
Method Article
A Behavioral Screen for Heat-Induced Seizures in Mouse Models of Epilepsy
In This Article
Summary
The goal of the method is to screen for hyperthermia or heat-induced seizures in mouse models. The protocol describes the use of a custom-built chamber with continuous monitoring of the body temperature to determine whether elevated body temperature leads to seizures.
Abstract
Transgenic mouse models have proved to be powerful tools in studying various aspects of human neurological disorders, including epilepsy. The SCN1A-associated genetic epilepsies comprise a wide spectrum of seizure disorders with incomplete penetrance and clinical variability. SCN1A mutations can result in a large variety of seizure phenotype ranging from simple, self-limited fever-associated febrile seizures (FS), moderate-level genetic epilepsy with febrile seizures plus (GEFS+) to more severe Dravet Syndrome (DS). Although FS are commonly seen in children below 6-7 years of age who do not have genetic epilepsy, FS in GEFS+ patients continue to occur into adulthood. Traditionally, experimental FS have been induced in mice by exposing the animal to a stream of dry air or heating lamps, and the rate of change in body temperature is often not well controlled. Here, we describe a custom-built heating chamber, with a plexiglass front, that is fitted with a digital temperature controller and a heater-equipped electric fan, which can send heated forced air into the test arena in a temperature-controlled manner. The body temperature of a mouse placed in the chamber, monitored through a rectal probe, can be increased to 40-42 °C in a reproducible manner by increasing the temperature inside the chamber. Continual visual monitoring of the animals during the heating period demonstrates induction of heat-induced seizures in mice carrying an FS mutation at a body temperature that does not elicit behavioral seizures in wild-type litter mates. Animals can be easily removed from the chamber and placed on a cooling pad to rapidly return body temperature to normal. This method provides for a simple, rapid, and reproducible screening protocol for the occurrence of heat-induced seizures in epilepsy mouse models.
Introduction
Epilepsy, the fourth most common family of neurological disorders in the United States1, are characterized by an imbalance of excitatory and inhibitory drive in the CNS that leads to recurrent seizures. Febrile seizures (FS) or fever associated seizures can occur in the general population, most often in children as early as 3 months up through 6 -7 years of age. However, in some individuals with genetic mutations, most often in a sodium channel gene, FS can persist beyond the age of 7 years into adulthood. This condition is referred to as febrile seizures plus or FS+. Rapid advances in genome sequencing has identified over 1,300 mutations in the human sodium ion channel gene SCN1A, making it a hotspot for epilepsy mutations. SCN1A mutations have been linked to a wide spectrum of seizure disorders, including febrile seizures (FS), genetic epilepsy with febrile seizures plus (GEFS+), and Dravet Syndrome (DS)2,3,4,5,6. About 20% of SCN1A missense mutations leads to GEFS+5,7,8. Pediatric history of complex or prolonged FS in childhood can subsequently develop into more debilitating forms of epilepsy such as temporal lobe epilepsy (TLE)9,10,11. Dravet Syndrome arises due to truncation mutations or loss of function mutations in SCN1A and is a severe form of intractable epilepsy, with childhood onset of febrile seizures that develop into refractory seizures, and is often associated with cognitive, developmental, and motor impairments2,5,12. Since many individuals with GEFS+ and/or DS exhibit febrile seizures, it becomes imperative to develop novel therapies to better combat these seizure disorders.
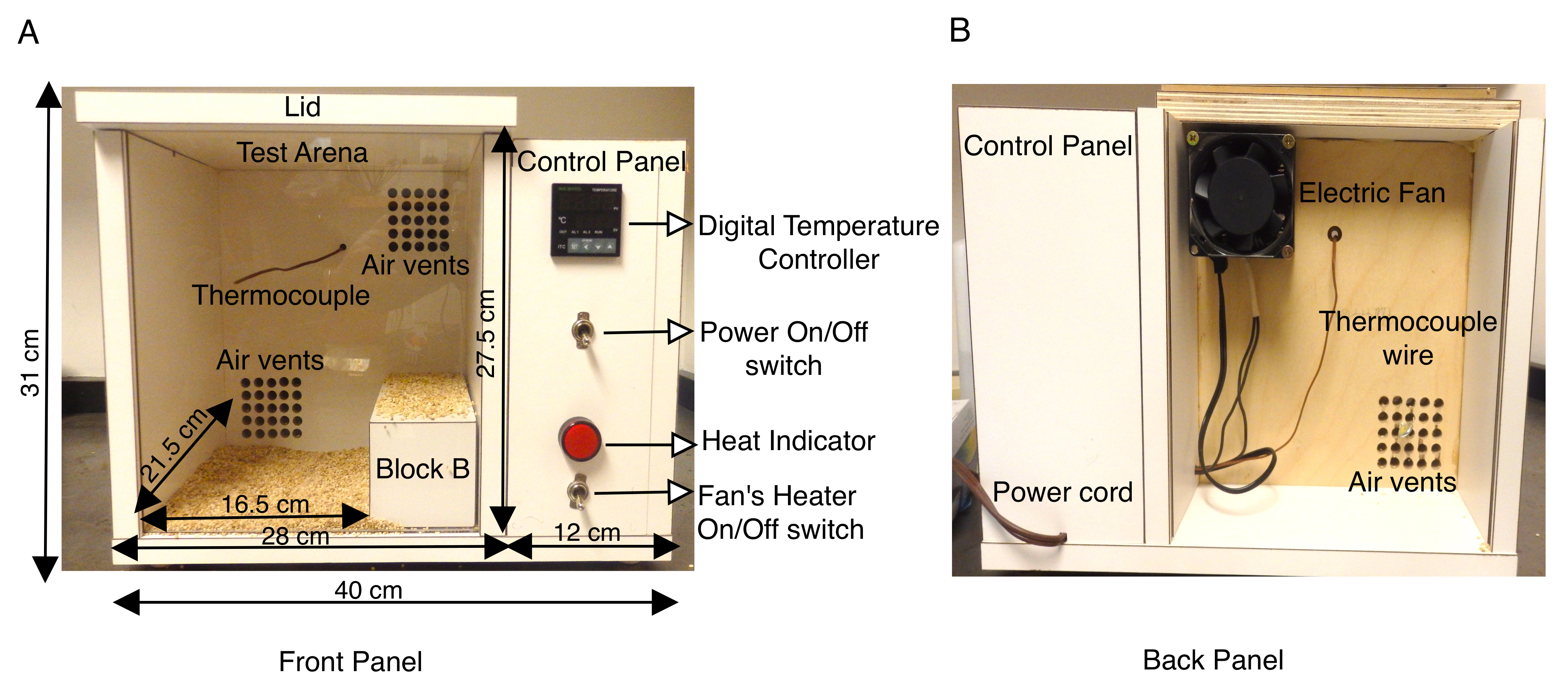
Animal models of SCN1A associated epilepsy have proven invaluable in characterizing different types of seizures (febrile vs generalized) and dissecting the neuronal mechanism of seizure generation13,14,15,16,17,18. While the study of spontaneous seizures via EEG/EMG recordings in rodent brains is well established and is a very useful tool, only a few studies have attempted to mimic febrile seizures in mouse models14,16,19,20,21,22,23. Previous studies have used a jet of heated dry air, or a methacrylate cylinder fitted with a thermal system, or heat lamps with a temperature controller in enclosed test arenas9,16,21,22,23,24 to induce seizures via hyperthermia. In order to increase body temperature in a more controlled environment, the protocol described here uses a custom-built chamber with a temperature-controlled heating system that allowed reproducible rates of increase in the body temperature of a mouse inside the chamber. The heat chamber was constructed from wood (length 40 cm x width 34 cm x height 31 cm) and was fitted with a digital temperature controller with a K thermocouple. A small axial fan equipped with a heater at the back panel of the chamber directs heated air into the chamber regulated by a digital temperature controller. This forced air heating system enables one to control the rate at which the chamber temperature increases. (Figure 1A,B). The K thermocouple located inside the wooden heat chamber sends feedback to the digital temperature controller, to maintain constant temperatures inside of the box during the assay. Setting the temperature on the digital temperature controller, enables the electric fan to send heated forced air through vents to uniformly heat the chamber (Figure 1A). The front panel of the heat chamber is a clear plexiglass sheet to enable easy video recording of the trials.
Adult (P30-P40) mice, heterozygous for a missense mutation in SCN1A that causes GEFS+ and an equal number of wild-type litter mates to serve as the control group, were selected for each experiment. Animals, both male and female, used in these studies weighed at least 15 g as wild-type mice weighing less were more sensitive to heat-induced seizures than heavier animals of the same age. In the pilot study, both mutant and wild-type mice were observed to seek out the cooler corners of the chamber at the back and remained there for prolonged periods of time. To circumvent this, effective floor size inside the heat chamber test arena was reduced to length 16.5 cm x width 21.5 cm x height 27.5 cm by placing a wooden block B (dimensions 20 cm x 8cm x 7.2 cm) at the right side of the chamber (Figure 1A). The heat chamber was constructed from 1.9 cm thick plywood (length 40 cm x width 34 cm x height 31 cm) covered with white laminate and fitted with a digital temperature controller with a K thermocouple. The laminate surface of the chamber walls is impermeable and can be easily sanitized between trials by wiping down with 70% ethanol. The temperature of the heat chamber was initially set at 50 °C and preheated for at least 1 h before the start of the experiment, to ensure uniform heating inside the chamber. Each mouse was fitted with a rectal thermometer for continuous monitoring of body temperature throughout the experiment. A single mouse was placed in the chamber at a time and the temperature was kept at 50 °C between 1st-10th minute. The temperature was then raised to 55 °C for 11th-20th minute, and finally raised to 60 °C for 21st-30th minute. This resulted in a reproducible rate of increase in the mouse body temperature (Figure 2A). Each trial was video-taped and behavioral analysis was conducted offline.
The heating protocol can be easily modified to change the initial temperature of the heat chamber and the rate that the chamber is heated, which in turn changes how quickly the body temperature of the mouse is elevated during the assay. Thus, this method provides more flexibility over traditional methods in setting up the behavioral screens involving heat-induced seizures. The heat-induced seizure protocol can also be used to screen for anti-epileptic drugs that make mutant mice more resistant to heat-induced seizures or increase the threshold temperature at which seizures are observed. Similarly, beneficial effects of restrictive diet regimes such as keto diet on heat-induced seizures can be examined in normal chow-fed vs keto-fed mice.

Figure 1: Description of the custom-built mouse heat chamber. (A) The front panel of the wooden mouse heat chamber shows the side control panel containing Power ON/OFF switch that turns on digital temperature controller, K thermocouple, fan heater's ON/OFF switch and heat indicator. The outer dimensions of the box and the inner test arena are shown in cm. A wooden block B used to effectively reduce test arena surface is also shown. The bottom of the test arena is covered with cob bedding to prevent mice from directly coming in contact with heated wooden surfaces. (B) The back panel of the heat chamber shows the fan mounted on the top air vent and the power cord to supply electricity to the chamber. This figure is modified from Figure 3 in Das et al., 2021, eNeuro14. Please click here to view a larger version of this figure.
{kind=link}
Protocol
All animal procedures were performed in accordance with the guidelines of Institutional Animal Care and Use Committee (IACUC) at University of California, Irvine.
1. Preparation for the heat-induced seizure assay
- Switch on the Power On button on the heat chamber, followed by the Heat On button.
- Set the temperature of the heat chamber at 50 °C using the keypad on the digital temperature controller.
- Wait for a minimum of 1 h to preheat the chamber at 50 °C before introducing the first mouse into the chamber. Preheating ensures uniform heating inside the chamber.
- Line the floor of the mouse heat chamber with cob bedding.
- Mount a video recording camera in front of the heat chamber for recording each heat-induced seizure assay trial.
- Line a 140 mm diameter Petri dish with thick layers of tissue paper and place it on ice to serve as a cooling pad.
NOTE: At the end of the assay, individual mouse will be transferred on the prechilled cooling pad to help bring down their elevated body temperature.
2. Preparing the mouse for heat-induced seizure assay
- Select 10 adult mice (P30-P40), 5 that carry the epilepsy causing mutation and 5 of the wild-type litter mates for heat-induced seizure screening assay.
NOTE: Wild-type mice, not harboring any epilepsy causing mutation do not exhibit heat-induced seizures at temperatures below 44 °C and serve as the control group. - Weigh each mouse to be used for the screening assay and record its body weight. Only mice weighing 15 g or more should be used for the assay.
- Screen one mouse at a time in the mouse heat chamber.
- Briefly anesthetize the mouse for 10-15 s using a few drops of isoflurane at the bottom of a bell jar.
- Take the animal out of the bell jar and place it on a paper towel.
- Ensure that the mouse is completely anesthetized by checking that the mouse is unresponsive to a noxious toe pinch.
- Coat the metal tip of the rectal temperature probe with a lubricant (such as petroleum jelly) and gently insert it into the mouse at a depth of less than or equal to 2 cm.
- Secure the rectal probe to the mouse's tail with tape, so the probe does not come out during the assay.
NOTE: Alternatively, place the animal in a mouse restrainer cone and insert the rectal temperature probe. Secure it by taping to the tail. - Ensure the rectal probe is connected to a multimeter that displays internal body temperature of the mouse.
- Place the animal in a fresh cage lined with cob bedding, i.e., the recovery cage.
- Start a timer and wait for 5 min. Observe the mouse until it has completely recovered from anesthesia and the mouse is active and grooming.
- Simultaneously, monitor the core body temperature of the mouse till it stabilizes at 35-36 °C.
- At the end of 5 min, note the body temperature of the mouse. This is the initial body temperature at time "0" min.
NOTE: If the core body temperature of the mouse is below 35 °C, wait for additional time for the animal to recover from anesthesia-induced hypothermia. - Quickly, transfer the individual mouse into the preheated mouse chamber. This marks the START of the experiment trial. Only one mouse is screened at a given time.
3. Heat-induced seizure assay
- After gently placing the mouse on the floor of the pre-heated mouse heat chamber, close the plexiglass door and start the camera for video recording the experiment.
- Start the stopwatch. Record the body temperature of the mouse from the rectal thermometer at 1 min intervals for the duration of the experiment.
- At regular intervals, increase the temperature of the mouse heat chamber such that the body temperature of the mouse increases at a rate of 0.25-0.5 °C /min.
NOTE: Rapid increases in body temperature can lead to heat stroke or death and should be avoided. - Following this protocol, increase the temperature of the mouse heat chamber by 5 °C every 10 min as shown in Figure 2A.
- At 9.5 min, set the temperature of the heat chamber to 55 °C, to stabilize the temperature of the heat chamber to 55 °C by the 10th min as shown on the digital temperature display.
- Similarly, increase the temperature to 60 °C at 19.5 min to stabilize the temperature of the heat chamber to 60 °C by the 20th min. Each seizure screening trial lasts for 30 min.
- If the mouse has a seizure (vocalizes, shows head nodding, forelimb clonus, hindlimb extension, falls on its side, or experiences generalized tonic/clonic convulsions), record the following information.
- Record the body temperature of the mouse during the seizure (seizure threshold temperature) from the rectal temperature thermometer.
- Record the seizure behavior characteristics such as head nodding, forelimb clonus, hind limb extension, falling on the side, and/or generalized tonic/clonic seizures (GTCS) displayed by the mouse.
- Quickly but gently pick up the mouse from the chamber and place it on the cooling pad prepared in step 1.6.
NOTE: If a mouse is experiencing Racine scale 5 seizures and exhibiting uncontrolled jumping, it can be difficult to pick up the animal from the heat chamber and transferred to the cooling pad outside. However, a typical heat-induced seizure lasts between 30 s to 60 s.Thus, the mouse should be taken out of the heat-chamber and put on the cooling pad within 60 s of start of heat-induced seizure episode. - Wait for the mouse body temperature to come down to 36-37 °C, before transferring it to a recovery cage. Only one mouse is placed in a recovery cage at a time.
NOTE: Do not mix mice that are yet to be used for heat-induced screening with the mouse that has already experienced the heat-induced seizure experiment trial. - Gently and carefully, cut the tape between the mouse tail and rectal probe wire with a pair of scissors to remove the rectal probe from the mouse.
- Wipe clean the metal tip of the rectal probe with 70% alcohol and soft tissue wipes to keep it ready for the next trial.
- Continue to observe the mouse in the recovery cage until it resumes normal activity (walking, grooming, etc.), before returning the mouse to its home cage. This marks the END of the experiment trial for this mouse.
- Record the animal status after the assay-alive and recovered from the test session or dead. High intensity seizures involving uncontrolled jumping and generalized tonic/clonic seizures can sometimes result in the death of the mouse.
- If a mouse does not experience heat-induced seizures within the 30 min observation period or the body temperature of the mouse reaches 44 °C, remove the mouse from the heat chamber and place on the cooling pad till the body temperature of the mouse returns to 36-37 °C.
- Reset the temperature of the mouse heat chamber to 50 °C and allow it to equilibrate till the display temperature on the digital temperature controller shows 50 °C.
- Change cob bedding between individual mouse trials.
- Prepare the next mouse for screening trial as described in section 2 and repeat the steps from section 3.
4. Euthanizing the animals
- Although most animals recover post heat-induced seizures, in our experience, a few of the mice undergo SUDEP (Sudden Unexplained Death in EPilepsy) in their home cage within 24-48 hrs of heat-induced seizures. After concluding the screening on all the mice individually for heat-induced seizures following the 30 min trial, euthanize all the mice as per the institution's IACUC guidelines.
5. Analyzing the heat-induced seizure data
- After completing the screening of a cohort of animals, calculate the percentage of mice in a given genotype showing seizures using the following formula:

- Estimate the mean seizure threshold temperature of mice within a given genotype by averaging the seizure threshold temperature of all mice (noted in step 3.7) in that genotype that exhibit heat-induced seizures.
- While still being blind to the identity and genotype, replay the video recordings of each of the mouse during the heat-induced seizure assay screening on a computer screen to score severity of seizure bouts.
- Give scores to individual mouse exhibiting heat-induced seizure behavior by using the modified Racine scale13 as described by previous studies13,14. See Table 1 for details.
- If a mouse, while experiencing heat-induced seizures, only shows head nodding, give it a score of 2. If a mouse starts a seizure episode with head nodding but also exhibits forelimb clonus, falling over, and/or jumping give it a score of 5.
- Record the maximum score for each mouse using the modified Racine scale13 as described above.
- Plot a scatter graph of maximum Racine scores exhibited by all mice in a given genotype.
- Statistically compare maximum Racine scores among different mouse groups as a method to determine the severity of behavioral seizures such as heat-induced seizures.
NOTE: Racine scores are helpful to compare seizure characteristics between different mutant mice groups or genotypes. It is expected that the wild-type mice would not undergo heat-induced seizures and would not have to be considered for Racine score comparisons. - Based on experimental design, perform appropriate statistical analysis to determine if the percentage of mice exhibiting seizures between wild-type and mutant mice, and their mean seizure threshold temperature values are significantly different from each other.
| Racine Score | Seizure characteristics | ||
| 0 | No seizures | ||
| 1 | Mouth and facial movements | ||
| 2 | Head nodding | ||
| 3 | Forelimb clonus, usually one limb | ||
| 4 | Forelimb clonus with rearing | ||
| 5 | Generalized tonic-clonic seizure, rearing, jumping, falling over | ||
Table 1: Racine scores.
Results
Animal models with febrile seizure mutations are expected to undergo heat-induced seizures at elevated body temperatures that do not induce seizures in the wild-type litter mates. SCN1A mutations have been linked with febrile seizures, including K1270T GEFS+ patients, who display both febrile and afebrile generalized seizures7. We screened CRISPR generated SCN1A K1270T GEFS+ mutant mice recently described in a study14 for the occurrence of heat seizures in two genetic backgrounds - seizure resistant 129X1/SvJ (129X1) and seizure susceptible C57BL/NJ (B6N) backgrounds. Age matched wild-type litter mates in the mouse heat chamber which do not harbor any GEFS+ mutations and thus, are not expected to exhibit heat-induced seizures, served as the control group. The rate of the body temperature change over time was evaluated by plotting mean body temperature of mice recorded every minute during the assay. There was no difference in the rate of change of body temperature between heterozygous mutant mice and wild-type litter mates tested in respective 129X1 and B6N genetic backgrounds (Figure 2B,C). This suggests that thermoregulation is not altered in K1270T GEFS+ heterozygous mutant mice.
All heterozygous mutant mice from 129X1 (n = 15) or B6N (n = 9) genetic backgrounds exhibited heat-induced seizures (Figure 2D). None of the wild-type mice in the 129X1 enriched background (n = 13) exhibited heat-induced seizures (Figure 2D). In contrast, a third of the mice tested (n = 3 out of the 9 mice) in the seizure sensitive B6N background exhibited heat-induced seizures. Statistical comparison shows that percentage of heterozygous mutant mice exhibiting heat-induced seizures was significantly higher than their respective wild-type counterpart mice in both the 129X1and B6N genetic backgrounds (Figure 2D, Fisher's exact test, 129X1 p < 0.0001; B6NJ p = 0.009). The average seizure threshold temperature between the heterozygous mutant mice in 129X1 and B6N genetic backgrounds was similar. 129X1 mutant mice have a mean seizure threshold temperature of 42.6 ± 0.20 °C, which was not significantly different from the mean seizure threshold temperature of 42.7 ± 0.06 °C seen in B6N mice (Figure 2E; two-tailed unpaired Student's t-test, p = 0.782). It is important to note that the mean seizure threshold temperature of three B6N wild-type mice that exhibited heat-induced seizures was 43.7 ± 0.08 °C and significantly higher than the mean seizure threshold of 42.7 ± 0.06 °C displayed by B6N heterozygous mutant mice (Figure 2E, two-tailed unpaired Student's t-test, p < 0.0001).
The plexiglass front of the chamber makes it possible to do continuous video recordings during the assay that can be used later to score for seizure severity in each mouse on a modified Racine scale as described previously14,20. During a typical assay, heterozygous mutant mice would display heat-induced seizures with vocalization and/or head-nodding (Racine score 2), and rapidly transition to forelimb clonus, falling on side, jumping, hindlimb extension, and/or generalized tonic/clonic seizures (Racine scores 3-5) when body temperature reached about 42 °C. The maximum Racine score represents the most severe heat-induced seizure behavior among the mutant mice. The maximum Racine score of heterozygous mutant mice in 129X1 enriched background (n = 15) is not different from heterozygous mutant mice in B6N (n = 9) genetic background (Figure 2F; Mann-Whitney test, p > 0.9999). This suggests that heat-induced seizure behavior characteristics in K1270T GEFS+ mutant mice are independent of strain background.
Taken together, the data demonstrate that all mutant mice exhibit heat-induced seizures with similar frequency, seizure threshold temperature, and behavioral seizure severity in a strain-independent manner. The majority of wild-type litter mates do not exhibit such seizures at or below 44 °C. About one-third of the wild-type control mice in a seizure sensitive B6N background did display heat-induced seizures (possibly due to genetic background effects) but the seizure threshold temperature was significantly higher compared to mutant mice in the same background. These results suggest that mutant mice in B6N genetic background are susceptible to heat-induced seizures at lower temperature thresholds due to the SCN1A GEFS+ mutation they harbor. Thus, using this protocol, one can evaluate heat-induced seizures in epilepsy mutant mice and distinguish from wild-type litter mate mice, which either do not undergo heat-induced seizures or display heat seizures at significantly higher temperatures.

Figure 2: Mutant mice exhibit heat-induced seizures. (A) The heating protocol for behavioral screening of heat-induced seizures in mice. (B-C) Mean body temperature of mice across time in wild-type (Scn1a+/+ - black triangles) and heterozygous mutant (Scn1aKT/+ - orange circles) mice in two genetic backgrounds 129X1 and B6N, respectively. (D) Percentage of mice showing heat-induced seizures in both genetic backgrounds. Wild-type (Scn1a+/+) and heterozygous (Scn1aKT/+) mice are represented by black and orange bars, respectively. Heterozygous mutants in 129X1 and B6N backgrounds are shown in orange solid bars and orange bars with black stripes, respectively. (E) Seizure temperature threshold to heat-induced seizures in wild-type (Scn1a+/+) and heterozygous mutant (Scn1aKT/+) mice in both strains. (F) Scatter distribution of maximum Racine scores of heat-induced seizures exhibited by heterozygous (Scn1aKT/+) mice in both genetic backgrounds. Each dot represents maximum Racine score in a single mouse. Number of animals in each genotype is shown within parentheses. Data shown in panels B-F are mean ± S.E.M. This figure is modified from Figure 3 in Das et al., 2021, eNeuro14. Please click here to view a larger version of this figure.
{kind=link}
Discussion
We describe a simple and effective protocol to screen for occurrence of heat-induced seizures in mice, the behavioral equivalent of febrile seizures in human patients. The assay evaluates several parameters - including the percentage of mice showing seizures, seizure threshold, severity of seizures on a Racine scale, in order to compare the sensitivity of control and test mice groups to increases in body temperature.
A critical step in this protocol involves increasing the heat in the chamber while continuously monitoring the body temperature of the mouse. It is imperative that the maximum body temperature the mice will experience in these assays is 44 °C because wild-type animals can undergo heat-induced seizures at body temperatures >44 °C. Pretreatment with general anesthesia or analgesics might reduce core body temperature of animals or interfere with thermoregulation, which in turn would confound seizure threshold temperature data collection. Thus, mice under this screening protocol could not be provided with these agents during the 30 min trial window. All procedures must be approved by the institution's IACUC committee. To ensure continuous monitoring of core body temperature of the mouse during the assay, securely tape the rectal temperature probe to the tail of the mice. If during the assay, the mouse body temperature is found to remain unchanged for prolonged periods of time even after increasing the temperature of the mouse chamber, ensure that the rectal temperature probe has not come out of the mouse or is attached loosely to the tail.
Genetic background of mouse models can affect sensitivity to the SCN1A mutation and pharmacologically induced seizures18,25,26,27. As discussed in the results above, the genetic background of the mice can influence their susceptibility to heat-induced seizures. Scn1a K1270T GEFS+ mutant mice were tested in two genetic backgrounds - 129X1 and B6NJ, and a small percentage of wild-type mice (33%) in the seizure sensitive B6NJ background, were also observed to undergo heat-induced seizures. However, in comparison to the heterozygous mutant Scn1aKT/+ mice, the B6NJ wild-type mice experienced heat-induced seizures at a significantly higher temperature threshold. This confirms that the genetic mutation (Scn1a K1270T) that was introduced by CRISPR knock-in makes the mutant mice more susceptible to hyperthermia-induced seizures.
There are several advantages of adopting this protocol, which are summarized below. First, unlike the use of stream of dry air or heated lamps, a temperature-controlled forced air set up within an enclosed space provides the experimenter more control over heating up the test arena at a desired rate. The steps in the heating protocol can be easily modified to increase/decrease the starting temperature, duration of each step, etc. to screen older mice that are heavier or larger rodents such as rats. Second, continuous monitoring of mouse body temperature via the attached rectal probe, gives valuable information about the rate of body temperature change in individual mouse, throughout the assay. This allows the experimenter to closely observe that the rate of temperature change in the mouse does not exceed 0.25-0.5 °C/min (which might be stressful for the animals), when adapting this protocol to other test arenas. Importantly, the rate of change of body temperature across time in different mice groups can shed light on their ability to thermoregulate and could be helpful to understand whether febrile seizure causing mutations also alter thermoregulation in mice. Third, continuous body temperature monitoring ensures that the seizure threshold temperature measurements using this protocol are accurate, since they are recorded concurrently with the first bout of seizure experienced by the mouse. If the body temperature of the animal is not continuously monitored or seizure threshold temperature is measured after taking the animal out of the test arena, seizure threshold values can vary due to the time taken to handle the mice post seizures. Finally, this method circumvents the need to use invasive methods to induce fever (by injecting pathogens) in mice to mimic febrile seizures in human patients.
One of the limitations of this protocol is that it is difficult to screen juvenile (less than P30 in age) mice for heat-induced seizures. The protocol was developed to screen for sensitivity of adult mice (P30-P40 and above) to heat- or hyperthermia-induced seizures. In our experience, the younger wild-type mice, especially those weighing below 15 g, are more likely to undergo heat-induced seizures, which could be due to underdeveloped thermoregulation mechanisms, physiological thermal stress, or a combination of both. Hence, it is not ideal to perform the heat-induced seizure screen on juvenile mice using this protocol.
Future studies that combine EEG monitoring while subjecting the mouse to heat-induced seizures can shed light on EEG seizure patterns of heat-induced seizures, similar to a previous study19. Neuronal activity in specific areas in the mouse brain can be traced by combining optogenetic approaches and immunohistochemistry-based studies after harvesting the brain tissue. Also, the effects of restrictive diets such as keto diet on reducing febrile seizures can be evaluated by subjecting keto-fed mice and normal chow-fed mice to heat-induced seizure protocol. Similarly, epilepsy drug screening paradigms can be developed to test and identify candidate anti-epileptic drugs that ameliorate or suppress heat-induced seizures in drug-fed or treated mice when compared to vehicle-fed or control mice.
Disclosures
The authors declare no conflicts of interest.
Acknowledgements
We would like to thank Connor J. Smith for his help in building the customized mouse heat chamber. We acknowledge the help of O'Dowd lab members, Lisha Zeng and Andrew Salgado for standardizing the heating protocol during the early stages of the assay development. We also thank Danny Benavides and Kumar Perinbam for video recording parts of the experimental procedure for the manuscript. This work was supported by the NIH grant (NS083009) awarded to D.O.D.
Materials
| Name | Company | Catalog Number | Comments |
| Axial fan | Farnam | AF20-200-120-xx10-3.1 | Farnam custom products -Axial Fan Heater with Fan |
| Digital temperature controller | Inkbird | ITC-100RH | Inkbird digital PID temperature controller ITC-100RH with K thermocouple |
| Mouse rectal temperature probe | ThermoWorks, Braintree Scientific, Inc | RET-3 | Mouse rectal temperature probe with thermometer |
References
- Hirtz, D., et al. How common are the 'common' neurologic disorders. Neurology. 68, 326-337 (2007).
- Catterall, W. A. Sodium Channel Mutations and Epilepsy. Jasper's Basic Mechanisms of the Epilepsies. , Center for Biotechnology Information. US. Internet (2012).
- Mantegazza, M., Broccoli, V. SCN 1A /Na V 1.1 channelopathies: Mechanisms in expression systems, animal models, and human iPSC models. Epilepsia. 60, (2019).
- Stafstrom, C. E. Persistent Sodium Current and Its Role in Epilepsy. Epilepsy Currents. 7, 15-22 (2007).
- Schutte, S. S., Schutte, R. J., Barragan, E. V., O'Dowd, D. K. Model systems for studying cellular mechanisms of SCN1A-related epilepsy. Journal of Neurophysiology. 115, 1755-1766 (2016).
- Wei, F., et al. Ion Channel Genes and Epilepsy: Functional Alteration, Pathogenic Potential, and Mechanism of Epilepsy. Neuroscience Bulletin. 33, 455-477 (2017).
- Abou-Khalil, B., et al. Partial and generalized epilepsy with febrile seizures plus and a novel SCN1A mutation. Neurology. 57, 2265-2272 (2001).
- Zhang, Y. -H., et al. Genetic epilepsy with febrile seizures plus: Refining the spectrum. Neurology. 89, 1210-1219 (2017).
- Patterson, K. P., et al. Enduring memory impairments provoked by developmental febrile seizures are mediated by functional and structural effects of neuronal restrictive silencing factor. Journal of Neuroscience. 37, 3799-3812 (2017).
- Rossi, M. A. SCN1A and febrile seizures in mesial temporal epilepsy: An early signal to guide prognosis and treatment. Epilepsy Currents. 14, 189-190 (2014).
- Zhang, Y., et al. Altered gut microbiome composition in children with refractory epilepsy after ketogenic diet. Epilepsy Research. 145, 163-168 (2018).
- Meng, H., et al. The SCN1A mutation database: Updating information and analysis of the relationships among genotype, functional alteration, and phenotype. Human Mutation. 36, 573-580 (2015).
- Cheah, C. S., et al. Specific deletion of NaV1.1 sodium channels in inhibitory interneurons causes seizures and premature death in a mouse model of Dravet syndrome. Proceedings of the National Academy of Science U.S.A. 109, 14646-14651 (2012).
- Das, A., et al. Interneuron dysfunction in a new mouse model of SCN1A GEFS. eNeuro. , (2021).
- Kalume, F., et al. Sudden unexpected death in a mouse model of Dravet syndrome. Journal of Clinical Investigations. 123, 1798-1808 (2013).
- Martin, M. S., et al. Altered function of the SCN1A voltage-gated sodium channel leads to gamma-aminobutyric acid-ergic (GABAergic) interneuron abnormalities. Journal of Biological Chemistry. 285, 9823-9834 (2010).
- Rubinstein, M., et al. Dissecting the phenotypes of Dravet syndrome by gene deletion. Brain. 138, 2219-2233 (2015).
- Yu, F. H., et al. Reduced sodium current in GABAergic interneurons in a mouse model of severe myoclonic epilepsy in infancy. Nature Neuroscience. 9, 1142-1149 (2006).
- Dutton, S. B. B., et al. Early-life febrile seizures worsen adult phenotypes in Scn1a mutants. Experimental Neurology. 293, 159-171 (2017).
- Cheah, C. S., et al. Specific deletion of NaV1.1 sodium channels in inhibitory interneurons causes seizures and premature death in a mouse model of Dravet syndrome. Proceedings of the National Academy of Science U.S.A. 109, 14646-14651 (2012).
- Oakley, J. C., Cho, A. R., Cheah, C. S., Scheuer, T., Catterall, W. A. Synergistic GABA-enhancing therapy against seizures in a mouse model of Dravet Syndrome. Journal of Pharmacology and Experimental Therapeutics. 345, 215-224 (2013).
- Ricobaraza, A., et al. Epilepsy and neuropsychiatric comorbidities in mice carrying a recurrent Dravet syndrome SCN1A missense mutation. Scientific Reports. 9, (2019).
- Warner, T. A., Liu, Z., Macdonald, R. L., Kang, J. -Q. Heat induced temperature dysregulation and seizures in Dravet Syndrome/GEFS+ Gabrg2+/Q390X mice. Epilepsy Research. 134, 1-8 (2017).
- Eun, B. -L., Abraham, J., Mlsna, L., Kim, M. J., Koh, S. Lipopolysaccharide potentiates hyperthermia-induced seizures. Brain and Behavior. 5, 00348(2015).
- Miller, A. R., Hawkins, N. A., McCollom, C. E., Kearney, J. A. Mapping genetic modifiers of survival in a mouse model of Dravet syndrome. Genes Brain and Behavior. 13, 163-172 (2013).
- Mistry, A. M., et al. Strain- and age-dependent hippocampal neuron sodium currents correlate with epilepsy severity in Dravet syndrome mice. Neurobiology of Disease. 65, 1-11 (2014).
- Ogiwara, I., et al. Nav1.1 localizes to axons of parvalbumin-positive inhibitory interneurons: a circuit basis for epileptic seizures in mice carrying an Scn1a gene mutation. Journal of Neuroscience. 27, 5903-5914 (2007).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved