым. Врезная панель представляет собой участок изображения увеличен в 3 раза. (B) Количество пятен на клетку фагоцитированных мертвых SH-SY5Ys через 5 ч, с использованием различных количеств добавок груза к диким типам iPSC-макрофагов. Среднее ± стандартной погрешности среднего значения (SEM), для N = 3 урожая. (C)Фагоцитоз (3 ч) ингибируется 10 мкМ цитохалазина D (Cyt), 1 мкМ бафиломицина A1 (Baf), 1 мкМ джасплакинолида (с 1 ч предварительной обработкой; Jas) и 13 мкг/мл рекомбинантного аннексина V (добавляют одновременно к мертвому SH-SY5Ys; Анна). iPSC-макрофаги без добавления SH-SY5Ys использовались в качестве отрицательного контроля (-ve), а положительный (+ve) контроль представляет собой необработанные iPSC-макрофаги с добавлением SH-SY5Ys. Данные нормализовали до среднего значения для повторения эксперимента. Означает ± SEM, для N = 3-6 урожаев и с двумя клеточными линиями дикого типа (SFC840-03-03, характеристика этой линии описана в (Fernandes et al.21 и BIONi010-C). 1-way ANOVA с пост-специальным тестом Даннетта, сравнения с необработанными клетками. *p < 0,05, ***p < 0,001. (D)Свежефиксированное окрашивание SH-SY5Ys равномерно для дисплея фосфатидилсерина (приложение V-FITC) и ограниченное проницаемость клеток (йодид пропидия). Живые SH-SY5Ys не окрашиваются для аннексина V-FITC или йодида пропидия, за исключением очагового окрашивания, присутствующего на нескольких мертвых клетках в культуре. Цифры воспроизводятся с разрешения Alzheimer's Research & Therapy17. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 5:Фагоцитоз снижается в TREM2 KO, но не в R47H TREM2 iPSC-макрофагах. Анализ фагоцитоза с высоким содержанием проводится с 40 000 SH-SY5Ys на скважину с поэтапными добавками. Средние были количественно определены для параметров: количество пятен на клетку, сумма площадей пятен (мкм2)на клетку и процент клеток, содержащих фагоцитированные частицы на поле. Данные были нормализованы для обозначения каждого генотипа в эксперименте. Среднее ± SEM, для N = 3 урожая. Повторные измерения 2-стороннего ANOVA, пост-хок теста Даннетта, парные сравнения с WT для каждого раза: *p < 0,05, **p < 0,01, ***p < 0,001. Цифры воспроизводятся с разрешения Alzheimer's Research & Therapy17. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
Дополнительный рисунок S1: Пример контроля качества для приготовления SH-SY5Ys. Диссоциированные SH-SY5Ys фиксировали в течение 10 мин с 0% (живые клетки), 1% и 2% параформальдегида (PFA), затем промывали. Клетки окрашивали аннексином V-FITC и йодидом пропидия (PI) и немедленно измеряли проточной цитометрией. Точечные графики цветовой плотности были созданы в программном обеспечении для анализа проточной цитометрии с использованием одноокрашенных и незапятнанных элементов управления для размещения квадрантных ворот. Квадранты аннотируются процентом событий в этом квадранте. Живые клетки находятся в основном в Q4, а фиксированные клетки в основном в Q2. Q1 = приложение V-/PI-, Q2 = приложение V+/PI+, Q3 = приложение V+/PI-, Q4 = приложение V-/PI- (живые ячейки). Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Дополнительное видео: Фагоцитоз живых клеток замедленной съемки. Репрезентативное покадровое видео фагоцитозов SH-SY5Ys дикими iPSC-макрофагами BIONi010-C (ECACC ID: 66540023). Снимки делались каждые 5 мин в течение 3 ч. Видео обрезается и выполняется со скоростью 3 кадра в секунду, показывая последние 1,5 ч анализа. Чувствительные к кислоте красители SH-SY5Y показаны красным цветом, интенсивность сигнала увеличивается с подкислением фагосом. Клеточные ядра, окрашенные Hoechst 33342, показаны синим цветом. Пожалуйста, нажмите здесь, чтобы загрузить это видео.
Обсуждение
Микроглии обладают важными функциями, которые влияют на инициацию и прогрессирование нейродегенеративных заболеваний, включая фагоцитоз апоптотических нейронов. Нарушение фагоцитоза микроглии и неадекватный фагоцитоз синапсов были связаны с нейродегенеративными заболеваниями, хотя основные механизмы и причинно-следственная связь не совсем понятны4,23. В этой статье описывается анализ фагоцитоза для измерения фагоцитоза апоптотических клеток iPSC-макрофагами с помощью либо считывания покадровой визуализации живых клеток, либо с микроскопией с фиксированными клетками с высоким содержанием, либо комбинацией обоих на одном анализе. Эта универсальность означает, что анализ может быть использован для изучения отдельных фагоцитарных событий с течением времени в нескольких скважинах или использован для скрининга с высоким содержанием с несколькими состояниями или методами лечения. Поскольку анализ с высоким содержанием фиксируется в одной точке времени, одновременно могут быть подготовлены несколько пробирных пластин. Анализ с высоким содержанием имеет потенциальную полезность для характеристики макрофагов / микроглии с генетическими вариантами, связанными с заболеванием, или скрининга ингибиторов малых молекул на наличие изменений фагоцитоза. Анализ также может быть легко адаптирован для изучения фагоцитоза других моделей микроглии или потенциально астроцитов. Анализ фагоцитоза потенциально может быть мультиплексирован пятнами визуализации живых клеток, например, индикаторами митохондрий, кальция или АФК, и может быть выполнено постфиксационное иммунофлуоресцентное окрашивание для белков, представляющих интерес. По сравнению с существующими анализами фагоцитоза, которые используют апоптотические нейрональные клетки, основные преимущества, которые дает этот протокол, заключается в том, что подготовка фагоцитарного груза относительно проста и быстра и приводит к однородным продуктам. Другие анализы индуцируют апоптоз нейронов или SH-SY5Ys с S-нитрозо-L-цистеином в течение 2 ч25,окадаевой кислотой в течение 3 ч22,ставроспорином в течение 4-16 ч26,27,28,29 или УФ-облучением в течение 24 ч30,и могут приводить к клеткам на разных стадиях апоптоза. Кроме того, изображения живых клеток и показания изображений с высоким содержанием ранее не были описаны, насколько известно авторам. Основным ограничением использования параформальдегид-фиксации для приготовления фагоцитарного груза является то, что он не полностью рекапитулирует процесс апоптоза, поскольку фиксация препятствует расщеплению клеток на апоптотические тела, которые, вероятно, будут фагоцитозироваться быстрее из-за их меньших размеров. Неизвестно, какое влияние оказывает фиксация на секрецию нуклеотидных сигналов «найди меня» (например, АТФ, УДФ) от клетки-мишени, которая притягивает фагоциты. Подобно апоптотическим клеткам, фиксированные SH-SY5Ys проявляют некоторую мембранную проницаемость для йодида пропидия. Мембранная проницаемость связана с выделением сигналов «найди меня»; однако это не было изучено в фиксированных SH-SY5Ys, и если нуклеотиды высвобождаются слишком быстро, они будут смыты до того, как SH-SY5Ys будут добавлены к iPSC-макрофагам.
Первым критическим шагом в протоколе является окрашивание мертвого SH-SY5Ys эфиром STP чувствительного к pH красного флуоресцентного красителя. Этот краситель быстро и ковалентно реагирует со свободными первичными аминами на поверхности мертвого SH-SY5Ys. Длительность окрашивания не нужно оптимизировать; однако необходимо соблюдать осторожность при обращении с красителем перед маркировкой. Реакция маркировки не должна проводиться в буферах, содержащих свободные амины. Кроме того, существует риск осадков, если запас ДМСО разбавляется в холодном водном буфере или в высокой конечной концентрации. Осадки будут появляться в виде плотных темных объектов под микроскопом. Кроме того, pH-чувствительный раствор красителя прилипает к обычным пластиковым центрифужным трубкам и медленно смывается; поэтому на этапе маркировки рекомендуются трубки с низким связыванием. Использование pH-чувствительного красителя вместо постоянно флуоресцентного красителя помогает идентифицировать поглощенные частицы по сравнению с частицами, которые соседствуют с плазматической мембраной. Поскольку существует некоторая флуоресценция при нейтральном рН, плотность фагоцитарного груза и iPSC-макрофагов должна быть достаточно низкой для точной сегментации, хотя и достаточно высокой, чтобы захватывались многочисленные фагоцитарные события. Высокосодержимая микроскопия была способна точно идентифицировать фагоцитоз со средней плотностью груза в скважине (более 2 SH-SY5Ys на iPSC-макрофаг). И наоборот, из-за более слабой чувствительности микроскопа в темно-красном спектре сегментация iPSC-макрофагов в данных покадровой визуализации живых клеток была менее уверенной и необходимо было использовать очень низкую плотность груза для снижения вероятности ложных срабатываний (1 SH-SY5Y на каждые два iPSC-макрофага). Валидация надлежащей сегментации и плотности груза должна проводиться со сравнением необработанных скважин и скважин, обработанных цитохалазином D. В хорошо оптимизированном анализе цитохалазин D должен уменьшить среднее количество пятен на клетку на 90% по сравнению с необработанными образцами.
Другим важным шагом в протоколе является окрашивание iPSC-макрофагов, которое позволяет идентифицировать и сегментировать клетку в анализе изображений, так что любые внешние SH-SY5Y исключаются из подсчета. Рекомендуемый краситель является клеточно-проницаемым, преобразованным в нерастворимый флуоресцентный продукт внутри цитоплазмы, поправимым и нетоксичным (см. Таблицу материалов). Этап окрашивания был оптимизирован для использования iPSC-макрофагов с анализом фагоцитоза с высоким содержанием изображения, и мы предлагаем его повторно оптимизировать, если используются другие типы клеток. Продолжительность окрашивания клеток может быть увеличена для улучшения осаждения нерастворимого флуоресцентного продукта в клетках. Если концентрация красителя оптимизирована, следует позаботиться о том, чтобы избежать токсичных уровней органического растворителя.
Третьим критическим фактором успеха анализа является анализ данных. Предоставляемые конвейеры анализа предназначены для руководства, а не для предписания, поскольку различия в интенсивности окрашивания или морфологии клеток могут снизить эффективность сегментации конвейеров, как написано. Поэтому потребуются некоторые оптимизации с тестированием конвейера на соответствующих положительных и отрицательных элементах управления, а параметры, которые должны быть оптимизированы, указаны в тексте протокола. Отрицательный контроль должен включать состояние, при котором iPSC-макрофаги предварительно обрабатываются мощным ингибитором фагоцитоза, таким как цитохалазин D, перед добавлением SH-SY5Ys. Другим возможным негативным контролем является добавление SH-SY5Ys к ранее необработанным скважинам iPSC-макрофагов в конце анализа, за 10 минут до фиксации, что позволяет некоторое осаждение груза, но слишком коротко для того, чтобы произошло значительное количество фагоцитоза. Событие фагоцитоза определяется как красно-флуоресцентный объект в границах iPSC-макрофага, определяемый программным алгоритмом с использованием темно-красного флуоресцентного канала. Если сегментация клеток плохая(рисунок 2),многие нефагоцитозные SH-SY5Ys в непосредственной близости от iPSC-макрофагов могут быть ошибочно включены в анализ, т.е. ложноположительные результаты. Важнейшим фактором достижения хорошей сегментации является строгое разграничение iPSC-макрофагов. Сегментация для обоих анализов автоматизирована, поэтому невозможно получить идеальную сегментацию для каждой ячейки; однако некоторые параметры могут быть скорректированы, чтобы сделать сегментацию более оптимальной, используя несколько тестовых изображений в качестве эталона. Контроль цитохалазина D важен для оценки оптимальной сегментации, поскольку большое количество фагоцитарных событий, обнаруженных в этом состоянии, указывает на то, что сегментация является неоптимальной. Оптимизация конвейера анализа данных в идеале должна повторяться до тех пор, пока количество фагоцитарных событий на клетку не будет на 80-90% ниже в состоянии цитохалазина D по сравнению с отсутствием ингибитора.
Наиболее вероятными проблемами с анализом фагоцитоза являются: (1) слабая рН-чувствительная флуоресценция в положительном контроле, (2) редкое или неравномерное распределение макрофагов в конце анализа или (3) большое количество ложноположительных результатов в анализе из нефагоцитозных SH-SY5Ys. При устранении неполадок слабой pH-чувствительной флуоресценции следует сначала проверить, что окрашивание SH-SY5Ys привело к получению ячейки с сильным пурпурным цветом. Если цвет слабый, убедитесь, что используется свежий запас красителя, убедитесь, что буфер маркировки не содержит аминов, добавьте дополнительную промывку в SH-SY5Ys перед окрашиванием, проверьте, было ли окрашено правильное количество SH-SY5Y, убедитесь, что нет осадков красителя, и оптимизируйте маркировку концентрации красителя. Если SH-SY5Y сильно окрашены, проверьте, правильна ли концентрация, добавленная к анализной пластине, и убедитесь, что iPSC-макрофаги здоровы и не слишком стары. Второй тип проблемы, неравномерное распределение макрофагов, может возникнуть в результате потери клеток во время пипетки, и следует предпринять шаги для уменьшения сил пипетки, испытываемых клетками, избегая узкобородых кончиков. Если проблема остается, сократите время инкубации загрузки iPSC-макрофагов клеточно-проницаемым красителем. Третья проблема, касающаяся ошибочного включения в анализ нефагоцитированных частиц, указывает на то, что требуется большая оптимизация конвейера анализа. Устранение неполадок должно быть сосредоточено в первую очередь на сегментации ячеек и на том, включает ли программное обеспечение соседние объекты. Конкретные параметры, которые могут быть скорректированы, предлагаются в примечаниях ниже соответствующих этапов (этапы 6.1.11-6.1.15 для покадрового анализа живых клеток и этапы 6.2.4-6.2.8 для анализа высокого содержания). Если сегментация клеток не может быть дополнительно улучшена, анализ высокого содержания имеет дополнительный шаг (шаг 6.2.8), который исключает неправильно сегментированные iPSC-макрофаги. Кроме того, модуль, который фильтрует принятые пятна pH-чувствительной флуоресценции в iPSC-макрофагах, может быть оптимизирован, увеличивая пороговую интенсивность принимаемых объектов, что должно помочь исключить нефагоцитозные SH-SY5Ys (шаг 6.1.17 для покадрового анализа живых клеток и шаг 6.2.11 для анализа высокого содержания).
Мы разработали два типа считывания микроскопии для анализа фагоцитоза, каждый из которых имеет преимущества и ограничения. Покадровая визуализация живых клеток имеет преимущества предоставления дополнительной информации о кинетике фагоцитоза и более широко доступна, чем платформы визуализации с высоким содержанием. Рекомендуемое программное обеспечение с открытым исходным кодом не соответствует источнику микроскопа и может использоваться с любым высококачественным флуоресцентным микроскопом, с возможностью замедленной съемки живых клеток или без нее. Основным ограничением визуализации живых клеток является ограниченная чувствительность и оптика, что затрудняет обнаружение и выполнение хорошей сегментации iPSC-макрофагов. Это ограничение может быть смягчено либо путем увеличения продолжительности окрашивания iPSC-макрофагов, либо путем переключения на более чувствительный микроскоп, если таковой имеется. Анализ фагоцитоза с высоким содержанием является рекомендуемым считыванием, если доступна система визуализации с высоким содержанием. Системы визуализации с высоким содержанием обеспечивают более высокую пропускную способность и более надежные данные, что позволяет использовать этот анализ для скрининга, в котором ожидается надежный Z' ≥0,7 для выхода «количество пятен на ячейку»20. По сравнению с методом замедленной съемки живых клеток, считывание микроскопии с высоким содержанием имеет более высокую чувствительность, более высокую степень автоматизации и скорости, может быть обработано больше скважин и полей визуализации, и производятся конфокальные изображения с высоким разрешением. Сегментация клеток более эффективна при хороших изображениях, а сегментации дополнительно помогает программное обеспечение для анализа изображений с высоким содержанием, обеспечивающее больше методов сегментации клеток, подходящих для клеток с высокой неправильной формой. Программное обеспечение для анализа изображений с высоким содержанием также рассчитало больше параметров фагоцитоза по сравнению с программным обеспечением с открытым исходным кодом, таким как процент фагоцитарных клеток. Основным ограничением анализа фагоцитоза с высоким содержанием является стоимость и доступность системы визуализации и программного обеспечения для анализа.
В заключение, количественный анализ фагоцитоза, представленный в данной работе, является полезным инструментом для моделирования фагоцитоза микроглии мертвых нейронов in vitro. Микроглия моделируется iPSC-макрофагами, а мертвые нейроны моделируются параформальдегид-фиксированным SH-SY5Ys. Хотя это не самые аутентичные модели микроглии и мертвых / апоптотических нейронов, они просты в подготовке и масштабируемы. Сам анализ очень универсален, с двумя типами считывания изображений, и он может быть адаптирован для использования с различными моделями монокультуры микроглии / макрофагов или другим типом клеток, чтобы действовать в качестве фагоцитарного груза. Считывание изображений с высоким содержанием выгодно для получения количественных данных и может быть масштабировано до анализа модуляторов малых молекул фагоцитоза или скрининга генетических вариантов в iPSC-макрофагах. Однако, поскольку системы визуализации с высоким содержанием являются дорогостоящими и большими, альтернативное считывание изображений было включено в протокол с использованием покадрового микроскопа с живыми клетками, который при необходимости может быть заменен любым обычным флуоресцентным микроскопом хорошего качества.
Раскрытие информации
Авторам нечего раскрывать.
Благодарности
Авторы благодарят доктора Вэла Миллара и доктора Сохаиба Низами за их помощь в микроскопии с высоким содержанием, а также доктора Даниэля Эбнера за доступ к микроскопам с высоким содержанием. Кроме того, авторы благодарят доктора Эмму Мид за советы по разработке анализа и миссис Кэти Браун за поддержку iPSC. Эта работа была поддержана Оксфордским институтом поиска лекарств (ARUK ODDI, ссылка на грант ARUK-2020DDI-OX) при дополнительной поддержке Центра стволовых клеток Джеймса Мартина в Оксфорде (S.A.C.) из Оксфордской школы Мартина LC0910-004; Премия Monument Trust Discovery Award от Паркинсона Великобритании (J-1403); MRC Dementias Platform UK Stem Cell Network Capital Equipment MC_EX_MR/N50192X/1, Partnership MR/N013255/1 и Momentum MC_PC_16034 Awards.
Материалы
| Name | Company | Catalog Number | Comments |
| 15 mL conical centrifuge tube | Falcon | 352096 | For centrifugation of cells |
| 2-20 µL, 20-200 µL, 100-1000 µL single-channel micropipettes | |||
| 2-mercaptoethanol 50 mM | Gibco | 31350010 | Component of Factory media |
| 4% paraformaldehyde in PBS | Alfa Aesar | J61899 | For fixation of cells |
| 6-well plate, tissue culture treated | |||
| AggreWell-800 24-well plate | STEMCELL Technologies | 34815 | Microwell low-adherence 24-well plate for formation of embryoid bodies |
| Annexin V-FITC Apoptosis Staining / Detection Kit | Abcam | ab14085 | Kit for annexin V-FITC staining , as an assay for quality control of fixed SH-SY5Ys. Kit contains annexin binding buffer, annexin V-FITC, and propidium iodide. |
| Automated cell counter | |||
| Benchtop centrifuge | |||
| Benchtop microcentrifuge | |||
| CellCarrier-96 Ultra Microplates, tissue culture treated, black, 96-well with lid | Perkin Elmer | 6055302 | 96-well tissue culture (TC)-treated microplate with black well walls and an optically-clear bottom, for phagocytosis assay |
| CellProfiler software | Open-source software for analysis of phagocytosis images obtained by live-cell time-lapse microscope. Download for free from website (http://cellprofiler.org/), this protocol used version 2.2.0. | ||
| CellTracker Deep Red dye | Thermo Fisher | C34565 | Deep red-fluorescent, cell-permeant, succinimidyl ester-reactive dye for staining cytoplasm of iPS-macrophages. Dissolve CellTracker Deep Red dye in DMSO to 2 mM (1.4 mg/mL). Use at 1 μM, by dilution of DMSO stock with Macrophage media. |
| Class 2 laminar air flow safety cabinet | |||
| CO2 gas bottle | Accessory for EVOS FL Auto | ||
| CO2 incubator, set to 37°C and 5 % CO2 | |||
| Columbus Image Data Storage and Analysis System | Perkin Elmer | Columbus | Data storage and analysis platform for Opera Phenix. Supports all major high content screening instruments. |
| Cytochalasin D | Cayman | 11330 | Negative control treatment for phagocytosis assay. Reconstitute in DMSO to 10 mM and store aliquots at -20°C, avoid further freeze-thaw cycles. Use at final concentration 10 µM. |
| DMEM/F12 | Gibco | 11320074 | Component of SH-SY5Y media |
| DMSO | Sigma | D8418 | Solvent for CellTracker and pHrodo dyes |
| EVOS FL Auto Imaging System | Thermo Fisher | AMF4300 | Live-cell time-lapse imaging microscope |
| EVOS Light Cube CY5 | Thermo Fisher | AMEP4656 | Accessory for EVOS FL Auto |
| EVOS Light Cube DAPI | Thermo Fisher | AMEP4650 | Accessory for EVOS FL Auto |
| EVOS Light Cube RFP | Thermo Fisher | AMEP4652 | Accessory for EVOS FL Auto |
| EVOS Onstage Incubator | Thermo Fisher | AMC1000 | Accessory for EVOS FL Auto |
| Fetal Bovine Serum | Sigma | F4135 | Component of SH-SY5Y media |
| Flow cytometer | |||
| Flow cytometry analysis software | |||
| Geltrex LDEV-Free, hESC-Qualified, Reduced Growth Factor Basement Membrane Matrix | Invitrogen | A1413302 | hESC-qualified basement membrane matrix for iPSC culture |
| GlutaMAX Supplement | Gibco | 35050-038 | Component of both Factory and Macrophage media |
| HBSS | Lonza | BE 10-547F | Hank’s balanced salt solution for washing steps |
| Human recombinant BMP4 | Gibco | PHC9534 | Component of Embryoid Body media |
| Human recombinant IL-3 | Gibco | PHC0033 | Component of both Factory and Macrophage media |
| Human recombinant SCF | Miltenyi Biotech | 130-096-695 | Component of Embryoid Body media |
| Human recombinant VEGF | Gibco | PHC9394 | Component of Embryoid Body media |
| Live Cell Imaging Solution | Thermo Fisher | A14291DJ | Phenol red-free HEPES-buffered media for labelling dead SH-SY5Ys |
| Low protein binding 2 mL tubes | Eppendorf | 30108.132 | For staining SH-SY5Ys |
| M-CSF | Thermo Fisher | PHC9501 | Component of both Factory and Macrophage media |
| mTeSR1 Medium | STEMCELL Technologies | 85850 | iPSC media |
| Multichannel 20-200 uL pipette | For liquid handling of 96-well plate | ||
| NucBlue Live ReadyProbes Reagent | Thermo Fisher | R37605 | Hoechst 33342 formulation in a dropper bottle for staining nuclei of iPS-macrophages, use 0.5 drops/mL in Macrophage media. |
| Opera Phenix High-Content Screening System | Perkin Elmer | HH14000000 | High-content imaging microscope, used with Harmony software version 4.9. |
| Penicillin-Streptomycin | Gibco | 15140-122 | Component of Factory, Macrophage, and SH-SY5Y media |
| pHrodo iFL Red STP-Ester | Thermo Fisher | P36011 | pH-sensitive red fluorescent dye for labelling dead SH-SY5Ys. Reconstitute pHrodo iFL Red STP Ester powder in DMSO to a 5 mg/mL concentration. For each 1 million SH-SY5Ys, add 2.5 μL (12.5 μg) of pHrodo iFL Red STP Ester stock to pre-warmed cells suspended in Live Cell Imaging Solution. |
| Serological pipette filler | |||
| T175 flask, tissue culture treated | Vessel for differentiations of iPSC-macrophage precursors, known as "Factories" | ||
| T75 flask | Vessel for SH-SY5Y culture | ||
| Transparent plate sealers | Greiner Bio-One | 676001 | For assay plate storage and transportation |
| TrypLE Express (1X), no phenol red | Gibco | 12604013 | Cell dissociation buffer containing recombinant trypsin-like enzymes and 1.1 mM EDTA, use neat. |
| Water bath, set to 37°C | |||
| X-VIVO 15 Medium with L-glutamine, gentamicin, and phenol red | Lonza | BE04-418F | Component of Factory and Macrophage media |
| X-VIVO 15 Medium with L-glutamine; without gentamicin or phenol red | Lonza | 04-744Q | Phenol red-free macrophage media, for use in phagocytosis without additives or growth factors |
Ссылки
- Hochreiter-Hufford, A., Ravichandran, K. S. Clearing the dead: apoptotic cell sensing, recognition, engulfment, and digestion. Cold Spring Harbour Perspectives in Biology. 5, 008748 (2013).
- Freeman, S. A., Grinstein, S. Phagocytosis: receptors, signal integration, and the cytoskeleton. Immunological Reviews. 2262, 193-215 (2014).
- Hickman, S., Izzy, S., Sen, P., Morsett, L., El Khoury, J. Microglia in neurodegeneration. Nature Neuroscience. 21 (10), 1359-1369 (2018).
- Galloway, D. A., Phillips, A. E. M., Owen, D. R. J., Moore, C. S. Phagocytosis in the brain: Homeostasis and disease. Frontiers in Immunology. 10, 790 (2019).
- Nizami, S., Hall-Roberts, H., Warrier, S., Cowley, S. A., Di Daniel, E. Microglial inflammation and phagocytosis in Alzheimer's disease: potential therapeutic targets. British Journal of Pharmacology. 176 (18), 3515-3532 (2019).
- Hong, S., et al. Complement and microglia mediate early synapse loss in Alzheimer mouse models. Science. 352 (6286), 712-716 (2016).
- Neher, J. J., et al. Inhibition of microglial phagocytosis is sufficient to prevent inflammatory neuronal death. The Journal of Immunology. 186 (8), 4973-4983 (2011).
- Brown, G. C., Neher, J. J. Microglial phagocytosis of live neurons. Nature Reviews Neuroscience. 15 (4), 209-216 (2014).
- Scott-Hewitt, N., et al. Local externalization of phosphatidylserine mediates developmental synaptic pruning by microglia. The EMBO Journal. 39 (16), 105380 (2020).
- Li, T., et al. A splicing isoform of GPR56 mediates microglial synaptic refinement via phosphatidylserine binding. The EMBO Journal. 39 (16), 104136 (2020).
- Sapar, M. L., et al. Phosphatidylserine externalization results from and causes neurite degeneration in Drosophila. Cell Reports. 24 (9), 2273-2286 (2018).
- Skjesol, A., et al. The TLR4 adaptor TRAM controls the phagocytosis of Gram-negative bacteria by interacting with the Rab11-family interacting protein 2. PLOS Pathogens. 15 (3), 1007684 (2019).
- Wong, K., Li, X., Ma, Y. Paraformaldehyde induces elevation of intracellular calcium and phosphatidylserine externalization in platelets. Thrombosis Research. 117 (5), 537-542 (2006).
- van Wilgenburg, B., Browne, C., Vowles, J., Cowley, S. A. Efficient, long term production of monocyte-derived macrophages from human pluripotent stem cells under partly-defined and fully-defined conditions. PLoS One. 8 (8), (2013).
- Haenseler, W., et al. A highly efficient human pluripotent stem cell microglia model displays a neuronal-co-culture-specific expression profile and inflammatory response. Stem Cell Reports. 8 (6), 1727-1742 (2017).
- Buchrieser, J., James, W., Moore, M. D. Human induced pluripotent stem cell-derived macrophages share ontogeny with MYB-independent tissue-resident macrophages. Stem Cell Reports. 8 (2), 334-345 (2017).
- Hall-Roberts, H., et al. TREM2 Alzheimer's variant R47H causes similar transcriptional dysregulation to knockout, yet only subtle functional phenotypes in human iPSC-derived macrophages. Alzheimer's Research & Therapy. 12, 151 (2020).
- Friedman, B. A., et al. Diverse brain myeloid expression profiles reveal distinct microglial activation states and aspects of Alzheimer's disease not evident in mouse models. Cell Reports. 22 (3), 832-847 (2018).
- Aziz, M., Yang, W. L., Wang, P. Measurement of phagocytic engulfment of apoptotic cells by macrophages using pHrodo succinimidyl ester. Current Protocols in Immunology. 100, 1-8 (2013).
- Atmaramani, R., Pancrazio, J. J., Black, B. J. Adaptation of robust Z' factor for assay quality assessment in microelectrode array based screening using adult dorsal root ganglion neurons. Journal of Neuroscience Methods. 339, 108699 (2020).
- Fernandes, H. J. R., et al. ER Stress and Autophagic Perturbations Lead to Elevated Extracellular α-Synuclein in GBA-N370S Parkinson's iPSC-Derived Dopamine Neurons. Stem Cell Reports. 6 (3), 342-356 (2016).
- Takahashi, K., Rochford, C. D. P., Neumann, H. Clearance of apoptotic neurons without inflammation by microglial triggering receptor expressed on myeloid cells-2. Journal of Experimental Medicine. 201 (4), 647-657 (2005).
- Kober, D. L., Brett, T. J. TREM2-ligand interactions in health and disease. Journal of Molecular Biology. 429 (11), 1607-1629 (2017).
- Hong, S., et al. Complement and microglia mediate early synapse loss in Alzheimer mouse models. Science. 352 (6286), 712-716 (2016).
- Witting, A., Müller, P., Herrmann, A., Kettenmann, H., Nolte, C. Phagocytic clearance of apoptotic neurons by microglia/brain macrophages in vitro: Involvement of lectin-, integrin-, and phosphatidylserine-mediated recognition. Journal of Neurochemistry. 75 (3), 1060-1070 (2000).
- Hsieh, C. L., et al. A role for TREM2 ligands in the phagocytosis of apoptotic neuronal cells by microglia. Journal of Neurochemistry. 109 (4), 1144-1156 (2009).
- Beccari, S., Diaz-Aparicio, I., Sierra, A. Quantifying microglial phagocytosis of apoptotic cells in the brain in health and disease. Current Protocols in Immunology. 122 (1), 49 (2018).
- Diaz-Aparicio, I., et al. Microglia actively remodel adult hippocampal neurogenesis through the phagocytosis secretome. Journal of Neuroscience. 40 (7), 1453-1482 (2020).
- Shirotani, K., et al. Aminophospholipids are signal-transducing TREM2 ligands on apoptotic cells. Scientific Reports. 9 (1), 1-9 (2019).
- Garcia-Reitboeck, P., et al. Human induced pluripotent stem cell-derived microglia-like cells harboring TREM2 missense mutations show specific deficits in phagocytosis. Cell Reports. 24 (9), 2300-2311 (2018).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеThis article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены