
Method Article
Поведенческий экран для тепловых припадков в мышиных моделях эпилепсии
В этой статье
Резюме
Целью метода является скрининг на гипертермию или вызванные теплом судороги в мышиных моделях. Протокол описывает использование специально построенной камеры с непрерывным мониторингом температуры тела, чтобы определить, приводит ли повышенная температура тела к судорогам.
Аннотация
Трансгенные мышиные модели оказались мощными инструментами в изучении различных аспектов неврологических расстройств человека, включая эпилепсию. SCN1A-ассоциированные генетические эпилепсии включают широкий спектр судорожных расстройств с неполной пенетрантностью и клинической вариабельностью. Мутации SCN1A могут привести к большому разнообразию фенотипа судорог, начиная от простых, самоограниченных фебрильных судорог, связанных с лихорадкой (FS), генетической эпилепсии умеренного уровня с фебрильными судорогами плюс (GEFS +) до более тяжелого синдрома Драве (DS). Хотя ФС обычно наблюдаются у детей в возрасте до 6-7 лет, у которых нет генетической эпилепсии, ФС у пациентов с ГЭФ+ продолжают возникать во взрослой жизни. Традиционно экспериментальные ФС индуцировались у мышей путем воздействия на животное потока сухого воздуха или нагрева ламп, и скорость изменения температуры тела часто плохо контролируется. Здесь мы описываем изготовленную на заказ нагревательную камеру с передней частью из плексигласа, которая оснащена цифровым регулятором температуры и электрическим вентилятором, оснащенным нагревателем, который может отправлять нагретый принудительный воздух на испытательную арену с контролируемой температурой. Температура тела мыши, помещенной в камеру, контролируемая через ректальный зонд, может быть увеличена до 40-42 °C воспроизводимым образом путем повышения температуры внутри камеры. Постоянный визуальный мониторинг животных в течение отопительного периода демонстрирует индукцию вызванных теплом судорог у мышей, несущих мутацию FS при температуре тела, которая не вызывает поведенческих судорог у пометников дикого типа. Животные могут быть легко удалены из камеры и помещены на охлаждающую площадку, чтобы быстро вернуть температуру тела к норме. Этот метод обеспечивает простой, быстрый и воспроизводимый протокол скрининга для возникновения тепловых припадков в моделях мышей эпилепсии.
Введение
Эпилепсия, четвертое наиболее распространенное семейство неврологических расстройств в Соединенных Штатах1, характеризуется дисбалансом возбуждающего и тормозного влечения в ЦНС, что приводит к рецидивирующим судорогам. Фебрильные судороги (ФС) или судороги, связанные с лихорадкой, могут возникать в общей популяции, чаще всего у детей уже в возрасте от 3 месяцев до 6-7 лет. Тем не менее, у некоторых людей с генетическими мутациями, чаще всего в гене натриевого канала, FS может сохраняться после возраста 7 лет во взрослой жизни. Это состояние называется фебрильными судорогами плюс или FS +. Быстрые достижения в секвенировании генома выявили более 1 300 мутаций в гене ионного канала натрия человека SCN1A, что делает его горячей точкой для мутаций эпилепсии. Мутации SCN1A были связаны с широким спектром судорожных расстройств, включая фебрильные судороги (FS), генетическую эпилепсию с фебрильными судорогами плюс (GEFS+) и синдром Драве (DS)2,3,4,5,6. Около 20% мутаций SCN1A приводит к GEFS+5,7,8. Детский анамнез сложных или длительных ФС в детском возрасте может впоследствии развиться в более изнурительные формы эпилепсии, такие как височная эпилепсия (TLE)9,10,11. Синдром Драве возникает из-за мутаций усечения или потери функциональных мутаций в SCN1A и является тяжелой формой трудноизлечимой эпилепсии, с детским началом фебрильных судорог, которые перерастают в рефрактерные припадки, и часто связан с когнитивными, развивающими и двигательными нарушениями2,5,12 . Поскольку многие люди с GEFS+ и / или DS демонстрируют фебрильные судороги, становится необходимым разработать новые методы лечения для лучшей борьбы с этими судорожными расстройствами.
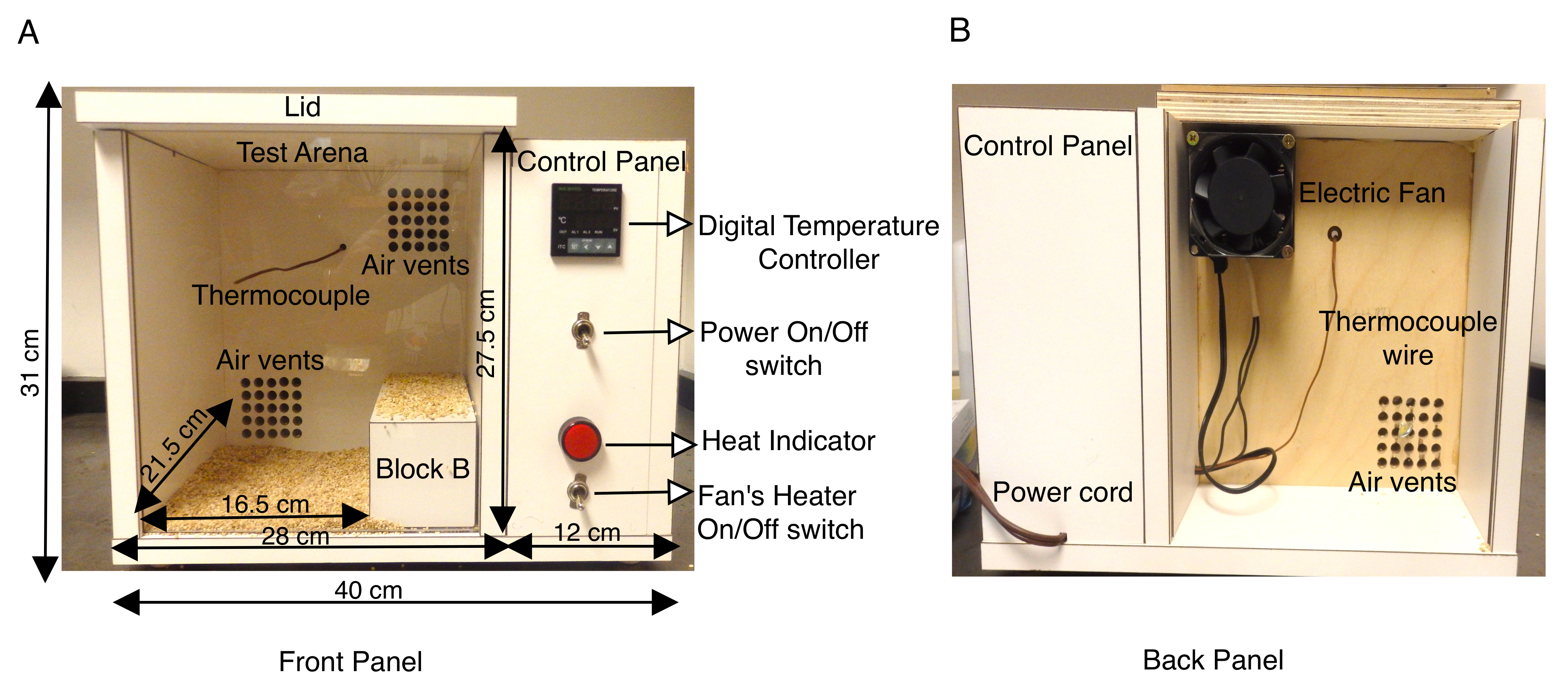
Животные модели эпилепсии, связанной с SCN1A, оказались неоценимыми в характеристике различных типов судорог (фебрильные и генерализованные) и препарировании нейронного механизма генерации судорог13,14,15,16,17,18. В то время как изучение спонтанных судорог с помощью записей ЭЭГ / ЭМГ в мозге грызунов хорошо известно и является очень полезным инструментом, только несколько исследований пытались имитировать фебрильные судороги на моделях мышей14,16,19,20,21,22,23 . В предыдущих исследованиях использовалась струя нагретого сухого воздуха, или метакрилатный цилиндр, оснащенный тепловой системой, или тепловые лампы с регулятором температуры в закрытых испытательных аренах9,16,21,22,23,24 для индуцирования судорог через гипертермию. Чтобы повысить температуру тела в более контролируемой среде, протокол, описанный здесь, использует специально построенную камеру с системой нагрева с контролируемой температурой, которая позволяла воспроизводить темпы повышения температуры тела мыши внутри камеры. Тепловая камера была построена из дерева (длина 40 см x ширина 34 см x высота 31 см) и была оснащена цифровым регулятором температуры с термопарой K. Небольшой осевой вентилятор, оснащенный нагревателем на задней панели камеры, направляет нагретый воздух в камеру, регулируемую цифровым регулятором температуры. Эта система принудительного нагрева воздуха позволяет контролировать скорость, с которой температура камеры увеличивается. (Рисунок 1А,В). Термопара K, расположенная внутри деревянной тепловой камеры, отправляет обратную связь на цифровой регулятор температуры, чтобы поддерживать постоянную температуру внутри коробки во время анализа. Установка температуры на цифровом регуляторе температуры позволяет электрическому вентилятору отправлять нагретый принудительный воздух через вентиляционные отверстия для равномерного нагрева камеры (рисунок 1А). Передняя панель тепловой камеры представляет собой прозрачный лист из плексигласа, позволяющий легко записывать испытания на видео.
Для каждого эксперимента были отобраны взрослые (P30-P40) мыши, гетерозиготные для мутации в SCN1A, которая заставляет GEFS+ и равное количество пометников дикого типа служить контрольной группой. Животные, как самцы, так и самки, используемые в этих исследованиях, весили не менее 15 г, поскольку мыши дикого типа с меньшим весом были более чувствительны к тепловым судорогам, чем более тяжелые животные того же возраста. В пилотном исследовании было замечено, что мыши как мутантного, так и дикого типа искали более холодные углы камеры сзади и оставались там в течение длительных периодов времени. Чтобы обойти это, эффективный размер пола внутри испытательной арены тепловой камеры был уменьшен до длины 16,5 см x ширины 21,5 см x высоты 27,5 см путем размещения деревянного блока B (размеры 20 см x 8 см x 7,2 см) в правой части камеры (рисунок 1A). Тепловая камера была построена из фанеры толщиной 1,9 см (длина 40 см x ширина 34 см x высота 31 см), покрытой белым ламинатом и оснащенной цифровым регулятором температуры с термопарой K. Ламинированная поверхность стенок камеры непроницаема и может быть легко продезинфицирована между испытаниями путем протирания 70% этанолом. Температуру тепловой камеры первоначально устанавливали на уровне 50 °C и предварительно нагревали в течение не менее 1 ч до начала эксперимента, чтобы обеспечить равномерный нагрев внутри камеры. Каждая мышь была оснащена ректальным термометром для непрерывного мониторинга температуры тела на протяжении всего эксперимента. Одну мышь помещали в камеру за один раз, и температура поддерживалась на уровне 50 ° C между 1-10-й минутой. Затем температуру поднимали до 55 °C на 11-20-й минуте и, наконец, поднимали до 60 °C на 21-30-й минуте. Это привело к воспроизводимой скорости повышения температуры тела мыши (рисунок 2А). Каждое испытание было записано на видео, а поведенческий анализ проводился в автономном режиме.
Протокол нагрева может быть легко изменен для изменения начальной температуры тепловой камеры и скорости нагрева камеры, что, в свою очередь, изменяет скорость повышения температуры тела мыши во время анализа. Таким образом, этот метод обеспечивает большую гибкость по сравнению с традиционными методами в настройке поведенческих экранов, включающих тепловые судороги. Протокол теплового припадка также может быть использован для скрининга противоэпилептических препаратов, которые делают мышей-мутантов более устойчивыми к тепловым судорогам или повышают пороговую температуру, при которой наблюдаются судороги. Аналогичным образом, благотворное влияние ограничительных режимов диеты, таких как кето-диета, на судороги, вызванные жарой, могут быть изучены у нормальных мышей, которых кормили чау- и кето-кормили.

Рисунок 1: Описание изготовленной на заказ тепловой камеры мыши. (A) На передней панели деревянной тепловой камеры мыши показана боковая панель управления, содержащая переключатель Power ON/OFF, который включает цифровой регулятор температуры, термопару K, переключатель ON/OFF тепловентилятора и индикатор тепла. Внешние размеры коробки и внутренней испытательной арены указаны в см. Также показан деревянный блок B, используемый для эффективного уменьшения поверхности испытательной арены. Нижняя часть тестовой арены покрыта початковой подстилкой, чтобы мыши не вступали в непосредственный контакт с нагретыми деревянными поверхностями. (B) На задней панели тепловой камеры показан вентилятор, установленный на верхнем вентиляционном отверстии, и шнур питания для подачи электроэнергии в камеру. Эта цифра изменена по сравнению с рисунком 3 в Das et al., 2021, eNeuro14. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
протокол
Все процедуры для животных были выполнены в соответствии с руководящими принципами Институционального комитета по уходу за животными и их использованию (IACUC) в Калифорнийском университете в Ирвине.
1. Подготовка к анализу теплового захвата
- Включите кнопку Power On в тепловой камере, а затем кнопку Heat On .
- Установите температуру тепловой камеры на уровне 50 °C с помощью клавиатуры цифрового регулятора температуры.
- Подождите не менее 1 часа, чтобы предварительно разогреть камеру при 50 °C, прежде чем вводить первую мышь в камеру. Предварительный нагрев обеспечивает равномерный нагрев внутри камеры.
- Выровняйте пол тепловой камеры мыши подложкой из початков.
- Установите видеорегистратор перед тепловой камерой для записи каждого испытания по анализу захвата, вызванного теплом.
- Выровняйте чашку Петри диаметром 140 мм толстыми слоями папиросной бумаги и поместите ее на лед, чтобы она служила охлаждающей подушкой.
ПРИМЕЧАНИЕ: В конце анализа отдельные мыши будут перенесены на предварительно охлажденную охлаждающую подушку, чтобы помочь снизить их повышенную температуру тела.
2. Подготовка мыши к анализу судорог, вызванному теплом
- Выберите 10 взрослых мышей (P30-P40), 5, которые несут эпилепсию, вызывающую мутацию, и 5 партнеров по помету дикого типа для скринингового анализа судорог, вызванных теплом.
ПРИМЕЧАНИЕ: Мыши дикого типа, не питающиеся эпилепсией, вызывающей мутацию, не проявляют вызванных теплом судорог при температуре ниже 44 ° C и служат контрольной группой. - Взвесьте каждую мышь, которая будет использоваться для скринингового анализа, и запишите ее массу тела. Для анализа следует использовать только мышей весом 15 г и более.
- Экран по одной мыши за раз в тепловой камере мыши.
- Ненадолго обезболивайте мышь в течение 10-15 с, используя несколько капель изофлурана на дне банки-колокольчика.
- Выньте животное из банки-колокольчика и положите его на бумажное полотенце.
- Убедитесь, что мышь полностью обезболена, проверив, что мышь не реагирует на вредное защемление пальца ноги.
- Покройте металлический наконечник ректального температурного зонда смазкой (например, вазелином) и аккуратно вставьте его в мышь на глубине менее или равной 2 см.
- Закрепите ректальный зонд на хвосте мыши скотчем, чтобы зонд не выходил во время анализа.
ПРИМЕЧАНИЕ: В качестве альтернативы поместите животное в конус удерживающего устройства мыши и вставьте ректальный температурный зонд. Закрепите его, приклеив к хвосту. - Убедитесь, что ректальный зонд подключен к мультиметру, который отображает внутреннюю температуру тела мыши.
- Поместите животное в свежую клетку, выстланную початковой подстилкой, т.е. в клетку восстановления.
- Запустите таймер и подождите 5 минут. Наблюдайте за мышью до тех пор, пока она полностью не оправится от анестезии и мышь не будет активной и груминговой.
- Одновременно контролируйте температуру тела мыши до тех пор, пока она не стабилизируется при 35-36 °C.
- По истечении 5 мин обратите внимание на температуру тела мыши. Это начальная температура тела в момент времени «0» мин.
ПРИМЕЧАНИЕ: Если температура тела мыши ниже 35 °C, подождите дополнительное время, пока животное оправится от гипотермии, вызванной анестезией. - Быстро перенесите отдельную мышь в предварительно нагретую камеру мыши. Это знаменует собой НАЧАЛО экспериментального испытания. В данный момент времени проверяется только одна мышь.
3. Анализ судорог, вызванных теплом
- Аккуратно положив мышь на пол предварительно нагретой мышиной тепловой камеры, закройте дверцу из плексигласа и запустите камеру для видеозаписи эксперимента.
- Запустите секундомер. Записывайте температуру тела мыши с ректального термометра с интервалом в 1 мин на время эксперимента.
- Через равные промежутки времени повышают температуру тепловой камеры мыши таким образом, чтобы температура тела мыши повышалась со скоростью 0,25-0,5 °C/мин.
ПРИМЕЧАНИЕ: Быстрое повышение температуры тела может привести к тепловому удару или смерти, и его следует избегать. - Следуя этому протоколу, увеличивайте температуру тепловой камеры мыши на 5 °C каждые 10 минут, как показано на рисунке 2A.
- Через 9,5 мин установите температуру тепловой камеры на 55 °C, чтобы стабилизировать температуру тепловой камеры до 55 °C к 10-й минуте , как показано на цифровом температурном дисплее.
- Аналогично, увеличьте температуру до 60 °C через 19,5 мин, чтобы стабилизировать температуру тепловой камеры до 60 °C к 20-й минуте . Каждое испытание скрининга судорог длится в течение 30 минут.
- Если у мыши есть припадок (вокализирует, показывает кивок головой, клонус передней конечности, разгибание задней конечности, падает на бок или испытывает генерализованные тонические / клонические судороги), запишите следующую информацию.
- Запишите температуру тела мыши во время припадка (температуру порога припадка) с ректального термометра температуры.
- Запишите характеристики судорожного поведения, такие как кивок головы, клонус передней конечности, разгибание задних конечностей, падение на бок и / или генерализованные тонические / клонические судороги (GTCS), отображаемые мышью.
- Быстро, но осторожно поднимите мышь из камеры и поместите ее на охлаждающую подушку, подготовленную на шаге 1.6.
ПРИМЕЧАНИЕ: Если мышь испытывает судороги шкалы Расина 5 и демонстрирует неконтролируемые прыжки, может быть трудно поднять животное из тепловой камеры и перенести на охлаждающую площадку снаружи. Однако типичный тепловой припадок длится от 30 с до 60 с. Таким образом, мышь должна быть извлечена из тепловой камеры и помещена на охлаждающую подушку в течение 60 с после начала эпизода теплового припадка. - Подождите, пока температура тела мыши снизится до 36-37 °C, прежде чем переносить ее в клетку для восстановления. Одновременно в клетку для восстановления помещается только одна мышь.
ПРИМЕЧАНИЕ: Не смешивайте мышей, которые еще не используются для теплового скрининга, с мышью, которая уже испытала испытание эксперимента с судорогами, вызванными теплом. - Аккуратно и осторожно отрежьте ленту между хвостом мыши и ректальным зондом с помощью ножниц, чтобы удалить ректальный зонд у мыши.
- Протрите металлический наконечник ректального зонда 70% спиртом и салфетками для мягких тканей, чтобы подготовить его к следующему испытанию.
- Продолжайте наблюдать за мышью в клетке восстановления, пока она не возобновит нормальную активность (ходьба, груминг и т. Д.), Прежде чем вернуть мышь в ее домашнюю клетку. Это знаменует собой КОНЕЦ экспериментального испытания для этой мыши.
- Запишите состояние животного после анализа живым и восстановленным после тестового сеанса или мертвым. Судороги высокой интенсивности, включающие неконтролируемые прыжки и генерализованные тонические / клонические припадки, иногда могут привести к смерти мыши.
- Если мышь не испытывает тепловых судорог в течение 30-минутного периода наблюдения или температура тела мыши достигает 44 °C, выньте мышь из тепловой камеры и поместите на охлаждающую подушку до тех пор, пока температура тела мыши не вернется к 36-37 °C.
- Сбросьте температуру тепловой камеры мыши до 50 °C и позвольте ей уравновеситься до тех пор, пока температура дисплея на цифровом регуляторе температуры не покажет 50 °C.
- Меняйте постельное белье между отдельными испытаниями на мышах.
- Подготовьте следующую мышь к скрининговому испытанию, как описано в разделе 2, и повторите шаги из раздела 3.
4. Усыпление животных
- Хотя большинство животных восстанавливаются после приступов, вызванных жарой, по нашему опыту, некоторые из мышей подвергаются SUDEP (внезапной необъяснимой смерти в EPilepsy) в своей домашней клетке в течение 24-48 часов после судорог, вызванных теплом. После завершения скрининга на всех мышах индивидуально на предмет тепловых судорог после 30-минутного испытания усыпьте всех мышей в соответствии с руководящими принципами IACUC учреждения.
5. Анализ данных об изъятиях, вызванных теплом
- После завершения скрининга когорты животных рассчитайте процент мышей в данном генотипе, показывающих судороги, используя следующую формулу:

- Оцените среднюю пороговую температуру захвата мышей в пределах данного генотипа путем усреднения пороговой температуры захвата всех мышей (отмеченной на этапе 3.7) в том генотипе, который демонстрирует тепловые индуцированные судороги.
- Все еще будучи слепым к личности и генотипу, воспроизводите видеозаписи каждой из мышей во время скрининга на экран компьютера, вызванного теплом, на экране компьютера, чтобы оценить тяжесть приступов судорог.
- Дайте оценки отдельным мышам, демонстрирующим вызванное теплом судорожное поведение, используя модифицированную шкалу Расина13, как описано в предыдущих исследованиях13,14. Подробную информацию см. в таблице 1.
- Если мышь, испытывая приступы, вызванные жарой, показывает только кивок головой, дайте ей оценку 2. Если мышь начинает эпизод припадка с кивком головы, но также демонстрирует клонус передней конечности, падение и / или прыжок, дайте ей оценку 5.
- Запишите максимальный балл для каждой мыши, используя модифицированную шкалу Расина13 , как описано выше.
- Постройте график рассеяния максимальных баллов Расина, демонстрируемых всеми мышами в данном генотипе.
- Статистически сравните максимальные баллы Расина среди различных групп мышей в качестве метода определения тяжести поведенческих припадков, таких как судороги, вызванные жарой.
ПРИМЕЧАНИЕ: Оценки Расина полезны для сравнения характеристик судорог между различными группами мутантных мышей или генотипами. Ожидается, что мыши дикого типа не будут подвергаться тепловым судорогам и не должны будут рассматриваться для сравнения показателей Расина. - Основываясь на экспериментальном дизайне, выполните соответствующий статистический анализ, чтобы определить, значительно ли отличается процент мышей, демонстрирующих судороги между мышами дикого типа и мутантами, и их средние пороговые значения температуры захвата друг от друга.
| Расин Партитура | Характеристики судорог | ||
| 0 | Без судорог | ||
| 1 | Движения во рту и лице | ||
| 2 | Кивок головой | ||
| 3 | Передний клонус, обычно одна конечность | ||
| 4 | Передний клонус с выращиванием | ||
| 5 | Генерализованный тонико-клонический припадок, вздыбление, прыжки, падение | ||
Таблица 1: Расин забивает.
Результаты
Ожидается, что животные модели с фебрильными мутациями судорог будут подвергаться вызванным жарой судорогам при повышенных температурах тела, которые не вызывают судорог у партнеров по помету дикого типа. Мутации SCN1A были связаны с фебрильными судорогами, включая пациентов с K1270T GEFS+, у которых наблюдаются как фебрильные, так и афебрильные генерализованные судороги7. Мы проверили CRISPR генерируемых SCN1A K1270T GEFS+ мутантных мышей, недавно описанных в исследовании14 , на возникновение тепловых судорог в двух генетических фонах - устойчивых к судорогам 129X1 / SvJ (129X1) и восприимчивых к судорогам C57BL / NJ (B6N) фонах. Соответствующие возрасту пометные товарищи дикого типа в тепловой камере мыши, которые не содержат никаких мутаций GEFS+ и, следовательно, не должны проявлять вызванные теплом судороги, служившие в качестве контрольной группы. Скорость изменения температуры тела с течением времени оценивали путем построения средней температуры тела мышей, регистрируемой каждую минуту во время анализа. Не было выявлено различий в скорости изменения температуры тела между гетерозиготными мутантными мышами и пометниками дикого типа, протестированными в соответствующих генетических фонах 129X1 и B6N (рисунок 2B, C). Это говорит о том, что терморегуляция не изменяется у гетерозиготных мутантных мышей K1270T GEFS+.
Все гетерозиготные мыши-мутанты из генетического фона 129X1 (n = 15) или B6N (n = 9) демонстрировали тепловые судороги (рисунок 2D). Ни у одной из мышей дикого типа на обогащенном фоне 129X1 (n = 13) не наблюдались теплово-индуцированные судороги (рисунок 2D). Напротив, треть протестированных мышей (n = 3 из 9 мышей) в чувствительном к судорогам фоне B6N демонстрировали судороги, вызванные теплом. Статистическое сравнение показывает, что процент гетерозиготных мутантных мышей, демонстрирующих тепловые припадки, был значительно выше, чем у их соответствующих мышей-коллег дикого типа как в генетическом фоне 129X1, так и b6N (рисунок 2D, точный тест Фишера, 129X1 p < 0,0001; B6NJ p = 0,009). Средняя пороговая температура захвата между гетерозиготными мутантными мышами в генетическом фоне 129X1 и B6N была схожей. Мыши-мутанты 129X1 имеют среднюю пороговую температуру захвата 42,6 ± 0,20 °C, что существенно не отличалось от средней пороговой температуры припадка 42,7 ± 0,06 °C, наблюдаемой у мышей B6N (Рисунок 2E; двуххвостый непарный t-тест Стьюдента, p = 0,782). Важно отметить, что средняя пороговая температура судорог у трех мышей дикого типа B6N, у которых наблюдались тепловые судороги, составляла 43,7 ± 0,08 °C и значительно выше, чем средний порог судороги 42,7 ± 0,06 °C, показанный гетерозиготными мутантными мышами B6N (рисунок 2E, двухвостый непарный t-тест Стьюдента, < p 0,0001).
Передняя часть камеры из плексигласа позволяет делать непрерывные видеозаписи во время анализа, которые могут быть использованы позже для оценки тяжести судорог у каждой мыши по модифицированной шкале Расина, как описано ранее14,20. Во время типичного анализа гетерозиготные мыши-мутанты демонстрировали вызванные жарой судороги с вокализацией и / или кивком головы (балл Расина 2) и быстрый переход в клонус передней конечности, падение на бок, прыжки, разгибание задних конечностей и / или генерализованные тонические / клонические судороги (баллы Расина 3-5), когда температура тела достигала около 42 ° C. Максимальный балл Расина представляет собой наиболее тяжелое судорожное поведение, вызванное жарой, среди мышей-мутантов. Максимальный показатель Расина гетерозиготных мутантных мышей в 129X1 обогащенном фоне (n = 15) не отличается от гетерозиготных мутантных мышей в генетическом фоне B6N (n = 9) (рисунок 2F; Тест Манна-Уитни, p > 0,9999). Это говорит о том, что индуцированные теплом характеристики судорожного поведения у мутантных мышей K1270T GEFS+ не зависят от фона штамма.
Взятые вместе, данные показывают, что все мыши-мутанты демонстрируют вызванные теплом судороги с одинаковой частотой, температурой порога судорог и тяжестью поведенческих припадков независимо от напряжения. Большинство самцов помета дикого типа не проявляют таких припадков при 44 °C или ниже. Около трети контрольных мышей дикого типа в чувствительном к судорогам B6N фоне действительно демонстрировали тепловые судороги (возможно, из-за эффектов генетического фона), но температура порога судорог была значительно выше по сравнению с мутантными мышами на том же фоне. Эти результаты свидетельствуют о том, что мыши-мутанты в генетическом фоне B6N восприимчивы к тепловым судорогам при более низких температурных порогах из-за мутации SCN1A GEFS+, которую они содержат. Таким образом, используя этот протокол, можно оценить тепловые припадки у мышей-мутантов эпилепсии и отличить от мышей дикого типа, которые либо не подвергаются тепловым припадкам, либо демонстрируют тепловые судороги при значительно более высоких температурах.

Рисунок 2: Мыши-мутанты демонстрируют приливы, вызванные теплом. (A) Протокол нагрева для поведенческого скрининга тепловых индуцированных судорог у мышей. (Б-С) Средняя температура тела мышей во времени у диких (Scn1a+/+ - черные треугольники) и гетерозиготных мутантных (Scn1aKT/+ - оранжевые круги) мышей в двух генетических фонах 129X1 и B6N соответственно. (D) Процент мышей, у которых наблюдались тепловые судороги в обоих генетических фонах. Дикие (Scn1a+/+) и гетерозиготные (Scn1aKT/+) мыши представлены черными и оранжевыми полосами соответственно. Гетерозиготные мутанты на фонах 129X1 и B6N показаны в оранжевых сплошных полосах и оранжевых полосах с черными полосами соответственно. (E) Порог судорожной температуры для индуцированных теплом судорог у мышей дикого типа (Scn1a+/+) и гетерозиготных мутантов (Scn1aKT/+) у обоих штаммов. (F) Распределение рассеяния максимальных показателей Расина по тепловым индуцированным судорогам, демонстрируемым гетерозиготными (Scn1aKT/+) мышами в обоих генетических фонах. Каждая точка представляет максимальный балл Расина за одну мышь. Количество животных в каждом генотипе показано в скобках. Данные, показанные на панелях B-F, являются средними ± S.E.M. Эта цифра изменена по сравнению с рисунком 3 в Das et al., 2021, eNeuro14. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
Обсуждение
Мы описываем простой и эффективный протокол скрининга возникновения тепловых судорог у мышей, поведенческий эквивалент фебрильных судорог у пациентов с человеческими заболеваниями. Анализ оценивает несколько параметров - в том числе процент мышей, показывающих судороги, порог судорог, тяжесть судорог по шкале Расина, чтобы сравнить чувствительность контрольных и тестовых групп мышей к повышению температуры тела.
Критический шаг в этом протоколе включает в себя увеличение тепла в камере при постоянном мониторинге температуры тела мыши. Крайне важно, чтобы максимальная температура тела, которую мыши будут испытывать в этих анализах, составляла 44 ° C, потому что дикие животные могут подвергаться тепловым судорогам при температуре тела >44 ° C. Предварительная обработка общей анестезией или анальгетиками может снизить температуру тела животных или помешать терморегуляции, что, в свою очередь, приведет к нарушению сбора данных о температуре судорожного порога. Таким образом, мыши в соответствии с этим протоколом скрининга не могли быть обеспечены этими агентами в течение 30-минутного испытательного окна. Все процедуры должны быть одобрены комитетом IACUC учреждения. Чтобы обеспечить непрерывный мониторинг температуры тела мыши во время анализа, надежно прикрепите ректальный температурный зонд к хвосту мышей. Если во время анализа температура тела мыши остается неизменной в течение длительных периодов времени даже после повышения температуры мышиной камеры, убедитесь, что ректальный температурный зонд не вышел из мыши или не прикреплен к хвосту.
Генетический фон мышиных моделей может влиять на чувствительность к мутации SCN1A и фармакологически индуцированным судорогам18,25,26,27. Как обсуждалось в результатах выше, генетический фон мышей может влиять на их восприимчивость к тепловым судорогам. Scn1a Мутантные мыши K1270T GEFS+ были протестированы на двух генетических фонах - 129X1 и B6NJ, и небольшой процент мышей дикого типа (33%) в чувствительном к судорогам фоне B6NJ также подвергался тепловым припадкам. Однако, по сравнению с гетерозиготными мутантными мышами Scn1aKT/+, мыши дикого типа B6NJ испытывали тепловые судороги при значительно более высоком температурном пороге. Это подтверждает, что генетическая мутация (Scn1a K1270T), которая была введена с помощью CRISPR knock-in, делает мутантных мышей более восприимчивыми к судорогам, вызванным гипертермией.
Принятие этого протокола имеет ряд преимуществ, которые кратко излагаются ниже. Во-первых, в отличие от использования потока сухого воздуха или нагреваемых ламп, контролируемый температурой принудительный воздух, установленный в закрытом пространстве, обеспечивает экспериментатору больший контроль над нагревом испытательной арены с желаемой скоростью. Этапы в протоколе нагрева могут быть легко изменены для увеличения / уменьшения начальной температуры, продолжительности каждого шага и т. Д. Для скрининга старых мышей, которые являются более тяжелыми или более крупными грызунами, такими как крысы. Во-вторых, непрерывный мониторинг температуры тела мыши с помощью прикрепленного ректального зонда дает ценную информацию о скорости изменения температуры тела у отдельных мышей на протяжении всего анализа. Это позволяет экспериментатору внимательно наблюдать, что скорость изменения температуры у мыши не превышает 0,25-0,5 °C/ мин (что может быть стрессом для животных), при адаптации этого протокола к другим тестовым аренам. Важно отметить, что скорость изменения температуры тела с течением времени в разных группах мышей может пролить свет на их способность к терморегуляции и может быть полезна для понимания того, изменяют ли фебрильные судороги, вызывающие мутации, терморегуляцию у мышей. В-третьих, непрерывный мониторинг температуры тела гарантирует, что измерения пороговой температуры захвата с использованием этого протокола являются точными, поскольку они регистрируются одновременно с первым приступом припадка, испытываемым мышью. Если температура тела животного не контролируется непрерывно или температура порога судорог измеряется после выведения животного из тестовой арены, пороговые значения судорог могут варьироваться из-за времени, затрачиваемого на обработку мышей после судорог. Наконец, этот метод обходит необходимость использования инвазивных методов для индуцирования лихорадки (путем инъекции патогенов) у мышей, чтобы имитировать фебрильные судороги у пациентов.
Одним из ограничений этого протокола является то, что трудно проводить скрининг ювенильных (менее P30 в возрасте) мышей на предмет судорог, вызванных теплом. Протокол был разработан для скрининга чувствительности взрослых мышей (P30-P40 и выше) к судорогам, вызванным теплом или гипертермией. По нашему опыту, молодые мыши дикого типа, особенно те, которые весят менее 15 г, с большей вероятностью подвергаются теплово-индуцированным судорогам, что может быть связано с недостаточно развитыми механизмами терморегуляции, физиологическим тепловым стрессом или комбинацией того и другого. Следовательно, не идеально выполнять скрининг судорог, вызванный теплом, на молодых мышах с использованием этого протокола.
Будущие исследования, которые сочетают мониторинг ЭЭГ при одновременном воздействии на мышь теплово-индуцированных судорог, могут пролить свет на модели ЭЭГ-судорог, вызванных теплом, подобно предыдущему исследованию19. Активность нейронов в определенных областях мозга мыши может быть прослежена путем объединения оптогенетических подходов и исследований на основе иммуногистохимии после сбора ткани мозга. Кроме того, влияние ограничительных диет, таких как кето-диета, на снижение фебрильных судорог можно оценить, подвергая мышей, которых кормили кето, и нормальных мышей, которых кормили чау, протоколу судорог, вызванному теплом. Аналогичным образом, парадигмы скрининга лекарств от эпилепсии могут быть разработаны для тестирования и идентификации потенциальных противоэпилептических препаратов, которые улучшают или подавляют вызванные теплом судороги у мышей, которых кормили лекарствами или лечили, по сравнению с мышами, которых кормили транспортным средством или контрольными мышами.
Раскрытие информации
Авторы заявляют об отсутствии конфликта интересов.
Благодарности
Мы хотели бы поблагодарить Коннора Смита за его помощь в создании индивидуальной тепловой камеры мыши. Мы признаем помощь членов лаборатории O'Dowd, Лиши Цзэн и Эндрю Салгадо в стандартизации протокола нагрева на ранних этапах разработки анализа. Мы также благодарим Дэнни Бенавидеса и Кумара Перинбама за видеозапись частей экспериментальной процедуры рукописи. Эта работа была поддержана грантом NIH (NS083009), присужденным D.O.D.
Материалы
| Name | Company | Catalog Number | Comments |
| Axial fan | Farnam | AF20-200-120-xx10-3.1 | Farnam custom products -Axial Fan Heater with Fan |
| Digital temperature controller | Inkbird | ITC-100RH | Inkbird digital PID temperature controller ITC-100RH with K thermocouple |
| Mouse rectal temperature probe | ThermoWorks, Braintree Scientific, Inc | RET-3 | Mouse rectal temperature probe with thermometer |
Ссылки
- Hirtz, D., et al. How common are the 'common' neurologic disorders. Neurology. 68, 326-337 (2007).
- Catterall, W. A. Sodium Channel Mutations and Epilepsy. Jasper's Basic Mechanisms of the Epilepsies. , Center for Biotechnology Information. US. Internet (2012).
- Mantegazza, M., Broccoli, V. SCN 1A /Na V 1.1 channelopathies: Mechanisms in expression systems, animal models, and human iPSC models. Epilepsia. 60, (2019).
- Stafstrom, C. E. Persistent Sodium Current and Its Role in Epilepsy. Epilepsy Currents. 7, 15-22 (2007).
- Schutte, S. S., Schutte, R. J., Barragan, E. V., O'Dowd, D. K. Model systems for studying cellular mechanisms of SCN1A-related epilepsy. Journal of Neurophysiology. 115, 1755-1766 (2016).
- Wei, F., et al. Ion Channel Genes and Epilepsy: Functional Alteration, Pathogenic Potential, and Mechanism of Epilepsy. Neuroscience Bulletin. 33, 455-477 (2017).
- Abou-Khalil, B., et al. Partial and generalized epilepsy with febrile seizures plus and a novel SCN1A mutation. Neurology. 57, 2265-2272 (2001).
- Zhang, Y. -H., et al. Genetic epilepsy with febrile seizures plus: Refining the spectrum. Neurology. 89, 1210-1219 (2017).
- Patterson, K. P., et al. Enduring memory impairments provoked by developmental febrile seizures are mediated by functional and structural effects of neuronal restrictive silencing factor. Journal of Neuroscience. 37, 3799-3812 (2017).
- Rossi, M. A. SCN1A and febrile seizures in mesial temporal epilepsy: An early signal to guide prognosis and treatment. Epilepsy Currents. 14, 189-190 (2014).
- Zhang, Y., et al. Altered gut microbiome composition in children with refractory epilepsy after ketogenic diet. Epilepsy Research. 145, 163-168 (2018).
- Meng, H., et al. The SCN1A mutation database: Updating information and analysis of the relationships among genotype, functional alteration, and phenotype. Human Mutation. 36, 573-580 (2015).
- Cheah, C. S., et al. Specific deletion of NaV1.1 sodium channels in inhibitory interneurons causes seizures and premature death in a mouse model of Dravet syndrome. Proceedings of the National Academy of Science U.S.A. 109, 14646-14651 (2012).
- Das, A., et al. Interneuron dysfunction in a new mouse model of SCN1A GEFS. eNeuro. , (2021).
- Kalume, F., et al. Sudden unexpected death in a mouse model of Dravet syndrome. Journal of Clinical Investigations. 123, 1798-1808 (2013).
- Martin, M. S., et al. Altered function of the SCN1A voltage-gated sodium channel leads to gamma-aminobutyric acid-ergic (GABAergic) interneuron abnormalities. Journal of Biological Chemistry. 285, 9823-9834 (2010).
- Rubinstein, M., et al. Dissecting the phenotypes of Dravet syndrome by gene deletion. Brain. 138, 2219-2233 (2015).
- Yu, F. H., et al. Reduced sodium current in GABAergic interneurons in a mouse model of severe myoclonic epilepsy in infancy. Nature Neuroscience. 9, 1142-1149 (2006).
- Dutton, S. B. B., et al. Early-life febrile seizures worsen adult phenotypes in Scn1a mutants. Experimental Neurology. 293, 159-171 (2017).
- Cheah, C. S., et al. Specific deletion of NaV1.1 sodium channels in inhibitory interneurons causes seizures and premature death in a mouse model of Dravet syndrome. Proceedings of the National Academy of Science U.S.A. 109, 14646-14651 (2012).
- Oakley, J. C., Cho, A. R., Cheah, C. S., Scheuer, T., Catterall, W. A. Synergistic GABA-enhancing therapy against seizures in a mouse model of Dravet Syndrome. Journal of Pharmacology and Experimental Therapeutics. 345, 215-224 (2013).
- Ricobaraza, A., et al. Epilepsy and neuropsychiatric comorbidities in mice carrying a recurrent Dravet syndrome SCN1A missense mutation. Scientific Reports. 9, (2019).
- Warner, T. A., Liu, Z., Macdonald, R. L., Kang, J. -Q. Heat induced temperature dysregulation and seizures in Dravet Syndrome/GEFS+ Gabrg2+/Q390X mice. Epilepsy Research. 134, 1-8 (2017).
- Eun, B. -L., Abraham, J., Mlsna, L., Kim, M. J., Koh, S. Lipopolysaccharide potentiates hyperthermia-induced seizures. Brain and Behavior. 5, 00348(2015).
- Miller, A. R., Hawkins, N. A., McCollom, C. E., Kearney, J. A. Mapping genetic modifiers of survival in a mouse model of Dravet syndrome. Genes Brain and Behavior. 13, 163-172 (2013).
- Mistry, A. M., et al. Strain- and age-dependent hippocampal neuron sodium currents correlate with epilepsy severity in Dravet syndrome mice. Neurobiology of Disease. 65, 1-11 (2014).
- Ogiwara, I., et al. Nav1.1 localizes to axons of parvalbumin-positive inhibitory interneurons: a circuit basis for epileptic seizures in mice carrying an Scn1a gene mutation. Journal of Neuroscience. 27, 5903-5914 (2007).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеThis article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены