
Для просмотра этого контента требуется подписка на Jove Войдите в систему или начните бесплатную пробную версию.
Method Article
Графический пользовательский интерфейс для отслеживания концентрации белка в программном сопровождении в динамических клеточных выпячиваниях
В этой статье
Резюме
Мы представляем программное решение для полуавтоматического отслеживания относительной концентрации белка по длине динамических клеточных выступов.
Аннотация
Филоподии являются динамическими, пальцевидными клеточными выпячиваниями, связанными с миграцией и клеточными клеточными коммуникациями. Чтобы лучше понять сложные сигнальные механизмы, лежащие в основе филоподиального инициирования, удлинения и последующей стабилизации или ретракции, крайне важно определить пространственно-временную активность белка в этих динамических структурах. Чтобы проанализировать функцию белка в филоподии, мы недавно разработали полуавтоматический алгоритм отслеживания, который адаптируется к изменениям филоподиальной формы, что позволяет проводить параллельный анализ динамики выступов и относительной концентрации белка по всей филоподиальной длине. Здесь мы представляем подробный пошаговый протокол для оптимизации обработки ячеек, сбора изображений и анализа программного обеспечения. Мы также предоставляем инструкции по использованию дополнительных функций во время анализа изображений и представления данных, а также рекомендации по устранению неполадок для всех критических шагов на этом пути. Наконец, мы также включаем сравнение dПрограммное обеспечение для анализа изображений с другими программами, доступными для количественной оценки филоподий. Вместе представленный протокол обеспечивает основу для точного анализа динамики белка в филоподиальных выпячиваниях с использованием программного обеспечения для анализа изображений.
Введение
Пространственно-временное управление регуляторными белками актина ассоциируется с филоподиарной динамикой 1 , 2 . Таким образом, отслеживание пространственно разрешенной концентрации белка по всей филоподиальной длине во времени имеет решающее значение для продвижения нашего понимания механизмов, лежащих в основе инициации, удлинения, стабилизации или коллапса этих динамических структур 3 , 4 . В отличие от анализа белков в цитозоле, где многие изменения формы клетки происходят в большем масштабе, филоподии представляют собой динамические микроструктуры, которые постоянно пряжу 5 и изгибаются, тем самым исключая анализ с использованием простого подхода, такого как сканирование линии.
Доступны различные программные решения для отслеживания филоподиальной формы 6 , 7 , 8 , 9 . LikewIse, было разработано программное обеспечение для дозиметрического отслеживания динамики белка в теле клетки 10 , 11 . Чтобы объединить автоматическое отслеживание филоподиальной формы и пространственно-временного анализа белков, мы недавно разработали программное обеспечение для анализа изображений, основанное на алгоритме выпуклого корпуса 12 . Этот новый метод анализа, который работает через графический интерфейс пользователя (GUI), впервые объединяет относительную концентрацию белка по длине филоподии и скорости роста, что позволяет точно измерять пространственно-временное распределение белка независимо от движения этих Динамические структуры 12 .
Идея программного обеспечения (исходный код свободно доступен, см. Ниже) заключается в том, что одна из вершин выпуклой оболочки будет совпадать с кончиком филоподия ( рис. 1А ). Посмотрев в следующем кадре foR ближайшая вершина выпуклой оболочки, движущийся кончик можно отследить на протяжении всего фильма. Как только наконечник обнаружен в каждом кадре, его положение используется для рисования оси путем соединения кончика с опорной точкой у основания филоподия ( рис. 1В ). Наконец, используя эквидистантные узловые точки, позиции которых определяются медианным пикселем с максимальной интенсивностью вдоль линии, ортогональной оси, используются для определения основы, которая следует за филоподиальной формой. Используя эту адаптивную основу, создается кимограф для отслеживания филоподиального роста и концентрации белка для трех каналов вдоль филоподиальной длины ( рис. 1С ).

Рисунок 1: Принцип работы программного обеспечения для анализа изображений. ( A ) Алгоритм, лежащий в основеПрограммное обеспечение. На шаге 1 пользователь указывает ссылку (основание) и вершину (наконечник) филоподия. На шаге 2-1 основа филоподия получена с использованием медианного пикселя с максимальным значением интенсивности. На этапе 3 магистраль используется для профиля интенсивности пространственного белка. На шаге 2-2 программное обеспечение автоматически отслеживает кончик в следующем кадре. Вся процедура повторяется. ( B ) Снимок алгоритма с реальным филоподием, представляющий важные элементы, такие как выпуклый корпус, который используется для отслеживания. ( C ) Обзор параметров, которые можно измерить с помощью алгоритма. Эта цифра была изменена из ссылки 12 . Нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
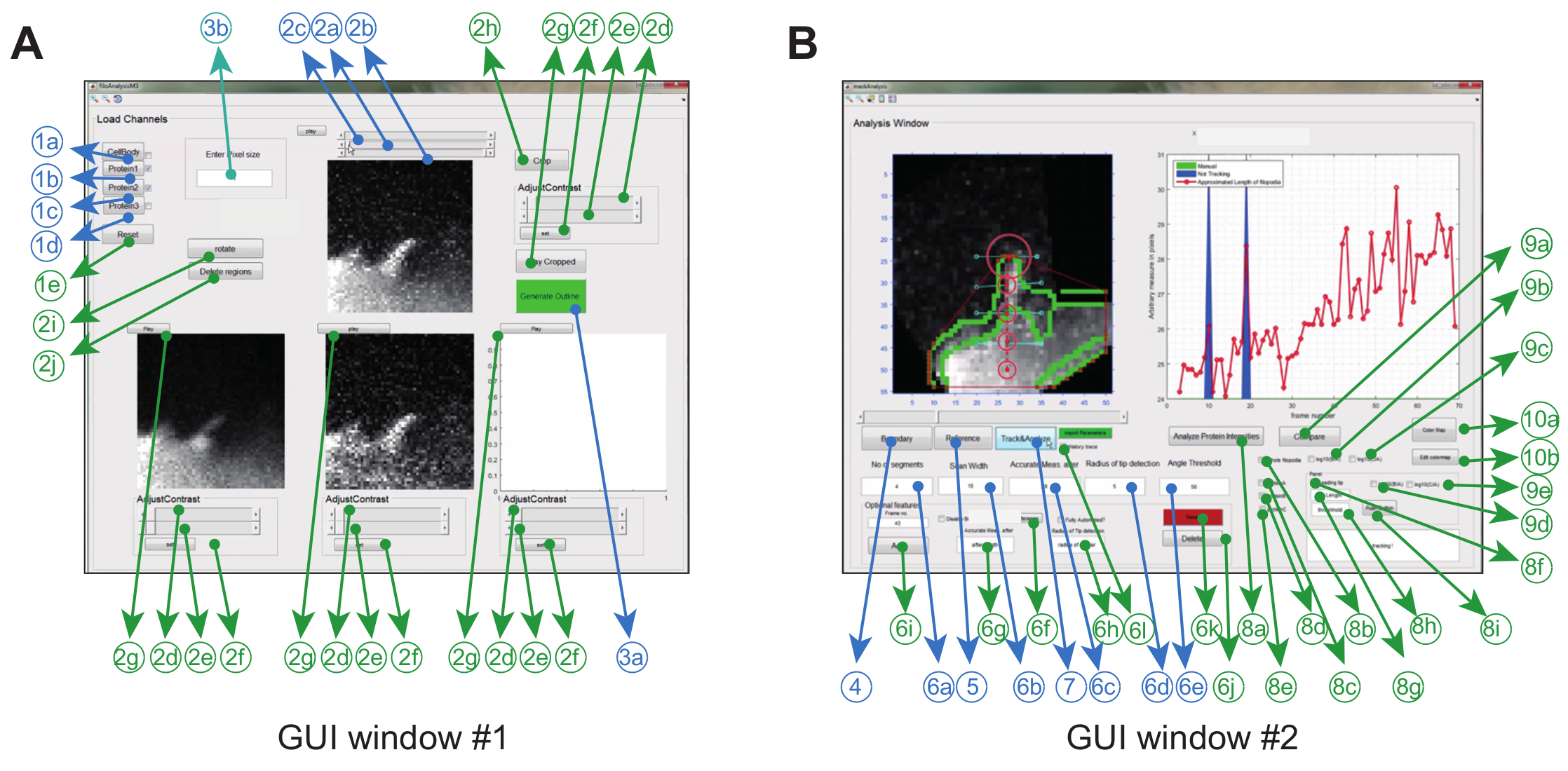
Программное обеспечение для анализа изображений работает в Matlab (называемом программным обеспечением) посредством графического использованияR. Чтобы максимизировать гибкость и надежность для конкретной экспериментальной установки, пользователь может настроить ряд параметров отслеживания ( например, разрешенный угол изгиба и межкадровое перемещение), а также внести некоторые поправки в фильмы ( например, обрезка, поворот, удаление нежелательных объектов) ( Рисунок 2A и Таблица 1) .
| графический интерфейс пользователя | Нет . | Обязательное | Описание | Имя (в графическом интерфейсе) | ||||
| # 1 | 1a | Y | Загрузка сложного файла .tiff, представляющего тело ячейки (с включенным полем) или создание наложенного тела ячейки из каналов | Тело клетки | ||||
| # 1 | 1б | Y | Загрузка файла с образцами изображений, соответствующего белку 1 | Протеин 1 | ||||
| # 1 | 1c | Y | Загрузка файла с образцами изображений, соответствующего белку 2 | Протеин 2 | ||||
| # 1 | 1d | Y | Загрузка файла с образцами изображений, соответствующего белку 3 | Протеин 3 | ||||
| # 1 | 1e | N | Сбрасывает все в предварительно загруженные файлы сложенными изображениями | Сброс | ||||
| # 1 | 2а | Y | Полоса прокрутки для определения начального кадра для анализа в окне графического интерфейса # 2 | Не Доступно | ||||
| # 1 | 2b | Y | Полоса прокрутки для определения окончательного кадра для анализа в окне графического интерфейса # 2 | Не Доступно | ||||
| # 1 | 2с | Y | Полоса прокрутки, представляющая curreКадр nt | Не Доступно | ||||
| # 1 | 2d | N | Серое значение пикселей, ниже которых все пиксели будут установлены на ноль | Не Доступно | ||||
| # 1 | 2e | N | Серое значение пикселей, над которым все пиксели будут установлены на максимальные значения | Не Доступно | ||||
| # 1 | 2f | N | Задайте значения интенсивности пикселей, указанных в <2e> & <2f> | Задавать | ||||
| # 1 | 2g | N | Воспроизведение фильма с регулировкой интенсивности | Играть | ||||
| # 1 | 2h | N | Обрезать изображение | урожай | ||||
| # 1 | 2i | N | Повернуть изображение | Поворот | ||||
| # 1 | 2j | N | Удаление областей во всем стеке | Удалить регионы | ||||
| # 1 | 3a | Y | Нажмите, чтобы открыть окно «Анализ» (окно № 2 графического интерфейса пользователя) | Окно отслеживания | ||||
| # 1 | 3b | Y | Введите размер пикселя в микрон | Введите размер пикселя | ||||
| # 2 | 4 | Y | Нажмите, чтобы сгенерировать изображение границы / края наложенного тела ячейки | граничный | ||||
| # 2 | 5 | Y | Нажмите, чтобы выбрать основание и конец филоподии | Referernce | ||||
| # 2 | 6а | Y | Введите количество сегментов или узлов | Нет сегментов | ||||
| # 2 | 6b | Y | Введите длину сканирования (перпендикулярно оси) | Ширина сканирования | ||||
| # 2 | 6с | Y | Введите длину, выше которой филоподия начинает изгиб | Точность измерения после | ||||
| # 2 | 6d | Y | Введите радиус круга обнаружения наконечника (т.е. область, где вершина может быть локализована в следующем кадре) | Радиус обнаружения наконечника | ||||
| # 2 | 6e | Y | Введите максимальный угол, из которого филоподия может изгибаться с вертикальной оси | Пороговое значение угла | ||||
| # 2 | 6f | N | Добавить опорные точки для основания и кончика для этого конкретного кадра | Выберите ссылку | ||||
| # 2 | 6g | N | Введите длину, выше которой филоподия начинает изгиб для этого конкретного кадра | Точность измерения после | ||||
| # 2 | 6h | N | Введите радиус круга обнаружения для этого конкретного кадра | Радиус обнаружения наконечника | ||||
| # 2 | 6i | N | После ввода всех параметров для конкретного кадра нажмите, чтобы сохранить значения в памяти и файл для дальнейшей справки | Добавить | ||||
| # 2 | 6j | N | Нажмите, чтобы удалить заданные вручную параметры для этого кадра. | Удалить | ||||
| # 2 | 6k | N | Нажмите, чтобы удалить все параметры, сохраненные вручную, используя «панель дополнительных функций» для всех фреймов | Сброс | ||||
| # 2 | 6l | N | Заходите перед отслеживанием, чтобы сохранить все результаты отслеживания в памяти для дальнейшего использования | История следа | ||||
| # 2 | 7 | Y | Нажмите, чтобы начать отслеживание | Отслеживание и анализ | ||||
| # 2 | 8а | N | Нажмите, чтобы начать отслеживать интенсивность белка | Анализ интенсивности протеина | ||||
| # 2 | 8б | N | Заходите, чтобы отслеживать интенсивность белкового канала вдоль филоподиальной длины | Целая филоподия | ||||
| # 2 | 8в | N | Заезд для отслеживания контрольного белка или белка А | ProteinA | ||||
| # 2 | 8d | N | Заезд для отслеживания белка B | ProteinB | ||||
| # 2 | 8е | N | Заезд для отслеживания белка C | ProteinC | ||||
| # 2 | 8е | N | Заходите, чтобы отслеживать среднюю интенсивность белка в thПодсказка | Ведущий наконечник | ||||
| # 2 | 8g | N | Введите длину наконечника | Длина кончика | ||||
| # 2 | 8h | N | Введите минимальное расстояние от основания, над которым начинается наконечник | порог | ||||
| # 2 | 8i | N | Нажмите, чтобы сохранить результаты анализа ведущих вершин в файле | Нажать кнопку | ||||
| # 2 | 9а | N | Нажмите, чтобы инициировать анализ с помощью ратиометрического белка | сравнить | ||||
| # 2 | 9b | N | Проверьте, чтобы сравнить белок B по отношению к A | log10 (В / А) | ||||
| # 2 | 9c | N | Проверьте, чтобы сравнить белок C по отношению к A | log10 (С / А) | ||||
| # 2 | 9г | N | Проверьте, чтобы сравнить белок B по отношению к A на кончике | log10 (В / А) | ||||
| # 2 | 9e | N | Проверьте, чтобы сравнить белок C по отношению к A на кончике | log10 (С / А) | ||||
| # 2 | 10a | N | Выберите другую цветовую карту (по умолчанию: Jetplot) | Цветная карта | ||||
| # 2 | 10b | N | Редактирование цветовой карты | Изменить цветную карту | ||||
Таблица 1: Обзор всех функций, присутствующих в графическом интерфейсе Windows # 1 и # 2.
Как только это будет достигнуто, программа создает выпуклый корпус и автоматически отслеживает подсказку во всем фильме. Параметры, извлеченные из фильма, такие как ратиометрический кимограф, скорость роста и филоподиальная длина aПовторно отображаться и также сохраняться в рабочей папке в виде изображений и файлов данных. Другие параметры, такие как время жизни, скорость роста и скорость отвода, могут быть затем извлечены и дополнительно проанализированы из хранимых файлов данных ( рисунок 2B ).

Рисунок 2: Графический пользовательский интерфейс для использования программного обеспечения для анализа изображений. ( A ) Окно № 1 графического интерфейса используется для загрузки и обработки изображений. Программа может загружать до 3-х белковых каналов, при этом 2 канала сравниваются по-разному. Окно поставляется с обязательными (синими) и дополнительными функциями (зеленым) для предварительной обработки изображений перед отслеживанием ( B ) Окно № 2 графического интерфейса используется для отслеживания филоподиума, а также пространственно-временного и количественного анализа белков. Опять же, дополнительные функции отмечены зеленым цветом. Эта цифра была измененаИз ссылки 12 . Нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
Здесь мы приводим подробный протокол для подготовки проб и обработки программного обеспечения. Мы начинаем с подробных инструкций по выращиванию клеток и приобретению фильмов, оптимизированных для анализа изображений. Этот раздел по сбору данных сопровождается подробным описанием работы программного обеспечения для анализа изображений. Во всем протоколе мы вводим критические шаги и дополнительные функции, которые следует учитывать при сборе и обработке данных. Наконец, мы анализируем филоподии из разных модельных систем с помощью программного обеспечения для анализа изображений, прежде чем закрывать с помощью сравнения описанного программного обеспечения для анализа изображений с другими программами, доступными для количественной количественной оценки и обсуждения ограничений и будущего направления.
протокол
1. Культура клеток
- Культурные клетки HeLa или COS в модифицированной среде Дульбекко Eagle Medium (DMEM), содержащей 4,5 г / л D-глюкозы, дипептид L-аланин-L-глутамина, пируват, 10% фетальную бычью сыворотку и 10 единиц / мл пенициллина / стрептомицина. Культурные нейроны в культуральных средах без L-глутамина, глутаминовой кислоты или аспарагиновой кислоты, дополненные 0,5 мМ дипептидом L-аланин-L-глутамина, добавками нейронов без сыворотки и 10 единиц / мл пенициллина / стрептомицина.
- После достижения 40% -ной слияния трансфектные клетки выбирают конструкцию с использованием трансфекционного реагента в соответствии с инструкциями производителя. Хранить трансфецированные клетки в инкубаторе при 37 ° C и 5% CO 2 в течение 15-18 часов.
- Чтобы уменьшить изменения pH и осмолярности (из-за испарения) во время получения изображения, заполните камеры клеточной культуры (до получения изображений) до 90% при 37 ° С предварительно нагретой среде, содержащей 20 мМ 4- (2-гидроксиэтил) -1 -пиперазинэтансульфоновой кислоты (HEPES). Чтобы запечататьКрышку, нанесите тонкий слой вакуумной смазки на внутреннюю часть крышки и осторожно нажмите на нее в камеру для культивирования, содержащую трансфицированные клетки.
2. Приобретение изображения
ПРИМЕЧАНИЕ. Длина филоподий колеблется от 2 до 10 мкм 13 . Филоподии растут со средней скоростью 0,05-0,1 мкм / с 13 , 14 .
- Приобретайте изображения с помощью микроскопа с объективом 60X или 100X и без пиксельного биннинга (здесь конфокальный микроскоп с вращающимся диском). Используйте скорости приема более 1 Гц (Гц) для отслеживания динамики филоподиона. Чтобы свести к минимуму артефакты вне фокуса, изображение филоподии близко к базальной мембране ( т.е. поверхности субстрата).
- Чтобы обеспечить плавное отслеживание, отрегулируйте время экспозиции камеры и интенсивность лазера так, чтобы отношение сигнал / шум (SNR) было больше 4. Избегайте насыщения отдельных каналов ( т.е. значения пикселей255 для 8-битных изображений и 65 535 для 16-разрядных изображений), так как это предотвратит последующий анализ изображения.
- Чтобы избежать просачивания, используйте только флуоресцентные метки, совместимые с лазерными линиями и фильтрами микроскопа (подробнее см. Ссылку 15 ).
3. Предварительная обработка изображения
ПРИМЕЧАНИЕ. Используйте ImageJ или другое доступное программное обеспечение для предварительной обработки изображений 16 , 17 .
- Если образец перемещается, исправьте боковой сдвиг с использованием доступного программного обеспечения ( например, https://github.com/NMSchneider/fixTranslation-Macro_for_ImageJ) перед анализом. Исключить фильмы с аксиальным дрейфом (т. Е. Движение в направлении z).
- Правильный фон ( т. Е. Серые значения областей за пределами ячейки), отбеливание ( т.е. непрерывная потеря при интенсивности флуоресценции из-за повреждения флуоресцентных белков) и возможного кровотока ( т. Е. Сигнал от одного гриппаВ обоих каналах) с использованием доступного программного обеспечения ( например, http://imagej.net/Category:Plugins и 16 , 17 ).
Примечание. Интенсивность флуоресценции отдельных каналов не изменяется программным обеспечением. - Чтобы обеспечить последующий анализ изображения, сохраните фильмы, соответствующие конкретным белковым каналам, в сером цвете, как «.tiff» в формате файла в рабочей папке.
ПРИМЕЧАНИЕ. Размеры ( т.е. размер, длина) стеков должны быть одинаковыми для всех каналов.
4. Анализ изображения - Шаг 1: Загрузка изображений
ПРИМЕЧАНИЕ. Программное обеспечение, описанное здесь, было написано в Matlab (называемом программным обеспечением) и будет работать только с этой программой.
- Загрузите ZIP-архив, содержащий все необходимые файлы для анализа изображений, со следующего сайта: https://campus.uni-muenster.de/en/einrichtungen/impb0/nanoscale-forces-in-cells/softwнаходятся/. Разархивируйте и скопируйте файлы в рабочую папку.
- После установки откройте программное обеспечение и запустите «filopodiaAnalysisM3.fig». Окно GUI 1 откроется, как показано на рисунке 2A .
- Загрузите сохраненные сложенные файлы .tiff, соответствующие конкретному интересующему белку в окне GUI 1, используя кнопки <1b> для белка A, <1c> для белка B, <1d> для белка C. См. Рисунок 2 и таблицу 1 для деталей.
ПРИМЕЧАНИЕ. Протеин А, показанный в окне графического интерфейса пользователя №1, выступает в качестве эталонного канала для окончательного рациометрического анализа. - Создайте наложенное изображение ячейки из белковых каналов, нажав <1a>.
- Нажмите <2a>, чтобы назначить первый кадр и <2b> для последнего кадра, используемого для анализа.
- Необязательно, обрезайте интересующую область (ROI), содержащую интересующий филоподиум, с помощью кнопки <2h>, поверните изображение uНажмите кнопку <2i> или удалите ненужные области, используя инструмент рисования с свободной рукой <2j>.
ПРИМЕЧАНИЕ. Для оптимизации анализа изображения рекомендуется изолировать ROI ( то есть обрезку, поворот, удаление) вместе с другими шагами предварительной обработки (вычитание фона, поправки на отбеливание и т. Д.) С помощью ImageJ. - Передвиньте ползунок <2c> для каждого кадра для контроля качества и проверьте, остается ли филоподий четко видным на протяжении всего фильма.
5. Анализ изображений. Шаг 2. Создание трассировки.
- Нажмите кнопку <3a> в окне GUI № 1, чтобы открыть окно GUI № 2 (см. Рисунок 2B ).
- Нажмите кнопку <4> в окне # GUI для генерации маски наложенного тела ячейки (сгенерированного в окне графического интерфейса пользователя 1 после нажатия <1a>). Программа также генерирует границу маски, где выпуклая оболочка реализована для получения точек вершин.
- КлиCk button <5>; Появится курсор. Используйте курсор, чтобы выбрать основание (от которого измеряется расстояние от филоподиального наконечника), а затем конец филоподии в кадре, где он впервые появляется. Для этого переместите ползунок в окне №2).
ПРИМЕЧАНИЕ. Чтобы свести к минимуму ошибки в выходе данных, поместите базовую точку вертикально ниже филоподиального наконечника вдоль оси. Позиционирование базовой точки в других местах ( например, сдвиг в поперечном направлении) может привести к смещению направления в значениях флуоресценции внутри тела ячейки. - Выберите пороговую длину (выше которой филоподии будут изгибаться) с помощью <6c>.
Примечание. Эта длина определяется как расстояние между базовой точкой (выбрано с помощью <5>) и границей тела ячейки ( т. Е. Области, от которой начинается филоподия). Расстояние измеряется в пикселях. - Укажите количество сегментов, используемых для аппроксимации формы филоподии в поле <6a>.
ПРИМЕЧАНИЕ.Минимальное количество сегментов зависит от максимальной длины, достигаемой филоподием, а также от того, как филоподиум изгибается. Не выбирайте число сегментов, большее, чем число пикселей между базой и кончиком ( т.е. пороговая длина). Выбор большего количества сегментов, чем пороговая длина, определенная на шаге 5.4. ( То есть количество пикселей между основанием и кончиком) приведет к переоценке длины филоподия. - Укажите ширину сканирования, которая действует как горизонтальный сканер для размещения узлов, в <6b>.
ПРИМЕЧАНИЕ. Эти узлы используются программой для соответствия линии, соединяющей основание и кончик с телом филоподиума ( т. Е. Для создания основы). В качестве отправной точки поставьте значение в пикселях, равное максимальной длине, умноженной на коэффициент ,
, - Укажите радиус сканирования (в пикселях) в поле <6d>. Используйте значение, которое приблизительно на 50% больше, чем obОбслуживаемое перемещение между кадрами.
ПРИМЕЧАНИЕ. Использование очень большого радиуса сканирования может захватывать нежелательные точки с выпуклым корпусом в кадрах, где нет реального филоподиального наконечника ( например , из движения плоскости или низкого SNR). - Укажите угол изгиба в поле <6e>.
ПРИМЕЧАНИЕ. Порог угла определяется максимальным углом, из которого филоподия изгибается во время всего анализа. Указание порога угла помогает программному обеспечению исключить отслеживание нежелательных структур, растущих со стороны филоподий, когда филоподии сгибаются к телу клетки. Программа работает надежно для филоподий с углами наклона менее 45 градусов от вертикальной оси. - Чтобы начать отслеживание, нажмите кнопку в окне графического интерфейса # 2. Нажмите «История трассировки» в окне графического интерфейса пользователя №2, чтобы сохранить весь протокол отслеживания для дальнейшего использования.
ПРИМЕЧАНИЕ. После завершения процедуры отслеживания длина филоподия в каждом кадре сохраняется вЛист с именем «length_vel» для «dynamic.xlsx» для будущих ссылок. Аналогично, все остальные параметры отслеживания сохраняются в листе с параметрами «dynamic.xlsx». - При желании, если филоподия не будет автоматически обнаружена во всех кадрах, выполните следующие действия, чтобы вручную ее исправить.
- Посетите соответствующий кадр с помощью ползунка в окне №2 и вручную выберите кончик филоподия.
Примечание. Фреймы, в которых не обнаружены точки выпуклой оболочки, будут представлены синей областью в окне отслеживания окна GUI Window 2. Обязательно проверять кнопку «История трассировки» до того, как была запущена программа отслеживания для доступа к узловым координатам в этом фрейме. - Выберите опорные точки (база, за которой следует подсказка) с помощью <6f> в окне графического интерфейса # 2. Укажите другие параметры, такие как «длина сканирования», «точное измерение после» и «максимальный угол изгиба»,; Для этого кадра, как описано в шагах 5.4-5.8.
- Нажмите кнопку <6i>, чтобы сохранить новые параметры (относящиеся к одному кадру). Этапы должны повторяться для всех кадров, обозначенных синей областью.
- По завершении повторите инициализацию процедуры отслеживания с помощью <7>.
ПРИМЕЧАНИЕ. Эта коррекция также может быть использована, если программа внезапно переключается с одного филоподиального на другой в фильме. В качестве альтернативы рассмотрим фильмы обрезки, содержащие несколько филоподий.
- Посетите соответствующий кадр с помощью ползунка в окне №2 и вручную выберите кончик филоподия.
6. Анализ изображения. Шаг 3: Анализ пространственно-временного белка.
- Для пространственно-временного анализа выберите поле, представленное <8b>, за которым следует интересующий белковый канал (<8c> и / или <8d> и / или <8e>).
Примечание. Обязательно выбрать белок A перед анализом (<9a>, <9b> и / или <9c>), поскольку в качестве эталона используется белок A. - Нажмите <8a>Инициировать отслеживание белка вдоль филоподиальной длины с использованием следа, полученного на этапе 5.9.
7. Анализ изображения. Этап 4. Анализ соотношения белков и метрических белков.
- Установите флажок <9b> или <9c> и щелкните по кнопке <9a>, чтобы получить пространственно-временный рациометрический график.
ПРИМЕЧАНИЕ. Чтобы избежать искажения относительной концентрации белка, ратиометрическое изображение не отображается как X / Y, а как log (X / Y). Для будущего использования графитированный график экспортируется в файлы формата «.png» и «.fig», а необработанные данные графика хранятся в файле «dynamic.xlsx».
8. Анализ изображения. Шаг 5: Анализ филоподиальных подсказок.
- Установите флажок <8f> и укажите длину наконечника и длину порога из базы с помощью <8g> и <8h>. Нажмите <кнопка>, чтобы сохранить данные в файл «dynamic.xlsx». Нажмите <Анализ интенсивности протеина>, чтобы сгенерироватьСлед интенсивности белков филоподиального наконечника и сохранение для рациометрического анализа.
ПРИМЕЧАНИЕ. Длина наконечника определяет количество пикселей на кончике, используемом для анализа. Программное обеспечение вернет среднее значение интенсивности пикселей на кончике над каждым фреймом. Пороговая длина определяет минимальное расстояние от основания до кончика, над которым начинает расти филоподий (филоподиальный наконечник) (и когда программа начинает отслеживать значение интенсивности наконечника). Поэтому длина наконечника должна быть меньше пороговой. - Нажмите на требуемое соотношение для анализа с помощью <9d> или <9e>.
- Нажмите кнопку сравнения <9a>, чтобы сгенерировать данные с данными.
Результаты
Используя клетки COS, трансфецированные маркером для нитевидного актина (f-тракта 18 , красный) и цитозольной ссылки (зеленый), мы обнаружили актино-богатые филоподиальные выступы ( рис. 3A , верхняя панель). Временные ряды показали, что филоподи...
Обсуждение
Здесь мы представляем подробный протокол для отслеживания динамики филоподиального роста и анализа относительных концентраций белка в этих динамических структурах с помощью алгоритма с выпуклым корпусом. Используя программное обеспечение, до трех каналов можно сравнивать поровну ?...
Раскрытие информации
Авторам нечего раскрывать.
Благодарности
Авторы признают финансирование от DFG (EXC-1003 до MG).
Материалы
| Name | Company | Catalog Number | Comments |
| DMEM | Life Technologies | 31966-021 | |
| 10% Fetal bovine serum | Biochrom AG | L11-044 | |
| Lipofectamine 2000 | Life Technologies | 11668-027 | |
| 1% penicillin/streptomycin | Biochrom AG | 12212 | |
| Neurobasal Medium | Life Technologies | 21103-049 | |
| B27 | Life Technologies | 17504-044 | |
| HEPES (1M stock solution) | Life Technologies | 15630 | |
| Citrine-N1 | Addgene | 54593 | |
| Labtech | Thermo | 155411 | |
| Glutamax-I | Thermo | 35050-061 | |
| Hela | Leibniz Institute DSMZ | ACC-57 | |
| COS 7 | Leibniz Institute DSMZ | ACC-60 | |
| 3T3 cells | Leibniz Institute DSMZ | ACC-59 | |
| Microscope | Nicon Eclipse | ||
| Camera | Andor | DU888 Ultra | |
| Confocal Unit | Yokagawa | CSU-X1 | |
| Pyruvate | Gibco | 31966-021 |
Ссылки
- Dunaevsky, A., Tashiro, A., Majewska, A., Mason, C., Yuste, R. Developmental regulation of spine motility in the mammalian central nervous system. Proc Natl Acad Sci U S A. 96 (23), 13438-13443 (1999).
- Matus, A., Brinkhaus, H., Wagner, U. Actin dynamics in dendritic spines: a form of regulated plasticity at excitatory synapses. Hippocampus. 10 (5), 555-560 (2000).
- Galic, M., et al. Dynamic recruitment of the curvature-sensitive protein ArhGAP44 to nanoscale membrane deformations limits exploratory filopodia initiation in neurons. Elife. 3, e03116 (2014).
- Hotulainen, P., et al. Defining mechanisms of actin polymerization and depolymerization during dendritic spine morphogenesis. J Cell Biol. 185 (2), 323-339 (2009).
- Leijnse, N., Oddershede, L. B., Bendix, P. M. Helical buckling of actin inside filopodia generates traction. Proc Natl Acad Sci U S A. 112 (1), 136-141 (2015).
- Tsygankov, D., et al. CellGeo: a computational platform for the analysis of shape changes in cells with complex geometries. J Cell Biol. 204 (3), 443-460 (2014).
- Xiong, Y., et al. Automated characterization of cell shape changes during amoeboid motility by skeletonization. BMC Syst Biol. 4, 33 (2010).
- Styner, M., Gerig, G., Lieberman, J., Jones, D., Weinberger, D. Statistical shape analysis of neuroanatomical structures based on medial models. Med Image Anal. 7 (3), 207-220 (2003).
- Blum, H., Wathen-Dunn, W. A transformation for extracting new descriptors of shape. Models for the Perception of Speech and Visual Form: Proceedings of a Symposium. , 362-380 (1967).
- Barry, D. J., Durkin, C. H., Abella, J. V., Way, M. Open source software for quantification of cell migration, protrusions, and fluorescence intensities. J Cell Biol. 209 (1), 163-180 (2015).
- Machacek, M., et al. Coordination of Rho GTPase activities during cell protrusion. Nature. 461 (7260), 99-103 (2009).
- Saha, T., et al. Automated analysis of filopodial length and spatially resolved protein concentration via adaptive shape tracking. Mol Biol Cell. 27 (22), 3616-3626 (2016).
- Argiro, V., Bunge, M. B., Johnson, M. I. A quantitative study of growth cone filopodial extension. J Neurosci Res. 13 (1-2), 149-162 (1985).
- Mogilner, A., Rubinstein, B. The physics of filopodial protrusion. Biophys J. 89 (2), 782-795 (2005).
- Shaner, N. C., Steinbach, P. A., Tsien, R. Y. A guide to choosing fluorescent proteins. Nat Methods. 2 (12), 905-909 (2005).
- Abramoff, M. D., Magalhaes, P. J., Ram, S. J. Image Processing with ImageJ. Biophotonics International. 11, 36-42 (2004).
- Courtney, J., Woods, E., Scholz, D., Hall, W. W., Gautier, V. W. MATtrack: A MATLAB-Based Quantitative Image Analysis Platform for Investigating Real-Time Photo-Converted Fluorescent Signals in Live Cells. PLoS One. 10 (10), e0140209 (2015).
- Schell, M. J., Erneux, C., Irvine, R. F. Inositol 1,4,5-trisphosphate 3-kinase A associates with F-actin and dendritic spines via its N terminus. J Biol Chem. 276 (40), 37537-37546 (2001).
- Korobova, F., Svitkina, T. Molecular architecture of synaptic actin cytoskeleton in hippocampal neurons reveals a mechanism of dendritic spine morphogenesis. Mol Biol Cell. 21 (1), 165-176 (2010).
- Cheadle, L., Biederer, T. The novel synaptogenic protein Farp1 links postsynaptic cytoskeletal dynamics and transsynaptic organization. J Cell Biol. 199 (6), 985-1001 (2012).
- Tarnok, K., et al. A new tool for the quantitative analysis of dendritic filopodial motility. Cytometry A. 87 (1), 89-96 (2015).
- Hendricusdottir, R., Bergmann, J. H. F-dynamics: automated quantification of dendrite filopodia dynamics in living neurons. J Neurosci Methods. 236, 148-156 (2014).
- Fanti, Z., Martinez-Perez, M. E., De-Miguel, F. F. NeuronGrowth, a software for automatic quantification of neurite and filopodial dynamics from time-lapse sequences of digital images. Dev Neurobiol. 71 (10), 870-881 (2011).
- Costantino, S., et al. Semi-automated quantification of filopodial dynamics. J Neurosci Methods. 171 (1), 165-173 (2008).
- Nilufar, S., Morrow, A. A., Lee, J. M., Perkins, T. J. FiloDetect: automatic detection of filopodia from fluorescence microscopy images. BMC Syst Biol. 7, 66 (2013).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены