
Bu içeriği görüntülemek için JoVE aboneliği gereklidir. Oturum açın veya ücretsiz deneme sürümünü başlatın.
Method Article
Tek molekül floresan ın Situ hibridizasyon (SM-balık) kullanmak için fare yumurta miktarını ve Localize mRNA
Bu Makalede
Özet
Tekrarlanarak mRNA'ların içinde bireysel yumurtalar, tek molekül RNA Floresans in situ olarak sayıları saymak için hibridizasyon (RNA-balık) yapışık olmayan hücreler için optimize edildi. Yumurtalar toplanmıştır, transkript belirli probları ile melezleşmiştir ve bir görüntü miktar yazılımı kullanarak sayılabilir.
Özet
Geçerli yöntemleri düzenli olarak yumurta ve embriyo mRNA ölçmek için kullanılan dijital ters transkripsiyon Polimeraz zincir reaksiyonu (dPCR), nicel, gerçek zamanlı RT-PCR (RT-qPCR) ve RNA sıralama içerir. Bu tekniklerin bir tek yumurta veya embriyo kullanılarak gerçekleştirildiğinde, düşük-kopya mRNA'ların güvenilir bir şekilde tespit edilmedi. Bu sorunu çözmek için yumurta veya embriyo birlikte analiz için havuza; Ancak, bu kez yüksek değişkenlik örnekleri arasında yol açar. Bu protokol için Floresans in situ hibridizasyon (balık) dallı DNA kimya kullanarak içinde nasıl kullanılacağını açıklar. Bu teknik mRNA'ların tek tek hücreleri içinde kayma desenini tanımlar. Teknik nokta bulma ve izleme bilgisayar yazılım ile birleştiğinde, mRNA'ların Hücre bolluğu da sayısal. Bu tekniği kullanarak, bir deney grubu içinde azaltılmış değişkenlik var ve daha az sayıda yumurta ve embriyo deneysel grupları arasında önemli farklılıklar tespit etmek gerekir. Piyasada bulunan dallı DNA SM-kesitli dokularda veya slaytlardaki yapışık hücreleri mRNA'ların algılamak için optimize balık kitleri. Ancak, yumurta etkili slaytlara uymayan ve bazı reaktif kitinde oosit lizis kaynaklanan çok sert. Bu lizis önlemek için birçok değişikliği balık seti için yapılmıştır. Özellikle, yumurta permeabilization ve yıkama arabellekleri yumurta ve embriyo ayirt için tasarlanmış özel arabellekleri yerine. 6-iyi levha permeabilization, yıkama ve incubations sonda ve amplifikatör ile gerçekleştirilen ve yumurta montaj medya iletişim kuralı'nı sonundaki slaytlarda yerleştirildi. Bu değişiklikler ticari olarak mevcut kit özellikle, oosit lizis sınırlamaları aşmak başardık. Doğru ve tekrarlanarak mRNA'ların tek tek yumurta içinde saymak için bilgisayar yazılımı kullanılmıştır. Birlikte, bu iletişim kuralını PCR ve sıralamanın Tek Kişilik hücrelerde belirli transkript ifadesi karşılaştırmak için bir alternatif temsil eder.
Giriş
Ters transkriptaz Polimeraz zincir tepkimesi (PCR) mRNA Nefelometri için altın standart olmuştur. İki deneyleri, dijital PCR (dPCR)1 ve nicel, real time PCR (qPCR)2 şu anda kullanılır. İki PCR teknikleri, dPCR qPCR tek hücrelerinde mRNA bereket ölçmek için kullanılabilir olduğu ima daha büyük duyarlılığa sahiptir. Ancak, bizim elimizde, deneysel her örnek başına 5-10 yumurta havuzlarda düşük bereket mRNA'ların analizini dPCR veri düşük tekrarlanabilirlik ve yüksek varyasyon3ile üretti. Bu RNA ayıklama ve ters transkripsiyon verimliliği ile ilgili deneysel hata nedeniyle muhtemeldir. RNA sıralama aynı zamanda tek bir fare ve insan yumurta4,5kullanılarak yapılmıştır. Bu teknik büyük olasılıkla bir deney grubu içinde değişkenlik artar Kütüphane üretimi için gerekli cDNA amplifikasyon adımları gerektirir. Ayrıca, düşük bereket transkript tespit olmayabilir. Her ne kadar sıralama fiyatlar son birkaç yıldır gitti, hala maliyet engelleyici Biyoinformatik analizi yüksek maliyet nedeniyle olabilir. Son olarak, mRNA yerelleştirme protein işlevi6için katkıda kayma değişikliklerle dinamik bir süreçtir. Bu nedenle, doğru ve tekrarlanabilir nicel ölçüler ve tek yumurta içinde bireysel mRNA'ların yerelleştirilmesini üretecektir bir teknik benimsemeye Vadisi'ni gezmeye gidiyoruz.
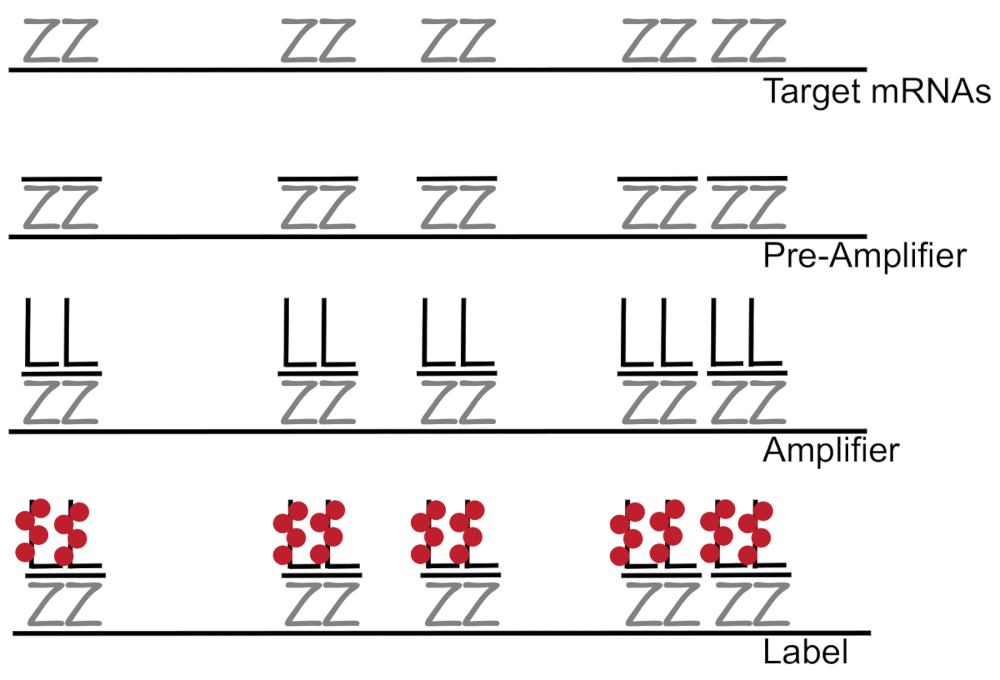
Dallı DNA Floresans in situ hibridizasyon için birleştiğinde yükselterek RNA/cDNA tek tek hücreler 7,8,9tek mRNA'ların etkinleştirme tespiti yerine Floresans sinyali güçlendirir. Tahlil hibridizasyon, amplifikasyon (dallı DNA'yı kullanarak) ve floresan sinyal7yükseltmek için adımları etiketleme floresan bir dizi aracılığıyla gerçekleştirilir. Teknik bir belirli mRNA3,8,10' a tamamlayıcı 18 - 25 Bankası Oligonükleotid sonda çiftlerinin bağlama ile başlar. On beş-yirmi sonda çiftleri her transkript sağlanması özgüllük hedef transkript için tasarlanmıştır. MRNA özgü hibridizasyon dallı bir yapılandırma oluşturan ön amfi ve amplifikatör probları tarafından takip edilir. Yaklaşık olarak, 400 etiket fluorophores 8000-fold artış bireysel mRNA (şekil 1)11olarak algılanmasını sağlayan Floresans sonuçlanan her amplifikatör bağlamak.

Şekil 1: SM-balık protokolünün şematik. Transkript belirli sonda, sıralı hibridizasyon dallı DNA amplifikatör ve mRNA gösterilir bir hedefe fluorophore. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.
{kind=link}
Önceki çalışmalarda in situ hibridizasyon (SM-balık) Lokalize β-aktin mRNA bireysel nöronlar12 ve servikal kanser HPV DNA molekülünü Floresans kullanarak satırları7hücre. Bilgisayar yazılımı nokta bulma ve izleme programı bireysel punctate floresan sinyal tanımlar ve başarıyla mRNA'ların her hücre3,13sayısını ölçmek için kullanılır.
SM-balık transkript düzeyleri fare yumurta ve embriyo düşük bereket mRNA'ların da dahil olmak üzere quantitate için yararlı bir araç olduğunu kanıtlayacak ki nöronlar12mRNA tespiti sonuçlarına dayanarak, biz onaylanmadığına karar. Ancak, teknik yapisan sabit hücreleri ile kullanım için optimize edilmiş ve parafin sabit formaldehit (FFPE) doku bölümleri gömülü. Bile Poly-L-lizin ile kaplı olan yumurtalar bir slayt için uygun değil. Ayrıca, onlar somatik hücre ve doku bölümleri ne zaman piyasada kitleri3özel arabellekleri bazıları tabi hücre lizis sonuçlanan daha kırılgan. Bu zorlukları aşmak için yumurta sabit ve el ile arabellekleri damla arasında transfer. Ayrıca, permeabilization ve yıkama arabellekleri kitleri hücre lizis azaltmak için değiştirildi. Önceden tasarlanmış probları balık seti ile birlikte satın alınan veya belirli transkript talep edilebilir. Her özel yoklama kümesi birinde üç Floresans kanal (C1, C2 ve C3) çoğullama için izin kullanılabilir. Geçerli deneyde, çift lekeli ve quantified C2 MicroRNAs sonda ve C3 Pou5f1 sonda kullanarak fare yumurta vardı. Bu sonda bildirilen ifadede yumurta ve embriyo MicroRNAs ve Pou5f1 göre seçildi. Hibridizasyon adımlar sonucunda, yumurta damla uygulama histolojik slaytlar için anti-fade montaj ortamının yerleştirildi. Confocal görüntüleri bireysel mRNA'ların temsil punctate floresan sinyallerini sayısını ölçmek için kullanılmıştır. MRNA'ların, görüntüleme Ayrıca belirli mRNA kayma dağılımını hücre içinde gösterdi. miktarının yanı sıra hangi diğer RNA miktar yöntemleri elde edemiyoruz. Bu teknik deneysel grupları3arasındaki önemli farkları belirlemek için her deney grubu içinde yumurta daha küçük sayıda kullanımına izin veren bir deney grubu içinde düşük değişkenlik var kanıtladı.
Protokol
Hayvan yordamları gözden geçirilmiş ve kurumsal hayvan bakım ve kullanım Komitesi Üniversitesi Nebraska-Lincoln adlı tarafından onaylanmış ve tüm yöntemleri ilgili kurallara ve düzenlemelere uygun şekilde gerçekleştirilmiştir. Bu çalışma için CD-1 outbred fareler vardı ad libitum erişim normal kemirgen chow ve su; Onlar 12: 12'karanlık muhafaza: ışık döngüsü.
1. gerekli ortam hazırlanması
- (OMM) temel medya için 100 mM NaCl, 5 mM KCl, 0,5 mM KH2PO4ve 1.7 mM CaCl2-2 H2O 100 mL steril su ekleyin.
Not: OMM orta için en fazla 1 ay depolanabilir. - 20 mM 3-morpholinopropane-1-sulfonic asit (paspas), 1.2 mM MgSO4-7 H2O, 0,5 mM glikoz, 6 mM L-laktat, 1 mM ala-gln, 0,1 mM taurin, 1 x esansiyel olmayan amino asitler (NEAA), 0.01 mM ethylenediaminetetraacetic asit ((OMOPS) tam ortamı için ekleyin EDTA), 10 µM Alfa lipoik asit, 10 µg/mL su katılmamış gentamisin, 21 mM 1 M NaOH, 5 mM NaHCO3, 0.2 mM Pyruvate, 0,5 mM sitrat, 1:10 4 mg/mL FAF BSA için toplam hacmi 100 mL steril su OMM seyreltme. Orta 0,22 µM filtre ile sterilize.
Not: OMOPS 1 hafta kadar saklanabilir. - Holding orta (HM) için % 5 fetal sığır serum OMOPS için ekleyin. Fare başına 2 mL HM olun.
- Hyaluronidase çözüm için 0.1 mg/mL sığır testis 1 mL HM elde hyaluronidase ekleyin.
- Fiksasyon arabelleği için % 4 paraformaldehyde 10 ml 1 x PBS ile birlikte % 0,1 embriyo sınıf polyvinylpyrrolidone (PVP)14birleştirin.
- 50 mL yıkama arabellek (WB) hazırlamak için % 0,1 iyonik olmayan yüzey aktif ve % 0,1 ekleyin 1 PBS14x PVP.
- Hazırlamak 10 mL permeabilization arabelleğe, 1 x PBS14' e %1 non - iyonik yüzey aktif ekleyin.
Not: yukarıda açıklanan yıkama ve permeabilization arabellekleri özel arabellekleri piyasada kitleri değiştirin.
2. ovulated yumurtalar dişi fareler gelen topluluğu
- Hazırlanışı:
- 5 IU insan Korionik gonadotropin (hCG) 44-48 h tarafından takip 5 IU at koryonik gonadotropin (EKG), mayi (IP) enjeksiyonla 5-8 haftalık dişi fareler teşvik daha sonra15,16.
- 35 mm Petri yemekler HM 2 mL 37 ° C ısınma plaka üzerinde içeren tutmak. Bir pipet, HM içeren 100 µL açılan üç, HM 50 µL damla olmadan 60 mm petri kabına hyaluronidase ardından hyaluronidase sulandırılmış. 37 ° C ısınma içeren Damlalar kullanılmadan önce plaka tabak koyun.
Not: Hyaluronidase damla sadece her ovidukt çifti diseksiyon önce buharlaşma ve konsantrasyon ile ya da ezelî hyaluronidase HM bileşenlerinin önlemek için yapılmalıdır.
- Fareler, hCG IP enjeksiyon sonra 16 h ötenazi, isoflurane aşırı doz kullanarak tarafından servikal çıkığı izledi.
- % 70 etanol kullanarak fare temiz. Karın boşluğu ortaya çıkarmak ve kadın üreme sistemi görselleştirmek. Yumurtalık Forseps ile tutun ve rahim bağ ve yumurtalık çevresinde aşırı yağ dokusu kaldırın. Rahim üzerinden ovidukt kesmek ve 35 mm tabak sıcak HM yumurtalık-ovidukt çifti yer.
- Yumurtalık ve çevresindeki herhangi bir yağ dokusu kaldırın. ½ inç 27'lik iğne kullanarak ovidukt şişmiş ampulla gözyaşı. Gözyaşı sitesinde ovidukt itmek ve kümülüs hücre-oosit kompleksleri (COCs) ihraç edilecektir. Metafaz II (MII) mayoz, ağzını pipet (Şekil 2) kullanarak hyaluronidase ile HM ortamı içeren 100 μL damlasına içinde olduğu kabul ovulated yumurta aktarın.

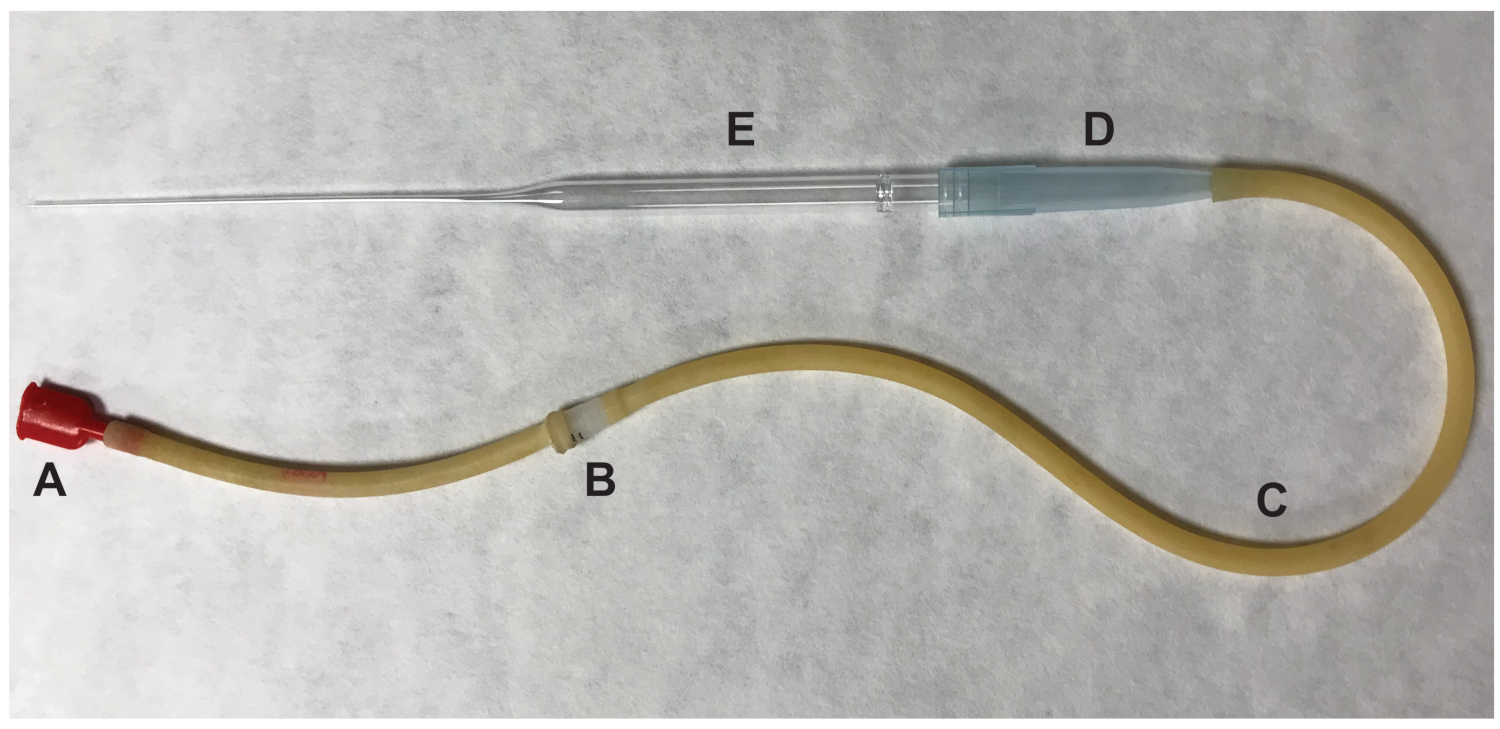
Resim 2 : Yumurta aktarmak için kullanılan ağız pipettor parçaları. (A) ağız parçası (B) 0.22 um, 4 mm filtre (C) aspiratör boru (D) 1000 μL damlalıklı ipucu (E) 9" Pasteur damlalıklı. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.
{kind=link}
- MII oosit-kümülüs hücre kompleksleri yukarı ve aşağı HM kümülüs hücreleri çıkarmak için ağzını pipet ile içeren hyaluronidase pipette. Her yumurta transferi, kümülüs hücreleri bir yıkama için yoksun olduklarını sonra içeren HM sadece ağzını pipet kullanarak bırakın. Her yıkama damlacık için bu işlemi yineleyin. Parçalanmış veya şeffaf yumurta15aktarılmaz.
Not: Yumurta ile 35 mm çanak olarak mümkün olduğunca küçük HM her düşüş aktarmak önemlidir. Bu her aktarma iletişim kuralı için geçerlidir. MII yumurta HM orta bir dakikadan daha içeren hyaluronidase kalmamalı.
3. SM-balık yumurta boyama
- Yumurta 500 µL fiksasyon arabelleği içeren bir 6-şey plaka bireysel bir kuyuda düzeltmek. 20 yumurta daldırın ya da daha az iyi. Oda sıcaklığında 20 dk için kuluçkaya.
Not: Her SM-balık boyama adım 6-şey konik bir tabak bireysel bir kuyuda meydana gelmektedir. Yumurtalar tamamen arabelleklerindeki batık ve değil üstünde tepe-in tampon kayan olduğundan emin olun. Her adım 20 yumurta ile gerçekleştirilen veya her şey daha az olmalıdır. - Sabit yumurta yıkama arabelleği (1.6 adımda açıklanan WB) 500 μL transfer için 10 dak. Tekrarlayın 2 kez daha.
- Yumurta oda sıcaklığında 30 dakika permeabilization arabellekte kuluçkaya.
Not: 1.7. adımda anlatılan permeabilization arabellek görgü permeabilization arabellek yerini alır.- Sonda kümelerini toplamak ve hızlı bir şekilde onları bir microcentrifuge spin aşağı. 10 dk 40 ° C su banyosu veya KULUÇKA için ayarla her sonda sıcak. Oda sıcaklığına kadar ne yapıyorsun?
Not: Bu adımı permeabilization kuluçka sırasında yapılmalıdır
- Sonda kümelerini toplamak ve hızlı bir şekilde onları bir microcentrifuge spin aşağı. 10 dk 40 ° C su banyosu veya KULUÇKA için ayarla her sonda sıcak. Oda sıcaklığına kadar ne yapıyorsun?
- Yumurtalar WB 500 µL oda sıcaklığında 10 dakika yıkayın.
- Yumurtalar proteaz III seyreltilmiş 1:8 1 X PBS, oda sıcaklığında 30 dakika içinde arabelleğin (--dan belgili tanımlık teçhizat elde edilebilir), 80 μL aktarın.
Not: 80 µL birim yeterince bireysel bir kuyunun 6-şey plaka kapsar. - Yumurtalar WB 500 µL oda sıcaklığında 10 dakika yıkayın.
- Isıtılan sonda kümeleri için MicroRNAs, Pou5f1 ve DapB (bir negatif kontrol gen) seyreltik, 1:50 de sonda seyreltici. 2 saat 40 ° C'de transkript özgü inceleyebilirsek 80 μL yumurtalar kuluçkaya
Not: Her özel yoklama kümesi içinde bir-in üç Floresans kanal (C1, C2 ve C3) kullanılabilir. MicroRNAs ve Pou5f1 sondalar C2 ve C3, sırasıyla etiketli. - Özel, amplifikatör 1 (AMP 1), amplifikatör 2 (AMP2), amplifikatör 3 (AMP3) ve amplifikatör 4-floresan (AMP 4-FL) oda sıcaklığında ısıtın.
Not: Bu adım 2 saatlik transkript özgü sonda kuluçka sırasında yapılmalıdır. - WB 500 μL için yumurta transferi ve oda sıcaklığında 10 dakika için kuluçkaya.
- Yumurtalar amplifikasyon arabelleklerindeki sırayla kuluçkaya.
- AMP1 80 µL oda sıcaklığında 10 dakika WB 40 ° C. Transfer yumurta 500 µL için 30 dk içinde yumurtalar kuluçkaya.
- AMP2 80 µL 40 ° C'de 15 dakika içinde yumurtalar kuluçkaya Yumurtalar WB 500 µL oda sıcaklığında 10 dakika aktarın.
- AMP3 80 µL 40 ° C'de 30 dk içinde yumurtalar kuluçkaya Yumurtalar WB 500 µL oda sıcaklığında 10 dakika aktarın.
Not: AMP-FL fluorophore içerdiği için protokol kalan karanlıkta gerçekleştirilir. Diseksiyon mikroskop altında çalışırken, ışık mümkün olduğu kadar azaltmak. - AMP4-FL 80 μL 40 ° C'de 15 dakika yumurta eklemek
Not: AMP4-FL alternatif olarak arabellek-A sağlanır (Alt-A), Alt-B veya Alt-C. AMP4-FL arabellek bağımlı üzerinde hangi emisyon dalga boyu isteniyorsa seçin.
- Yumurtalar WB 500 µL oda sıcaklığında 10 dakika yıkayın. DAPI 80 µL oda sıcaklığında 20 dakika içinde yumurtalar kuluçkaya. Yumurtalar WB 500 µL oda sıcaklığında 5 dakika yıkayın.
- Anti-solma montaj medya merkezi bir slaydın üzerine 12 µL pipet için reaktif kabarcıklar eklemeden. Yumurta ile mümkün olduğunca küçük WB olarak montaj ortam transfer ve bir coverslip uygulayın.
- Coverslip bir açıyla eğilme ve sıvı slayt üzerinde yavaş yavaş ve nazikçe yerleştirin. Coverslip yumurta ve kabarcıklar getirilmesi bozulmasını önlemek zor tuşuna basarak kaçının.
- Slaytları koyu kutu için Kuru gecede oda sıcaklığında saklayın. Coverslip imzalamaya açık oje slaytları kenarlarını kat.
- Standart bir mikroskop ile kalıcı bir kalem yumurta slayt ve daire bulmak için kullanın.
Not: Bu adım gerekli değildir ancak Slayt üzerindeki yerini yumurta geliştirir. En iyi sonuçlar için floresan sinyal olarak 1-5 gün içinde Resim Slayt solmaya başlar.
4. görüntü işleme
- Z adım confocal mikroskobu kullanarak 3 boyutlu yumurta görüntü.
Not: görüntüleri doğru bir şekilde analiz etmek, her z adım 1.0 µm/dilim olmalıdır. - Confocal görüntüleri sıkıştırılmış bir nd2 olarak veya tek tek kaydedin. TIFF dosyaları her yumurta için. Her iki görüntü türü ile açık kaynak görüntü işleme programı, Fiji uyumludur.
- Karşıdan yükleyip açık erişim Fiji yazılım (https://imagej.net/Fiji/Downloads).
- ND2 dosyaları Fiji sürükleyin ve hyperstackseçin. Confocal görüntü olarak kaydedilmişse. TIFF dosyaları 4.4 adıma atlayın.
Not: Nd2 dosyasının Fiji kesildiğinde hyperstack açılan otomatik olarak görüntülenmesi gerekir. - Görüntü sekmesini tıklatın, rengiseçin ve nd2 dosyasının floresan kanalları ayırmak için Split kanalları ' yi tıklatın.
- Bireysel oluşturmak. TIFF dosyaları için oosit floresan her kanaldaki her z-dilim. Görüntü sekmesini tıklatın, yığınlarseçin ve yığın görüntüleri içintıklayın. Görüntü sekmesini tıklatın, türünüseçin ve her z dilim bireysel RGB renk görüntüyü dönüştürmek için RGB Renk'i tıklatın.
Not: RGB renk yapay ve seçilmiş olarak her emisyon dalga boyu için istenen olmak. - Dönüştürülen her resmi kaydet. TIFF dosyası. Floresan her kanal için tek bir yumurta görüntülerden dikiş (adımı 4.3) karışıklığı önlemek için yeni bir klasöre yerleştirin.
- ND2 dosyaları Fiji sürükleyin ve hyperstackseçin. Confocal görüntü olarak kaydedilmişse. TIFF dosyaları 4.4 adıma atlayın.
- Her normalleştirmek. TIFF görüntü Pou5f1 ve negatif kontrol görüntüleri (DapB) kullanarak MicroRNAs için.
Not: Normalleştirme bir fotoğraf düzenleme programı kullanılarak gerçekleştirilir. Arka plan Floresans aynı düzeyde her denetim yansımadan kaldırmak emin olun. - Açık her normalleştirilmiş. TIFF dosyası olarak Fiji tüm z-dilimler her dalga boyu her yumurta için birleştirmek için.
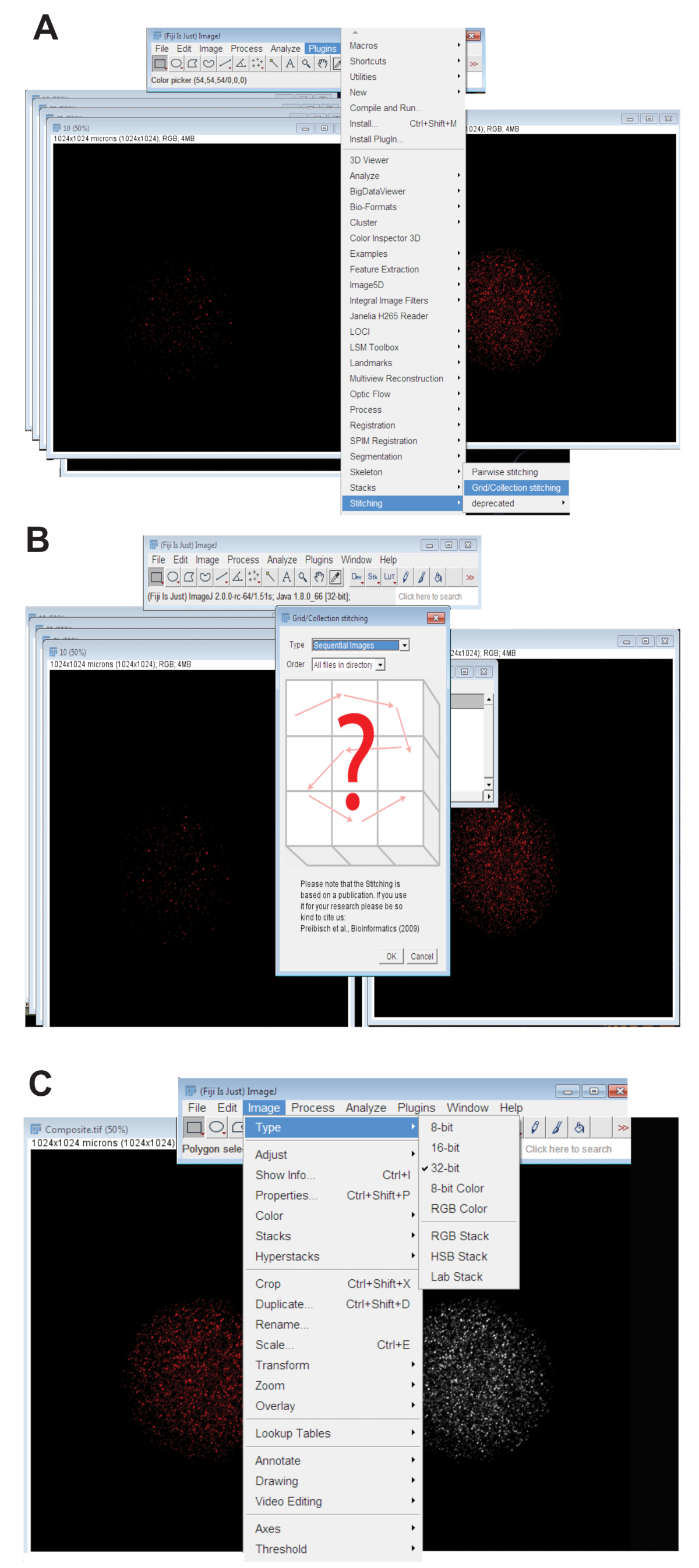
- Eklentiler sekmesini tıklatın, dikişseçin ve Kılavuz/koleksiyon (şekil 3A)'ı tıklatın. Sıralı görüntüleri damla-aşağı yemek listesi ve Tamam'ı tıklatın (şekil 3B) seçin.
- Dizine göz atın ve bir dalga boyu, bireysel bir yumurta için z dilimli görüntüleri içeren klasörü seçin (bkz. Adım 4.3.4). Tamam' ı tıklatın.
- Uygun renk kanalı için kullanılan dalga boyu dikişli görüntü altındaki kaydırıcıyı taşıyın ve son RGB dikişli görüntü görüntü tipi, seçme, tıklatarak oluşturun ve RGB rengitıklatın.
Not: Bu görüntü adım 4,6 açıklanan Floresans Nefelometri için kullanılacaktır.
- Dikişli görüntü 32-bit en fazla öngörülen resme dönüştürmek. Tıklayın görüntü türünüseçin ve 32-bit (şekil 3 c) tıklatın. Bu resmi bir yeni kaydedin. TIFF dosyası.

Şekil 3 : Birlikte yumurta confocal z-serisi görüntülerini dikiş. (A) eklentisi kılavuz/toplama aracı oosit bileşik görüntüler üretmek için kullanılan Fiji'de gösterilen ekran görüntüsü. (B) arasında sıralı Floresans örtüşme sıralı görüntüleri kullanır. TIFF dosyaları bir kompozit görüntü oluşturmak için. (C) bileşik görüntü bir 32-bit olarak kaydedildi. TIFF dosyası. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.
{kind=link}
- Karşıdan yükleyip Dr Larson, bir araştırmacı ulusal kurumları, sağlık Ulusal Kanser Enstitüsü (https://ccr.cancer.gov/Laboratory-of-Receptor-Biology-and-Gene-Expression/daniel-r-larson) için Web üzerinden kullanılabilir nokta bulma ve izleme programı13. Download ve install açık erişim sanal makine için etkileşimli verileri dil (IDL) iş sistem hangi nokta bulma ve izleme programı (http://www.spacewx.com/pdf/idlvm.pdf) çalıştırmak için gerekli.
- Adım 4.6 (şekil 4A), nokta bulma ve izleme programı içinde oluşturulan 32-bit, dikişli görüntüyü açın. Localize açılan seçin ve hangi görüntünün içinde bulunan sayı noktalar hesaplar Localize (şekil 4B),'ı tıklatın.
Not: bireysel bir mRNA sayılan her spot temsil eder. Grup geçişi ve foton eşik ayarları ekran (şekil 4B) gösterilmiştir. Bu iletişim kuralı için her eşik ayarı için varsayılan değer kullanıldı. Temsilcisi olumlu ve arka plan noktalar (şekil 4 c) gösterilir.

Şekil 4 : Yer bulucu kullanarak mRNA'ların miktar ve izleme. (A) bireysel z-serisi görüntüleri şekil 3 ' te açıklandığı gibi birlikte dikişli ve öngörülen bir 32-bit en yüksek sayı olarak kaydedilir. TIFF dosyası. (B) birçok parçalardan oluşan imge yer bulucu ve izleme açıldı. Yerelleştirmek floresan noktalar (kırmızı kutu) saymak için kullanıldı. Grup geçişi ve foton eşik mavi kutu tarafından belirtilir. (C) (eşiğin) olumlu bir sinyal mavi ok işaret eder. Beyaz ok bir floresan eşiğin altına nokta gösterir ve bu nedenle, sayılmaz. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.
{kind=link}
Sonuçlar
İletişim kuralı tamamlandıktan sonra confocal z-serisi (şekil 4A ve şekil 5), dikişli görüntüleri (şekil 4 c), tek tek görüntüleri sonuç olacaktır ve mRNA sayar (şekil 4B). Çoğullama gerçekleştirildiğinde, Ayrıca birleştirilmiş görüntüleri iki farklı mRNA'ların (...
Tartışmalar
Protokol sırasında küçük adımlar bir dizi başarılı Floresans ve mRNA'ların doğru sayar sağlayacaktır. İlk olarak, protokol koleksiyonu ve yumurta fiksasyonu hemen sonra gerçekleştirilmelidir. PVP yumurta birbirine yapışmasını önlemek için %4 paraformaldehyde fiksaj arabelleğin eklenir unutmayın. Biz deneme koleksiyonu ve yumurta fiksasyonu hemen sonra gerçekleştirmek gerekli bulundu. Herhangi bir gecikme transkript undercounting neden olacak bir çok daha düşük Floresans sinyal sonuçlanır. ...
Açıklamalar
Yazarlar ilan yok
Teşekkürler
Dr Daniel R. Larson yükleme ve 13 nokta bulma ve izleme programı kullanımı ve Nebraska Üniversitesi Lincoln mikroskobu çekirdek confocal mikroskobu görüntüleme için teknik destek ile cömert onun yardım için teşekkür. Bu çalışmada bir katkı Nebraska tarımsal araştırma bölümü, Lincoln, Nebraska Üniversitesi'nin temsil eder ve UNL Hatch fonlar (NF'leri-26-206/katılım numarası-232435 ve NF'leri-26-231/katılım numarası-1013511) tarafından desteklenmiştir.
Malzemeler
| Name | Company | Catalog Number | Comments |
| (±)-α-Lipoic acid | Sigma-Aldrich | T1395 | Alpha Lipoic Acid |
| Albumin, Bovine Serum, Low Fatty Acid | MP Biomedicals, LLC | 199899 | FAF BSA |
| BD 10mL TB Syringe | Becton, Dickinson and Company | 309659 | 10 mL syringe |
| BD PrecisionGlide Needle | Becton, Dickinson and Company | 305109 | 27 1/2 gauge needle |
| Calcium chloride dihydrate | Sigma-Aldrich | C7902 | CaCl2-2H2O |
| Citric acid | Sigma-Aldrich | C2404 | Citrate |
| D-(+)-Glucose | Sigma-Aldrich | G6152 | Glucose |
| Disodium phosphate | Na2HPO4 | ||
| Easy Grip Petri Dish | Falcon Corning | 351008 | 35 mm dish |
| Edetate Disodium | Avantor | 8994-01 | EDTA |
| Extra Fine Bonn Scissors | Fine Science Tools | 14084-08 | Straight, Sharp/Sharp, non-serrated, 13mm cutting edge scissors |
| Fetal Bovine Serum | Atlanta biologicals | S10250 | FBS |
| Gentamicin Reagent Solution | gibco | 15710-064 | Gentamicin |
| GlutaMAX-I (100X) | gibco | 35050-061 | Glutamax |
| Gold Seal Micro Slides | Gold Seal | 3039 | 25 x 75mm slides |
| Gonadotropin, From Pregnant Mares' Serum | Sigma | G4877 | eCG |
| hCG recombinant | NHPP | AFP8456A | hCG |
| Hyaluronidase, Type IV-S: From Bovine Testes | Sigma-Aldrich | H3884 | Hyaluronidase |
| Jewelers Style Forceps | Integra | 17-305X | Forceps 4-3/8", Style 5F, Straight, Micro Fine Jaw |
| L-(+)-Lactic Acid, free acid | MP Biomedicals, LLC | 190228 | L-Lactate |
| Magnesium sulfate heptahydrate | Sigma-Aldrich | M2773 | MgSO4-7H2O |
| MEM Nonessential Amino Acids | Corning | 25-025-Cl | NEAA |
| Microscope Cover Glass | Fisher Scientific | 12-542-C | 25 x 25x 0.15 mm cover slips |
| Mm-Nanog-O2-C2 RNAscope Probe | Advanced Cell Diagnostics | 501891-C2 | Nanog Probe |
| Mm-Pou5f1-O1-C3 RNAscope Probe | Advanced Cell Diagnostics | 501611-C3 | Pou5f1 Probe |
| MOPS | Sigma-Aldrich | M3183 | |
| Paraformaldehyde | Sigma-Aldrich | P6148 | Paraformaldehyde |
| PES 0.22 um Membrane -sterile | Millex-GP | SLGP033RS | 0.22 um filters |
| Polyvinylpyrrolidone | Sigma-Aldrich | P0930 | PVP |
| Potassium chloride | Sigma-Aldrich | 60128 | KCl |
| Potassium phosphate monobasic | Sigma-Aldrich | 60218 | KH2PO4 |
| Prolong Gold antifade reagent | invitrogen | P36934 | Antifade reagent without DAPI |
| RNAscope DAPI | Advanced Cell Diagnostics | 320858 | DAPI |
| RNAscope FL AMP 1 | Advanced Cell Diagnostics | 320852 | Amplifier 1 |
| RNAscope FL AMP 2 | Advanced Cell Diagnostics | 320853 | Amplifier 2 |
| RNAscope FL AMP 3 | Advanced Cell Diagnostics | 320854 | Amplifier 3 |
| RNAscope FL AMP 4 ALT A | Advanced Cell Diagnostics | 320855 | Amplifier 4 ALT A |
| RNAscope FL AMP 4 ALT B | Advanced Cell Diagnostics | 320856 | Amplifier 4 ALT B |
| RNAscope FL AMP 4 ALT C | Advanced Cell Diagnostics | 320857 | Amplifier 4 ALT C |
| RNAscope Fluorescent Multiplex Detection Reagents Kit | Advanced Cell Diagnostics | 320851 | FISH Reagent Kit |
| RNAscope Probe 3-plex Negative Control Probe | Advanced Cell Diagnostics | 320871 | Negative Control |
| RNAscope Probe 3-plex Positive Control | Advanced Cell Diagnostics | 320881 | Positive Control |
| RNAscope Probe Diluent | Advanced Cell Diagnostics | 300041 | Probe Diluent |
| RNAscope Protease III | Advanced Cell Diagnositics | 322337 | Protease III |
| RNAscope Protease III & IV Reagent Kit | Advanced Cell Diagnostics | 322340 | FISH Protease Kit |
| RNAscope Protease IV | Advanced Cell Diagnostics | 322336 | Protease IV |
| S/S Needle with Luer Hub 30G | Component Supply Co. | NE-301PL-50 | blunt 30 gauge needle |
| Sodium bicarbonate | Sigma-Aldrich | S6297 | NaHCO3 |
| Sodium chloride | Sigma-Aldrich | S6191 | NaCl |
| Sodium hydroxide | Sigma-Aldrich | 306576 | NaOH |
| Sodium pyruvate, >= 99% | Sigma-Aldrich | P5280 | Pyruvate |
| Solution 6 Well Dish | Agtechinc | D18 | 6 well dish |
| Taurine | Sigma-Aldrich | T8691 | Taurine |
| Tissue Culture Dish | Falcon Corning | 353002 | 60 mm dish |
| Triton X-100 | Sigma-Aldrich | X100 | Triton X-100 |
Referanslar
- Vogelstein, B., Kinzler, K. W. Digital PCR. Proceedings of the National Academy of Sciences. 96 (16), 9236-9241 (1999).
- MacK, E. M., Smith, J. E., Kurz, S. G., Wood, J. R. CAMP-dependent regulation of ovulatory response genes is amplified by IGF1 due to synergistic effects on Akt phosphorylation and NF-kB transcription factors. Reproduction. 144 (5), 595-602 (2012).
- Xie, F., Timme, K. A., Wood, J. R. Using Single Molecule mRNA Fluorescent in Situ Hybridization (RNA-FISH) to Quantify mRNAs in Individual Murine Oocytes and Embryos. Scientific Reports. 8 (1), 7930 (2018).
- Ruebel, M. L., et al. Obesity modulates inflammation and lipidmetabolism oocyte gene expression: A single-cell transcriptome perspective. Journal of Clinical Endocrinology and Metabolism. 102 (6), 2029-2038 (2017).
- Borensztein, M., Syx, L., Servant, N., Heard, E. . Mouse Oocyte Development. 1818, 51-65 (2018).
- Jansova, D., Tetkova, A., Koncicka, M., Kubelka, M., Susor, A. Localization of RNA and translation in the mammalian oocyte and embryo. PLoS ONE. 13 (3), 1-25 (2018).
- Player, A. N., Shen, L. P., Kenny, D., Antao, V. P., Kolberg, J. A. Single-copy gene detection using branched DNA (bDNA) in situ hybridization. Journal of Histochemistry and Cytochemistry. 49 (5), 603-611 (2001).
- Wang, F., et al. RNAscope: A novel in situ RNA analysis platform for formalin-fixed, paraffin-embedded tissues. Journal of Molecular Diagnostics. 14 (1), 22-29 (2012).
- Itzkovitz, S., van Oudenaarden, A. Validating transcripts with probes and imaging technology. Nature Methods. 8 (4), S12-S19 (2011).
- Derti, A., et al. ProbeDesigner: for the design of probesets for branched DNA (bDNA) signal amplification assays. Bioinformatics. 15 (5), 348-355 (1999).
- Larson, B., Malayter, D., Shure, M. Multiplexed Detection of Cytokine Cancer Biomarkers using Fluorescence RNA In Situ Hybridization and Cellular Imaging. BioTek Application Notes. , 1-5 (2016).
- Buxbaum, A. R., Wu, B., Singer, R. H. Single β-Actin mRNA Detection in Neurons Reveals a Mechanism for Regulating Its Translatability. Science. 343 (6169), 419-422 (2014).
- Thompson, R. E., Larson, D. R., Webb, W. W. Precise nanometer localization analysis for individual fluorescent probes. Biophysical Journal. 82 (5), 2775-2783 (2002).
- Herrick, J. R., Paik, T., Strauss, K. J., Schoolcraft, W. B., Krisher, R. L. Building a better mouse embryo assay: effects of mouse strain and in vitro maturation on sensitivity to contaminants of the culture environment. Journal of Assisted Reproduction and Genetics. 33 (2), 237-245 (2016).
- Pohlmeier, W. E., Xie, F., Kurz, S. G., Lu, N., Wood, J. R. Progressive obesity alters the steroidogenic response to ovulatory stimulation and increases the abundance of mRNAs stored in the ovulated oocyte. Molecular Reproduction and Development. 81 (8), 735-747 (2014).
- Xie, F., Anderson, C. L., Timme, K. R., Kurz, S. G., Fernando, S. C., Wood, J. R. Obesity-dependent increases in oocyte mRNAs are associated with increases in proinflammatory signaling and gut microbial abundance of lachnospiraceae in female mice. Endocrinology. 157 (4), 1630-1643 (2016).
- Hirao, Y., Yanagimachi, R. Detrimental effect of visible light on meiosis of mammalian eggs in vitro. Journal of Experimental Zoology. , (1978).
- Takenaka, M., Horiuchi, T., Yanagimachi, R. Effects of light on development of mammalian zygotes. Proceedings of the National Academy of Sciences. 104 (36), 14293-14293 (2007).
- Komminoth, P., Werner, M. Target and signal amplification: Approaches to increase the sensitivity of in situ hybridization. Histochemistry and Cell Biology. 108 (4-5), 325-333 (1997).
- Hornick, J. E., Duncan, F. E., Shea, L. D., Woodruff, T. K. Multiple follicle culture supports primary follicle growth through paracrine-acting signals. Reproduction. 145 (1), 19-32 (2013).
- Vlasova-St. Louis, I., Bohjanen, P. Feedback Regulation of Kinase Signaling Pathways by AREs and GREs. Cells. 5 (1), 4 (2016).
- Gilbert, C., Svejstrup, J. Q. RNA Immunoprecipitation for Determining RNA-Protein Associations In Vivo. Current Protocols in Molecular Biology. 75 (1), 27.4.1-27.4.11 (2006).
- Kwon, S., Chin, K., Nederlof, M., Gray, J. W. Quantitative, in situ analysis of mRNAs and proteins with subcellular resolution. Scientific Reports. 7 (1), 16459 (2017).
- Voigt, F., et al. Single-Molecule Quantification of Translation-Dependent Association of mRNAs with the Endoplasmic Reticulum. Cell Reports. 21 (13), 3740-3753 (2017).
- Halstead, J. M., Wilbertz, J. H., Wippich, F., Lionnet, T., Ephrussi, A., Chao, J. A. TRICK: A Single-Molecule Method for Imaging the First Round of Translation in Living Cells and Animals. Methods in Enzymology. 572, (2016).
- Cookson, W., Liang, L., Abecasis, G., Moffatt, M., Lathrop, M. Mapping complex disease traits with global gene expression. Nature Reviews Genetics. 10 (3), 184-194 (2009).
- Houle, D., Govindaraju, D. R., Omholt, S. Phenomics: the next challenge. Nature Reviews Genetics. 11, 855 (2010).
- Freimer, N., Sabatti, C. The Human Phenome Project. Nature Genetics. 34, 15 (2003).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiThis article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır