
Bu içeriği görüntülemek için JoVE aboneliği gereklidir. Oturum açın veya ücretsiz deneme sürümünü başlatın.
Method Article
Zebra Balıklarında Omurgalı Segmentasyonunu İncelemek için 3-D Kuyruk Eksplant Kültürü
Bu Makalede
Özet
Burada, omurgalı segmentasyonunun canlı çalışmasını sağlayan zebra balığı arka vücut ekseninin 3 boyutlu doku kültürü protokolünü sunuyoruz. Bu eksplant model eksen uzama, morfojen kaynaklarının değiştirilmesi ve hücre altı çözünürlük doku düzeyinde canlı görüntüleme üzerinde kontrol sağlar.
Özet
Omurgalı embriyolar ana vücut eksenlerini tekrarlayan somitler, omurların, kasların ve cildin öncülleri olarak düzenlerler. Embriyonun kuyruk ucu arka olarak uzadıkça presomitik mezodermden (PSM) giderek bölünür. Düzenli periyodiklik ve boyut ölçeği ile somites oluşur. Zebra balığı, genetik olarak çekişli olduğu ve canlı görüntülemeye izin veren şeffaf embriyolara sahip olduğu için popüler bir model organizmadır. Bununla birlikte, somitogenez sırasında, balık embriyoları büyük, yuvarlatılmış bir yumurta sarısı etrafına sarılır. Bu geometri, zebra balığı embriyolarındaki PSM dokusunun canlı görüntülenmesini, özellikle de yakın objektif çalışma mesafesi gerektiren daha yüksek çözünürlüklerde sınırlar. Burada zebra balığı kuyruk eksplantlarının canlı görüntülenmesi için düzleştirilmiş 3 boyutlu doku kültürü yöntemi sunuyoruz. Kuyruk eksplantları, eksen uzamasının orantılı bir yavaşlamasını ve rostrocaudal somit uzunluklarının kısalmasıyla bozulmamış embriyoları taklit eder. Eksen uzama hızını eksplant kültürü ile daha da durdurabiliyoruz. Bu, ilk kez, akssal uzamanın mekanistik girdisinden sinyal gradyanlarının kimyasal girişini çözememizi sağlar. Gelecekteki çalışmalarda, bu yöntem, zaman kontrollü farmasötik pertürbasyonlara veya omurgalı segmentasyonunun herhangi bir ilaç penetrasyon endişesi olmadan taranmasına izin vermek için mikroakışkan bir kurulumla birleştirilebilir.
Giriş
Organizmaların metamerik segmentasyonu doğada yaygın olarak kullanılmaktadır. Tekrarlanan yapılar, omurlar, kaslar, sinirler, damarlar, uzuvlar veya bir vücut planındaki yapraklar gibi yanal organların işlevselliği için gereklidir1. Eksenel simetrinin bu tür fizyolojik ve geometrik kısıtlamalarının bir sonucu olarak, Bilateria'nın annelidler, eklembacaklılar ve akorlar gibi çoğu phyla embriyonik dokularının (örneğin, ektoderm, mesoderm) antero-posteriorly segmentasyonunu sergiler.
Omurgalı embriyolar, paraksiyal mezodermlerini ana vücut ekseni boyunca sırayla türlere özgü aralıklar, sayımlar ve boyut dağılımları ile somitlere ayırırlar. Bir tür içindeki bireysel embriyolar arasında bu kadar sağlam olmasına rağmen, omurgalı türler arasında somit segmentasyonu çok yönlüdür. Segmentasyon, geniş bir zaman aralıkları rejiminde (zebra balıklarında 25 dakikadan insanlarda 5 h'ye kadar), boyutlarda (zebra balıklarının kuyruk somitlerinde ~20 μm'den farelerin gövde somitlerinde ~ 200 μm'ye) ve sayımlarda (zebra balıklarında 32'den mısır yılanlarında ~ 300'e)2. Daha ilginçtir ki, balık embriyoları çok çeşitli sıcaklıklarda (zebra balıkları için ~20,5 °C'den 34 °C'ye kadar) gelişebilirken, hem segmentasyon aralıklarını hem de eksenel uzama hızlarını telafi ederek uygun boyut dağılımlarıyla somitlerini sağlam tutarlar. Bu ilginç özelliklerin ötesinde zebra balığı, kardeş embriyolarının bir bolluğu ve erişilebilir genetik araçlarının dış, senkron ve şeffaf gelişimi nedeniyle omurgalılarda segmentasyonu incelemek için yararlı bir model organizma olarak kalır. Mikroskopi açısından olumsuz olarak, teleost embriyoları hacimli küresel bir yumurta sarısı üzerinde gelişir, etrafındaki gastrulating dokusunu gererek ve yuvarlar (Şekil 1A). Bu yazıda zebra balığı kuyrukları için düzleştirilmiş 3 boyutlu doku eksplant kültürü sunuyoruz. Bu eksplant sistem, yumurta sarısı kütlesinin küresel kısıtlamalarını atlatarak somit desenleme için balık embriyolarının yüksek çözünürlüklü canlı görüntülemesine erişim sağlar.

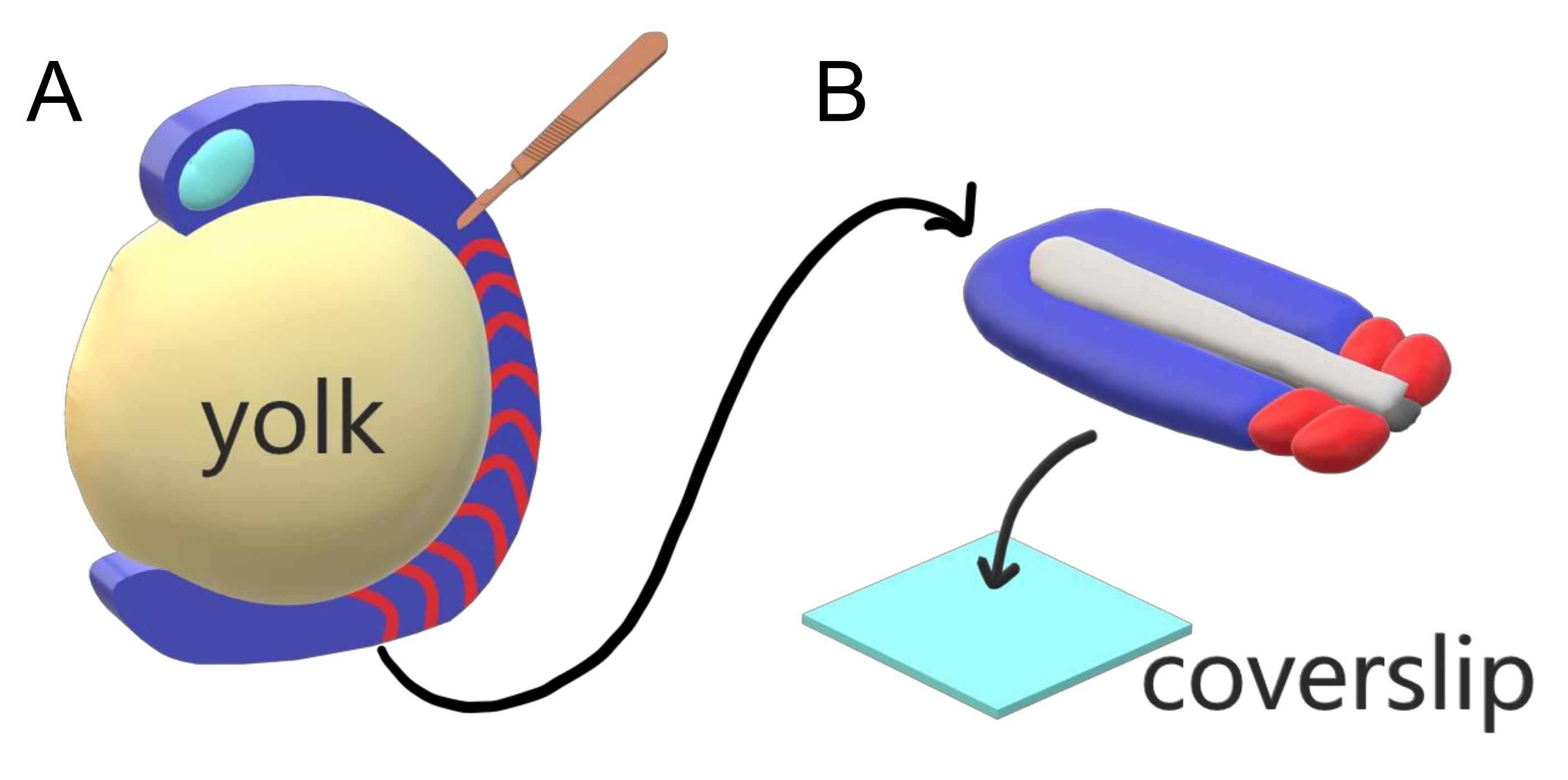
Şekil 1: Zebra Balığı Embriyoları için Slayt Odası Eksplant Sistemi. (A) Zebra balığı embriyolarının canlı görüntüleme için avantajları vardır, örneğin gastrulating embriyonik dokunun şeffaflığı (mavi), ancak doku, sağlam embriyolarda neredeyse objektif, yüksek çözünürlüklü görüntülemeyi önleyen hacimli küresel bir yumurta sarısı kütlesi (sarı) etrafında oluşur. Kuyruk eksplantları, somitlerin doku ön kısmından (kırmızı) kesilen mikrocerrahi bir bıçakla (kahverengi) başlayarak ve arka plan sarısı ile sınırda devam ederek kesilebilir. (B) Diseksiyonlu kuyruk eksplantları bir kapak (açık mavi) dorsoventrally üzerine yerlenebilir; üstte sinir dokusu (açık gri), altta notochord (koyu gri) tutmak. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
Protokol
Bu protokol, döllenme sonrası 1 günden daha genç canlı omurgalı embriyoların kullanımını içerir. Tüm hayvan deneyleri Cincinnati Çocuk Hastanesi Tıp Merkezi'nin etik kuralları altında gerçekleştirildi; hayvan protokolleri Kurumsal Hayvan Bakım ve Kullanım Komitesi tarafından gözden geçirilmiş ve onaylanmıştır (Protokol # 2017-0048).
1. Embriyo koleksiyonu
- Embriyo toplama gününden bir gece önce geçiş tanklarına zebra balığı çiftleri kurun. Embriyo gelişiminin hassas evreleme kontrolü için çiftleşme çiftleri arasındaki engelleri kullanın.
- Tercih edilen yumurtlama süresinden önce bariyerleri kaldırın ve 100 mm Petri kabında 15 dakika içinde yumurta toplayın.
- Petri kabının enkazını temizleyin. Tek bir debriyajdan 50'den fazla embriyo toplanırsa, debriyajı buna göre birden fazla Petri kabına bölün.
- Embriyoları% 50 epiboly evreye ulaşana kadar 28 ° C'de balık sistemi suyunda kuluçkaya yatırın (döllenme sonrası 5 saat). 3.2. basama kadar akvaryum sistemi suyu yerine E3 gibi standartlaştırılmış embriyo büyüme ortamı da kullanılabilir.
- Döllenmemiş yumurtaları stereoskop altında çıkarın ve embriyoları bir gecede 23,5 °C inkübatöre (O/N) taşıyın. Embriyolar, toplama gününü takip eden sabah 8-10 somites aşamasında olmalıdır.
2. Takım hazırlama
- Mikrocerrahi bıçağını, iğne uçlarını (dokunun diseksiyonu için kullanılır) ve cam Pasteur pipeti% 100 etanol (EtOH) ve ateş camında ıslatarak sterilize edin.
- 25 mm x 75 mm mikroskop slaytlarda iki kat şeffaf bant (~100-120 μm kalınlığı) kullanın. Her slaydın bant kaplamasının ortasında neşterle ~18 mm x 18 mm kare kuyular kesin.
- Hazırlanan slayt odalarını %70 EtOH ile silin. Bu kuyular ~ 40 μL orta tutacaktır.
3. Örnek hazırlama
- Stereoskop altında iki iğne şırıngasının ucunu kullanarak embriyoları deforionat. Embriyoları durulamak için balık sistemi suyu ile ayrı bir Petri kabına aktarın.
- Ateşle sırlı steril cam Pasteur pipet kullanarak embriyoları diseksiyon ortamı içeren 6 cm Petri kabına aktarın (Fenol Kırmızısı içermeyen L-Glutamin ile Leibovitz-15 hücre kültürü ortamı, 0.8 mM CaCl2 ve 1× antibiyotik-antimykotik çözelti).
NOT: Bu adımı izleyen tüm transferler için sterilize edilmiş bir cam pipet kullanmaya devam edin.- Diseksiyon sırasında polistiren talaşlardan kaçınmak için çıkarma prosedürü için cam bir Petri kabı kullanın.
- Slayt odasına 50 μL doku büyüme ortamı (diseksiyon ortamı ve% 10 FBS) koyun.
- Stereoskop altında diseksiyon için bir embriyoyu arka beyin yakınındaki yumurta sarısı dokusu kavşağında bir iğne ucu ile stabilize edin.
- Embriyonik dokuyu bir iğne ile sabit tutmak, mikrocerrahi bıçağı 45° 'de tutulan bıçakla kullanarak doku önünü arka beyine ayırın ve ön taraftan başlayıp kuyruk tomurcuklarına doğru hareket eden embriyonik dokunun sarısını soyun (Şekil 1A).
NOT: Sarısını temizlerken cilt dokusunu kaybetmemeye dikkat edin. Deri, diseksiyon sırasında embriyonun etrafında tek katmanlı elastik doku olarak kolayca soyulur, bu nedenle tanınması kolaydır. - Yumurta sarısı embriyonik vücuttan tamamen çıkarıldıktan sonra, yan cilt dokusunu kuyruk kapağından kesin. Son oluşan 3-4 somit'i sağlam tutarak, daha ön dokuyu kesin (tam eksenli eksplant).
- Yumurta sarısı esas olarak bu prosedürden sağlam çıkmalıdır. Yırtılmış bir yumurta sarısı durumunda, önemli yumurta sarısı granülleri dokunun ventral yüzeyine bağlı kalabilir. Öyleyse, kalan yumurta sarısı granüllerini nazikçe temizlemek için bir kirpik aracı kullanın.
NOT: Bir eksizyonun yan taraflarındaki cilt dokularının dengesizliği, dokunun düz bir büyüme yönünü korumasına izin vermez. Bunun yerine explant daha gerilmiş cildin yanına doğru bükülecektir. Bu dengesizlik mikrocerrahi bıçak yardımıyla cilt tabakası yırtılarak stereoskop altında düzeltilebilir. - Derisiz eksplantlar için, iğne ile cilt tabakasının bir ucuna bastırın ve mikrocerrahi bıçakla eksplante olan dokuyu soyun. Bu assiler kültürde vücut eksenlerini uzatmayacaklardır.
- Tam eksen eksplantlarına ek olarak, bu adımda alternatif eksplantlar yapılabilir. Örneğin, mikrocerrahi bıçağı (tam PSM eksplantları) kullanarak zaten parçalanmış somitleri parçalara ayırın veya PSM'yi yarı anteroposterioruna (yarı PSM eksplantları) ayırın. Bu tür alternatif eksplantların uygulanması için lütfen bölüm 5.1'e bakın.
- Yumurta sarısı esas olarak bu prosedürden sağlam çıkmalıdır. Yırtılmış bir yumurta sarısı durumunda, önemli yumurta sarısı granülleri dokunun ventral yüzeyine bağlı kalabilir. Öyleyse, kalan yumurta sarısı granüllerini nazikçe temizlemek için bir kirpik aracı kullanın.
- Diseksiyonlu eksplantı hemen görüntülemenin gerçekleştirileceği 22 mm x 22 mm'lik bir kapak kapağına aktarın.
- Eksplantı dorsoventral eksende, kapak ucuna dokunan ventral tarafta düzenleyin (Şekil 1B). 20 μL filtreli uç pipet kullanarak doku eksplant etrafındaki fazla ortamı yavaşça çıkarın.
NOT: Parçalanmış eksplantların kapak kapağına gecikmeli transferi, sarısının geometrik kısıtlamalarından kurtulduğundan dokuda deformasyonlara neden olur.
- Eksplantı dorsoventral eksende, kapak ucuna dokunan ventral tarafta düzenleyin (Şekil 1B). 20 μL filtreli uç pipet kullanarak doku eksplant etrafındaki fazla ortamı yavaşça çıkarın.
- Kapak kapağını büyüme ortamı dolu slayt odasının üzerine eksplant ile hızlı ve dikkatlice çevirin.
- Kabarcık oluşumunu önlemek için, kare örtünün bir tarafını bant haznesine yerleştirin ve diğer tarafı nazikçe serbest bırakın. Bu adımda explant'ı hareket ettirmemeye/deforme etmemeye özen edin.
- Bir laboratuvar dokusu üzerindeki slayt odasına basarak fazla media kanamasını yavaşça odadan çıkarın. Kapak, herhangi bir sızdırmazlık olmadan sıvı ortamın yüzey gerilimi nedeniyle canlı görüntüleme için slayt odasına sıkıca oturacaktır.
- Uzun süreli kültleme için (>6 saat), daha büyük bir oda kullanın. Bu gibi durumlarda slaytlarda iki paralel bant katmanı şeridi ile birlikte 22 mm x 50 mm dikdörtgen kapaklar kullanılabilir. Büyüme ortamına hava erişimini kolaylaştırmak için iki bant şeridi arasında ~1 mm genişliğinde bir boşluk bırakılabilir.
- Daha fazla eksplant hazırlamak için 3.3-3.8 adımlarını yineleyin. Hazırlanan eksplantlar ortalama ~30 μm/h hız ile A-P gövde eksenlerini uzatacak ve somitlerini 25 °C'de ~40 dk aralıklarla segmentlere ayıracaktır (Şekil 2A, Video 1).
- Uzun olmayan eksplantlar için, bir laboratuvar dokusunda fazla medyayı emerken numuneyi 3.8.2 adımda tutan slaytın kenarlarına hafif basınç uygulayın. Alternatif olarak, eksplantlar tek bant katmanlı slayt odalarında kültürlenebilir. Ayrıca, slayt odası yüzeyinin Tip I Kollajen ile kimyasal olarak aktive edilmesi, uzamayan eksplantlara neden olacaktır (Şekil 2B, Video 2).
- Kayma odalarını 1 saat boyunca oda sıcaklığında 15-20 mL önceden seyreltilmiş kollajen çözeltisi ile tamamen kaplayarak odanın Tip I Kollajen ile kaplamasını önceden gerçekleştirin. Steriliteyi korumak için bu protokol için bir laminer akış başlığı kullanın. Odaları sonunda diseksiyon ortamı ile dikkatlice durulayın.
- 15 somit aşamasından daha eski embriyolar için kuyruk eksplant dokusunu düz (dorsoventral) bir montaj yerine yanal olarak monte edin (Video 3). Kas seğirmesini önlemek için, kültür medyasına anestezik ajan olarak% 0.004 trikain çözeltisi ekleyin3.
- Uzun olmayan eksplantlar için, bir laboratuvar dokusunda fazla medyayı emerken numuneyi 3.8.2 adımda tutan slaytın kenarlarına hafif basınç uygulayın. Alternatif olarak, eksplantlar tek bant katmanlı slayt odalarında kültürlenebilir. Ayrıca, slayt odası yüzeyinin Tip I Kollajen ile kimyasal olarak aktive edilmesi, uzamayan eksplantlara neden olacaktır (Şekil 2B, Video 2).
4. Canlı görüntü alımı
- Görüntü örnekleri, somite segmentasyon boyutlarının ve dönemlerinin geniş alan iletilmiş ışık görüntülemesi için bir diseksiyon kapsamında veya transgenik muhabir balık hatları kullanılarak yapılandırılmış aydınlatma / konfokal / ışık tabakası mikroskopisi ile.
- Doku eksplantlarının sıcaklığını görüntüleme odası sıcaklığı ile en az 15 dakika boyunca dengelayın.
- Daha hassas sıcaklık kontrolü için ters mikroskop üzerine monte edilmiş ticari bir sıcaklık kontrol sistemi kullanın.
- Görüntü alma kare aralıklarını, biyolojik ilgi sürecine bağlı olarak 2 - 10 dakika olarak ayarlayın.
NOT: Zebra balığı somit segmentasyonu, tüm embriyolarda 30 °C ila 21,5 °C'lik canlı sıcaklıklar için 20 - 55 dakika arasında değişen hızlı bir işlemdir. Explants, tüm embriyolardan ~%30 daha yavaş bir şekilde uzayacak ve segmentlere ayıracaktır.- Canlı dokuya olası fototoksisiten kaçınmak için kanal edinim setleri arasında yeterli gecikme bırakmaya dikkat edin. Görüntüleme süresinin yarısından fazlası için dokuyu ekscitasyon ışınlarına maruz bırakmayın ve ışın yoğunluğunu mümkün olduğunca düşüremeyin.
NOT: Reaktif oksijen türlerinin (ROS) birikmesi genellikle canlı örneklerde fototoksiliğin başlıca nedenidir4. Ros çöpçü olarak askorbik asit, ROS aktivitesini tamponlamak ve fototoksisiyi hafifletmek için 4 mM konsantrasyonda büyüme ortamına desteklenebilir. Canlı görüntüleme sırasında fototoksisinin olumsuz etkilerini fark etmek zor olabilir. Kuyruk eksplantları bu açıdan avantajlıdır, çünkü mitotik arrest, engellenmiş doku gelişimi (yani, somit oluşumu, kuyruk uzaması) ve parçalanan doku gibi fototoksioksisitenin bazı görsel belirteçlerini fark etmek daha kolaydır. Ayrıntılı bir tartışma için lütfen sağlanan referans4'e bakın.
- Canlı dokuya olası fototoksisiten kaçınmak için kanal edinim setleri arasında yeterli gecikme bırakmaya dikkat edin. Görüntüleme süresinin yarısından fazlası için dokuyu ekscitasyon ışınlarına maruz bırakmayın ve ışın yoğunluğunu mümkün olduğunca düşüremeyin.
- Hücresel çözünürlük düzeyinde segmentlere ayırılması ve analiz edilmesi amaçlanan 4 boyutlu görüntüler elde etmek için tek hücreli evre RNA enjekte edilmiş embriyoları kullanın.
- 300 pg RNA'yı in vitro transkript membranlardan ve pCS-membran-ceruleanFP (Addgene plazmid #53749) veya pCS-memb-mCherry gibi nükleer floresan muhabir marker plazmidlerinden kullanın (Addgene plazmid #53750) pCS2+ H2B-mTagBFP2 (Addgene plazmid #99267) veya pCS2+ H2B-TagRFP-T (Addgene plazmid #99271) enjeksiyonlarında. Hücre zarı ve çekirdek işaretleyicileri olan örnek bir film için lütfen Video 4' e bakın.
NOT: PSM dokusunun ortalama hücre büyüklüğü, çekirdekleri ~3 - 4 μm'den oluşan yaklaşık 5 μm çapındadır. Uygun hücre segmentasyonu için ~0,5 μm piksel boyutu ve ~1 μm z kesitleme kaydedilmelidir.
- 300 pg RNA'yı in vitro transkript membranlardan ve pCS-membran-ceruleanFP (Addgene plazmid #53749) veya pCS-memb-mCherry gibi nükleer floresan muhabir marker plazmidlerinden kullanın (Addgene plazmid #53750) pCS2+ H2B-mTagBFP2 (Addgene plazmid #99267) veya pCS2+ H2B-TagRFP-T (Addgene plazmid #99271) enjeksiyonlarında. Hücre zarı ve çekirdek işaretleyicileri olan örnek bir film için lütfen Video 4' e bakın.
5. Kuyruk eksplantlarının immünostaining
NOT: Çeşitli diseksiyon senaryolarından sonra yetiştirilen dokular (uzama, uzama, kuyruk tomurcukları diseksiyon, yarı PSM vb.) düz monteli kuyruk eksplantlarıolarak 5, ilgi çekici proteinlerin daha fazla immün olarak ölçülmesi için slayt odalarından geri kazanılabilir. Burada, di-fosforilize hücre dışı sinyal düzenlenmiş kinaz (ppERK) eksplantların boyanması için kullanılan protokolü FGF sinyal gradyan okuma olarak sunuyoruz.
- İstenilen aşamaya kadar somitlerin oluşumundan sonra, kapak kapağını kaldırmadan slayt odasının köşesine dikkatlice kaydırın.
- Cam pasteur pipette ~100 μL ek diseksiyon ortamı yardımıyla, eksplantları slayttan kurtarın ve 64 kuyu hücreli bir hücre kültürü plakasına aktarın.
NOT: Bu adımdan başlayarak, tüm çözelti değiştirmeleri sabit numuneler için ayrı bir cam pipet ile diseksiyon kapsamı altında gerçekleştirilebilir. Bu, kuyulardaki eksplant dokuların kaybedilmesini veya aralarında aktarılmasını sağlayacaktır. - Tüm eksplantları aktardıktan sonra, aşırı ortamı kuyulardan tek tek emin ve PBS'deki (PFA) % 4 paraformaldehitin 100 μL'sini her kuyuya koyun.
DİkKAT: PFA kanserojen etkileri olan toksik bir çözeltidir. Kullanım yaparken uygun KKD kullanılmalıdır. - Eksplantları oda sıcaklığında 64 kuyulu bir plakaya bir çalkalayıcı üzerinde 1 saat sabitlayın.
- Doku eksplantları deformasyonlara karşı tüm embriyolara göre daha hassastır. Çalkalayıcı hızını buna göre ayarlayın.
- Fiksatifi 150 μL PBS-Tw (PBS'de % 0,1 Tween20) ile üç kez yıkayın. İlk yıkamayı belirli bir "PFA Atık" kabında toplayın.
- Her seferinde ~40 μL çözeltiyi %100 metanol (MeOH) ile değiştirerek 4×5 dakikalık adımlarla susuz kalsın.
DİkKAT: MeOH uçucu ve yanıcı toksik bir kimyasaldır. İyi havalandırılmış bir alanda çalışın ve kullanım için uygun KKD kullanın. - Dehidratasyonun son adımı olarak, tüm çözeltiyi kuyulardan çıkarın ve 100 μL MeOH ile değiştirin. 15 dakika boyunca -20 °C'de kuluçkaya yatır.
NOT: 5.11. adıma kadar çözümleri toplamak için belirli bir "MeOH Atık" kabı kullanın. - 50 μL MeOH ekleyin ve oda sıcaklığında 5 dakika çalkalayın.
- Her seferinde ~40 μL çözeltiyi PBS-T (PBS'de %0×,1 Triton-X 100) ile değiştirerek 4,5 dakikalık adımlarla eksplantları yeniden sulaşın. Çözümleri toplamak için belirli bir "MeOH Atık" kabı kullanın.
- Rehidrasyonun son adımı olarak, tüm çözeltiyi kuyulardan çıkarın ve 100 μL PBS-T ile değiştirin.
- Doku permeabilizasyonu için PBS'de %1,5 Triton-X 100 olan eksplantları çalkalayıcıdaki oda sıcaklığında 20 dakika boyunca tedavi edin.
- Numuneleri MAB-D-T (%0,1 Triton-X 100 deterjan ve %1 dimetil sülfit (DMSO) ile 150 mM NaCl 100 mM maleik asit tampon pH 7,5) 3×5 dk ile yıkayın.
DİkKAT: DMSO yanıcı ve zehirli bir mutajendir. Kullanım yaparken uygun KKD kullanılmalıdır. - Oda sıcaklığında 2 saat boyunca 100 μL/kuyu serum bloke çözeltisinde (MAB-D-T'de %2 fetal sığır serumu) eksplantları inkübasyona edin.
- Tüm blokaj çözeltisini 50-100 μL/kuyu birincil antikor çözeltisi (serum bloğunda ppERK'ya karşı monoklonal fare antikorunun 1:1000 seyreltilmesi) ile değiştirin. Çalkalayıcıda 4 °C'de O/N (>16 h) örnekleri kuluçkaya yatırın.
- Birincil antikor çözeltisini MAB-D-T ile 5×5 dk yıkayın.
- İkincil antikor çözeltisinde (Alexa Fluor 597 keçi anti-fare IgG2b (1:200) ve Hoechst 33342 (1:5000) MAB-D-T'de) O/N'de 4 °C'de bir çalkalayıcıda veya oda sıcaklığında 3 saat boyunca örnekleri kuluçkaya bırakın.
NOT: Bu adımdan başlayarak, ikincil antikorla tedavi edilen numunelerin ışığa maruz kalmasını önlemek için 64 kuyu plakasını alüminyum folyo ile örtün.
DİkKAT: Hoechst 33342 potansiyel bir kanserojendir. Kullanım yaparken uygun KKD kullanılmalıdır. - İkincil antikor çözeltisini PBS-Tw 3×5 dk ile yıkayın.
- Numuneleri ODA sıcaklığında 15 dakika PFA ile sabitleyin.
- Fiksatifi PBS-Tw ile yıkayın ve örnekleri% 60 gliserol içinde dengeleyin. Mikroskop slaytlarına oje ve görüntüleme için% 60 gliserol ile eksplantlar monte edin. Temsili immünostaining sonuçları için lütfen Şekil 3'ebakınız.
Sonuçlar
Bu protokol, canlı zebra balığı kuyruk eksplantlarının düz geometrik kültünü sağlar. Doku kültürü tüm embriyolara göre üç büyük avantaj sunar: 1) eksen uzama hızının kontrolü, 2) basit diseksiyon ile çeşitli sinyal (morfojen) kaynakları üzerinde kontrol ve 3) nesnel, yüksek büyütme ve yüksek NA canlı görüntüleme.
Kimyasal olarak işlem görmemiş slayt odaları, kuyruk eksplantının altındaki dokuya sarılan deri ektoderm tarafından ana eksenini (
Tartışmalar
Bu makalede zebra balığı embriyolarıiçin son zamanlarda geliştirdiğimiz ve kullandığımız bir doku kültürü eksplant tekniğinin ayrıntılı bir protokolü sunulmaktadır. Tekniğimiz civciv8 ve zebra balığı 9 ,10,11model organizmalarda önceki eksplant yöntemleri üzerine inşa edilmektedir. Bu protokolle hazırlanan kuyruk eksplantları, basit bir slayt odasında...
Açıklamalar
Yazarların açıklayacak ve çıkar çatışması ilan edecek hiçbir şeyi yoktur.
Teşekkürler
Balık bakımı için AECOM Zebrafish Core Facility ve Cincinnati Çocuk Veterinerlik Hizmetlerine, teknik yardım için Cincinnati Çocuk Görüntüleme Çekirdeğine, video prodüksiyona yardım için Didar Saparov'a ve el yazmasını düzenlediği için Hannah Seawall'a teşekkür ederiz. Bu yayında bildirilen araştırmalar, Ulusal Sağlık Enstitüleri Ulusal Genel Tıp Bilimleri Enstitüsü tarafından R35GM140805 ödül numarası altında E.M.Ö. İçerik sadece yazarların sorumluluğundadır ve Ulusal Sağlık Enstitülerinin resmi görüşlerini temsil etmek zorunda değildir.
Malzemeler
| Name | Company | Catalog Number | Comments |
| 1 mL Sub-Q Syringe with PrecisionGlide Needle | Becton, Dickinson and Co. | REF 309597 | for dechorionating embryos and manipulations |
| 200 Proof Ethanol, Anhydrous | Decon Labs | 2701 | for immunostaining |
| Antibiotic Antimycotic Solution (100×) | Sigma-Aldrich | A5955 | for tissue dissection media |
| Calcium Chloride Anhydrous, Powder | Sigma-Aldrich | 499609 | for tissue dissection media |
| Dimethylsulfoxide | Sigma-Aldrich | D5879 | for immunostaining |
| Disposable Scalpel, #10 Stainless Steel | Integra-Miltex | MIL4-411 | for preparing tape slide wells |
| Ethyl 3-aminobenzoate methanesulfonate salt (Tricaine) | Sigma-Aldrich | 886-86-2 | (optional) for anesthesizing tissues older than 20 somites stage |
| Fetal Bovine Serum (FBS) | ThermoFisher | A3160601 | additional for tissue culture media |
| Goat anti-Mouse IgG2b, Alexa Fluor 594 | Invitrogen | Cat#A-21145; RRID: AB_2535781 | secondary antibody for immunostaining |
| L-15 Medium with L-Glutamine w/o Phenol Red | GIBCO | 21083-027 | for tissue dissection media |
| Methanol | Sigma-Aldrich | 179337 | for immunostaining |
| Microsurgical Corneal Knife 2.85 mm Angled Tip Double Bevel Blade | Surgical Specialties | 72-2863 | for tissue dissection |
| Mouse monoclonal anti-ppERK | Sigma-Aldrich | Cat#M8159; RRID:AB_477245 | for ppERK immunostaining |
| NucRed Live 647 ReadyProbes Reagent | Invitrogen | R37106 | (optional) for live staining of cell nuclei |
| Paraformaldehyde Powder, 95% | Sigma-Aldrich | 158127 | for fixation of samples for immunostaining |
| Rat Tail Collagen Coating Solution | Sigma-Aldrich | 122-20 | (optional) for chemically activating slide chambers |
| Stage Top Incubator | Tokai Hit | tokai-hit-stxg | (optional) for temperature control during live imaging |
| Transparent Tape 3/4'' | Scotch | S-9782 | for preparing tape slide wells |
| Triton X-100 | Sigma-Aldrich | X100 | for immunostaining |
| Tween 20 | Sigma-Aldrich | P1379 | for immunostaining |
| Zebrafish: Tg(actb2:2xMCP-NLS-EGFP) | Campbell et al., 2015 | ZFIN: ZDB-TGCONSTRCT-150624-4 | transgenic fish with nuclear localized EGFP |
| Zebrafish: Tg(Ola.Actb:Hsa.HRAS-EGFP) | Cooper et al., 2005 | ZFIN: ZDB-TGCONSTRCT-070117-75 | transgenic fish with cell membrane localized EGFP |
Referanslar
- Assheton, R. . Growth in length: Embryological Essays. , (1916).
- Gomez, C., et al. Control of segment number in vertebrate embryos. Nature. 454 (7202), 335-339 (2008).
- Westerfield, M. . The Zebrafish Book: a guide for the laboratory use of zebrafish (Danio rerio), 3rd edition. , (1995).
- Icha, J., Weber, M., Waters, J. C., Norden, C. Phototoxicity in live fluorescence microscopy, and how to avoid it. BioEssays. 39 (1700003), (2017).
- Simsek, M. F., Ozbudak, E. M. Spatial fold change of Fgf signaling encodes positional information for segmental determination in zebrafish. Cell Reports. 24 (1), 66-78 (2018).
- Dubrulle, J., Pourquié, O. fgf8 mRNA decay establishes a gradient that couples axial elongation to patterning in the vertebrate embryo. Nature. 427 (6973), 419-422 (2004).
- Diez del Corral, R., et al. Opposing FGF and Retinoid Pathways Control Ventral Neural Pattern, Neuronal Differentiation, and Segmentation during Body Axis Extension. Neuron. 40 (1), 65-79 (2003).
- Stern, H. M., Hauschka, S. D. Neural tube and notochord promote in vitro myogenesis in single somite explants. Developmental Biology. 167 (1), 87-103 (1995).
- Langenberg, T., Brand, M., Cooper, M. S. Imaging brain development and organogenesis in zebrafish using immobilized embryonic explants. Developmental Dynamics. 228 (3), 464-474 (2003).
- Picker, A., Roellig, D., Pourquié, O., Oates, A. C., Brand, M. Tissue micromanipulation in zebrafish embryos. Methods in molecular biology. 546 (11), 153-172 (2009).
- Manning, A. J., Kimelman, D. Tbx16 and Msgn1 are required to establish directional cell migration of zebrafish mesodermal progenitors. Developmental Biology. 406 (2), 172-185 (2015).
- Kimmel, C. B., Ballard, W. W., Kimmel, S. R., Ullmann, B., Schilling, T. F. Stages of embryonic development of the zebrafish. Developmental Dynamics. 203 (3), 253-310 (1995).
- Kaufmann, A., Mickoleit, M., Weber, M., Huisken, J. Multilayer mounting enables long-term imaging of zebrafish development in a light sheet microscope. Development. 139, 3242-3247 (2012).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiThis article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır