
Method Article
Optik Cımbız Kullanarak Hapsetmede Hücre Altı Mekaniğinin Doğrudan Kuvvet Ölçümleri
Bu Makalede
Özet
Burada, izole embriyonik zebra balığı hücrelerinin hücre içi mekanik özelliklerini optik bir tuzakla doğrudan kuvvet ölçümü ile üç boyutlu hapsetmede araştırmak için bir protokol sunuyoruz.
Özet
Çok hücreli bir organizmanın gelişimi sırasında, tek bir döllenmiş hücre bölünür ve çeşitli işlevlere sahip birden fazla dokuya yol açar. Doku morfogenez, hücre altı mekanik özelliklerin varyasyonlarına neden olan tek hücre düzeyinde moleküler ve yapısal değişikliklerle elde gider. Sonuç olarak, aynı hücre içinde bile, farklı organeller ve bölmeler mekanik gerilmelere farklı dirençler; ve mekanotransdüksiyon yolları mekanik özelliklerini aktif olarak düzenleyebilir. Bir hücrenin doku nişinin mikroçevriciliğine uyum sağlama yeteneği kısmen mekanik gerilmeleri hissetme ve yanıt verme yeteneğinden kaynaklanmaktadır. Yakın zamanda, nükleer deformasyon ve konumlandırmanın bir hücrenin fiziksel 3D ortamı ölçmesini sağladığı ve hücre şeklindeki değişiklikleri çözmek için hücreye propriyosepsiyon duygusu sağladığı yeni bir mekanosensasyon paradigması önerdik. Bu yazıda, canlı hücrelerin içindeki hücre çekirdeğini şekillendiren, yapışık hücreler ve mekanik olarak sınırlı hücreler üzerinde örneklendirilen kuvvetleri ve malzeme özelliklerini ölçmek için yeni bir yöntem açıklıyoruz. Ölçümler, hücrelerin içindeki optik tuzaklarla invaziv olmayan bir şekilde gerçekleştirilebilir ve kuvvetlere ışık momentumunun kalibrasyonsuz olarak algılanması yoluyla doğrudan erişilebilir. Bu, çekirdeğin mekaniğini hücre yüzeyi deformasyonlarından bağımsız olarak ölçmeye ve dışlayıcı ve interoceptif mekanotransdüksiyon yollarının diseksiyona izin verir. Daha da önemlisi, yakalama deneyi, sitoskeleton, kalsiyum iyonları veya nükleer morfolojinin floresan görüntülemesi kullanılarak hücresel yanıtı ve hücre altı dinamikleri araştırmak için optik mikroskopi ile birleştirilebilir. Sunulan yöntemin uygulanması kolaydır, kuvvet ölçümleri için ticari çözümlerle uyumludur ve mitokondri, stres lifleri ve endozomlar gibi diğer hücre altı bölmelerin mekaniğini araştırmak için kolayca genişletilebilir.
Giriş
Doku morfogenez, biyokimyasal sinyallerin ve fiziksel kuvvetlerin mekansal olarak koordine olduğu karmaşık bir süreçtir. Gelişmekte olan embriyoda, biyokimyasal sinyal faktörlerinin gradyanları kader spesifikasyonunu belirler ve doğru doku desenini sağlar1,2. Aynı zamanda, içsel ve dışsal kuvvetler embriyonun mimarisinin inşasında rol oynar3,4. Hücre korteksi mekaniğinin bu bağlamdaki etkisi kapsamlı bir şekilde incelenmiştir5,6. Morfogenez sırasında mekano-kimyasal işlemler arasındaki sıkı bağlantı, doku mikroçevrimindeki mekanik kuvvetleri hissetme ve yanıt verme özelliklerine dayanır. Hücreler, böylece mekanik bilgileri hücre davranışını, hücre kaderini ve hücre mekaniğini kontrol eden belirli sinyal yollarına dönüştürücü kuvvete duyarlı hücre altı ve moleküler elementlerin varlığıyla mekanik sinyallerin kodunu çözer.
Gelişimsel süreçlerin ayırt edici özelliği, hücrelerin çok hücreli yapılar oluşturmak için gruplar olarak düzenlenmesidir. Bu nedenle, tek hücreler nadiren yeniden düzenlenir ve tek başına hareket eder, ancak supracellular migration7, (un) jamming transitions8,9 veya blastocyst compaction10 gibi kolektif davranışlar gösterdikleri sıkı bir sosyotopta ilişkilidir. Hücreler içinde ve hücreler arasında üretilen mekanik kuvvetler, kolektif hücre dinamiklerini öğretmek için önemli ipuçları olarak hizmet eder7,11. Ancak hücreler tek başlarına hareket etseler bile, doku tabakaları veya dar doku nişleri arasında yol alan progenitör hücreler gibi, üç boyutlu bir ortamda gezinirken geniş anizotropik mekanik kuvvetler yaşarlar. Hücreler üzerindeki bu mekanik gerilmeler hücresel davranış üzerinde derin sonuçlar doğurur12,13. Yoğun bir 3D doku ortamında göç sırasında pasif veya aktif mekanik eleman olarak ana mekanotransdüksiyon elemanı14,15 olarak çekirdek üzerinde birleşen çeşitli mekanizmalar araştırılmıştır15,16.
Yakın zamanda çekirdekleri elastik hücre içi mekano-ölçer olarak kullanarak şekil deformasyonlarını ölçmek için hücreleri donatan bir mekanizma önerdik12. Bir hücredeki en büyük organel olan çekirdek, hücreler mekanik esneme, hapsetme veya ozmotik stres altında polarize olduğunda, göç ettiğinde veya şekillerini değiştirdiğinde büyük deformasyonlara uğrar16,17,18,19. Çekirdeklerin hücre içi konumlandırılması ile birlikte nükleer zarfın hücrelere hücre deformasyonunun büyüklüğü ve türü (hücre sıkışması ve hücre şişmesi gibi) hakkında bilgi sağladığını bulduk. Çekirdeğin gerilmesi, INM'de kalsiyuma bağımlı cPLA2 (sitozolizik fosfolipaz A2) lipaz aktivitesini destekleyen iç nükleer zarın (INM) açılmasıyla ve ardından Araşidonik Asit (AA) salınımı ve hücre korteksinde miyozin II'nin hızlı aktivasyonu ile ilişkilidir. Bu, hücre kontrtinaklığının artmasına ve amoeboid hücre göçünün kortikal kontrtinaklık eşiğinin üzerine çıkarılmasına neden olur6. Hücre deformasyonuna verilen mekanosensitive yanıt bir dakikadan kısa bir sürede gerçekleşir ve hapsedildikten sonra geri döndürülebilir, bu da çekirdeğin mekanik stres koşullarında adaptif hücre davranışını düzenleyen hücresel propriyosepsiyon için bir gerinim ölçer görevi görür. Bu mekanosensitive yolunun zebra balığı embriyolarından elde edilen progenitör kök hücrelerde, hem pluripotent hem de soyuna bağlı hücrelerde aktif olduğu gösterilmiştir12 ve farklı türlerde ve hücre hatlarında korunur20.
Bir hücre-mekanosensor olarak nükleer özelliklere ek olarak, nükleer mimari ve mekanik, geliştirme sırasında ve hücre kaderi spesifikasyonuna yanıt olarak özünde düzenlenir21, bu nedenle hücresel mekano duyarlılığını ayarlar22,23. Bunun sonucu, morfolojik değişikliklere ve bir önsegratory'den göçmen durumuna geçişlere izin veren nükleer uyumdaki bir değişiklik olabilir ve bunun tersi de olabilir.
Atomik kuvvet mikroskopisi24,25, mikropipette aspirasyonu26,27, mikroakışkan teknoloji28 ve mikroneedles29 gibi hücre çekirdeği mekaniğini ölçmek için çeşitli teknikler uygulanmıştır. Bununla birlikte, bu tekniklerin çoğu, tüm hücrenin deforme olması gerektiği anlamında istilacıdır, mekanik özelliklerin ölçümünü ve çekirdeğin kendisinin kuvvete bağlı yanıtlarını sınırlar. Hücre yüzeyinin ve mekanosensitive hücre korteksinin eşzamanlı deformasyonunu atlatmak için izole çekirdekler çeşitli bağlamlarda çalışılmıştır31,32. Bununla birlikte, nükleer izolasyonun mekanik çekirdek özelliklerindeki bir değişiklik ve bunların düzenlenmesiyle ilişkili olduğu göz ardı edilemez (referans24 ve yayınlanmamış gözlemler).
Optik cımbızlar (OTs), hücre mekanobiyolojisinde çok sayıda deneye izin veren ve moleküler makinelerin kimyasalı mekanik enerjiye nasıl dönüştürdüğünü anlamamızda etkili olan çok yönlü bir teknolojidir33,34. Optik cımbız, çevredeki medium33'ten daha yüksek kırma indeksine sahip dielektrik parçacıklara optik kuvvetler uygulamak için sıkı bir şekilde odaklanmış bir lazer ışını kullanır. Bu tür kuvvetler yüzlerce pico-Newton'un sırasına göre olabilir ve parçacığın lazer tuzak odağı içinde etkili bir şekilde hapsedilmesine neden olarak sıkışan parçacığın üç boyutta manipülasyonu sağlar. Ölçümlerin canlı hücrelerin içinde invaziv olmayan bir şekilde yapılabilmesinde ışık kullanımı önemli bir avantaja sahiptir. Optik manipülasyonlar lazer ışınının tuzak odağı ile daha da sınırlıdır. Bu nedenle, manipülasyon çevredeki hücresel zarları uyarmadan gerçekleştirilebilir ve iyon kanallarının kuvvete bağlı aktivasyonu gibi plazma zarındaki aktisin korteksi veya mekanosensitif süreçleri etkilemez.
Optik cımbız yaklaşımının zorluğu, equipartition teoremine veya lazer gücüne bağımlı bir kaçış kuvvetini ölçmek için tanımlanmış Stokes sürükleme kuvvetlerinin kullanımına dayanan klasik yaklaşımları kullanarak mikrosfere uygulanan kuvvetleri tam olarak belirlemektir35. Bu yöntemlerin bir in vitro deneyde uygulanması kolayken, genellikle hücresel bir ortama çevrilemezler. Momentum korumanın ilk ilkelerinden türetilen doğrudan kuvvet kalibrasyonuna dayanan çeşitli stratejiler sahaya sürünmiştir36,37. Diğer kuvvet spektroskopisi yaklaşımlarının aksine, kuvvet ölçümleri keyfi şekilli sıkışmış parçacık38,39 ile yerel bir ışık momentumu değişiminden elde edilir. Deneysel kurulumumuzda, optik kuvvetlerden kaynaklanan ışık momentumundaki değişiklikler, yerinde tuzak kalibrasyonuna gerek kalmadan doğrudan ölçülür40,41,42,43. Böylece, ölçümler hücrenin içi gibi viskoz bir ortamda veya hatta bir doku içinde mümkün hale gelir ve kuvvetler pN seviyesine kadar kolayca ölçülebilir.
Bu protokolde, hücre içi organelleri veya yapıları mekanik olarak manipüle etmek ve mekanik özelliklerini optik bir cımbız kurulumu ile nicel olarak değerlendirmek için bir test açıklıyoruz. Bu kurulum, hücresel davranışın veya hücre içi dinamiklerin paralel görüntülenmesini sağlayan dönen bir disk floresan mikroskobuna entegre edilmiştir. Test, çekirdek gibi belirli hücresel bölmelerin mekanik özelliklerinin karakterizasyonuna izin verirken, aynı zamanda deformasyonun bir sonucu olarak moleküler sinyal yollarının olası mekanoresponse ve aktivasyonunu inceler. Ayrıca, enjekte edilen mikrobeadların hücreler içinde optik olarak hapsedilmesi, polistiren boncukunun (n = 1.59) çekirdeğin iç kırılma kontrastı44 ile kstoplazma (n ~ 1.38) ile karşılaştırıldığında önemli ölçüde daha yüksek bir kırılma indeksi sayesinde girinti kuvvetinde bir artışa izin verir. Sunulan strateji, diğer hücre içi yapıların ve organellerin çalışmasına, aktif mikrorheolojiyi içeren diğer yaklaşımlara, aynı/farklı alt hücresel yapıları aynı anda araştırmak için birden fazla optik tuzağın kullanılmasına ve canlı embriyodaki hücre mekanobiyolojisini hedefleyen ölçümlere kolayca uyarlanabilir.
Protokol
Kullanılan tüm protokoller Kurumsal Hayvan Bakım ve Kullanım Etik Kurulu (PRBB-IACUEC) tarafından onaylanmış ve ulusal ve Avrupa yönetmeliklerine göre uygulanmıştır. Tüm deneyler 3R'lerin ilkelerine uygun olarak gerçekleştirildi. Zebra balığı (Danio rerio) daha önce açıklandığı gibi muhafaza edildi.
1. İzole primer embriyonik zebra balığı progenitor kök hücrelerinin hazırlanması
- Mikropipette ve agarose hazırlama
NOT: Tam bir zebra balığı embriyo mikroenjeksiyon protokolü için bkz.- Bir mikropipette puller ile, iki iğne elde etmek için 1.0 mm cam kılcal damarı çekin45. Kullanılmayan iğneleri, ince ucu taşıma sırasında hasardan korumak için bir oyun yatayı minderine bağlı 150 mm Petri kabında veya içten dışa bir laboratuvar bant halkasında saklayın.
- E3'te %1 ultra saf agarose eritin (5 mM NaCl, 0,17 mM KCl, 0,33 mM CaCl2, 0,33 mM MgSO4) 10 s için standart bir mutfak/laboratuvar mikrodalgasında. Agarose eriyene kadar karışımı kısa süreler (birkaç saniye) boyunca tekrar tekrar ısıtın.
- Agarose tamamen eridiğinde, kısa bir süre soğumaya bırakın ve ardından 10 cm'lik bir Petri kabına dökün. Kabarcıkların ortaya çıkmasını önleyerek agarose'un üstüne üçgen mikroenjeksiyon kalıbını (bkz. Malzeme Tablosu) yavaşça ekleyin. Agarose yüzeyinde kalmasını sağlayarak kalıbı itmeyin.
- Agarose tamamen katılaştığında, agaroseda herhangi bir kırılmayı önlemek için yumuşak bir kuvvet uygulayarak üçgen kalıbı çok yavaş bir şekilde çıkarın. Plaka 2-4 hafta boyunca 4 °C'de baş aşağı saklanabilir.
- Mikroenjeksiyondan 30 dakika önce, plakayı buzdolabından alın ve oda sıcaklığında stabilize olması için önceden ısıtılmış E3'ü 28 °C'ye ekleyin.
- Enjeksiyon karışımı hazırlığı
- Enjeksiyon karışımını hazırlamak için, RNase içermeyen suda 1 μm mikrobeadları (polistiren, floresan olmayan) 1:5 oranında seyreltin.
- Floresan belirteçlerin geçici ekspresyyürü veya rekombinant gen yapılarının ekspresy ekspresyolü ve/veya morfotinin istenen konsantrasyonda birlikte enjeksiyonu için mRNA'yı hazırlayın.
NOT: Mikrobeadların birlikte enjekte edilmesi için tipik bir enjeksiyon karışımı ve etiketlenecek embriyo başına 100 pg mRNA, örneğin, H2A-mCherry ile çekirdek: 1 μL boncuk + 1 μL mRNA (stok konsantrasyonu 1 μg / μL' dir) + 2,5 μL RNA içermeyen su + 0,5 μL fenol kırmızısı (stok çözeltisi% 0,5, fenol kırmızısı zorunlu değildir; enjekte edilen damlanın daha iyi görselleştirilmesi için kullanılır, ancak etiketlenmemiş enjeksiyon damla, deneyimli bir deneyci için de görülebilir). RNA enjeksiyonu, enjekte edilen embriyoları seçmek için de yararlı olabilir. Floresan mikrobeadlar, floresan olmayanlar yerine, onları görselleştirmek için enjekte edilebilir.
- Mikroenjeksiyon iğne yükleme ve kalibrasyon
- Zaman Kapılı seçeneğini kullanarak mikroinjektörü açın. Enjeksiyon hacmini düzgün bir şekilde kalibre etmek için bu ayar çok önemlidir. Gating süresini yaklaşık 500 ms olarak ayarlayın.
- Enjeksiyon karışımının 3 μL'sini bir mikro yükleyici pipet kullanarak iğneye yükleyin.
- İğneyi mikromanipülatöre yerleştirin ve sıkıca kapatın. Mikromanipülatörnün iyi bir konumda olup olmadığını ve enjeksiyon plakasında x-y yönünde hareket etmek için yeterli özgürlüğe sahip olup olmadığını kontrol edin.
- Bir mikrometre kaydırağı (5 mm/100 bölme) kullanarak, üstüne bir damla mineral yağ ve enjeksiyon karışımının bir damlasını doğrudan mineral yağa atarak düşme boyutunu ölçün.
- Keskin sivri bir uç oluşturmak için iğneyi dik bir açıda keskin tokmaklarla kırpın. Damla boyutunu 0,5 nL enjekte edilen malzemeye karşılık gelen 0,1 mm'ye ayarlayın.
NOT: İğne kesilerek bu hacim aşılırsa, kalibrasyon prosedürünün yeni bir iğne ile yeniden yapılması önerilir. Mikroinjektörün gating süresi, düşme hacmine uyacak şekilde hafifçe ayarlanabilir; bununla birlikte, kısa gating süreleri embriyolara potansiyel olarak zarar verebilecek büyük bir iğne çapına karşılık gelir.
- Zebra balığı embriyolarının tek hücreli aşamada mikroenjeksiyon
- Boncuk karışımının ilk hücre bölünmesi gerçekleşmeden önce doğrudan tek hücreli (zigot) evre embriyosuna mikroenjeksiyonu için döllenmeden kısa bir süre sonra zebra balığı embriyolarını toplayın.
NOT: Bu, mikrokürelerin doğru dağılımını ve deneylerin yapıldığı daha sonraki gelişim aşamalarında (blastula-gastrula aşaması) hücre başına en az bir mikrosfer ile izole edilmiş blastomerlerin yeterince yüksek verimini sağlar. Hücre içinde iki küre varsa girinti deneyleri hala yapılabilir, ancak boncuk içermeyen hücreler hariç tutulmalıdır (küresiz girinti mümkün olsa bile). Bu protokolde AB wildtype suşları kullanılmıştır, ancak tl gibi diğer suşlar kullanılabilir. - Tek hücreli evre embriyoları (zigot) plastik bir Pasteur pipet kullanarak Şekil 1A'da gösterildiği gibi önceden ısınmış üçgen şekilli% 1 agarose kalıbına yerleştirin.
- Embriyoların etrafta dolaşmasını önlemek için aynı pipetle ekstra ortamı çıkarın. Embriyoları bir fırça ile üçgen kalıba hafifçe itin. Doğru oryantasyonu kolaylaştırmak için embriyolar arasında biraz boşluk tutun (Şekil 1B).
- Embriyoları bir fırça ile hafifçe hizalayın, böylece embriyolar yanal olarak yönlendirilir, zigotların bir hücresi Şekil 1B'de gösterildiği gibi açıkça görülebilir. Embriyonun bir hücresi iğne yönüne (embriyonun hayvan kutbundan enjeksiyon) veya şekil 1C'de gösterildiği gibi yumurta sarısı hücresine (embriyonun vegetal kutbundan enjeksiyon) bakan ters yönde olduğunda mikroenjeksiyon için ideal bir yönelime ulaşılır.
- Bir elinizle çanağı tutun ve diğer elimizi mikromanipülatör denetleyicisini kullanarak iğne ucunu konumlandırmak için kullanın. İğne ucunu embriyolara doğru küt.
- Koroniti delin ve stereomikroskop aracılığıyla prosedürü izlerken iğne ile tek hücreli embriyoya girin. İğnenin doğru yerleştirilmesini ve enjekte edildikten sonra Şekil 1C'de gösterildiği gibi enjekte edilen damlanın doğru konumunu sağlayın.
- Tüm embriyolar için tekrarlayın: iğneyi yukarı hareket ettinin, bir sonraki embriyo ortalanana kadar embriyolarla birlikte yemeği kaydırın, iğneyi küt küt küt küt koyun ve enjekte edin.
- Tüm embriyo seti enjekte edildikten sonra, embriyoları biraz E3 yıkayarak agarose kalıb / Petri kabından çıkarın ve plastik bir Pasteur pipet kullanarak yeni bir Petri kabına koyun. Mikroenjeksiyon işlemi sırasında embriyoların kurumasını önlemek için enjeksiyon plakasına yeterli ortam yerleştirilmesi önerilir.
- İstenilen embriyo sayısı enjekte edilene kadar işlemi tekrarlayın. Boncukların maksimal ve homojen yayılmasını sağlamak için embriyolar tek hücre aşamasında olmalıdır.
NOT: Bu prosedür erken blastula embriyoları için optimize edilmiştir ve farklı gelişim evreleri araştırılacaksa muhtemelen optimize edilmesi gerekir. - Enjekte edilen embriyoları, birincil hücre kültürü protokolüne geçmeden önce yaklaşık 4 saat boyunca veya istenen aşamaya (Şekil 1D) kadar 28-31 °C'de bir inkübatörün içine yerleştirin.
NOT: İsteğe bağlı olarak, embriyoların hayatta kalmayı sağlamak ve toksisite eserlerini ekarte etmek için blastula aşamasının (veya istenen ölçüm zaman noktasının) ötesinde gelişmesine izin verin. Larva aşamalarında, % 0.75 agarose trikain ile uyuşturularak uyuşturularak larvalar monte edin ve mikrokürelerin çeşitli dokulardaki dağılımını görüntüleyin. Stok çözeltisi yapmak için karıştırın: 97,9 mL damıtılmış suda 400 mg trikain tozu, yaklaşık 2,1 mL 1 M TRIS bazlı (pH 9) ve pH 7'ye ayarlayın. Bu çözelti 4 °C'de saklanabilir. Trikaini anestezik olarak kullanmak için, 4,2 mL stok çözeltisini 100 mL yumurta ortamında (veya istenen ortamda) seyreltin; bu durumda E3 kullanılmıştır. Ayrıntılar için reference46'ya bakın.
- Boncuk karışımının ilk hücre bölünmesi gerçekleşmeden önce doğrudan tek hücreli (zigot) evre embriyosuna mikroenjeksiyonu için döllenmeden kısa bir süre sonra zebra balığı embriyolarını toplayın.
2. Tek hücreli hazırlama ve boyama
- Küre evre embriyolarını (döllenmeden saatler sonra) plastik bir Pasteur pipet kullanarak bir cam tabağa yerleştirin. Enjekte edilen boncukların sinyali için pozitif olan ve mRNA enjeksiyonu durumunda floresan proteini ifade eden embriyoları seçin. Bazı embriyolar yüksek boncuk kümelenmesi gösterebilir ve hariç tutulabilir.
- Embriyoları mansiyonları kullanarak manuel olarak kaynatın. Cam pastör pipet kullanarak yaklaşık 10-15 embriyoyu 1,5 mL reaksiyon kaplarına aktarın.
NOT: Embriyolar dekonsiyone edildiğinde plastiğe bağlanırlar ve cam eşyaların kullanılması gerekir. Cam plakaya alternatif olarak,% 1 agarose ince bir tabakaya sahip plastik bir Petri kabı kullanılabilir. Hücre yüzey proteinlerinde proteolitik hasarı ve mekanik hücre ve doku özelliklerinde olası değişiklikleri önlemek için enziptik Pronaz tedavisi yerine manuel dekonsiyon tercih edilmeli ve uzun iyileşme süreleri önlenmelidir47.
- Embriyoları mansiyonları kullanarak manuel olarak kaynatın. Cam pastör pipet kullanarak yaklaşık 10-15 embriyoyu 1,5 mL reaksiyon kaplarına aktarın.
- E3 ortamını çıkarın ve önceden ısıtılmış CO2-bağımsız doku kültürü ortamına 500 μL ekleyin (DMEM-F12; L-glutamin ve 15 mM HEPES ile, sodyum bikarbonat ve fenol kırmızısı olmadan 10 ünite penisilin ve 10 mg / L streptomisin ile desteklenir).
NOT: Mikroskop inkübatörü kullanılmadığı sürece CO2'ye bağımlı ortam kullanmayın. Karbonat tamponlu koşullarda RPMI kullanımı ortamın pH'ında değişikliklere neden olur ve hücre sağkalımını etkileyebilir. Bir diğer önemli husus da serum içeren kültür medyasından kaçınmaktır. Serum, rho/rock yolunun güçlü bir aktivatörü olan ve progenitör kök hücrelerde hücresel kontrtiyansı ve hareketliliği kontrol edebilen Lizosfatidik asit (LPA) içerebilir6. Nükleer morfoloji veya mekaniğe müdahale edebilecek ozmotik zorlukları önlemek için ortamın ozmolaritesi 300 mOsm'de tutulmalıdır12. - Tüpü hafifçe sallayarak hücreleri manuel olarak ayırın. Tüpün içeriğinin gözle görülebilen büyük parçalar olmadan bulanık hale gelmesini sağlayın. Hücrelerin hasarını ve kaybını en aza indirmek için kabarcıkların oluşumundan kaçının.
- 3 dakika boyunca 200 x g'da santrifüj. Pelet açıkça görülebilir olmalıdır.
- Üstnatant kaldırmak ve aşağıda ayrıntılı adımlardan birini izleyin.
- Boyamaya gerek yoksa, 500 μL DMEM ekleyin. Peletin üzerine sıvı bir jet hedef alarak 200 μL pipetle hafifçe yeniden diriltin. Hücrelere aşırı kesme kuvveti uygulamayın. Köpürme hücrelerin zarar gördüğünü gösterir.
- Çekirdeği Hoechst gibi DNA boyalarıyla etiketlemek için, 1.000 μL DMEM'de 0,5 μL DNA-Hoechst (stok 2 mg/mL) karıştırarak 1 μg/mL nihai konsantrasyon elde edin. Bu boyama çözeltisinin 500 μL'lik kısmını hücrelere ekleyin ve yavaşça yeniden biriktirin. Karanlıkta 7 dakika kuluçkaya yaslanın.
- Hücreleri floresan kimyasal kalsiyum göstergesi Calbryte-520 ile boyamak için, DMEM'deki 5 μM konsantrasyonuna Calbryte-520 ekleyin. Karanlıkta 20 dakika kuluçkaya yaslanın.
NOT: 2.5.2 ve 2.5.3 adımlarında belirtilen protokoller bu özel ürünler için optimize edilmiştir. Diğer boyamalar üretici tarafından belirtilen protokoller kullanılarak yapılabilir.
- Adım 2.4 ile aynı ayarları kullanarak tekrar santrifüjleyin; üstnatantı çıkarın ve süspansiyondaki numuneler için 50 μL DMEM'de veya hapsedilen hücreler için 20 μL DMEM'de hücreleri (küme oluşumunu önlemek için) hafifçe yeniden biriktirin.
3. Polidimetilsiloksan (PDMS) aralığı kullanılarak optik bindirme odalarının hazırlanması
NOT: Işık momentumu algılamaya dayalı optik kuvvet ölçümleri, optik tuzaklardan çıkan tüm ışığın yakalanmasını gerektirir40. Α (pN/V) sabit kalibrasyon faktörünün sağlamlığı için, optik kuvvet sensörünün arka odak düzleminde (BFP) ışık dağılımı foton momentumu ile doğru bir yazışmayı taşımalıdır. Bu, toplama lensinin yüzeyinden bindirme düzlemine, optik bindirme odalarının maksimum yüksekliği olan yaklaşık 2 mm'ye olan mesafeyi belirler.
- 1,5 numaralı cam alt tabakların PDMS spin kaplaması.
NOT: Yaklaşık 40 yemek için aşağıdaki tarif verilmiştir. Elde edilen mikro şarampol, deneylerin askıya alınmış veya kapalı hücreler üzerinde yapılıp yapılmayacağına bağlı olarak farklı yüksekliklere sahip olacaktır (Şekil 1D).- 50 mL konik tüpte 9 mL baz polimer PDMS ve 1 mL PDMS kürleme maddesi karıştırın. Kür maddesinin doğru dağılımını sağlamak için iki ürünü aktif olarak karıştırın.
- Vakum pompası kullanarak kabarcıkları önlemek için karışımı gazdan arındırın. Konik tüpü bir vakum şişesinde tanıtın ve odayı boşaltın. Karışımda kabarcıklar bulunmayana kadar bekleyin.
NOT: PDMS'nin şahin tüpünden köpürmesini ve dökülmesini önlemek için vakumun yavaşça açılmasını açın. - Cam alt kabı spin-coater aynasının üzerine yerleştirin (Şekil 2A). Tırmalamamak, parmak izi almamak veya tabağı kirletmemek için nazik olun. Spin-coater kutusunu alüminyum folyo ile PDMS sızıntılarından koruyun.
- Süspansiyondaki hücreler üzerinde deneyler için OT odaları için, alt kabın ortasına yaklaşık 250 μL PDMS karışımı ekleyin ve 1 dakika boyunca 750 rpm'de döndürün. PDMS katmanının yüksekliği yaklaşık 50 μm 48 olacaktır.
- Sınırlı hücreler üzerinde deneyler için OT odaları için, küçük bir PDMS damlası (yaklaşık 50 μL) ekleyin ve 5 dakika boyunca 4.000 rpm'de döndürün. PDMS katmanının yüksekliği yaklaşık 10 μm olacaktır. Farklı PDMS kalınlıklarının nasıl eldeılacağı hakkında ayrıntılı bir protokol için bkz.
- PDMS kaplı cam tabanlı yemekleri 70 °C'de 1 saat boyunca tedavi edin.
- PDMS tabakasına neşterle 1 x 1 cm kare kesin ve cımbızla soyun (Şekil 2C). Sınırlı hücreler durumunda, PDMS kalıntılarını izopropanol ile yıkayın.
- Süspansiyonda hafifçe tutturulmuş hücrelerle yapılan deneyler için oda kaplaması
- Kare boşluğun tüm yüzeyini kaplamak ve 30 dakika boyunca kuluçkaya yatmasını sağlamak için 0,5 mg / mL'de 100 μL Concanavalin A (ConA) ekleyin.
NOT: ConA, hücre yüzeyi şekerlerine bağlanan ve tek tek hücreleri kapak yüzeyine bağlayan bir kürsüdür. - ConA damlasını çıkarın ve ConA işlem görmüş yüzeyi çizmeden DMEM ortamı ile yüzeyi dikkatlice durulayın.
- Daha önce hazırlanmış numunenin 30 μL'sini (adım 2.6) kuyuya ekleyin ve herhangi bir hücre kümesinden kurtulmak için hafifçe yeniden diriltin.
- PDMS jantlarının üzerine hafifçe 22 x 22 mm #1,5 kapak camı yerleştirerek boşluğu kapatın (aniden düşmesine izin vermekten kaçının, mümkünse toparlama kullanın, Şekil 2B,C).
NOT: Herhangi bir kapak kılıfı kalınlığı üst cam kapak için işe yarar (toplama lensinin çalışma mesafesi 2 mm'dir).
- Kare boşluğun tüm yüzeyini kaplamak ve 30 dakika boyunca kuluçkaya yatmasını sağlamak için 0,5 mg / mL'de 100 μL Concanavalin A (ConA) ekleyin.
- Hapsedilen hücrelerle deneyler için oda hazırlığı
- Kare boşluğa hücre içeren 10 μL'lik bir çözelti damlası (adım 2.6) koyun (Şekil 2B).
- Çok nazikçe, numuneyi 22 x 22 mm'lik bir kapak camı ile sandviç edin, böylece damla tüm alana yayılır ve kabarcıklar gözlenmez. Yine, kapak camının aniden düşmesini önlemek için Şekil 2C'de gösterildiği gibi asa kullanmak uygundur.
4. OT oda aralığı için alternatif seçenekler
NOT: Mikrofabrikasyon atölyesi veya spin kaplayıcı yoksa bu adımlar izlenebilir.
- Süspansiyondaki hücrelerle deneyler için oda hazırlığı
NOT: Spin kaplayıcı olmaması durumunda, normal, çift taraflı viski bandı (yaklaşık 100 μm yüksekliğinde) kullanılarak bir aralayıcı yapılabilir.- Ortada yaklaşık 10 cm x 10 cm kare delikli bir çift taraflı viski bandı kesin (PDMS ile aynı boyutlar, Şekil 2B).
- Bandın koruyucu katmanlarından birini soyup çıkarın ve bandın açık tarafını 1,5 H cam tabanlı bir kabın ortasına yerleştirin. Hava kabarcıklarından kaçınırken tüm yüzeyi cama yapıştırmak için hafifçe bastırın ve ardından bandın kalan koruyucu tabakasını soyup çıkarın.
- 3.2. adımdaki yönergeleri izleyin.
- Hapsedilen hücrelerle deneyler için oda hazırlığı
NOT: Hücreleri hassas bir şekilde sınırlamak için, bilinen bir çapa sahip monodisperse mikropartikülleri, iki kapaklı gözlük arasında ara eleman olarak kullanılabilir.- Askıya alınan hücrelere 104 boncuk/μL konsantrasyonda 10 μm polistiren boncuk ekleyin.
- 22 x 60 mm'lik bir kapak camına hücre ve boncuk içeren 10 μL'lik bir çözelti damlası koyun.
- Çok nazikçe, numuneyi başka bir 22 x 60 mm kapak camı ile sandviç edin, böylece damla tüm alana yayılır ve kabarcıklar gözlenmez. Üst kapak camını nazikçe konumlandırmak için (aniden düşmesini önlemek), asalar kullanmak uygundur.
- Numune kuruyabileceği için, hazırlığın hızlı bir şekilde yapılması önerilir.
5. Hücre içi ölçümler için optik tuzağın kurulması
NOT: Aşağıdaki adımlar, acousto-optik sapmaya (AOD) dayalı bir optik mikromanipülasyon modülü ve ışık momentumu değişikliklerinin doğrudan algılanmasına dayanan bir optik kuvvet sensöründen oluşan ticari bir optik cımbız platformu için optimize edilmiştir (Şekil 2, referans12,40,49). Kurulumun ayrıntıları ve optik bileşenleri Şekil 2F'de bulunabilir. Optik cımbız manipülasyonları sırasında kuvvet kaynaklı deformasyonu gözlemlemek için, nipkow iplik disk konfokal mikroskobu, çift renkli floresan görüntüleme için ters mikroskobun sol bağlantı noktasına bağlanır. Genellik eksikliği olmadan, bu protokol ışık momentumu algılamaya dayalı doğrudan kuvvet ölçümleri ile donatılmış herhangi bir dinamik OTs sistemi ile uygulanabilir. In vivo uygulamalar için ev yapımı optik degrade tuzakları oluşturmak için ayrıntılı adım adım yordamlar mevcuttur50. AOD modülasyonuna dayananlar, birden fazla tuzak ve hızlı ölçümlerle yapılan nihai deneyler için öne çıkıyor51,52. Literatürde ışık momentumu tabanlı bir enstrüman oluşturmak için çeşitli protokoller mevcuttur36,39,40,53 ve diğer görüntüleme modalitesi (diferansiyel girişim kontrastı, geniş alan floresan vb.) kullanılabilir.
- Optik cımbız başlatma
- Çıkış gücü kararlılığını optimize etmek için, deneyden en az 30 dakika önce lazeri önemli ölçüde yüksek güçte (örneğin, 3 W) açın.
- Optik mikromanipülasyon ve kuvvet ölçüm ünitelerinin elektronik modülünü açın.
NOT: Tüm lazer güvenlik önlemlerini uygulayın ve sadece kurumsal kurul tarafından onaylanan ekipmanları kullanın. Lazer açıkken optik mikroskobun göz merceği kullanmayın. Her zaman onaylı IR koruma gözlüklerini kullanın (950-1080 nm aralığında OD7), IR lazer ışığını epifluoresans bağlantı noktası 2'deki deklanşörle engelleyin ve 5.3 adımından sonra optik kuvvet sensörü hizalamasını bitirene kadar optik bindirme yazılımını çalıştırmayın. Genel olarak, arka yansıma lazerde hasara neden olabileceği için son derece yansıtıcı bir örnek kullanmayın. - Optik mikromanipülasyon modülünün girişindeki dönen HWP (Şekil 2F) ile tuzak gücünü kontrol edin.
NOT: Bu protokolde kullanılan ticari optik mikromanipülasyon modülü zaten bu özelliği içerir. Ev inşa optik bindirme sistemleri için, daha yüksek ve daha kararlı lazer güçlerinin kullanılabilmesi için güç kontrolü için bu aracı entegre edin.
- Kalibrasyon için boş bir mikrochamber kullanın
- Çift taraflı bir viski bandına 1 x 1 cm kare kesin ve 1 mm kalınlığında bir mikroskop kaydırağı üzerine takın.
- Kareye su ekleyin ve 1,5 numaralı kapak camı (22 x 22 mm) ile üstten kapatın. Biraz daha yüksek hacimli su ekleme, örneğin, 30-40 μL kapalı odanın içindeki kabarcıkları önlemeniz önerilir. İçinden su dökülmesi durumunda kalibrasyon haznesini nazikçe silin.
- Optik kuvvet sensörünün hizalanması
- 60x/1.2 su daldırma hedefine bir damla su koyun. Kalibrasyon haznesini hedefe bakan 1,5 numaralı kapak camı ile sahneye yerleştirin. Hücre örneklerinin sonunda olacağı alt yüzeye odaklanın.
- Numuneyi kaplayan üst cam kaydırağın üzerine bir damla daldırma yağı ekleyin (Şekil 2D). Kuvvet sensörü ünitesinin toplama lensini yağ damlacığını temas edene kadar dikkatlice küstür.
NOT: Damlacık, tuzaklardan çıkan lazer ışığını toplayan tüm lensi kaplayacak kadar büyük olmalıdır. Genellikle, 200 μL tüm yüzeyi kaplamak ve kararlı bir daldırma teması sağlamak için yeterlidir. Muhafazakar olun ve örneğe sızabileceği için aşırı doldurmayı önleyin. - Üreticinin optik kuvvet sensörü hizalaması protokolünü izleyerek, OTs'yi konumlandırmak için kullanılacak yardımcı kameradaki örnek düzlem görüntüsüne bakın (AUX, Şekil 2F). Çok nazikçe, alan durması (FS, Şekil 2F-G) örnek düzlemde eşlenmiş görünene kadar optik kuvvet sensörünü kübünleyin. Bu, ışık momentumu değişikliklerinin numune değişmez tespitinden uygun doğrudan kuvvet ölçümlerini sağlayacaktır40.
NOT: FS'yi, görüntüsü görüş alanından (FOV) daha küçük olacak şekilde yeterince kapatın, bu nedenle görünür. Ekstra dikkatli olun ve optik kuvvet sensörünün toplama lensini numuneye doğru itmeyin. Optik kuvvet sensörünün dikey konumu alternatif olarak, tanımlanmış sayısal diyafram açıklığına (NA) sahip ışık konileri için BFP'deki bindirme ışığı dağılımının analizinden belirlenebilir. - Yağ damlacığı içinde hava kabarcıkları olmadığından emin olun; bunlar kuvvet ölçümlerini doğrudan etkileyebilir. Hava kabarcıklarını kontrol etmek için Bertrand lensi yerine yerleştirin (BL, Şekil 2G) ve göz merceğindeki görüntüleme yolunu gözlemleyin. Herhangi bir kir veya hava kabarcığı görünürse veya daha fazla yağ gerekiyorsa (Şekil S1A), lensi ve hazneyi tozsuz lens dokusuyla temizleyin ve prosedürü 5.3.2 ve 5.3.3 adımlarında tekrarlayın. Şekil S1B'de engelsiz bir optik yol resmedilmiştir.
- Optik kuvvet sensörünün tutucusuna yerleştirilen yanal vidaları kullanarak FS'yi FOV'a ortalayın. Doğruluk için FS'yi açın, böylece yardımcı kamerada görünen FOV'yi neredeyse doldurur (AUX, Şekil 2F).
6. Optik cımbız optimizasyonu
NOT: Doğrudan kuvvet ölçümü sadece sıkışan parçacık üzerine uygulanan kuvvetten kaynaklanan ışık momentumunun değişimine dayanır ve bu nedenle dolaylı yöntemlerin aksine, tuzak sertliğinin her deneyden önce kalibre edilmesi gerekmez. Sapma/kuvvet faktörünün (α; pN/V, reference41) cihaza özgü dönüşümü üretici tarafından kalibre edilir ve bu nedenle deneme değişmezdir. Bununla birlikte, lazer noktası 70 μm x 70 μm'lik bir alan üzerinde manipüle edildiği için, optimum bindirme ve güç kararlılığını sağlamak için 6.2-6.5 adımları kritik öneme sahiptir. Aşağıdaki adımlar, OT'lerin çalışma alanı üzerinde yarı otomatik bir şekilde optimize edilmesini sağlayan üretici yazılımında sağlanır.
- OTs yazılımını ve kamera AUX için satın alma yazılımını başlatın.
- Optik cımbız sürüş yazılımının Sistem Kalibrasyon alt menmenündeki Adım 1: Elektronik Ofset adımına tıklayarak ilk voltaj taban çizgisini çıkarın.
- OT çalışma alanında tuzak gücü düzleştirmesi yapmak için, HWP'yi buna göre döndürerek tuzak gücünü maksimumun yarısına ayarlayın. Lazer çıkışını değiştirerek değil, dönen HWP ile tuzak gücünü değiştirmeyin (Şekil 2F). Adım 2'ye tıklayın: Tuzak gücü düzleştirme için otomatik yordamı başlatmak için güç.
NOT: Bu, TUZAK gücünün OTs çalışma alanındaki değişimini telafi etmek için kritik bir adımdır (Şekil S1D). Başarılı bir rutin, TUZAK güç değişimini OTs çalışma alanında% 2'ye düşürür ve 2 dakika sonra yakınsar. - Tuzak konumu kalibrasyonu gerçekleştirmek için IR filtresini çıkarın, böylece lazerdeki ışık kamerada görünür. Görüntü düzlemini mikro şarampolün alt yüzeyine odaklanarak IR noktasını bulun. Kamera AUX alma yazılımındaki görüntü düzlemini (nesnel konum) ve histogram kontrastını ayarlayarak mümkün olan en küçük IR noktasını elde edin. Gerekirse, HWP'yi döndürerek optik tuzağın gücünü azaltın (Şekil 2F). Adım 3'e tıklayın: Otomatik rutin veya bindirme konumlandırma kalibrasyonunu başlatmak için konum.
NOT: Bu rutin, OT'nin kamera AUX'deki konum koordinatlarının AOD direksiyon açılarıyla hassas bir şekilde yazışmasını sağlar. Başarılı bir yordam, birkaç saniye içinde açıdan konuma eşleme oluşturur. - İlk momentum telafisi
NOT: Optik tuzağın numune boyunca hareketi, BFP'deki ışık momentum dağılımında değişikliklere neden olur (Şekil S1E, F). Bu, tuzak gücü 6.3 adımında olduğu gibi düzleştirilmiş olsa da, çalışma alanı üzerindeki lazer konumuyla ilgili kuvvetten bağımsız sinyal değişikliklerine yol açar. Sonuç, her deneyden önce düzeltilmesi gereken konum (optik olarak sıkışmış boncuk üzerinde hareket eden gerçek bir kuvvetin bağımsız) nedeniyle kuvvet taban hattındaki bir varyasyondur.- HWP'yi döndürerek denemelerde kullanılacak tuzak gücünü ayarlayın (Şekil 2F).
- Araçlar alt menmenunda Genel Ofset seçeneğine tıklayın. Bu, ilk momentum temelini düzelten optik cımbız yazılımının Ofset İptal yardımcısını açar.
- Öteleme | tıklayın Pozisyon-varyant başlangıç momentumunu düzeltmek için telafi edin.
NOT: Devam eden haftalarda hiçbir değişiklik optik yolu etkilemezse, tuzak gücü düzleştirme (adım 6.3) ve konum (adım 6.4) haritaları değişmez kalır. Bu nedenle, lazer tuzak yolunu etkileyebilecek optik elemanların (dikroik aynalar, filtreler vb.) her zaman aynı kombinasyonunu kullanmanızı veya yeni bir tuzak gücü düzleştirme rutini gerçekleştirmenizi öneririz. İlk momentum telafisi (adım 6.5) ile ilgili olarak, OTs platformunun üreticisi, her yeni bindirme gücü ve deneysel oturum için değiştirilmesi gereken anında bir kalibrasyon sağlar. Adım 6.3 ve 6.4, 5.2 adımında açıklanan boş kalibrasyon slaytında gerçekleştirilmelidir. Hücre veya diğer nesneleri içeren bir örnekte, adım 6.5, OTs çalışma alanındaki ışık saçılımını değiştirebilecek nesnelerden arındırılmış olarak gerçekleştirilmelidir.
- İsteğe bağlı olarak, bir mikro küreyi hapsedin ve kuvvet sinyalini kaydederken tuzağı bilinen bir hızda hareket ettiresiniz. Örneğin, üçgen salınım yapmak için tuzağı ayarlayın: kaydedilen kuvvet sinyali kare bir sinyal olacaktır.
NOT: Kuvvet değeri, boncuk üzerinde hareket eden sürükleme kuvvetine göre hız ile doğrusal olarak artmalıdır. Bu test, kuvvet ölçümlerinin doğru şekilde yapıldığının pozitif bir kontrolü görevi görür38. Alternatif olarak, optik kuvvet sensörü optik bindirme sertliğini, φ [pN/μm] ve [μm/V] β konum kalibrasyon faktörünü güç spektral analizinden elde etmek için kullanılabilir35. Doğru hizalama altında, üretici tarafından sağlanan sabit kalibrasyon faktörü α = φ·β [pN/V]' dir.- Üretici yazılımındaki Ölçüler alt menmenunda Çizim 1'e tıklayarak gerçek zamanlı bir kuvvet okuması başlatın. Bu, mevcut optik bindirme kuvvetinin ve gücünün okunmasını sağlayacaktır.
- Araçlar alt menüyünden Salınım Parametreleri iletişim kutusunu açın. Şekil ve Tür seçici halkalarında sırasıyla üçgen boşluk dalga biçimi şekli ayarlayın. Örnek olarak, 10 μm'lik bir genlik ve 3 Hz frekans ayarlayın. Bu, 1 μm38 çapında bir mikrobead üzerinde yaklaşık 1 pN viskoz bir kuvvetle sonuçlanacaktır.
- Kameranın AUX penceresinde, mikrobead'a sağ tıklayın ve Salınımı Başlat'ı seçin. Kuvvet okuması, ±1 pN'de platolarla kare kuvvet sinyaline dönüşecektir.
- Mikrobead'a sağ tıklayın ve Salınımı Durdur'u seçin.
7. Dönen disk konfokal mikroskopisi
- Dönen disk konfokal mikroskobu ve aksesuar ekipmanlarını, entegre lazer motorlarını ve alım kameralarını açın.
- Görüntüleme yazılımını başlatın.
- Hücre plazma zarı için çekirdek ve GFP'nin Hoechst lekesi için görüntüleme kanallarını ayarlayın.
- 405 nm ve 488 nm heyecan lazer hatlarını etkinleştirin.
- Eksize edilmeyi yansıtmak için çok bantlı bir dikroik ekleyin ve bu da yayılan ışığın kameralara geçmesini sağlar.
- Floresan emisyonu 500 nm uzunluğunda geçiş kenarı dikroik ayna ile bölün.
- sırasıyla iki alım kamerasının önünde DAPI/BFP (~445 nm) ve GFP (~521 nm) emisyon filtrelerini kullanın. Şekil 2F,G'ye bakın.
- Pozlama süresini her kanal için 100 ms olarak ayarlayın.
- Örnek düzlemde 5 mW güç elde etmek için lazer emisyonu ayarlayın. Gücü ölçmek için ticari bir güç ölçer kullanın.
- Görüntüleme protokolünü ayarlayın. Hoechst kanalından GFP kanalına spektral kanamayı önlemek için, iki boyanın ardışık olarak görüntülenmesi gerekir.
NOT: Optik yakalamanın AOD'leri ile kamera alımı arasında bir donanım senkronizasyonu varsa, tetik polaritesinin doğru ayarlendiğinden emin olun. Şüpheniz varsa, tesis yöneticinize veya mikroskop üreticinize danışın.
8. Çekirdek girintisi deneylerinin gerçekleştirilmesi
NOT: Kuvvet sensörü modülunu kaldırırken ve numuneyi değiştirirken hem yazılım kullanarak hem de epifluoresans bağlantı noktası 2 üzerindeki deklanşörü kapatarak optik tuzakları her zaman kapatın. Değilse, optik elemanlara ve deneyciye ciddi hasarlar oluşabilir. Lensin sahne/kültür çanağında çarpmaması için hücre ararken lens tutucu ile alt çanak kenarı arasındaki yanal mesafeye dikkat edin (Şekil 2).
- Örneği mikroskopa yerleştirin ve bu iletişim kuralının 5.3 adımını izleyin.
- Dönen HWP 'yi (Şekil 2F) kullanarak, araştırılan çekirdeğin veya hücre içi yapının sertliği bilinmiyorsa, tuzak gücünü başlangıç değeri olarak 200 mW'a ayarlayın. 6,5 adım boyunca ilk momentum taban çizgisini telafi etmek için OTs çalışma alanını (mikroskop aşamasını kullanarak) hücrelerden arındırılmış bir yere çevirin.
NOT: Hücre altı yapının sertliğine bağlı olarak, tuzak güç değeri benzer bir girinti derinliği elde etmek için daha düşük veya daha yüksek değerlere ayarlanmalıdır. - Mikroskop aşaması yazılım denetleyicisini kullanarak, iletilen brightfield mikroskopisi aracılığıyla bir veya iki boncuklu bir hücre arayın (Şekil 3A).
- Bir tuzak yörüngesi tanımlayın.
- Araçlar alt menüsündeki Yörünge iletişim kutusunu açın ve Yörünge Türü seçici halkasında Yer Değiştirme'yi seçin.
- Sayısal sayfaya, sonraki her yörünge adımının yer değiştirmesini ve zamanını yazın. İşte iki örnek.
- Bir stres gevşeme deneyi için, Şekil 3B'de gösterildiği gibi program yamuk yükler. Tablo S1'de, 5 μm seyahat mesafesi ile iki yamuk girinti uygulandı; 5 μm/s hız; geri çekilmeden önce bekleme süresi: 10 s.
- Çekirdek üzerinde uzun süre kalmadan üçgen bir rutin elde etmek için sabit hızda tekrarlayan bir girinti deneyi için, yörünge genliğini ayarlayın, örneğin, 5 μm ve adım için süre, örneğin, 2,5 μm / s hız için 2 s. Tablo S2'de, bu aynı hızda sekiz kez uygulanır.
NOT: Bu değerlerin her hücre tipi ve deney için belirlenmesi gerekir, ancak bir yamuk rutininin aşağıdaki parametreleri burada sunulan deneydeki en önemli dinamikleri yakalar. Bekleme süresi, çekirdeğin girintiden sonra tam stres gevşemesini göstermesi için yeterli olmalıdır.
- Mikro küreyi bindirme
- Mikroskop aşaması yazılım denetleyicisi ile görüntü düzlemini boncuk üzerinde biraz ayarlayın.
- OTs yazılımını kullanarak tuzakları etkinleştirin ve kamera AUX görüntüleme penceresindeki boncuk üzerine tıklayın (adım 6.4'ü izleyerek kalibre edildi). Boncukların optik tuzak tarafından başarılı bir şekilde hapsedilmesi, boncuk hareketini güçlü bir şekilde azaltacaktır.
- Boncukları sitoplazm boyunca tıklayıp sürükleyin ve nükleer zarftan ~2 μm uzaklıkta yerleştirin (Şekil 3A). Yörüngenin boncuk girintisinin nükleer zara dik olacak şekilde ayarlı olduğundan emin olun.
- İsteğe bağlı olarak, tuzağın konum ölçümleri için gerekirse, bindirme sertliğini belirlemek için tuzağı boncuk boyunca tarayın, k [pN/μm]54, böylece Δxbead = -F/k (bkz. Tartışma). Bu protokolde kullanılan optik mikromanipülasyon modülü bu amaçla yerleşik bir rutine sahiptir.
- Araçlar alt menüsündeki Parçacık Taraması iletişim kutusunu açın.
- Tarama Yöntemi olarak taramak istediğiniz tuzağı ve Yüksek Frekans'ı seçin. Boncuk tarama ölçümü için girinti yörüngesinin yönünü (x veya y) seçin.
- Bindirme sertliğinin ölçümü ile bir pencere görünecektir. Grafikte, F = -kx'e karşılık gelen doğrusal bindirme alanını seçmek için iki imleci sürükleyin. Seçili veri bölümüne doğrusal uyum otomatik olarak yenilenir.
NOT: Orta hücreli arayüzdeki ışık momentum sapmaları kuvvet ölçümlerinin uygunluğunu etkilediğinden, boncukun başlangıç konumunu hücre zarından (~5 μm) uzağa ayarlayın. Çekirdek hücre zarı çok yakın bir yerde bulunuyorsa, çekirdeği karşı bölgeden girintilemeye çalışın. Mümkün değilse hücreyi atın.
- Görüntüleme yazılımındaki alma düğmesine tıklayarak görüntü almaya başlayın.
- Veri | tıklayarak bindirme konumunu başlatın ve ölçüm verilerini kaydetmeye zorlayın Gerçek zamanlı kuvvet okuma penceresinde tasarruf edin (adım 6.6.1'de olduğu gibi açılır).
NOT: Optik tuzak, kameranın zamanlama çıkışına bağlanabilen bir tetik girişi ile donatılmıştır. Böylece, görüntü ve kuvvet verileri donanımla senkronize edilir ve elektronik, bindirme döngülerini alma sırasında görüntülerin kare sayısıyla eşleyebilir. - Boncuk üzerine sağ tıklayarak ve Yörüngeyi Başlat'ı seçerek önceden yüklenmiş yörüngeyi başlatın.
- Yörünge bitene ve sistem dengelenene kadar bekleyin.
- Tuzak kuvveti ölçüm veri tasarrufünü durdurun. Veri kaydetme iletişim kutusu açılır.
NOT: Veri depolamayı en iyi duruma getirmek için, bu iletişim kutusundaki (10, 100 veya 1000) yok edici parametre seçilerek veriler yok edilebilir. - Görüntü alımını durdurun ve sonuçları kullanıcının seçtiği işlemeyin yazılımında çizin.
- Mikroküreği rutin sırasında kaybolursa ve çekirdek girintilenemezse (Şekil S2), ölçümü atın ve gücü artırın. 6.5 adımının yinelenmeli olduğunu unutmayın. Elimizde, rutinlerin en az% 95'i tuzaktan boncuk kaybetmeden başarıyla tamamlanır.
Sonuçlar
Tuzak boncuklarının mikroenjeksiyon:
Tek hücreli zebra balığı embriyosuna enjekte edilen mikroküreler morfogenez sırasında tüm hayvan kapağına yayılır. Daha net bir görselleştirme için enjeksiyon protokolünü kırmızı floresan mikrobeadlarla tekrarladık ve farklı gelişim aşamalarında konfokal mikroskopumuzla hacimsel görüntüler çektik. Şekil 4A-D'de enjekte edilen boncuklar progenitör kök hücrelerin sitoplazmında 5 hfp'de in vivo olarak görselleştirilir. Daha sonra, mikroküreler 24 hpf'de tüm embriyoya yayılmış olarak ortaya çıktı (Şekil 4E). Her iki aşamadaki embriyolar normal olarak gelişti ve sağkalım oranları enjekte edilmeyen veya sahte enjekte edilen embriyoların kontrolü ile karşılaştırılabilirdi (bkz. Şekil S3). Bu, boncuk enjekte edilen zebra balıklarının döllenmeden 5 güne kadar bozulmamış hayatta kalmalarını bildiren diğer çalışmalarla tutarlıdır55.
İplik diski konfokal mikroskobumuz çok kanallı floresan mikrocopisi ile uyumludur. Şekil 5A'da sitoplazmda bir veya iki boncuk bulunan izole kök hücreler gösteriyoruz. Hücrenin farklı yönlerini araştırmak için birden fazla floresan etiket kullanılabilir (Şekil 5B). Nükleer morfoloji bir Hoechst boyası ile veya bir H2A::mCherry mRNA ifadesi kullanılarak izlenebilirken, iç nükleer membran Lap2b-eGFP12 ile analiz edilebilir. Aktomiyosin korteksinin dinamikleri ve hücre içi kalsiyum düzeyleri sırasıyla My12.1::eGFP transgenik line56 ve Calbryte-520 inkübasyonu ile gözlenebilir. Burada açıklanan protokol, yapışkan substratlar (daha sonra süspansiyon olarak adlandırılır) ve mekanik hapsetmede hareketsiz wildtype hücrelerinin hücre çekirdeği mekaniğini karşılaştırmayı amaçlamaktadır. 10 μm yüksekliğindeki mikro şarampollerde sınırlı olan izole kök hücreler, iç nükleer zarın (INM) kısmi açılmasını ve akromiyosin kontraktilitesinde daha sonra bir artış gösterdi12. Şekil 5C'de sitoplazmada bir veya iki boncuk bulunan sınırlı hücreler gösterilmiştir. Başarılı hapsetme, çekirdeğin daha geniş bir kesitine sahip düzleştirilmiş, genişletilmiş hücreler aracılığıyla görülebilir. Nükleer membran sınırlı hücrelerde daha da açılır ve süspansiyondaki hücrelere kıyasla yumuşatılmış görünmelidir (Şekil 5C).
Kuvvet-zaman ve kuvvet-deformasyon analizi
Elde edilen sonuçların analizi, araştırılan örneğe ve ilgi sorusuna bağlıdır ve bu nedenle burada genelleştirilemezler. Örnek olarak, girinti ölçümünü analiz etmenin yaygın bir yolu, değiştirilmiş bir Hertz modelini kuvvet girintisi verilerine sığdırarak bir Young modülü çıkarmaktır57. Bununla birlikte, böyle bir tedavi için varsayımın dikkatlice değerlendirilmesi gerekir ve her zaman düzgün bir şekilde haklı olmayabilir (araştırılan yapının izotropik, homojen, doğrusal elastikiyet ve girintilerin boncuk yarıçapından daha küçük olması gibi). Bu nedenle, burada sadece araştırılan yapının mekanik davranışının farklı deneysel senaryolar arasında karşılaştırılmasını sağlayan model bağımsız ölçümlerini dikkate alıyoruz.
Başlangıç noktası olarak, kuvvet-yer değiştirme eğrisinin eğimini belirli bir girinti derinliğinde ölçmek, çekirdeğin model bağımsız yapısal sertliğinin bir ölçüsünü sağlar58 . Bu değer daha sonra birden fazla örnekten toplanabilir ve değişen deneysel ayarlar ile örnek pertürbasyonları arasında karşılaştırılabilir.
Girinti ölçümü
Aşağıdaki satırlarda, hücre deformasyonu sırasında hücre çekirdeğinin hücre hapsinde mekanik tepkisine odaklanıyoruz. Bu protokolün 8. Bununla birlikte, bu değerler hücre tipine ve deneysel koşullara bağlı olarak büyük ölçüde farklı olabilir ve daha yumuşak çekirdekler belirli bir girinti için daha düşük kuvvete yol açabilir. Böylece, hücre çekirdeğinin doğru bir mekanik karakterizasyonu için nükleer deformasyonun kuvvetle birlikte doğru bir şekilde ölçülmesi gerekir. Bu bölümde hücre nükleer sertliğini temsili kuvvet girintisi ölçümlerinden elde edeceğiz.
Şekil 6'da, askıda ve kapalı bir hücrede çekirdeğin distal ve proksimal kenarlarındaki deformasyonları gösteriyoruz. Zengin bir mekanik davranış gözlenebilir. Yapışkan bir substrat üzerindeki tipik bir askıya alınmış hücrede, çekirdek boncuk tarafından güçlü bir şekilde girintilenmiş, aynı zamanda tekrarlayan itme olayları üzerine hafifçe yer değiştirmiş. Hoechst lekeli hücre çekirdeklerinin floresan görüntülemesinden elde edilen kimografları analiz ederek çekirdek üzerindeki boncuk girintisini ölçtük. Kymographs, girinti yönü boyunca Fiji'nin Multi Kymograph eklentisi kullanılarak kolayca hesaplandı (Şekil 6A,B) ve daha fazla işlem için Matlab'a (Sürüm 2021, Mathworks) ithal edildi. Girinti rutininin yörüngesi boyunca çekirdeğin sınırlayıcı kenarlarını izlemek amacıyla ham yoğunluk profiline bir adım fonksiyonu takıldı. Görüldüğü gibi, nükleer şekil değişikliği hakkında doğru bilgiler taşır (Şekil 6 ve Şekil S2). Bir adım işlevinin analitik bir sürümü olarak aşağıdaki çift sigmoid eğrisini kullandık:
 (Denklem 1)
(Denklem 1)
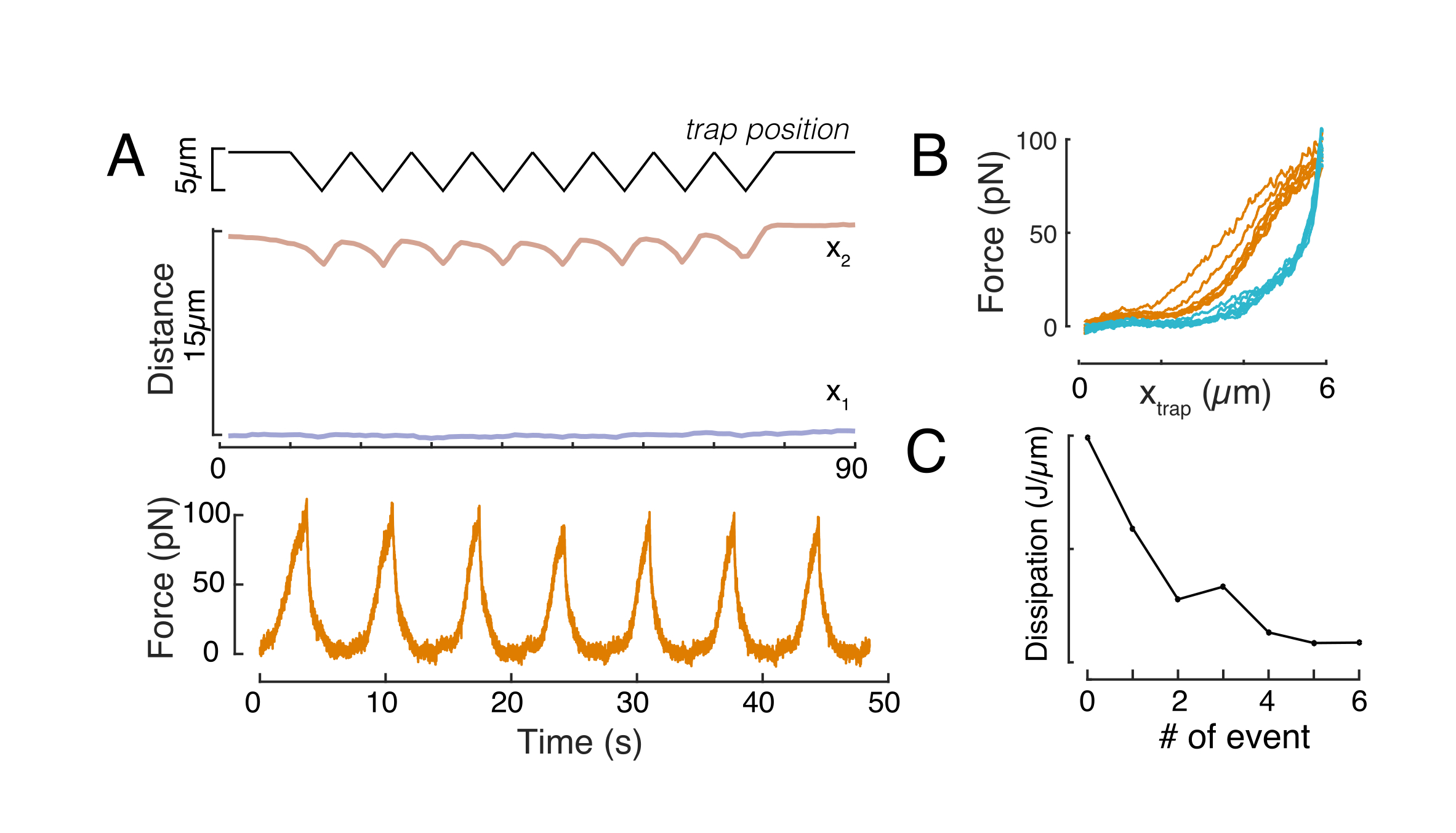
Burada x1 ve x2 çekirdeğin distal ve proksimal kenarlarını gösterirken, A ve B görüntünün mavi kanalının (Hoechst boyası) maksimum ve arka plan gri değerleridir (Şekil 6B). Kenar genişliği göz önünde bulundurulmuştır (e0 = 0,25 mm). Girintili, proksimal çekirdek kenarı (x2) mikroküreği-çekirdek temasından sonra optik bindirme rutini tarafından uygulanan yörüngeyi takip ederken, tam tersi, distal kenar (x1) sitoplazma gibi viskoelastik bir malzeme için beklendiği gibi gevşeme dinamikleri görüntüler (Şekil 6D). Buna karşılık, 10 μm yüksekliğinde mikroçaplarla sınırlı hücrelerdeki çekirdekler, hücre içindeki girinti üzerine çekirdeğin bu tür translokasyon davranışını sergilemez (Şekil 6B,D). Ayrıca Şekil 6D'de gösterilen çekirdeklerin arka kenarları, büyük olasılıkla hücre konttilasyonundan ve girinti kuvvetine karşı hareket eden sürtünmeden kaynaklanan daha güçlü kuvvetler nedeniyle, proksimal taraftan itilen boncuk tarafından değişmeden kalır. Doğru deformasyon derinliğini elde etmek için, x1 deplase girintili ölçü x2'den çıkarılır: Δx = x2 - x1 (ayrıca bkz. Şekil 6D).
Veri çözümlemesine zorla
Nükleer deformasyona neden olan kuvvet, optik olarak sıkışmış mikroptan kaynaklanan ışık momentumundaki değişimden ölçtürüldüğünde (Şekil 7A). Yamuk yörüngeler (adım 8.4.3, Şekil 7B) uygulandıktan sonraki kuvvet başlangıçta tuzak hareket etmeyi bırakana kadar doğrusal olarak arttı, ancak daha sonra sabit bir durum değerine gevşedi. Bu davranış, kayıp ve depolama modülü sergileyen viskoelastik bir malzemeyi gösteriyordu. Girinti olayından hemen sonra, kuvvet bir zirve değeri olan Fp'ye ulaştı ve ardından bir stres gevşemesi (Şekil 7C):
 (Denklem 2)
(Denklem 2)
burada F0 elastik bileşen için depolanan kuvvettir ve f(t) boyutsuz bir gevşeme işlevidir. Bu davranışı üç şekilde analiz ettik:
1. Üstel stres gevşemesi olan standart doğrusal bir katı göz önüne alındığında, yani, f(t) = e-t/φ, Şekil 7C inset'te şematik olarak temsil edilir.
2. Genel, çift üstel bir çürüme kullanarak:
F(t) = A + B1e-t/φ1 + B2e-t/φ2.
3. Üstel bir çürümenin ardından bir güç yasası kullanmak59:
f(t) = t-pe-t/φ, Şekil 7C'ye monte edilmiştir.
Model 1 için uygunluk basit bir şekilde gerçekleştirilebilirken, sırasıyla 2 ve 3 modelleri için (φ1, φ2) ve (p, φ) için ilk tahminleri tahmin etmenizi öneririz. Bu, sırasıyla, logaritmik ve doğrusal (Şekil 7D, sol) ve logaritmik-logaritmik (Şekil 7D, sağ) ölçeklerinde verilere çizgiler sığdırarak gerçekleştirilebilir. Tablo S3, Şekil 7'de analiz edilen örneğin sonuçlarını özetler. Aşağıdaki bölümde, hücre çekirdeği mekaniğinin karakterizasyonu için bir güç yasası ve üstel bir yasanın kombinasyonunu ele alacağız.
Yer değiştirme ilişkisini zorla
Aynı şekilde, açıklanan deneysel kurulum, birden fazla girinti olayının kuvvet-yer değiştirme ilişkisini elde etmek için kullanılabilir. Üçgen rutinler gerçekleştirerek (adım 8.4.4, Şekil 8A), kuvveti deformasyonla ilişkilendirmek ve bir kuvvet girinti eğrisi çizmek mümkündür. Boncuk çekirdeğe temas ettikten sonra düz bir taban çizgisinin eğimi sorunsuz bir şekilde değiştirdiği Şekil 8B'de örnek bir sonuç gösterilmiştir. Gürültülü verilerdeki gerçek temas noktasını belirlemek zordur ve temas bölgesinin elastik modellere uygun olup olmadığını görmek için dikkatli olunmalıdır60. Bu özel deneyde, sonraki girintilerin daha derin temas noktalarına sahip eğrilere, boncuk geri çekilmesinden sonra çok yavaş nükleer şekil iyileşmesine işaret eden ve çekirdek viskoelastik malzeme özellikleri tarafından tanımlanan histerik döngüde bir değişikliğe neden olduğu da görülebilir (Şekil 8C). Bu nedenle, araştırmacı bunun olup olmadığının farkında olmalı ve bunu analitik işlem hattına dahil etmeli veya sonraki ölçümlerin sayısını bu etkinin ölçümü değiştirmemesi için kısıtlamalıdır.
Süspansiyondaki ve 10 μm'nin altındaki hücrelerde çekirdek mekaniği
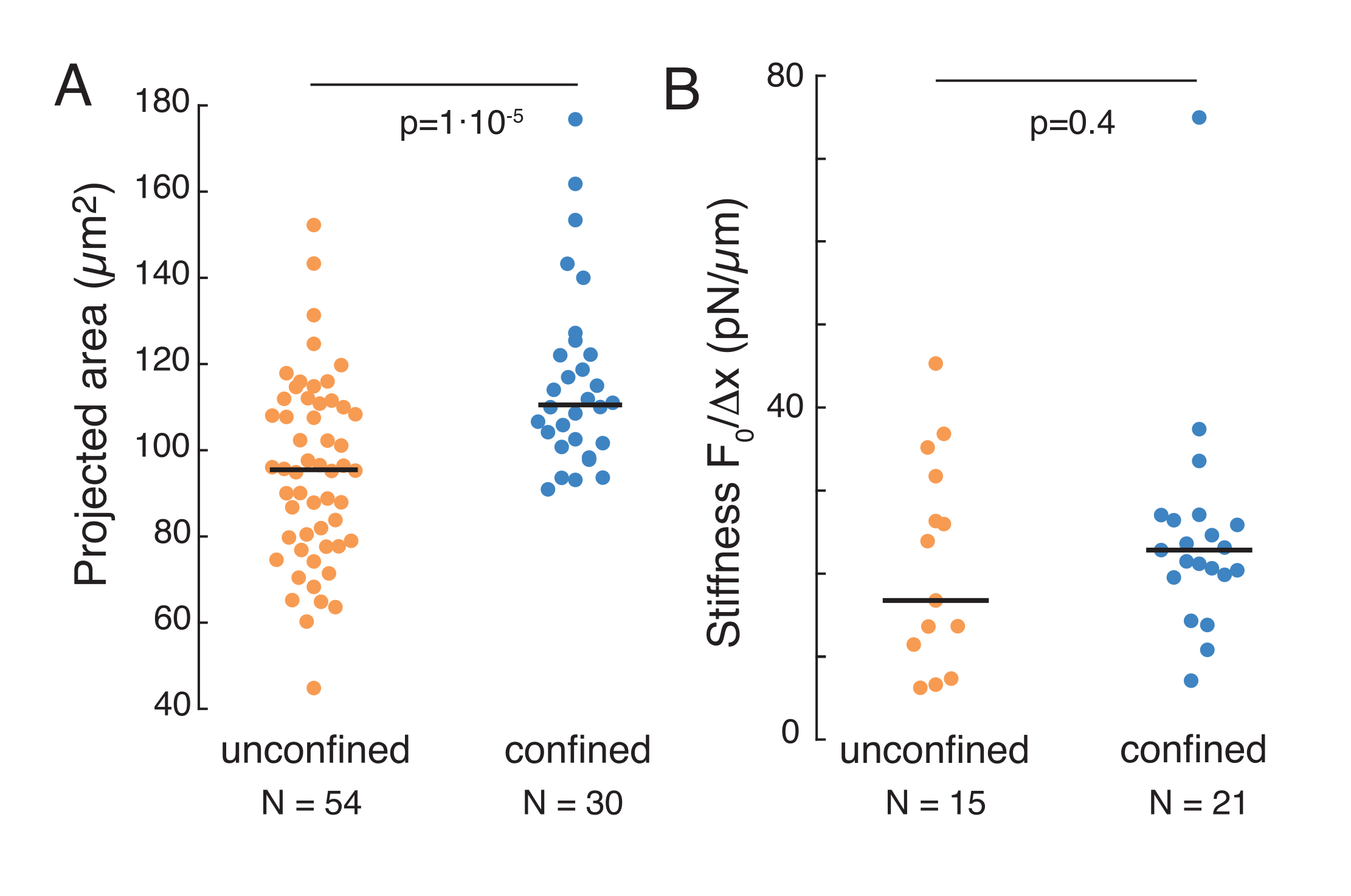
Yukarıda belirtilen yaklaşım, yapışkan substratlar ve sınırlı hücreler üzerindeki askıya alınmış hücrelerde çekirdek stresi gevşeme dinamiklerini analiz etmek için kullanılmıştır. Sonuçlarımız, hapsürün öngörülen alanın genişlemesiyle sonuçlendiğini göstermektedir (Şekil 9A), ancak nükleer sertlikte önemsiz bir değişiklik (Şekil 9B). Benzer gevşemeyi 6,08 ± 1,1 sn (sınırlanmamış) ve φ = 4,00 ± 0,6 sn (hapsetme) ile ölçtük, bu da hızlı viskoelastik dağılım olduğunu gösterir, ardından çekirdeğin elastik modülüne karşılık gelen depolanmış bir kuvvet değeri. Girinti rutinlerindeki farklı başlangıç koşulları tarafından üretilebilen deneysel varyasyonları hesaba katmak için, ölçülen depolanmış kuvvetler girinti derinliğine normalleştirildi.  Bu parametre çekirdek sertliğini açıklar ve belirli bir girinti için gerekli olan kuvveti veya stresi açıklar. Benzer sertliği hapsetme altında ve sınırlanmamış hücrelerde elde ettik: = sırasıyla 20.1 ± 12.6 pN/μm ve = 24.6 ± 13.6 pN/μm (ortalama ± standart sapma).
Bu parametre çekirdek sertliğini açıklar ve belirli bir girinti için gerekli olan kuvveti veya stresi açıklar. Benzer sertliği hapsetme altında ve sınırlanmamış hücrelerde elde ettik: = sırasıyla 20.1 ± 12.6 pN/μm ve = 24.6 ± 13.6 pN/μm (ortalama ± standart sapma).

Şekil 1: Zebra balığı embriyolarının tek hücreli (zigot) aşamada mikroenjeksiyonu. (A) Enjeksiyon plakası: Enjeksiyon için üçgen şeklinde bir enjeksiyon plakası kullanılır. Plaka E3'te (Yumurtanın ortası)% 1 ultra saf agarosedan yapılmıştır. Üst ve yan görünümler sağda gösterilir. (B) Embriyo konumlandırma: embriyoları bir fırça kullanarak hafifçe yönlendirin ve tek hücrenin iğne ile açıkça görülebileceği ve kolayca erişilebileceği şekilde yönlendirin. Embriyoları, çizimde gösterildiği gibi iğnenin karşı tarafında bulunan hücre ile yönlendirmeyi öneriyoruz. (C) Tek hücreli evre embriyoya enjeksiyon prosedürü: embriyoyu çevreleyen korona ve tek hücreyi iğne ile delin. İğnenin ucunun hücrenin içinde olduğundan emin olun ve enjekte etmek için basıncı serbest bırakın. (D) Embriyoları blastula (küre) aşamasına (4 hpf) kadar gelişene kadar 28-31 °C'de kuluçkaya yatırın. Hücre izolasyon protokolünü ve hücre boyama işlemini gerçekleştirin (adım 2) ve optik bindirme odasını, ilgili substrat yüzey kaplaması (adım 3) ile birlikte süspansiyon ve/veya hapsedilmiş izole hücrelerle hazırlayın. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 2: Optik cımbız aparatının hazırlanması. (A) Cam alt tabaklara tanımlanmış bir yüksekliğe sahip PDMS'nin spin kaplama katmanları. PDMS düşüşü, santrifüj kuvveti nedeniyle eşit olarak yayılacak. (B) Numune odasının PDMS katmanından hazırlanması. 1: neşterli bir kare kesin, 2: iç kuyuyu concanavalin A (ConA), yıkama ve tohum hücreleri ile kapla; 3: kuyuyu kapatmak için cam bir slayt veya kapak kayması ile örtün. (C) Neşterle kesen ve PDMS'yi kümeslerle iyi bir şekilde çıkaran karenin resmi. (D) Optik kuvvet sensörünün toplama lensinin bindirme odasının üzerine monte edilir. Bir damla daldırma yağı, toplama lensi ile üst cam kapak arasında bir daldırma ortamı görevi görür. Ölçeklememek için şematik. Numune kabının cam kapağına dokunmamak için toplama lensini indirirken dikkatli olun. (E) Numune ile temas eden kuvvet algılama ünitesinin resmi. (F) Deneysel kurulumun şeması. Optik mikromanipülasyon modülü, yarım dalga plakası (HWP) ve polarize ışın ayırıcı (BS) aracılığıyla güç kontrolüne sahip sürekli dalga lazer ışını (5W, φ = 1064 nm) kullanır. Bir çift AOD ile modüle edildikten sonra, ters bir mikroskobun üst epifluoresans bağlantı noktasına bağlanır. Lazer ışını daha sonra 950 nm kısa geçişli dikroik ayna (IR-DM) ile yansıtılır ve floresan eksilme ve emisyonun iletilmesini sağlar. Bindirme lazeri mikroskobun (üst kule) arka, epifluoresans portuna yönlendirilir. OT'ler, suya batırma hedef lensinin odak düzlemsinde oluşturulur (60x, NA = 1.2). Optik kuvvet sensörü mikroskop kulesi tarafından tabi tutulur ve OT'lerden çıkan lazer ışığını yüksek NA, yağ daldırma lensi ile yakalar. Aynı zamanda, kuvvet sensörü parlak alan aydınlatması sağlar. Dönen disk konfokal ünitesi sol bağlantı noktasına bağlanmış. Yedi floresan heyecan lazerini kontrol eden iki entegre lazer motoru (İLE) ve iki arka aydınlatmalı sCMOS kamera ile donatılmıştır, paralel Abb'de çift florofor görüntüleme sağlar: TI, Transilluminator; FS, saha durdurma; AOD, a vesayetçi saptırıcı; HWP, yarım dalga plakası; CAM, kamera (G) Optik bindirme ekipmanının fotoğrafı. Kırmızı daire, manuel olarak optik yola geçilebilen Bertrand lensi gösterir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
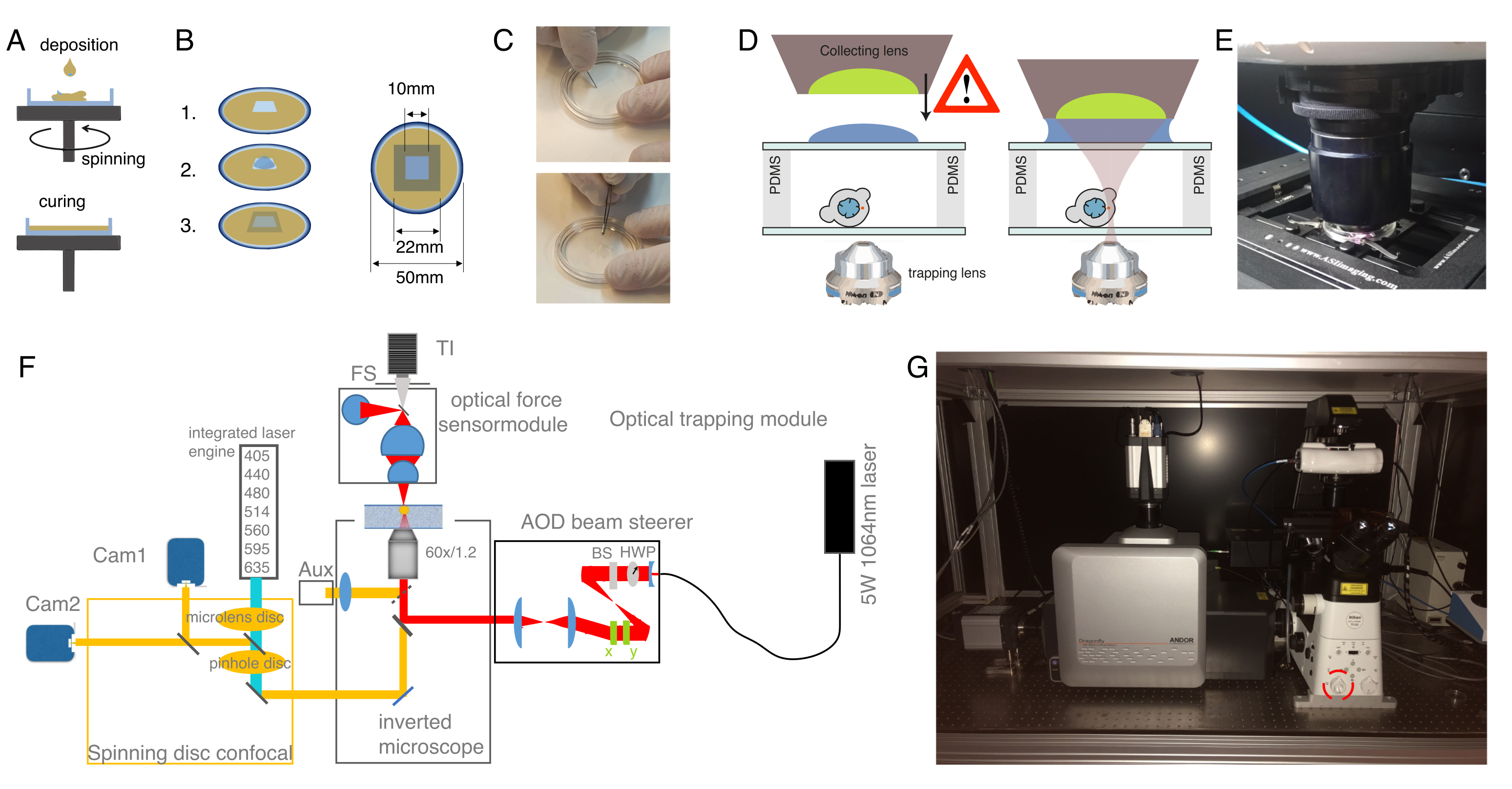{kind=link}

Şekil 3: Doğru örneklerin ve parametrelerin seçilmesi. (A) Girinti deneyini gerçekleştirmek için çekirdeğe yeterince yakın konumlandırılmış tek bir mikrokürece sahip izole zebra balığı progenitor kök hücresinin temsili görüntüsü. Ölçek çubuğu = 10 μm. (B) Örnek tuzak yörüngesi; girinti derinliği 5 μm; girinti hızı = 5 μm/s; gevşeme süresi 10 s. Bu rakamın daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
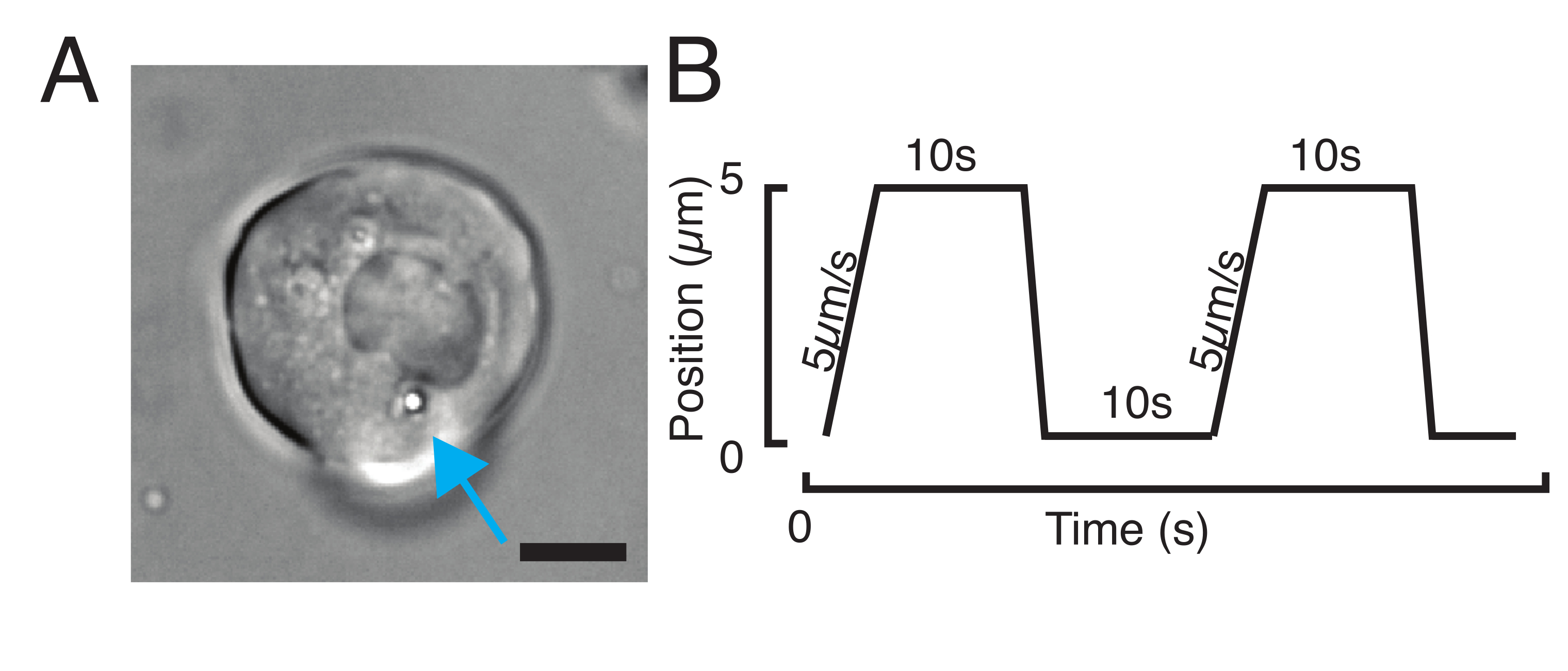{kind=link}

Şekil 4: Geliştirme sırasında zebra balığı embriyolarının içinde mikrobead lokalizasyonu. Boncuk lokalizasyonlarını görselleştirmek için WT embriyolarında 1 μm kırmızı floresan boncukların 0,5 nL'si GPI-GFP mRNA (100 pg/embriyo, plazma membran) ile birlikte enjekte edilir. (A-D) %0.75 agarose monte edilmiş bir embriyonun içine mikrosfer 5 saat enjeksiyon sonrası dağılımı. (A) Brightfield ve floresan görüntü. Boncuklar, konfokal mikrografide görüldüğü gibi embriyo dokusuna homojen bir şekilde dağılır. (B) Konfokal floresan z-yığınının maksimum projeksiyonu. Boncuklar, görüntü yığınındaki z konumlarına göre mordan sarıya renk kodludur. Mor/macenta en dış boncuklara/hücrelere (EVL; epitel saran tabaka; veya EVL yüzeyine yakın bulunan progenitör kök hücrelere) karşılık gelir, sarı sağdaki çizimde gösterildiği gibi iç boncuklara (progenitör derin hücreler) karşılık gelir. (C) Turuncu kutudaki bölgeye karşılık gelen bir (B) alt yığınının kesilmesi ve maksimum projeksiyonu: derin hücrelerin büyük bir kısmı 1-2 boncuk içerir. (D) Macenta kutusuna karşılık gelen bir (B) alt yığınının kesilmesi ve maksimum projeksiyonu: bazı EVL hücreleri 1-2 boncuk içerir. (E) Brightfield görüntüsü ve %0,75 agarose monte edilmiş ve trikain ile uyuşturulmuş 24 hpf embriyonun z-yığınının maksimum projeksiyonu. Embriyolar 15 dakika boyunca trikain ile ön kuluçkaya alındı. Soldan sağa: mikroküreler (1 μm çap), GPI-GFP ve görüntü üst üste biniyor. Boncuklar embriyonun tüm vücuduna dağılmış. Her panelde belirtilen ölçek çubuğu boyutu. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
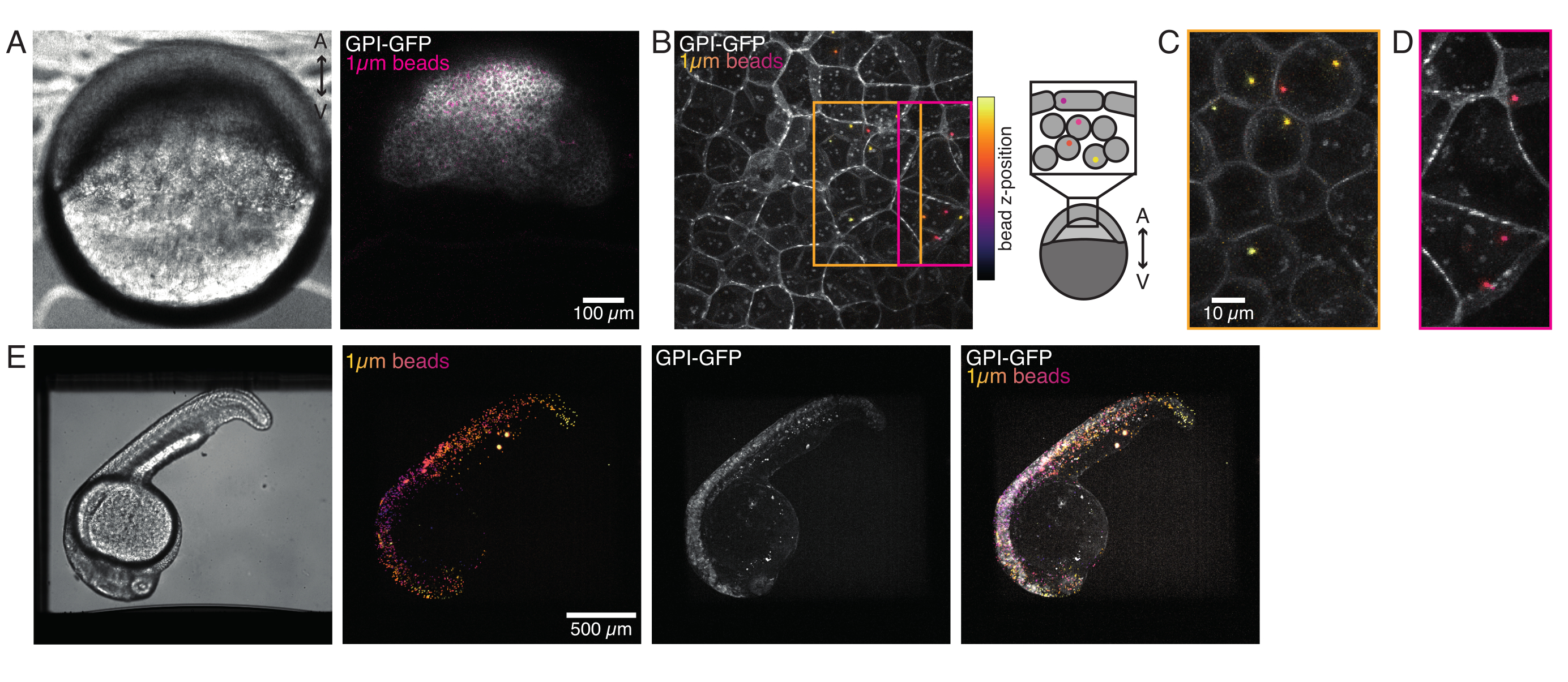{kind=link}

Şekil 5: Farklı etiketlemeye sahip izole zebra balığı progenitor kök hücreleri. (A) 1 (üst) veya 2 (altta) enjekte boncuklu süspansiyon hücrelerinin iletim ışığı mikroskopi görüntüsü. Siyan okları boncukları işaret eder. (B) Farklı boyamalara sahip süspansiyon hücrelerinin floresan konfokal görüntüleri. Sol üst: Lap2b-eGFP (iç nükleer membran, 80 pg/embriyo) ve H2A-mCherry. Sağ üst: GPI-GFP (plazma membran, 100 pg/embriyo) ve DNA-Hoechst (bölüm 2'de açıklandığı gibi lekeli). Sol alt: MyI12.1-eGFP (transgenik çizgi) ve DNA-Hoechst. Sağ alt: Calbryte488 ve DNA-Hoechst (bölüm 2'de açıklandığı gibi lekelenmiş). (C) 1 (üst) veya 2 (alt) enjekte boncuk ile sınırlı hücrelerin iletim ışığı mikroskopi görüntüsü. Siyan okları boncukları işaret eder. Ölçek çubukları = 10 μm. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 6: Dönen disk filmlerinden nükleer deformasyonun tahmin edilmesi. (A,B) (A) askıya alınmış bir hücredeki çekirdeğin ve (B) kapalı bir hücredeki çekirdeğin girinti denemesinin zaman atlamalı. Ölçek çubuğu 10 μm. Hoechst etiketli bir çekirdeğin temsili anlık görüntüleri, optik olarak sıkışmış bir mikrokürece (beyaz ok ucu) ile girintiden önce, sırasında ve 5 sn sonra 5 s gösterilir. Girinti segmenti boyunca kimograflar (kırmızı çizgi, sağ panel). x1 ve x2, yoğunluk profilinin denklem 1'e sığdırılmasından çıkarılan girinti deneyi sırasında çekirdeğin distal ve proksimal (boncuka yakın) sınırlarıdır. (C) Üç farklı kare için girinti segmenti boyunca yoğunluk profilleri (girinti öncesi, sırasında ve sonrasında) ve çekirdek kenarlarının distal, x1 ve proksimal, x2 konumlarını değerlendirmek için Denklem 1'e takılır. (D) Askıda ve kapalı hücrelerin (10 μm) girinti deneyi sırasında mavi renkte x1(t) ve kehribarda x2(t) temsili yörüngeler. Gölgeli alanlar girintiyi, x1 ile x2 arasındaki mesafe çekirdeğin çapını gösterir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
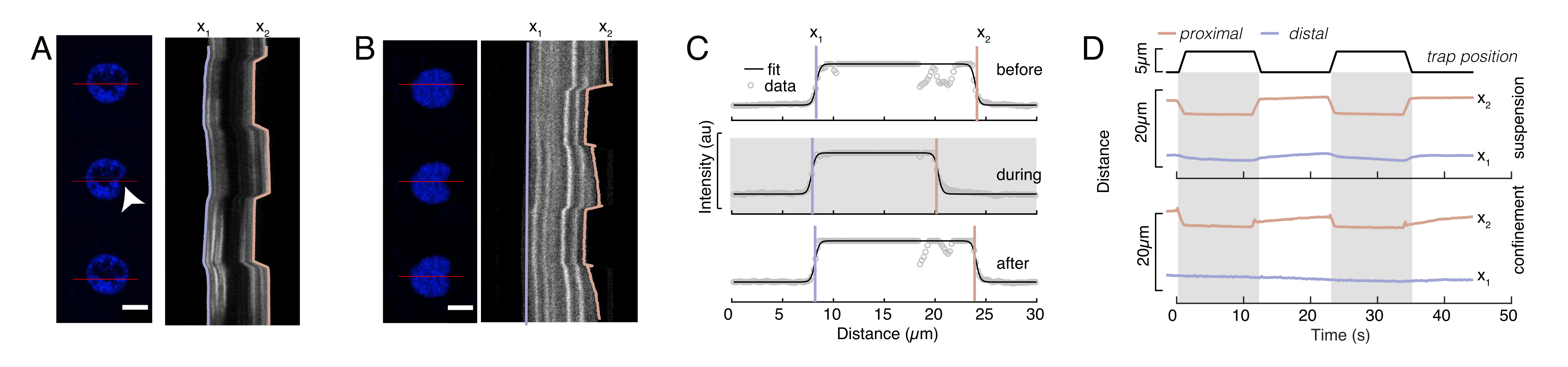{kind=link}

Şekil 7: Güç sinyali işleme. (A) Girinti üzerine hücre çekirdeğini deforme eden optik olarak sıkışmış bir mikro kürenin şeması. Nükleer membran ve optik kuvvetler siyah oklarla gösterilir. Işın momentumundaki değişim yeşil ok Somurtması ile gösterilir. (B) Tekrarlanan bir nükleer girinti deneyi sırasında optik olarak sıkışmış mikro kürenin yaşadığı tuzak yörüngesi (üst) ve kuvvet (alt). (C) Maksimum girinti derinliğinde kuvvet zirvesinden sonra gevşeme çürümesini zorlayın. Inset, dinamikleri buradaki fenomenolojik gözlemlere yaklaşık olarak sahip standart doğrusal katının şemasını gösterir. (D) Sol: normalleştirilmiş kuvvetin logaritm ve zamana karşı. Gölgeli alanlar, çift üstel bozunmaya (kırmızı çizgiler) uymak için kullanılan veri bölümünü gösterir. Doğru: normalleştirilmiş kuvvetin logaritm'i ile zamanın logaritmının. Gölgeli alan, güç yasasına uymak için kullanılan veri bölümünü gösterir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 8: Üçgen tuzak yer değiştirmeleri ile girinti rutinini zorlayın. (A) 10 μm hapsetme yüksekliğindeki bir hücrede yapılan üçgen girinti deneyi sırasında mavi renkte x1(t) ve kehribarda x2(t) temsili yörünge. Üst: Tuzak konumu. Orta: Çekirdek şekli analizi. x1 ve x2 arasındaki mesafe çekirdek çapını gösterir. Alt: Kuvvet sinyali. (B) Ardışık sekiz girinti için kuvvet ve tuzak pozisyonu. (C) F-d eğrisinin yaklaşım ve çekilme kısmı arasındaki histegenezden türetilen dağılımın evrimi, sonraki her girinti olayı için çekirdeğin. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 9. Süspansiyondaki hücrelerin nükleer özellikleri (yapışkan yüzey) ve yamuk rutinlerden hapsedilir. (A) Çekirdeğin süspansiyonlu ve 10 μm'nin altındaki hücrelerden öngörülen alanı. Siyah çubuk ortancayı temsil eder. (B) Süspansiyondaki ve hapsedilen hücrelerin nükleer sertliği. Siyah çubuk ortancayı temsil eder. MatLab kullanılarak Kruskal-Wallis testinden elde edilen P değerleri. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
Ek Tablo 1: Optik cımbız yazılımı tarafından tanımlanan trapez yörüngesi. birinci (ikinci) satır, tuzağın doğrusal olarak yer değiştirileceği x (y) mesafesidir. Üçüncü satırda, belirli bir adımın süresi saniye cinsinden ayarlanır. Bu yörünge yedi noktadan oluşur ve Şekil 7B'deki çekirdeğe karşı iki kez yüklenen yamuklara karşılık gelir. Bu Tabloyu indirmek için lütfen tıklayınız.
Ek Tablo 2: Optik cımbız yazılımı tarafından tanımlanan üçgen yörünge. Tablo 2'ye benzer şekilde, bu yörünge 5 μm derinlikte sekiz girinti olayına ve 2,5 μm / s hıza karşılık gelen 16 noktadan oluşur.
Ek Tablo 3: Şekil 7'deki veriler için parametrelerin takılması. IG: ilk tahmin. Bu Tabloyu indirmek için lütfen tıklayınız.
Ek Şekil S1: Optik kuvvet sensörü hizalaması ve momentum taban çizgisi telafisi. (A) Bertrand objektifinden yardımcı kamerada (AUX, Şekil 2) alan durdurma görüntüsü. Daldırma yağının içinde gözle görülemeyen bir hava kabarcığı görünür. (B) Optik yolu temizleyin. Doğru hizalama için, alan durağını açın ve NA = 1.2 ışık konisi ile çakışmasını sağlayın. (C) Örnek düzlemin görüntüsü. Kırmızı kare OT çalışma alanını gösterir. Ölçek çubuğu: 20 μm. (D) FOV boyunca ölçülen tuzak gücü, C ile belirtilen beyaz çift oklar boyunca. Kırmızı renkte, düzeltme uygulanmadığında güç değişimlerini hapsedin. Mavi, tuzak gücü tüm görüş alanı üzerinde düzeltildi. (E) Momentum taban çizgisinin X bileşeni aynı aralık boyunca. Kırmızı, düzeltilmemiş iz. Mavide, izleme tuzak gücü için düzeltildi. Yeşil olarak, izleme, üreticinin yazılımındaki Genel Ofset Telafisi kullanılarak momentum temeli için düzeltildi. (F) E ile aynı, Y bileşeni için. Normal çalışma altında, gölgeli bileşenlerin mekanik ve kuvvet ölçümleri için kullanıldığını unutmayın, örneğin x koordinatı boyunca hareket sırasında x kuvvet bileşeni ve y ekseni boyunca hareket sırasında y kuvvet bileşeni. Tüm düzeltmeler uygulandıktan sonra, <0,5 pN RMSD gürültüsü elde edilir. Bu Dosyayı indirmek için lütfen tıklayınız.
Ek Şekil S2: Zayıf tuzaklar nedeniyle başarısız bir rutin. (A) Başarısız bir rutinden çekirdek girintisi gösteren kimograf. Boncukların tuzaktan kaçması nedeniyle sadece kısa, geçici deformasyonlar görülebilir. Daha da önemlisi, bindirme lazeri önceden tanımlanmış yörüngeyi (yeşil noktalı çizgi) tamamlamak için boncuksuz hareket etmeye devam eder. Ölçek çubuğu = 10 μm. (B) Üst: Tuzak konumu ve zamana karşı. Orta: Girintili proksimal ve distal çekirdek kenarının kenar izleme sonucu. Distal kenarın, yapışkan substratlardaki yalıtılmış hücrelerde tamamlanmış rutinler için yaygın olarak gözlemlendiği gibi girinti olmadan hareket etmediğini unutmayın. Alt: Termal gürültüde azalma ve ani bir düşüşle belirtilen mikrosferin kaybını gösteren zamana karşı kuvvet. Bu Dosyayı indirmek için lütfen tıklayınız.
Ek Şekil S3: Enjekte edilen embriyoların hayatta kalması. Protokolde belirtilen konsantrasyonlarda 1 μm boncuk ve 100 pg/embriyo mRNA ile enjekte edilen embriyolar, incelenmemiş embriyolarla karşılaştırıldı ve döllenme sonrası 24 saat önemli bir fark göstermedi. N ile yapılan üç bağımsız deneyin ortalama ve standart sapması, her deney için durum başına 21 embriyo >. Bu Dosyayı indirmek için lütfen tıklayınız.
Tartışmalar
Bu protokolde, hücre çekirdeğinin canlı hücrelerin içindeki mekanik özelliklerini sorgulamak için benzersiz bir yöntem açıklıyoruz. Diğer kuvvet spektroskopisi tekniklerinden farklı olarak, non-invaziv optik bindirme, hücre zarı ve sitoskeletonun hücre nükleer sertliğinden katkısını ayırmamızı sağladı. Daha da önemlisi, optik mikromanipülasyon multimodal mikroskopi ile uyumludur, bu da deneycinin hücre nükleer mekanobiyolojisinde yer alan farklı süreçleri incelemesine izin verecektir. Temsili bir sonuç olarak, birkaç yüzlerce picoNewton düzenindeki kuvvetler tarafından gerçekleştirilen girinti üzerine çekirdek deformasyonunu ölçmek için DNA-Hoechst lekesini kullandık.
Yöntemimizin bu protokolde belirtilen örneklerin ötesinde potansiyel uygulamaları
Canlı hücrelerin içindeki ölçümlerden, dış pertürbasyon olmadan nicel mekanik bilgiler çıkarma imkanı, yeni keşfedilmeye başlanan çok sayıda benzeri görülmemiş fırsat sağlar. Böylece, optik mikromanipülasyon platformumuzun sunulan protokolü, çok yönlülükle daha karmaşık deneylere genişletilebilir. Acousto optik deflektörler (AOD), farklı hücre konumlarında senkron kuvvet ölçümleri için birden fazla optik tuzak üretebilir ve geniş bir frekans aralığında aktif mikrorheoloji için kullanılabilir51,61. Belirtildiği gibi, girinti üzerine kuvvet tepkisi maksimum bindirme kuvvetinin üstesinden gelebilir ve boncukların optik tuzaktan kaçmasına yol açabilir. Bu durumda, optik kuvveti sıkıştırmak için AOD ile bir kuvvet geri bildirimi yapılandırılabilir. Sonuç olarak, bu protokolde açıklanan stres gevşemesi gibi birden fazla mikrorheolojik yaklaşım, aynı zamanda aktif mikrorheoloji veya sürünme uyumluluğu, bu platformla deneysel olarak elde edilebilir ve yeni yazılım paketleri tarafından iyice analiz edilebilir61,62,63,64,65 . Ayrıca, kuvvetlerin uygulanması çekirdekle sınırlı değildir, ancak prensip olarak çeşitli hücre içi yapıları ölçmek için ve bozulmamış kan damarlarının içine akan kırmızı kan hücrelerini hapsetmek için gösterildiği gibi karmaşık dokularda gerçekleştirilebilir66,67 veya kloroplastları ve mitokondrileri hapsetmek ve deforme etmek68 . Işık momentumu kalibrasyonu, sıkışan nesnenin şeklinden ve boyutundan bağımsızdır, bu nedenle rastgele şekle sahip herhangi bir kuvvet probunda doğrudan kuvvet ölçümleri sağlar38,39. Enjekte edilen mikroküreçlerin kullanımı, hücresel yapıların doğrudan manipülasyonuna kıyasla nispeten düşük lazer gücüyle çekirdeğe yüksek kuvvetler uygulamamızı sağladı69,70,71. Bununla birlikte, yeterince yüksek bir kırılma indeksi farkı göz önüne alındığında, dışarıdan uygulanan bir kuvvet probu gerekli değildir ve hücre içi organeller enjekte edilmiş boncuklar (yayınlanmamış gözlemler ve referans70) olmadan doğrudan manipüle edilebilir.
Uygulamaları genişletmek için yöntemimizin olası değişiklikleri
Deneye bağlı olarak farklı boyutlarda mikrobead enjekte edilebilir, ancak göreli kontroller yapılmalıdır. Örneğin, hücreleri daha sonraki aşamalarda incelemek için daha küçük boncuklar enjekte edilebilir. Bu, optik tuzak tarafından uygulanabilecek maksimum gücü azaltacaktır (referans55'te gösterildiği gibi). Daha yüksek kuvvetler uygulamak için daha büyük boncuklar enjekte edilebilir, ancak bunlar büyüklüklerine veya ilgi aşamalarına bağlı olarak embriyo gelişimini etkileyebilir. Mikrobead enjeksiyonunun bir seçenek olmadığı deneylerde, sitoplazma ile karşılaştırıldığında kırılma endeksleri farklılıkları gösteren çeşitli organeller hala optik olarak manipüle edilebilir ve ışık momentumu değişikliklerinden ölçülebilir optik kuvvetlere yol açabilir42. Yukarıda belirtildiği gibi, bu yöntemler Bambardekar ve ark. tarafından Drosophila embriyosunda hücre-hücre bağlantılarını deforme etmek için 70'tir. Aynı şekilde, hücrenin çekirdeği çevredeki medium44'ten daha düşük bir kırılma indeksine sahiptir, bu da daha düşük bindirme gücüne sahip olsa bile boncuksuz girintiye (yayınlanmamış gözlemler ve referans72) izin verir. Böylece çekirdek kolayca tuzağa düşürülemez ve tuzaktan kaçar.
Spin kaplı PDMS aralayıcı, kullanışlı ve hızlı bir yöntemle üretilmiştir, ancak mikro/nanofabrikasyon tesisine veya mühendislik laboratuvarlarına erişimi olmayan laboratuvarlar için ulaşılamaz olabilir. Böylece, aralayıcı laboratuvar bandından veya parafilmden kolayca monte edilebilir (adım 4). Protokol ayrıca, tek hücrelerin önceden tanımlanmış ölçüm kuyularına veya aynı numune içindeki hapsetme etkisini tahmin etmek için tanımlanmış yüksekliğe sahip bir odaya teslimini otomatikleştiren mikroakışkan kanallar üretilerek de uyarlanabilir. Bununla birlikte, bu tür mikroakışkan cihazlar, mikroskop hedefi ile optik kuvvet sensörünün toplama lensi arasındaki boşluğu yaklaşık 2 mm'ye sığacak şekilde tasarlanmalıdır (bkz. adım 3). Optik kuvvet sensörünün uygun yüksekliğe yerleştirilmesi gerektiğini unutmayın, böylece odaklamadan kaynaklanan optik sapmalar foton momentum ölçümünü etkilemez.
Diğer değişiklikler arasında biyolojik muhabirlerin değiştirilmesi de yer alabilir. Hoechst floresan'ın GFP kanalına spektral olarak aktığı tespit ettik ve böylece iki floresan kanalda eşzamanlı ölçüm için nükleer bir işaretleyici olarak mCherry etiketli histon ile kombinasyonu tercih ediyoruz. Alternatif olarak, nükleer deformasyon Lap2b-GFP gibi iç nükleer membranı hedefleyen bir etiketle kolayca takip edilebilir (Şekil 2).
Hücre çekirdeğine girinti, kırınım sınırlı iplik diski konfokal mikroskopisinin görüntü analiziyle doğru bir şekilde ölçebildiğimiz 2-3 mikronluk bir sıradaydı. Daha sert çekirdekler veya daha küçük kuvvetler için, girinti bu yaklaşım kullanılarak zar zor ölçülebilir olacaktır. Bununla birlikte, mutlak kuvvet kalibreli optik cımbızlar, nanometre doğruluğunda BFP interferometri kullanılarak sıkışan boncuk in situ'nun konum ölçümleri için de kalibre edilebilir51. Bu yaklaşım kullanılarak, voltaj sinyali ve optik kuvvet sensörü β [nm/V] parametresi aracılığıyla sıkışmış probun konumuna çevrilebilirken, [pN/V] α sabit parametre yukarıda belirtilen ışık momentumu kalibrasyonu41 aracılığıyla kuvvet değerleri verir (ayrıntılar için aşağıya bakın).
Sorun giderme
Deney sırasında aşağıdaki zorlukların ortaya çıkabileceğini gördük:
Kararlı bir tuzak oluşmaz ve mikroküre kolayca kaçar
Mikroskop hedefteki herhangi bir kir veya yanlış hizalanmış bir düzeltme tasması, kararlı bir tuzağın arızalanmasına neden olabilir. Hemen bir çözüm bulunamazsa, objektif lensin nokta yayma işlevini ölçün. İlgi örneği optik olarak yoğun bir dokunun derinliklerindeyse, lazer odağı kararsız bindirmeye yol açan ciddi optik sapmalarla karşılaşabilir (bu etki genellikle izole hücrelerde ihmal edilebilir, ancak daha kalın dokularda daha belirgin hale gelir). Yüksek sertlik için, çekirdeğin geri yükleme kuvveti tuzağın kaçış kuvvetini aşabilir, böylece mikroküreği kaybolur ve girinti rutini başarısız olur. Başlangıçta, optik kapanın proksimal nükleer membran kenarı neredeyse girintilenir (Şekil S2A). Bu durumda, bindirme lazeri artık kuvvet ve Brownian hareketinden etkilenmez, bu da kuvvetin sıfıra düşmesine ve sinyal gürültüsünün azalmasına neden olur (Şekil S2B). Bunun olması durumunda, lazer gücü daha güçlü bir tuzağa sahip olacak şekilde artırılabilir, boncukları çekirdeğe iten yamuk yörüngesinin genliği azaltılabilir veya sıkışan mikrobun başlangıç konumu çekirdeğin daha uzağına ayarlanabilir.
Hücre uyarılma sırasında hareket ediyor
Hücreler yeterince bağlı değilse, optik degrade tuzağı hücre içi girinti rutinini gerçekleştirirken hücreleri hareket ettirir, böylece çekirdeğin kuvvetleri ve altta yatan mekaniği artetseldir. Tüm hücrenin yer değiştirmesini önlemek için, örneğin ConA gibi yüzeydeki hücre yapışma moleküllerinin konsantrasyonunu artırmanızı öneririz.
İlk momentum telafisi
OTs platformunda (adım 6.5) ilk momentum dengeleme yordamı mevcut değilse, yapay, kuvvet bağımsız bir taban çizgisi sinyalinin düzeltilmesi gerekir. Bu, boncuk sıkışmamış olsa bile kuvvet eğrisinde bir eğim olarak görülebilir (Şekil S1E). Düzeltmeyi yapmak için, aynı yörüngenin bir boncuk olmadan, hücrenin dışında tam olarak aynı konumda yapılması gerekir. Bunun için, sahne alanı denetimini kullanarak hücreyi tuzaktan uzaklaştırın. Referans olarak, kuvvet uzaklığı sistemimizde 200 mW'ta FOV genelinde 5 pN değişir; böylece, kısa yörüngeler için ihmal edilebilir hale gelir. Alternatif olarak, örnekteki hücreleri hareket ettirmek için lazer konumunu sabit bırakarak bir piezo tarama aşaması kullanılabilir.
Sunulan protokolün kritik adımları
Embriyo üzerinde maksimum dağılımı sağlamak için mikroküreçler sağ, 1 hücreli aşamada enjekte edilmelidir. Boncuklar floresan olmamalıdır, böylece görüntüleme için kullanılan floresan kanallara ışık sızmaz. Örneğin, tipik kırmızı floresan boncuklar bile parlaklıkları nedeniyle Hoechst lekelenmesinden sonra hücre çekirdeğini görüntülemek için kullanılan mavi kanalda açıkça görülebilir (heyecan: 405 nm; emisyon: 445 nm). Hücrenin alt tabakaya kararlı bir şekilde bağlanması, girinti rutini sırasında yanal yer değiştirmeyi önlemek için kritik öneme sahiptir. Hücre rutin sırasında hareket ederse, kuvvetler hafife edilir. Bu sık sık gerçekleşirse, ek protokolünü en iyi duruma getirin. Doku kültürü hücreleri için fibronektin, kollajen veya poli-L lizin gibi diğer hücre yapışıklık proteinleri tatmin edici bağlanmaya (yayınlanmamış gözlemler) yol açar. Hapsetme sırasında hücreler ani ve şiddetli mekanik strese maruz kalır. Bu, hücrelere zarar verebilir ve prosedür dikkatli bir şekilde gerçekleştirilmezse deneyci sık sık patlayan hücrelerle karşılaşır. Ayrıca, hapsetme yüksekliği çok küçükse, tüm hücreler nükleer sarmal kırılma veya geri dönüşü olmayan hasardan muzdarip olacaktır. Bunları azaltmak için üst kapak kapağını daha yavaş küseyin ve/veya kapak arasındaki aralığı artırın.
Tekniğin sınırlamaları ve bunların üstesinden gelmek için öneriler
Tekniğin açık bir sınırlaması, lazer ışığının dokunun derin bölümlerine nüfuz etmesidir, bu da sapmalara ve dengesiz bindirmeye yol açar. Bu nedenle, penetrasyon derinliğinin bir alt sınırı, numunenin netliğine,73 kullanılabilen sapma düzeltmeye ve uygulanan lazer gücüne bağlıdır. Daha yüksek bir lazer gücünün mikrokürenin çevresindeki numunenin termal olarak çıkarılmasına yol açtığı dikkate alınmalıdır. Bununla birlikte, 1064 nm dalga boyu lazer noktasının neden olduğu numunenin ısıtılması, biyolojik numunelerimize ısı ile ilgili makul stresi önlemek için en aza indirilmektedir74.
Diğer bir sınırlama, ölçülebilecek maksimum kuvvettir. Doğrudan ışık momentumu algılaması, optik trap40,41'in doğrusal tepki rejiminin çok ötesinde kuvvet ölçümleri sağlasa da, maksimum uygulanan kuvvet birkaç yüz picoNewtons sırasına göredir. Bu, lazer gücü ve buna bağlı olarak yumuşak biyolojik malzemenin hasar eşiği ve normalde 0,1 veya 0,344'ten büyük olmayan kırılma indeksi farklılıkları ile sınırlıdır. Güç algılama sınırını artırmak için çeşitli yöntemler önerilmiştir, örneğin yapılandırılmış light75, yansıma önleyici kaplamalı mikroküreler76, yüksek kırılma indeksi parçacıkları77 veya yüksek doped kuantum noktaları78 kullanılarak.
BFP interferometrisi aracılığıyla nanometre ölçeğinde konum ölçümleri için kullanılabilir, böylece boncukun tuzak içindeki konumu Δx = β Sx'tir, burada Sx sensörün voltaj sinyalidir ve β [μm/V] farklı protokoller35,54'ü izleyerek anında kalibre edilebilir. Optik kuvvet sensörü için, [pN/V] α gerilimden kuvvete sabit dönüştürme faktörünün doğrudan β ve tuzak sertliği ile ilişkili olduğu kanıtlanabilir, k [pN/μm], α = kβ 37) Optik görüntülemeden tespit edilemeyecek kadar küçük boncuk yer değiştirmeleri ile yapılan deneylerde, bu strateji kuvvet ölçümlerini küçük konum tespiti ile tamamlamak için kullanılabilir. Burada sunulan deneysel rutinlerin, makul lazer güçlerindeki (200-500 mW) kuvvetlerin yeterince büyük girinti değerlerini teşvik etmek için yeterli olmadığı çok sert çekirdeklere uygulanması bir örnektir. Bu durumda, boncukun çekirdekle temas etmesi ve bindirme sertliğinin ölçümden önce kalibre edilmesi gerekir (adım 8.6). Çekirdeğin kuvvet fonksiyonu olarak girintisi dolaylı olarak şu şekilde belirlenebilir:
d = xtrap - F/k
burada xtrap tuzak konumudur. [pN/V] α değişmez ışık momentum faktöründen farklı olarak, parçacık boyutu, optik bindirme noktası boyutu ve göreli kırılma indeksleri gibi bindirme dinamiklerini belirleyen birçok yerel değişkene bağlı olduğundan, [pN/V] faktör β [μm/V] her deneyden önce kalibre edilmesi gerekir.
Açıklamalar
Yazarların açıklayacak bir şeyi yok.
Teşekkürler
MK, İspanya Ekonomi ve Rekabetçilik Bakanlığı'nın Nacional (PGC2018-097882-A-I00), FEDER (EQC2018-005048-P), Severo Ochoa Ar-Ge Mükemmeliyet Merkezleri programı (CEX2019-000910-S; RYC-2016-21062), Fundació Privada Cellex, Fundació Mir-Puig ve Generalitat de Catalunya'dan CERCA ve Research programı (2017 SGR 1012) aracılığıyla, ERC (MechanoSystems) ve HFSP (CDA00023/2018) aracılığıyla finansmana ek olarak. V.R., İspanya Bilim ve Yenilik Bakanlığı'nın EMBL ortaklığı Centro de Excelencia Severo Ochoa, MINECO'nun Plan Nacional (BFU2017-86296-P, PID2020-117011GB-I00) ve Generalitat de Catalunya'ya (CERCA) desteğini kabul ediyor. V.V., 665884 Marie Skłodowska-Curie hibe anlaşması kapsamında Avrupa Birliği'nin Horizon 2020 araştırma ve yenilik programı tarafından finanse edilen ICFOstepstone Doktora Programı'ndan destek kabul ediyor. Arnau Farré'ye makalenin eleştirel okuması için teşekkür ederiz; Maria Marsal, 24 hpf embriyonun görüntülenmesi ve montajı için yardım için; Senda Jiménez-Delgado zebra balığı mikronjeksiyonları ile destek için.
Malzemeler
| Name | Company | Catalog Number | Comments |
| #1.5 22 mm cover glasses | Ted Pella | 260148 | |
| #1.5 22x60 mm Coverglasses | Ted Pella | 260152 | |
| #1.5H glass bottom dishes | Willco | GWST-5040 | |
| 10-um beads | Supelco | 72986 | |
| 1-mm glass capillaries | Harvard Apparatus | 30-0020 GC100F-15 | |
| 1-um polystyrene microbeads | Sigma | 89904 | |
| 1-um red-fluorescent beads | ThermoFisher | F8816 | |
| Agar | ThermoFisher | 16500500 | |
| Aqcuisition cameras sCMOS | Andor | Sona-4BV11-UNI | |
| Auxiliary camera (Figure 3, AUX) | Blackfly, FLIR | BFS-U3-200S6M-C | |
| Calbryte 520 | AAT Bioquest | 520 AM | |
| Centrifuge | Eppendorf | 5453000011 | |
| Concanavalin A | Sigma | C5275 | |
| DMEM | Sigma | D2906 | |
| DNA-Hoechst | ThermoFisher | 33342 | |
| Double scotch tape | Biesse Adesivi | ||
| E3 | 5 mM NaCl. 0.17 mM KCl. 0.33 mM CaCl2. 0.33 mM MgSO4 | ||
| Eclipse Ti2 | Nikon | ||
| Forceps | Fine Science Tools | 11252-20 | |
| GPI-GFP | |||
| H2A-mCh | |||
| Image acquisition software | Fusion-Andor | ||
| Immersion Oil | Cargille | Type B: 16484 | |
| IR protection googles | Thorlabs | LG1 | |
| Lap2b-eGFP | |||
| Micro loader pipette | Eppendorf | GELoader | |
| Microinjector | World Precision Instruments | SYS-PV820 | |
| MicroManager 2.0 | |||
| Micromiter slide | ID5243 GXMGRAT-5 5mm/100 divisions | ||
| Mineral oil | Sigma | M3616 | |
| Motorized stage | ASI | ||
| Needle puller | Sutter instrument Co. | Model P-97 | |
| Optical tweezers platform | Impetux Optics | Sensocell | |
| OTs software (LightAce) | Impetux Optics | ||
| PDMS | Sigma | Sylgard 184 | |
| PDMS Curing agent | Sigma | Sylgard 184 | |
| Post processing software (Matlab) | Mathworks | ||
| RNAse free water | Thermofisher | AM9937 | |
| Short-pass dichroic mirror (Figure 3, IR-F) | Semrock | FF01-950/SP-25 | |
| Spin-coater | Specialty Coating Systems | Spincoat G3P-8 | |
| Spinning-disk confocal microscope | Andor | DragonFly 502 | |
| Stereomicroscope | Leica M80 | ||
| Triangular microinjection mold | Adaptive Science Tools | TU1 | |
| Universal oven | Memmert | UNB 200 | |
| Water immersion objective | Nikon | MRD07602 |
Referanslar
- Chan, C. J., Heisenberg, C. P., Hiiragi, T. Coordination of Morphogenesis and Cell-Fate Specification in Development. Current Biology. 27 (18), 1024-1035 (2017).
- Heller, E., Fuchs, E. Tissue patterning and cellular mechanics. Journal of Cell Biology. 211 (2), 219-231 (2015).
- Heisenberg, C. P., Bellaïche, Y. Forces in tissue morphogenesis and patterning. Cell. 153 (5), 948-962 (2013).
- Petridou, N. I., Spiró, Z., Heisenberg, C. P. Multiscale force sensing in development. Nature Cell Biology. 19 (6), 581-588 (2017).
- Krieg, M., et al. Tensile forces govern germ-layer organization in zebrafish. Nature Cell Biology. 10 (4), 429-436 (2008).
- Ruprecht, V., et al. Cortical contractility triggers a stochastic switch to fast amoeboid cell motility. Cell. 160 (4), 673-685 (2015).
- Shellard, A., Mayor, R. Supracellular migration - Beyond collective cell migration. Journal of Cell Science. 132 (8), (2019).
- Mongera, A., et al. A fluid-to-solid jamming transition underlies vertebrate body axis elongation. Nature. 561 (7723), 401-405 (2018).
- Atia, L., et al. Geometric constraints during epithelial jamming. Nature Physics. 14 (6), 613-620 (2018).
- Turlier, H., Maître, J. -. L. Mechanics of tissue compaction. Seminars in Cell & Developmental Biology. 47-48, 110-117 (2015).
- Ladoux, B., Mège, R. M. Mechanobiology of collective cell behaviours. Nature Reviews Molecular Cell Biology. 18 (12), 743-757 (2017).
- Venturini, V., et al. The nucleus measures shape changes for cellular proprioception to control dynamic cell behavior. Science. 370 (6514), (2020).
- Charras, G., Sahai, E. Physical influences of the extracellular environment on cell migration. Nature Reviews Molecular Cell Biology. 15 (12), 813-824 (2014).
- Kirby, T. J., Lammerding, J. Emerging views of the nucleus as a cellular mechanosensor. Nature Cell Biology. 20 (4), 373-381 (2018).
- Lee, H. P., et al. The nuclear piston activates mechanosensitive ion channels to generate cell migration paths in confining microenvironments. Science Advances. 7 (2), (2021).
- Friedl, P., Wolf, K., Lammerding, J. Nuclear mechanics during cell migration. Current Opinion in Cell Biology. 23 (1), 55-64 (2011).
- Versaevel, M., Riaz, M., Grevesse, T., Gabriele, S. Cell confinement: Putting the squeeze on the nucleus. Soft Matter. 9 (29), 6665-6676 (2013).
- Zuela-Sopilniak, N., et al. Measuring nucleus mechanics within a living multicellular organism: Physical decoupling and attenuated recovery rate are physiological protective mechanisms of the cell nucleus under high mechanical load. Molecular Biology of the Cell. 31 (17), 1943-1950 (2020).
- Kim, D. H., Wirtz, D. Cytoskeletal tension induces the polarized architecture of the nucleus. Biomaterials. 48, 161-172 (2015).
- Lomakin, A. J., et al. The nucleus acts as a ruler tailoring cell responses to spatial constraints. Science. 370 (6514), (2020).
- Hampoelz, B., et al. Microtubule-induced nuclear envelope fluctuations control chromatin dynamics in Drosophila embryos. Development. 138 (16), 3377-3386 (2011).
- Heo, S. J., et al. Differentiation alters stem cell nuclear architecture, mechanics, and mechano-sensitivity. eLife. 5, 1-21 (2016).
- Cosgrove, B. D., et al. Nuclear envelope wrinkling predicts mesenchymal progenitor cell mechano-response in 2D and 3D microenvironments. Biomaterials. 270, 120662 (2021).
- Liu, H., et al. In situ mechanical characterization of the cell nucleus by atomic force microscopy. ACS Nano. 8 (4), 3821-3828 (2014).
- Hobson, C. M., et al. Correlating nuclear morphology and external force with combined atomic force microscopy and light sheet imaging separates roles of chromatin and lamin A/C in nuclear mechanics. Molecular Biology of the Cell. 31 (16), 1788-1801 (2020).
- Pajerowski, J. D., Dahl, K. N., Zhong, F. L., Sammak, P. J., Discher, D. E. Physical plasticity of the nucleus in stem cell differentiation. Proceedings of the National Academy of Sciences of the United States of America. 104 (40), 15619-15624 (2007).
- Rowat, A. C., Lammerding, J., Ipsen, J. H. Mechanical properties of the cell nucleus and the effect of emerin deficiency. Biophysical Journal. 91 (12), 4649-4664 (2006).
- Davidson, P. M., et al. High-throughput microfluidic micropipette aspiration device to probe time-scale dependent nuclear mechanics in intact cells. Lab on a Chip. 19 (21), 3652-3663 (2019).
- Lombardi, M., Zwerger, M., Lammerding, J. Biophysical assays to probe the mechanical properties of the interphase cell nucleus: Substrate strain application and microneedle manipulation. Journal of Visualized Experiments: JoVE. (55), (2011).
- Luo, T., Mohan, K., Iglesias, P. A., Robinson, D. N. Molecular mechanisms of cellular mechanosensing. Nature Materials. 12 (11), 1064-1071 (2013).
- Dahl, K. N., Engler, A. J., Pajerowski, J. D., Discher, D. E. Power-law rheology of isolated nuclei with deformation mapping of nuclear substructures. Biophysical Journal. 89 (4), 2855-2864 (2005).
- Guilluy, C., et al. Isolated nuclei adapt to force and reveal a mechanotransduction pathway in the nucleus. Nature Cell Biology. 16 (4), 376-381 (2014).
- Bustamante, C. J., Wang, M. D. Optical tweezers in single-molecule biophysics. Nature Reviews Methods Primers. , 1-29 (2021).
- Svoboda, K., Block, S. M. Force and velocity measured for single kinesin molecules. Cell. 77 (5), 773-784 (1994).
- Berg-Sørensen, K., Flyvbjerg, H. Power spectrum analysis for optical tweezers. Review of Scientific Instruments. 75 (3), 594-612 (2004).
- Smith, S. B., Cui, Y., Bustamante, C. Optical-trap force transducer that operates by direct measurement of light momentum. Methods in Enzymology. 361 (1994), 134-162 (2003).
- Farré, A., Montes-Usategui, M. A force detection technique for single-beam optical traps based on direct measurement of light momentum changes. Optics Express. 18 (11), 11955 (2010).
- Català, F., Marsà, F., Montes-Usategui, M., Farré, A., Martín-Badosa, E. Extending calibration-free force measurements to optically-trapped rod-shaped samples. Scientific Reports. 7, 1-10 (2017).
- Bui, A. A. M., et al. Calibration of force detection for arbitrarily shaped particles in optical tweezers. Scientific Reports. 8 (1), 1-12 (2018).
- Farré, A., Marsà, F., Montes-Usategui, M. Beyond the hookean spring model: Direct measurement of optical forces through light momentum changes. Methods in Molecular Biology. 1486, (2017).
- Farré, A., Marsà, F., Montes-Usategui, M. Optimized back-focal-plane interferometry directly measures forces of optically trapped particles. Optics Express. 20 (11), 12270 (2012).
- Jun, Y., Tripathy, S. K., Narayanareddy, B. R. J., Mattson-Hoss, M. K., Gross, S. P. Calibration of optical tweezers for in vivo force measurements: How do different approaches compare. Biophysical Journal. 107 (6), 1474-1484 (2014).
- Mas, J., Farré, A., Sancho-Parramon, J., Martín-Badosa, E., Montes-Usategui, M. Force measurements with optical tweezers inside living cells. Optical Trapping and Optical Micromanipulation XI. 9164, (2014).
- Schürmann, M., Scholze, J., Müller, P., Guck, J., Chan, C. J. Cell nuclei have lower refractive index and mass density than cytoplasm. Journal of Biophotonics. 9 (10), 1068-1076 (2016).
- Rosen, J. N., Sweeney, M. F., Mably, J. D. Microinjection of zebrafish embryos to analyze gene function. Journal of Visualized Experiments: JoVE. (25), (2009).
- Westerfield, M. . The Zebrafish Book. A Guide for the Laboratory Use of Zebrafish. (Danio rerio), 5th Edition. , (2007).
- Schubert, R., et al. Assay for characterizing the recovery of vertebrate cells for adhesion measurements by single-cell force spectroscopy. FEBS Letters. 588 (19), 3639-3648 (2014).
- Koschwanez, J. H., Carlson, R. H., Meldrum, D. R. Thin PDMS films using long spin times or tert-butyl alcohol as a solvent. PLoS One. 4 (2), 2-6 (2009).
- Das, R., et al. Mechanical stretch inhibition sensitizes proprioceptors to compressive stresses. bioRxiv. , (2021).
- Chardès, C., Clement, R., Blanc, O., Lenne, P. F. Probing cell mechanics with bead-free optical tweezers in the drosophila embryo. Journal of Visualized Experiments: JoVE. (141), (2018).
- Staunton, J. R., Blehm, B., Devine, A., Tanner, K. In situ calibration of position detection in an optical trap for active microrheology in viscous materials. Optics Express. 25 (3), 1746 (2017).
- Bola, R., Treptow, D., Marzoa, A., Montes-Usategui, M., Martin-Badosa, E. Acousto-holographic optical tweezers. Optics Letters. 45 (10), 2938-2941 (2020).
- Thalhammer, G., Obmascher, L., Ritsch-Marte, M. Direct measurement of axial optical forces. Optics Express. 23 (5), 6112 (2015).
- Vermeulen, K. C., et al. Calibrating bead displacements in optical tweezers using acousto-optic deflectors. Review of Scientific Instruments. 77 (1), 1-6 (2006).
- Dzementsei, A., Barooji, Y. F., Ober, E. A., Oddershede, L. B. Foregut organ progenitors and their niche display distinct viscoelastic properties in vivo during early morphogenesis stages. bioRxiv. , 1-35 (2021).
- Behrndt, M., et al. Forces driving epithelial spreading in zebrafish gastrulation. Science. 338 (6104), 257-260 (2012).
- Krieg, M., et al. Atomic force microscopy-based mechanobiology. Nature Reviews Physics. , (2018).
- A-Hassan, E., et al. Relative microelastic mapping of living cells by atomic force microscopy. Biophysical Journal. 74 (3), 1564-1578 (1998).
- Khalilgharibi, N., et al. Stress relaxation in epithelial monolayers is controlled by the actomyosin cortex. Nature Physics. 15, (2019).
- Crick, S. L., Yin, F. C. Assessing micromechanical properties of cells with atomic force microscopy: importance of the contact point. Biomechanics and Modeling in Mechanobiology. 6 (3), 199-210 (2007).
- Hurst, S., Vos, B. E., Betz, T. Intracellular softening and fluidification reveals a mechanical switch of cytoskeletal material contributions during division. bioRxiv. , 425761 (2021).
- Kaplan, J. L., Bonfanti, A., Kabla, A. RHEOS.jl - A Julia package for rheology data analysis. arXiv. 4, 1-5 (2020).
- Bonfanti, A., Kaplan, J. L., Charras, G., Kabla, A. Fractional viscoelastic models for power-law materials. Soft Matter. 16 (26), 6002-6020 (2020).
- Rivas-Barbosa, R., Escobedo-Sánchez, M. A., Tassieri, M., Laurati, M. i-Rheo: determining the linear viscoelastic moduli of colloidal dispersions from step-stress measurements. Physical Chemistry Chemical Physics: PCCP. 22 (7), 3839-3848 (2020).
- Tassieri, M., et al. i-Rheo: Measuring the materials' linear viscoelastic properties "in a step". Journal of Rheology. 60 (4), 649-660 (2016).
- Zhong, M. C., Wei, X. B., Zhou, J. H., Wang, Z. Q., Li, Y. M. Trapping red blood cells in living animals using optical tweezers. Nature Communications. 4, 1767-1768 (2013).
- Harlepp, S., Thalmann, F., Follain, G., Goetz, J. G. Hemodynamic forces can be accurately measured in vivo with optical tweezers. Molecular Biology of the Cell. 28 (23), 3252-3260 (2017).
- Bayoudh, S., Mehta, M., Rubinsztein-Dunlop, H., Heckenberg, N. R., Critchley, C. Micromanipulation of chloroplasts using optical tweezers. Journal of Microscopy. 203 (2), 214-222 (2001).
- Favre-Bulle, I. A., Stilgoe, A. B., Rubinsztein-Dunlop, H., Scott, E. K. Optical trapping of otoliths drives vestibular behaviours in larval zebrafish. Nature Communications. 8 (1), 630 (2017).
- Bambardekar, K., Clément, R., Blanc, O., Chardès, C., Lenne, P. F. Direct laser manipulation reveals the mechanics of cell contacts in vivo. Proceedings of the National Academy of Sciences of the United States of America. 112 (5), 1416-1421 (2015).
- Ferro, V., Chuai, M., McGloin, D., Weijer, C. J. Measurement of junctional tension in epithelial cells at the onset of primitive streak formation in the chick embryo via non-destructive optical manipulation. Development (Cambridge). 147 (3), (2020).
- Hörner, F., et al. Holographic optical tweezers-based in vivo manipulations in zebrafish embryos. Journal of Biophotonics. 10 (11), 1492-1501 (2017).
- Zhong, M. -. C., Wang, Z. -. Q., Li, Y. -. M. Aberration compensation for optical trapping of cells within living mice. Applied Optics. 56 (7), 1972 (2017).
- Català, F., Marsà, F., Montes-Usategui, M., Farré, A., Martín-Badosa, E. Influence of experimental parameters on the laser heating of an optical trap. Scientific Reports. 7 (1), 1-9 (2017).
- Taylor, M. A., Waleed, M., Stilgoe, A. B., Rubinsztein-Dunlop, H., Bowen, W. P. Enhanced optical trapping via structured scattering. Nature Photonics. 9 (10), 669-673 (2015).
- Bormuth, V., et al. Optical trapping of coated microspheres. Optics Express. 16 (18), 13831-13844 (2008).
- Sudhakar, S., et al. Germanium nanospheres for ultraresolution picotensiometry of kinesin motors. Science. 371 (6530), (2021).
- Shan, X., et al. Optical tweezers beyond refractive index mismatch using highly doped upconversion nanoparticles. Nature Nanotechnology. 16 (5), 531-537 (2021).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiDaha Fazla Makale Keşfet
This article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır