
É necessária uma assinatura da JoVE para visualizar este conteúdo. Faça login ou comece sua avaliação gratuita.
Method Article
Uma cultura de explanta de cauda 3D para estudar segmentação de vertebrados em zebrafish
Neste Artigo
Resumo
Aqui, apresentamos o protocolo para a cultura tecidual 3D do eixo corporal posterior do zebrafish, possibilitando o estudo ao vivo da segmentação de vertebrados. Este modelo de explant fornece controle sobre o alongamento do eixo, alteração de fontes de morfógeno e imagem viva de nível de tecido de resolução subcelular.
Resumo
Embriões vertebrados padronizam seu eixo corporal principal como somites repetitivos, os precursores de vértebras, músculos e pele. As somitas se segmentam progressivamente do mesoderme presomótico (PSM) à medida que a extremidade traseira do embrião alonga posteriormente. As somitas formam-se com periodicidade regular e escala em tamanho. O zebrafish é um organismo modelo popular, pois é geneticamente tratável e possui embriões transparentes que permitem imagens vivas. No entanto, durante a somitogênese, os embriões de peixe são enrolados em torno de uma grande gema arredondada. Esta geometria limita a imagem viva do tecido PSM em embriões de zebrafish, particularmente em resoluções mais altas que requerem uma distância de trabalho objetiva próxima. Aqui, apresentamos um método de cultura de tecido 3D achatado para imagens vivas de explants de cauda de zebrafish. Explantas de cauda imitam embriões intactos exibindo uma desaceleração proporcional do alongamento do eixo e encurtamento dos comprimentos de somite rostrocaudal. Ainda podemos parar a velocidade de alongamento do eixo através da cultura explant. Isso, pela primeira vez, nos permite desembaraçar a entrada química de gradientes de sinalização da entrada mecanicista do alongamento axial. Em estudos futuros, este método pode ser combinado com uma configuração microfluida para permitir perturbações farmacêuticas controladas pelo tempo ou triagem da segmentação de vertebrados sem qualquer preocupação com a penetração de medicamentos.
Introdução
A segmentação metamérica de organismos é amplamente utilizada na natureza. Estruturas repetidas são essenciais para a funcionalidade de órgãos laterais como vértebras, músculos, nervos, vasos, membros ou folhas em um plano corporal1. Como resultado de tais restrições fisiológicas e geométricas da simetria axial, a maioria da phyla de Bilateria- como annelids, artrópodes e cordatas-exposição segmentação de seus tecidos embrionários (por exemplo, ectoderme, mesoderme) antero-posteriormente.
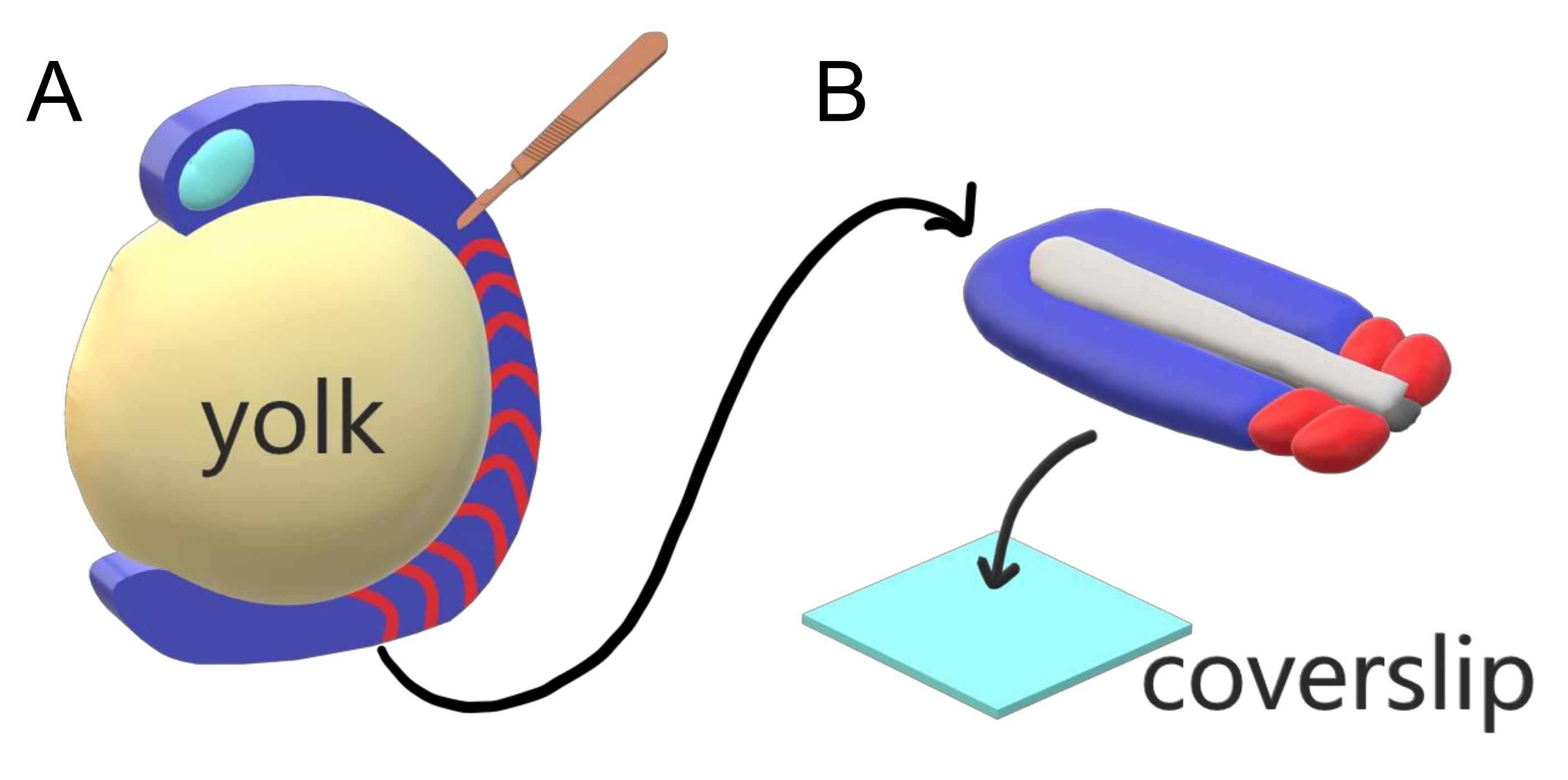
Embriões vertebrados segmentam sequencialmente seu mesoderme paraxial ao longo do eixo principal do corpo em somitas com intervalos específicos de espécies, contagens e distribuições de tamanho. Apesar de tal robustez entre embriões individuais dentro de uma espécie, a segmentação de somite é versátil entre espécies de vertebrados. A segmentação acontece em um vasto regime de intervalos de tempo (de 25 minutos em zebrafish a 5 h em humanos), tamanhos (de ~20 μm em somitas de cauda de zebrafish a ~200 μm em somitas tronco de camundongos) e conta (de 32 em zebrafish a ~300 em cobras de milho)2. O mais interessante é que os embriões de peixe podem desenvolver-se em uma ampla gama de temperaturas (de ~20,5 °C até 34 °C para zebrafish) mantendo seus somitas intactos com distribuições de tamanho adequado, compensando tanto os intervalos de segmentação quanto as velocidades de alongamento axial. Além dessas características interessantes, o zebrafish permanece como um organismo modelo útil para estudar a segmentação em vertebrados devido ao desenvolvimento externo, síncrono e transparente de uma plenitude de embriões irmãos, bem como suas ferramentas genéticas acessíveis. Adversamente a partir de uma perspectiva de microscopia, os embriões teleost desenvolvem-se em uma gema esférica volumosa, esticando e arredondando o tecido gastrulating ao seu redor(Figura 1A). Neste artigo, apresentamos uma cultura de explante de tecido 3D achatada para caudas de zebrafish. Este sistema de explant contorna as restrições esféricas da massa de gema, permitindo o acesso a imagens vivas de alta resolução de embriões de peixes para padronização de somite.

Figura 1: Sistema de Explantação de Câmara de Slides para Embriões de Zebrafish. (A) Os embriões de zebrafish têm vantagens para imagens vivas, como a transparência do tecido embrionário gastrulating (azul), mas o tecido se forma em torno de uma massa de gema esférica volumosa (amarela) que previne imagens quase objetivas e de alta resolução em embriões intactos. Explantas de cauda podem ser dissecadas começando com uma faca microcirúrgica (marrom) cortada do tecido anterior de somites (vermelho) e continuando na borda com a gema posteriormente. (B) Explantas traseiras dissecadas podem ser colocadas em um deslizamento (azul claro) dorsoventrally; mantendo tecido neural (cinza claro) em cima e notochord (cinza escuro) na parte inferior. Clique aqui para ver uma versão maior desta figura.
{kind=link}
Protocolo
Este protocolo envolve o uso de embriões vertebrados vivos com menos de 1 dia de fertilização. Todos os experimentos em animais foram realizados sob as diretrizes éticas do Centro Médico hospitalar infantil de Cincinnati; os protocolos animais foram revisados e aprovados pelo Comitê Institucional de Cuidados e Uso de Animais (Protocolo nº 2017-0048).
1. Coleção de embriões
- Coloque pares de zebrafish em tanques de travessia na noite anterior ao dia da coleta de embriões. Para um controle preciso do desenvolvimento de embriões, use barreiras entre pares de acasalamento.
- Levante barreiras antes do tempo de desova preferido e colete ovos dentro de 15 minutos em uma placa de Petri de 100 mm.
- Limpar detritos da placa de Petri. Se mais de 50 embriões forem coletados de uma única embreagem, divida a embreagem em várias placas de Petri em conformidade.
- Incubar embriões em água do sistema de peixes a 28 °C até atingirem 50% de estágio epiboly (5 horas após a fertilização). Um meio de crescimento padronizado de embriões como o E3 também pode ser usado em vez da água do sistema de aquário até o passo 3.2.
- Remova ovos não fertilizados sob um estereóscópio e mova os embriões para uma incubadora de 23,5 °C durante a noite (O/N). Os embriões devem estar em 8-10 somites estágio na manhã seguinte dia de coleta.
2. Preparação da ferramenta
- Esterilizar a lâmina da faca de microcirurgia, as pontas da agulha (usada para dissecção do tecido) e a pipeta pasteur de vidro, de molho em 100% etanol (EtOH) e vidros de fogo.
- Use duas camadas de fita transparente (~100-120 μm de espessura) em lâminas de microscópio de 25 mm x 75 mm. Corte ~18 mm x 18 mm de poços quadrados no centro da cobertura de fita de cada slide com um bisturi.
- Limpe as câmaras de slides preparadas com 70% de EtOH. Estes poços conterão ~40 μL de médio.
3. Preparação da amostra
- Embriões descorrionatos usando a ponta de duas seringas de agulha sob um estereoscópio. Transfira embriões para uma placa de Petri separada com água do sistema de peixes para enxaguar.
- Usando uma pipeta pasteur de vidro estéril com vidro de vidro, transfira embriões em uma placa de Petri de 6 cm contendo meio de dissecção (meio de cultura celular Leibovitz-15 com L-Glutamina sem Phenol Red, 0,8 mM CaCl2 e 1× solução antimicocítica antibiótico).
NOTA: Continue a usar uma pipeta de vidro esterilizada para todas as transferências após esta etapa.- Use uma placa de vidro Petri para o procedimento de explanagem para evitar lascas de poliestireno durante a dissecção.
- Coloque 50 μL de meio de crescimento tecidual (médio de dissecção e 10% FBS) na câmara de slides.
- Estabilize um embrião para dissecção sob o estereóscópio com uma ponta de agulha na intersecção de tecido de gema perto do cérebro traseiro.
- Mantendo o tecido embrionário estável com uma agulha, use a faca microcirúrgica com a lâmina presa a 45° para cortar o tecido anterior ao cérebro traseiro e descascar a gema do tecido embrionário que parte do anterior e movendo-se em direção ao broto da cauda(Figura 1A).
NOTA: Tenha cuidado para não perder o tecido da pele durante a limpeza da gema. A pele facilmente descascaria como um tecido elástico de camada única ao redor do embrião durante a dissecação, por isso é fácil de reconhecer. - Uma vez que a gema é totalmente removida do corpo embrionário, corte o tecido da pele flanqueando do farol. Mantendo intactas as últimas 3-4 somites, corte o tecido mais anterior (explantação de eixo completo).
- A gema deve sair principalmente intacta deste procedimento. No caso de uma gema rompida, grânulos significativos de gema podem permanecer ligados à superfície ventral do tecido. Se assim for, use uma ferramenta de chicote para limpar os grânulos restantes da gema suavemente.
NOTA: Um desequilíbrio dos tecidos da pele nas laterais laterais de uma explanta não permitiria que o tecido mantivesse uma direção de crescimento reto. A explanta, em vez disso, se curvará para o lado da pele mais esticada. Esse desequilíbrio pode ser corrigido sob o estereoscópio rompendo a camada da pele com o auxílio da faca microcirúrgica. - Para explantas sem pele, pressione uma ponta da camada de pele com agulha e retire o tecido com a faca microcirúrgica. Essas explantas não alongarão seu eixo corporal na cultura.
- Além das explantas do eixo completo, explants alternativos podem ser feitas nesta etapa. Por exemplo, dissecar somites já segmentados usando a faca microcirúrgica (explantas full-PSM) ou dissecar o PSM em sua metade anteroposterior (explantas meia-PSM). Consulte a seção 5.1 para uma aplicação dessas explantas alternativas.
- A gema deve sair principalmente intacta deste procedimento. No caso de uma gema rompida, grânulos significativos de gema podem permanecer ligados à superfície ventral do tecido. Se assim for, use uma ferramenta de chicote para limpar os grânulos restantes da gema suavemente.
- Transfira imediatamente a explanta dissecada para um deslizamento de cobertura de 22 mm x 22 mm no qual a imagem será realizada.
- Disponha a explanta plana no eixo dorsoventral, tora o lado ventral tocando o deslizamento(Figura 1B). Remova suavemente o excesso de mídia ao redor da explanta tecidual usando uma pipeta de ponta filtrada de 20 μL.
NOTA: A transferência retardada de explantas dissecadas para o deslizamento de cobertura resulta em deformações do tecido, pois é aliviada das restrições geométricas da gema.
- Disponha a explanta plana no eixo dorsoventral, tora o lado ventral tocando o deslizamento(Figura 1B). Remova suavemente o excesso de mídia ao redor da explanta tecidual usando uma pipeta de ponta filtrada de 20 μL.
- De forma rápida e cuidadosa, gire o deslizamento com a explanta sobre a câmara de slides preenchida com meio de crescimento.
- Para evitar a formação de bolhas, coloque um lado da tampa quadrada na câmara de fita e solte o outro lado suavemente. Tome cuidado para não mover/deformar a explanta nesta etapa.
- Remova suavemente o sangramento do excesso de mídia da câmara pressionando a câmara de slides em um tecido de laboratório. A mancha de cobertura sentará-se firmemente na câmara de slides para imagens ao vivo, devido à tensão superficial do meio líquido sem qualquer vedação.
- Para cultivo a longo prazo (>6 horas), use uma câmara maior. Tampas retangulares de 22 mm x 50 mm juntamente com duas faixas paralelas de camadas de fita em slides podem ser utilizadas nesses casos. Uma abertura de ~1 mm de largura pode ser deixada entre duas faixas de fita para facilitar o acesso aéreo ao meio de crescimento.
- Repita as etapas 3.3-3.8 para preparar mais explants. As explanações preparadas alongarão seu eixo corporal A-P com uma velocidade média de ~30 μm/h e segmentarão seus somites com intervalos de ~40 min a 25 °C(Figura 2A, Vídeo 1).
- Para explants não alongantes, aplique pressão suave nas laterais do slide segurando a amostra na etapa 3.8.2 enquanto suga a mídia em excesso em um tecido de laboratório. Alternativamente, as explantas podem ser cultivadas em câmaras de slides de camada de fita única. Além disso, ativar quimicamente a superfície da câmara de slides com colágeno tipo I resultará em explicações não alongadoras(Figura 2B, Vídeo 2).
- Realize o revestimento da câmara com colágeno tipo I com antecedência, cobrindo totalmente as câmaras de slides com 15-20 mL de solução de colágeno pré-diluída à temperatura ambiente por 1h. Use uma capa de fluxo laminar para este protocolo para manter a esterilidade. Enxágue cuidadosamente as câmaras com o meio de dissecção no final.
- Para embriões com mais de 15 anos, monte o tecido de explanta da cauda lateralmente em vez de uma montagem plana (dorsoventral)(Vídeo 3). Para evitar contrações musculares, inclua 0,004% de solução tricaina na mídia cultural como agente anestésico3.
- Para explants não alongantes, aplique pressão suave nas laterais do slide segurando a amostra na etapa 3.8.2 enquanto suga a mídia em excesso em um tecido de laboratório. Alternativamente, as explantas podem ser cultivadas em câmaras de slides de camada de fita única. Além disso, ativar quimicamente a superfície da câmara de slides com colágeno tipo I resultará em explicações não alongadoras(Figura 2B, Vídeo 2).
4. Aquisição de imagem ao vivo
- Amostras de imagem em um escopo de dissecção para imagens de luz transmitidas em campo largo de tamanhos e períodos de segmentação de somite, ou com iluminação estruturada/microscopia de folha de luz/folha de luz usando linhas de peixes repórteres transgênicos.
- Equilibre a temperatura das explantas teciduais com a temperatura ambiente de imagem por pelo menos 15 minutos.
- Para um controle de temperatura mais preciso, use um sistema comercial de controle de temperatura montado em um microscópio invertido.
- Ajuste os intervalos do quadro de aquisição de imagens para 2 - 10 min, dependendo do processo biológico de interesse.
NOTA: A segmentação de somite de zebrafish é um processo rápido, variando entre 20 - 55 min para temperaturas viáveis de 30 °C a 21,5 °C em embriões inteiros. Explantas alongarão e segmentarão ~30% mais lento que os embriões inteiros.- Preste atenção para deixar bastante atraso entre conjuntos de aquisições de canais para evitar possível fototoxicidade para o tecido vivo. Não exponha o tecido ao feixe de excitação por mais da metade da duração da imagem e reduza a intensidade do feixe o máximo possível.
NOTA: O acúmulo de espécies reativas de oxigênio (ROS) é geralmente a principal causa de fototoxicidade em amostras vivas4. O ácido ascórbico como um catador ros pode ser complementado ao meio de crescimento a 4 mM de concentração para tampão da atividade ROS e aliviar a fototoxicidade. Efeitos adversos da fototoxicidade podem ser difíceis de notar durante a imagem ao vivo. As explanações da cauda são vantajosas nesse aspecto, uma vez que alguns marcadores visuais de fototoxicidade, como a parada mitótica, o desenvolvimento de tecidos impedidos (ou seja, formação de somites, alongamento da cauda) e tecido desintegrador são mais fáceis de notar. Consulte o cet previamente à contratação do jornal4.
- Preste atenção para deixar bastante atraso entre conjuntos de aquisições de canais para evitar possível fototoxicidade para o tecido vivo. Não exponha o tecido ao feixe de excitação por mais da metade da duração da imagem e reduza a intensidade do feixe o máximo possível.
- Use embriões injetados de RNA em estágio celular único para adquirir imagens 4D destinadas a serem segmentadas e analisadas em nível de resolução celular.
- Use 300 pg de RNA de membranas transcritas in vitro e plasmídeos de marcadores fluorescentes fluorescentes nucleares, como pCS-membrana-ceruleanFP (Addgene plasmid #53749) ou pCS-memb-mCherry (Addgene plasm #53750) em combinação com pCS2+ H2B-mTagBFP2 (Addgene plasmid #99267) ou pCS2+ H2B-TagRFP-T (Addgene plasmid #99271) em injeções. Para um filme de amostra com membrana celular e marcadores de núcleos, consulte o Vídeo 4.
NOTA: O tamanho médio da célula do tecido PSM tem cerca de ~5 μm de diâmetro, dos quais os núcleos compreendem ~3 - 4 μm. Um tamanho de pixel de ~0,5 μm e uma seção z de ~1 μm devem ser registrados para segmentação celular adequada.
- Use 300 pg de RNA de membranas transcritas in vitro e plasmídeos de marcadores fluorescentes fluorescentes nucleares, como pCS-membrana-ceruleanFP (Addgene plasmid #53749) ou pCS-memb-mCherry (Addgene plasm #53750) em combinação com pCS2+ H2B-mTagBFP2 (Addgene plasmid #99267) ou pCS2+ H2B-TagRFP-T (Addgene plasmid #99271) em injeções. Para um filme de amostra com membrana celular e marcadores de núcleos, consulte o Vídeo 4.
5. Imunostaining de explants tail
NOTA: Os tecidos cultivados após vários cenários de dissecção (alongamento, não alongamento, broto de cauda dissecado, metade PSM etc.) como explants de cauda de montada plana5 podem ser recuperados de câmaras de slides para quantificações mais imunossumanas de proteínas de interesse. Aqui, apresentamos o protocolo utilizado para a coloração de kinase regulada por sinal extracelular di-fosforilado (ppERK) de explants como leitura gradiente de sinalização FGF.
- Após a formação de somites até o estágio desejado, mude cautelosamente o deslizamento de cobertura para o canto da câmara de slides sem levantar.
- Com a ajuda de ~100 μL de meio de dissecção suplementar em uma pipeta Pasteur de vidro, recupere as explantas do slide e transfira para uma placa de cultura celular de 64 poços.
NOTA: A partir desta etapa, todas as substituições de solução podem ser realizadas sob um escopo de dissecção com uma pipeta de vidro separada para amostras fixas. Isso garantirá não perder tecidos explantos em poços ou transferi-los no meio. - Depois de transferir todas as explantas, suge o meio excessivo dos poços um por um e coloque 100 μL de 4% de paraformaldeído em PBS (PFA) em cada poço.
ATENÇÃO: PFA é uma solução tóxica com efeitos cancerígenos. Epi adequado deve ser usado durante o manuseio. - Fixar explantes em uma placa de 64 poços à temperatura ambiente por 1 h em um shaker.
- Explantas teciduais são mais sensíveis a deformações do que embriões inteiros. Ajuste a velocidade do agitador de acordo.
- Lave o fixador com 150 μL de PBS-Tw (0,1% Tween20 na PBS) três vezes. Colete a primeira lavagem em um recipiente específico "PfA Waste".
- Desidratar explantes em passos de 4×5 min, substituindo ~40 μL de solução cada vez por 100% de metanol (MeOH).
ATENÇÃO: MeOH é um químico tóxico que é volátil e inflamável. Trabalhe em um espaço bem ventilado e use EPI adequado para manuseio. - Como última etapa de desidratação, remova toda a solução dos poços e substitua por 100 μL de MeOH. Incubar a -20 °C por 15 min.
NOTA: Use um recipiente específico "MeOH Waste" para coletar as soluções até o passo 5.11. - Adicione 50 μL de MeOH e agite à temperatura ambiente por 5 minutos.
- Reidratar explantes em passos de 4×5 min, substituindo ~40 μL de solução cada vez por PBS-T (0,1% Triton-X 100 em PBS). Use um recipiente específico "MeOH Waste" para coletar as soluções.
- Como última etapa de reidratação, remova toda a solução dos poços e substitua por 100 μL de PBS-T.
- Para permeabilização tecidual tratar explants com Triton-X 100 de 1,5% em PBS por 20 minutos à temperatura ambiente no shaker.
- Lave amostras com MAB-D-T (0,1% Triton-X 100 detergente e 1% de sulfóxido de dimetil (DMSO) em 150 mM NaCl 100 mM mM tampão de ácido macho pH 7,5) 3×5 min.
ATENÇÃO: DMSO é inflamável e um mutagênico tóxico. Epi adequado deve ser usado durante o manuseio. - Incubar explantes em 100 μL/solução de bloqueio de soro de poço (2% de soro bovino fetal em MAB-D-T) por 2 horas em temperatura ambiente.
- Substitua toda a solução de bloqueio por uma solução de anticorpos primários de 50-100 μL/well (diluição de 1:1000 de anticorpo monoclonal do rato contra ppERK no bloco sérico). Incubar amostras O/N (>16 h) a 4 °C no shaker.
- Lave a solução de anticorpos primária com MAB-D-T 5×5 min.
- Incubar amostras em solução secundária de anticorpos (Alexa Fluor 597 anti-rato de cabra IgG2b (1:200) e Hoechst 33342 (1:5000) em MAB-D-T) O/N a 4 °C em um shaker ou por 3 h à temperatura ambiente.
NOTA: A partir desta etapa, cubra a placa de 64 poços com papel alumínio para evitar a exposição leve de amostras tratadas com anticorpos secundários.
ATENÇÃO: Hoechst 33342 é um potencial cancerígeno. Epi adequado deve ser usado durante o manuseio. - Lave a solução de anticorpos secundários com PBS-Tw 3×5 min.
- Fixar amostras com PFA por 15 minutos em temperatura ambiente.
- Lave o fixador com PBS-Tw e equilibre amostras dentro de 60% de glicerol. Monte explantas em slides de microscópio com esmalte e 60% glicerol para imagem. Para obter resultados representativos de imunossuagem, consulte a Figura 3.
Resultados
Este protocolo permite a colheita geométrica plana de explantes de cauda de zebrafish vivos. A cultura tecidual apresenta três grandes vantagens sobre embriões inteiros: 1) controle da velocidade de alongamento do eixo, 2) controle sobre várias fontes de sinalização (morfógeno) por dissecção simples, e 3) quase objetivo, alta ampliação e alta imagem viva na NA.
Câmaras de slides quimicamente não tratadas permitem que a explanta da cauda alongue seu eixo principal (
Discussão
Este artigo apresenta um protocolo detalhado de uma técnica de explant de cultura de tecido que desenvolvemos e usamos recentemente5 para embriões de zebrafish. Nossa técnica baseia-se nos métodos anteriores de explantagem emfilhotes 8 e zebrafish9,10,11 organismos modelo. Explantas de cauda preparadas com este protocolo podem sobreviver desde >12 h em uma simples câmara de sl...
Divulgações
Os autores não têm nada a revelar e não declaram conflitos de interesse.
Agradecimentos
Agradecemos ao AECOM Zebrafish Core Facility e ao Serviço Veterinário Infantil de Cincinnati pela manutenção de peixes, ao Núcleo de Imagem Infantil de Cincinnati por assistência técnica, Didar Saparov pela assistência com a produção de vídeo e Hannah Seawall para a edição do manuscrito. A pesquisa relatada nesta publicação foi apoiada pelo Instituto Nacional de Ciências Médicas Gerais dos Institutos Nacionais de Saúde sob o Prêmio Número R35GM140805 a E.M.Ö. O conteúdo é de responsabilidade exclusiva dos autores e não representa necessariamente as opiniões oficiais dos Institutos Nacionais de Saúde.
Materiais
| Name | Company | Catalog Number | Comments |
| 1 mL Sub-Q Syringe with PrecisionGlide Needle | Becton, Dickinson and Co. | REF 309597 | for dechorionating embryos and manipulations |
| 200 Proof Ethanol, Anhydrous | Decon Labs | 2701 | for immunostaining |
| Antibiotic Antimycotic Solution (100×) | Sigma-Aldrich | A5955 | for tissue dissection media |
| Calcium Chloride Anhydrous, Powder | Sigma-Aldrich | 499609 | for tissue dissection media |
| Dimethylsulfoxide | Sigma-Aldrich | D5879 | for immunostaining |
| Disposable Scalpel, #10 Stainless Steel | Integra-Miltex | MIL4-411 | for preparing tape slide wells |
| Ethyl 3-aminobenzoate methanesulfonate salt (Tricaine) | Sigma-Aldrich | 886-86-2 | (optional) for anesthesizing tissues older than 20 somites stage |
| Fetal Bovine Serum (FBS) | ThermoFisher | A3160601 | additional for tissue culture media |
| Goat anti-Mouse IgG2b, Alexa Fluor 594 | Invitrogen | Cat#A-21145; RRID: AB_2535781 | secondary antibody for immunostaining |
| L-15 Medium with L-Glutamine w/o Phenol Red | GIBCO | 21083-027 | for tissue dissection media |
| Methanol | Sigma-Aldrich | 179337 | for immunostaining |
| Microsurgical Corneal Knife 2.85 mm Angled Tip Double Bevel Blade | Surgical Specialties | 72-2863 | for tissue dissection |
| Mouse monoclonal anti-ppERK | Sigma-Aldrich | Cat#M8159; RRID:AB_477245 | for ppERK immunostaining |
| NucRed Live 647 ReadyProbes Reagent | Invitrogen | R37106 | (optional) for live staining of cell nuclei |
| Paraformaldehyde Powder, 95% | Sigma-Aldrich | 158127 | for fixation of samples for immunostaining |
| Rat Tail Collagen Coating Solution | Sigma-Aldrich | 122-20 | (optional) for chemically activating slide chambers |
| Stage Top Incubator | Tokai Hit | tokai-hit-stxg | (optional) for temperature control during live imaging |
| Transparent Tape 3/4'' | Scotch | S-9782 | for preparing tape slide wells |
| Triton X-100 | Sigma-Aldrich | X100 | for immunostaining |
| Tween 20 | Sigma-Aldrich | P1379 | for immunostaining |
| Zebrafish: Tg(actb2:2xMCP-NLS-EGFP) | Campbell et al., 2015 | ZFIN: ZDB-TGCONSTRCT-150624-4 | transgenic fish with nuclear localized EGFP |
| Zebrafish: Tg(Ola.Actb:Hsa.HRAS-EGFP) | Cooper et al., 2005 | ZFIN: ZDB-TGCONSTRCT-070117-75 | transgenic fish with cell membrane localized EGFP |
Referências
- Assheton, R. . Growth in length: Embryological Essays. , (1916).
- Gomez, C., et al. Control of segment number in vertebrate embryos. Nature. 454 (7202), 335-339 (2008).
- Westerfield, M. . The Zebrafish Book: a guide for the laboratory use of zebrafish (Danio rerio), 3rd edition. , (1995).
- Icha, J., Weber, M., Waters, J. C., Norden, C. Phototoxicity in live fluorescence microscopy, and how to avoid it. BioEssays. 39 (1700003), (2017).
- Simsek, M. F., Ozbudak, E. M. Spatial fold change of Fgf signaling encodes positional information for segmental determination in zebrafish. Cell Reports. 24 (1), 66-78 (2018).
- Dubrulle, J., Pourquié, O. fgf8 mRNA decay establishes a gradient that couples axial elongation to patterning in the vertebrate embryo. Nature. 427 (6973), 419-422 (2004).
- Diez del Corral, R., et al. Opposing FGF and Retinoid Pathways Control Ventral Neural Pattern, Neuronal Differentiation, and Segmentation during Body Axis Extension. Neuron. 40 (1), 65-79 (2003).
- Stern, H. M., Hauschka, S. D. Neural tube and notochord promote in vitro myogenesis in single somite explants. Developmental Biology. 167 (1), 87-103 (1995).
- Langenberg, T., Brand, M., Cooper, M. S. Imaging brain development and organogenesis in zebrafish using immobilized embryonic explants. Developmental Dynamics. 228 (3), 464-474 (2003).
- Picker, A., Roellig, D., Pourquié, O., Oates, A. C., Brand, M. Tissue micromanipulation in zebrafish embryos. Methods in molecular biology. 546 (11), 153-172 (2009).
- Manning, A. J., Kimelman, D. Tbx16 and Msgn1 are required to establish directional cell migration of zebrafish mesodermal progenitors. Developmental Biology. 406 (2), 172-185 (2015).
- Kimmel, C. B., Ballard, W. W., Kimmel, S. R., Ullmann, B., Schilling, T. F. Stages of embryonic development of the zebrafish. Developmental Dynamics. 203 (3), 253-310 (1995).
- Kaufmann, A., Mickoleit, M., Weber, M., Huisken, J. Multilayer mounting enables long-term imaging of zebrafish development in a light sheet microscope. Development. 139, 3242-3247 (2012).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados