
Method Article
Phenotipizzazione del ceppo di lievito ad alta velocità effettiva con sequenziamento dell'RNA basato su droplet
In questo articolo
Riepilogo
Un collo di bottiglia nel ciclo "design-build-test" dell'ingegneria microbica è la velocità con cui possiamo eseguire schermi funzionali di ceppi. Descriviamo un metodo ad alto throughput per lo screening della deformazione applicato a centinaia a migliaia di cellule di lievito per esperimento che utilizza il sequenziamento dell'RNA basato sulle gocciole.
Abstract
I potenti strumenti disponibili per modificare i genomi dei lieviti hanno reso questo microbo una preziosa piattaforma per l'ingegneria. Sebbene sia ora possibile costruire librerie di milioni di ceppi geneticamente distinti, lo screening per un fenotipo desiderato rimane un ostacolo significativo. Con le tecniche di screening esistenti, esiste un compromesso tra l'output delle informazioni e la velocità effettiva, con lo screening ad alta velocità effettiva in genere eseguito su un prodotto di interesse. Pertanto, presentiamo un approccio per accelerare lo screening della deformazione adattando il sequenziamento dell'RNA a singola cellula a colonie di picoliter ipogeni di ceppi di lievito geneticamente ingegnerizzati. Per affrontare le sfide uniche dell'esecuzione del sequenziamento dell'RNA sulle cellule di lievito, coltiviamo colonie di lieviti isogenici all'interno di idrogel e spheroplast prima di eseguire il sequenziamento dell'RNA. I dati di sequenziamento dell'RNA possono essere utilizzati per dedurre i fenotipi del lievito e ordinare percorsi ingegnerizzati. La scalabilità del nostro metodo affronta un ostacolo critico nell'ingegneria microbica.
Introduzione
Un obiettivo primario dell'ingegneria microbica è quello di modificare i microbi per indurli a produrre preziosi composti1,2. S. cerevisiae è stato l'organismo primario per l'ingegneria microbica grazie alla sua facilità di cultura e all'ampiezza degli strumenti disponibili per l'ingegneria del suo genoma3,4,5. Tuttavia, rimane un ostacolo nell'esecuzione di schermi funzionali sul lievito modificato: la produttività dello screening è in ritardo rispetto all'ingegneria genomica per ordini di grandezza. Lo screening prevede in genere l'isolamento dei ceppi in piastre di microwell e la loro fenotipizzazione misurando la produzione di un composto specifico6,7. La produttività di questo processo è limitata dalle grandi quantità di reagente necessarie per sodire singoli ceppi in cento reazioni al microlitro. La microfluidica a goccia fornisce una soluzione interessante per aumentare la produttività dello screening del lievito per ordini di grandezza da reazioni di downscaling normalmente eseguite in piastre ben8. Tuttavia, come per gli schermi di piastra, gli schermi delle goccioline in genere rilevano composti di singoli prodotti, il che fornisce informazioni limitate sulla funzione globale del percorso ingegnerizzato9,10,11.
Il sequenziamento dell'RNA (RNA-seq) può consentire una caratterizzazione più completa del funzionamento della via, consentendo di valutare simultaneamente i livelli di espressione di tutti i geni pertinenti12,13. Inoltre, i metodi delle goccioline consentono di profilare migliaia di celle per ogni esperimento, fornendo la velocità effettiva necessaria alle librerie di screening delle varianti ingegnerizzate14,15. Tuttavia, i metodi RNA-seq sono ottimizzati per le cellule dei mammiferi; lievito, in confronto, hanno meno mRNA per cellula e una parete cellulare che è difficile rimuovere16, precludendo il loro sequenziamento con metodi esistenti. Se si potesse ideare un metodo di gocciolina ad alto contenuto di velocità effettiva per consentire l'rna-seq di lievito, fornirebbe una piattaforma di fenotipizzazione scalabile, conveniente e ricca di informazioni per l'ingegneria del lievito.
Vi presentiamo un protocollo dettagliato del nostro metodo recentemente sviluppato per il sequenziamento delle cellule di lievito utilizzando microfluidics gocciolina ad alta produttività17. Per superare la sfida dell'RNA limitato, incapsulamo e coltiviamo singole cellule di lievito nelle sfere di idrogel picoliter. La coltura duplica le cellule, producendo centinaia di copie che condividono lo stesso percorso ingegnerizzato; questo riduce la variazione dovuta all'espressione genica a singola cellula, aumentando significativamente la quantità di RNA disponibile per il sequenziamento. Dopo l'amplificazione basata sulla coltura, sferova le cellule, rimuovendo la parete cellulare attraverso la digestione ezimatica sfusa. Le membrane cellulari rimangono intatte, in modo che ogni colonia isogenica e l'mRNA associato rimangano incapsulati nelle loro sfere di idrogel. Questo ci permette di accoppiare le singole colonie con reagenti di cattura dell'mRNA e tampone di lisi, e l'mRNA da catturare, codificare a barre e sequenziare in seguito al flusso di lavoro Drop-Seq14. Il nostro metodo consente lo screening a livello trascrittoma di migliaia di colonie di lieviti isogenici per esperimento.
Protocollo
1. Fabbricazione di dispositivi microfluidici
- Fabbricazione di mastro SU-8
- Progettare la maschera negativa per i canali microfluidici per i dispositivi A e B (File supplementari 1 e 2) utilizzando un software di progettazione assistita da computer e stamparli su pellicola di circuito con risoluzione di almeno 10 m.
- Mettere un wafer di silicio pulito da 75 mm su un rivestimento di spin e versare circa 1 mL di SU-8 al centro. Accendere l'aspirapolvere per fissare il wafer al mandrino.
- Per il dispositivo A, spin coat SU-8 2150 a 500 rpm per 30 s, seguito da 30 s a 2.750 rpm. Per il dispositivo B, spin-coat SU-8 2100 a 500 rpm per 30 s, seguito da 30 s a 2.500 rpm. Questo produrrà SU-8 strati di spessore 200 m e 120 m, rispettivamente.
- Togliere il wafer dallo spin-coater e mettere su una piastra calda a 95 gradi centigradi per 60 min a cuocere a morbido.
- Togliere il wafer dalla piastra calda e lasciarlo raffreddare a temperatura ambiente. Posizionare la maschera sopra il wafer, ed esporre sotto un collimator 190 mW, 365 nm UV LED per 2 min.
- Mettere il wafer su un set a piastra calda a 95 gradi centigradi per 5 min per la cottura post-esposizione.
- Rimuovere il wafer e lasciarlo raffreddare a temperatura ambiente. Mettere il wafer in un bagno di propilene glicol monomethyl eteato (PGMEA) per 20 min.
- Sciacquare il wafer con PGMEA seguito da isopropanol. Se durante questo processo è visibile un residuo opaco, ripetere il risciacquo con PGMEA e isopropanolo. Asciugare all'aria il wafer.
- Mettere il wafer su una piastra calda a 95 gradi centigradi per 3 min.
- Rimuovere e posizionare il wafer in un piatto Petri di 90 mm di diametro.
- Casting polidimetilsiloxane (PDMS) su master SU-8
- Mescolare insieme un rapporto di massa 10:1 della base in silicone con l'agente di polimerità. Degas il PDMS dopo la miscelazione per circa 30 min.
- Versare PDMS degassato sulla parte superiore del master SU-8 fino a quando uno strato spesso almeno 5 mm si forma sulla parte superiore del wafer.
- Degas il PDMS in cima al wafer per circa 30 min.
- Mettere il wafer in un forno a 65 gradi centigradi per almeno 80 min per curare il PDMS.
- Tagliare la lastra PDMS curata dal wafer.
- Posizionare la lastra PDMS con le caratteristiche microfluidiche rivolte verso l'alto e perforare i fori di inslizione e di uscita con un punzone biopsia da 0,75 mm.
- Pulire un vetrino di vetro da 50 mm x 75 mm con isopropanolo e rimuovere tutta la polvere dal lato delle caratteristiche microfluidiche della lastra PDMS con nastro adesivo.
- Esporre il vetrino di vetro pulito e la lastra PDMS con le caratteristiche microfluidiche faccia fino a 100 Pa (1 mbar O2)plasma per 1 min.
- Posizionare la lastra PDMS con le caratteristiche a faccia in giù sul vetrino di vetro per consentire l'incollaggio. Mettere il vetrino in un forno a 65 gradi centigradi per almeno 30 minuti per completare l'incollaggio.
- Trattare tutti i canali microfluidici vunerando con un fluido di trattamento superficiale fluorurato. Cuocere il dispositivo in un forno a 65 gradi centigradi per almeno 10 min per far evaporare il fluido.
2. Incapsulamento di lievito negli idrogel utilizzando il dispositivo A
- Prendere il lievito che cresce in una coltura di sospensione e contare su un emocitometro.
- Risospendere le cellule in salina tampone fosfato (PBS) a una concentrazione di circa 750 k/mL. Questo assicura che 30% di idrogel avrà una cella di lievito in loro. Solo circa la metà delle cellule di lievito cresce in colonie, portando al 15% degli idrogel contenenti colonie di lieviti.
- Mescolare il punto di fusione ultrabasso è salito al 2% w/v in PBS e riscaldare a 90 gradi centigradi fino a sciogliersi. Questo richiede 10 dollari min.
- Caricare la miscela di agarose in una siringa con un filtro da 0,22 m collegato in una pompa di siringa davanti al riscaldatore spaziale impostato a 80 gradi centigradi.
- Caricare una siringa riempita con la sospensione del lievito e una siringa contenente olio fluorurato con 2% w/v fluorosurfactant ionico18 in pompe di siringa.
- Prendi il dispositivo splitter a goccia di flusso fatto nella sezione 1 e collega il tubo dalle siringhe al dispositivo. Guidare il tubo dalla presa in un tubo conico 15 mL in un secchio di ghiaccio per la raccolta delle gocce.
- Scorrere le tre soluzioni nel dispositivo con le seguenti velocità di flusso:
- Scorrere la sospensione del lievito con la portata di 3 mL/h.
- Scorrere la miscela di agarose alla portata di 3 mL/h.
- Scorrere l'olio fluorurato alla portata di 15 mL/h.
- Raccogliere circa 1 mL di emulsione. Attendere altri 5 min per consentire all'agarose di impostare completamente.
3. Rottura e lavaggio gel per la cultura
- Aggiungere all'emulsione un volume uguale del 20% di perfluorooctanolo (PFO) nell'olio fluorurato. Invertire il tubo conico un paio di volte per consentire la miscelazione.
- Girare verso il basso l'emulsione rotta a 2.000 x g per 2 min. I gel si fermano sopra le fasi dell'olio e del PFO.
- Rimuovere la fase dell'olio e aggiungere 2 mL di buffer TE-TW (10 mM MM Tris pH : 8,0, 1 mM EDTA, 0.01% Tween-20) per risospendere i gel. Trasferire la sospensione in un nuovo tubo conico da 15 mL.
- Pellet giù i gel come nel passo 3.2 e lavare ancora una volta in TE-TW per un totale di due lavaggi.
- Rimuovere i gel supernatali e risospendere in 2 mL di supporti. Trasferire in un tubo di coltura 5 mL.
- Incubare a 30 gradi durante la notte sotto agitazione.
NOTA: Dopo l'incubazione notturna, gli idrogel di lievito possono essere conservati a 4 gradi centigradi per diversi giorni.
4. Lisi colonia di lievito
- Trasferire gel su un tubo conico da 15 ml e idrogel a pellet a 2.000 x g per 2 min.
- Lavare gli idrogel in PBS 2x.
- Lavare in 1x tampone spheroplasting 1x.
- Eseguire una diluizione da 2 a 50 volte l'enzima spheroplastingr in tampone spheroplastinge e aggiungere 1 mL agli idrogel.
- Incubare a 37 gradi centigradi per 1 ora. Il lievito trattato avrà un aspetto più trasparente (Figura 3A).
- Prendere il fondo 0,8 mL di sospensione idrogel e trasferire in una siringa non con serto da 1 mL.
- Posizionare la siringa nel supporto della siringa stampata in 3D (File supplementare 3) e ruotare a 2.000 x g per 2 min. Questo farà sì che gli idrogel chiudano pacco nella testa della siringa.
5. cattura di mRNA da colonie di lievito litonato utilizzando il dispositivo B
- Prendi 240.000 perline Drop-Seq e trasferiscili in un tubo conico da 15 mL.
- Pellet Drop-Seq filatura verso il basso a 1.000 x g per 1 min.
- Rimuovere il supernatante e risospendere le perline in 2 mL di 0,9x lievito buffer con 500 mM di cloruro di sodio per una concentrazione di sospensione perline di 120.000 perline/mL.
- Trasferire la sospensione del tallone in una siringa da 3 mL con una barra di agitazione inserita.
- Preparare una siringa contenente diversi millilitri del 2% w/v perfluoropolyether-polyethylene glicole (PFPE-PEG) in olio fluorurato.
- Evacuare tutta la testa acquosa della siringa contenente idrogel imballati e chiudere la siringa.
- Inserire l'idrogel, la sospensione del tallone e le siringhe di olio nelle pompe di siringhe e collegare tramite tubi nel dispositivo di incapsulamento realizzato nella sezione 1.
- Collegare dal tubo di uscita in un tubo conico 50 mL sul ghiaccio.
- Scorrere le tre soluzioni nel dispositivo con le seguenti velocità di flusso:
- Far scorrere gli idrogel a 0,4 mL/h.
- Scorrono la sospensione del tallone a 0,4 mL/h.
- Scorrere l'olio fluorurato a 1,6 mL/h.
- Raccogliere 1 mL di emulsione o eseguire il dispositivo fino a quando non ci sono più idrogels sinistra.
6. generazione di cDNA, preparazione della libreria di sequenziamento e sequenziamento
- Aggiungere 30 mL di buffer SSC 6x e 1 mL di PFO all'emulsione raccolta come indicato nel protocollo Drop-Seq14.
- Continua a seguire il protocollo Drop-Seq per generare cDNA da mRNA catturati su perline, preparazione della libreria di sequenziamento e analisi dei dati di sequenziamento.
Risultati
Abbiamo adattato il flusso di lavoro Drop-Seq pubblicato in precedenza14 per il sequenziamento delle colonie isogeniche (ICO-seq) per eseguire la profilazione dell'espressione genica delle colonie di lieviti inogenici. Abbiamo isolato singole cellule di lievito e le abbiamo incapsulate in microgel di agarose (Figura 1A). Dopo l'incubazione notturna di microgel, queste cellule di lievito incapsulate sono cresciute in colonie isogeniche. Prima di caricare i gel in un secondo dispositivo microfluidico per la cattura dell'mRNA, abbiamo digerito la parete cellulare del lievito per rendere l'mRNA più accessibile (Figura 1B, a sinistra). Abbiamo ravvicinato questi microgel e abbiamo fuso le perline di cattura dell'mRNA e il buffer di lisi. Alcune goccioline contenevano esattamente una perla abbinata a una colonia di lieviti lisci. Tutte le perle nell'emulsione sono state raccolte e il cDNA sintetizzato e sequenziato secondo il protocollo Drop-Seq.
Abbiamo generato colonie di lievito isogenico attraverso l'incapsulamento di una singola cella di lievito all'interno di microgel di agarose utilizzando un dispositivo microfluidico di coecapsulazione con uno splitter a otto gocce collegato (Figura 2A). Abbiamo diluito la sospensione del lievito di ingresso ad una concentrazione di 750.000 dollari/mL in modo che il 30% dei microgel abbia esattamente un lievito. Prima di inserire la temperatura di fusione ultrabassa nel dispositivo, l'abbiamo sciolta a una temperatura elevata e mantenuto la siringa a questa temperatura per evitare la gelazione prematura. Alla giunzione di generazione di goccia (Figura 2B), le cellule di lievito sono state inizialmente incapsulate in 160 goccioline di 160 m. Dopo la giunzione di generazione goccia una splitter otto pieghe ha diviso queste goccioline in otto goccioline da 80 m (Figura 2C). Un filtro di siringa è stato attaccato all'agarose fuso per evitare la formazione di zoccoli all'interno dei canali, che può essere stretto come 37 m durante la caduta-splitting. Abbiamo raccolto l'emulsione sul ghiaccio, che ha immediatamente iniziato il processo di gelazione delle agarose. È stata calcolata la polidispersionità di un'emulsione tipica in modo che sia del 6%(figura supplementare 1),anche se i valori di polidispersionità fino al 10% sono accettabili. Una volta che i gel di agarose set, abbiamo rotto l'emulsione e rimosso la fase di olio. I gel sono stati lavati in tampone acquoso prima dell'immersione nei media di crescita. L'incubazione notturna dei microgel ha portato alla crescita di colonie isogeniche all'interno di alcuni dei microgel (Figura 2D). La percentuale di idrogel contenenti colonie di almeno 20 cellule dipendeva dalle condizioni di coltura, tra cui il tempo di incubazione e la composizione dei media. Nella nostra dimostrazione con C. albicans, abbiamo determinato che circa il 15% degli idrogel conteneva una colonia dopo 20 h di coltura delle sospensioni.
Un secondo dispositivo di coeincapsulazione ha estratto l'mRNA dalle colonie isogeniche (Figura 3A). Prima di caricare i microgel di lievito nel dispositivo microfluidico, abbiamo lavato e immerso i gel in una soluzione per digerire le pareti delle cellule di lievito. La corretta digestione delle cellule di lievito è stata verificata mediante microscopia, con lievito trattato con una morfologia più riflettente (Figura 3B). Abbiamo chiudere i microgel in una siringa e abbiamo sintonizzato la portata dell'ingresso del gel in modo che un gel fosse in ogni goccia. Flusso di perline di cattura mRNA nel buffer di lisi mescolato con il flusso di gel ravvicinato prima della giunzione di drop-making (Figura 3C). Abbiamo raccolto un'emulsione risultante di 160 goccioline di m, e le colonie hanno cominciato a mentire e rilasciare il loro contenuto cellulare. Abbiamo caricato perline con una diluizione limitante per ridurre al minimo il numero di gocce contenenti più perline, ma l'imballaggio ravvicinato dei gel durante la consegna ha provocato circa il 10% delle gocce raccolte contenenti una perlina con una colonia lisciviata (Figura 3D).
Abbiamo analizzato l'espressione genica di C. albicans, una specie di lievito presente nel microbioma intestinale umano, utilizzando il flusso di lavoro ICO-seq. C. albicans è noto per la sua capacità di passare tra due diversi stati cellulari, chiamato bianco e opaco19. Usiamo un ceppo ingegnerizzato di albicani C, ceppo R-Y122, che sostituisce una copia del gene WH11, attivo solo nelle cellule bianche con YFP20. Abbiamo ottenuto una serie di profili di espressione genica usando il flusso di lavoro e li abbiamo usati per l'analisi delle colonie che esprimono almeno 300 geni unici. Come set di dati di riferimento, abbiamo usato i dati dell'espressione di C. Albicans ottenuti da uno studio pubblicato in precedenza17 e filtrato le colonie che esprimevano meno di 600 geni unici. Dopo aver eseguito l'analisi del componente principale (PC) e unariduzionedella dimensionalità tSNE (TSNE) tstocatico ( tSNE), abbiamo trovato la concordanza generale tra il set di dati di esempio e il riferimento (Figura 4A). L'analisi del PC ha rivelato che YFP e WH11 hanno contribuito in modo significativo ai primi due PC. Inoltre, l'analisi tSNE ha rivelato tre cluster(Figura 4B). Mentre il cluster 2 era costituito prevalentemente da celle del set di dati campione, i cluster 0 e 1 erano costituiti da cellule di entrambi i campioni. Sovrapponendo l'espressione WH11 sul tSNE (Figura 4C, pannello superiore), abbiamo determinato che il cluster 1 probabilmente conteneva colonie bianche. È stato inoltre rilevato che l'espressione STF2 è aumentata nel cluster 1 (Figura 4C, pannello inferiore), coerente con i dati ottenuti in precedenza17. Nei cluster 0 e 2, WH11 e STF2 sono stati significativamente downregolamentat rispetto al cluster 1 (Figura 4D). I geni coinvolti nella fermentazione, come ad esempio l'ADH1,sono stati upregolati nel cluster 0, in linea con precedenti studi sulle cellule opache22. Abbiamo scoperto che le colonie nel cluster 2 avevano diminuito l'RNA ribosomico rispetto alle colonie negli ammassi 0 e 1. Anche se il campione e i set di dati di riferimento sono stati ottenuti utilizzando lo stesso stock di cellule, questo risultato suggerisce che anche sottili differenze nella manipolazione sperimentale possono influenzare l'espressione genica.

Figura 1: Panoramica del flusso di lavoro ICO-seq. (A) La coltivazione del lievito in una coltura di sospensione è stata diluita in tamponati e coencapsulateta con agarose fusa in un dispositivo generatore di goccioline a fuoco dinamico per consentire il caricamento di Poisson di microgel agarose con celle a lievito singolo. I gel impostati quando l'agarose si è raffreddata, la sospensione olio/acqua era rotta e l'olio è stato rimosso, producendo una sospensione di perline di gel in acqua. Dopo la coltura durante la notte, le cellule di lievito sono cresciute in colonie isogeniche all'interno dei microgel. (B) Le colonie sono state sottoposte a un buffer di degradazione della parete cellulare, dopo di che sono state vicine e coinziate con perline di cattura dell'mRNA in un secondo dispositivo microfluidico. L'imballaggio stretto dei microgel ha assicurato che ogni goccia aveva un gel, mentre il caricamento di Poisson delle perline riduceva la possibilità di perline multiple all'interno di una goccia. Le gocce raccolte sono state elaborate per la sintesi cDNA e la generazione di una libreria di sequenziamento. Fare clic qui per visualizzare una versione più grande di questa figura.
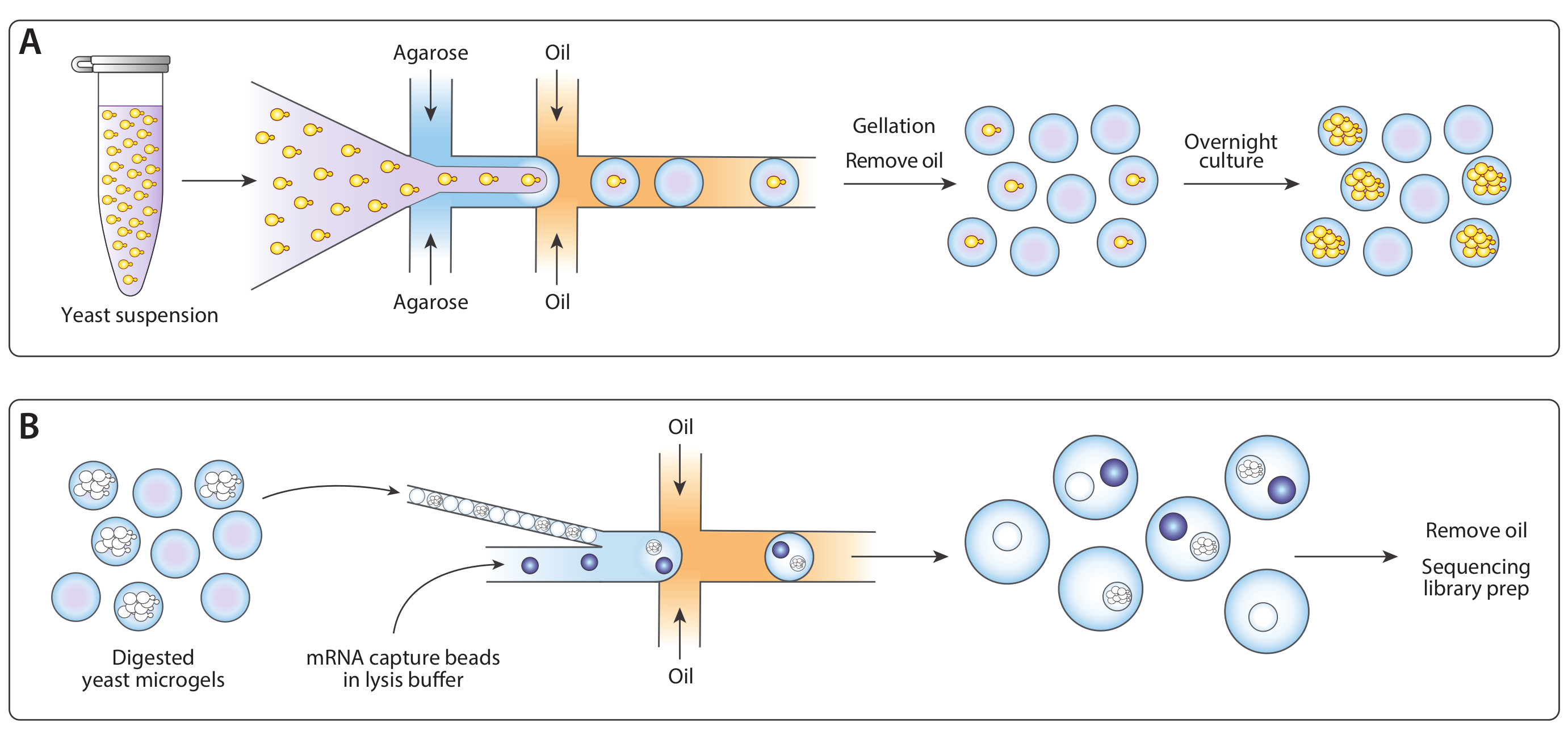{kind=link}

Figura 2: Generazione di colonie di lieviti inogeni all'interno di microgel di agarose utilizzando il dispositivo A. (A) Schematica del dispositivo microfluidico, che mostra le posizioni dei tre ingressi e delle porte di uscita. La giunzione drop-making è evidenziata in rosso. (B) Primo piano del nodo di drop-making durante il normale funzionamento del dispositivo. (C) Micrografo di goccioline raccolte, con un primo piano di una goccia contenente una cella incapsulata (insetto). (D) Micrografo di colonie di lievito isogenico in microgel di agarose a seguito di un'incubazione di 24 ore, con un primo piano di due colonie (insetto). Tutte le barre della scala - 100 m. Si prega di fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 3: Lisis e cattura di mRNA da colonie isogeniche utilizzando il dispositivo B. (A) Schematica del dispositivo microfluidico, che mostra le posizioni dei tre ingressi e delle porte di uscita. La giunzione drop-making è evidenziata in rosso. (B) Micrografo di colonie di lieviti a seguito della digestione della parete cellulare, con un primo piano di una colonia (insetto). (C) Primo piano del nodo di drop-making durante il normale funzionamento del dispositivo. (D) Micrografo delle emulsioni raccolte a seguito dell'abbinamento di microgel e perline, con un primo piano che mostra una goccia con un tallone e una colonia lisci (inset). Tutte le barre della scala - 100 m. Si prega di fare clic qui per visualizzare una versione più grande di questa figura.
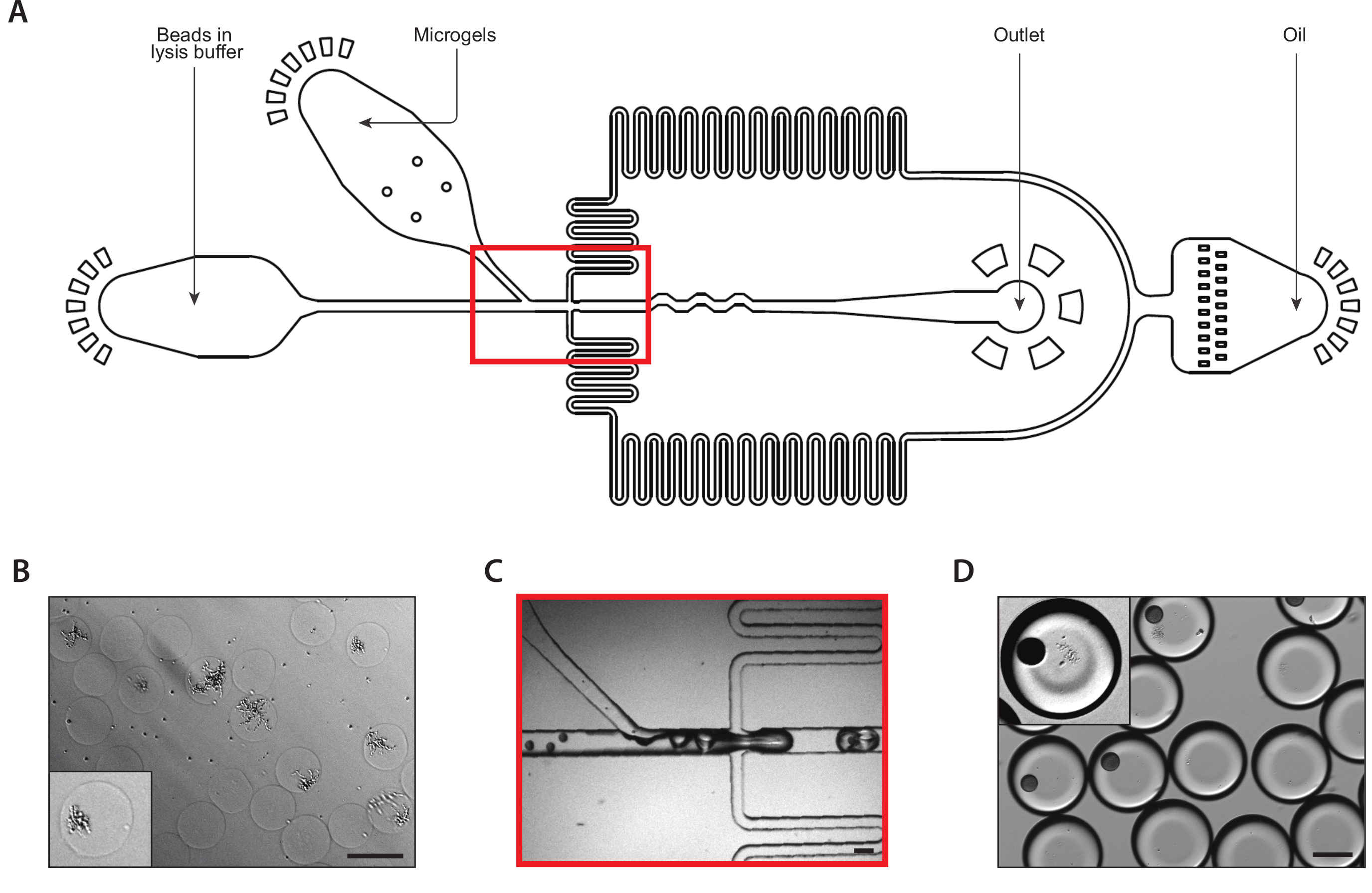{kind=link}

Figura 4: Analisi della risposta di commutazione bianco-opaco in C. albicans. (A) grafico tSNE di un set di dati di esempio combinato con un set di dati di riferimento di Liu17. (B) Il raggruppamento dei trascrittomi rivela tre cluster visualizzati su un grafico tSNE. (C) I geni chiave coinvolti nella risposta di commutazione bianco-opaco hanno contribuito alla variazione determinata attraverso l'analisi dei componenti principali. (D) Le trame di violino dei livelli di espressione normalizzati di YFP e WH11 da cluster contrassegnati sulla trama di tSNE. : indica p <<< 0,05 e : indica p << 0,05. Fare clic qui per visualizzare una versione più grande di questa figura.
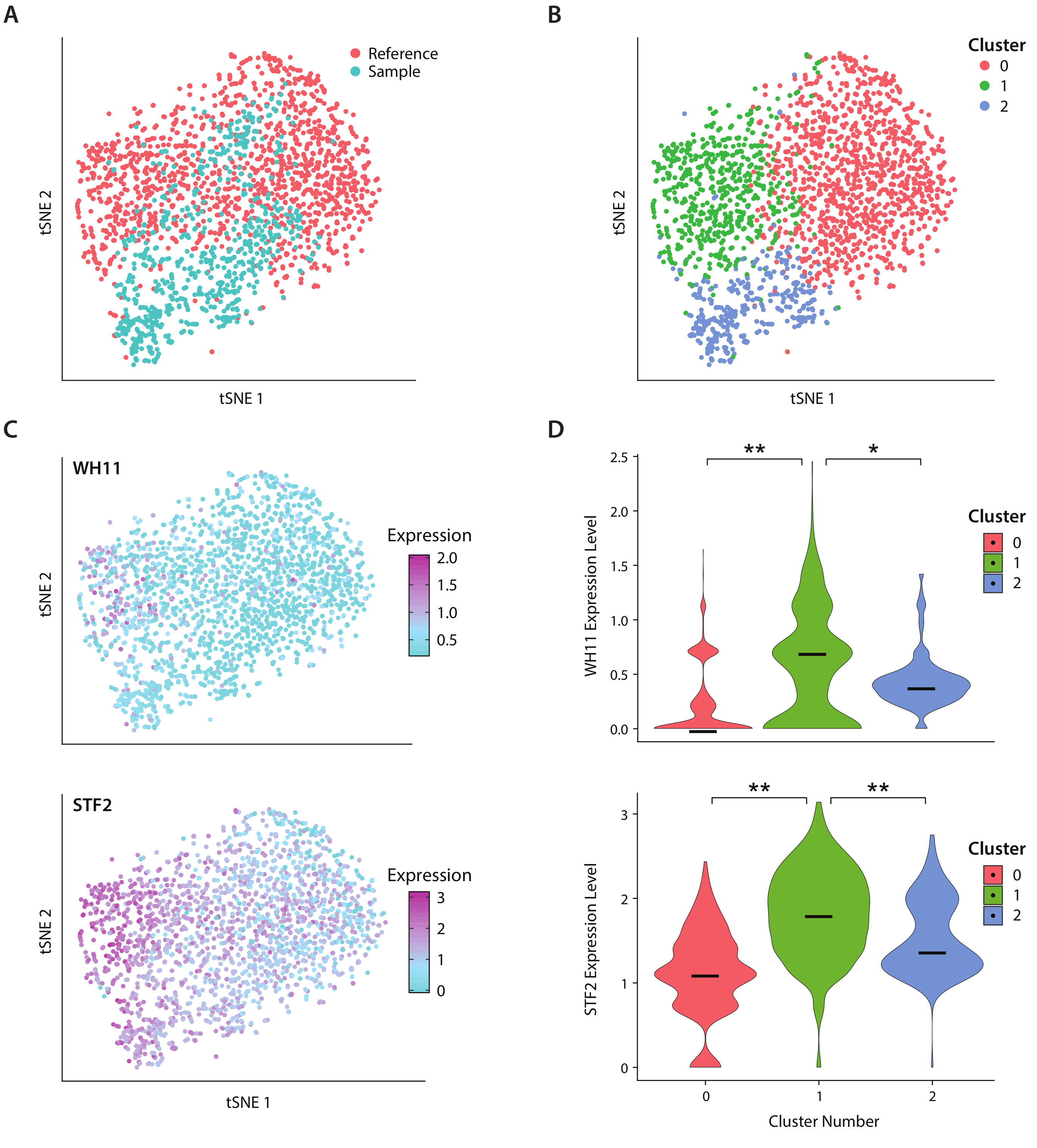{kind=link}
Figure supplementari 1 e 2. Clicca qui per scaricare questi filgures.
File supplementari 1-3. Fare clic qui per scaricare questi file.
Discussione
Il nostro metodo per il sequenziamento dell'RNA (ICO-seq) della colonia di lievito isogenico adatta una piattaforma di sequenziamento dell'RNA a singola cellula pubblicata, Drop-Seq, per lo screening ad alto throughput di ceppi di lievito ingegnerizzati. Le cellule di lievito contengono meno del 10% delle copie di mRNA di una tipica cellula mammaria e hanno una parete cellulare che deve essere degradata prima della cattura dell'mRNA16. Questi due fattori precludono l'applicazione diretta del lievito a Drop-Seq o ad altre piattaforme scRNA-seq a base di gocciolamento. Per risolvere questi problemi, incapsulamo singole cellule all'interno di idrogel e le cresciamo in colonie per fornire materiale di input sufficiente per il sequenziamento dell'RNA e digeriamo la parete cellulare del lievito per generare sferoplasti prima della lisi e della cattura di mRNA. Queste modifiche aggiungono ulteriore complessità nel flusso di lavoro ICO-seq rispetto al flusso di lavoro Drop-Seq originale e sono passaggi critici che gli utenti devono garantire di procedere senza intoppi.
Il corretto funzionamento del dispositivo A è necessario per incapsulare singole cellule di lievito all'interno di idrogel di agarose. È necessario seguire un corretto conteggio della sospensione del lievito di ingresso per ridurre al minimo il numero di idrogel con più di una cella di lievito, garantendo al contempo che un numero sufficiente di idrogel contenga una singola cella per garantire una ragionevole efficienza di cattura delle cellule durante la cattura di mRNA. Durante il funzionamento del dispositivo microfluidico, la miscela di gel di agarose deve essere ben disciolta e passata attraverso un filtro di siringa per ridurre al minimo la possibilità di intasamento del dispositivo. La miscela di gel di agarose è viscosa e la regione in cui un singolo canale si divide in otto è particolarmente incline all'intasamento. Centrando una telecamera ad alta velocità per visualizzare il funzionamento del dispositivo in quella regione del dispositivo, gli utenti possono monitorare l'uniformità delle goccioline che emergono da ciascuno degli otto canali e reagire rapidamente se l'uniformità cambia a causa degli zoccoli in uno dei canali. L'ispezione di una piccola quantità di emulsione raccolta al microscopio fornisce un metodo secondario per confermare un'emulsione di alta qualità.
Dopo la crescita delle colonie di lieviti all'interno degli idrogel, sono necessarie diverse precauzioni per garantire l'estrazione di mRNA di qualità a livello di singola colonia. È importante ottimizzare il lievito temporale sono in coltura idrogel, perché se il lievito viene lasciato in coltura per troppo tempo, molti sfuggiranno ai confini degli idrogel, portando ad un segnale di fondo più alto durante il sequenziamento dell'RNA e una minore sensibilità quando si discrimina tra i tipi di cellule. Una corretta generazione di sferoplasti che utilizzano la zimolisasi assicura che l'mRNA venga rilasciata in seguito all'esposizione cellulare al buffer di lisi. L'ispezione visiva delle colonie di lieviti che seguono la zimolisa si dovrebbe produrre cellule di lievito più brillanti. Una digestione impropria della parete cellulare porterà a una minore efficienza di cattura dell'RNA. Infine, gli idrogel devono essere strettamente imballati in quanto vengono iniettati nel dispositivo B. Il monitoraggio dell'ingresso dell'idrogel con una telecamera ad alta velocità consentirà la terminazione della raccolta dell'emulsione una volta che gli idrogel non saranno più imballati al momento dell'input nel dispositivo, altrimenti l'efficienza di acquisizione sarà influenzata.
Una potenziale preoccupazione con il nostro metodo è che la coltura microgel del lievito può alterare significativamente l'espressione genica. Il lavoro precedente che studia l'espressione genica del lievito nei microgel e sugli agar dimostrano differenze nelle medie dell'espressione genica, ma nel complesso una correlazione positiva17, anche se un'ulteriore indagine di questa affermazione su una varietà di ceppi di lievito è prudente. Il metodo ha anche una limitata efficienza di acquisizione cellulare a causa del caricamento stocastico di perline di cattura mRNA seguendo le statistiche di Poisson14. Attualmente circa il 10% delle gocce contiene una perlina e una colonia, e il tasso di incapsulamenti doppi dovrebbe essere inferiore all'1%. I doppi incapsulamenti portano a elementi di confusione durante l'analisi dei dati RNA-seq e il loro filtraggio rimane impegnativo23; un tasso di cattura del 25% porterebbe ad un corrispondente aumento di doppi incapsulamenti al 5%(Figura supplementare 2). Anche se dimostriamo ICO-seq utilizzando la piattaforma Drop-Seq, ci sono altre piattaforme di RNA-seq goccioline che introducono perline di cattura dell'mRNA in modo deterministico piuttosto che statisticamente, come la piattaforma 10x Genomics Chromium disponibile in commercio15,24. L'integrazione di queste piattaforme con ICO-seq potrebbe aumentare l'efficienza di cattura al di là di ciò che le statistiche di Poisson consentono. Infine, una limitazione fondamentale delle gocciolate RNA-seq è l'incapacità di recuperare le cellule di interesse dopo il sequenziamento. Questa limitazione deve essere presa in considerazione quando si considerano i tipi di librerie di lievito da analizzare utilizzando questo metodo.
L'eterogeneità tra cellule è stata dimostrata a livello clonale per microbi come E. coli25 e S. cerevisiae26 rivelando nuove cellule afferma che un'analisi a livello di massa sarebbe altrimenti mascherare. Le analisi bulk di RNA-seq eseguite su C. albicans tendono a osservare i cambiamenti del trascrittoma a livello di popolazione, o le cellule bianche e opache come due popolazioni separate27,28. L'applicazione di ICO-seq potrebbe portare alla scoperta di ulteriori sottostati e fornire un quadro analitico per scoprire nuovi stati cellulari all'interno di altre specie di lieviti. Tuttavia, la crescita delle cellule all'interno degli idrogel non è limitata al lievito: altri tipi di cellule, come mammiferi, batterie e altre cellule fungine possono anche essere coltivate all'interno di idrogel29,30. Il sequenziamento delle colonie isogeniche rispetto alle singole cellule porta alla media del rumore biologico a causa della variazione da cellula a cellula, migliorando la discriminazione tra i tipi di cellule. Questo può essere utile quando si analizzano le cellule in cui la diversità genetica si concentra su specifici percorsi di sintesi. Le possibilità ampliate di input di tipo cellulare per ICO-seq e la sua potenziale integrazione con piattaforme rna-seq gocciolabili disponibili in commercio posiziona ICO-seq come una piattaforma promettente per sezionare l'eterogeneità cellulare a livello genetico.
Divulgazioni
Gli autori non dichiarano conflitti di interesse.
Riconoscimenti
Questo progetto è stato sostenuto dal National Science Foundation Career Award DBI-1253293, dal National Institutes of Health New Innovator Award DP2AR068129 e dalla concessione R01HG008978, dalla National Science Foundation Technology Center DBI-1548297 e dal Centro cellulare per la costruzione dell'UCSF. ARA e JG sono i ricercatori di Chan-'uckerberg Biohub.
Materiali
| Name | Company | Catalog Number | Comments |
| 0.22 um syringe filter | Milipore Sigma | SLGP033RS | |
| 0.5M EDTA, pH 8.0 | Thermo-Fisher | 15575020 | Used to make TE-TW buffer |
| 0.75 mm biopsy punch | World Precision Instruments | 504529 | |
| 1 mL syringes | BD | 309628 | |
| 1H,1H,2H-Perfluoro-1-Octanol (PFO) | Sigma-Aldrich | 370533 | |
| 1M Tris-HCI, pH 8.0 | Thermo-Fisher | 15568025 | Used to make TE-TW buffer |
| 27 gauge needles | BD | 305109 | |
| 3 mL syringes | BD | 309657 | |
| 3" silicon wafers, P type, virgin test grade | University Wafers | 447 | |
| 3D-printed centrifuge syringe holder | (custom) | (custom) | See Supplemental Files for 3D print file |
| Agarose, low gelling temperature | Sigma-Aldrich | a9414 | |
| Aquapel (hydrophobic glass treatment) | Pittsburgh Glass Works | 47100 | |
| Drop-Seq Beads | ChemGenes | MACOSKO-2011-10 | |
| Glass microscope slides (75 mm x 50 mm) | Corning | 294775X50 | |
| Ionic Krytox Surfactant | Synthesis instructions in ref 14. Can substitute with PEG-PFPE surfactant. | ||
| Isopropanol | Sigma-Aldrich | 109827 | |
| NaCl | Sigma-Aldrich | S9888 | |
| Novec 7500 | 3M | 98-0212-2928-5 | Commonly knowns as HFE 7500 |
| PBS | Fisher Scientific | BP243820 | |
| PE-2 polyethylene tubing | Scientific Commodities | B31695-PE/2 | |
| PEG-PFPE surfactant | Ran Biotechnologies | 008-FluoroSurfactant | |
| PGMEA developer | Sigma-Aldrich | 484431 | |
| Photomasks | CadArt Servcies | (custom) | See Supplemental Files for mask designs |
| Spin coater | Specialty Coating Systems | G3P-8 | |
| SSC Buffer | Sigma-Aldrich | S6639 | |
| SU-8 2100 | MicroChem | Y111075 | |
| SU-8 2150 | MicroChem | Y111077 | |
| Sylgard 184 silicone elastomer kit | Krayden | 4019862 | |
| Tween-20 | Sigma-Aldrich | P1379 | Used to make TE-TW buffer |
| YR Digestion buffer | Zymo Research | R1001-1 | Spheroplasting buffer |
| YR Lysis Buffer | Zymo Research | R1001-2 | |
| Zymolyase | Zymo Research | E1005 | Spheroplasting enzyme mixture |
Riferimenti
- Peralta-Yahya, P. P., Zhang, F., Del Cardayre, S. B., Keasling, J. D. Microbial engineering for the production of advanced biofuels. Nature. 488 (7411), 320-328 (2012).
- Curran, K. A., Alper, H. S. Expanding the chemical palate of cells by combining systems biology and metabolic engineering. Metabolic Engineering. 14 (4), 289-297 (2012).
- Botstein, D., Chervitz, S. A., Cherry, J. M. Yeast as a model organism. Science. 277 (5330), 1259-1260 (1997).
- Mager, W. H., Winderickx, J. Yeast as a model for medical and medicinal research. Trends in Pharmacological Sciences. 26 (5), 265-273 (2005).
- Krivoruchko, A., Siewers, V., Nielsen, J. Opportunities for yeast metabolic engineering: Lessons from synthetic biology. Biotechnology Journal. 6 (3), 262-276 (2011).
- Vanella, R., et al. Yeast-based assays for screening 11β-HSD1 inhibitors. Microbial Cell Factories. 15 (1), (2016).
- Zhuang, X., Chappell, J. Building terpene production platforms in yeast. Biotechnology and Bioengineering. 112 (9), 1854-1864 (2015).
- Agresti, J. J., et al. Ultrahigh-throughput screening in drop-based microfluidics for directed evolution. Proceedings of the National Academy of Sciences. 107 (9), 4004-4009 (2010).
- Wang, G., et al. RNAi expression tuning, microfluidic screening, and genome recombineering for improved protein production in Saccharomyces cerevisiae. Proceedings of the National Academy of Sciences. 116 (19), 9324-9332 (2019).
- Beneyton, T., et al. Droplet-based microfluidic high-throughput screening of heterologous enzymes secreted by the yeast Yarrowia lipolytica. Microbial Cell Factories. 16 (1), 18(2017).
- Sjostrom, S. L., et al. High-throughput screening for industrial enzyme production hosts by droplet microfluidics. Lab on a Chip. 14 (4), 806-813 (2014).
- Nadal-Ribelles, M., et al. Sensitive high-throughput single-cell RNA-seq reveals within-clonal transcript correlations in yeast populations. Nature Microbiology. 4 (4), 683-692 (2019).
- Gasch, A. P., et al. Single-cell RNA sequencing reveals intrinsic and extrinsic regulatory heterogeneity in yeast responding to stress. PLoS Biology. 15 (12), 2004050(2017).
- Macosko, E. Z., et al. Highly parallel genome-wide expression profiling of individual cells using nanoliter droplets. Cell. 161 (5), 1202-1214 (2015).
- Klein, A. M., et al. Droplet barcoding for single-cell transcriptomics applied to embryonic stem cells. Cell. 161 (5), 1187-1201 (2015).
- von der Haar, T. A quantitative estimation of the global translational activity in logarithmically growing yeast cells. BMC Systems Biology. 2, 87(2008).
- Liu, L., Dalal, C. K., Heineike, B. M., Abate, A. R. High throughput gene expression profiling of yeast colonies with microgel-culture Drop-seq. Lab on a Chip. 19 (10), 1838-1849 (2019).
- Fallah-Araghi, A., Baret, J. C., Ryckelynck, M., Griffiths, A. D. A completely in vitro ultrahigh-throughput droplet-based microfluidic screening system for protein engineering and directed evolution. Lab on a Chip. 12 (5), 882-891 (2012).
- Berman, J., Sudbery, P. E. Candida albicans: A molecular revolution built on lessons from budding yeast. Nature Reviews Genetics. 3 (12), 918-930 (2002).
- Srikantha, T., Soll, D. R. A white-specific gene in the white-opaque switching system of Candida albicans. Gene. 131 (1), 53-60 (1993).
- van der Maaten, L., Hinton, G. Visualizing data using t-SNE. Journal of Machine Learning Research. 9, 2579-2605 (2008).
- Sun, Y., et al. Deletion of a yci1 domain protein of Candida albicans allows homothallic mating in MTL heterozygous cells. mBio. 7 (2), 00465-00516 (2016).
- McGinnis, C. S., Murrow, L. M., Gartner, Z. J. DoubletFinder: Doublet Detection in Single-Cell RNA Sequencing Data Using Artificial Nearest Neighbors. Cell Systems. 8 (4), 329-337 (2019).
- Baran-Gale, J., Chandra, T., Kirschner, K. Experimental design for single-cell RNA sequencing. Briefings in Functional Genomics. 17 (4), 233-239 (2018).
- Silander, O. K., et al. A Genome-Wide Analysis of Promoter-Mediated Phenotypic Noise in Escherichia coli. PLoS Genetics. 8 (1), 1002443(2012).
- Newman, J. R. S., et al. Single-cell proteomic analysis of S. cerevisiae reveals the architecture of biological noise. Nature. 441 (7095), 840-846 (2006).
- Tuch, B. B., et al. The Transcriptomes of Two Heritable Cell Types Illuminate the Circuit Governing Their Differentiation. PLoS Genetics. 6 (8), 1001070(2010).
- Romo, J. A., et al. Global Transcriptomic Analysis of the Candida albicans Response to Treatment with a Novel Inhibitor of Filamentation. mSphere. 4 (5), 00620(2019).
- Huang, H., et al. Generation and manipulation of hydrogel microcapsules by droplet-based microfluidics for mammalian cell culture. Lab on a Chip. 17 (11), 1913-1932 (2017).
- Lin, X., Nishio, K., Konno, T., Ishihara, K. The effect of the encapsulation of bacteria in redox phospholipid polymer hydrogels on electron transfer efficiency in living cell-based devices. Biomaterials. 33 (33), 8221-8227 (2012).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneThis article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati