
JoVE 비디오를 활용하시려면 도서관을 통한 기관 구독이 필요합니다. 전체 비디오를 보시려면 로그인하거나 무료 트라이얼을 시작하세요.
Method Article
얼룩말피에서 척추동물 세분화를 연구하는 3-D 꼬리 이식 문화
요약
여기서, 우리는 척추 동물 분할의 라이브 연구를 가능하게, 제브라피시 후방 체축의 3-D 조직 배양에 대한 프로토콜을 제시한다. 이 각성 모델은 축 신장, 형태항원 변형, 세포외 분해능 조직 수준 라이브 이미징에 대한 제어를 제공합니다.
초록
척추 동물 배아는 반복적인 솜투, 척추, 근육 및 피부의 선구자로서 주요 신체 축을 패턴으로 합니다. 소마이트는 배아의 꼬리 끝이 후방으로 길게 하는 동안 전소성 중구(PSM)로부터 점진적으로 분과한다. 소미트는 정기적인 주기율과 크기가 크기가 있는 형태를 이룹니다. Zebrafish는 유전적으로 유전적으로 견딜 수 있고 살아있는 화상 진찰을 허용하는 투명한 배아를 가지고 있기 때문에 인기있는 모델 유기체입니다. 그럼에도 불구하고, somitogenesis 동안, 물고기 배아는 크고 둥근 노른자 주위에 싸여 있습니다. 이 기하학은 특히 가까운 객관적인 작업 거리를 필요로 하는 더 높은 해상도에서 제브라피시 배아에 있는 PSM 조직의 살아있는 화상 진찰을 제한합니다. 여기서, 우리는 제브라피시 꼬리 의 살아있는 화상 진찰을 위한 평평한 3-D 조직 배양 방법을 제시합니다. 꼬리 는 축 신장의 비례 감속과 로스트로코달 소미트 길이의 단축을 표시하여 그대로 배아를 모방. 우리는 또한 절제 된 문화를 통해 축 신장 속도를 지연 시킬 수 있습니다. 이를 통해 처음으로 축 신장의 기계성 입력에서 신호 그라데이션의 화학적 입력을 풀 수 있습니다. 향후 연구에서, 이 방법은 약물 침투 우려 없이 시간 조절 된 제약 동요 또는 척추 동물 세분화의 검열을 허용 하기 위해 미세 유체 설정과 결합 될 수 있습니다.
서문
유기체의 메타메황 세분화는 자연에서 널리 사용됩니다. 반복된 구조물은 바디 계획1에서척추, 근육, 신경, 혈관, 사지 및 잎과 같은 측면 기관의 기능에 필수적입니다. 축 대칭의 이러한 생리적 및 기하학적 제약의 결과로, 아네리드, 절지동물 및 그들의 배아 조직의 초다트 전시 세분화와 같은 Bilateria의 대부분의 필라(예: 자궁 절제술, 중음근) antero-후방.
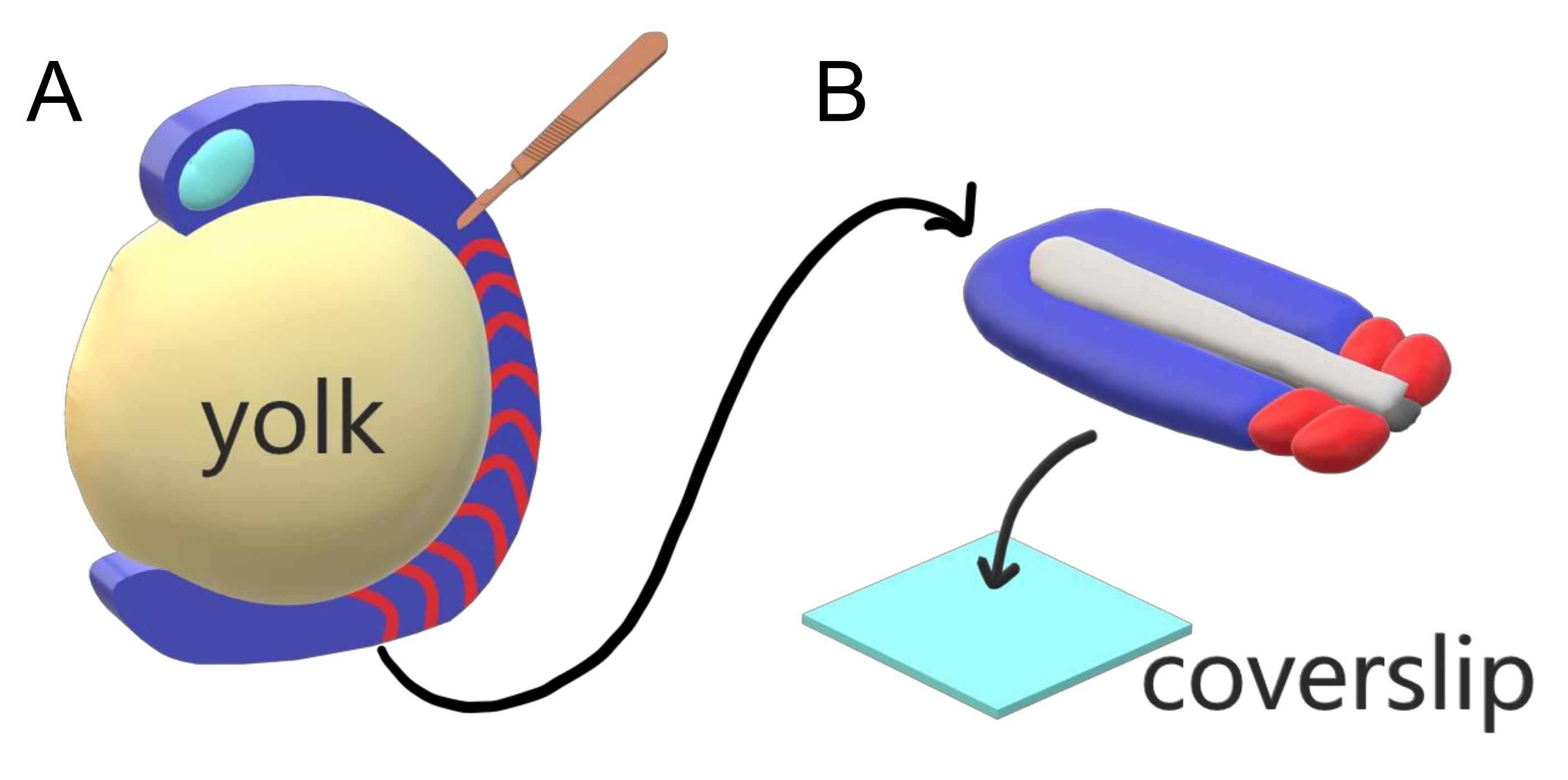
척추 동물 배아는 주 체축을 따라 종별 간격, 개수 및 크기 분포를 가진 솜투로 순차적으로 분사합니다. 종 내의 개별 적인 태아 중 이러한 견고함에도 불구 하 고, somite 세분화는 척추 동물 종 사이 다재 다능 한. 세분화는 시간 간격의 광대 한 정권에서 발생 (제브라피시에서 25 분에서 5 인간의 시간), 크기 (제브라피시의 꼬리 솜톤의 ~ 20 μm에서 ~200 μm 마우스의 트렁크 솜트에서 ~ 200 μm) 및 카운트 (제브라피시 에서 32에서 ~300 옥수수 뱀에 ~ 300)2. 더 흥미롭게도, 물고기 배아는 세분화 간격과 축 신장 속도 모두에 대해 보상하여 적절한 크기 분포로 소크를 그대로 유지하면서 광범위한 온도 (~20.5 °C에서 얼룩말 피시의 경우 34 °C까지)에서 개발할 수 있습니다. 이러한 흥미로운 특징 외에도 Zebrafish는 외부, 동기 및 형제 배아의 풍요롭고 투명한 발달뿐만 아니라 접근 가능한 유전 도구로 인해 척추 동물의 세분화를 연구하는 유용한 모델 유기체로 유지됩니다. 현미경 관점에서 불리하게, 텔레오스트 배아는 부피가 큰 구형 노른자에서 발전하고, 그 주위의 가스루 조직을 스트레칭하고 반올림한다(그림1A). 이 기사에서는 제브라피시 꼬리에 대한 평평한 3D 조직 식배 문화를 제시합니다. 이 절제 시스템은 노른자 질량의 구형 제약을 우회하여 소미트 패터닝을 위해 물고기 배아의 고해상도 라이브 이미징에 접근할 수 있게 합니다.

그림 1: 제브라피시 배아용 슬라이드 챔버 엑스플랜트 시스템. (A)제브라피시 배아는 배아 조직(blue)의 투명도와 같은 살아있는 이미징에 장점이 있지만, 부피가 큰 구형 노른자 질량(yellow)을 중심으로 조직이 형성되어 그대로 배아에서 거의 객관적이고 고해상도영상을 방지한다. 꼬리 각질은 소크라이트(빨강)의 조직 전방에서 절단된 미세 수술 칼(brown)으로 시작하여 노른자와 의 국경에서 후불로 계속해낼 수 있다. (B)해부된 꼬리 이출은 커버슬립(라이트 블루)에 등등불구해 배치될 수 있다; 상단에 신경 조직 (밝은 회색)을 유지하고 하단에 notochord (어두운 회색)를 유지합니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
프로토콜
이 프로토콜은 1 일 보다 더 젊은 살아있는 척추 동물 태아의 사용을 포함 수정 후 수정. 모든 동물 실험은 신시내티 아동 병원 의료 센터의 윤리적 지침에 따라 수행되었다; 동물 프로토콜은 기관 동물 관리 및 사용 위원회 (프로토콜 # 2017-0048)에 의해 검토되고 승인되었습니다.
1. 배아 수집
- 배아 수집 일 전날 밤 교차 탱크에 제브라피 쌍을 설정합니다. 배아 발달의 정밀한 준비 제어를 위해 짝짓기 쌍 사이의 장벽을 사용합니다.
- 선호하는 산란 시간 전에 장벽을 높이고 100mm 페트리 접시에서 15분 이내에 계란을 수집합니다.
- 페트리 접시의 깨끗한 파편. 하나의 클러치에서 50개 이상의 배아를 수집하는 경우 클러치를 여러 페트리 접시로 나눕히면 그에 따라 분리합니다.
- 어필 스테이지(5시간 후 수정)에 도달할 때까지 28°C의 물고기 계통 물에서 배아를 배양합니다. E3와 같은 표준화된 배아 성장 배지는 3.2단계까지 수족관 시스템 물 대신에도 사용할 수 있다.
- 스테레오스코프 하에서 수정되지 않은 계란을 제거하고 배아를 하룻밤 동안 23.5 °C 인큐베이터로 이동시다(O/N). 배아는 수집 일 다음 날 아침에 8-10 somites 단계에 있어야합니다.
2. 공구 준비
- 미세 수술 칼 날, 바늘 팁 (조직의 해부에 사용), 유리 파스퇴르 파이펫을 100 % 에탄올 (EtOH) 및 화재 유약에 담그고 살균하십시오.
- 25mm x 75mm 현미경 슬라이드에 투명 테이프(~100-120 μm 두께)의 두 층을 사용하십시오. 각 슬라이드 테이프의 중앙에 18mm x 18mm 정사각형 우물을 잘라 메스로 덮습니다.
- 준비된 슬라이드 챔버를 70% EtOH로 닦으십시오. 이 우물은 중간 ~ 40 μL을 보유합니다.
3. 샘플 준비
- 스테레오스코프 아래 두 개의 바늘 주사기의 끝을 사용하여 배아를 데초리오네이트. 배아를 물고기 시스템 물로 분리된 페트리 접시에 넣고 헹구는 것입니다.
- 불유리 멸균 유리 파스퇴피를 사용하여, 해부 배지를 함유한 6cm 페트리 접시에 배아를 전달합니다(레보비츠-15 세포 배양 배지 는 Phenol Red 없이 L-글루타민을 함유하고, 0.8 mMM CaCl2 및 1× 항생제 항mycotic 용액).
참고: 이 단계 다음의 모든 전송에 멸균 유리 파이펫을 계속 사용합니다.- 해부 중에 폴리스티렌 칩을 피하기 위해 유리 페트리 접시를 사용하여 절차를 내포하십시오.
- 슬라이드 챔버에 조직 성장 배지(해부 매체 및 10% FBS)의 50 μL을 넣습니다.
- 힌드뇌 근처의 노른자 조직 교차로에서 바늘 팁으로 입체 범위 아래해부를 위한 배아를 안정화시하십시오.
- 배아 조직을 바늘로 안정적으로 유지시키고, 45°에서 블레이드를 가진 미세 수술용 칼을 사용하여 조직 전방을 힌뇌로 잘라내고 전방에서 시작하여 꼬리 싹을 향해 이동하는 배아 조직에서 노른자를 벗겨낸다(도1A).
참고 : 노른자를 청소하는 동안 피부 조직을 잃지 않도록주의하십시오. 피부는 해부 도중 배아의 주위에 측면 단층 탄성 조직으로 쉽게 벗겨질 것입니다, 그래서 인식하기 쉽습니다. - 노른자는 배아 몸에서 완전히 제거되면 테일 버드에서 측면 피부 조직을 잘라냅니다. 마지막 형성 된 3-4 솜릿을 그대로 유지, 밖으로 더 전방 조직을 잘라 (전체 축 절제).
- 노른자는 이 절차에서 주로 그대로 벗겨져야 합니다. 파열 된 노른자의 경우, 노른자의 중요한 과립은 조직의 복부 표면에 부착 유지 될 수 있습니다. 그렇다면 속눈썹 도구를 사용하여 남은 노른자 과립을 부드럽게 청소하십시오.
참고: 절제의 측면에 피부 조직의 불균형은 조직이 직선 성장 방향을 유지하는 것을 허용하지 않을 것입니다. 대신 간구는 더 뻗은 피부의 측면을 향해 구부러집니다. 이 불균형은 미세 수술 칼의 도움으로 피부 층을 파열하여 입체 범위하에서 교정 될 수있다. - 피부가 없는 이장의 경우, 바늘로 피부 층의 끝을 누르고 미세 수술 칼로 짜는 조직을 벗겨냅니다. 이 이 흥분은 문화에서 그들의 바디 축을 길게 하지 않습니다.
- 전체 축 이외에도 이 단계에서 대체 축을 만들 수 있습니다. 예를 들어, 미세 수술 칼 (전체 PSM 이출)을 사용하여 이미 세분화 된 솜투를 해부하거나 PSM을 절반 의 anteroposterior (반 PSM 이출)로 해부하십시오. 그러한 대체 구역의 적용은 5.1항을 참조하십시오.
- 노른자는 이 절차에서 주로 그대로 벗겨져야 합니다. 파열 된 노른자의 경우, 노른자의 중요한 과립은 조직의 복부 표면에 부착 유지 될 수 있습니다. 그렇다면 속눈썹 도구를 사용하여 남은 노른자 과립을 부드럽게 청소하십시오.
- 해부된 해동을 이미징이 수행되는 22mm x 22mm 커버슬립으로 즉시 전달합니다.
- 등소벤탈 축에 평평하게 익히면, 커버슬립(도1B)을터치하는 복부 측. 20 μL 여과 팁 파이펫을 사용하여 조직 주위의 과잉 매체를 부드럽게 제거합니다.
참고: 해부된 각출물의 지연된 전달은 노른자의 기하학적 제약에서 벗어나므로 조직의 변형을 초래합니다.
- 등소벤탈 축에 평평하게 익히면, 커버슬립(도1B)을터치하는 복부 측. 20 μL 여과 팁 파이펫을 사용하여 조직 주위의 과잉 매체를 부드럽게 제거합니다.
- 성장 중간 채워진 슬라이드 챔버 를 통해 explant와 신속 하고 신중하게 커버 슬립을 뒤집습니다.
- 거품 형성을 방지하기 위해, 테이프 챔버에 사각형 커버 슬립의 측면을 배치하고 부드럽게 다른 쪽을 해제합니다. 이 단계에서 각기를 이동/변형하지 않도록 주의하십시오.
- 실험실 조직에 슬라이드 챔버를 눌러 챔버에서 출혈 과잉 미디어를 부드럽게 제거합니다. 커버슬립은 밀봉 없이 액체 매체의 표면 장력으로 인해 라이브 이미징을 위해 슬라이드 챔버에 안정적으로 앉을 것입니다.
- 장기 배양 (>6 시간)의 경우 더 큰 챔버를 사용합니다. 22mm x 50mm 직사각형 커버립과 슬라이드에 테이프 레이어의 두 개의 평행 차선이 이러한 경우에 활용할 수 있습니다. ~1mm 너비의 간격은 성장 매체로의 공기 접근을 용이하게 하기 위해 두 개의 테이프 레인 사이에 남을 수 있습니다.
- 3.3-3.8 단계를 반복하여 더 많은 절제를 준비합니다. 준비된 이종은 평균 ~30 μm/h 속도로 A-P 체형을 길게 하고 25°C(그림2A,비디오 1)에서~40분 간격으로 소크를 분단합니다.
- 비 장간 이식의 경우, 실험실 조직에 과잉 매체를 빨아 동안 3.8.2 단계에서 샘플을 들고 슬라이드의 측면에 부드러운 압력을 적용합니다. 또는, 이식은 단일 테이프 층 슬라이드 챔버에서 배양 될 수있다. 또한, 타입 I 콜라겐으로 슬라이드 챔버 표면을 화학적으로 활성화하면 비 길쭉한이장(도 2B,비디오 2)이생성됩니다.
- 1시간 동안 실온에서 15-20mL의 미리 희석된 콜라겐 용액으로 슬라이드 챔버를 완전히 덮어 제1형 I 콜라겐으로 챔버의 코팅을 수행한다. 이 프로토콜에 라미나르 플로우 후드를 사용하여 멸균을 유지합니다. 끝에 해부 매체로 챔버를 조심스럽게 헹구는 다.
- 15개 이상의 소미트 스테이지의 경우, 꼬리 절제 조직을 평평한(dorsoventral) 마운트 대신 측면으로 장착한다(비디오3). 근육 경련을 방지하기 위해, 마취제로 문화 매체에 0.004% 트리카인 용액을 포함한다3.
- 비 장간 이식의 경우, 실험실 조직에 과잉 매체를 빨아 동안 3.8.2 단계에서 샘플을 들고 슬라이드의 측면에 부드러운 압력을 적용합니다. 또는, 이식은 단일 테이프 층 슬라이드 챔버에서 배양 될 수있다. 또한, 타입 I 콜라겐으로 슬라이드 챔버 표면을 화학적으로 활성화하면 비 길쭉한이장(도 2B,비디오 2)이생성됩니다.
4. 라이브 이미지 수집
- 이미지 샘플은 솜라이트 세분화 크기와 기간의 광화상 광 이미징을 위한 해부 범위 또는 형질 기자 어항을 이용한 구조화된 조명/공초점/광시트 현미경으로 샘플링합니다.
- 적어도 15 분 동안 이미징 실온과 함께 조직의 온도를 평형화.
- 보다 정밀한 온도 제어를 위해 반전된 현미경에 장착된 상용 온도 제어 시스템을 사용하십시오.
- 생물학적 관심 과정에 따라 이미지 수집 프레임 간격을 2-10분으로 설정합니다.
참고: Zebrafish somite 분할은 전체 배아에서 30°C내지 21.5°C의 실행 가능한 온도에 대해 20-55분 사이의 빠른 공정입니다. 엑셀은 전체 배아보다 ~30% 느리게 줄어들며 세그먼트를 길게 합니다.- 살아있는 조직에 가능한 광독성을 피하기 위해 채널 인수 세트 사이에 충분한 지연을 남기는 데주의를 기울이십시오. 이미징 지속 시간의 절반 이상을 외리 빔에 노출시키고 가능한 한 빔 강도를 낮춥시게 하지 마십시오.
참고: 반응성 산소 종(ROS)의 축적은 일반적으로 살아있는 샘플4에서광독성의 주요 원인이다. ROS 스캐빈거로서의 아스코르브산은 ROS 활성을 완충하고 광독성을 완화하기 위해 4m 농도의 성장 배지로 보충될 수 있다. 광독성의 부작용은 라이브 이미징 중에 알아차리기 어려울 수 있습니다. 꼬리 이식은 미토알 검거, 조직 발달 장애(즉, 소미트, 꼬리 신장 형성) 및 붕괴 조직과 같은 광독성의 일부 시각적 마커가 쉽게 발견되기 때문에 이러한 측면에서 유리하다. 자세한 내용은 제공된 참조4를 참조하십시오.
- 살아있는 조직에 가능한 광독성을 피하기 위해 채널 인수 세트 사이에 충분한 지연을 남기는 데주의를 기울이십시오. 이미징 지속 시간의 절반 이상을 외리 빔에 노출시키고 가능한 한 빔 강도를 낮춥시게 하지 마십시오.
- 단일 세포 단계 RNA 주입 된 배아를 사용하여 세포 분해수준에서 분할 및 분석될 4 차원 이미지를 습득하십시오.
- 시험관 내 전사막 및 핵 형광 기자 마커 플라스미드(pCS-membrane-ceruleanFP(addgene plasmid #53749) 또는 pCS-memb-mCherry(추가)에서 300pg의 RNA를 사용하십시오( 추가 pCS2+ H2B-mTagBFP2(애드진 플라스미드 #99267) 또는 pCS2+ H2B-TagRFP-T(Addgene plasmid #99271)와 함께 #53750. 세포막과 핵 마커가 있는 샘플 영화의 경우 비디오 4를참조하십시오.
참고: PSM 조직의 평균 세포 크기는 직경약 ~5μm이며, 그 중 핵은 ~3-4 μm을 포함한다. ~0.5 μm의 픽셀 크기와 ~1 μm의 z 단면은 적절한 세포 세분화를 위해 기록되어야 합니다.
- 시험관 내 전사막 및 핵 형광 기자 마커 플라스미드(pCS-membrane-ceruleanFP(addgene plasmid #53749) 또는 pCS-memb-mCherry(추가)에서 300pg의 RNA를 사용하십시오( 추가 pCS2+ H2B-mTagBFP2(애드진 플라스미드 #99267) 또는 pCS2+ H2B-TagRFP-T(Addgene plasmid #99271)와 함께 #53750. 세포막과 핵 마커가 있는 샘플 영화의 경우 비디오 4를참조하십시오.
5. 꼬리 의 면역 염색
참고: 다양한 해부 시나리오(길쭉한, 비길쭉한, 꼬리 싹 해부, 반 PSM 등)를 거쳐 자란 조직은 5개의 평평한 꼬리이출로 슬라이드 챔버에서 회수하여 관심 있는 단백질의 정제를 더욱 면역화할 수 있다. 여기서, 우리는 FGF 신호 그라데이션 판독으로 이종의 디인세포 신호 조절 키나아제 (ppERK) 염색에 사용되는 프로토콜을 제시한다.
- 원하는 단계까지 소암이 형성된 후, 커버슬립을 들어올리지 않고 슬라이드 챔버의 구석쪽으로 조심스럽게 이동합니다.
- 유리 파스퇴르 파이펫에서 보충 해부 배지의 ~100 μL의 도움으로, 슬라이드에서 이출을 복구하고 64 웰 세포 배양 판으로 전송.
참고: 이 단계를 시작으로 고정 시료에 대한 별도의 유리 파이펫이 있는 해부 범위 하에서 모든 솔루션 교체를 수행할 수 있습니다. 이것은 우물에서 각제 조직을 잃거나 그 사이에 전송하지 않도록합니다. - 모든 이양을 옮김한 후, 우물에서 과도한 배지를 하나씩 빨아내고 PBS(PFA)에 4%의 파라포름알데히드100 μL을 각 우물에 넣습니다.
주의: PFA는 발암 효과를 가진 독성 용액입니다. 적절한 PPE를 처리하는 동안 사용해야 합니다. - 셰이커에서 1 시간 동안 실온에서 64 웰 플레이트에 담기 고정하십시오.
- 조직 이질은 전체 배아보다 변형에 더 민감합니다. 셰이커 속도를 그에 따라 조정합니다.
- PBS-Tw의 150 μL(PBS에서 0.1% Tween20)로 고정을 세번 씻으시면 세 번 씻으십니다. 특정 "PFA 폐기물"용기에서 첫 번째 세척을 수집합니다.
- 탈수는 매번 100% 메탄올(MeOH)으로 용액의 ~40 μL을 대체하여 4×5분 단계로 이내합니다.
주의: MeOH는 휘발성 및 인화성 독성 화학 물질입니다. 통풍이 잘 되는 공간에서 작업하고 적절한 PPE를 사용하여 처리를 하십시오. - 탈수의 마지막 단계로, 우물에서 모든 용액을 제거하고 MeOH의 100 μL로 대체하십시오. -20°C에서 15분 동안 배양합니다.
참고: 특정 "MeOH 폐기물" 용기를 사용하여 5.11 단계까지 솔루션을 수집합니다. - MeOH 50 μL을 추가하고 실온에서 5 분 동안 흔들어 주세요.
- PBS-T(PBS에서 0.1% 트리톤-X 100)로 매번 용액의 ~40 μL을 교체하여 4×5분 단계로 재수화합니다. 특정 "MeOH 폐기물" 용기를 사용하여 솔루션을 수집합니다.
- 재수화의 마지막 단계로, 우물에서 모든 용액을 제거하고 PBS-T의 100 μL로 대체합니다.
- 조직 과구화치료의 경우 PBS에서 1.5% 트리톤-X 100으로 분해하여 셰이커의 실온에서 20분 동안 이질합니다.
- MAB-D-T(0.1% 트리톤-X 100 세제, 1% 디메틸 설산화물(DMSO)로 시료를 150m NaCl 100mM MM 남성산 완충 pH 7.5) 3×5분으로 세척한다.
주의: DMSO는 인화성 및 독성 돌연변이원입니다. 적절한 PPE를 처리하는 동안 사용해야 합니다. - 100 μL/웰 혈청 차단 용액(MAB-D-T의 태아 소 세럼 2%)에서 실온에서 2시간 동안 배양합니다.
- 모든 차단 용액을 50-100 μL/잘 1차 항체 용액(혈청 블록에서 ppERK에 대한 단일 클론 마우스 항체의 1:1000 희석)으로 교체하십시오. 셰이커에 4 °C에서 샘플 O / N (>16 h)를 배양.
- MAB-D-T 5×5분으로 1차 항체 용액을 세척합니다.
- 이차 항체 용액(Alexa Fluor 597 염소 안티마우스 IgG2b(1:200) 및 MAB-D-T의 Hoechst 33342(1:5000)에서 샘플을 증식하거나 실온에서 3시간 동안 배양한다.
참고: 이 단계에서 시작하여 알루미늄 호일로 64웰 플레이트를 덮어 이차 항체 처리 시료의 광 노출을 방지합니다.
주의: Hoechst 33342는 잠재적인 발암 물질입니다. 적절한 PPE를 처리하는 동안 사용해야 합니다. - PBS-Tw 3×5분으로 이차 항체 용액을 세척합니다.
- 실온에서 15분 동안 PFA로 샘플을 수정합니다.
- PBS-Tw로 고정을 세척하고 60 % 글리세롤 내에서 샘플을 평형화하십시오. 매니큐어와 60% 글리세롤현미경 슬라이드에 이식합니다. 대표적인 면역염색 결과는 그림 3을참조하십시오.
결과
이 프로토콜은 살아있는 제브라피시 꼬리 의 평평한 기하학적 배양이 가능합니다. 조직 배양은 전체 배아에 비해 세 가지 주요 장점을 제시합니다: 1) 축 신장 속도의 제어, 2) 간단한 해부에 의한 다양한 신호(morphogen) 소스에 대한 제어, 3) 거의 목표, 높은 배율 및 높은 NA 라이브 이미징.
화학적으로 처리되지 않은 슬라이드 챔버는 꼬리가 그 아래 조직 주위에 감싸는 피부 자궁...
토론
이 문서는 우리가 개발하고 제브라피시 배아를 위해 최근에 사용 된 조직 배양 각기 기술의 상세한 프로토콜을 제시합니다. 우리의 기술은 병아리8과 제브라피시9,10,11 모델 유기체의 이전 각성 방법에 기반을 두고 있습니다. 이 프로토콜로 준비된 꼬리 이식은 간단한 슬라이드 챔버에서 >12h...
공개
저자는 아무 것도 공개하고 이해 상충을 선언할 수 없습니다.
감사의 말
AECOM Zebrafish 코어 시설과 신시내티 아동 수의학 서비스, 기술 지원을 위한 신시내티 어린이 이미징 코어, 디다르 사파로프(Didar Saparov) 비디오 제작 지원, 원고 편집을 위한 한나 씨월에게 감사드립니다. 이 간행물에서 보고된 연구는 E.M.Ö에 상 번호 R35GM140805에 국가의 건강 연구소의 일반 의학 과학의 국가 학회에 의해 지원되었습니다. 이 내용은 전적으로 저자의 책임이며 반드시 국립 보건원의 공식 견해를 나타내는 것은 아닙니다.
자료
| Name | Company | Catalog Number | Comments |
| 1 mL Sub-Q Syringe with PrecisionGlide Needle | Becton, Dickinson and Co. | REF 309597 | for dechorionating embryos and manipulations |
| 200 Proof Ethanol, Anhydrous | Decon Labs | 2701 | for immunostaining |
| Antibiotic Antimycotic Solution (100×) | Sigma-Aldrich | A5955 | for tissue dissection media |
| Calcium Chloride Anhydrous, Powder | Sigma-Aldrich | 499609 | for tissue dissection media |
| Dimethylsulfoxide | Sigma-Aldrich | D5879 | for immunostaining |
| Disposable Scalpel, #10 Stainless Steel | Integra-Miltex | MIL4-411 | for preparing tape slide wells |
| Ethyl 3-aminobenzoate methanesulfonate salt (Tricaine) | Sigma-Aldrich | 886-86-2 | (optional) for anesthesizing tissues older than 20 somites stage |
| Fetal Bovine Serum (FBS) | ThermoFisher | A3160601 | additional for tissue culture media |
| Goat anti-Mouse IgG2b, Alexa Fluor 594 | Invitrogen | Cat#A-21145; RRID: AB_2535781 | secondary antibody for immunostaining |
| L-15 Medium with L-Glutamine w/o Phenol Red | GIBCO | 21083-027 | for tissue dissection media |
| Methanol | Sigma-Aldrich | 179337 | for immunostaining |
| Microsurgical Corneal Knife 2.85 mm Angled Tip Double Bevel Blade | Surgical Specialties | 72-2863 | for tissue dissection |
| Mouse monoclonal anti-ppERK | Sigma-Aldrich | Cat#M8159; RRID:AB_477245 | for ppERK immunostaining |
| NucRed Live 647 ReadyProbes Reagent | Invitrogen | R37106 | (optional) for live staining of cell nuclei |
| Paraformaldehyde Powder, 95% | Sigma-Aldrich | 158127 | for fixation of samples for immunostaining |
| Rat Tail Collagen Coating Solution | Sigma-Aldrich | 122-20 | (optional) for chemically activating slide chambers |
| Stage Top Incubator | Tokai Hit | tokai-hit-stxg | (optional) for temperature control during live imaging |
| Transparent Tape 3/4'' | Scotch | S-9782 | for preparing tape slide wells |
| Triton X-100 | Sigma-Aldrich | X100 | for immunostaining |
| Tween 20 | Sigma-Aldrich | P1379 | for immunostaining |
| Zebrafish: Tg(actb2:2xMCP-NLS-EGFP) | Campbell et al., 2015 | ZFIN: ZDB-TGCONSTRCT-150624-4 | transgenic fish with nuclear localized EGFP |
| Zebrafish: Tg(Ola.Actb:Hsa.HRAS-EGFP) | Cooper et al., 2005 | ZFIN: ZDB-TGCONSTRCT-070117-75 | transgenic fish with cell membrane localized EGFP |
참고문헌
- Assheton, R. . Growth in length: Embryological Essays. , (1916).
- Gomez, C., et al. Control of segment number in vertebrate embryos. Nature. 454 (7202), 335-339 (2008).
- Westerfield, M. . The Zebrafish Book: a guide for the laboratory use of zebrafish (Danio rerio), 3rd edition. , (1995).
- Icha, J., Weber, M., Waters, J. C., Norden, C. Phototoxicity in live fluorescence microscopy, and how to avoid it. BioEssays. 39 (1700003), (2017).
- Simsek, M. F., Ozbudak, E. M. Spatial fold change of Fgf signaling encodes positional information for segmental determination in zebrafish. Cell Reports. 24 (1), 66-78 (2018).
- Dubrulle, J., Pourquié, O. fgf8 mRNA decay establishes a gradient that couples axial elongation to patterning in the vertebrate embryo. Nature. 427 (6973), 419-422 (2004).
- Diez del Corral, R., et al. Opposing FGF and Retinoid Pathways Control Ventral Neural Pattern, Neuronal Differentiation, and Segmentation during Body Axis Extension. Neuron. 40 (1), 65-79 (2003).
- Stern, H. M., Hauschka, S. D. Neural tube and notochord promote in vitro myogenesis in single somite explants. Developmental Biology. 167 (1), 87-103 (1995).
- Langenberg, T., Brand, M., Cooper, M. S. Imaging brain development and organogenesis in zebrafish using immobilized embryonic explants. Developmental Dynamics. 228 (3), 464-474 (2003).
- Picker, A., Roellig, D., Pourquié, O., Oates, A. C., Brand, M. Tissue micromanipulation in zebrafish embryos. Methods in molecular biology. 546 (11), 153-172 (2009).
- Manning, A. J., Kimelman, D. Tbx16 and Msgn1 are required to establish directional cell migration of zebrafish mesodermal progenitors. Developmental Biology. 406 (2), 172-185 (2015).
- Kimmel, C. B., Ballard, W. W., Kimmel, S. R., Ullmann, B., Schilling, T. F. Stages of embryonic development of the zebrafish. Developmental Dynamics. 203 (3), 253-310 (1995).
- Kaufmann, A., Mickoleit, M., Weber, M., Huisken, J. Multilayer mounting enables long-term imaging of zebrafish development in a light sheet microscope. Development. 139, 3242-3247 (2012).
재인쇄 및 허가
JoVE'article의 텍스트 или 그림을 다시 사용하시려면 허가 살펴보기
허가 살펴보기더 많은 기사 탐색
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. 판권 소유