
Method Article
Измерения прямой силы субклеточной механики в конфайнменте с помощью оптического пинцета
В этой статье
Резюме
Здесь мы представляем протокол исследования внутриклеточных механических свойств изолированных эмбриональных клеток рыбок данио в трехмерном заключении с прямым измерением силы оптической ловушкой.
Аннотация
Во время развития многоклеточного организма одна оплодотворенная клетка делится и дает начало множественным тканям с разнообразными функциями. Тканевый морфогенез идет рука об руку с молекулярными и структурными изменениями на уровне одной клетки, которые приводят к изменениям субклеточных механических свойств. Как следствие, даже внутри одной клетки разные органеллы и компартменты по-разному противостоят механическим воздействиям; и пути механотрансдукции могут активно регулировать свои механические свойства. Таким образом, способность клетки адаптироваться к микроокружению тканевой ниши частично обусловлена способностью ощущать и реагировать на механические напряжения. Недавно мы предложили новую парадигму механосенсации, в которой ядерная деформация и позиционирование позволяют клетке измерять физическую 3D-среду и наделяют клетку чувством проприоцепции для декодирования изменений в форме клеток. В этой статье мы опишем новый метод измерения сил и свойств материала, которые формируют ядро клетки внутри живых клеток, на примере адгезивных клеток и механически ограниченных клеток. Измерения могут быть выполнены неинвазивно с оптическими ловушками внутри клеток, а силы напрямую доступны через безкалибровочное обнаружение светового импульса. Это позволяет измерять механику ядра независимо от деформаций клеточной поверхности и позволяет рассеивать пути экстероцептивной и интероцептивной механотрансдукции. Важно отметить, что эксперимент по улавливанию может быть объединен с оптической микроскопией для исследования клеточного ответа и субклеточной динамики с использованием флуоресцентной визуализации цитоскелета, ионов кальция или ядерной морфологии. Представленный метод прост в применении, совместим с коммерческими решениями для измерения силы и может быть легко расширен для исследования механики других субклеточных компартментов, например, митохондрий, стресс-волокон и эндосом.
Введение
Тканевый морфогенез представляет собой сложный процесс, при котором биохимические сигналы и физические силы пространственно-временно координируются. У развивающегося эмбриона градиенты биохимических сигнальных факторов диктуют специфику судьбы и обеспечивают правильную структуру тканей1,2. В то же время внутренние и внешние силы играют определенную роль в построении архитектуры эмбриона3,4. Влияние механики клеточной коры в этом контексте было широко изучено5,6. Тесная взаимосвязь между механохимическими процессами во время морфогенеза зависит от свойств отдельных клеток ощущать и реагировать на механические силы в микроокружении их тканей. Клетки, таким образом, декодируют механические сигналы через присутствие чувствительных к силе субклеточных и молекулярных элементов, которые преобразуют механическую информацию в конкретные сигнальные пути, контролирующие поведение клеток, судьбу клеток и клеточную механику.
Отличительной чертой процессов развития является то, что клетки организуются в группы для построения многоклеточных структур. Таким образом, одиночные клетки редко перестраиваются и движутся в одиночку, но связаны в плотном социотопе, в котором они демонстрируют коллективное поведение, такое как надклеточная миграция7, (не)глушащие переходы8,9 или уплотнение бластоцисты10. Механические силы, генерируемые внутри и между клетками, служат важными сигналами для управления коллективной динамикой клеток7,11. Но даже когда клетки движутся в одиночку, такие как клетки-предшественники, которые протискиваются между тканевыми листами или узкими тканевыми нишами, они испытывают обширные анизотропные механические силы при навигации по трехмерной среде. Эти механические нагрузки на клетки оказывают глубокое воздействие на поведение клеток12,13. Было исследовано несколько механизмов, которые сходятся на ядре как основной элемент механотрансдукции14,15, как пассивный или активный механический элемент во время миграции в плотной среде 3D-ткани15,16.
Недавно мы предложили механизм, который оснащает клетки для измерения деформаций формы с использованием ядра в качестве эластичного внутриклеточного механо-калибра12. Ядро, будучи самой большой органеллой в клетке, подвергается большим деформациям, когда клетки поляризуются, мигрируют или изменяют свою форму при механическом растяжении, ограничении или осмотическом напряжении16,17,18,19. Мы обнаружили, что растяжение ядерной оболочки вместе с внутриклеточным позиционированием ядра предоставляет клеткам информацию о величине и типе клеточной деформации (например, сжатие клеток против клеточного набухания). Растяжение ядра связано с развертыванием внутренней ядерной мембраны (INM), что способствует кальций-зависимой активности cPLA2 (цитозольная фосфолипаза A2) липазы в INM с последующим высвобождением арахидоновой кислоты (AA) и быстрой активацией миозина II в клеточной коре. Это приводит к повышенной сократимости клеток и миграции амебоидных клеток выше порога сократимости коры6. Механочувствительная реакция на деформацию клеток происходит менее чем за минуту и обратима при высвобождении удержания, предполагая, что ядро действует как тензорезистор клеточной проприоцепции, регулирующий адаптивное поведение клеток в условиях механического стресса. Показано, что этот механочувствительный путь активен в стволовых клетках-предшественниках, полученных из эмбрионов рыбок данио, как в плюрипотентных, так и в линейных клетках12 и сохраняется у различных видов и клеточных линий20.
В дополнение к ядерным свойствам в качестве клеточного механосенсора, ядерная архитектура и механика внутренне регулируются во время разработки и в ответ на спецификацию клеточной судьбы21, следовательно, настраивая клеточную механочувствительность22,23. Следствием этого может стать изменение в ядерном соответствии, которое допускает морфологические изменения и переходы из преимущественного в миграционное состояние и наоборот8.
Было применено несколько методов измерения механики клеточного ядра, таких как атомно-силовая микроскопия24,25, аспирация микропипетки26,27, микрофлюидная технология28 и микроиглы29. Однако многие из этих методов являются инвазивными в том смысле, что вся клетка должна быть деформирована, ограничивая измерение механических характеристик и силозависимых реакций самого ядра. Для обхода одновременной деформации поверхности клетки и ее механочувствительной клеточной коры30 изучали изолированные ядра в различных контекстах31,32. Однако нельзя исключать, что ядерная изоляция связана с изменением свойств механических ядер и их регуляции (ссылка24 и собственные неопубликованные наблюдения).
Оптический пинцет (OT) является универсальной технологией, которая позволила провести множество экспериментов в клеточной механобиологии и сыграла важную роль в нашем понимании того, как молекулярные машины преобразуют химическую энергию в механическую33,34. Оптический пинцет использует плотно сфокусированный лазерный луч для воздействия оптических сил на диэлектрические частицы, которые имеют показатель преломления выше, чем окружающая среда33. Такие силы могут составлять порядка сотен пиконьютонов и приводить к эффективному удержанию частицы в фокусе лазерной ловушки, что позволяет манипулировать захваченной частицей в трех измерениях. Использование света имеет важное преимущество в том, что измерение может быть выполнено неинвазивно внутри живых клеток. Оптические манипуляции дополнительно ограничиваются фокусом ловушки лазерного луча. Следовательно, манипуляция может быть выполнена без стимуляции окружающих клеточных мембран и не нарушает актиновую кору или механочувствительные процессы на плазматической мембране, такие как силозависимая активация ионных каналов.
Сложность подхода оптического пинцета заключается в точном определении сил, приложенных к микросфере, с использованием классических подходов, которые полагаются на калибровку косвенной силы на основе теоремы о равнораспределении или использование определенных сил сопротивления Стокса для измерения лазерной силы убегания35. Хотя эти методы легко реализовать в эксперименте in vitro, они обычно не могут быть переведены в клеточную среду. В этой области было введено несколько стратегий, основанных на калибровке прямой силы, полученной из первых принципов сохранения импульса36,37. В отличие от других подходов силовой спектроскопии, измерения силы выводятся из локального обмена световым импульсом с захваченной частицей произвольной формы38,39. В нашей экспериментальной установке изменения светового импульса, возникающие в результате оптических сил, непосредственно измеряются без необходимости калибровки ловушки in situ40,41,42,43. Таким образом, измерения становятся возможными в вязкой среде, такой как внутренняя часть клетки или даже внутри ткани, и силы могут быть легко количественно оценены вплоть до уровня pN.
В этом протоколе мы описываем анализ для механического манипулирования внутриклеточными органеллами или структурами и количественной оценки их механических свойств с помощью оптической установки пинцета. Эта установка интегрирована во вращающийся дисковый флуоресцентный микроскоп, позволяющий параллельно визуализировать поведение клеток или внутриклеточную динамику. Анализ позволяет охарактеризовать механические свойства конкретных клеточных компартментов, таких как ядро, одновременно изучая возможный механореспонгс и активацию молекулярных сигнальных путей в результате самой деформации. Кроме того, оптическое улавливание введенных микрошариков внутри клеток позволяет увеличить силу углубления благодаря значительно более высокому показателю преломления полистирольной бусины (n = 1,59) по сравнению с внутренним преломляющим контрастом44 ядра (n ~ 1,35) по сравнению с цитоплазмой (n ~ 1,38). Представленная стратегия может быть легко адаптирована к изучению других внутриклеточных структур и органелл, а также других подходов, включающих активную микрореологию, использование нескольких оптических ловушек для одновременного зондирования одних и тех же/разных субклеточных структур и измерения, нацеленные на клеточную механобиологию у живого эмбриона.
протокол
Все используемые протоколы были одобрены Институциональным комитетом по уходу за животными и их использованию (PRBB-IACUEC) и реализованы в соответствии с национальными и европейскими правилами. Все эксперименты проводились в соответствии с принципами 3Р. Рыбки данио (Danio rerio) содержались так, как описано ранее.
1. Получение изолированных первичных эмбриональных стволовых клеток-предшественников рыбок данио
- Препарат микропипетки и агарозы
ПРИМЕЧАНИЕ: Полный протокол микроинъекции эмбриона рыбки данио см. в ссылке45.- С помощью съемника микропипетки потяните стеклянный капилляр толщиной 1,0 мм, чтобы получить две иглы45. Храните неиспользованные иглы в 150-миллиметровой чашке Петри, прикрепленной к подушке для теста, или в вывернутом наизнанку лабораторном ленточном кольце, чтобы защитить тонкий наконечник от повреждений во время транспортировки.
- Расплавить 1% сверхчистой агарозы в E3 (5 мМ NaCl, 0,17 мМ KCl, 0,33 мМ CaCl2, 0,33 мМ MgSO4) в стандартной кухонной/лабораторной микроволновой печи в течение 10 с. Нагревайте смесь многократно в течение коротких периодов времени (несколько секунд), пока агароза не расплавится.
- Когда агароза полностью расплавится, дайте ей ненадолго остыть, а затем вылейте в чашку Петри на 10 см. Медленно добавьте треугольную микроинъекционную форму (см. Таблицу материалов) на верхнюю часть агарозы, избегая появления пузырьков. Не давите на плесень, следя за тем, чтобы она оставалась на агароновой поверхности.
- Когда агароза полностью затвердеет, удалите треугольную плесень очень медленно, приложив мягкую силу, чтобы избежать каких-либо разрывов в агарозе. Пластину можно хранить вверх ногами при 4 °C в течение 2-4 недель.
- За 30 минут до микроинъекции выньте тарелку из холодильника и добавьте E3 предварительно расплавленным до 28 °C, чтобы она стабилизировалась при комнатной температуре.
- Приготовление смеси для инъекций
- Для приготовления инъекционной смеси разводят 1 мкм микрошариков (полистирол, нефлуоресцентный) в соотношении 1:5 в воде, свободной от РНКазы.
- Подготовка мРНК для преходящей экспрессии флуоресцентных маркеров или экспрессии рекомбинантных генных конструкций и/или совместной инъекции морфолиноса в желаемой концентрации.
ПРИМЕЧАНИЕ: Типичная инъекционная смесь для совместной инъекции микрогранул вместе со 100 пг мРНК на эмбрион для маркировки, например, ядра с H2A-mCherry составляет: 1 мкл шариков + 1 мКЛ мРНК (концентрация запаса составляет 1 мкг/мкл) + 2,5 мкл свободной от РНК воды + 0,5 мкл фенольного красного (стоковый раствор 0,5%, фенол красный не является обязательным; он используется для лучшей визуализации введенной капли, но немаркированной инъекции капля также видна опытному экспериментатору). Инъекция РНК также может быть полезна для отбора введенных эмбрионов. Флуоресцентные микрошарики могут быть введены, вместо нефлуоресцентных, чтобы визуализировать их.
- Микроинъекционная загрузка и калибровка иглы
- Включите микроинжектор с помощью параметра Time-Gated . Эта настройка очень важна для правильной калибровки объема впрыска. Установите время затвора приблизительно на 500 мс.
- Загрузите 3 мкл инъекционной смеси в иглу с помощью микрозагрузочной пипетки.
- Вставьте иглу в микроманипулятор и плотно запечатайте. Проверьте, находится ли микроманипулятор в хорошем положении и имеет ли он достаточную свободу для перемещения в направлении x-y на инъекционной пластине.
- Измерьте размер капли с помощью микрометрового слайда (5 мм/100 делений) с каплей минерального масла сверху45 и выбросив каплю смеси для инъекций непосредственно в минеральное масло.
- Обрежьте иглу острыми щипцами под крутым углом, чтобы получить острый заостренный кончик. Отрегулируйте размер капли до 0,1 мм, что соответствует 0,5 нЛ вводимого материала.
ПРИМЕЧАНИЕ: Если при разрезании иглы этот объем превышен, рекомендуется переделать процедуру калибровки новой иглой. Время нагнетания микроинжектора может быть слегка отрегулировано в соответствии с объемом падения; однако короткое время затвора соответствует большому диаметру иглы, что потенциально повреждает эмбрионы.
- Микроинъекция эмбрионов рыбок данио на одноклеточной стадии
- Собирайте эмбрионы рыбок данио вскоре после оплодотворения для микроинъекции бисерной смеси непосредственно в одноклеточный (зиготный) эмбрион до того, как произойдет первое деление клеток.
ПРИМЕЧАНИЕ: Это обеспечивает правильное распределение микросфер и достаточно высокий выход изолированных бластомеров с по меньшей мере одной микросферой на клетку на более поздних стадиях развития, в которых проводятся эксперименты (стадия бластула-гаструла). Эксперименты по углублению все еще могут быть выполнены, если в клетке есть две сферы, но клетки, у которых нет бусин, должны быть исключены (хотя отступ без сфер возможен). В этом протоколе использовались штаммы дикого типа AB, но можно использовать любой другой штамм, например, TL. - Поместите одноклеточные эмбрионы стадии (зиготу) в предварительно сваренную треугольную форму 1% агарозы, как показано на рисунке 1А, используя пластиковую пипетку Пастера.
- Удалите лишнюю среду той же пипеткой, чтобы избежать плавания эмбрионов. Осторожно протолкните эмбрионы в треугольную форму с помощью щетки. Держите некоторое пространство между эмбрионами, чтобы облегчить правильную ориентацию (рисунок 1B).
- Аккуратно выровняйте эмбрионы кистью, чтобы эмбрионы были ориентированы в боковом направлении, при этом одна клетка зиготы была хорошо видна, как показано на рисунке 1B. Идеальная ориентация для микроинъекции достигается, когда одна клетка эмбриона обращена в направлении иглы (инъекция через полюс животного эмбриона) или противоположным образом лицом к желтковой клетке (инъекция через растительный полюс эмбриона), как показано на рисунке 1C.
- Держите блюдо одной рукой, а другой рукой позиционируйте кончик иглы с помощью контроллера микроманипулятора. Опустите кончик иглы к эмбрионам.
- Проткните хорион и введите одноклеточный эмбрион с помощью иглы, наблюдая за процедурой через стереомикроскоп. Обеспечьте правильное размещение иглы и, после инъекции, правильное расположение введенной капли, как показано на рисунке 1C.
- Повторите для всех эмбрионов: переместите иглу вверх, сдвиньте блюдо с эмбрионами, пока следующий эмбрион не будет центрирован, опустите иглу и введите ее.
- После того, как весь набор эмбрионов введен, удалите эмбрионы из агарозной формы / чашки Петри, промыв немного E3 и поместите их в новую чашку Петри с помощью пластиковой пипетки Пастера. Рекомендуется размещать достаточное количество сред на инъекционной пластине, чтобы избежать высыхания эмбрионов во время процедуры микроинъекции.
- Повторяйте процедуру до тех пор, пока не будет введено нужное количество эмбрионов. Эмбрионы должны находиться на одной клеточной стадии, чтобы обеспечить максимальное и однородное распространение шариков.
ПРИМЕЧАНИЕ: Эта процедура оптимизирована для ранних эмбрионов бластулы и, вероятно, должна быть оптимизирована, если необходимо исследовать различные стадии развития. - Поместите введенные эмбрионы внутрь инкубатора при 28-31 °C в течение приблизительно 4 ч или до желаемой стадии (рисунок 1D), прежде чем приступить к протоколу для первичной клеточной культуры.
ПРИМЕЧАНИЕ: Необязательно, пусть эмбрионы развиваются за пределами стадии бластулы (или желаемой точки времени измерения), чтобы обеспечить выживание и исключить артефакты токсичности. На личиночных стадиях устанавливают обезболенные личинки трикаином в 0,75% агарозы и снимают распределение микросфер в различных тканях. Чтобы сделать запасной раствор, смешайте: 400 мг порошка трикаина в 97,9 мл дистиллированной воды, примерно 2,1 мл 1 М TRIS-основания (рН 9), и отрегулируйте до рН 7. Этот раствор можно хранить при температуре 4 °C. Чтобы использовать трикаин в качестве анестетика, разводят 4,2 мл бульонного раствора в 100 мл среды яйца (или желаемой среды); в данном случае использовался E3. Подробности см. в reference46 .
- Собирайте эмбрионы рыбок данио вскоре после оплодотворения для микроинъекции бисерной смеси непосредственно в одноклеточный (зиготный) эмбрион до того, как произойдет первое деление клеток.
2. Одноклеточная подготовка и окрашивание
- Поместите эмбрионы сферической стадии (4 hpf, часы после оплодотворения) в стеклянную посуду с помощью пластиковой пипетки Пастера. Выберите эмбрионы, которые являются положительными для сигнала введенных шариков и которые экспрессируют флуоресцентный белок в случае инъекции мРНК. Некоторые эмбрионы могут демонстрировать высокую кластеризацию шариков и могут быть исключены.
- Вручную дехорионировать эмбрионы с помощью щипцов. Перенесите приблизительно 10-15 эмбрионов в реакционные контейнеры объемом 1,5 мл с помощью стеклянной пипетки Пастера.
ПРИМЕЧАНИЕ: Когда эмбрионы дехорионированы, они прикрепляются к пластику, и требуется использование стеклянной посуды. В качестве альтернативы стеклянной пластине можно использовать пластиковую чашку Петри с тонким слоем 1% агарозы. Ручная дехорионация должна быть предпочтительнее ферментативной обработки проназой, чтобы предотвратить протеолитическое повреждение белков клеточной поверхности и потенциальные изменения механических свойств клеток и тканей, предотвращая длительное время восстановления47.
- Вручную дехорионировать эмбрионы с помощью щипцов. Перенесите приблизительно 10-15 эмбрионов в реакционные контейнеры объемом 1,5 мл с помощью стеклянной пипетки Пастера.
- Удалите среду E3 и добавьте 500 мкл предварительно нагретой CO2-независимой тканевой питательной среды (DMEM-F12; с L-глутамином и 15 мМ HEPES, без бикарбоната натрия и фенол-красного с добавлением 10 единиц пенициллина и 10 мг / л стрептомицина).
ПРИМЕЧАНИЕ: Не используйте CO2-зависимые среды, если не используется микроскопический инкубатор. Использование, например, RPMI в карбонатно-буферизованных условиях вызывает изменения рН среды и может повлиять на выживание клеток. Другим ключевым аспектом является избегание культурных сред, содержащих сыворотку. Сыворотка может содержать лизофосфатидную кислоту (LPA), мощный активатор пути Rho/ROCK, способный контролировать клеточную сократимость и подвижность в стволовых клетках-предшественниках6. Осмолярность среды должна поддерживаться на уровне 300 мОсм, чтобы избежать осмотических проблем, которые могут помешать ядерной морфологии или механике12. - Вручную диссоциируют клетки, осторожно встряхивая трубку. Убедитесь, что содержимое трубки становится мутным без больших кусков, видимых глазом. Избегайте образования пузырьков, чтобы свести к минимуму повреждение и потерю клеток.
- Центрифуга при 200 х г в течение 3 мин. Гранула должна быть хорошо видна.
- Удалите супернатант и выполните одно из шагов, описанных ниже.
- Если окрашивание не требуется, добавьте 500 мкл DMEM. Осторожно повторно суспендируйте с помощью пипетки объемом 200 мкл, нацеливая струю жидкости на гранулу. Не прикладывайте к клеткам чрезмерную силу сдвига. Вспенивание указывает на повреждение клеток.
- Для маркировки ядра ДНК-красителями, такими как Hoechst, смешайте 0,5 мкл ДНК-Hoechst (запас 2 мг/мл) в 1000 мкл DMEM для получения 1 мкг/мл конечной концентрации. Добавьте 500 мкл этого окрашивающего раствора к клеткам и осторожно повторно суспендируйте. Инкубировать в течение 7 мин в темноте.
- Чтобы окрасить клетки флуоресцентным химическим индикатором кальция Calbryte-520, добавьте Calbryte-520 к концентрации 5 мкМ в DMEM. Инкубировать в течение 20 мин в темноте.
ПРИМЕЧАНИЕ: Протоколы, указанные на этапах 2.5.2 и 2.5.3, были оптимизированы для этих конкретных продуктов. Другое окрашивание может быть выполнено с использованием протоколов, указанных производителем.
- Центрифуга снова с использованием тех же настроек, что и на шаге 2.4; удалить надосадочный агент и осторожно повторно суспендировать клетки (чтобы избежать образования кластеров) в 50 мкл DMEM для образцов в суспензии или 20 мкл DMEM для клеток в заключении.
3. Подготовка оптических улавливающих камер с использованием расстояния полидиметилсилоксана (PDMS)
ПРИМЕЧАНИЕ: Измерения оптической силы, основанные на обнаружении светового импульса, требуют захвата всего света, выходящего из оптических ловушек40. Для обеспечения надежности инвариантного калибровочного коэффициента α (pN/V) распределение света в задней фокальной плоскости (BFP) датчика оптической силы должно точно соответствовать импульсу фотона. Это определяет расстояние от поверхности собирающей линзы до плоскости улавливания примерно до 2 мм, что является максимальной высотой оптических камер улавливания.
- PDMS спин-покрытие стеклянной нижней посуды No1.5.
ПРИМЕЧАНИЕ: Следующий рецепт предусмотрен примерно для 40 блюд. Полученная микрокамера будет иметь разную высоту в зависимости от того, будут ли проводиться эксперименты на взвешенных или ограниченных клетках (рисунок 1D).- Смешайте 9 мл базового полимера PDMS и 1 мл отверждающего агента PDMS в конической трубке объемом 50 мл. Активно смешивайте два продукта, чтобы обеспечить правильное распределение отверждающего агента.
- Дегазируйте смесь, чтобы избежать пузырьков, используйте вакуумный насос. Введите коническую трубку в вакуумную бутылку и отсоедините камеру. Подождите, пока в смеси не появятся пузырьки.
ПРИМЕЧАНИЕ: Медленно открывайте вакуум, чтобы предотвратить вспенивание и разливы PDMS из соколиной трубки. - Поместите стеклянную нижнюю посуду на отжимной патрон (рисунок 2А). Будьте осторожны, чтобы не поцарапать, не снять отпечатки пальцев или не испачкать посуду. Защитите отвинчивающуюся коробку от утечек PDMS алюминиевой фольгой.
- Для камер OT для экспериментов на ячейках в суспензии добавьте приблизительно 250 мкл смеси PDMS в центре нижней чашки и вращайте ее при 750 об/мин в течение 1 мин. Высота слоя PDMS составит примерно 50 мкм48.
- Для камер OT для экспериментов на замкнутых клетках добавьте небольшую каплю PDMS (примерно 50 мкл) и вращайте ее при 4000 об/мин в течение 5 мин. Высота слоя PDMS составит приблизительно 10 мкм. Подробный протокол о том, как получить различные толщины PDMS, см. в ссылке 48.
- Отверждайте посуду со стеклянным дном с покрытием PDMS при 70 °C в течение 1 ч.
- Вырежьте скальпелем квадрат размером 1 х 1 см на слой PDMS и очистите его пинцетом (рисунок 2C). В случае замкнутых клеток промывайте мусор PDMS изопропанолом.
- Покрытие камеры для экспериментов с легко прикрепленными клетками в суспензии
- Добавьте 100 мкл конканавалина А (ConA) по 0,5 мг/мл, чтобы покрыть всю поверхность квадратной полости, и дайте ему инкубироваться в течение 30 мин.
ПРИМЕЧАНИЕ: ConA представляет собой лектин, который связывается с сахарами клеточной поверхности и связывает отдельные клетки с поверхностью покровного стекла. - Удалите каплю ConA и тщательно промойте поверхность средой DMEM, не царапая поверхность, обработанную ConA.
- Добавьте 30 мкл предварительно подготовленного образца (этап 2.6) в лунку и осторожно повторно суспендируйте, чтобы избавиться от любых кластеров клеток.
- Закройте полость, осторожно поместив крышку 22 x 22 мм #1.5 поверх ободков PDMS (не позволяйте ему резко упасть, используйте щипцы, если это возможно, рисунок 2B, C).
ПРИМЕЧАНИЕ: Любая толщина крышки будет работать для верхней стеклянной крышки (собирающая линза имеет рабочее расстояние 2 мм).
- Добавьте 100 мкл конканавалина А (ConA) по 0,5 мг/мл, чтобы покрыть всю поверхность квадратной полости, и дайте ему инкубироваться в течение 30 мин.
- Камерная подготовка к экспериментам с клетками в конфайнменте
- Поместите 10 мкл капли раствора, содержащего клетки (шаг 2.6), в квадратную полость (рисунок 2B).
- Очень аккуратно сэндвич образец с покрытием стекла размером 22 х 22 мм таким образом, чтобы капля распространялась по всей площади и не наблюдалось пузырьков. Опять же, удобно использовать щипцы, как показано на рисунке 2C, чтобы предотвратить резкое падение покровного стекла.
4. Альтернативные варианты интервала между камерами OT
ПРИМЕЧАНИЕ: Эти шаги могут быть выполнены, если нет мастерской микропроизводства или прядильного коатера.
- Камерная подготовка к экспериментам с клетками в суспензии
ПРИМЕЧАНИЕ: В случае, если спин-коатер недоступен, распорка может быть изготовлена с использованием обычной, двусторонней скотча (высота около 100 мкм).- Вырежьте кусок двусторонней скотча с квадратным отверстием размером примерно 10 см х 10 см в центре (те же размеры, что и в PDMS, рисунок 2B).
- Снимите один из защитных слоев ленты, сняв его, и поместите непокрытую сторону ленты в центр тарелки со стеклянным дном No 1,5 H. Осторожно надавите, чтобы вся поверхность прилипла к стеклу, избегая пузырьков воздуха, а затем удалите оставшийся защитный слой ленты, отклеив его.
- Следуйте инструкциям в шаге 3.2.
- Камерная подготовка к экспериментам с клетками в конфайнменте
ПРИМЕЧАНИЕ: Для точного ограничения клеток монодисперсные микрочастицы с известным диаметром могут быть использованы в качестве распорок между двумя закрывающими стеклами.- Добавьте 10 мкм полистирольных шариков в взвешенные ячейки в концентрации 104 шарика/мкл.
- Нанесите каплю раствора размером 10 мкл, содержащую ячейки и шарики, на покровное стекло размером 22 х 60 мм.
- Очень аккуратно сэндвич образец с еще одним 22 х 60 мм покрытым стеклом так, чтобы капля распространялась по всей площади и не наблюдалось пузырьков. Чтобы аккуратно расположить верхнюю крышку стекла (не допускайте, чтобы оно резко опускалось вниз), удобно использовать щипцы.
- Поскольку образец может высохнуть, рекомендуется выполнять подготовку быстро.
5. Настройка оптической ловушки для внутриклеточных измерений
ПРИМЕЧАНИЕ: Следующие шаги оптимизированы для коммерческой платформы оптического пинцета, состоящей из модуля оптической микроманипуляции на основе акустооптического отклонения (AOD) и датчика оптической силы, основанного на прямом обнаружении изменений светового импульса (рисунок 2, ссылка12,40,49). Подробную информацию и оптические компоненты установки можно найти на рисунке 2F. Для наблюдения вызванной силой деформации во время оптических манипуляций пинцетом конфокальный микроскоп Nipkow spinning-disk соединен с левым портом инвертированного микроскопа для визуализации двойной цветной флуоресценции. Без отсутствия общности этот протокол может применяться к любой динамической системе КН, оснащенной прямыми измерениями силы на основе обнаружения светового импульса. Подробные пошаговые процедуры доступны для создания самодельных оптических градиентных ловушек для приложений in vivo50. Те, которые основаны на модуляции AOD, выделяются для возможных экспериментов с несколькими ловушками и быстрыми измерениями51,52. В литературе существует несколько протоколов для построения прибора на основе светового импульса36,39,40,53, и можно использовать любую другую модальность визуализации (дифференциальный интерференционный контраст, широкоугольная флуоресценция и т.д.).
- Запуск оптического пинцета
- Чтобы оптимизировать стабильность выходной мощности, включите лазер со значительно высокой мощностью (например, 3 Вт) не менее чем за 30 минут до начала эксперимента.
- Включите электронный модуль единиц оптического микроманипуляции и измерения силы.
ПРИМЕЧАНИЕ: Применяйте все меры лазерной безопасности и используйте только оборудование, одобренное институциональным советом. Никогда не используйте окуляры оптического микроскопа, когда лазер включен. Всегда используйте утвержденные очки ИК-защиты (OD7 в диапазоне 950-1080 нм), блокируйте ИК-лазерный свет затвором в порту эпифлуоресценции 2 и не выполняйте программное обеспечение оптического захвата до завершения выравнивания оптического датчика силы после шага 5.3. В общем, не используйте образец с высокой отражающей способностью, так как обратное отражение может привести к повреждению лазера. - Управляйте мощностью ловушки с помощью вращающегося HWP (рисунок 2F) на входе в модуль оптической микроманипуляции.
ПРИМЕЧАНИЕ: Коммерческий оптический модуль микроманипуляции, используемый в этом протоколе, уже включает в себя эту функцию. Для самодельных оптических систем улавливания интегрируйте этот инструмент для управления мощностью, чтобы можно было использовать более высокие и стабильные мощности лазера.
- Использование пустой микрокамеры для калибровки
- Вырежьте квадрат размером 1 х 1 см на двусторонней скотч-ленте и прикрепите его к слайду микроскопа толщиной 1 мм.
- Добавьте воду в квадрат и закройте его сверху крышкой #1.5 (22 x 22 мм). Добавление немного большего объема воды, например, 30-40 мкл, рекомендуется избегать пузырьков внутри закрытой камеры. Аккуратно протрите калибровочную камеру в случае выплескивания из нее воды.
- Выравнивание оптического датчика силы
- Поместите каплю воды на цель погружения в воду 60x/1.2. Поместите калибровочную камеру на сцену со стеклянным покрытием No1.5, обращенным к объективу. Сосредоточьтесь на нижней поверхности, где в конечном итоге окажутся образцы клеток.
- Добавьте каплю погружного масла поверх верхнего стеклянного слайда, покрывающего образец (рисунок 2D). Осторожно опустите собирающую линзу блока датчика силы, пока она не соприкоснется с каплей масла.
ПРИМЕЧАНИЕ: Капля должна быть достаточно большой, чтобы она покрывала всю линзу, которая собирает лазерный свет, выходящий из ловушек. Обычно 200 мкл достаточно, чтобы покрыть всю поверхность и обеспечить стабильный погружной контакт. Будьте консервативны и избегайте переполнения, так как оно может просочиться в образец. - Следуя протоколу производителя для выравнивания оптического датчика силы, посмотрите на изображение плоскости образца на вспомогательной камере, которое будет использоваться для позиционирования ОТ (AUX, рисунок 2F). Очень осторожно опустите датчик оптической силы до тех пор, пока на плоскости образца не появится точка поля (FS, рисунок 2F-G). Это обеспечит надлежащие измерения прямой силы от инвариантного обнаружения изменений светового импульса40.
ПРИМЕЧАНИЕ: Закройте FS достаточно, чтобы его изображение стало меньше поля зрения (FOV), следовательно, видимым. Будьте особенно осторожны и не прижимайте собирающую линзу датчика оптической силы к образцу. Вертикальное положение датчика оптической силы может быть альтернативно определено на основе анализа распределения улавливающего света на BFP для световых конусов с определенной числовой апертурой (NA). - Убедитесь, что в капле масла нет пузырьков воздуха; они могут непосредственно влиять на измерения силы. Чтобы проверить наличие пузырьков воздуха, поставьте линзу Бертрана на место (BL, рисунок 2G) и наблюдайте за траекторией изображения через окуляр. Если видны какие-либо пузырьки грязи или воздуха или требуется больше масла (рисунок S1A), очистите линзу и камеру беспыльной тканью хрусталика и повторите процедуру на этапах 5.3.2 и 5.3.3. Беспрепятственный оптический путь изображен на рисунке S1B.
- Используя боковые винты, размещенные на держателе оптического датчика силы, центрируйте FS в FOV. Для точности откройте FS так, чтобы он почти заполнил FOV, видимый на вспомогательной камере (AUX, рисунок 2F).
6. Оптимизация оптического пинцета
ПРИМЕЧАНИЕ: Измерение прямой силы опирается исключительно на изменение светового импульса, возникающего в результате силы, оказываемой на захваченную частицу, и, таким образом, в отличие от косвенных методов, жесткость ловушки не нужно калибровать перед каждым экспериментом. Конкретное для прибора преобразование коэффициента отклонения/силы (α; pN/V, reference41) калибруется изготовителем и, таким образом, является инвариантом эксперимента. Однако, поскольку лазерным пятном манипулируют на площади 70 мкм х 70 мкм, шаги 6,2-6,5 имеют решающее значение для обеспечения оптимального улавливания и стабильности мощности. Следующие шаги поставляются в программном обеспечении производителя, чтобы OT были оптимизированы над рабочей зоной полуавтоматическим способом.
- Запустите программное обеспечение OTs и программное обеспечение для приобретения для камеры AUX.
- Вычтите начальную базовую линию напряжения, щелкнув шаг Шаг 1: Смещение электроники в подменю «Калибровка системы» программного обеспечения для управления оптическим пинцетом.
- Чтобы выполнить выравнивание мощности ловушки в рабочей зоне OT, установите мощность ловушки на половину ее максимума, соответственно повернув HWP. Изменяйте мощность ловушки не путем изменения выхода лазера, а с помощью вращающегося HWP (рисунок 2F). Нажмите на Шаг 2: Питание , чтобы инициировать автоматическую процедуру сглаживания мощности ловушки.
ПРИМЕЧАНИЕ: Это критический шаг для компенсации изменения мощности ловушки в рабочей зоне КН (рисунок S1D). Успешная процедура снижает изменение мощности ловушки до 2% в рабочей зоне OT и сходится через 2 минуты. - Чтобы выполнить калибровку положения ловушки, снимите ИК-фильтр так, чтобы свет от лазера был виден на камере. Найдите ИК-пятно, установив плоскость изображения, сфокусированную на нижней поверхности микрокамеры. Получите наименьшее возможное ИК-пятно, настроив плоскость изображения (положение объектива) и контрастность гистограммы в программном обеспечении для сбора AUX камеры. При необходимости уменьшите мощность оптической ловушки, вращая HWP (рисунок 2F). Нажмите на Шаг 3: Позиционирование , чтобы начать автоматическую калибровку процедуры или позиционирования ловушки.
ПРИМЕЧАНИЕ: Эта процедура позволяет точно соответствовать координатам положения OT в камере AUX углам поворота AOD. Успешная процедура создает сопоставление угла с положением за несколько секунд. - Компенсация начального импульса
ПРИМЕЧАНИЕ: Движение оптической ловушки по образцу вызывает изменения в распределении светового импульса на BFP (рисунок S1E, F). Это приводит к независимым от силы изменениям сигнала, связанным с положением лазера над рабочей зоной, даже если мощность ловушки была сглажена, как на шаге 6.3. Следствием этого является изменение исходного уровня силы из-за положения (независимо от фактической силы, действующей на оптически захваченную бусину), которое необходимо корректировать перед каждым экспериментом.- Установите мощность ловушки, которая будет использоваться в экспериментах, вращая HWP (рисунок 2F).
- Нажмите на опцию «Глобальное смещение » в подменю «Инструменты ». Это откроет помощник Offset Cancel программного обеспечения оптического пинцета, который корректирует исходный уровень импульса.
- Нажмите на смещение | Компенсировать для коррекции начального импульса позиционного варианта.
ПРИМЕЧАНИЕ: Если в течение текущих недель на оптический путь не повлияет никакая модификация, карты мощности ловушки (шаг 6.3) и положения (шаг 6.4) останутся инвариантными. Поэтому мы рекомендуем всегда использовать одну и ту же комбинацию оптических элементов (дихроичные зеркала, фильтры и т. Д.), Которые могут повлиять на траекторию лазерной ловушки, или выполнять новую процедуру сплющивания мощности ловушки. Что касается компенсации начального импульса (этап 6.5), то производитель платформы ОТ обеспечивает калибровку на лету, которая должна изменяться для каждой новой мощности улавливания и экспериментальной сессии. Этапы 6.3 и 6.4 должны выполняться на пустом калибровочном слайде, описанном на этапе 5.2. В образце, содержащем ячейки или другие объекты, этап 6.5 должен осуществляться без объектов, которые могут изменять рассеяние света в рабочей зоне КН.
- Опционально поймайте микросферу и переместите ловушку с известной скоростью во время записи сигнала силы. Например, установите ловушку для выполнения треугольного колебания: регистрируемый силовой сигнал будет квадратным сигналом.
ПРИМЕЧАНИЕ: Значение силы должно линейно увеличиваться со скоростью, в соответствии с силой сопротивления, действующей на шарик. Этот тест служит положительным контролем того, что измерения силы проводятся правильно38. Альтернативно, датчик оптической силы может быть использован для получения жесткости оптического улавливания, κ [pN/μm], и коэффициента калибровки положения, β [мкм/В], из спектрального анализа мощности35. При правильном выравнивании инвариантный калибровочный коэффициент, предоставленный заводом-изготовителем, составляет α = κ·β [pN/V].- Инициируйте считывание силы в режиме реального времени, щелкнув график 1 в подменю «Меры» в программном обеспечении производителя. Это обеспечит считывание текущей оптической силы улавливания и мощности.
- Откройте диалоговое окно Параметры колебаний в подменю Сервис . Задайте форму сигнала треугольного пространства в кольцах селектора «Фигура» и «Тип» соответственно. В качестве примера задайте амплитуду 10 мкм и частоту 3 Гц. Это приведет к вязкой силе примерно 1 пН на микрогранулу диаметром 1 мкм38.
- В окне AUX камеры щелкните правой кнопкой мыши микрошарик и выберите Начать осциллирование. Показания силы станут квадратным силовым сигналом с плато на ±1 пН.
- Щелкните правой кнопкой мыши на микрогрануле и выберите Остановить колебание.
7. Вращающаяся дисковая конфокальная микроскопия
- Включите вращающийся дисковый конфокальный микроскоп и вспомогательное оборудование, интегрированные лазерные двигатели и камеры сбора.
- Запустите программное обеспечение для создания образов.
- Установка каналов визуализации для окрашивания ядра Hoechst и GFP для клеточной плазматической мембраны.
- Активируйте линии возбуждения 405 нм и 488 нм.
- Добавьте многодиапазонный дихроик, чтобы отразить возбуждение к образцу, и это позволит излучаемому свету проходить к камерам.
- Разделите флуоресцентное излучение с помощью дихроичного зеркала с длинным проходным краем 500 нм.
- Используйте эмиссионные фильтры DAPI/BFP (~445 нм) и GFP (~521 нм) перед двумя камерами сбора данных соответственно. См. рисунок 2F,G.
- Установите время экспозиции равным 100 мс для каждого канала.
- Установите лазерное излучение для получения мощности 5 мВт в плоскости образца. Чтобы измерить мощность, используйте коммерческий измеритель мощности.
- Задайте протокол обработки изображений. Чтобы избежать спектрального кровотечения из канала Хёхста в канал GFP, два красителя должны быть изображены последовательно.
ПРИМЕЧАНИЕ: Если существует аппаратная синхронизация между АОД оптической ловушки и камерой захвата, убедитесь, что полярность триггера настроена правильно. Если вы сомневаетесь, проконсультируйтесь с менеджером вашего объекта или производителем микроскопа.
8. Проведение экспериментов по углублению ядра
ПРИМЕЧАНИЕ: Всегда выключайте оптические ловушки - как с помощью программного обеспечения, так и закрывая затвор на порте эпифлуоресценции 2 - при подъеме модуля датчика силы и замене образца. Если нет, то могут произойти серьезные повреждения оптических элементов и экспериментатора. Будьте осторожны с боковым расстоянием между держателем линзы и краем нижней тарелки при поиске клеток, чтобы избежать столкновения линзы со сценой / чашкой для культивирования (рисунок 2).
- Поместите образец в микроскоп и выполните шаг 5.3 этого протокола.
- Используя вращающийся HWP (рисунок 2F), установите мощность ловушки на 200 мВт в качестве начального значения, если жесткость исследуемого ядра или внутриклеточной структуры неизвестна. Переведите рабочую область КН (с помощью ступени микроскопа) в место, свободное от клеток, чтобы компенсировать начальную базовую линию импульса на этапе 6.5.
ПРИМЕЧАНИЕ: В зависимости от жесткости субклеточной структуры значение мощности ловушки должно быть скорректировано до более низких или более высоких значений для получения аналогичной глубины отступа. - Используя программный контроллер ступени микроскопа, ищите ячейку с одной или двумя шариками с помощью передаваемой микроскопии яркого поля (рисунок 3A).
- Определите траекторию ловушки.
- Откройте диалоговое окно «Траектория » в подменю «Инструменты» и выберите «Смещение» в кольце селектора «Тип траектории ».
- В числовом листе запишите смещение и время каждого последующего шага траектории. Вот два примера.
- Для эксперимента по расслаблению стресса запрограммируйте трапециевидные нагрузки, как показано на рисунке 3B. В таблице S1 были нанесены два трапециевидных углубления с расстоянием перемещения 5 мкм; скорость 5 мкм/с; время ожидания перед втягиванием: 10 с.
- Для повторяющегося эксперимента по отступу с постоянной скоростью для получения треугольной рутины без времени пребывания на ядре задайте амплитуду траектории, например, 5 мкм, и время для шага, например, 2 с для скорости 2,5 мкм/с. В таблице S2 это применяется восемь раз при одинаковой скорости.
ПРИМЕЧАНИЕ: Эти значения должны быть определены для каждого типа клеток и эксперимента, но следующие параметры трапециевидной процедуры отражают наиболее важную динамику в эксперименте, представленном здесь. Время ожидания должно быть достаточным для того, чтобы ядро показало полное расслабление напряжения после отступа.
- Захват микросферы
- Установите плоскость изображения немного выше бусины с помощью программного контроллера ступени микроскопа.
- Активируйте ловушки с помощью программного обеспечения OTs и нажмите на шарик в окне изображения камеры AUX (откалибровано после шага 6.4). Успешное удержание шарика оптической ловушкой сильно уменьшит движение шарика.
- Нажмите и перетащите шарик через цитоплазму и поместите его на расстояние ~2 мкм от ядерной оболочки (рисунок 3A). Убедитесь, что траектория установлена таким образом, чтобы углубление шарика было перпендикулярно ядерной мембране.
- Опционально, если это необходимо для измерения положения шарика относительно ловушки, сканируйте ловушку поперек шарика, чтобы определить жесткость улавливания, k [pN/μm]54, тем самым Δxbead = -F/k (см. Обсуждение). Оптический модуль микроманипуляции, используемый в этом протоколе, имеет встроенную процедуру для этой цели.
- Откройте диалоговое окно «Сканирование частиц » в подменю «Инструменты ».
- Выберите ловушку, которую вы хотите сканировать, и Высокую частоту в качестве метода сканирования. Выберите направление (x или y) траектории отступа для измерения сканирования шариков.
- Появится окно с измерением жесткости улавливания. На графике перетащите два курсора, чтобы выбрать область линейного треппинга, соответствующую F = -kx. Линейная подгонка к выбранной части данных будет обновлена автоматически.
ПРИМЕЧАНИЕ: Установите начальное положение шарика вдали от клеточной мембраны (~5 мкм), так как отклонения светового импульса на границе раздела средней ячейки влияют на целесообразность измерений силы. Если ядро расположено слишком близко к клеточной мембране, попробуйте отступить ядро от противоположного участка. Удалите ячейку, если это невозможно.
- Начните сбор изображений, нажав на кнопку получения в программном обеспечении для обработки изображений.
- Начните экономию данных о положении ловушки и измерении силы, нажав на | Сохраните в окне принудительного считывания в реальном времени (открывается, как в шаге 6.6.1).
ПРИМЕЧАНИЕ: Оптическая ловушка оснащена триггерным входом, который может быть подключен к выходу синхронизации камеры. Таким образом, данные изображения и силы аппаратно синхронизированы, и электронный способен отображать циклы ловушек с количеством кадров изображений во время съемки. - Инициируйте ранее загруженную траекторию, щелкнув правой кнопкой мыши по шарику и выбрав Стартовая траектория.
- Подождите, пока траектория не закончится и система стабилизируется.
- Сохранение данных измерения силы остановки ловушки. Появится диалоговое окно сохранения данных.
ПРИМЕЧАНИЕ: Чтобы оптимизировать хранение данных, данные могут быть уничтожены, выбрав параметр децимации в этом диалоговом окне (10, 100 или 1000). - Остановите получение изображений и отобразите результаты в программном обеспечении постобработки по выбору пользователя.
- Если микросфера теряется во время рутины и ядро не может быть вдавлено (рисунок S2), отбросьте измерение и увеличьте мощность. Обратите внимание, что шаг 6.5 необходимо повторить. В наших руках не менее 95% процедур успешно выполняются без потери бусины из ловушки.
Результаты
Микроинъекция улавливающих шариков:
Микросферы, вводимые в одноклеточный эмбрион рыбки данио, распространяются по всей шляпке животных во время морфогенеза. Для более четкой визуализации мы повторили протокол инъекции с красными флуоресцентными микрошариками и сделали объемные изображения с помощью нашего конфокального микроскопа на разных этапах развития. На рисунке 4A-D введенные шарики визуализируются в цитоплазме стволовых клеток-предшественников in vivo при 5 hfp. Позже микросферы оказались распределенными по всему эмбриону при 24 hpf (рисунок 4E). Эмбрионы на обеих стадиях развивались нормально, и показатели выживаемости были сопоставимы с контрольными эмбрионами без инъекций или имитацией инъекций (см. Рисунок S3). Это согласуется с другими исследованиями, которые сообщают о невозмутимой выживаемости рыбок данио, введенных бисером, до 5 дней после оплодотворения55.
Наш вращающийся дисковый конфокальный микроскоп совместим с многоканальной флуоресцентной микроскопией. На рисунке 5А мы показываем изолированные стволовые клетки с одной или двумя шариками в цитоплазме. Несколько флуоресцентных этикеток могут быть использованы для исследования различных аспектов ячейки (рисунок 5B). Ядерная морфология может быть отслежена с помощью красителя Hoechst или с использованием экспрессии мРНК H2A::mCherry, в то время как внутренняя ядерная мембрана может быть проанализирована с помощью Lap2b-eGFP12. Динамику кортекса актомиозина, а также внутриклеточные уровни кальция можно наблюдать при инкубации трансгенной линии My12.1::eGFP56 и Calbryte-520 соответственно. Протокол, который был описан здесь, направлен на сравнение механики клеточного ядра иммобилизованных клеток дикого типа на адгезивных субстратах (позже называемых суспензией) и в механическом заключении. Изолированные стволовые клетки, заключенные в микрокамеры высотой 10 мкм, демонстрировали частичное развертывание внутренней ядерной мембраны (INM) и последующее увеличение сократимости актомиозина12. На рисунке 5С показаны замкнутые клетки с одной или двумя шариками в цитоплазме. Успешное удержание будет видно через уплощенные, расширенные клетки с более широким поперечным сечением ядра. Ядерная мембрана дополнительно разворачивается в ограниченных клетках и должна казаться сглаженной по сравнению с клетками в суспензии (рисунок 5C).
Анализ силы-времени и деформации силы
Анализ полученных результатов сильно зависит от исследуемого образца и интересующего их вопроса и поэтому они не могут быть обобщены здесь. Например, распространенным способом анализа измерения отступа является извлечение модуля Юнга путем подгонки модифицированной модели Герца к данным силового отступа57. Однако предположение о такой обработке нуждается в тщательной оценке и не всегда может быть должным образом обосновано (например, исследуемая структура является изотропной, однородной, с линейной упругостью и углублениями, меньшими, чем радиус шарика). Таким образом, мы рассматриваем здесь только независимые от модели измерения, которые позволяют сравнивать механическое поведение исследуемой структуры между различными экспериментальными сценариями.
В качестве отправной точки измерение наклона кривой силы-смещения при определенной глубине отступа обеспечивает измерение моделью независимой структурной жесткости58 ядра. Затем это значение может быть собрано из нескольких образцов и сравнено между различными экспериментальными условиями и возмущениями образца.
Измерение отступов
В следующих строках мы сосредоточимся на механической реакции клеточного ядра во время деформации клетки в заключении. Эксперименты на этапе 8 этого протокола обычно приводят к пикам силы до 200 пН для глубины отступа приблизительно 2-3 мкм. Однако эти значения могут быть в значительной степени различными, в зависимости от типа клетки и экспериментальных условий, причем более мягкие ядра приводят к снижению силы для данного углубления. Таким образом, необходимо точно измерить ядерную деформацию вместе с силой для точной механической характеристики ядра клетки. В этом разделе мы получим жесткость ядра ячейки из измерений отступа репрезентативной силы.
На рисунке 6 показаны деформации дистальной и проксимальной сторон ядра во взвешенной и замкнутой клетке. Можно наблюдать богатое механическое поведение. В типичной взвешенной клетке на адгезивной подложке ядро было сильно вдавлено шариком, но также слегка смещено при повторяющихся событиях толкания. Мы измерили углубление шарика на ядре, проанализировав кимографы, полученные из флуоресцентной визуализации ядер клеток, окрашенных Хёхстом. Кимографы были легко вычислены с использованием плагина Multi Kymograph Fiji вдоль направления отступа (рисунок 6A, B) и импортированы в Matlab (версия 2021, Mathworks) для дальнейшей обработки. К необработанному профилю интенсивности была установлена ступенчатая функция с целью отслеживания разграничительных краев ядра по траектории процедуры отступа. Как видно, он содержит точную информацию об изменении формы ядерного оружия (рисунок 6 и рисунок S2). В качестве аналитической версии ступенчатой функции мы использовали следующую двойную сигмовидную кривую:
 (Уравнение 1)
(Уравнение 1)
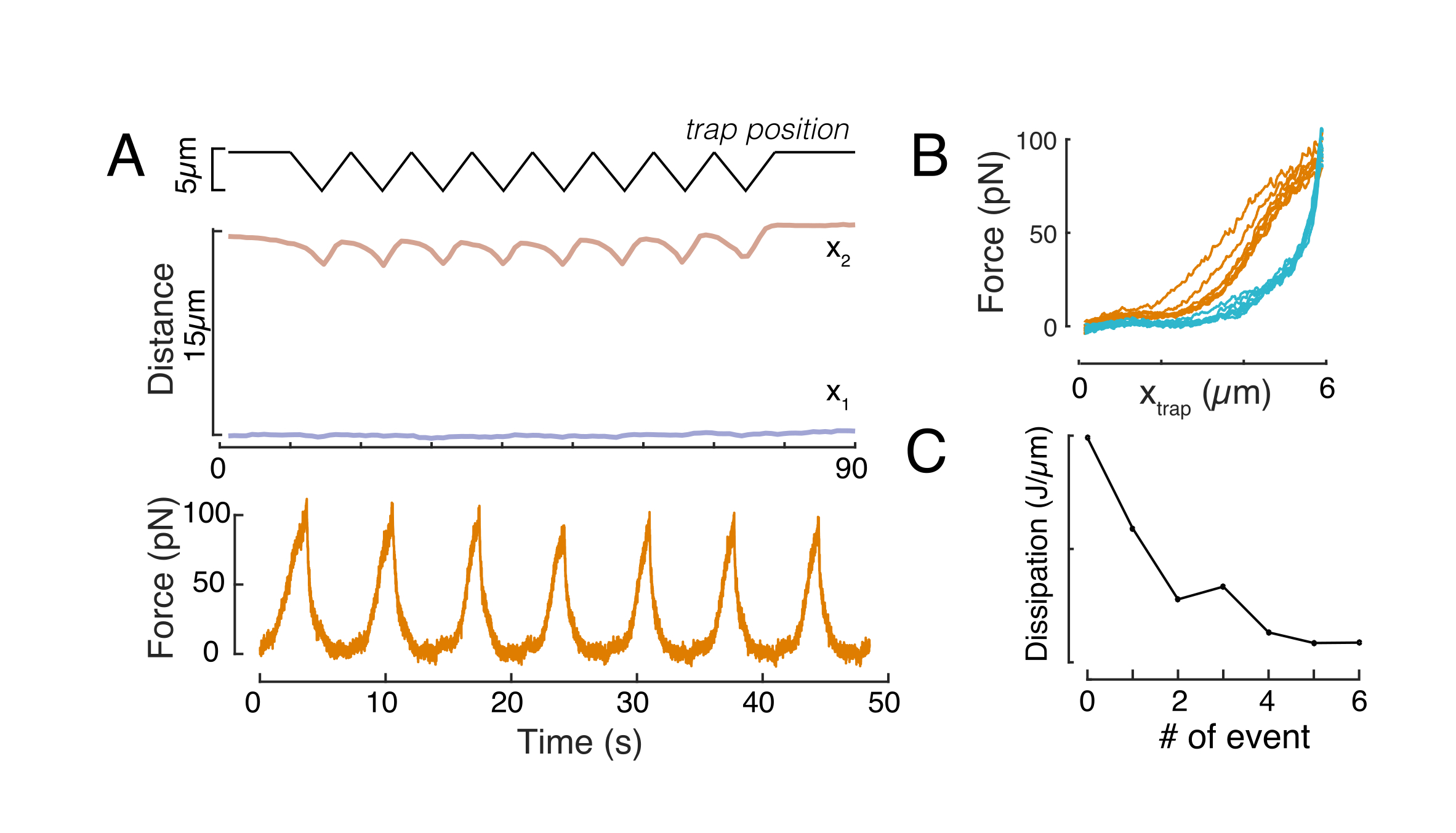
Здесь x1 и x2 обозначают дистальный и проксимальный края ядра, в то время как A и B являются максимальными и фоновыми серыми значениями синего канала (краситель Hoechst) изображения (рисунок 6B). Рассмотрена ширина кромки (e0 = 0,25 мм). В то время как отступ, проксимальный край ядра (x2) следовал траектории, применяемой процедурой оптической ловушки после контакта микросферы с ядром, противоположный, дистальный край (x1) отображает динамику релаксации, как и ожидалось для вязкоупругого материала, такого как цитоплазма (рисунок 6D). Напротив, ядра в клетках, заключенных в микрокамерах высотой 10 мкм, не проявляют такого транслокационного поведения ядра при углублении внутри клетки (рисунок 6B, D). Также показанные на рисунке 6D, задние края ядер остаются неизменными при выталкивании шарика с проксимальной стороны, скорее всего, из-за более сильных сил, возникающих из-за сократимости клеток и трения, действующих против силы отступа. Чтобы получить правильную глубину деформации, смещение x1 вычитали из меры с отступом x2: Δx = x2 - x1 (см. также рисунок 6D).
Анализ силовых данных
Сила, вызывающая ядерную деформацию, измерялась изменением светового импульса, происходящего в оптически захваченной микрогрануле (рисунок 7А). Сила при применении трапециевидных траекторий (шаг 8.4.3, рисунок 7В) первоначально увеличивалась линейно до тех пор, пока ловушка не переставала двигаться, но затем расслаблялась до значения устойчивого состояния. Такое поведение указывало на то, что вязкоупругий материал проявляет потери и модули хранения. Сразу после события отступа сила достигла пикового значения, Fp, за которым последовало расслабление напряжения (рисунок 7C):
 (Уравнение 2)
(Уравнение 2)
где F0 — хранимая сила для упругой составляющей, а f(t) — безразмерная релаксационная функция. Мы проанализировали это поведение тремя способами:
1. Рассмотрим стандартное линейное твердое тело с экспоненциальной релаксацией напряжений, т.е. f(t) = e-t/τ, схематично представленное на рисунке 7C вставке.
2. Использование общего, двойного экспоненциального распада:
F(t) = A + B1e-t/τ1 + B2e-t/τ2.
3. Использование степенного закона с последующим экспоненциальным распадом59:
f(t) = t-pe-t/τ, показанный на рисунке 7C.
Хотя подгонка для модели 1 может быть выполнена напрямую, мы рекомендуем оценить начальные догадки для (τ1, τ2) и (p, τ) для моделей 2 и 3 соответственно. Это может быть выполнено, соответственно, путем подгонки линий к данным в логарифмических и линейных (рисунок 7D, слева) и логарифмических против логарифмических (рисунок 7D, справа) масштабах. В таблице S3 обобщены результаты для примера, проанализированного на рисунке 7. В следующем разделе мы рассмотрим сочетание степенного закона и экспоненциального закона для характеристики механики клеточного ядра.
Отношение смещения силы
Аналогичным образом, описанная экспериментальная установка может быть использована для получения соотношения сила-смещение множественных событий отступа. Выполняя треугольные процедуры (шаг 8.4.4, рисунок 8А), можно соотнести силу с деформацией и построить кривую силы-отступа. Примерный результат показан на рисунке 8B, в котором плоская базовая линия плавно изменяла наклон, как только шарик соприкасался с ядром. Идентификация истинной точки контакта в шумных данных является сложной задачей, и необходимо позаботиться о том, подходит ли область контакта к эластичным моделям60. В этом конкретном эксперименте также можно было увидеть, что последующие отступы приводят к кривым с более глубокими точками контакта, что свидетельствует о слишком медленном восстановлении ядерной формы после втягивания шарика и изменении гистеретического цикла, определяемого свойствами вязкоупругого материала ядра (рисунок 8C). Таким образом, исследователь должен знать, если это происходит, и включить это в аналитический конвейер или ограничить количество последующих измерений таким образом, чтобы этот эффект не изменил измерение.
Механика ядра в клетках в суспензии и с удержанием менее 10 мкм
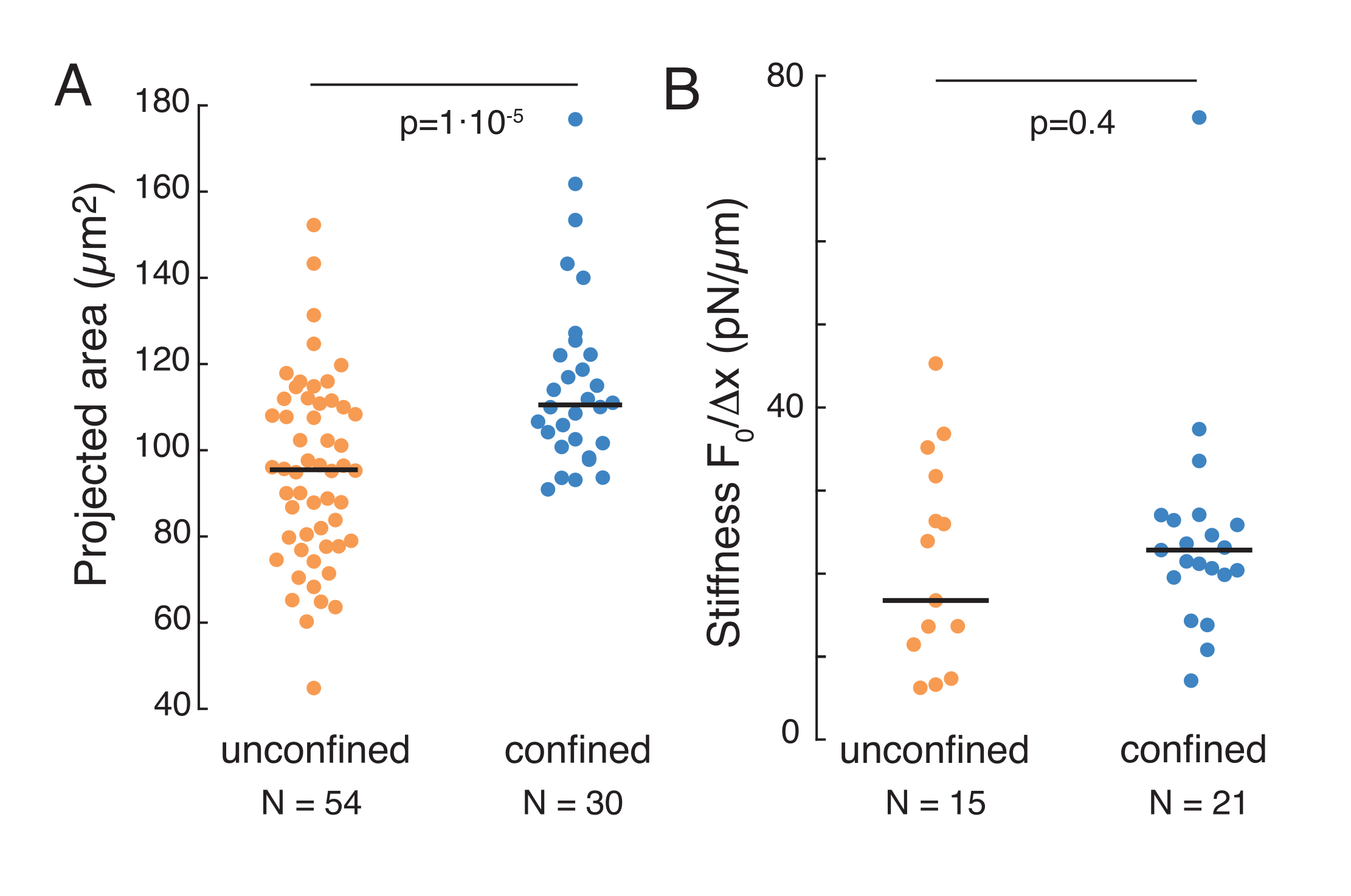
Вышеупомянутый подход был использован для анализа динамики релаксации напряжения ядра в взвешенных клетках на адгезивных субстратах и замкнутых клетках. Наши результаты показывают, что ограничение приводит к расширению прогнозируемой площади (рисунок 9A), но незначительному изменению жесткости ядра (рисунок 9B). Мы измерили аналогичную релаксацию с τ = 6,08 ± 1,1 с (неограниченная) и τ = 4,00 ± 0,6 с (удержание), что указывает на быстрое вязкоупругое рассеивание, за которым следует сохраненное значение силы, соответствующее модулю упругости ядра. Чтобы учесть экспериментальные вариации, которые могут быть вызваны различными начальными условиями в процедурах отступа, измеренные сохраненные силы были нормализованы до глубины отступа, как  . Этот параметр учитывает жесткость ядра и описывает силу, или напряжение, необходимое для определенного отступа. Мы получили аналогичную жесткость в условиях удержания и в неограниченных камерах: = 20,1 ± 12,6 пН/мкм и = 24,6 ± 13,6 пН/мкм (среднее ± стандартного отклонения) соответственно.
. Этот параметр учитывает жесткость ядра и описывает силу, или напряжение, необходимое для определенного отступа. Мы получили аналогичную жесткость в условиях удержания и в неограниченных камерах: = 20,1 ± 12,6 пН/мкм и = 24,6 ± 13,6 пН/мкм (среднее ± стандартного отклонения) соответственно.

Рисунок 1: Микроинъекция эмбрионов рыбок данио на одноклеточной (зиготной) стадии. (А) Инъекционная пластина: для инъекции используется инъекционная пластина треугольной формы. Пластина изготовлена из 1% сверхчистой агарозы в Е3 (среда яйца). Вид сверху и сбоку показаны справа. (B) Позиционирование эмбрионов: мягко ориентируйте эмбрионы с помощью кисти и ориентируйтесь таким образом, чтобы одноклеточный элемент был хорошо виден и легко доступен с помощью иглы. Мы предлагаем сориентировать эмбрионы клеткой, расположенной на противоположной стороне иглы, как показано на рисунке. (C) Процедура инъекции в эмбрион с одноклеточной стадией: прокалывание хориона, окружающего эмбрион, и одной клетки иглой. Убедитесь, что кончик иглы находится внутри клетки и отпустите давление для инъекции. (D) Инкубировать эмбрионы при 28-31 °C до тех пор, пока они не разовьются до стадии бластулы (сферы) (4 л.с.ф.). Выполняют протокол изоляции клеток и окрашивание клеток (этап 2) и подготавливают оптическую улавливающую камеру с изолированными ячейками в суспензии и/или конфайнменте в сочетании с соответствующим покрытием поверхности подложки (этап 3). Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 2: Подготовка аппарата оптического пинцета. (А) Спин-покрытие слоев PDMS с заданной высотой на стеклянную нижнюю посуду. Падение PDMS будет распределяться равномерно из-за центробежной силы. (B) Подготовка пробоотборной камеры вне слоя PDMS. 1: вырезать квадрат скальпелем, 2: покрыть внутренний колодец конканавалином А (ConA), промыть и посеять клетки; 3: накройте стеклянной горкой или крышкой, чтобы запечатать колодец. (C) Изображение квадратной резки скальпелем и снятия колодца PDMS щипцами. D) Установка съемной линзы датчика оптической силы над камерой улавливания. Капля погружного масла служит средой погружения между собирающей линзой и верхней стеклянной крышкой. Схема не масштабируется. Будьте осторожны при опускании коллекционной линзы, чтобы не касаться стеклянной крышки тарелки для образцов. Е) Изображение блока обнаружения силы, контактирующего с образцом. F) Схема экспериментальной установки. Модуль оптической микроманипуляции использует непрерывный волновой лазерный луч (5 Вт, λ = 1064 нм) с управлением мощностью через полуволновую пластину (HWP) и поляризационный разветвитель пучка (BS). После модуляции с помощью пары AOD он соединяется с верхним портом эпифлуоресценции инвертированного микроскопа. Затем лазерный луч отражается 950-нм короткочастотным дихроичным зеркалом (IR-DM), что обеспечивает пропускание флуоресцентного возбуждения и излучения. Улавливающий лазер направляется в задний, эпифлуоресцентный порт микроскопа (верхняя башня). ОТ создаются в фокальной плоскости объектива погружения в воду (60x, NA = 1,2). Оптический датчик силы подвергается воздействию башни микроскопа и захватывает лазерный свет, исходящий от ОТ, с помощью высоко-NA, масляной иммерсионной линзы. В то же время датчик силы обеспечивает ярко-полевую подсветку. Конфокальный блок вращающегося диска соединен с левым портом. Он оснащен двумя интегрированными лазерными двигателями (ILE), которые управляют семью флуоресцентными лазерами возбуждения и двумя камерами sCMOS с задней подсветкой, что позволяет выполнять двойную флуорофорную визуализацию параллельно Abb: TI, Transilluminator; ФС, полевая остановка; AOD, акустооптический дефлектор; HWP, полуволновая пластина; CAM, камера (G) Фотография оптического улавливающего оборудования. Красный круг указывает на линзу Бертрана, которую можно переключить на оптический путь вручную. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
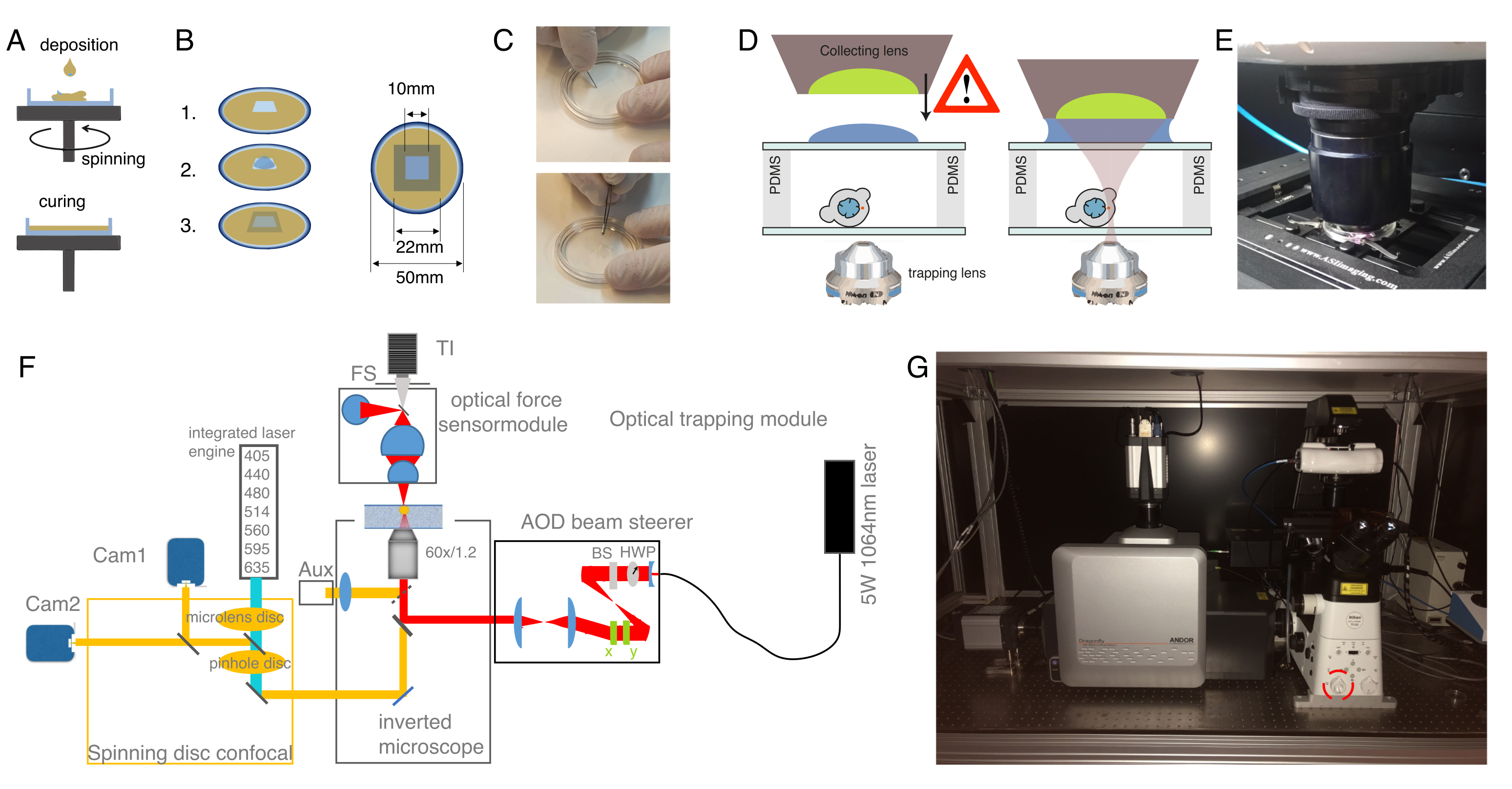{kind=link}

Рисунок 3: Выбор правильных образцов и параметров. (A) Репрезентативное изображение изолированной стволовой клетки-прародителя рыбки данио с одной микросферой, расположенной достаточно близко к ядру для выполнения эксперимента по углублению. Шкала стержня = 10 мкм. (B) Примерная траектория ловушки; глубина углубления 5 мкм; скорость отступа = 5 мкм/с; время расслабления 10 с. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
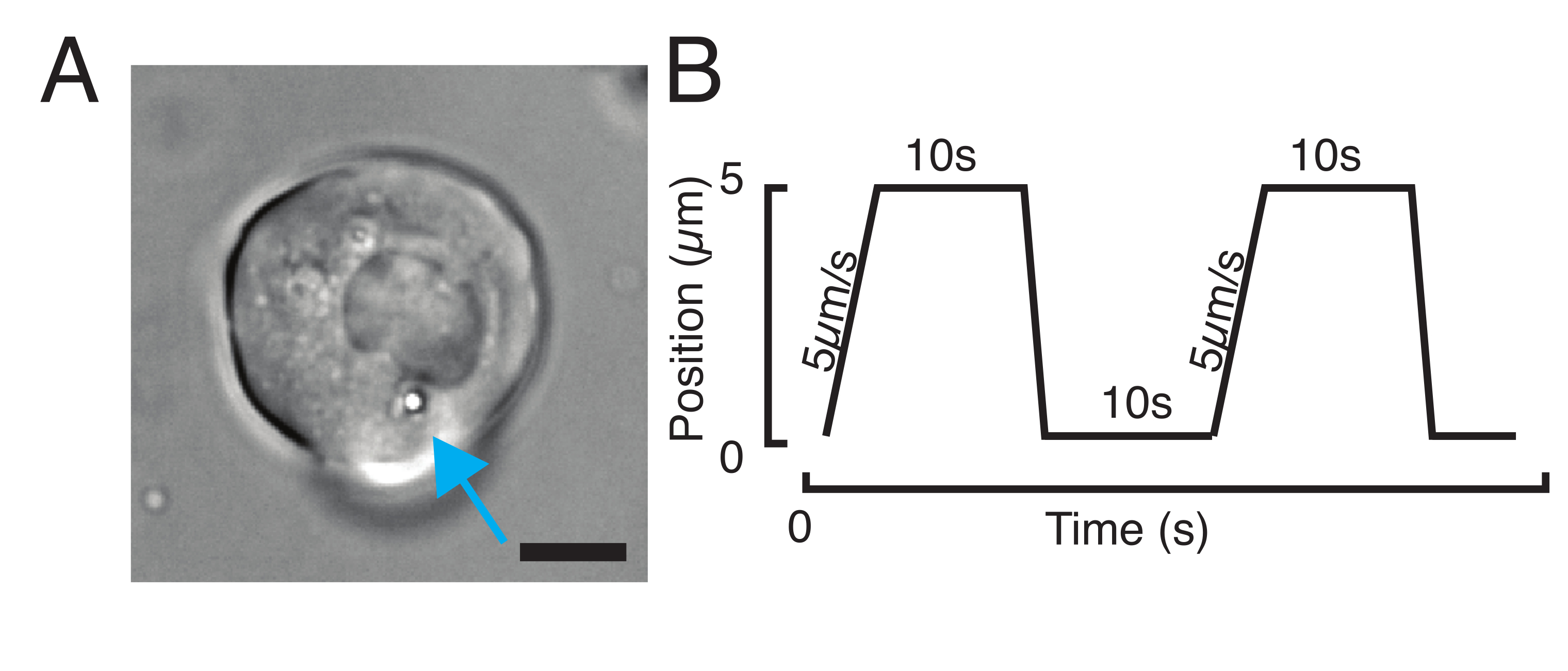{kind=link}

Рисунок 4: Локализация микрогранул внутри эмбрионов рыбок данио во время развития. 0,5 нл красных флуоресцентных шариков размером 1 мкм вводят вместе с мРНК GPI-GFP (100 пг/эмбрион, плазматическая мембрана) в эмбрионы WT для визуализации локализаций шариков. (А-Д) Распределение микросферы через 5 ч после инъекции внутри эмбриона, установленного в 0,75% агарозе. (A) Яркое поле и флуоресцентное изображение. Бусины однородно диспергированы по ткани эмбриона, как видно на конфокальной микрофотографии. (B) Максимальная проекция конфокальной флуоресценции z-стека. Бусины имеют цветовую маркировку от фиолетового до желтого в соответствии с их z-положением в стеке изображений. Фиолетовый/пурпурный соответствует большинству наружных шариков/клеток (EVL; эпителиальный обволакивающий слой; или стволовые клетки-предшественники, расположенные близко к поверхности EVL), желтый соответствует внутренним шарикам (глубинным клеткам-предшественникам), как показано на рисунке справа. (C) Разрезание и максимальная проекция подстега (B), соответствующего области в оранжевой коробке: большая часть глубоких ячеек содержит 1-2 шарика. (D) Вырезание и максимальная проекция подстега (B), соответствующего пурпурной коробке: некоторые ячейки EVL содержат 1-2 шарика. (E) Изображение Яркого поля и максимальная проекция z-стека эмбриона мощностью 24 л.с.ф., установленного в 0,75% агарозы и обезболенного трикаином. Эмбрионы предварительно инкубировали с трикаином в течение 15 мин. Слева направо: микросферы (диаметр 1 мкм), GPI-GFP и перекрытие изображения. Бусины распределены по всему телу эмбриона. Размер шкалы, указанный на каждой панели. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
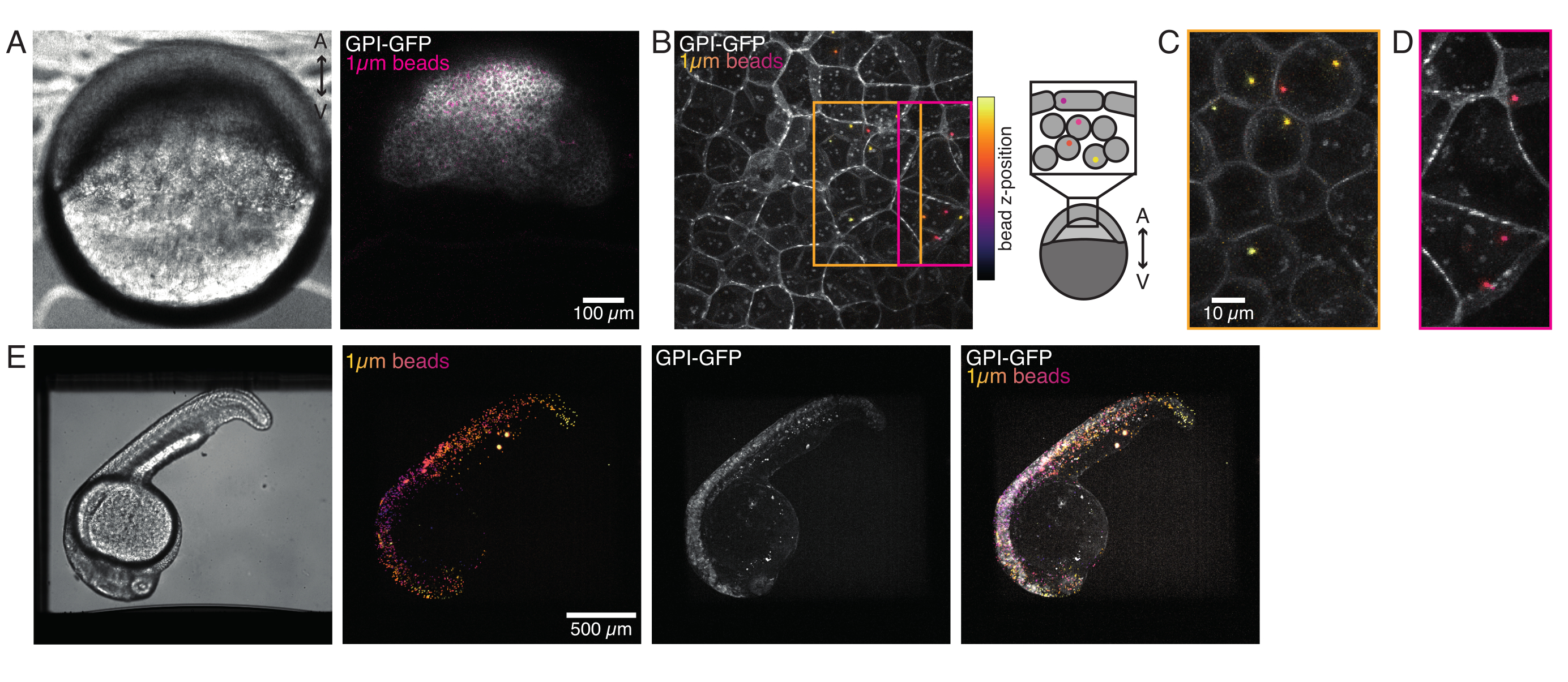{kind=link}

Рисунок 5: Изолированные стволовые клетки-предшественники рыбок данио с различной маркировкой. (A) Изображение проходящей световой микроскопии суспензионных ячеек с 1 (сверху) или 2 (снизу) введенными шариками. Голубые стрелки указывают на бусины. (B) Флуоресцентные конфокальные изображения суспензионных клеток с различными окрашиваниями. Вверху слева: Lap2b-eGFP (внутренняя ядерная мембрана, 80 пг/эмбрион) и H2A-mCherry. Вверху справа: GPI-GFP (плазматическая мембрана, 100 пг/эмбрион) и ДНК-Хёхст (окрашенный, как описано в разделе 2). Внизу слева: MyI12.1-eGFP (трансгенная линия) и ДНК-Hoechst. Внизу справа: Calbryte488 и DNA-Hoechst (окрашены, как описано в разделе 2). (C) Изображение при пропускающей световой микроскопии замкнутых клеток с 1 (сверху) или 2 (снизу) введенными шариками. Голубые стрелки указывают на бусины. Шкала = 10 мкм. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 6: Оценка ядерной деформации от фильмов вращающегося диска. (A,B) Замедленная съемка эксперимента по углублению ядра в (A) взвешенной клетке и (B) замкнутой клетке. Шкала стержня 10 мкм. Репрезентативные снимки меченых Хёхстом ядер показаны за 5 с до, во время и через 5 с после отступа с оптически захваченной микросферой (белый наконечник стрелы). Кимографы вдоль отрезка отступа (красная линия, правая панель). x1 и x2 представляют собой дистальную и проксимальную (близкую к шарику) границы ядра во время эксперимента с отступом, извлеченные из соответствия профиля интенсивности уравнению 1. (C) Профили интенсивности вдоль сегмента отступа для трех различных кадров (до, во время и после отступа) и установленные в уравнении 1 для оценки дистального, x1 и проксимального, x2, положений ребер ядра. D) репрезентативные траектории x1(t) в синем цвете и x2(t) в автожелтом цвете во время эксперимента по отступу взвешенных и замкнутых клеток (10 мкм). Затененные области указывают на отступ, расстояние между x1 и x2 указывает на диаметр ядра. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
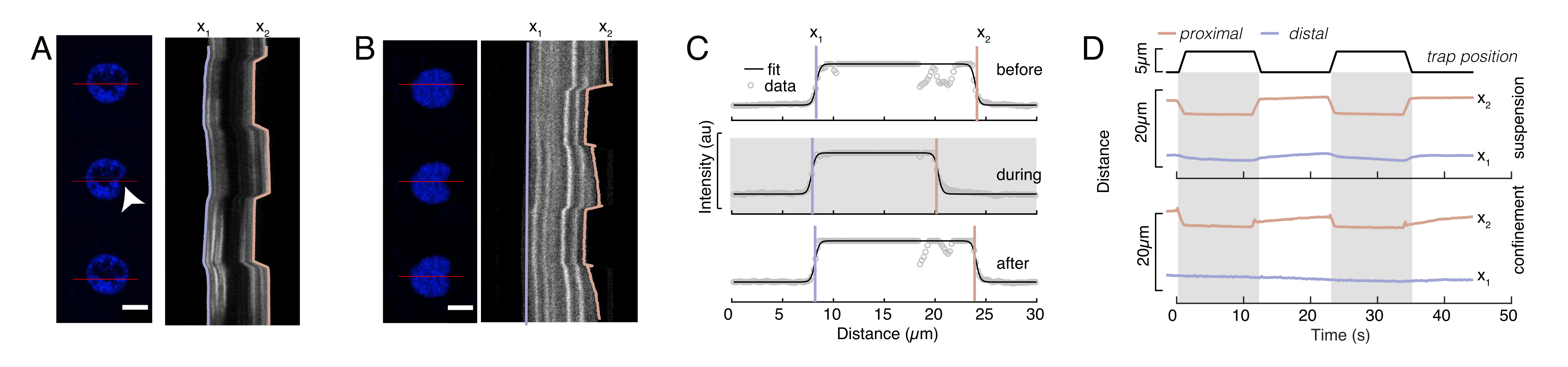{kind=link}

Рисунок 7: Силовая обработка сигнала. (А) Схема оптически захваченной микросферы, деформирующей ядро клетки при углублении. Ядерная мембрана и оптические силы обозначены черными стрелками. Изменение импульса луча обозначается зеленой стрелкой Pout. (B) Траектория ловушки (вверху) и сила (внизу), испытываемые оптически захваченной микросферой во время повторного эксперимента по ядерному углублению. (C) Силовой релаксационный распад после пика силы на максимальной глубине отступа. Вставка показывает схему стандартного линейного твердого тела, динамика которого приближается к феноменологическим наблюдениям здесь. (D) Слева: логарифм нормализованной силы против времени. Затененные области указывают на часть данных, используемую для соответствия двойному экспоненциальному распаду (красные линии). Справа: логарифм нормализованной силы против логарифма времени. Затененная область указывает на часть данных, используемую в соответствии с степенным законом. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 8: Процедура силового отступа с треугольными смещениями ловушек. (A) Репрезентативная траектория x1(t) в синем цвете и x2(t) в янтаре во время эксперимента по треугольному отступу, взятого на ячейке на высоте 10 мкм. Вверху: Положение ловушки. Середина: Анализ формы ядра. Расстояние между x1 и x2 указывает на диаметр ядра. Внизу: Сигнал силы. (B) Сила против положения ловушки для восьми последовательных отступов. (C) Эволюция диссипации, полученной из гистерезиса между приближением и частью изъятия кривой f-d, ядра для каждого последующего события отступа. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 9. Ядерные свойства клеток в суспензии (адгезивной поверхности) и удержания из трапециевидных процедур. (A) Проекционная площадь ядра из клеток, находящихся в суспензии и менее 10 мкм удержания. Черная полоса представляет медиану. (B) Ядерная жесткость камер в суспензии и под удержанием. Черная полоса представляет медиану. P-значения, полученные из теста Крускала-Уоллиса с использованием MatLab. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
Дополнительная таблица 1: Трапециевидная траектория, определенная программным обеспечением оптического пинцета. Первая (вторая) строка — это расстояние x (y), на котором ловушка будет линейно смещена. В третьей строке длительность заданного шага задается в секундах. Эта траектория состоит из семи точек и соответствует трапеции, нагруженной два раза против ядра на рисунке 7B. Пожалуйста, нажмите здесь, чтобы загрузить эту таблицу.
Дополнительная таблица 2: Треугольная траектория, определяемая программным обеспечением оптического пинцета. Аналогично таблице 2, эта траектория состоит из 16 точек, соответствующих восьми событиям отступа на глубине 5 мкм и скорости 2,5 мкм/с. Пожалуйста, нажмите здесь, чтобы загрузить эту таблицу.
Дополнительная таблица 3: Параметры подгонки для данных на рисунке 7. ИГ: первоначальная догадка. Пожалуйста, нажмите здесь, чтобы загрузить эту таблицу.
Дополнительный рисунок S1: Выравнивание оптического датчика силы и компенсация базовой линии импульса. (A) Полевой упор, изображенный на вспомогательной камере (AUX, рисунок 2) через объектив Бертрана. В погружном масле появляется воздушный пузырь, который не виден через окуляр. (B) Чистый оптический путь. Для точного выравнивания откройте полевую остановку и сделайте так, чтобы она совпадала с конусом NA = 1,2. (C) Изображение плоскости образца. Красный квадрат обозначает рабочую зону OT. Шкала шкалы: 20 мкм. (D) Мощность ловушки, измеренная по всему FOV, вдоль белых двойных стрелок, указанных в C. Красным цветом выделено изменение мощности ловушки, когда коррекция не применяется. Синим цветом сила ловушки скорректирована по всему полю зрения. (E) X-компонент исходного уровня импульса в том же диапазоне. Красным цветом, некорректированный след. Синим цветом трассировка скорректирована для мощности ловушки. Зеленым цветом трассировка скорректирована для базового уровня импульса с помощью глобальной компенсации смещения в программном обеспечении производителя. (F) То же самое, что и в E, для Y-компонента. Обратите внимание, что при нормальной работе затененные компоненты используются для механики и измерения силы, например, компонент силы x во время движения по координате x и компонент силы y во время движения по оси y. После того, как все поправки реализованы, получается rmSD-шум <0,5 пН. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Дополнительный рисунок S2: Неудачная процедура из-за слабых ловушек. (A) Кимограф, показывающий отступ ядра от неудачной процедуры. Видны только короткие, преходящие деформации из-за выхода шарика из ловушки. Важно отметить, что улавливающий лазер по-прежнему движется без бусины, чтобы завершить предопределенную траекторию (зеленая пунктирная линия). Шкала = 10 мкм. (B) Сверху: положение ловушки в зависимости от времени. Средний: результат отслеживания края вдавленного проксимального и дистального края ядра. Обратите внимание, что дистальный край не движется без отступа, как это обычно наблюдается при выполнении процедур на изолированных клетках на адгезивных подложках. Внизу: Сила против времени, показывающая потерю микросферы, о чем свидетельствует уменьшение теплового шума и внезапное падение до нулевой силы. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Дополнительный рисунок S3: Выживаемость введенных эмбрионов. Эмбрионы, которым вводили шарики 1 мкм и 100 пг/эмбрион мРНК в концентрациях, описанных в протоколе, сравнивали с неинъекционными эмбрионами и не показывали существенных различий через 24 ч после оплодотворения. Среднее и стандартное отклонение трех независимых экспериментов с N > 21 эмбриона на условие для каждого эксперимента. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Обсуждение
В этом протоколе мы описываем уникальный метод опроса механических свойств клеточного ядра внутри живых клеток. В отличие от других методов силовой спектроскопии, неинвазивное оптическое улавливание позволило отделить вклад клеточной мембраны и цитоскелета от жесткости клеточного ядра. Важно отметить, что оптическая микроманипуляция совместима с мультимодальной микроскопией, что позволит экспериментатору изучать различные процессы, участвующие в клеточной ядерной механобиологии. В качестве репрезентативного результата мы использовали окрашивание ДНК-Хёхста для измерения деформации ядра при вдавливании, выполненном силами порядка нескольких сотен пикоНьютонов.
Потенциальные применения нашего метода за пределами примеров, изложенных в этом протоколе
Возможность извлекать количественную механическую информацию из измерений внутри живых клеток без внешних возмущений обеспечивает множество беспрецедентных возможностей, которые только начинают изучаться. Таким образом, представленный протокол нашей оптической платформы микроманипуляции может быть распространен на более сложные эксперименты с большой универсальностью. Акустооптические дефлекторы (AOD) могут генерировать несколько оптических ловушек для синхронных измерений силы в различных местах расположения ячеек, а также могут использоваться для активной микрореологии в широком диапазоне частот51,61. Как уже упоминалось, силовой отклик при отступе может преодолеть максимальную силу захвата, что приводит к выходу шарика из оптической ловушки. В этом случае силовая обратная связь может быть сконфигурирована с помощью AOD для того, чтобы зажать оптическую силу. В целом, несколько микрореологических подходов, таких как релаксация стресса, описанная в этом протоколе, а также активная микрореология или соответствие ползучести, могут быть экспериментально получены с помощью этой платформы и тщательно проанализированы новыми пакетами программного обеспечения61,62,63,64,65 . Кроме того, применение сил не ограничивается ядром, но в принципе может быть осуществлено для измерения различных внутриклеточных структур и в сложных тканях, как показано для улавливания протекающих красных кровяных клеток внутри интактных кровеносных сосудов66,67 или захвата и деформирования хлоропластов и митохондрий68 . Калибровка светового импульса не зависит от формы и размера захваченного объекта, что позволяет проводить прямые измерения силы на любом силовом зонде произвольной формы38,39. Использование инъекционных микросфер позволило приложить к ядру высокие силы при относительно низкой мощности лазера по сравнению с прямым манипулированием клеточными структурами69,70,71. Однако, учитывая достаточно высокую разницу в показателях преломления, нет необходимости в внешнем силовом зонде, и внутриклеточными органеллами можно манипулировать непосредственно без инъекционных шариков (неопубликованные наблюдения и ссылка70).
Потенциальные модификации нашего метода для расширения приложений
Различные размеры микрошариков могут быть введены в зависимости от эксперимента, но относительный контроль должен быть выполнен. Например, для изучения клеток на более поздних стадиях могут быть введены более мелкие шарики. Это уменьшит максимальную силу, которая может быть приложена оптической ловушкой (как показано в ссылке 55). Большие бусины могут быть введены для оказания более высоких сил, но они могут повлиять на развитие эмбриона в зависимости от их размера или стадии интереса. В экспериментах, где инъекция микрогранул не является вариантом, различными органеллами, показывающими различия в показателях преломления по сравнению с цитоплазмой, все еще можно оптически манипулировать, что приводит к оптическим силам, измеряемым из изменений светового импульса42. Как упоминалось выше, эти методы были использованы Bambardekar et al. для деформирования клеточно-клеточных соединений в эмбрионе Drosophila70. Аналогичным образом, ядро клетки имеет более низкий показатель преломления, чем окружающая среда44, что позволяет проводить отступ без шариков (неопубликованные наблюдения и ссылка72), хотя и с более низкой силой захвата. Таким образом, ядро не может быть легко захвачено и выходит из ловушки.
Распорка PDMS со спиновым покрытием изготавливается удобным и быстрым методом, но может быть недоступна для лабораторий, не имеющих доступа к микро-/ нанопроизводственному оборудованию или инженерным лабораториям. Таким образом, распорка может быть легко собрана из лабораторной ленты или парапленки (шаг 4). Протокол также может быть адаптирован путем изготовления микрофлюидных каналов, которые автоматизируют доставку отдельных ячеек в предопределенные измерительные колодцы или в камеру с определенной высотой для оценки эффекта удержания в пределах одного и того же образца. Однако такие микрофлюидные устройства должны быть сконструированы таким образом, чтобы они соответствовали пространству между объективом микроскопа и собирающей линзой датчика оптической силы около 2 мм (см. шаг 3). Обратите внимание, что датчик оптической силы должен быть расположен на соответствующей высоте таким образом, чтобы оптические аберрации от расфокусировки не влияли на измерение импульса фотона.
Другие модификации могут включать смену биологических репортеров. Мы обнаружили, что флуоресценция Хёхста спектрально просачивается в канал GFP, и поэтому мы предпочитаем комбинацию с гистоном, помеченным mCherry, в качестве ядерного маркера для одновременного измерения в двух флуоресцентных каналах. В качестве альтернативы, ядерная деформация может быть легко отслежена с помощью метки, направленной на внутреннюю ядерную мембрану, такую как Lap2b-GFP (рисунок 2).
Углубление на ядро клетки составляло порядка 2-3 микрон, что мы могли точно измерить с помощью анализа изображений дифракционно-ограниченной спиннинг-дисковой конфокальной микроскопии. В случае более жестких ядер или меньших сил отступ будет едва ли измеримым при использовании этого подхода. Тем не менее, абсолютный силовой калиброванный оптический пинцет также может быть откалиброван для измерения положения захваченной бусины in situ с использованием интерферометрии BFP с нанометровой точностью51. Используя этот подход, сигнал напряжения и датчик оптической силы могут быть переведены в положение захваченного зонда через параметр β [нм / В], в то время как инвариантный параметр α [pN / V] дает значения силы через вышеупомянутую калибровку светового импульса41 (см. Ниже для деталей).
Устранение неполадок
Мы обнаружили, что во время эксперимента могут возникнуть следующие проблемы:
Не образуется стабильная ловушка, и микросфера легко выходит
Любая грязь на объективе микроскопа или смещенный коррекционный ошейник могут привести к выходу из строя стабильной ловушки. Если немедленное решение не найдено, измерьте функцию точечного распространения объектива. Если интересующий образец находится глубоко внутри оптически плотной ткани, лазерный фокус может испытывать серьезные оптические аберрации, приводящие к нестабильному захвату (этот эффект обычно незначителен в изолированных клетках, но становится более очевидным в более толстых тканях). Для высокой жесткости восстанавливающая сила ядра может превышать силу выхода ловушки, так что микросфера теряется, а процедура отступа выходит из строя. Первоначально край ядерной мембраны, близкий к оптической ловушке, почти не отступает (рисунок S2A). Когда это происходит, на улавливающий лазер больше не воздействуют сила и броуновское движение, что приводит к падению силы до нуля и уменьшению шума сигнала (рисунок S2B). В случае, если это произойдет, мощность лазера может быть увеличена, чтобы иметь более сильную ловушку, амплитуда трапециевидной траектории, толкающей шарик в ядро, может быть уменьшена, или исходное положение захваченной микрогранулы может быть установлено дальше от ядра.
Клетка движется во время стимуляции
Если клетки недостаточно прикреплены, оптическая градиентная ловушка будет перемещать клетки во время выполнения процедуры внутриклеточного отступа, так что силы и основная механика ядра являются артефактными. Чтобы предотвратить смещение всей клетки, мы рекомендуем увеличить концентрацию молекул клеточной адгезии на поверхности, например, ConA.
Компенсация начального импульса
Если в платформе КН отсутствует процедура компенсации начального импульса (этап 6.5), необходимо скорректировать искусственный, не зависящий от силы базовый сигнал. Это видно как наклон на кривой силы даже без захвата бусины (рисунок S1E). Чтобы сделать коррекцию, ту же траекторию нужно выполнить без шарика, вне ячейки в точно таком же положении. Для этого отодвиньте ячейку от ловушки с помощью элемента управления stage. В качестве ориентира, смещение силы изменяется на 5 пН по FOV при 200 мВт в нашей системе; таким образом, она становится незначительной для коротких траекторий. Альтернативно, этап пьезосканирования может быть использован для перемещения клеток на образце, оставляя положение лазера постоянным.
Критические шаги представленного протокола
Микросферы должны быть введены на правой, 1-клеточной стадии, чтобы обеспечить максимальное распределение по эмбриону. Бусины не должны быть флуоресцентными, чтобы свет не просачивался в флуоресцентные каналы, используемые для визуализации. Например, даже типичные красно-флуоресцентные шарики хорошо видны в синем канале, используемом для визуализации ядра клетки после окрашивания Хёхста, благодаря их яркости (возбуждение: 405 нм; излучение: 445 нм). Стабильное прикрепление клетки к субстрату имеет решающее значение для предотвращения бокового смещения во время процедуры отступа. Если клетка движется во время рутины, силы недооцениваются. Если это происходит часто, оптимизируйте протокол вложения. Для клеток тканевой культуры другие белки клеточной адгезии, такие как фибронектин, коллаген или поли-L-лизин, приводят к удовлетворительному прикреплению (неопубликованные наблюдения). Во время содержания клетки подвергаются внезапному и сильному механическому воздействию. Это может привести к повреждению клеток, и часто экспериментатор столкнется с лопнувшими клетками, если процедура не проводится осторожно. Кроме того, если высота удержания слишком мала, все клетки пострадают от разрыва ядерной оболочки или необратимого повреждения. Чтобы смягчить их, опустите верхнюю крышку медленнее и/или увеличьте расстояние между крышками.
Ограничения техники и предложения по их преодолению
Явным ограничением методики является проникновение лазерного света в глубокие участки ткани, что приводит к аберрациям и нестабильному улавливанию. Таким образом, нижний предел глубины проникновения зависит от четкости образца, коррекции аберрации, которая может быть использована73 , и применяемой мощности лазера. Следует учитывать, что более высокая мощность лазера приводит к тепловому возбуждению образца в непосредственной близости от микросферы. Тем не менее, нагрев образца, вызванного лазерным пятном длиной волны 1064 нм, сведен к минимуму, чтобы избежать правдоподобного теплового стресса на наших биологических образцах74.
Другим ограничением является максимальная сила, которую можно измерить. Несмотря на то, что прямое обнаружение светового импульса позволяет измерять силу далеко за пределами линейного режима отклика оптической ловушки40,41, максимальная приложенная сила составляет порядка нескольких сотен пиконьютонов. Это ограничено мощностью лазера и последующим порогом повреждения мягкого биологического материала и различиями показателей преломления, которые обычно не превышают 0,1 или 0,344. Было предложено несколько методов для увеличения предела обнаружения силы, например, с использованием структурированного света75, микросфер с антибликовым покрытием76, частиц с высоким показателем преломления77 или высоколегированных квантовых точек78.
ОТ могут использоваться для измерения положения в нанометровом масштабе с помощью интерферометрии BFP, так что положение шарика внутри ловушки составляет Δx = β Sx, где Sx - сигнал напряжения датчика, а β [мкм/В] может быть откалибровано на лету в соответствии с различными протоколами35,54. Для оптического датчика силы можно доказать, что инвариантный коэффициент преобразования напряжения в силу α [pN/V] напрямую связан с β и жесткостью ловушки, k [pN/μm], через α = kβ 37) В экспериментах со смещениями шариков, которые слишком малы, чтобы их можно было обнаружить с помощью оптической визуализации, эта стратегия может быть использована для дополнения измерений силы с обнаружением малого положения. Примером может служить применение экспериментальных процедур, представленных здесь, к очень жестким ядрам, для которых сил при разумной мощности лазера (200-500 мВт) недостаточно для того, чтобы вызвать достаточно большие значения отступа. В этом случае шарик должен быть приведен в контакт с ядром, а жесткость улавливания должна быть откалибрована до измерения (этап 8.6). Отступ d ядра как функция силы может быть косвенно определен как:
d = xtrap - F/k
где xtrap — позиция ловушки. В отличие от инвариантного коэффициента светового импульса α [pN/V], коэффициент β [мкм/В] необходимо калибровать перед каждым экспериментом, поскольку он зависит от многих локальных переменных, определяющих динамику улавливания, таких как размер частиц, размер пятна оптической ловушки и относительные показатели преломления.
Раскрытие информации
Авторам нечего раскрывать.
Благодарности
МК признает финансовую поддержку со стороны Министерства экономики и конкурентоспособности Испании в рамках программы Plan Nacional (PGC2018-097882-A-I00), FEDER (EQC2018-005048-P), Severo Ochoa для центров передового опыта в области исследований и разработок (CEX2019-000910-S; RYC-2016-21062), от Fundació Privada Cellex, Fundació Mir-Puig и от Generalitat de Catalunya через CERCA и исследовательскую программу (2017 SGR 1012), в дополнение к финансированию через ERC (MechanoSystems) и HFSP (CDA00023/2018). V.R. признает поддержку со стороны Министерства науки и инноваций Испании партнерству EMBL, Centro de Excelencia Severo Ochoa, Plan Nacional MINECO (BFU2017-86296-P, PID2020-117011GB-I00) и Женералитату Каталонии (CERCA). V.V. признает поддержку со стороны программы ICFOstepstone PhD, финансируемой исследовательской и инновационной программой Европейского Союза Horizon 2020 в рамках грантового соглашения Марии Склодовской-Кюри 665884. Мы благодарим Арнау Фарре за критическое прочтение рукописи; Марии Марсал за помощь в визуализации и монтаже эмбриона мощностью 24 л.с.л.с.; Сенда Хименес-Дельгадо за поддержку микроинъекций рыбок данио.
Материалы
| Name | Company | Catalog Number | Comments |
| #1.5 22 mm cover glasses | Ted Pella | 260148 | |
| #1.5 22x60 mm Coverglasses | Ted Pella | 260152 | |
| #1.5H glass bottom dishes | Willco | GWST-5040 | |
| 10-um beads | Supelco | 72986 | |
| 1-mm glass capillaries | Harvard Apparatus | 30-0020 GC100F-15 | |
| 1-um polystyrene microbeads | Sigma | 89904 | |
| 1-um red-fluorescent beads | ThermoFisher | F8816 | |
| Agar | ThermoFisher | 16500500 | |
| Aqcuisition cameras sCMOS | Andor | Sona-4BV11-UNI | |
| Auxiliary camera (Figure 3, AUX) | Blackfly, FLIR | BFS-U3-200S6M-C | |
| Calbryte 520 | AAT Bioquest | 520 AM | |
| Centrifuge | Eppendorf | 5453000011 | |
| Concanavalin A | Sigma | C5275 | |
| DMEM | Sigma | D2906 | |
| DNA-Hoechst | ThermoFisher | 33342 | |
| Double scotch tape | Biesse Adesivi | ||
| E3 | 5 mM NaCl. 0.17 mM KCl. 0.33 mM CaCl2. 0.33 mM MgSO4 | ||
| Eclipse Ti2 | Nikon | ||
| Forceps | Fine Science Tools | 11252-20 | |
| GPI-GFP | |||
| H2A-mCh | |||
| Image acquisition software | Fusion-Andor | ||
| Immersion Oil | Cargille | Type B: 16484 | |
| IR protection googles | Thorlabs | LG1 | |
| Lap2b-eGFP | |||
| Micro loader pipette | Eppendorf | GELoader | |
| Microinjector | World Precision Instruments | SYS-PV820 | |
| MicroManager 2.0 | |||
| Micromiter slide | ID5243 GXMGRAT-5 5mm/100 divisions | ||
| Mineral oil | Sigma | M3616 | |
| Motorized stage | ASI | ||
| Needle puller | Sutter instrument Co. | Model P-97 | |
| Optical tweezers platform | Impetux Optics | Sensocell | |
| OTs software (LightAce) | Impetux Optics | ||
| PDMS | Sigma | Sylgard 184 | |
| PDMS Curing agent | Sigma | Sylgard 184 | |
| Post processing software (Matlab) | Mathworks | ||
| RNAse free water | Thermofisher | AM9937 | |
| Short-pass dichroic mirror (Figure 3, IR-F) | Semrock | FF01-950/SP-25 | |
| Spin-coater | Specialty Coating Systems | Spincoat G3P-8 | |
| Spinning-disk confocal microscope | Andor | DragonFly 502 | |
| Stereomicroscope | Leica M80 | ||
| Triangular microinjection mold | Adaptive Science Tools | TU1 | |
| Universal oven | Memmert | UNB 200 | |
| Water immersion objective | Nikon | MRD07602 |
Ссылки
- Chan, C. J., Heisenberg, C. P., Hiiragi, T. Coordination of Morphogenesis and Cell-Fate Specification in Development. Current Biology. 27 (18), 1024-1035 (2017).
- Heller, E., Fuchs, E. Tissue patterning and cellular mechanics. Journal of Cell Biology. 211 (2), 219-231 (2015).
- Heisenberg, C. P., Bellaïche, Y. Forces in tissue morphogenesis and patterning. Cell. 153 (5), 948-962 (2013).
- Petridou, N. I., Spiró, Z., Heisenberg, C. P. Multiscale force sensing in development. Nature Cell Biology. 19 (6), 581-588 (2017).
- Krieg, M., et al. Tensile forces govern germ-layer organization in zebrafish. Nature Cell Biology. 10 (4), 429-436 (2008).
- Ruprecht, V., et al. Cortical contractility triggers a stochastic switch to fast amoeboid cell motility. Cell. 160 (4), 673-685 (2015).
- Shellard, A., Mayor, R. Supracellular migration - Beyond collective cell migration. Journal of Cell Science. 132 (8), (2019).
- Mongera, A., et al. A fluid-to-solid jamming transition underlies vertebrate body axis elongation. Nature. 561 (7723), 401-405 (2018).
- Atia, L., et al. Geometric constraints during epithelial jamming. Nature Physics. 14 (6), 613-620 (2018).
- Turlier, H., Maître, J. -. L. Mechanics of tissue compaction. Seminars in Cell & Developmental Biology. 47-48, 110-117 (2015).
- Ladoux, B., Mège, R. M. Mechanobiology of collective cell behaviours. Nature Reviews Molecular Cell Biology. 18 (12), 743-757 (2017).
- Venturini, V., et al. The nucleus measures shape changes for cellular proprioception to control dynamic cell behavior. Science. 370 (6514), (2020).
- Charras, G., Sahai, E. Physical influences of the extracellular environment on cell migration. Nature Reviews Molecular Cell Biology. 15 (12), 813-824 (2014).
- Kirby, T. J., Lammerding, J. Emerging views of the nucleus as a cellular mechanosensor. Nature Cell Biology. 20 (4), 373-381 (2018).
- Lee, H. P., et al. The nuclear piston activates mechanosensitive ion channels to generate cell migration paths in confining microenvironments. Science Advances. 7 (2), (2021).
- Friedl, P., Wolf, K., Lammerding, J. Nuclear mechanics during cell migration. Current Opinion in Cell Biology. 23 (1), 55-64 (2011).
- Versaevel, M., Riaz, M., Grevesse, T., Gabriele, S. Cell confinement: Putting the squeeze on the nucleus. Soft Matter. 9 (29), 6665-6676 (2013).
- Zuela-Sopilniak, N., et al. Measuring nucleus mechanics within a living multicellular organism: Physical decoupling and attenuated recovery rate are physiological protective mechanisms of the cell nucleus under high mechanical load. Molecular Biology of the Cell. 31 (17), 1943-1950 (2020).
- Kim, D. H., Wirtz, D. Cytoskeletal tension induces the polarized architecture of the nucleus. Biomaterials. 48, 161-172 (2015).
- Lomakin, A. J., et al. The nucleus acts as a ruler tailoring cell responses to spatial constraints. Science. 370 (6514), (2020).
- Hampoelz, B., et al. Microtubule-induced nuclear envelope fluctuations control chromatin dynamics in Drosophila embryos. Development. 138 (16), 3377-3386 (2011).
- Heo, S. J., et al. Differentiation alters stem cell nuclear architecture, mechanics, and mechano-sensitivity. eLife. 5, 1-21 (2016).
- Cosgrove, B. D., et al. Nuclear envelope wrinkling predicts mesenchymal progenitor cell mechano-response in 2D and 3D microenvironments. Biomaterials. 270, 120662 (2021).
- Liu, H., et al. In situ mechanical characterization of the cell nucleus by atomic force microscopy. ACS Nano. 8 (4), 3821-3828 (2014).
- Hobson, C. M., et al. Correlating nuclear morphology and external force with combined atomic force microscopy and light sheet imaging separates roles of chromatin and lamin A/C in nuclear mechanics. Molecular Biology of the Cell. 31 (16), 1788-1801 (2020).
- Pajerowski, J. D., Dahl, K. N., Zhong, F. L., Sammak, P. J., Discher, D. E. Physical plasticity of the nucleus in stem cell differentiation. Proceedings of the National Academy of Sciences of the United States of America. 104 (40), 15619-15624 (2007).
- Rowat, A. C., Lammerding, J., Ipsen, J. H. Mechanical properties of the cell nucleus and the effect of emerin deficiency. Biophysical Journal. 91 (12), 4649-4664 (2006).
- Davidson, P. M., et al. High-throughput microfluidic micropipette aspiration device to probe time-scale dependent nuclear mechanics in intact cells. Lab on a Chip. 19 (21), 3652-3663 (2019).
- Lombardi, M., Zwerger, M., Lammerding, J. Biophysical assays to probe the mechanical properties of the interphase cell nucleus: Substrate strain application and microneedle manipulation. Journal of Visualized Experiments: JoVE. (55), (2011).
- Luo, T., Mohan, K., Iglesias, P. A., Robinson, D. N. Molecular mechanisms of cellular mechanosensing. Nature Materials. 12 (11), 1064-1071 (2013).
- Dahl, K. N., Engler, A. J., Pajerowski, J. D., Discher, D. E. Power-law rheology of isolated nuclei with deformation mapping of nuclear substructures. Biophysical Journal. 89 (4), 2855-2864 (2005).
- Guilluy, C., et al. Isolated nuclei adapt to force and reveal a mechanotransduction pathway in the nucleus. Nature Cell Biology. 16 (4), 376-381 (2014).
- Bustamante, C. J., Wang, M. D. Optical tweezers in single-molecule biophysics. Nature Reviews Methods Primers. , 1-29 (2021).
- Svoboda, K., Block, S. M. Force and velocity measured for single kinesin molecules. Cell. 77 (5), 773-784 (1994).
- Berg-Sørensen, K., Flyvbjerg, H. Power spectrum analysis for optical tweezers. Review of Scientific Instruments. 75 (3), 594-612 (2004).
- Smith, S. B., Cui, Y., Bustamante, C. Optical-trap force transducer that operates by direct measurement of light momentum. Methods in Enzymology. 361 (1994), 134-162 (2003).
- Farré, A., Montes-Usategui, M. A force detection technique for single-beam optical traps based on direct measurement of light momentum changes. Optics Express. 18 (11), 11955 (2010).
- Català, F., Marsà, F., Montes-Usategui, M., Farré, A., Martín-Badosa, E. Extending calibration-free force measurements to optically-trapped rod-shaped samples. Scientific Reports. 7, 1-10 (2017).
- Bui, A. A. M., et al. Calibration of force detection for arbitrarily shaped particles in optical tweezers. Scientific Reports. 8 (1), 1-12 (2018).
- Farré, A., Marsà, F., Montes-Usategui, M. Beyond the hookean spring model: Direct measurement of optical forces through light momentum changes. Methods in Molecular Biology. 1486, (2017).
- Farré, A., Marsà, F., Montes-Usategui, M. Optimized back-focal-plane interferometry directly measures forces of optically trapped particles. Optics Express. 20 (11), 12270 (2012).
- Jun, Y., Tripathy, S. K., Narayanareddy, B. R. J., Mattson-Hoss, M. K., Gross, S. P. Calibration of optical tweezers for in vivo force measurements: How do different approaches compare. Biophysical Journal. 107 (6), 1474-1484 (2014).
- Mas, J., Farré, A., Sancho-Parramon, J., Martín-Badosa, E., Montes-Usategui, M. Force measurements with optical tweezers inside living cells. Optical Trapping and Optical Micromanipulation XI. 9164, (2014).
- Schürmann, M., Scholze, J., Müller, P., Guck, J., Chan, C. J. Cell nuclei have lower refractive index and mass density than cytoplasm. Journal of Biophotonics. 9 (10), 1068-1076 (2016).
- Rosen, J. N., Sweeney, M. F., Mably, J. D. Microinjection of zebrafish embryos to analyze gene function. Journal of Visualized Experiments: JoVE. (25), (2009).
- Westerfield, M. . The Zebrafish Book. A Guide for the Laboratory Use of Zebrafish. (Danio rerio), 5th Edition. , (2007).
- Schubert, R., et al. Assay for characterizing the recovery of vertebrate cells for adhesion measurements by single-cell force spectroscopy. FEBS Letters. 588 (19), 3639-3648 (2014).
- Koschwanez, J. H., Carlson, R. H., Meldrum, D. R. Thin PDMS films using long spin times or tert-butyl alcohol as a solvent. PLoS One. 4 (2), 2-6 (2009).
- Das, R., et al. Mechanical stretch inhibition sensitizes proprioceptors to compressive stresses. bioRxiv. , (2021).
- Chardès, C., Clement, R., Blanc, O., Lenne, P. F. Probing cell mechanics with bead-free optical tweezers in the drosophila embryo. Journal of Visualized Experiments: JoVE. (141), (2018).
- Staunton, J. R., Blehm, B., Devine, A., Tanner, K. In situ calibration of position detection in an optical trap for active microrheology in viscous materials. Optics Express. 25 (3), 1746 (2017).
- Bola, R., Treptow, D., Marzoa, A., Montes-Usategui, M., Martin-Badosa, E. Acousto-holographic optical tweezers. Optics Letters. 45 (10), 2938-2941 (2020).
- Thalhammer, G., Obmascher, L., Ritsch-Marte, M. Direct measurement of axial optical forces. Optics Express. 23 (5), 6112 (2015).
- Vermeulen, K. C., et al. Calibrating bead displacements in optical tweezers using acousto-optic deflectors. Review of Scientific Instruments. 77 (1), 1-6 (2006).
- Dzementsei, A., Barooji, Y. F., Ober, E. A., Oddershede, L. B. Foregut organ progenitors and their niche display distinct viscoelastic properties in vivo during early morphogenesis stages. bioRxiv. , 1-35 (2021).
- Behrndt, M., et al. Forces driving epithelial spreading in zebrafish gastrulation. Science. 338 (6104), 257-260 (2012).
- Krieg, M., et al. Atomic force microscopy-based mechanobiology. Nature Reviews Physics. , (2018).
- A-Hassan, E., et al. Relative microelastic mapping of living cells by atomic force microscopy. Biophysical Journal. 74 (3), 1564-1578 (1998).
- Khalilgharibi, N., et al. Stress relaxation in epithelial monolayers is controlled by the actomyosin cortex. Nature Physics. 15, (2019).
- Crick, S. L., Yin, F. C. Assessing micromechanical properties of cells with atomic force microscopy: importance of the contact point. Biomechanics and Modeling in Mechanobiology. 6 (3), 199-210 (2007).
- Hurst, S., Vos, B. E., Betz, T. Intracellular softening and fluidification reveals a mechanical switch of cytoskeletal material contributions during division. bioRxiv. , 425761 (2021).
- Kaplan, J. L., Bonfanti, A., Kabla, A. RHEOS.jl - A Julia package for rheology data analysis. arXiv. 4, 1-5 (2020).
- Bonfanti, A., Kaplan, J. L., Charras, G., Kabla, A. Fractional viscoelastic models for power-law materials. Soft Matter. 16 (26), 6002-6020 (2020).
- Rivas-Barbosa, R., Escobedo-Sánchez, M. A., Tassieri, M., Laurati, M. i-Rheo: determining the linear viscoelastic moduli of colloidal dispersions from step-stress measurements. Physical Chemistry Chemical Physics: PCCP. 22 (7), 3839-3848 (2020).
- Tassieri, M., et al. i-Rheo: Measuring the materials' linear viscoelastic properties "in a step". Journal of Rheology. 60 (4), 649-660 (2016).
- Zhong, M. C., Wei, X. B., Zhou, J. H., Wang, Z. Q., Li, Y. M. Trapping red blood cells in living animals using optical tweezers. Nature Communications. 4, 1767-1768 (2013).
- Harlepp, S., Thalmann, F., Follain, G., Goetz, J. G. Hemodynamic forces can be accurately measured in vivo with optical tweezers. Molecular Biology of the Cell. 28 (23), 3252-3260 (2017).
- Bayoudh, S., Mehta, M., Rubinsztein-Dunlop, H., Heckenberg, N. R., Critchley, C. Micromanipulation of chloroplasts using optical tweezers. Journal of Microscopy. 203 (2), 214-222 (2001).
- Favre-Bulle, I. A., Stilgoe, A. B., Rubinsztein-Dunlop, H., Scott, E. K. Optical trapping of otoliths drives vestibular behaviours in larval zebrafish. Nature Communications. 8 (1), 630 (2017).
- Bambardekar, K., Clément, R., Blanc, O., Chardès, C., Lenne, P. F. Direct laser manipulation reveals the mechanics of cell contacts in vivo. Proceedings of the National Academy of Sciences of the United States of America. 112 (5), 1416-1421 (2015).
- Ferro, V., Chuai, M., McGloin, D., Weijer, C. J. Measurement of junctional tension in epithelial cells at the onset of primitive streak formation in the chick embryo via non-destructive optical manipulation. Development (Cambridge). 147 (3), (2020).
- Hörner, F., et al. Holographic optical tweezers-based in vivo manipulations in zebrafish embryos. Journal of Biophotonics. 10 (11), 1492-1501 (2017).
- Zhong, M. -. C., Wang, Z. -. Q., Li, Y. -. M. Aberration compensation for optical trapping of cells within living mice. Applied Optics. 56 (7), 1972 (2017).
- Català, F., Marsà, F., Montes-Usategui, M., Farré, A., Martín-Badosa, E. Influence of experimental parameters on the laser heating of an optical trap. Scientific Reports. 7 (1), 1-9 (2017).
- Taylor, M. A., Waleed, M., Stilgoe, A. B., Rubinsztein-Dunlop, H., Bowen, W. P. Enhanced optical trapping via structured scattering. Nature Photonics. 9 (10), 669-673 (2015).
- Bormuth, V., et al. Optical trapping of coated microspheres. Optics Express. 16 (18), 13831-13844 (2008).
- Sudhakar, S., et al. Germanium nanospheres for ultraresolution picotensiometry of kinesin motors. Science. 371 (6530), (2021).
- Shan, X., et al. Optical tweezers beyond refractive index mismatch using highly doped upconversion nanoparticles. Nature Nanotechnology. 16 (5), 531-537 (2021).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены