
Method Article
Damlacık Tabanlı RNA Sıralamaile Yüksek EldeLi Maya Zorlanma Fenotilik
Bu Makalede
Özet
Mikrobiyal mühendisliğin 'tasarım-yap-test' döngüsündeki bir darboğaz, suşların işlevsel ekranlarını gerçekleştirebildiğimiz hızdır. Damlacık bazlı RNA dizilimi kullanan deney başına yüzlerce ila binlerce maya hücresine uygulanan gerinim taraması için yüksek iş letme yöntemini tanımlıyoruz.
Özet
Maya genomlarını dilemek için kullanılan güçlü araçlar bu mikrobu mühendislik için değerli bir platform haline getirmiştir. Artık genetik olarak farklı milyonlarca suştan kütüphaneler inşa etmek mümkün olsa da, istenilen fenotip için tarama önemli bir engel olmaya devam etmektedir. Mevcut tarama teknikleri ile, bilgi çıktısı ve üretim arasında bir denge vardır ve yüksek verim taraması genellikle tek bir ilgi ürünü üzerinde gerçekleştirilir. Bu nedenle, genetik olarak tasarlanmış maya suşlarının izojenik pikolitre kolonilerine tek hücreli RNA dizilimini uyarlayarak gerilme taramasını hızlandırmak için bir yaklaşım savuruyoruz. Maya hücreleri üzerinde RNA dizilimi yapmanın benzersiz zorluklarını gidermek için, RNA dizilimi gerçekleştirmeden önce hidrojeller ve spheroplast içindeki izojenik maya kolonilerini kültürlendiriyoruz. RNA sıralama verileri maya fenotipleri çıkarmak ve mühendislik yolları sıralamak için kullanılabilir. Yöntemimizin ölçeklenebilirliği mikrobiyal mühendislikte kritik bir engeli gidermeözelliğidir.
Giriş
Mikrobiyal mühendisliğin temel amacı, mikropları değerli bileşikler 11,2üretmek için onları ikna etmek için değiştirmektir. S. cerevisiae, kültür kolaylığı ve genomu3,4,,5mühendislik için mevcut araçların genişliği nedeniyle mikrobiyal mühendislik için birincil organizma olmuştur. Ancak, modifiye maya üzerinde fonksiyonel ekranlar performans bir engel kalır: tarama iş kısmı büyüklük siparişleri ile genom mühendisliği arkasında geri kalıyor. Tarama genellikle mikrowell plakaları suşları izole içerir ve belirli bir bileşik üretimini ölçerek onları fenotilik6,7. Bu sürecin iş bilgili yüz mikrolitre reaksiyonlar bireysel suşları atamak için gerekli reaktif büyük miktarda sınırlıdır. Damlacık mikroakışkannormalde iyi plakalar8yapılan küçülme reaksiyonları tarafından büyüklük siparişleri ile maya tarama hacmini artırmak için cazip bir çözüm sağlar 8 . Ancak, iyi plaka ekranlar ile olduğu gibi, damlacık ekranlar genellikle tek ürün bileşikleri tespit, hangi mühendislik yolu9,,10,,11küresel fonksiyonu içine sınırlı bilgi sağlar.
RNA dizilemesi (RNA-seq), ilgili tüm genlerin ekspresyon düzeylerinin aynı anda değerlendirilmesine izin vererek yol çalışmasının daha kapsamlı bir karakterizasyonuna olanak sağlayabilir12,13. Ayrıca, damlacık yöntemleri, mühendislik varyantları14,15ekran kitaplıkları için gerekli iş sahibi sağlayarak, deney başına binlerce hücre profilli izin verir. Ancak, RNA-seq yöntemleri memeli hücreleri için optimize edilmiştir; maya, karşılaştırma, hücre başına daha az mRNA ve kaldırmak zor bir hücre duvarıvar, mevcut yöntemlerle sıralama engelleyen. Maya RNA-seq'yi etkinleştirmek için yüksek iş letiyetli bir damlacık yöntemi geliştirilebilseydi, maya mühendisliği için ölçeklenebilir, uygun maliyetli ve bilgi açısından zengin bir fenotileme platformu sağlar.
Biz yüksek iş letiyet mikroakışkanlar17kullanarak maya hücreleri sıralamak için son zamanlarda geliştirilen yöntem ayrıntılı bir protokol sıyoruz. Sınırlı RNA'nın zorluğunun üstesinden gelmek için, tek maya hücrelerini pikolitre hidrojel kürelerde kapsüller ve kültür. Kültür hücreleri kopyalar, aynı mühendislik yolu paylaşan yüzlerce kopya verir; bu, tek hücreli gen ekspresyonuna bağlı değişimi azaltırken, sıralama için kullanılabilir RNA miktarını önemli ölçüde artırır. Kültür tabanlı amplifikasyon sonra, biz toplu enzimatik sindirim yoluyla hücre duvarı kaldırarak, hücreleri spheroplast. Hücre zarları bozulmadan kalır, böylece her izojenik koloni ve ilişkili mRNA hidrojel kürelerinde kapsüllenmiş kalır. Bu bize mRNA yakalama reaktifler ve lysis tampon ve mRNA yakalanan, barkodlu ve Drop-Seq iş akışı14aşağıdaki sıralanır ile tek tek kolonileri eşleştirmek için izin verir. Yöntemimiz deney başına binlerce izojenik maya kolonisinin transkripsiyon çapında taranmasına olanak sağlar.
Protokol
1. Mikroakışkan cihaz imalatı
- SU-8 ana imalatı
- Bilgisayar destekli tasarım yazılımı kullanarak Cihaz A ve B(Ek Dosya 1 ve 2)için mikroakışkan kanallar için negatif maske tasarlayın ve en az 10 μm çözünürlüğe sahip devre kartı filmine yazdırın.
- Bir spin coater üzerine temiz bir 75 mm silikon gofret yerleştirin ve merkezine yaklaşık 1 mL SU-8 dökün. Gofret'i fırlatmak için vakumu açın.
- Cihaz A için, spin kat SU-8 2150 30 s için 500 rpm, 2.750 rpm 30 s izledi. Cihaz B için, spin-coat SU-8 2100 30 s için 500 rpm, 2.500 rpm 30 s izledi. Bu, su-8 kat kalınlığı 200 μm ve 120 m, sırasıyla verecektir.
- Spin-coater gelen gofret çıkarın ve yumuşak fırında 60 dakika için 95 °C'de bir ocak üzerine yerleştirin.
- Hotplate gelen gofret çıkarın ve oda sıcaklığına soğumaya bırakın. Maskeyi gofretin üzerine yerleştirin ve 2 dakika boyunca 190 mW, 365 nm UV LED'in altına yerleştirin.
- Pozlama sonrası pişirme için 5 dakika için 95 °C'de bir ocak seti üzerine gofret yerleştirin.
- Gofret çıkarın ve oda sıcaklığına soğumasını bekleyin. 20 dakika propilen glikol monometil eter asetat (PGMEA) bir banyo da gofret yerleştirin.
- PgMEA ile gofret durular izopropanol takip. Bu işlem sırasında herhangi bir opak kalıntı görünürse, PGMEA ve izopropanol ile durulama tekrarlayın. Hava gofret kuru.
- Gofret 95 °C'de bir ocak üzerine 3 dakika yerleştirin.
- Gofret'i 90 mm çapındaki Petri kabına çıkarın ve yerleştirin.
- SU-8 ustasında polidimethylsiloxane (PDMS) döküm
- Silikon tabanın 10:1 kütle oranını kürleme maddesine karıştırın. Degas PDMS yaklaşık 30 dakika karıştırıldıktan sonra.
- Gofretin üzerine en az 5 mm kalınlığında bir tabaka oluşana kadar SU-8 anasının üzerine gazdan arındırılmış PDMS dökün.
- Yaklaşık 30 dakika için gofret üstünde PDMS Degas.
- PDMS'yi tedavi etmek için gofreti en az 80 dakika boyunca 65 °C'lik bir fırına yerleştirin.
- Gofretten tedavi edilmiş PDMS levhasını kesin.
- PDMS levhasını mikroakışkan özelliklere ve 0,75 mm biyopsi zımbasıyla giriş ve çıkış deliklerine yerleştirin.
- 50 mm x 75 mm cam kaydırağı isopropanol ile temizleyin ve PDMS levhasının mikroakışkan özelliklerinden bantla tüm tozları çıkarın.
- Temizlenmiş cam kaydırağı ve mikroakışkan özelliklere sahip PDMS levhasını 1 dakika boyunca 100 Pa (1 mbar O2) plazmaya kadar yüzle tartın.
- Yapıştırma sağlamak için PDMS levhasını cam kaydıraktan aşağıya yüzüstü özellikleriyle yerleştirin. Yapıştırmayı tamamlamak için slaydı en az 30 dakika boyunca 65 °C'lik bir fırına yerleştirin.
- Florlu yüzey işleme sıvısı ile kızartarak tüm mikroakışkan kanalları tedavi edin. Cihazı 65 °C'lik fırında en az 10 dakika pişirin ve sıvıyı buharlaştırın.
2. A Cihazı kullanılarak hidrojellerde maya kapsüllemi
- Bir süspansiyon kültürü büyüyen maya alın ve bir hemositometre güvenin.
- Fosfat tamponlu salin (PBS) hücreleri yaklaşık 750 k/mL konsantrasyona geri alın. Bu hidrojellerin% 30 onları bir maya hücresi olacak sağlar. Maya hücrelerinin sadece yaklaşık yarısı koloniler halinde büyür, maya kolonileri içeren hidrojellerin ~% 15 yol.
- Ultralow erime noktası agarose pbs% 2 w / v ve ısı 90 °C eriyene kadar karıştırın. Bu ~ 10 dakika sürer.
- Agarose karışımını 0,22 μm'lik bir filtre yle şırınganın içine yükleyin.
- %2 w/v iyonik florosuraktif18 ile florlu yağ içeren bir şırınga ve maya süspansiyonu içeren bir şırınga yükleyin.
- Bölüm 1'de yapılan coflow drop splitter cihazını alın ve şırıngalardan gelen tüpü cihaza bağlayın. Damla toplama için bir buz kovasında 15 mL konik tüp içine çıkış boru kılavuzu.
- Aşağıdaki akış hızları ile cihaza üç çözüm akışı:
- 3 mL/h akış hızı ile maya süspansiyon akışı.
- Agarose karışımını 3 mL/h akış hızında nakışlayın.
- Florlu yağı 15 mL/h akış hızında nakışlayın.
- Yaklaşık 1 mL emülsiyon toplayın. Agarose tam olarak ayarlamak için izin vermek için ek bir 5 dk bekleyin.
3. Kültür için jel kırma ve yıkama
- Emülsiyona florlu yağda %20 perflorooctanol (PFO) eşit hacim ekleyin. Karıştırma için izin vermek için konik tüp birkaç kez ters.
- 2 dk için 2.000 x g kırık emülsiyon aşağı spin. Jeller yağ ve PFO aşamaları üzerinde pelet olacaktır.
- Jelleri yeniden askıya almak için yağ fazını çıkarın ve 2 mL TE-TW tampon (10 mM Tris pH = 8.0, 1 mM EDTA, %0.01 Ara-20) ekleyin. Süspansiyonu yeni bir 15 mL konik tüpe aktarın.
- Adım 3.2 gibi jelleri aşağı pelet ve iki yıkama toplam TE-TW bir kez daha yıkayın.
- 2 mL'lik ortamda süpernatant ve resuspend jelleri çıkarın. 5 mL'lik bir kültür tüpüne aktarın.
- Bir gecede 30 °C'de titreyerek kuluçkaya yatabilirsiniz.
NOT: Gece kuluçkadan sonra maya hidrojelleri 4 °C'de birkaç gün tutulabilir.
4. Maya kolonisi lysis
- Jelleri 2 dk için 2000 x g'da 15 mL konik tüpe ve pelet hidrojellerine aktarın.
- Hidrojelleri PBS 2x'te yıkayın.
- 1x sferoplasting tampon 1x yıkayın.
- Sferoplasting tamponda sferoplasting enzimin 2-50x seyreltilmesini gerçekleştirin ve hidrojellere 1 mL ekleyin.
- 37 °C'de 1 saat kuluçkaya yatırın. Tedavi edilen maya daha şeffaf görünecektir (Şekil 3A).
- En alttaki 0,8 mL hidrojel süspansiyonu alın ve 1 mL kapaksız şırıngaya aktarın.
- Şırıngayı 3D baskılı şırınga tutucuya yerleştirin(Ek dosya 3)ve 2 dk için 2.000 x g'de döndürün. Bu, hidrojellerin şırınga nın başındaki paketi kapatmasına neden olur.
5. Cihaz B kullanarak lysed maya kolonilerinden mRNA yakalama
- 240.000 Drop-Seq boncuk alın ve 15 mL konik tüp içine aktarın.
- Pelet Drop-Seq boncuklar 1 dk için 1.000 x g aşağı iplik tarafından.
- 120.000 boncuk/mL boncuk süspansiyon konsantrasyonu için 500 mM sodyum klorür ile 0.9x maya lysis tampon 2 mL supernatant ve resuspend boncuk çıkarın.
- Boncuk süspansiyonunu, bir karıştırma çubuğu takılı 3 mL şırıngaya aktarın.
- Florlu yağda %2 w/v perfloropolyether-polietilen glikol (PFPE-PEG) sürfaktan içeren bir şırınga hazırlayın.
- Yakın paketlenmiş hidrojeller içeren şırınganın tüm sulu kafasını boşaltın ve şırıngayı kapatın.
- Hidrojel, boncuk süspansiyonu ve yağ şırıngalarını şırınga pompalarına takın ve boru ile bölüm 1'de yapılan kapsülleme cihazına bağlanın.
- Çıkış borularından buz üzerinde 50 mL konik bir tüpe bağlanın.
- Aşağıdaki akış hızları ile cihaza üç çözüm akışı:
- Hidrojelleri 0,4 mL/h'de akıtın.
- Boncuk süspansiyonu 0,4 mL/h'de akar.
- Florlu yağı 1,6 mL/h'de akış.
- ~1 mL emülsiyon toplayın veya artık hidrojel kalmayana kadar cihazı çalıştırın.
6. cDNA üretimi, kütüphane hazırlama ve sıralama
- Drop-Seq protokolü14'tebelirtildiği gibi toplanan emülsiyona 6x SSC tamponunun 30 mL'sini ve 1 mL'sini PFO ekleyin.
- Boncuklar üzerinde yakalanan mRNA'dan cDNA oluşturmak, kitaplık hazırlama sıralamak ve veri analizini sıralamak için Drop-Seq protokolünü izlemeye devam edin.
Sonuçlar
Daha önce yayınlanmış Drop-Seq işakışını izojenik koloni dizilemesi (ICO-seq) için izojenik maya kolonilerinin gen ekspresyonu profillemesini gerçekleştirmek için uyarladık. Biz tek maya hücreleri izole ve agarose mikrojeller içine kapsüllenmiş (Şekil 1A). Mikrojellerin bir gecede kuluçkaya yatmasını takiben, bu kapsüllü maya hücreleri izojenik kolonilere dönüştü. Jelleri mRNA yakalaması için ikinci bir mikroakışkan cihaza yüklemeden önce, maya hücre duvarını sindirdik ve mRNA'yı daha erişilebilir hale getirmek için(Şekil 1B, solda). Bu mikrojelleri kapattık ve mRNA yakalama boncuklarını ve lysis tamponu birleştirdik. Bazı damlacıklar tam olarak bir boncuk bir lysed maya kolonisi ile eşleştirilmiş içeriyordu. Emülsiyondaki tüm boncuklar toplandı ve Drop-Seq protokolüne göre cDNA sentezlendi ve sıralandı.
Sekiz damla ayırıcı bağlı bir kokapsülasyon mikroakışkan cihazı kullanarak agarose mikrojelleri içinde tek maya hücre kapsülleme yoluyla izojenik maya kolonileri oluşturduk (Şekil 2A). Biz ~ 750.000/ mL bir konsantrasyon a girdi maya süspansiyon seyreltilmiş böylece ~ 30% mikrojeller onları tam olarak bir maya var. Cihaza ultradüşük erime sıcaklığı agarose eklemeden önce, biz yüksek bir sıcaklıkta eritilmiş ve erken jelleşme önlemek için bu sıcaklıkta şırınga muhafaza. Damla-nesil kavşakta(Şekil 2B),maya hücreleri başlangıçta 160 μm damlacıkiçine kapsüllenmiş edildi. Damla-nesil kavşaktaki sekiz kat bölünmüş lük bu damlacıkları sekiz adet 80 μm damlacıkolarak ikiye böldü(Şekil 2C). Erimiş agarose'a, damla bölme sırasında 37 μm'ye kadar dar olan kanallar içinde tıkanıklıkoluşmasını önlemek için bir şırınga filtresi bağlandı. Biz hemen agarose jelleşme işlemi başladı buz, emülsiyon toplandı. Tipik bir emülsiyonun polidispersitini ~%6(Ek Şekil 1)olarak hesapladık, ancak %10'a varan polidispersit değerleri kabul edilebilir. Bir kez agarose jeller ayarlamak, biz emülsiyon kırdı ve yağ fazı kaldırıldı. Jeller büyüme ortamına daldırma önce sulu tampon yıkandı. Mikrojellerin bir gecede kuluçkaya yatması, bazı mikrojellerde izojenik kolonilerin büyümesine neden olur(Şekil 2D). En az 20 hücreden oluşan kolonileri içeren hidrojellerin yüzdesi kuluçka süresi ve ortam kompozisyonu da dahil olmak üzere kültür koşullarına bağlıydı. C. albicanskullanarak yaptığımız gösteride, hidrojellerin yaklaşık %15'inin 20 saat süspansiyon kültüründen sonra bir koloni içerdiğini belirledik.
İkinci bir koenkapsülasyon cihazı mRNA'yı İzojenik kolonilerden çıkarttı (Şekil 3A). Maya mikrogellerini mikroakışkan cihaza yüklemeden önce, jelleri maya hücresi duvarlarını sindirmek için bir çözeltiye batırdık. Maya hücrelerinin doğru sindirimi mikroskopi ile doğrulandı, daha yansıtıcı morfolojisi olan tedavi edilen maya ile(Şekil 3B). Mikrojelleri bir şırıngayla kapattık ve jel giriş akış hızını her damlada bir jel olacak şekilde ayarladık. MRNA bir akış damla yapma kavşak(Şekil 3C)önce yakın paketlenmiş jel akışı ile karışık lysis tampon boncuk lar yakalamak . 160 μm'lik damlacıklardan oluşan bir emülsiyon topladık ve koloniler hücresel içeriklerini ortaya çıkmaya başladılar. Birden fazla boncuk içeren damla sayısını en aza indirmek için sınırlı bir seyreltme boncuk yükledik, ancak damla yapımı sırasında jellerin yakın ambalajı, lysed kolonisi olan bir boncuk içeren toplanan damlaların yaklaşık %10'unun toplanmasıyla sonuçlandı(Şekil 3D).
ICO-seq iş akışını kullanarak insan bağırsak mikrobiyomlarında bulunan bir maya türü olan C. albicans'ıngen ekspresyonunu analiz ettik. C. albicans iki farklı hücre durumları arasında geçiş yeteneği için dikkat edilir, beyaz ve opak olarak adlandıran19. Biz bir mühendislik C albicans suşu kullanın, rZY122, WH11 geninin bir kopyasını yerine, YFP ile beyaz hücrelerde sadece aktif20. İş akışını kullanarak bir dizi gen ekspresyonu profili elde ettik ve bunları en az 300 benzersiz geni ifade eden kolonilerin analizinde kullandık. Referans veri seti olarak, daha önce yayınlanmış bir çalışma17'den elde edilen C. Albicans ifade verilerini kullandık ve 600'den az benzersiz geni ifade eden kolonileri filtreledik. Ana bileşen (PC) analizi ve t-stochastic komşu katıştırma (tSNE) boyutsallık azaltma21yaptıktan sonra, örnek veri setimiz ile referans arasında genel uyum bulduk (Şekil 4A). PC analizi YFP ve WH11 önemli ölçüde ilk iki PC'nin katkıda bulundu. Ayrıca, tSNE analizinde üç küme saptandı(Şekil 4B). Küme 2 ağırlıklı olarak örnek veri kümesinden hücrelerden oluşurken, 0 ve 1 kümeleri her iki örnekteki hücrelerden oluşuyordu. WH11 ifadesini tSNE 'de(Şekil 4C, üst panel) üst üste biçerek, küme 1'in büyük olasılıkla beyaz koloniler içerdiğini belirledik. Ayrıca STF2 ekspresyonunun küme 1'de(Şekil 4C, alt panel) arttığını ve daha önce elde edilen verilerle tutarlı olduğunu tespit ettik17. 0 ve 2 kümelerinde WH11 ve STF2 küme 1(Şekil 4D)ile karşılaştırıldığında önemli ölçüde azaldı. Fermantasyonda yer alan ADH1gibi genler, opak hücrelerin önceki çalışmaları ile tutarlı, küme 0 upregulated edildi22. Küme 2'deki kolonilerin 0 ve 1 kümelerinde bulunan kolonilere göre ribozomal RNA'da azaldığını bulduk. Örnek ve referans veri setleri aynı hücre stoğu kullanılarak elde edilse de, bu sonuç deneysel işlemedeki ince farklılıkların bile gen ekspresyonunu etkileyebileceğini göstermektedir.

Şekil 1: ICO-seq iş akışına genel bakış. (A) Bir süspansiyon kültüründe büyüyen maya tampon seyreltilmiş ve tek maya hücreleri ile agarose mikrojellerin Poisson yükleme sağlamak için bir akış odaklama damlacık jeneratör cihaz erimiş agarose ile coenkapsüle edildi. Jeller agarose soğuduzaman ayarlayın, yağ / su süspansiyon bozuldu, ve yağ kaldırıldı, suda jel boncuk bir süspansiyon verim. Bir gecede kültürden sonra, maya hücreleri mikrojeller içinde izojenik koloniler haline geldi. (B) Koloniler bir hücre duvarı bozunma tamponuna maruz kaldılar, daha sonra ikinci bir mikroakışkan cihazda mRNA yakalama boncukları ile yakın paketlendiler ve coenkapsüle edildiler. Mikrojellerin yakın ambalaj her damla bir jel vardı sağladı, boncuk Poisson yükleme bir damla içinde birden fazla boncuk şansını azaltırken. Toplanan damlalar cDNA sentezi ve sıralama kütüphanesinin üretimi için işlendi. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
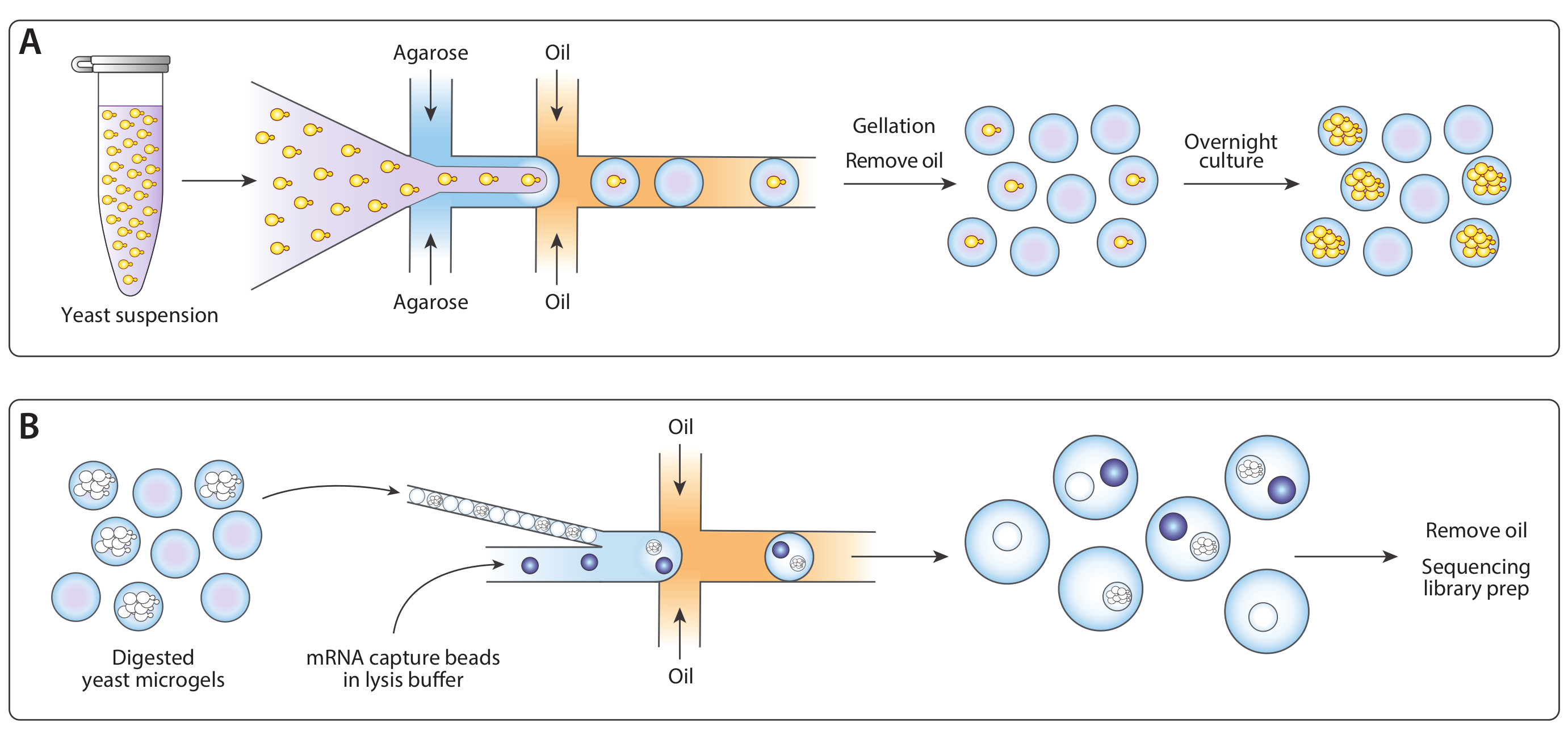{kind=link}

Şekil 2: A cihazı kullanılarak agarose mikrojelleri içinde iyojenik maya kolonilerinin üretimi. (A) Üç giriş ve çıkış portunun konumlarını gösteren mikroakışkan cihazın şeması. Bırakma bağlantı noktası kırmızı ile vurgulanır. (B) Normal cihaz çalışması sırasında açılan bağlantının yakın çekim. (C) Toplanan damlacıkların mikrografı, kapsüllü bir hücre (inset) içeren bir damlacık yakın çekim ile. (D) Agarose mikrojitlerinde 24 saatlik kuluçka sonrası izojenik maya kolonilerinin mikrografı, iki koloninin yakın çekimi (inset). Tüm ölçek çubukları = 100 μm. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 3: Cihaz B kullanarak iyojenik kolonilerden lysis ve mRNA yakalama. (A) Üç giriş ve çıkış portunun konumlarını gösteren mikroakışkan cihazın şeması. Bırakma bağlantı noktası kırmızı ile vurgulanır. (B) Hücre duvarı sindirimini takip eden maya kolonilerinin mikrografı, bir koloninin yakın çekimi (inset). (C) Normal cihaz çalışması sırasında açılan bağlantının yakın çekim. (D) Mikrojel ve boncuk eşleştirmesini takiben toplanan emülsiyonların mikrografı, boncuk ve lysed kolonisi (inset) ile bir damla gösteren yakın çekim. Tüm ölçek çubukları = 100 μm. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
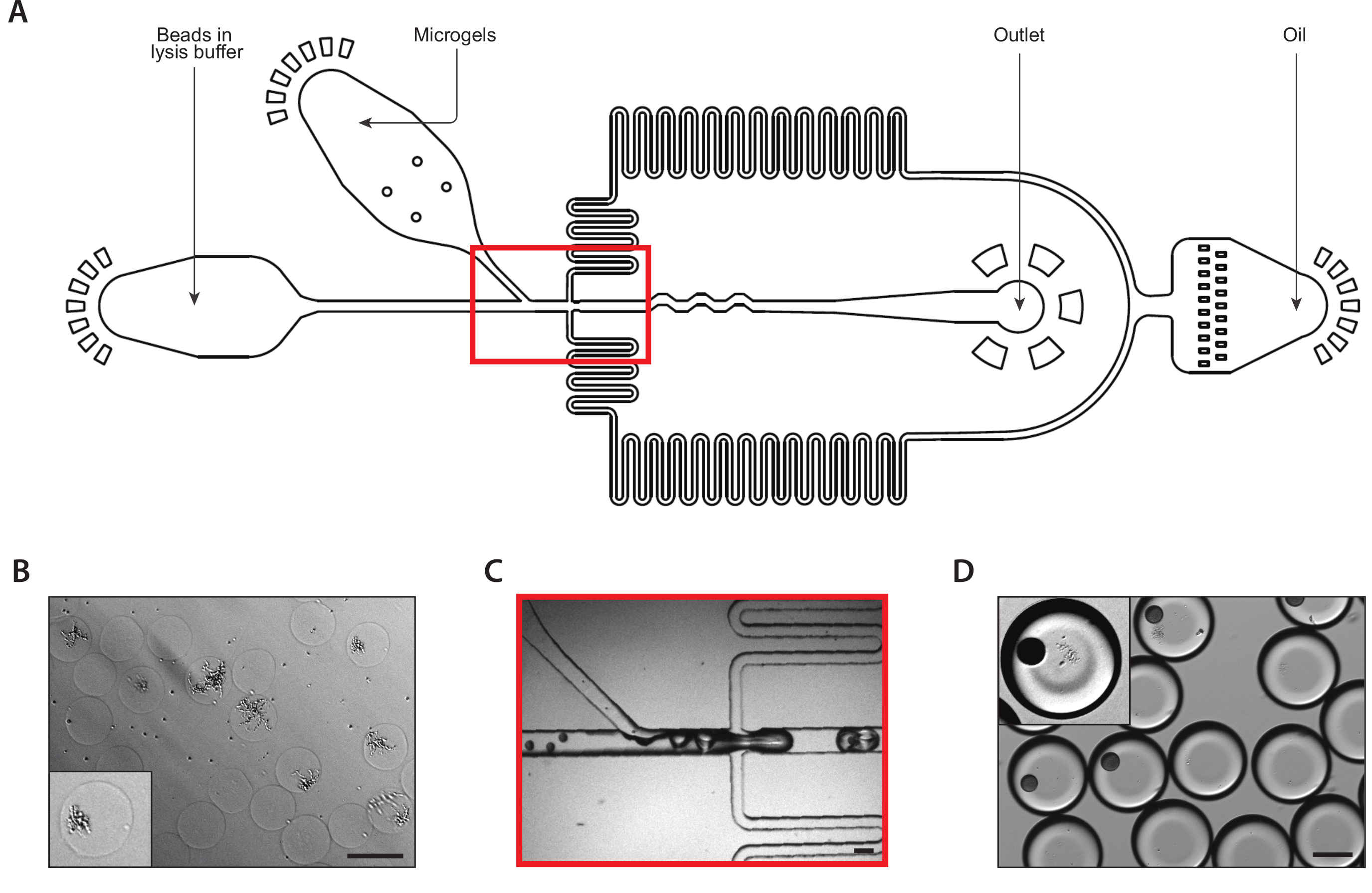{kind=link}

Şekil 4: C. albicansbeyaz-opak anahtarlama yanıtı analizi . (A) liu17bir referans veri kümesi ile birlikte bir örnek veri kümesi nin tSNE arsa . (B) Transkripsiyonların kümelanması, tSNE çiziminde görselleştirilen üç kümeyi ortaya çıkarır. (C) Beyaz-opak anahtarlama yanıtında yer alan anahtar genler, ana bileşen analizi ile belirlenen varyasyona katkıda bulunmuştur. (D) TSNE çiziminde işaretlenmiş kümeler tarafından YFP ve WH11 normalleştirilmiş ifade düzeylerinin keman çizimleri. **p <<< 0.05 ve * p << 0.05 gösterir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
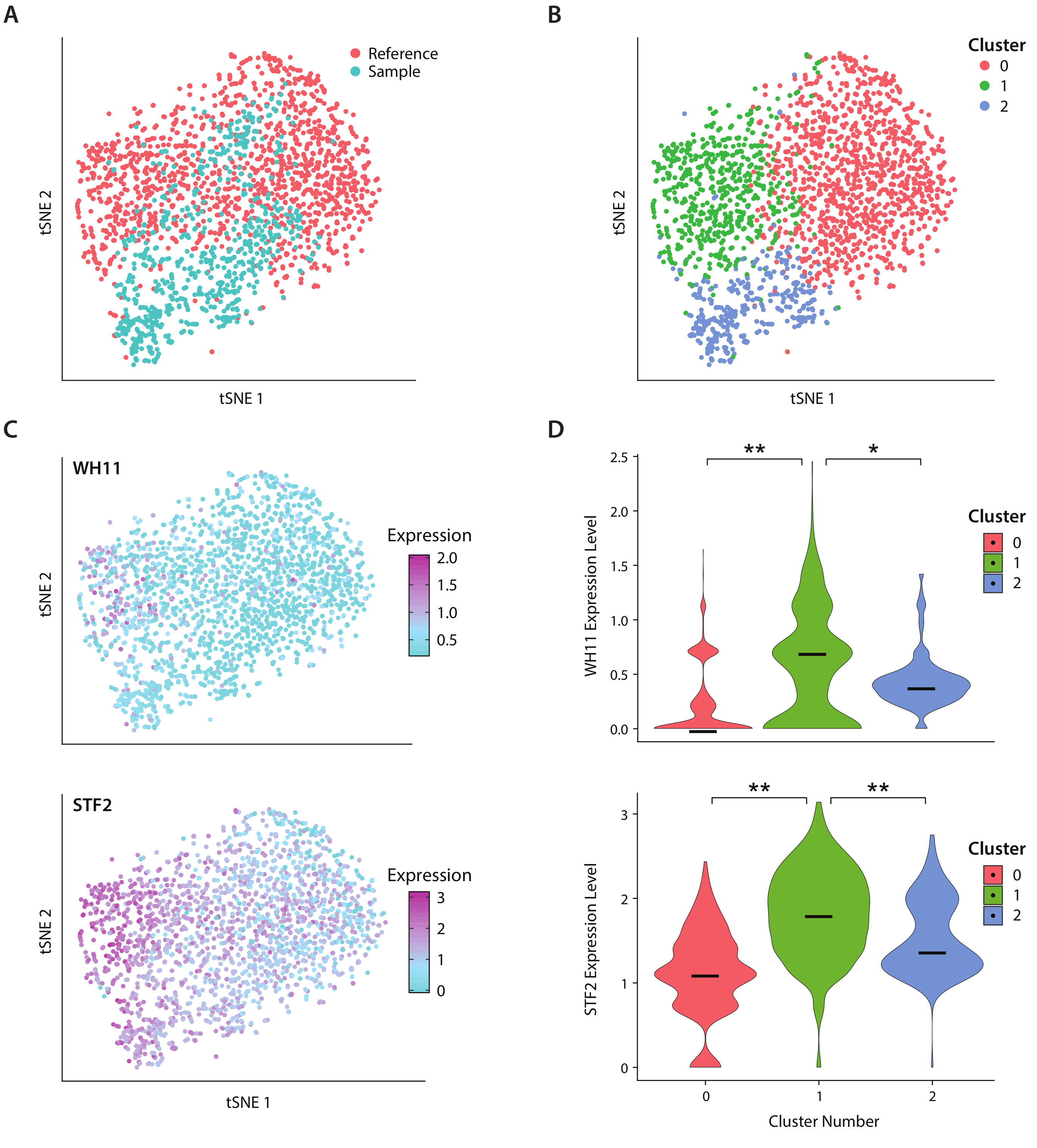{kind=link}
Ek Şekil 1 ve 2. Bu filgures indirmek için lütfen buraya tıklayın.
Ek Dosyalar 1-3. Bu dosyaları indirmek için lütfen buraya tıklayınız.
Tartışmalar
İzojenik maya kolonisi RNA dizilemesi (ICO-seq) için metodumuz, tasarlanmış maya suşlarının yüksek iş lenme alanı taraması için yayınlanmış tek hücreli RNA sıralama platformu Drop-Seq'yi uyarlar. Maya hücreleri tipik bir memeli hücresinin mRNA kopyalarının %10'undan daha azını içerir ve mRNA yakalamadan önce bozulması gereken bir hücre duvarına sahiptir16. Bu iki faktör mayanın Drop-Seq veya diğer damlacık tabanlı scRNA-seq platformlarına doğrudan uygulanmasını engellemektedir. Bu sorunları gidermek için hidrojellerin içindeki tek hücreleri kapsüllüyor ve rna dizilimi için yeterli giriş materyali sağlamak için koloniler halinde büyütüyor ve lizis ve mRNA yakalamadan önce spheroplastlar üretmek için maya hücre duvarını sindiriyoruz. Bu değişiklikler, orijinal Drop-Seq iş akışıyla karşılaştırıldığında ICO-seq iş akışında ek karmaşıklık ekler ve kullanıcıların sorunsuz bir şekilde ilerlemesini sağlamaları gereken kritik adımlardır.
A cihazının düzgün çalışması agarose hidrojelleri içinde tek maya hücrelerinin kapsüllemesi için gereklidir. Birden fazla maya hücresine sahip hidrojel sayısını en aza indirmek için giriş maya süspansiyonunun doğru sayımı takip edilmeli, aynı yandan yeterli hidrojelin mRNA yakalama sırasında makul bir hücre yakalama verimi sağlamak için tek bir hücre içermesini sağlamak gerekir. Mikroakışkan cihaz çalışması sırasında, agarose jel karışımı iyi çözülmüş olmalı ve cihaz tıkanma olasılığını en aza indirmek için bir şırınga filtresi nden geçirilmelidir. Agarose jel karışımı viskoz ve tek bir kanal sekiz ayrılır bölge özellikle tıkanmaya eğilimlidir. Cihazın o bölgesindeki cihaz işletimini görselleştirmek için yüksek hızlı bir kamerayı ortalayarak, kullanıcılar sekiz kanalın her birinden çıkan damlacıkların tekdüzeliğini izleyebilir ve herhangi bir kanaldaki tıkanıklıklara bağlı tekdüzelik değişirse hızlı bir şekilde tepki verebilir. Mikroskop altında toplanan emülsiyon küçük bir miktar incelenmesi yüksek kaliteli emülsiyon onaylamak için ikincil bir yöntem sağlar.
Hidrojeller içinde maya kolonilerinin büyümesini takiben, tek koloni düzeyinde kaliteli mRNA çıkarılmasını sağlamak için çeşitli önlemler gereklidir. Mayanın hidrojel kültüründe olduğu zamanı optimize etmek önemlidir, çünkü maya kültürde çok uzun süre bırakılırsa, birçok su jellerinin sınırlarından kaçacaktır, bu da RNA dizilimi sırasında daha yüksek bir arka plan sinyaline ve hücre tipleri arasında ayrım yaparken daha düşük hassasiyete yol açacaktır. Zymolyase kullanarak sferoplastların doğru üretimi, hücrenin lysis tamponuna maruz kaldıktan sonra mRNA'nın serbest bırakılmasını sağlar. Zymolyase'den sonra maya kolonilerinin görsel muayenesi daha parlak maya hücreleri vermelidir. Yanlış hücre duvarı sindirimi daha düşük RNA yakalama verimliliğine yol açacaktır. Son olarak, hidrojeller Cihaz B'ye enjekte edilirken yakın ambalajlı olmalıdır.
Bizim yöntem ile potansiyel bir endişe maya mikrogel kültürü önemli ölçüde gen ekspresyonunu değiştirebilir olmasıdır. Mikrojeller ve agar maya gen ekspresyonu araştıran önceki çalışma gen ekspresyonu ortalamaları farklılıkları göstermek ama genel olarak olumlu bir korelasyon17, maya suşları çeşitli bu iddianın daha fazla araştırma ihtiyatlı olmasına rağmen. Yöntem aynı zamanda Poisson istatistikleri14aşağıdaki mRNA yakalama boncuk stochastic yükleme nedeniyle sınırlı hücre yakalama verimliliği vardır. Şu anda damlaların yaklaşık % 10'u boncuk ve koloni içeriyor ve çift kapsülleme oranının %1'in altında olması bekleniyor. Çift kapsüllemeler RNA-seq veri analizi sırasında kafa karıştırıcı elemanlara yol açar ve bunların filtreleme zorlu kalır23; %25'lik bir yakalama oranı, çift kapsüllemelerin karşılık gelen artışa yol açacaktır 5%(Ek Şekil 2). Biz Drop-Seq platformu kullanarak ICO-seq göstermek rağmen, ticari olarak kullanılabilir 10x Genomik Krom platformu15,gibi, istatistiksel yerine deterministically boncuk yakalamak tanıtmak diğer damlacık RNA-seq platformları vardır,24. Bu platformların ICO-seq ile entegrasyonu, Poisson istatistiklerinin izin verdiğinden çok daha fazla yakalama verimliliğini artırabilir. Son olarak, damlacık RNA-seq temel bir sınırlama sıralama sonra ilgi hücreleri kurtarmak için yetersizliktir. Bu yöntem kullanılarak analiz edilecek maya kitaplıklarının türleri göz önünde bulundurulduğunda bu sınırlama dikkate alınmalıdır.
Hücreden hücreye heterojenite, E. coli25 ve Sgibi mikroplar için klonal düzeyde gösterilmiştir. cerevisiae26 yeni hücre devletlerin bir toplu düzey analizi aksi maske olacağını ortaya koymaktadır. C. albicans üzerinde yapılan toplu RNA-seq analizleri ya nüfus çapında transkripsiyon değişiklikleri bakmak eğilimindedir, ya da iki ayrı popülasyon olarak beyaz ve opak hücreler27,28. ICO-seq uygulaması ek alt devletlerin keşfine yol açabilir ve diğer maya türleri içinde yeni hücre durumları keşfetmek için analitik bir çerçeve sağlar. Ancak, hidrojeller içinde hücrelerin büyümesi maya ile sınırlı değildir: diğer hücre tipleri, memeli gibi, bakteriyel, ve diğer mantar hücreleri de hidrojeller içinde ekili olabilir29,30. Tek hücrelere karşı isojenik kolonilerin dizilişi, hücreden hücreye değişimine bağlı biyolojik gürültünün ortalamasına yol açarak hücre tipleri arasındaki ayrımcılığı n ilerletir. Bu, genetik çeşitliliğin belirli sentez yolları üzerinde odaklandığı hücreleri analiz ederken yardımcı olabilir. ICO-seq hücre tipi giriş genişletilmiş olanakları ve ticari olarak kullanılabilir damlacık RNA-seq platformları ile potansiyel entegrasyonu genetik düzeyde hücresel heterojenite diseksiyon için umut verici bir platform olarak ICO-seq pozisyonları.
Açıklamalar
Yazarlar çıkar çatışması beyan etmezler.
Teşekkürler
Bu proje Ulusal Bilim Vakfı Kariyer Ödülü DBI-1253293, Ulusal Sağlık Enstitüleri Yeni Yenilikçi Ödülü DP2AR068129 ve hibe R01HG0008978, Ulusal Bilim Vakfı Teknoloji Merkezi hibe DBI-1548297 ve UCSF Merkezi Hücresel İnşaat için desteklenmiştir. ARA ve ZJG Chan-Zuckerberg Biohub Müfettişler vardır.
Malzemeler
| Name | Company | Catalog Number | Comments |
| 0.22 um syringe filter | Milipore Sigma | SLGP033RS | |
| 0.5M EDTA, pH 8.0 | Thermo-Fisher | 15575020 | Used to make TE-TW buffer |
| 0.75 mm biopsy punch | World Precision Instruments | 504529 | |
| 1 mL syringes | BD | 309628 | |
| 1H,1H,2H-Perfluoro-1-Octanol (PFO) | Sigma-Aldrich | 370533 | |
| 1M Tris-HCI, pH 8.0 | Thermo-Fisher | 15568025 | Used to make TE-TW buffer |
| 27 gauge needles | BD | 305109 | |
| 3 mL syringes | BD | 309657 | |
| 3" silicon wafers, P type, virgin test grade | University Wafers | 447 | |
| 3D-printed centrifuge syringe holder | (custom) | (custom) | See Supplemental Files for 3D print file |
| Agarose, low gelling temperature | Sigma-Aldrich | a9414 | |
| Aquapel (hydrophobic glass treatment) | Pittsburgh Glass Works | 47100 | |
| Drop-Seq Beads | ChemGenes | MACOSKO-2011-10 | |
| Glass microscope slides (75 mm x 50 mm) | Corning | 294775X50 | |
| Ionic Krytox Surfactant | Synthesis instructions in ref 14. Can substitute with PEG-PFPE surfactant. | ||
| Isopropanol | Sigma-Aldrich | 109827 | |
| NaCl | Sigma-Aldrich | S9888 | |
| Novec 7500 | 3M | 98-0212-2928-5 | Commonly knowns as HFE 7500 |
| PBS | Fisher Scientific | BP243820 | |
| PE-2 polyethylene tubing | Scientific Commodities | B31695-PE/2 | |
| PEG-PFPE surfactant | Ran Biotechnologies | 008-FluoroSurfactant | |
| PGMEA developer | Sigma-Aldrich | 484431 | |
| Photomasks | CadArt Servcies | (custom) | See Supplemental Files for mask designs |
| Spin coater | Specialty Coating Systems | G3P-8 | |
| SSC Buffer | Sigma-Aldrich | S6639 | |
| SU-8 2100 | MicroChem | Y111075 | |
| SU-8 2150 | MicroChem | Y111077 | |
| Sylgard 184 silicone elastomer kit | Krayden | 4019862 | |
| Tween-20 | Sigma-Aldrich | P1379 | Used to make TE-TW buffer |
| YR Digestion buffer | Zymo Research | R1001-1 | Spheroplasting buffer |
| YR Lysis Buffer | Zymo Research | R1001-2 | |
| Zymolyase | Zymo Research | E1005 | Spheroplasting enzyme mixture |
Referanslar
- Peralta-Yahya, P. P., Zhang, F., Del Cardayre, S. B., Keasling, J. D. Microbial engineering for the production of advanced biofuels. Nature. 488 (7411), 320-328 (2012).
- Curran, K. A., Alper, H. S. Expanding the chemical palate of cells by combining systems biology and metabolic engineering. Metabolic Engineering. 14 (4), 289-297 (2012).
- Botstein, D., Chervitz, S. A., Cherry, J. M. Yeast as a model organism. Science. 277 (5330), 1259-1260 (1997).
- Mager, W. H., Winderickx, J. Yeast as a model for medical and medicinal research. Trends in Pharmacological Sciences. 26 (5), 265-273 (2005).
- Krivoruchko, A., Siewers, V., Nielsen, J. Opportunities for yeast metabolic engineering: Lessons from synthetic biology. Biotechnology Journal. 6 (3), 262-276 (2011).
- Vanella, R., et al. Yeast-based assays for screening 11β-HSD1 inhibitors. Microbial Cell Factories. 15 (1), (2016).
- Zhuang, X., Chappell, J. Building terpene production platforms in yeast. Biotechnology and Bioengineering. 112 (9), 1854-1864 (2015).
- Agresti, J. J., et al. Ultrahigh-throughput screening in drop-based microfluidics for directed evolution. Proceedings of the National Academy of Sciences. 107 (9), 4004-4009 (2010).
- Wang, G., et al. RNAi expression tuning, microfluidic screening, and genome recombineering for improved protein production in Saccharomyces cerevisiae. Proceedings of the National Academy of Sciences. 116 (19), 9324-9332 (2019).
- Beneyton, T., et al. Droplet-based microfluidic high-throughput screening of heterologous enzymes secreted by the yeast Yarrowia lipolytica. Microbial Cell Factories. 16 (1), 18(2017).
- Sjostrom, S. L., et al. High-throughput screening for industrial enzyme production hosts by droplet microfluidics. Lab on a Chip. 14 (4), 806-813 (2014).
- Nadal-Ribelles, M., et al. Sensitive high-throughput single-cell RNA-seq reveals within-clonal transcript correlations in yeast populations. Nature Microbiology. 4 (4), 683-692 (2019).
- Gasch, A. P., et al. Single-cell RNA sequencing reveals intrinsic and extrinsic regulatory heterogeneity in yeast responding to stress. PLoS Biology. 15 (12), 2004050(2017).
- Macosko, E. Z., et al. Highly parallel genome-wide expression profiling of individual cells using nanoliter droplets. Cell. 161 (5), 1202-1214 (2015).
- Klein, A. M., et al. Droplet barcoding for single-cell transcriptomics applied to embryonic stem cells. Cell. 161 (5), 1187-1201 (2015).
- von der Haar, T. A quantitative estimation of the global translational activity in logarithmically growing yeast cells. BMC Systems Biology. 2, 87(2008).
- Liu, L., Dalal, C. K., Heineike, B. M., Abate, A. R. High throughput gene expression profiling of yeast colonies with microgel-culture Drop-seq. Lab on a Chip. 19 (10), 1838-1849 (2019).
- Fallah-Araghi, A., Baret, J. C., Ryckelynck, M., Griffiths, A. D. A completely in vitro ultrahigh-throughput droplet-based microfluidic screening system for protein engineering and directed evolution. Lab on a Chip. 12 (5), 882-891 (2012).
- Berman, J., Sudbery, P. E. Candida albicans: A molecular revolution built on lessons from budding yeast. Nature Reviews Genetics. 3 (12), 918-930 (2002).
- Srikantha, T., Soll, D. R. A white-specific gene in the white-opaque switching system of Candida albicans. Gene. 131 (1), 53-60 (1993).
- van der Maaten, L., Hinton, G. Visualizing data using t-SNE. Journal of Machine Learning Research. 9, 2579-2605 (2008).
- Sun, Y., et al. Deletion of a yci1 domain protein of Candida albicans allows homothallic mating in MTL heterozygous cells. mBio. 7 (2), 00465-00516 (2016).
- McGinnis, C. S., Murrow, L. M., Gartner, Z. J. DoubletFinder: Doublet Detection in Single-Cell RNA Sequencing Data Using Artificial Nearest Neighbors. Cell Systems. 8 (4), 329-337 (2019).
- Baran-Gale, J., Chandra, T., Kirschner, K. Experimental design for single-cell RNA sequencing. Briefings in Functional Genomics. 17 (4), 233-239 (2018).
- Silander, O. K., et al. A Genome-Wide Analysis of Promoter-Mediated Phenotypic Noise in Escherichia coli. PLoS Genetics. 8 (1), 1002443(2012).
- Newman, J. R. S., et al. Single-cell proteomic analysis of S. cerevisiae reveals the architecture of biological noise. Nature. 441 (7095), 840-846 (2006).
- Tuch, B. B., et al. The Transcriptomes of Two Heritable Cell Types Illuminate the Circuit Governing Their Differentiation. PLoS Genetics. 6 (8), 1001070(2010).
- Romo, J. A., et al. Global Transcriptomic Analysis of the Candida albicans Response to Treatment with a Novel Inhibitor of Filamentation. mSphere. 4 (5), 00620(2019).
- Huang, H., et al. Generation and manipulation of hydrogel microcapsules by droplet-based microfluidics for mammalian cell culture. Lab on a Chip. 17 (11), 1913-1932 (2017).
- Lin, X., Nishio, K., Konno, T., Ishihara, K. The effect of the encapsulation of bacteria in redox phospholipid polymer hydrogels on electron transfer efficiency in living cell-based devices. Biomaterials. 33 (33), 8221-8227 (2012).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiThis article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır