
Method Article
Mediciones directas de la fuerza de la mecánica subcelular en confinamiento utilizando pinzas ópticas
En este artículo
Resumen
Aquí, presentamos un protocolo para investigar las propiedades mecánicas intracelulares de células embrionarias aisladas de pez cebra en confinamiento tridimensional con medición directa de la fuerza mediante una trampa óptica.
Resumen
Durante el desarrollo de un organismo multicelular, una sola célula fertilizada se divide y da lugar a múltiples tejidos con diversas funciones. La morfogénesis tisular va de la mano con cambios moleculares y estructurales a nivel de una sola célula que resultan en variaciones de las propiedades mecánicas subcelulares. Como consecuencia, incluso dentro de la misma célula, diferentes orgánulos y compartimentos resisten de manera diferente a las tensiones mecánicas; y las vías de mecanotransducción pueden regular activamente sus propiedades mecánicas. La capacidad de una célula para adaptarse al microambiente del nicho de tejido se debe en parte a la capacidad de detectar y responder a las tensiones mecánicas. Recientemente propusimos un nuevo paradigma de mecanosensición en el que la deformación y el posicionamiento nuclear permiten a una célula medir el entorno físico 3D y dotan a la célula de un sentido de propiocepción para decodificar los cambios en la forma de la célula. En este artículo, describimos un nuevo método para medir las fuerzas y propiedades del material que dan forma al núcleo celular dentro de las células vivas, ejemplificado en células adherentes y células confinadas mecánicamente. Las mediciones se pueden realizar de forma no invasiva con trampas ópticas dentro de las células, y las fuerzas son directamente accesibles a través de la detección sin calibración del momento de la luz. Esto permite medir la mecánica del núcleo independientemente de las deformaciones de la superficie celular y permitir la disección de las vías de mecanotransducción exteroceptivas e interoceptivas. Es importante destacar que el experimento de captura se puede combinar con la microscopía óptica para investigar la respuesta celular y la dinámica subcelular utilizando imágenes de fluorescencia del citoesqueleto, iones de calcio o morfología nuclear. El método presentado es fácil de aplicar, compatible con soluciones comerciales para mediciones de fuerza, y se puede ampliar fácilmente para investigar la mecánica de otros compartimentos subcelulares, por ejemplo, mitocondrias, fibras de estrés y endosomas.
Introducción
La morfogénesis tisular es un proceso complejo en el que las señales bioquímicas y las fuerzas físicas se coordinan espaciotemporalmente. En el embrión en desarrollo, los gradientes de factores de señalización bioquímica dictan la especificación del destino y aseguran el correcto patrón tisular1,2. Al mismo tiempo, las fuerzas intrínsecas y extrínsecas juegan un papel en la construcción de la arquitectura del embrión3,4. La influencia de la mecánica de la corteza celular en este contexto ha sido ampliamente estudiada5,6. La estrecha interconexión entre los procesos mecanoquímicos durante la morfogénesis se basa en las propiedades de las células individuales para detectar y responder a las fuerzas mecánicas en su microambiente tisular. Las células, por lo tanto, decodifican señales mecánicas a través de la presencia de elementos subcelulares y moleculares sensibles a la fuerza que transducen información mecánica en vías de señalización específicas que controlan el comportamiento celular, el destino celular y la mecánica celular.
Un sello distintivo de los procesos de desarrollo es que las células se organizan como grupos para construir estructuras multicelulares. Como tal, las células individuales rara vez se reorganizan y se mueven solas, sino que se asocian en un sociotopo apretado en el que muestran un comportamiento colectivo como la migración supracelular7, las transiciones (des)atasco8,9 o la compactación de blastocistos10. Las fuerzas mecánicas generadas dentro y entre las células sirven como señales importantes para instruir la dinámica celular colectiva7,11. Pero incluso cuando las células se mueven solas, como las células progenitoras que se abren paso entre las láminas de tejido o los nichos de tejido estrechos, experimentan extensas fuerzas mecánicas anisotrópicas cuando navegan por un entorno tridimensional. Estas tensiones mecánicas sobre las células tienen profundas consecuencias en el comportamiento celular12,13. Se han investigado varios mecanismos que convergen en el núcleo como elemento principal de mecanotransducción14,15, como elemento mecánico pasivo o activo durante la migración dentro de un entorno tisular 3D denso15,16.
Recientemente propusimos un mecanismo que equipa a las células para medir las deformaciones de forma utilizando el núcleo como un mecano-gauge intracelular elástico12. El núcleo, al ser el orgánulo más grande de una célula, sufre grandes deformaciones cuando las células se polarizan, migran o cambian su forma bajo estiramiento mecánico, confinamiento o estrés osmótico16,17,18,19. Encontramos que el estiramiento de la envoltura nuclear junto con el posicionamiento intracelular del núcleo proporciona a las células información sobre la magnitud y el tipo de deformación celular (como la compresión celular frente a la hinchazón celular). El estiramiento del núcleo se asocia con un despliegue de la membrana nuclear interna (INM), que promueve la actividad de la lipasa cPLA2 (fosfolipasa citosólica A2) dependiente del calcio en el INM, seguida de la liberación de ácido araquidónico (AA) y la activación rápida de la miosina II en la corteza celular. Esto conduce a un aumento de la contractilidad celular y a la migración de células ameboides por encima de un umbral de contractilidad cortical6. La respuesta mecanosensible a la deformación celular ocurre en menos de un minuto y es reversible tras la liberación por confinamiento, lo que sugiere que el núcleo actúa como un galga extensométrica para la propiocepción celular que regula el comportamiento celular adaptativo en condiciones de estrés mecánico. Esta vía mecanosensible ha demostrado ser activa en células madre progenitoras derivadas de embriones de pez cebra, tanto en células pluripotentes como comprometidas con el linaje12 y se conserva en diferentes especies y líneas celulares20.
Además de las propiedades nucleares como mecanosensor celular, la arquitectura y la mecánica nucleares están intrínsecamente reguladas durante el desarrollo y en respuesta a la especificación del destino celular21, por lo tanto, afinando la mecanosensibilidad celular22,23. La consecuencia podría ser un cambio en el cumplimiento nuclear que permita cambios morfológicos y transiciones de un estado premigratorio a un estado migratorio y viceversa8.
Se han aplicado diversas técnicas para medir la mecánica del núcleo celular, como la microscopía de fuerza atómica24,25, la aspiración de micropipetas26,27, la tecnología microfluídica28 y las microagujas29. Sin embargo, muchas de estas técnicas son invasivas en el sentido de que toda la célula debe deformarse, lo que limita la medición de las características mecánicas y las respuestas dependientes de la fuerza del propio núcleo. Para eludir la deformación simultánea de la superficie celular y su corteza celular mecanosensible30, se estudiaron núcleos aislados en diversos contextos31,32. Sin embargo, no se puede descartar que el aislamiento nuclear esté asociado a un cambio en las propiedades mecánicas del núcleo y su regulación (referencia24 y observaciones propias inéditas).
Las pinzas ópticas (OT) son una tecnología versátil que ha permitido una gran cantidad de experimentos en mecanobiología celular y han sido fundamentales en nuestra comprensión de cómo las máquinas moleculares convierten la energía química en energía mecánica33,34. Las pinzas ópticas utilizan un rayo láser estrechamente enfocado para ejercer fuerzas ópticas sobre partículas dieléctricas que tienen un índice de refracción más alto que el medio circundante33. Tales fuerzas pueden ser del orden de cientos de pico-Newtons y dar como resultado un confinamiento efectivo de la partícula dentro del foco de la trampa láser, lo que permite la manipulación de la partícula atrapada en tres dimensiones. El uso de la luz tiene una ventaja importante en que la medición se puede realizar de forma no invasiva dentro de las células vivas. Las manipulaciones ópticas se limitan aún más al foco de trampa del rayo láser. Por lo tanto, la manipulación se puede realizar sin estimular las membranas celulares circundantes y no perturba la corteza de actina o los procesos mecanosensibles en la membrana plasmática, como la activación dependiente de la fuerza de los canales iónicos.
La dificultad del enfoque de pinza óptica es determinar con precisión las fuerzas aplicadas a la microesfera utilizando enfoques clásicos que se basan en la calibración de fuerza indirecta basada en el teorema de equipartición o el uso de fuerzas de arrastre de Stokes definidas para medir una fuerza de escape dependiente de la potencia del láser35. Si bien estos métodos son fáciles de implementar en un experimento in vitro, generalmente no se pueden traducir a un entorno celular. Se han introducido varias estrategias en el campo que se basan en una calibración de fuerza directa, derivada de los primeros principios de conservación del momento36,37. A diferencia de otros enfoques de espectroscopia de fuerza, las mediciones de fuerza se deducen de un intercambio local de momento de luz con la partícula atrapada de forma arbitraria38,39. En nuestra configuración experimental, los cambios en el momento de la luz derivados de las fuerzas ópticas se miden directamente sin necesidad de calibración de trampa in situ40,41,42,43. Por lo tanto, las mediciones se vuelven posibles en un entorno viscoso como el interior de la célula o incluso dentro de un tejido, y las fuerzas se pueden cuantificar fácilmente hasta el nivel de pN.
En este protocolo, describimos un ensayo para manipular mecánicamente orgánulos o estructuras intracelulares y evaluar cuantitativamente sus propiedades mecánicas mediante una configuración de pinza óptica. Esta configuración está integrada en un microscopio fluorescente de disco giratorio que permite obtener imágenes paralelas del comportamiento celular o la dinámica intracelular. El ensayo permite la caracterización de las propiedades mecánicas de compartimentos celulares específicos, como el núcleo, al tiempo que estudia la posible mecanorespuesta y la activación de las vías de señalización molecular como resultado de la propia deformación. Además, el atrapamiento óptico de las microperlas inyectadas dentro de las células permite un aumento de la fuerza de indentación gracias a un índice de refracción considerablemente mayor de la cuenta de poliestireno (n = 1,59) en comparación con el contraste refractivo intrínseco44 del núcleo (n ~ 1,35) frente al citoplasma (n ~ 1,38). La estrategia presentada se puede adaptar fácilmente al estudio de otras estructuras intracelulares y orgánulos, así como a otros enfoques que involucran microrreología activa, el uso de múltiples trampas ópticas para sondear las mismas / diferentes estructuras subcelulares simultáneamente, y mediciones dirigidas a la mecanobiología celular en el embrión vivo.
Protocolo
Todos los protocolos utilizados han sido aprobados por el Comité Institucional de Ética de Cuidado y Uso de Animales (PRBB-IACUEC) e implementados de acuerdo con la normativa nacional y europea. Todos los experimentos se llevaron a cabo de acuerdo con los principios de las 3R. El pez cebra (Danio rerio) se mantuvo como se describió anteriormente.
1. Preparación de células madre progenitoras embrionarias primarias aisladas de pez cebra
- Preparación de micropipeta y agarosa
NOTA: Para un protocolo completo de microinyección de embriones de pez cebra, consulte la referencia45.- Con un extractor de micropipetas, tire de un capilar de vidrio de 1,0 mm para obtener dos agujas45. Guarde las agujas no utilizadas en una placa de Petri de 150 mm unida a un cojín de plastilina o en un anillo de cinta de laboratorio de adentro hacia afuera para proteger la punta delgada de daños durante el transporte.
- Derretir al 1% de agarosa ultrapura en E3 (5 mM NaCl, 0.17 mM KCl, 0.33 mM CaCl2, 0.33 mM MgSO4) en un microondas estándar de cocina/laboratorio durante 10 s. Calentar la mezcla repetidamente durante cortos períodos de tiempo (pocos segundos) hasta que la agarosa se derrita.
- Cuando la agarosa esté completamente derretida, deje que se enfríe brevemente y luego viértala en una placa de Petri de 10 cm. Añadir lentamente el molde de microinyección triangular (ver Tabla de Materiales) en la parte superior de la agarosa evitando la aparición de burbujas. No empuje el molde, asegurándose de que permanezca en la superficie de la agarosa.
- Cuando la agarosa se solidifique por completo, retire el molde triangular muy lentamente ejerciendo una fuerza suave para evitar cualquier rotura en la agarosa. La placa se puede almacenar boca abajo a 4 °C durante 2-4 semanas.
- 30 min antes de la microinyección, saca la placa de la nevera y añade el E3 precalentado a 28 °C para que se estabilice a temperatura ambiente.
- Preparación de la mezcla inyectable
- Para preparar la mezcla de inyección, diluya 1 μm de microperlas (poliestireno, no fluorescente) en proporción 1:5 en agua libre de RNasa.
- Preparar el ARNm para la expresión transitoria de marcadores fluorescentes o la expresión de construcciones de genes recombinantes y/o la coinyección de morfolino a la concentración deseada.
NOTA: Una mezcla de inyección típica para la coinyección de microperlas junto con 100 pg de ARNm por embrión para etiquetar, por ejemplo, el núcleo con H2A-mCherry es: 1 μL de perlas + 1 μL de ARNm (la concentración de stock es de 1 μg / μL) + 2.5 μL de agua libre de ARN + 0.5 μL de rojo fenol (solución madre 0.5%, rojo fenol no es obligatorio; se utiliza para una mejor visualización de la gota inyectada pero la inyección no etiquetada la gota también es visible para un experimentador experimentado). La inyección de ARN también puede ser útil para seleccionar embriones inyectados. Se pueden inyectar microperlas fluorescentes, en lugar de no fluorescentes, para visualizarlas.
- Carga y calibración de agujas de microinyección
- Encienda el microinyector con la opción Time-Gated . Esta configuración es muy importante para calibrar el volumen de inyección correctamente. Establezca el tiempo de cierre en aproximadamente 500 ms.
- Cargue 3 μL de la mezcla de inyección en la aguja utilizando una pipeta microcargadora.
- Inserte la aguja en el micromanipulador y selle herméticamente. Compruebe si el micromanipulador está en una buena posición y tiene suficiente libertad para moverse en dirección x-y en la placa de inyección.
- Mida el tamaño de la gota utilizando una corredera de micrómetro (5 mm/100 divisiones) con una gota de aceite mineral en la parte superior45 y expulsando una gota de la mezcla de inyección directamente en el aceite mineral.
- Recorte la aguja con pinzas afiladas en un ángulo pronunciado para generar una punta puntiaguda afilada. Ajuste el tamaño de la gota a 0,1 mm, correspondiente a 0,5 nL de material inyectado.
NOTA: Si al cortar la aguja, se excede este volumen, se recomienda rehacer el procedimiento de calibración con una aguja nueva. El tiempo de cierre del microinyector se puede ajustar ligeramente para que coincida con el volumen de caída; sin embargo, los tiempos de cierre cortos corresponden a un gran diámetro de aguja, lo que potencialmente daña los embriones.
- Microinyección de embriones de pez cebra en etapa unicelular
- Recolectar embriones de pez cebra poco después de la fertilización para la microinyección de la mezcla de cuentas directamente en el embrión en etapa de una célula (cigoto) antes de que ocurra la primera división celular.
NOTA: Esto asegura una distribución adecuada de las microesferas y un rendimiento lo suficientemente alto de blastómeros aislados con al menos una microesfera por célula en las etapas de desarrollo posteriores en las que se realizan experimentos (etapa de blástula-gastrula). Los experimentos de indentación aún se pueden realizar si hay dos esferas dentro de la célula, pero las células que no tienen perlas deben excluirse (aunque la sangría sin esferas es posible). En este protocolo se utilizaron cepas ab wildtype, pero se puede usar cualquier otra cepa, por ejemplo, TL. - Coloque los embriones en etapa de una célula (cigoto) en un molde de agarosa al 1% de forma triangular precalentada, como se muestra en la Figura 1A, utilizando una pipeta pasteur de plástico.
- Retire el medio adicional con la misma pipeta para evitar que los embriones floten alrededor. Empuje suavemente los embriones en el molde triangular a través de un cepillo. Mantenga un poco de espacio entre los embriones para facilitar la orientación correcta (Figura 1B).
- Alinee suavemente los embriones con un cepillo para que los embriones estén orientados lateralmente, con la única célula del cigoto siendo claramente visible, como se muestra en la Figura 1B. Una orientación ideal para la microinyección se alcanza cuando la célula del embrión está mirando hacia la dirección de la aguja (inyección a través del polo animal del embrión) o de la manera opuesta frente a la célula de la yema (inyección a través del polo vegetal del embrión), como se muestra en la Figura 1C.
- Sostenga el plato con una mano y use la otra mano para colocar la punta de la aguja con el controlador del micromanipulador. Baje la punta de la aguja hacia los embriones.
- Perfore el corion e ingrese el embrión de una célula con la aguja mientras monitorea el procedimiento a través del estereomicroscopio. Asegúrese de la colocación correcta de la aguja y, después de la inyección, la ubicación correcta de la gota inyectada como se muestra en la Figura 1C.
- Repita para todos los embriones: mueva la aguja hacia arriba, deslice el plato con los embriones hasta que el siguiente embrión esté centrado, baje la aguja e inyéctela.
- Una vez que se inyecta todo el conjunto de embriones, retire los embriones del molde de agarosa / placa de Petri enjuagando un poco de E3 y colóquelos en una nueva placa de Petri con una pipeta Pasteur de plástico. Se recomienda colocar suficientes medios en la placa de inyección para evitar el secado de los embriones durante el procedimiento de microinyección.
- Repita el procedimiento hasta que se inyecte el número deseado de embriones. Los embriones deben estar en una etapa celular para garantizar la propagación máxima y homogénea de las perlas.
NOTA: Este procedimiento está optimizado para embriones de blástula temprana y es probable que deba optimizarse si se van a investigar diferentes etapas de desarrollo. - Coloque los embriones inyectados dentro de una incubadora a 28-31 °C durante aproximadamente 4 h o hasta la etapa deseada (Figura 1D) antes de continuar con el protocolo para el cultivo celular primario.
NOTA: Opcionalmente, deje que los embriones se desarrollen más allá de la etapa de blástula (o el punto de tiempo de medición deseado) para garantizar la supervivencia y descartar artefactos de toxicidad. En estadios larvarios, monta larvas anestesiadas con tricaína en agarosa al 0,75% e imagina la distribución de microesferas en diversos tejidos. Para hacer una solución madre, mezcle: 400 mg de polvo de tricaína en 97.9 mL de agua destilada, aproximadamente 2.1 mL de 1 M de base TRIS (pH 9), y ajuste a pH 7. Esta solución se puede almacenar a 4 °C. Para usar tricaína como anestésico, diluya 4.2 ml de solución madre en 100 ml del medio del huevo (o medio deseado); en este caso, se utilizó E3. Consulte la referencia46 para obtener más detalles.
- Recolectar embriones de pez cebra poco después de la fertilización para la microinyección de la mezcla de cuentas directamente en el embrión en etapa de una célula (cigoto) antes de que ocurra la primera división celular.
2. Preparación y tinción de una sola célula
- Coloque los embriones de la etapa de esfera (4 hpf, horas después de la fertilización) en un plato de vidrio utilizando una pipeta Pasteur de plástico. Seleccionar los embriones que son positivos para la señal de las perlas inyectadas, y que expresan la proteína fluorescente en caso de inyección de ARNm. Algunos embriones pueden mostrar una alta agrupación de cuentas y pueden ser excluidos.
- Descoronar manualmente los embriones usando fórceps. Transfiera aproximadamente 10-15 embriones a recipientes de reacción de 1,5 ml utilizando una pipeta Pasteur de vidrio.
NOTA: Cuando los embriones son descoronados, se adhieren al plástico y se requiere el uso de cristalería. Como alternativa a la placa de vidrio, se puede utilizar una placa de Petri de plástico con una capa delgada de agarosa al 1%. Se debe preferir la descoronación manual al tratamiento con pronasa enzimática para prevenir el daño proteolítico a las proteínas de la superficie celular y los posibles cambios en las propiedades mecánicas de las células y los tejidos, evitando tiempos de recuperación prolongados47.
- Descoronar manualmente los embriones usando fórceps. Transfiera aproximadamente 10-15 embriones a recipientes de reacción de 1,5 ml utilizando una pipeta Pasteur de vidrio.
- Retire los medios E3 y agregue 500 μL de medio de cultivo tisular independiente de CO2 precalentado (DMEM-F12; con L-glutamina y 15 mM HEPES, sin bicarbonato de sodio y rojo fenol suplementado con 10 unidades de penicilina y 10 mg / L de estreptomicina).
NOTA: No utilice medios dependientes de CO2 a menos que se utilice una incubadora de microscopio. El uso de, por ejemplo, RPMI en condiciones tamponadas con carbonato causa cambios en el pH del medio y puede afectar la supervivencia celular. Otro aspecto clave es evitar los medios de cultivo que contienen suero. El suero puede contener ácido lisofosfatídico (LPA), un potente activador de la vía Rho/ROCK, capaz de controlar la contractilidad celular y la motilidad en las células madre progenitoras6. La osmolaridad del medio debe mantenerse a 300 mOsm para evitar desafíos osmóticos que puedan interferir con la morfología o mecánica nuclear12. - Disocie manualmente las células agitando suavemente el tubo. Asegúrese de que el contenido del tubo se vuelva turbio sin grandes trozos visibles para el ojo. Evitar la formación de burbujas para minimizar el daño y la pérdida de células.
- Centrifugadora a 200 x g durante 3 min. El pellet debe ser claramente visible.
- Retire el sobrenadante y siga uno de los pasos que se detallan a continuación.
- Si no se necesita tinción, agregue 500 μL de DMEM. Resuspender suavemente con una pipeta de 200 μL apuntando un chorro de líquido sobre el pellet. No ejerza una fuerza de cizallamiento excesiva sobre las células. La formación de espuma indica daño a las células.
- Para etiquetar el núcleo con colorantes de ADN como Hoechst, mezclar 0,5 μL de DNA-Hoechst (stock 2 mg/mL) en 1.000 μL de DMEM para obtener 1 μg/mL de concentración final. Agregue 500 μL de esta solución de tinción a las células y vuelva a suspender suavemente. Incubar durante 7 min en la oscuridad.
- Para teñir las células con un indicador químico fluorescente de calcio Calbryte-520, agregue Calbryte-520 a una concentración de 5 μM en DMEM. Incubar durante 20 min en la oscuridad.
NOTA: Los protocolos indicados en los pasos 2.5.2 y 2.5.3 se han optimizado para estos productos específicos. Otras tinciones se pueden realizar utilizando los protocolos indicados por el fabricante.
- Centrifugar de nuevo utilizando la misma configuración que en el paso 2.4; retirar el sobrenadante y resuspendir suavemente las células (para evitar la formación de grupos) en 50 μL de DMEM para muestras en suspensión o 20 μL de DMEM para células en confinamiento.
3. Preparación de cámaras de captura óptica utilizando espaciado de polidimetilsiloxano (PDMS)
NOTA: Las mediciones de fuerza óptica basadas en la detección del momento de la luz requieren la captura de toda la luz que emerge de las trampas ópticas40. Para la robustez del factor de calibración invariante α (pN/V), la distribución de la luz en el plano focal posterior (BFP) del sensor de fuerza óptica debe tener una correspondencia precisa con el momento del fotón. Esto determina la distancia desde la superficie de la lente colectora hasta el plano de captura a aproximadamente 2 mm, que es la altura máxima de las cámaras de captura óptica.
- PDMS spin-coating de platos con fondo de vidrio #1.5.
NOTA: La siguiente receta se proporciona para aproximadamente 40 platos. La microcámara resultante tendrá diferentes alturas dependiendo de si los experimentos se llevarán a cabo en células suspendidas o confinadas (Figura 1D).- Mezcle 9 ml del polímero base PDMS y 1 ml de agente de curado PDMS en un tubo cónico de 50 ml. Mezcle los dos productos activamente para garantizar la distribución adecuada del agente de curado.
- Desgasifica la mezcla para evitar burbujas utilizando una bomba de vacío. Introduzca el tubo cónico en una botella de vacío y evacue la cámara. Espere hasta que no haya burbujas presentes en la mezcla.
NOTA: Abra el vacío lentamente para evitar la formación de espuma y los derrames del PDMS fuera del tubo de halcón. - Coloque el plato con fondo de vidrio en el mandril de spin-coater (Figura 2A). Sea suave para no rascarse, tomar huellas dactilares o ensuciar el plato. Proteja la caja del revestimiento de centrifugado de fugas de PDMS con papel de aluminio.
- Para cámaras OT para experimentos en células en suspensión, agregue aproximadamente 250 μL de mezcla PDMS en el centro del plato inferior y gírelo a 750 rpm durante 1 min. La altura de la capa PDMS será de 50 μm aproximadamente48.
- Para cámaras OT para experimentos en células confinadas, agregue una pequeña gota de PDMS (aproximadamente 50 μL) y gírela a 4,000 rpm durante 5 minutos. La altura de la capa PDMS será de 10 μm aproximadamente. Para obtener un protocolo detallado sobre cómo obtener diferentes espesores de PDMS, consulte reference48.
- Cure los platos con fondo de vidrio recubiertos de PDMS a 70 °C durante 1 h.
- Cortar un cuadrado de 1 x 1 cm sobre la capa PDMS con un bisturí y despegarlo con pinzas (Figura 2C). En el caso de células confinadas, lave los residuos de PDMS con isopropanol.
- Recubrimiento de cámara para experimentos con células ligeramente unidas en suspensión
- Añadir 100 μL de Concanavalina A (ConA) a 0,5 mg/ml para cubrir toda la superficie de la cavidad cuadrada y dejar incubar durante 30 min.
NOTA: ConA es una lectina que se une a los azúcares de la superficie celular y acopla células individuales en la superficie del vidrio de cobertura. - Retire la gota de ConA y enjuague la superficie cuidadosamente con medio DMEM sin rayar la superficie tratada con ConA.
- Agregue 30 μL de la muestra previamente preparada (paso 2.6) en el pozo y vuelva a suspender suavemente para deshacerse de cualquier grupo celular.
- Cierre la cavidad colocando suavemente un vidrio de cubierta de 22 x 22 mm # 1.5 en la parte superior de las llantas PDMS (evite dejar que caiga abruptamente, use fórceps si es posible, Figura 2B, C).
NOTA: Cualquier grosor de la cubierta funcionaría para la cubierta superior de vidrio (la lente colectora tiene una distancia de trabajo de 2 mm).
- Añadir 100 μL de Concanavalina A (ConA) a 0,5 mg/ml para cubrir toda la superficie de la cavidad cuadrada y dejar incubar durante 30 min.
- Preparación de la cámara para experimentos con células en confinamiento
- Coloque una gota de 10 μL de solución que contenga células (paso 2.6) en la cavidad cuadrada (Figura 2B).
- Muy suavemente, empareje la muestra con un vaso de cubierta de 22 x 22 mm de tal manera que la gota se extienda en toda el área y no se observen burbujas. Una vez más, es conveniente usar fórceps, como se muestra en la Figura 2C, para evitar que el vidrio de la cubierta caiga abruptamente.
4. Opciones alternativas para el espaciado de la cámara OT
NOTA: Estos pasos se pueden seguir si no hay un taller de microfabricación o un revestimiento de centrifugado disponible.
- Preparación de la cámara para experimentos con células en suspensión
NOTA: En caso de que no haya un recubrimiento de centrifugado disponible, se puede hacer un espaciador con cinta adhesiva normal de doble cara (aproximadamente 100 μm de altura).- Cortar un trozo de cinta adhesiva de doble cara con un orificio cuadrado de aproximadamente 10 cm x 10 cm en el centro (mismas dimensiones que en PDMS, Figura 2B).
- Retire una de las capas protectoras de la cinta despegándola y coloque el lado descubierto de la cinta en el centro de un plato con fondo de vidrio # 1.5 H. Presione suavemente para que toda la superficie se adhiera al vidrio mientras evita las burbujas de aire, y luego retire la capa protectora restante de la cinta despegándola.
- Siga las instrucciones del paso 3.2.
- Preparación de la cámara para experimentos con células en confinamiento
NOTA: Para confinar con precisión las células, las micropartículas monodispersas con un diámetro conocido se pueden usar como espaciadores entre los dos vidrios de cubierta.- Añadir perlas de poliestireno de 10 μm a las células suspendidas a una concentración de 104 perlas/μL.
- Coloque una gota de solución de 10 μL que contenga células y perlas en un vidrio de cubierta de 22 x 60 mm.
- Muy suavemente, empareje la muestra con otro vaso de cubierta de 22 x 60 mm de tal manera que la gota se extienda en toda el área y no se observen burbujas. Para colocar el vidrio de la cubierta superior suavemente (evitar que se caiga bruscamente), es conveniente usar fórceps.
- Como la muestra puede secarse, se recomienda realizar la preparación rápidamente.
5. Configuración de la trampa óptica para mediciones intracelulares
NOTA: Los siguientes pasos están optimizados para una plataforma comercial de pinzas ópticas que comprende un módulo de micromanipulación óptica basado en la deflexión acusto-óptica (AOD) y un sensor de fuerza óptica basado en la detección directa de los cambios de momento de la luz (Figura 2, referencia12,40,49). Los detalles y los componentes ópticos de la configuración se pueden encontrar en la Figura 2F. Para observar la deformación inducida por la fuerza durante las manipulaciones de las pinzas ópticas, se acopla un microscopio confocal de disco giratorio de Nipkow en el puerto izquierdo del microscopio invertido para obtener imágenes de fluorescencia de doble color. Sin la falta de generalidad, este protocolo se puede aplicar con cualquier sistema de OT dinámico equipado con mediciones de fuerza directa basadas en la detección de momento de luz. Los procedimientos detallados paso a paso están disponibles para construir trampas de gradiente óptico construidas en casa para aplicaciones in vivo50. Las basadas en la modulación AOD destacan por eventuales experimentos con múltiples trampas y mediciones rápidas51,52. En la literatura existen varios protocolos para construir un instrumento basado en el momento de la luz36,39,40,53, y se puede emplear cualquier otra modalidad de imagen (contraste de interferencia diferencial, fluorescencia de campo ancho, etc.).
- Puesta en marcha de pinzas ópticas
- Para optimizar la estabilidad de la potencia de salida, encienda el láser a una potencia considerablemente alta (por ejemplo, 3 W) al menos 30 minutos antes del experimento.
- Encienda el módulo electrónico de las unidades ópticas de micromanipulación y medición de fuerza.
NOTA: Aplicar todas las medidas de seguridad láser y utilizar únicamente equipos homologados por la junta institucional. Nunca use los oculares del microscopio óptico cuando el láser esté encendido. Utilice siempre gafas de protección IR aprobadas (OD7 en el rango de 950-1080 nm), bloquee la luz láser IR con el obturador en el puerto de epifluorescencia 2 y no ejecute el software de captura óptica hasta que finalice la alineación del sensor de fuerza óptica después del paso 5.3. En general, no utilice una muestra altamente reflectante, ya que la retrorreflexión podría causar daños al láser. - Controle la potencia de la trampa con el HWP giratorio (Figura 2F) en la entrada del módulo de micromanipulación óptica.
NOTA: El módulo de micromanipulación óptica comercial utilizado en este protocolo ya incorpora esta característica. Para los sistemas de captura óptica de fabricación casera, integre esta herramienta para el control de potencia para que se puedan utilizar potencias láser más altas y estables.
- Utilice una microcámara vacía para la calibración
- Corte un cuadrado de 1 x 1 cm sobre una cinta adhesiva de doble cara y colóquelo en un portaobjetos de microscopio de 1 mm de grosor.
- Agregue agua al cuadrado y ciérrelo desde la parte superior con un vaso de cubierta # 1.5 (22 x 22 mm). Se recomienda agregar un volumen ligeramente mayor de agua, por ejemplo, 30-40 μL para evitar burbujas dentro de la cámara cubierta. Limpie la cámara de calibración suavemente en caso de que el agua se derrame fuera de ella.
- Alineación del sensor de fuerza óptica
- Coloque una gota de agua en el objetivo de inmersión en agua 60x/1.2. Coloque la cámara de calibración en el escenario con el vidrio de la cubierta # 1.5 frente al objetivo. Concéntrese en la superficie inferior, donde eventualmente estarán las muestras de células.
- Agregue una gota de aceite de inmersión en la parte superior de la diapositiva de vidrio superior que cubre la muestra (Figura 2D). Baje la lente colectora de la unidad del sensor de fuerza con cuidado hasta que entre en contacto con la gota de aceite.
NOTA: La gota debe ser lo suficientemente grande como para cubrir toda la lente que recoge la luz láser que emerge de las trampas. Por lo general, 200 μL es suficiente para cubrir toda la superficie y proporcionar un contacto de inmersión estable. Sea conservador y evite el llenado excesivo, ya que podría filtrarse en la muestra. - Siguiendo el protocolo del fabricante para la alineación del sensor de fuerza óptica, mire la imagen del plano de muestra en la cámara auxiliar que se utilizará para posicionar los OT (AUX, Figura 2F). Muy suavemente, baje el sensor de fuerza óptica hasta que el stop de campo (FS, Figura 2F-G) aparezca conjugado en el plano de muestra. Esto garantizará mediciones de fuerza directa adecuadas a partir de la detección invariante de muestras de cambios de momento de la luz40.
NOTA: Cierre el FS lo suficiente para que su imagen sea más pequeña que el campo de visión (FOV), por lo tanto, visible. Tenga mucho cuidado y no empuje la lente colectora del sensor de fuerza óptica contra la muestra. La posición vertical del sensor de fuerza óptica se puede determinar alternativamente a partir del análisis de la distribución de la luz de captura en el BFP para conos de luz con apertura numérica definida (NA). - Asegúrese de que no haya burbujas de aire en la gota de aceite; estos pueden afectar directamente las mediciones de fuerza. Para comprobar si hay burbujas de aire, coloque la lente Bertrand en su lugar (BL, Figura 2G) y observe la trayectoria de la imagen a través del ocular. Si hay suciedad o burbujas de aire visibles o se necesita más aceite (Figura S1A), limpie la lente y la cámara con tejido de lente libre de polvo y repita el procedimiento en los pasos 5.3.2 y 5.3.3. En la figura S1B se representa una ruta óptica sin obstáculos.
- Usando los tornillos laterales colocados en el soporte del sensor de fuerza óptica, centre el FS en el FOV. Para mayor precisión, abra el FS para que casi llene el FOV visible en la cámara auxiliar (AUX, Figura 2F).
6. Optimización de pinzas ópticas
NOTA: La medición de la fuerza directa se basa únicamente en el cambio del momento de la luz que surge de la fuerza ejercida sobre la partícula atrapada y, por lo tanto, a diferencia de los métodos indirectos, la rigidez de la trampa no necesita ser calibrada antes de cada experimento. La conversión específica del instrumento del factor de deflexión/fuerza (α; pN/V, referencia41) es calibrada por el fabricante y, por lo tanto, es invariante del experimento. Sin embargo, debido a que el punto láser se manipula en un área de 70 μm x 70 μm, los pasos 6.2-6.5 son críticos para garantizar una captura óptima y la estabilidad de la potencia. Los siguientes pasos se suministran en el software del fabricante para que los OT se optimicen en el área de trabajo de forma semiautomática.
- Inicie el software OTs y el software de adquisición para cámara AUX.
- Reste la línea de base de voltaje inicial haciendo clic en el paso Paso 1: Compensación electrónica en el submenú Calibración del sistema del software de conducción de pinzas ópticas.
- Para realizar el aplanamiento de la potencia de la trampa en el área de trabajo de OT, ajuste la potencia de la trampa a la mitad de su máximo girando el HWP en consecuencia. No cambie la potencia de la trampa cambiando la salida del láser, sino con el HWP giratorio (Figura 2F). Haga clic en Paso 2: Encendido para iniciar la rutina automatizada para el aplanamiento de la potencia de la trampa.
NOTA: Este es un paso crítico para compensar la variación de la potencia de la trampa en el área de trabajo de OT (Figura S1D). Una rutina exitosa reduce la variación de la potencia de la trampa al 2% en el área de trabajo de los OT y converge después de 2 minutos. - Para realizar la calibración de la posición de la trampa, retire el filtro IR para que la luz del láser sea visible en la cámara. Encuentre el punto IR configurando el plano de la imagen enfocado en la superficie inferior de la microcámara. Obtenga el punto IR más pequeño posible ajustando el plano de la imagen (posición del objetivo) y el contraste del histograma en el software de adquisición AUX de la cámara. Si es necesario, reduzca la potencia de la trampa óptica girando el HWP (Figura 2F). Haga clic en Paso 3: Posición para iniciar la rutina automatizada o la calibración de posicionamiento de trampa.
NOTA: Esta rutina permite la correspondencia precisa de las coordenadas de posición del OT en la cámara AUX con los ángulos de dirección del AOD. Una rutina exitosa genera el mapeo de ángulo a posición en varios segundos. - Compensación de impulso inicial
NOTA: El movimiento de la trampa óptica a través de la muestra causa variaciones en la distribución luz-momento en el BFP (Figura S1E, F). Esto conduce a cambios de señal independientes de la fuerza relacionados con la posición del láser sobre el área de trabajo, a pesar de que la potencia de la trampa se ha aplanado como en el paso 6.3. La consecuencia es una variación en la línea de base de la fuerza debido a la posición (independiente de una fuerza real que actúa sobre la cuenta atrapada ópticamente) que debe corregirse antes de cada experimento.- Establezca la potencia de captura que se utilizará en los experimentos, girando el HWP (Figura 2F).
- Haga clic en la opción Desplazamiento global en el submenú Herramientas . Esto abrirá el asistente Offset Cancel del software de pinzas ópticas que corrige la línea de base de momento inicial.
- Haga clic en | de desvío Compensar para corregir el momento inicial de la variante de posición.
NOTA: Si ninguna modificación afecta a la trayectoria óptica durante las semanas en curso, los mapas de aplanamiento de potencia de la trampa (paso 6.3) y posición (paso 6.4) permanecerán invariantes. Por lo tanto, recomendamos utilizar siempre la misma combinación de elementos ópticos (espejos dicroicos, filtros, etc.) que puedan afectar la trayectoria de la trampa láser o llevar a cabo una nueva rutina de aplanamiento de potencia de trampa. Con respecto a la compensación de impulso inicial (paso 6.5), el fabricante de la plataforma OTs proporciona una calibración sobre la marcha que debe cambiarse para cada nueva potencia de captura y sesión experimental. Los pasos 6.3 y 6.4 deben llevarse a cabo en la diapositiva de calibración vacía descrita en el paso 5.2. En una muestra que contenga células u otros objetos, el paso 6.5 debe llevarse a cabo libre de objetos que puedan alterar la dispersión de la luz en el área de trabajo de los OT.
- Opcionalmente, atrape una microesfera y mueva la trampa a una velocidad conocida mientras graba la señal de fuerza. Por ejemplo, configure la trampa para realizar una oscilación triangular: la señal de fuerza registrada será una señal cuadrada.
NOTA: El valor de la fuerza debe aumentar linealmente con la velocidad, de acuerdo con la fuerza de arrastre que actúa sobre la cuenta. Esta prueba sirve como un control positivo de que las mediciones de fuerza se están realizando correctamente38. Alternativamente, el sensor de fuerza óptica se puede utilizar para obtener la rigidez de atrapamiento óptico, κ [pN/μm], y el factor de calibración de posición, β [μm/V], a partir del análisis espectral de potencia35. Bajo la alineación correcta, el factor de calibración invariante proporcionado por el fabricante es α = κ·β [pN/V].- Inicie una lectura de fuerza en tiempo real haciendo clic en la gráfica 1 en el submenú Medidas del software del fabricante. Esto proporcionará una lectura de la fuerza y potencia de captura óptica actual.
- Abra el cuadro de diálogo Parámetros de oscilación desde el submenú Herramientas . Establezca una forma de onda de espacio triangular en los anillos selectores de forma y tipo, respectivamente. Como ejemplo, establezca una amplitud de 10 μm y una frecuencia de 3 Hz. Esto resultará en una fuerza viscosa de aproximadamente 1 pN sobre una microperla con un diámetro de 1 μm38.
- En la ventana AUX de la cámara, haga clic con el botón derecho en la microperla y seleccione Iniciar oscilación. La lectura de fuerza se convertirá en una señal de fuerza cuadrada con mesetas en ±1 pN.
- Haga clic con el botón derecho en la microperla y seleccione Detener oscilación.
7. Microscopía confocal de disco giratorio
- Encienda el microscopio confocal de disco giratorio y el equipo accesorio, los motores láser integrados y las cámaras de adquisición.
- Inicie el software de imágenes.
- Establezca canales de imágenes para la tinción de Hoechst del núcleo y GFP para la membrana plasmática celular.
- Active las líneas de láser de excitación de 405 nm y 488 nm.
- Añade un dicroico multibanda para reflejar la excitación a la muestra y que permita que la luz emitida pase a las cámaras.
- Divida la emisión de fluorescencia con un espejo dicroico de borde de paso largo de 500 nm.
- Utilice los filtros de emisión DAPI/BFP (~445 nm) y GFP (~521 nm) delante de las dos cámaras de adquisición, respectivamente. Consulte la Figura 2F,G.
- Establezca el tiempo de exposición en 100 ms para cada canal.
- Ajuste la emisión láser para obtener una potencia de 5 mW en el plano de muestreo. Para medir la potencia, use un medidor de potencia comercial.
- Establezca el protocolo de imágenes. Para evitar el sangrado espectral desde el canal hoechst hacia el canal GFP, los dos tintes deben visualizarse secuencialmente.
NOTA: Si existe una sincronización de hardware entre los AOD de la trampa óptica y la adquisición de la cámara, asegúrese de que la polaridad del disparador esté configurada correctamente. En caso de duda, consulte al gerente de su instalación o al fabricante del microscopio.
8. Realización de los experimentos de indentación del núcleo
NOTA: Apague siempre las trampas ópticas, tanto con software como cerrando el obturador en el puerto de epifluorescencia 2, al levantar el módulo del sensor de fuerza y cambiar la muestra. De lo contrario, podrían producirse daños graves en los elementos ópticos y en el experimentador. Tenga cuidado con la distancia lateral entre el soporte de la lente y el borde inferior del plato cuando busque células para evitar chocar la lente con el plato de escenario / cultivo (Figura 2).
- Coloque la muestra en el microscopio y siga el paso 5.3 de este protocolo.
- Utilizando el HWP giratorio (Figura 2F), establezca la potencia de la trampa en 200 mW como valor de partida si no se conoce la rigidez del núcleo o la estructura intracelular investigada. Traducir el área de trabajo de los OT (utilizando la etapa de microscopio) a un lugar libre de células para compensar la línea de base del momento inicial hasta el paso 6.5.
NOTA: Dependiendo de la rigidez de la estructura subcelular, el valor de potencia de la trampa debe ajustarse a valores más bajos o más altos para obtener una profundidad de indentación similar. - Usando el controlador de software de etapa de microscopio, busque una célula con una o dos perlas a través de microscopía de campo brillante transmitida (Figura 3A).
- Definir una trayectoria de trampa.
- Abra el cuadro de diálogo Trayectoria en el submenú Herramientas y elija Desplazamiento en el anillo selector Tipo de trayectoria .
- En la hoja numérica, escriba el desplazamiento y el tiempo de cada paso de trayectoria posterior. Aquí hay dos ejemplos.
- Para un experimento de relajación del estrés, programe cargas trapezoidales, como se muestra en la Figura 3B. En la Tabla S1, se aplicaron dos hendiduras trapezoidales con una distancia de recorrido de 5 μm; velocidad de 5 μm/s; tiempo de espera antes de la retracción: 10 s.
- Para un experimento de indentación repetitiva a una velocidad constante para obtener una rutina triangular sin tiempo de permanencia en el núcleo, establezca la amplitud de la trayectoria, por ejemplo, 5 μm, y el tiempo para el paso, por ejemplo, 2 s para una velocidad de 2.5 μm / s. En la Tabla S2, esto se aplica ocho veces a la misma velocidad.
NOTA: Estos valores deben determinarse para cada tipo de célula y experimento, pero los siguientes parámetros de una rutina trapezoidal capturan la dinámica más importante en el experimento presentado aquí. El tiempo de espera debe ser suficiente para que el núcleo muestre su relajación completa del estrés después de la hendidura.
- Atrapando una microesfera
- Coloque el plano de la imagen ligeramente por encima de la cuenta con el controlador de software de la etapa del microscopio.
- Active las trampas utilizando el software OTs y haga clic en la cuenta en la ventana de imágenes AUX de la cámara (calibrada siguiendo el paso 6.4). El confinamiento exitoso de la cuenta por la trampa óptica reducirá fuertemente el movimiento de la cuenta.
- Haga clic y arrastre la cuenta a través del citoplasma y colóquela a una distancia de ~ 2 μm de la envoltura nuclear (Figura 3A). Asegúrese de que la trayectoria esté establecida de modo que la hendidura de la perla sea perpendicular a la membrana nuclear.
- Opcionalmente, si es necesario para las mediciones de posición de la perla en relación con la trampa, escanee la trampa a través de la cuenta para determinar la rigidez de captura, k [pN/μm]54, por lo tanto Δxbead = -F/k (ver Discusión). El módulo de micromanipulación óptica utilizado en este protocolo tiene una rutina incorporada para este propósito.
- Abra el cuadro de diálogo Análisis de partículas en el submenú Herramientas .
- Seleccione la trampa que desea escanear y Alta frecuencia como método de escaneo. Seleccione la dirección (x o y) de la trayectoria de sangría para la medición de escaneo de perlas.
- Aparecerá una ventana con la medición de la rigidez de atrapamiento. En el gráfico, arrastre los dos cursores para seleccionar el área de reventado lineal correspondiente a F = -kx. El ajuste lineal a la parte de datos seleccionada se actualizará automáticamente.
NOTA: Establezca la posición inicial de la cuenta lejos de la membrana celular (~ 5 μm), ya que las deflexiones de momento de luz en la interfaz de celda media afectan la idoneidad de las mediciones de fuerza. Si el núcleo se encuentra demasiado cerca de la membrana celular, trate de sangrar el núcleo desde el sitio opuesto. Deseche la celda si no es posible.
- Inicie la adquisición de imágenes haciendo clic en el botón de adquisición en el software de imágenes.
- Comience a guardar los datos de medición de posición y fuerza de trampa haciendo clic en Data | Guardar en la ventana de lectura forzada en tiempo real (abierta como en el paso 6.6.1).
NOTA: La trampa óptica está equipada con una entrada de disparo que se puede conectar a la salida de temporización de la cámara. Por lo tanto, los datos de imagen y fuerza están sincronizados por hardware y el electrónico es capaz de mapear los ciclos de trampa con el número de fotogramas de las imágenes durante la adquisición. - Inicie la trayectoria cargada previamente haciendo clic con el botón derecho en la cuenta y seleccionando Iniciar trayectoria.
- Espere hasta que finalice la trayectoria y el sistema se estabilice.
- Ahorro de datos de medición de fuerza de trampa de parada. Aparecerá un cuadro de diálogo de ahorro de datos.
NOTA: Para optimizar el almacenamiento de datos, los datos se pueden diezmar seleccionando el parámetro de diezmado en este cuadro de diálogo (10, 100 o 1000). - Detenga la adquisición de imágenes y trace los resultados en el software de postprocesamiento de la elección del usuario.
- Si la microesfera se pierde durante la rutina y el núcleo no se puede sangrar (Figura S2), descarte la medición y aumente la potencia. Tenga en cuenta que el paso 6.5 debe repetirse. En nuestras manos, al menos el 95% de las rutinas se completan con éxito sin perder la cuenta de la trampa.
Resultados
Microinyección de perlas tramposas:
Las microesferas inyectadas en el embrión de pez cebra de una célula se extendieron por toda la tapa del animal durante la morfogénesis. Para una visualización más clara, repetimos el protocolo de inyección con microperlas fluorescentes rojas y tomamos imágenes volumétricas con nuestro microscopio confocal en diferentes etapas de desarrollo. En la Figura 4A-D, las perlas inyectadas se visualizan en el citoplasma de las células madre progenitoras in vivo a 5 hfp. Más tarde, aparecieron microesferas repartidas sobre todo el embrión a 24 hpf (Figura 4E). Los embriones en ambas etapas se desarrollaron normalmente y las tasas de supervivencia fueron comparables con los embriones controlados no inyectados o simulados (ver Figura S3). Esto es consistente con otros estudios que reportan una supervivencia imperturbable del pez cebra inyectado con cuentas hasta 5 días después de la fertilización55.
Nuestro microscopio confocal de disco giratorio es compatible con microcopia de fluorescencia multicanal. En la Figura 5A, mostramos células madre aisladas con una o dos perlas en el citoplasma. Se pueden usar múltiples etiquetas fluorescentes para investigar diferentes aspectos de la célula (Figura 5B). La morfología nuclear se puede rastrear con un tinte Hoechst o utilizando una expresión de ARNm H2A::mCherry, mientras que la membrana nuclear interna se puede analizar con Lap2b-eGFP12. La dinámica de la corteza actomiosina, así como los niveles de calcio intracelular, se pueden observar con una línea transgénica My12.1::eGFP56 y una incubación de Calbryte-520, respectivamente. El protocolo que se ha descrito aquí tiene como objetivo comparar la mecánica del núcleo celular de células silvestres inmovilizadas en sustratos adhesivos (más tarde denominados suspensión) y en confinamiento mecánico. Las células madre aisladas confinadas en microcámaras de 10 μm de altura exhibieron un despliegue parcial de la membrana nuclear interna (INM) y un posterior aumento de la contractilidad de la actomiosina12. En la Figura 5C, se muestran las células confinadas con una o dos perlas en el citoplasma. El confinamiento exitoso será visible a través de células aplanadas y expandidas con una sección transversal más ancha del núcleo. La membrana nuclear se despliega aún más en células confinadas y debe aparecer suavizada en comparación con las células en suspensión (Figura 5C).
Análisis fuerza-tiempo y fuerza-deformación
El análisis de los resultados obtenidos depende en gran medida del espécimen investigado y de la cuestión de interés y, por lo tanto, no se pueden generalizar aquí. Como ejemplo, una forma común de analizar la medición de la sangría es extraer un módulo de Young ajustando un modelo de Hertz modificado a los datos de fuerza-sangría57. Sin embargo, la suposición para tal tratamiento debe evaluarse cuidadosamente y no siempre puede justificarse adecuadamente (como que la estructura investigada sea isotrópica, homogénea, con elasticidad lineal y hendiduras más pequeñas que el radio de la perla). Por lo tanto, solo consideramos aquí mediciones independientes del modelo que permiten comparar el comportamiento mecánico de la estructura investigada entre diferentes escenarios experimentales.
Como punto de partida, la medición de la pendiente de la curva fuerza-desplazamiento a una cierta profundidad de indentación proporciona una medida de la rigidez estructural independiente de un modelo58 del núcleo. Este valor se puede recolectar de múltiples muestras y comparar entre diferentes configuraciones experimentales y perturbaciones de muestra.
Medición de sangría
En las siguientes líneas, nos centramos en la respuesta mecánica del núcleo celular durante la deformación celular en confinamiento. Los experimentos en el paso 8 de este protocolo generalmente conducen a picos de fuerza de hasta 200 pN para profundidades de indentación de aproximadamente 2-3 μm. Sin embargo, estos valores pueden ser muy diferentes, dependiendo del tipo de célula y las condiciones experimentales, con núcleos más blandos que conducen a una fuerza más baja para una hendidura dada. Por lo tanto, es necesario medir con precisión la deformación nuclear, junto con la fuerza, para una caracterización mecánica precisa del núcleo celular. En esta sección, obtendremos la rigidez nuclear de la célula a partir de mediciones representativas de la hendidura de fuerza.
En la Figura 6, mostramos las deformaciones de los lados distal y proximal de un núcleo en una célula suspendida y confinada. Se puede observar un rico comportamiento mecánico. En una célula suspendida típica sobre un sustrato adhesivo, el núcleo estaba fuertemente sangrado por la cuenta, pero también ligeramente desplazado por eventos repetitivos de empuje. Medimos la hendidura de perlas en el núcleo mediante el análisis de los kymographs obtenidos de imágenes de fluorescencia de núcleos celulares teñidos de Hoechst. Los kymographs se calcularon fácilmente utilizando el complemento Multi Kymograph de Fiji a lo largo de la dirección de sangría (Figura 6A, B) y se importaron a Matlab (Versión 2021, Mathworks) para su posterior procesamiento. Se ajustó una función de paso al perfil de intensidad bruta con el objetivo de rastrear los bordes delimitadores del núcleo a lo largo de la trayectoria de la rutina de indentación. Como se puede ver, contiene información precisa sobre el cambio de forma nuclear (Figura 6 y Figura S2). Utilizamos la siguiente curva de doble sigmoide como versión analítica de una función de paso:
 (Ecuación 1)
(Ecuación 1)
Aquí, x1 y x2 denotan los bordes distales y proximales del núcleo, mientras que A y B son los valores máximos y grises de fondo del canal azul (tinte hoechst) de la imagen (Figura 6B). Se ha considerado el ancho del borde (e0 = 0,25 mm). Mientras que el borde del núcleo proximal con sangría (x2) siguió la trayectoria aplicada por la rutina de trampa óptica después del contacto microesfera-núcleo, el borde distal opuesto (x1) muestra una dinámica de relajación como se esperaba para un material viscoelástico como el citoplasma (Figura 6D). Por el contrario, los núcleos en células confinadas en microcámaras de 10 μm de altura no exhiben tal comportamiento de translocación del núcleo tras la hendidura dentro de la célula (Figura 6B, D). También se muestra en la Figura 6D, los bordes posteriores de los núcleos permanecen inalterados por el empuje de la cuenta desde el lado proximal, muy probablemente debido a fuerzas más fuertes derivadas de la contractilidad celular y la fricción que actúan contra la fuerza de indentación. Para obtener la profundidad de deformación correcta, se restó el desplazamiento x1 de la medida con sangría x2: Δx = x2 - x1 (ver también Figura 6D).
Análisis de datos de fuerza
La fuerza que causa la deformación nuclear se midió a partir del cambio en el momento de la luz originado en la microperla atrapada ópticamente (Figura 7A). La fuerza al aplicar trayectorias trapezoidales (paso 8.4.3, Figura 7B) inicialmente aumentó linealmente hasta que la trampa dejó de moverse, pero luego se relajó a un valor de estado estacionario. Este comportamiento indicó un material viscoelástico que exhibe módulos de pérdida y almacenamiento. Justo después del evento de indentación, la fuerza alcanzó un valor máximo, Fp, seguido de una relajación por estrés (Figura 7C):
 (Ecuación 2)
(Ecuación 2)
donde F0 es la fuerza almacenada para el componente elástico y f(t) es una función de relajación sin dimensión. Hemos analizado este comportamiento de tres maneras:
1. Considerando un sólido lineal estándar con una relajación de tensión exponencial, es decir, f(t) = e-t/τ, representado esquemáticamente en el recuadro de la Figura 7C.
2. Usando un decaimiento general, doble exponencial:
F(t) = A + B1e-t/τ1 + B2e-t/τ2.
3. Usando una ley de potencia seguida de una decadencia exponencial59:
f(t) = t-pe-t/τ, incluido en la figura 7C.
Si bien el ajuste para el modelo 1 se puede llevar a cabo directamente, recomendamos estimar las conjeturas iniciales para (τ1, τ2) y (p, τ) para los modelos 2 y 3, respectivamente. Esto se puede realizar, respectivamente, ajustando líneas a los datos en escalas logarítmicas versus lineales (Figura 7D, izquierda) y logarítmicas versus logarítmicas (Figura 7D, derecha). La Tabla S3 resume los resultados del ejemplo analizado en la Figura 7. En la siguiente sección, consideraremos la combinación de una ley de potencia y una ley exponencial para la caracterización de la mecánica del núcleo celular.
Relación de desplazamiento de fuerza
Del mismo modo, la configuración experimental descrita se puede utilizar para obtener la relación fuerza-desplazamiento de múltiples eventos de sangría. Mediante la realización de rutinas triangulares (paso 8.4.4, Figura 8A), es posible relacionar la fuerza con la deformación y trazar una curva de fuerza-indentación. Un resultado ejemplar se muestra en la Figura 8B, en la que una línea de base plana cambió suavemente de pendiente una vez que la cuenta entró en contacto con el núcleo. Identificar el verdadero punto de contacto en los datos ruidosos es un desafío, y se debe tener cuidado para ver si la región de contacto se ajusta a los modelos elásticos60. En este experimento en particular, también se pudo ver que las hendiduras posteriores dan como resultado curvas con puntos de contacto más profundos, lo que indica una recuperación de la forma nuclear demasiado lenta después de la retracción de la perla y un cambio en el ciclo histérico definido por las propiedades del material viscoelástico del núcleo (Figura 8C). Por lo tanto, el investigador debe ser consciente de si esto sucede e incorporarlo en la tubería analítica, o restringir el número de mediciones posteriores de modo que este efecto no modifique la medición.
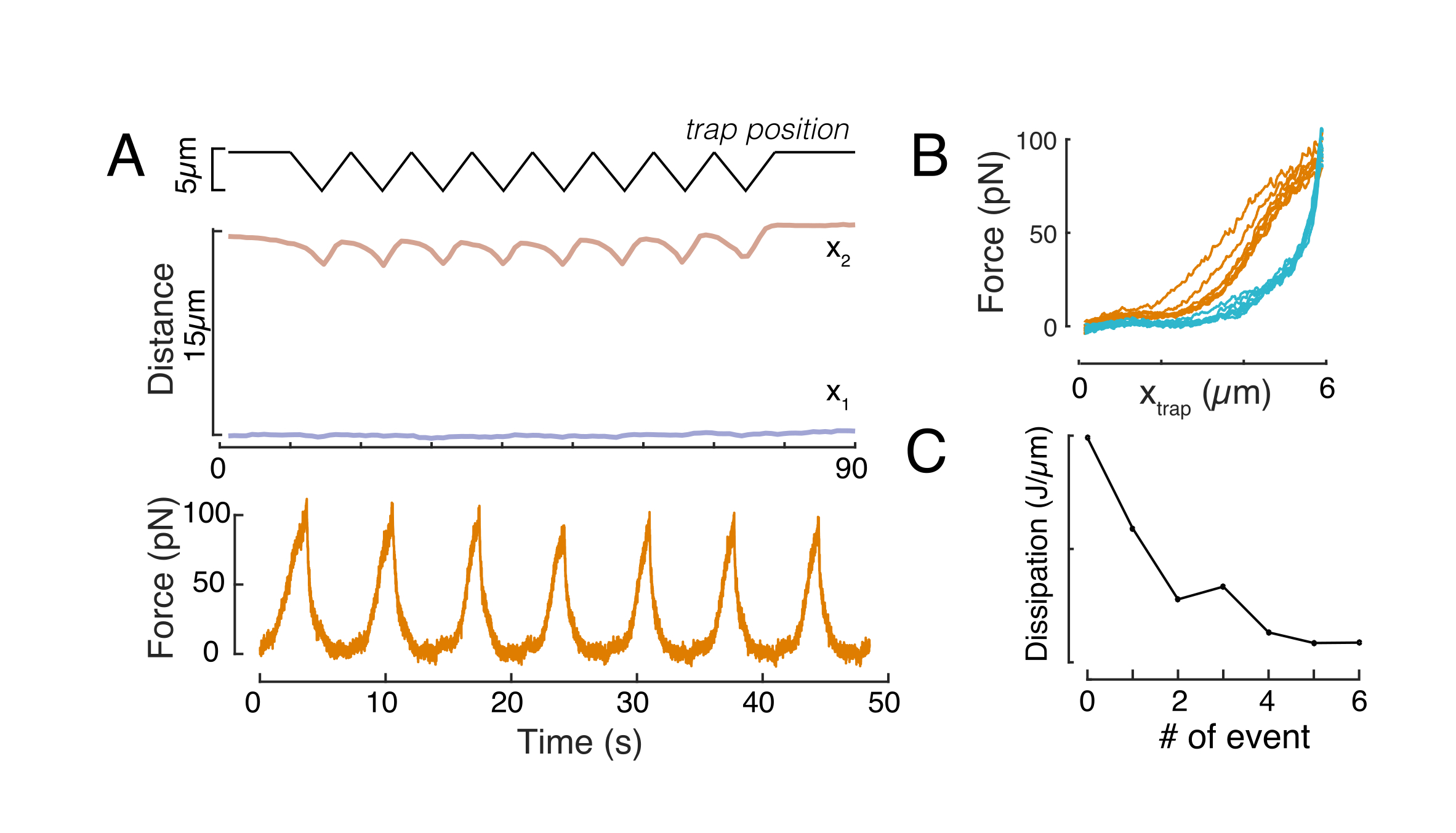
Mecánica del núcleo en células en suspensión y confinamiento por debajo de 10 μm
El enfoque antes mencionado se utilizó para analizar la dinámica de la relajación del estrés del núcleo en células suspendidas sobre sustratos adhesivos y células confinadas. Nuestros resultados muestran que el confinamiento resulta en una expansión del área proyectada (Figura 9A), pero un cambio insignificante en la rigidez nuclear (Figura 9B). Medimos una relajación similar con τ = 6,08 ± 1,1 s (no confinado) y τ = 4,00 ± 0,6 s (confinamiento), lo que indica una rápida disipación viscoelástica, seguida de un valor de fuerza almacenada que corresponde al módulo elástico del núcleo. Con el fin de tener en cuenta las variaciones experimentales, que pueden ser producidas por diferentes condiciones iniciales en las rutinas de sangría, las fuerzas almacenadas medidas se normalizaron a la profundidad de la sangría, como  . Este parámetro explica la rigidez del núcleo y describe la fuerza, o la tensión, necesaria para una determinada hendidura. Obtuvimos una rigidez similar bajo confinamiento y en celdas no confinadas: = 20,1 ± 12,6 pN/μm y = 24,6 ± 13,6 pN/μm (media ± desviación estándar), respectivamente.
. Este parámetro explica la rigidez del núcleo y describe la fuerza, o la tensión, necesaria para una determinada hendidura. Obtuvimos una rigidez similar bajo confinamiento y en celdas no confinadas: = 20,1 ± 12,6 pN/μm y = 24,6 ± 13,6 pN/μm (media ± desviación estándar), respectivamente.

Figura 1: Microinyección de embriones de pez cebra en la etapa de una célula (cigoto). (A) Placa de inyección: se utiliza una placa de inyección de forma triangular para la inyección. La placa está hecha de agarosa ultrapura al 1% en E3 (medio de huevo). Las vistas superior y lateral se muestran a la derecha. (B) Posicionamiento embrionario: oriente suavemente los embriones con un cepillo y oriente de tal manera que la célula sea claramente visible y fácilmente accesible con la aguja. Sugerimos orientar los embriones con la célula ubicada en el lado opuesto de la aguja, como se muestra en el boceto. (C) Procedimiento de inyección en el embrión en etapa de una célula: perfore el corion que rodea el embrión y la célula única con la aguja. Asegúrese de que la punta de la aguja esté dentro de la célula y libere la presión para inyectar. (D) Incubar los embriones a 28-31 °C hasta que se desarrollen hasta la etapa de blástula (esfera) (4 hpf). Realizar el protocolo de aislamiento celular y tinción celular (paso 2) y preparar la cámara de captura óptica con celdas aisladas en suspensión y/o confinamiento combinadas con el correspondiente recubrimiento superficial del sustrato (paso 3). Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 2: Preparación del aparato de pinza óptica. (A) Capas de recubrimiento por centrifugado de PDMS con una altura definida sobre platos con fondo de vidrio. La gota de PDMS se extenderá uniformemente debido a la fuerza centrífuga. (B) Preparación de la cámara de muestra fuera de la capa PDMS. 1: cortar un cuadrado con un bisturí, 2: cubrir bien el interior con concanavalina A (ConA), lavar y sembrar células; 3: cubra con un portaobjetos de vidrio o un resbalón de cubierta para sellar el pozo. (C) Imagen del cuadrado cortando con un bisturí y retirando bien el PDMS con fórceps. (D) Montaje de la lente colectora del sensor de fuerza óptica sobre la cámara de captura. Una gota de aceite de inmersión sirve como medio de inmersión entre la lente colectora y la cubierta superior de vidrio. Esquema no a escala. Tenga cuidado al bajar la lente colectora para no tocar la cubierta de vidrio del plato de muestra. (E) Imagen de la unidad de detección de fuerza en contacto con la muestra. F) Esquema de la configuración experimental. El módulo de micromanipulación óptica utiliza un rayo láser de onda continua (5W, λ = 1064 nm) con control de potencia a través de una placa de media onda (HWP) y un divisor de haz polarizador (BS). Después de ser modulado con un par de AODs, se acopla al puerto de epifluorescencia superior de un microscopio invertido. El rayo láser es reflejado por un espejo dicroico de paso corto de 950 nm (IR-DM), lo que permite la transmitancia de la excitación y emisión de fluorescencia. El láser de captura se guía hacia el puerto de epifluorescencia posterior del microscopio (torreta superior). Los OT se crean en el plano focal de una lente de objetivo de inmersión en agua (60x, NA = 1.2). El sensor de fuerza óptica es sometido por la torreta del microscopio y captura la luz láser que emerge de los OT con una lente de inmersión en aceite de alto NA. Al mismo tiempo, el sensor de fuerza permite la iluminación de campo brillante. La unidad confocal de disco giratorio está acoplada al puerto izquierdo. Está equipado con dos motores láser integrados (ILE) que controlan siete láseres de excitación de fluorescencia y dos cámaras sCMOS retroiluminadas, lo que permite obtener imágenes de fluoróforos duales en paralelo Abb: TI, Transilluminator; FS, parada de campo; AOD: deflector acustoóptico; HWP: placa de media onda; CAM, cámara (G) Fotografía del equipo de captura óptica. El círculo rojo indica la lente Bertrand, que se puede cambiar a la ruta óptica manualmente. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

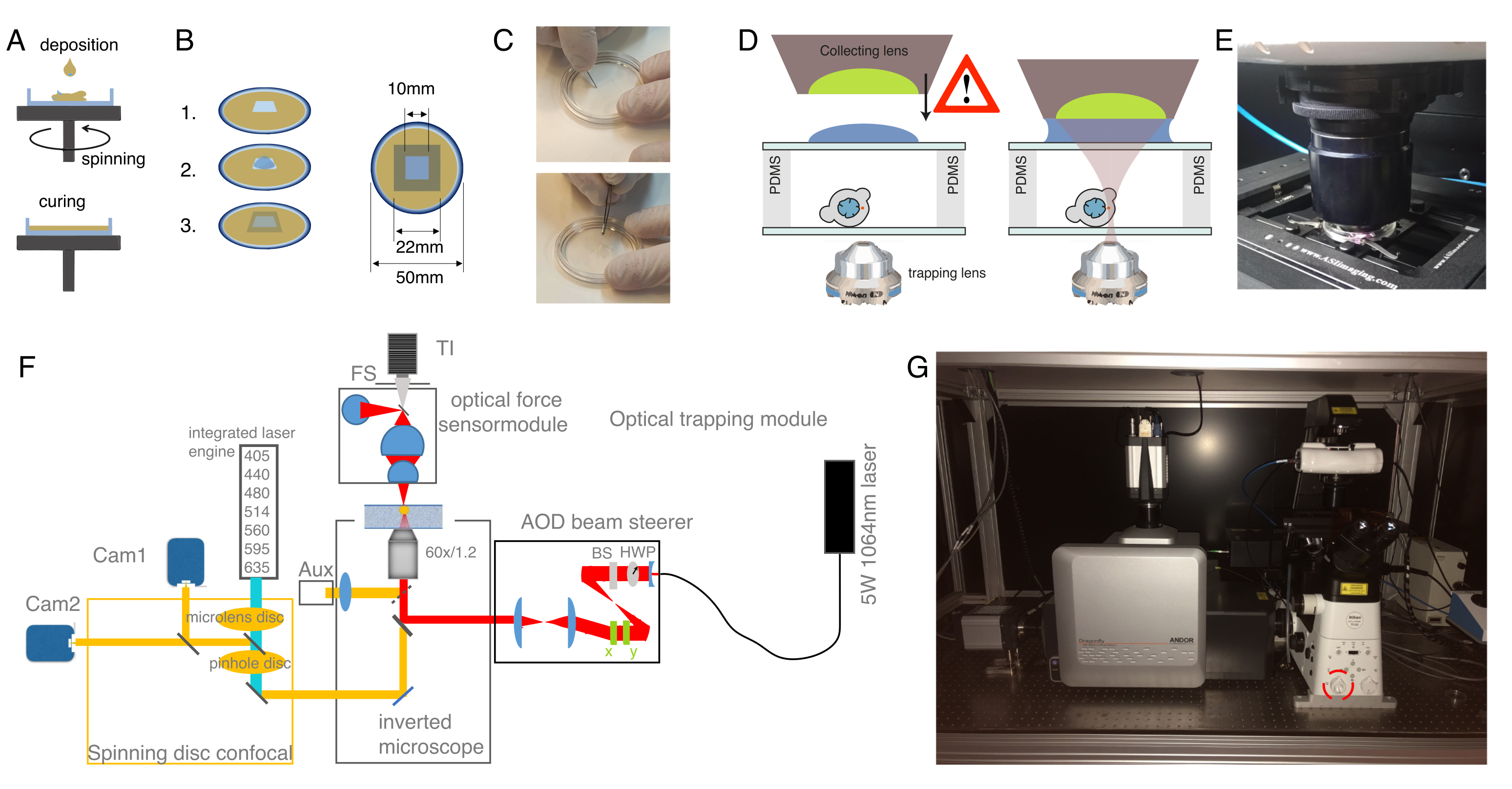
Figura 3: Elección de las muestras y parámetros correctos. (A) Imagen representativa de una célula madre progenitora de pez cebra aislada con una sola microesfera colocada lo suficientemente cerca del núcleo para realizar el experimento de indentación. Barra de escala = 10 μm. (B) Trayectoria de trampa ejemplar; profundidad de indentación 5 μm; velocidad de indentación = 5 μm/s; tiempo de relajación 10 s. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

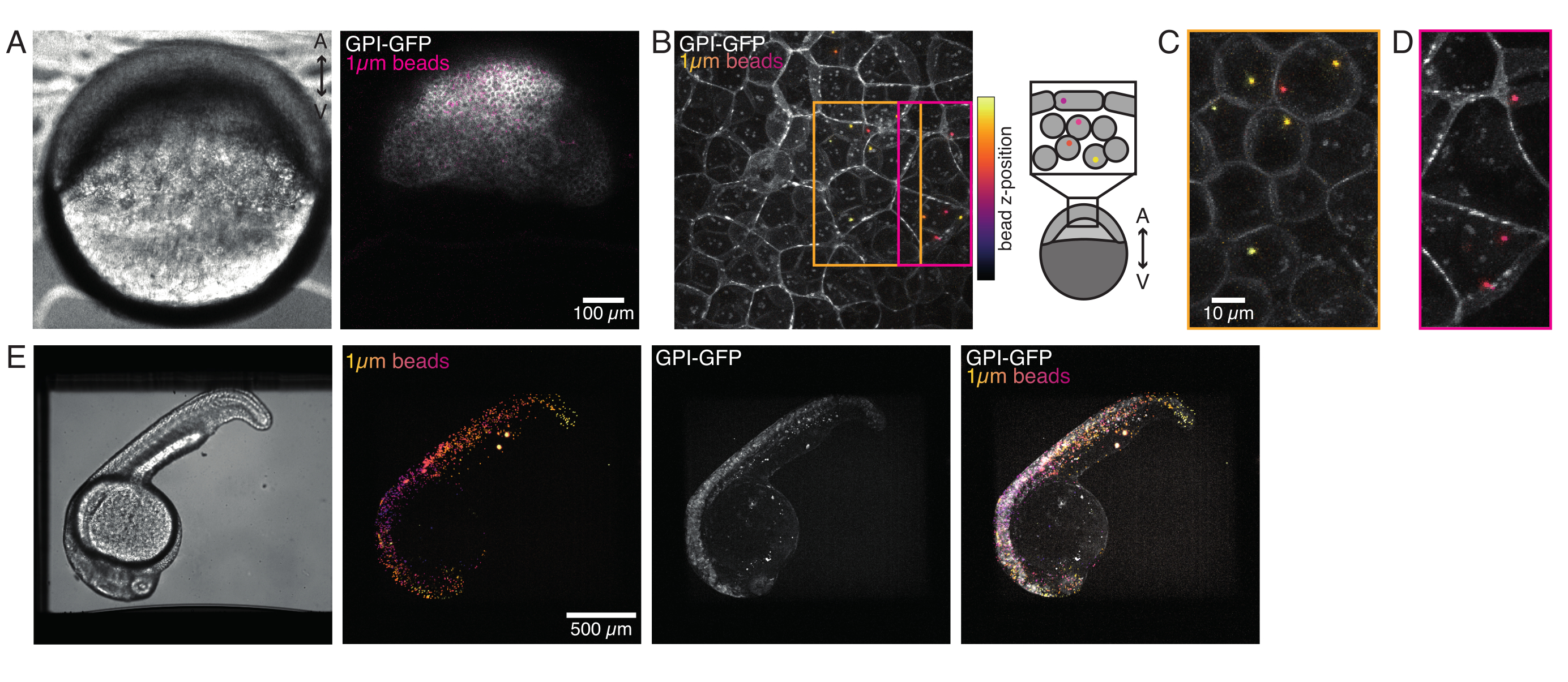
Figura 4: Localización de microperlas dentro de embriones de pez cebra durante el desarrollo. Se inyectan 0,5 nL de perlas fluorescentes rojas de 1 μm junto con ARNm GPI-GFP (100 pg/embrión, membrana plasmática) en embriones WT para visualizar las localizaciones de perlas. (A-D) Distribución de la microesfera 5 h después de la inyección dentro de un embrión montado en agarosa al 0,75%. (A) Imagen de campo brillante y fluorescencia. Las perlas se dispersan homogéneamente a través del tejido embrionario como se ve en una micrografía confocal. (B) Proyección máxima de la pila z de fluorescencia confocal. Las cuentas están codificadas por colores de púrpura a amarillo de acuerdo con su posición z en la pila de imágenes. Púrpura / magenta corresponde a la mayoría de las cuentas / células externas (EVL; capa envolvente epitelial; o células madre progenitoras ubicadas cerca de la superficie EVL), amarillo corresponde a cuentas internas (células profundas progenitoras), como se muestra en el boceto de la derecha. (C) Corte y proyección máxima de una subpila de (B) correspondiente a la región en la caja naranja: una gran fracción de celdas profundas contiene 1-2 cuentas. (D) Corte y proyección máxima de una subpila de (B) correspondiente a la caja magenta: algunas celdas EVL contienen 1-2 cuentas. (E) Imagen de campo brillante y proyección máxima de una pila z de un embrión de 24 hpf montado en agarosa al 0,75% y anestesiado con tricaína. Los embriones fueron preincubados con tricaína durante 15 min. De izquierda a derecha: microesferas (1 μm de diámetro), GPI-GFP y superposición de imágenes. Las cuentas se distribuyen por todo el cuerpo del embrión. Dimensión de la barra de escala indicada en cada panel. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 5: Células madre progenitoras aisladas de pez cebra con diferente etiquetado. (A) Imagen de microscopía de luz de transmisión de células de suspensión con 1 (arriba) o 2 (abajo) perlas inyectadas. Las flechas cian apuntan a las cuentas. (B) Imágenes confocales fluorescentes de células de suspensión con diferentes tinciones. Arriba a la izquierda: Lap2b-eGFP (membrana nuclear interna, 80 pg/embrión) y H2A-mCherry. Arriba a la derecha: GPI-GFP (membrana plasmática, 100 pg/embrión) y DNA-Hoechst (teñido como se describe en la sección 2). Abajo a la izquierda: MyI12.1-eGFP (línea transgénica) y DNA-Hoechst. Abajo a la derecha: Calbryte488 y DNA-Hoechst (teñidos como se describe en la sección 2). (C) Imagen de microscopía de luz de transmisión de celdas confinadas con 1 (arriba) o 2 (abajo) perlas inyectadas. Las flechas cian apuntan a las cuentas. Barras de escala = 10 μm. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

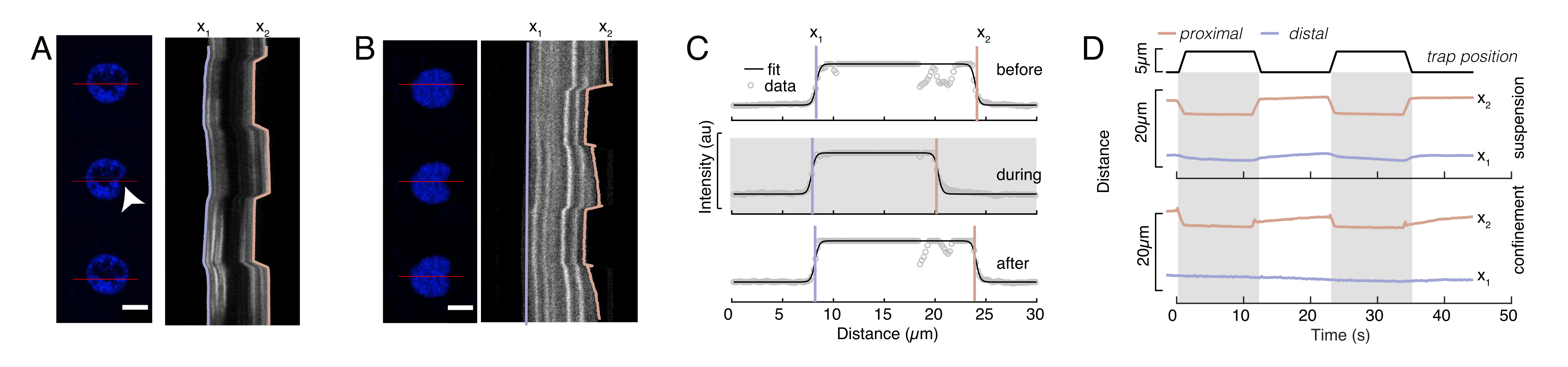
Figura 6: Estimación de la deformación nuclear a partir de películas de discos giratorios. (A, B) Lapso de tiempo de un experimento de indentación del núcleo en (A) una célula suspendida y (B) una célula confinada. Barra de escala 10 μm. Las instantáneas representativas de un núcleo marcado con Hoechst se muestran 5 s antes, durante y 5 s después de la sangría con una microesfera atrapada ópticamente (punta de flecha blanca). Kymographs a lo largo del segmento de sangría (línea roja, panel derecho). x1 y x2 son los límites distales y proximales (cerca de la perla) del núcleo durante el experimento de indentación extraído del ajuste del perfil de intensidad a la Ecuación 1. (C) Perfiles de intensidad a lo largo del segmento de sangría para tres fotogramas diferentes (antes, durante y después de la sangría) y ajustados a la Ecuación 1 para evaluar las posiciones distal, x1 y proximal, x2, de los bordes del núcleo. (D) Trayectorias representativas de x1(t) en azul y x2(t) en ámbar durante un experimento de indentación de células suspendidas y confinadas (10 μm). Las áreas sombreadas indican la sangría, la distancia entre x1 y x2 indica el diámetro del núcleo. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 7: Procesamiento de señales de fuerza. (A) Esquema de una microesfera atrapada ópticamente que deforma el núcleo celular tras la sangría. La membrana nuclear y las fuerzas ópticas están indicadas por las flechas negras. El cambio en el momento del haz se indica mediante la flecha verde Puchero. (B) Trayectoria de la trampa (arriba) y fuerza (abajo) experimentada por la microesfera atrapada ópticamente durante un experimento repetido de indentación nuclear. (C) Decaimiento de la relajación de la fuerza después del pico de fuerza en la profundidad máxima de indentación. El recuadro muestra un esquema de sólido lineal estándar cuya dinámica se aproxima a las observaciones fenomenológicas aquí. (D) Izquierda: logaritmo de la fuerza normalizada versus tiempo. Las áreas sombreadas indican la parte de datos utilizada para ajustarse a la doble desintegración exponencial (líneas rojas). Derecha: logaritmo de la fuerza normalizada versus el logaritmo del tiempo. El área sombreada indica la parte de datos utilizada para ajustarse a la ley de potencia. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 8: Rutina de indentación de fuerza con desplazamientos de trampa triangular. (A) Trayectoria representativa de x1(t) en azul y x2(t) en ámbar durante un experimento de indentación triangular tomado en una celda en altura de confinamiento de 10 μm. Arriba: Posición de trampa. Medio: Análisis de la forma del núcleo. La distancia entre x1 y x2 indica el diámetro del núcleo. Abajo: Señal de fuerza. (B) Fuerza vs posición de trampa durante ocho sangrías consecutivas. (C) Evolución de la disipación, derivada de la histéresis entre la parte de aproximación y retirada de la curva f-d, del núcleo para cada evento de indentación posterior. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 9. Propiedades nucleares de las células en suspensión (superficie adhesiva) y confinamiento de rutinas trapezoidales. (A) Área proyectada del núcleo a partir de células en suspensión y bajo confinamiento de 10 μm. La barra negra representa la mediana. B) Rigidez nuclear de las celdas en suspensión y bajo confinamiento. La barra negra representa la mediana. Valores P derivados de la prueba de Kruskal-Wallis utilizando MatLab. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
Tabla suplementaria 1: Trayectoria trapezoidal definida por el software de pinzas ópticas. La primera (segunda) fila es la distancia x (y) a la que se desplazará linealmente la trampa. En la tercera fila, la duración de un paso dado se establece en segundos. Esta trayectoria está compuesta por siete puntos y corresponde al trapecio cargado dos veces contra el núcleo de la Figura 7B. Haga clic aquí para descargar esta tabla.
Tabla suplementaria 2: Trayectoria triangular definida por el software de pinzas ópticas. Análogamente a la Tabla 2, esta trayectoria se compone de 16 puntos, correspondientes a ocho eventos de indentación a una profundidad de 5 μm y una velocidad de 2,5 μm/s. Haga clic aquí para descargar esta Tabla.
Tabla suplementaria 3: Parámetros de ajuste para los datos de la Figura 7. IG: conjetura inicial. Haga clic aquí para descargar esta tabla.
Figura suplementaria S1: Alineación del sensor de fuerza óptica y compensación de línea de base de momento. (A) Parada de campo fotografiada en la cámara auxiliar (AUX, Figura 2) a través de la lente Bertrand. Una burbuja de aire aparece visible en el aceite de inmersión, que no es visible a través del ocular. (B) Ruta óptica limpia. Para una alineación precisa, abra el tope de campo y haga que coincida con el cono de luz NA = 1.2. (C) Imagen del plano de muestra. El cuadrado rojo indica el área de trabajo de OT. Barra de escala: 20 μm. (D) Potencia de la trampa medida a través del FOV, a lo largo de flechas dobles blancas indicadas en C. En rojo, trapea la variación de potencia cuando no se aplica ninguna corrección. En azul, la potencia de la trampa se corrige en todo el campo de visión. (E) Componente X de la línea de base de momento a lo largo del mismo rango. En rojo, traza no corregida. En azul, traza corregida para la potencia de la trampa. En verde, traza corregida para la línea de base de impulso utilizando Global Offset Compensation en el software del fabricante. F) Igual que en E, para el componente Y. Tenga en cuenta que en condiciones normales de funcionamiento, los componentes sombreados se utilizan para la mecánica y las mediciones de fuerza, por ejemplo, el componente de fuerza x durante el movimiento a lo largo de la coordenada x y el componente de fuerza y durante el movimiento a lo largo del eje y. Después de implementar todas las correcciones, se obtiene un ruido RMSD de <0.5 pN. Haga clic aquí para descargar este archivo.
Figura suplementaria S2: Una rutina fallida debido a trampas débiles. (A) Kymograph que muestra una hendidura del núcleo de una rutina fallida. Solo las deformaciones cortas y transitorias son visibles debido a un escape de la cuenta de la trampa. Es importante destacar que el láser de captura todavía se mueve sin cuenta para completar la trayectoria predefinida (línea punteada verde). Barra de escala = 10 μm. (B) Superior: Posición de la trampa frente al tiempo. Medio: Resultado de seguimiento de borde del borde del núcleo proximal y distal con sangría. Tenga en cuenta que el borde distal no se mueve sin la hendidura como se observa comúnmente para las rutinas completadas en células aisladas en sustratos adhesivos. Abajo: Fuerza versus tiempo mostrando la pérdida de la microesfera indicada por una reducción en el ruido térmico y una caída repentina a fuerza cero. Haga clic aquí para descargar este archivo.
Figura suplementaria S3: Supervivencia de los embriones inyectados. Los embriones inyectados con perlas de 1 μm y 100 pg/embrión de ARNm a las concentraciones descritas en el protocolo se compararon con embriones no inyectados y no muestran diferencias significativas 24 h después de la fertilización. La media y la desviación estándar de tres experimentos independientes con N > 21 embriones por condición para cada experimento. Haga clic aquí para descargar este archivo.
Discusión
En este protocolo, describimos un método único para interrogar las propiedades mecánicas del núcleo celular dentro de las células vivas. A diferencia de otras técnicas de espectroscopia de fuerza, el atrapamiento óptico no invasivo nos permitió desacoplar la contribución de la membrana celular y el citoesqueleto de la rigidez nuclear celular. Es importante destacar que la micromanipulación óptica es compatible con la microscopía multimodal, lo que permitirá al experimentador estudiar diferentes procesos involucrados en la mecanobiología nuclear celular. Como resultado representativo, utilizamos la tinción de DNA-Hoechst para medir la deformación del núcleo tras la sangría realizada por fuerzas del orden de varios cientos de picoNewton.
Aplicaciones potenciales de nuestro método más allá de los ejemplos descritos en este protocolo
La posibilidad de extraer información mecánica cuantitativa de las mediciones dentro de las células vivas sin perturbaciones externas permite una gran cantidad de oportunidades sin precedentes que apenas están comenzando a explorarse. Así, el protocolo presentado de nuestra plataforma de micromanipulación óptica se puede extender a experimentos más complejos con gran versatilidad. Los deflectores acústico-ópticos (AOD) pueden generar múltiples trampas ópticas para mediciones de fuerza síncrona en diferentes ubicaciones celulares, así como también pueden usarse para microrreología activa en un amplio rango de frecuencias51,61. Como se ha mencionado, la respuesta de la fuerza a la sangría puede superar la fuerza de captura máxima, lo que lleva a un escape de la cuenta de la trampa óptica. En este caso, se puede configurar una retroalimentación de fuerza con el AOD para sujetar la fuerza óptica. Con todo, múltiples enfoques microrreológicos, como la relajación del estrés descrita en este protocolo, pero también la microrreología activa o el cumplimiento de la fluencia, se pueden obtener experimentalmente con esta plataforma y analizarse a fondo mediante nuevos paquetes de software61,62,63,64,65 . Además, la aplicación de fuerzas no se limita al núcleo, sino que en principio podría llevarse a cabo para medir diversas estructuras intracelulares y en tejidos complejos como se ha demostrado para atrapar glóbulos rojos que fluyen dentro de vasos sanguíneos intactos66,67 o atrapar y deformar cloroplastos y mitocondrias68 . La calibración del momento de la luz es independiente de la forma y el tamaño del objeto atrapado, lo que permite mediciones directas de la fuerza en cualquier sonda de fuerza con forma arbitraria38,39. El uso de microesferas inyectadas nos permitió aplicar altas fuerzas sobre el núcleo con una potencia láser relativamente baja en comparación con la manipulación directa de estructuras celulares69,70,71. Sin embargo, dada una diferencia de índice de refracción lo suficientemente alta, no es necesaria una sonda de fuerza aplicada externamente y los orgánulos intracelulares pueden manipularse directamente sin perlas inyectadas (observaciones no publicadas y referencia70).
Posibles modificaciones de nuestro método para ampliar las aplicaciones
Se pueden inyectar diferentes tamaños de microperlas dependiendo del experimento, pero se deben realizar los controles relativos. Por ejemplo, para estudiar las células en etapas posteriores se pueden inyectar perlas más pequeñas. Esto reducirá la fuerza máxima que puede ejercer la trampa óptica (como se muestra en la referencia55). Se pueden inyectar perlas más grandes para ejercer fuerzas más altas, pero estas pueden afectar el desarrollo embrionario dependiendo de su tamaño o etapa de interés. En experimentos en los que la inyección de microperlas no es una opción, varios orgánulos que muestran diferencias en los índices de refracción en comparación con el del citoplasma aún pueden manipularse ópticamente, dando lugar a fuerzas ópticas medibles a partir de los cambios de momento de la luz42. Como se mencionó anteriormente, estos métodos han sido empleados por Bambardekar et al. para deformar las uniones célula-célula en el embrión de Drosophila70. Asimismo, el núcleo de la célula tiene un índice de refracción inferior al medio circundante44, lo que permite una hendidura libre de perlas (observaciones inéditas y referencia72) aunque con una menor resistencia de atrapamiento. Por lo tanto, el núcleo no puede ser atrapado fácilmente y escapa de la trampa.
El espaciador PDMS con recubrimiento de espín se fabrica a través de un método conveniente y rápido, pero puede estar fuera del alcance de los laboratorios sin acceso a una instalación de micro / nanofabricación o laboratorios de ingeniería. Por lo tanto, el espaciador se puede ensamblar fácilmente a partir de cinta de laboratorio o parafilm (paso 4). El protocolo también se puede adaptar mediante la fabricación de canales microfluídicos que automatizan la entrega de células individuales en pozos de medición predefinidos o en una cámara con una altura definida para estimar el efecto de confinamiento dentro de la misma muestra. Sin embargo, dichos dispositivos microfluídicos deberán diseñarse de manera que se ajusten al espacio entre el objetivo del microscopio y la lente colectora del sensor de fuerza óptica, de alrededor de 2 mm (véase el paso 3). Tenga en cuenta que el sensor de fuerza óptica debe colocarse a la altura adecuada para que ninguna aberración óptica por desenfoque afecte la medición del momento del fotón.
Otras modificaciones podrían incluir el cambio de reporteros biológicos. Encontramos que la fluorescencia de Hoechst sangra espectralmente en el canal GFP y, por lo tanto, favorecemos la combinación con histona marcada con mCherry como marcador nuclear para la medición simultánea en dos canales fluorescentes. Alternativamente, la deformación nuclear se puede rastrear fácilmente con una etiqueta dirigida a la membrana nuclear interna, como Lap2b-GFP (Figura 2).
La indentación en el núcleo celular fue del orden de 2-3 micras, que pudimos medir con precisión mediante el análisis de imágenes de microscopía confocal de disco giratorio limitado por difracción. Para el caso de núcleos más rígidos o fuerzas más pequeñas, la hendidura será apenas medible utilizando este enfoque. Sin embargo, las pinzas ópticas calibradas por fuerza absoluta también se pueden calibrar para mediciones de posición de la cuenta atrapada in situ utilizando interferometría BFP con precisión nanométrica51. Utilizando este enfoque, la señal de voltaje y el sensor de fuerza óptica se pueden traducir en la posición de la sonda atrapada a través del parámetro β [nm/V], mientras que el parámetro invariante α [pN/V] produce valores de fuerza a través de la calibración de momento de luz antes mencionada41 (ver más abajo para más detalles).
Solución de problemas
Encontramos que los siguientes desafíos podrían ocurrir durante el experimento:
No se forma ninguna trampa estable y la microesfera se escapa fácilmente
Cualquier suciedad en el objetivo del microscopio o un collar de corrección desalineado podría conducir a una falla de una trampa estable. Si no se encuentra una solución inmediata, mida la función de dispersión de puntos de la lente del objetivo. Si la muestra de interés está profundamente dentro de un tejido ópticamente denso, el foco láser puede experimentar aberraciones ópticas graves que conducen a una captura inestable (este efecto suele ser insignificante en células aisladas, pero se hace más evidente en tejidos más gruesos). Para una alta rigidez, la fuerza de restauración del núcleo podría exceder la fuerza de escape de la trampa, de modo que la microesfera se pierda y la rutina de hendidura falle. Inicialmente, el borde de la membrana nuclear proximal a la trampa óptica apenas se sangra (Figura S2A). Cuando esto ocurre, el láser de captura ya no se ve afectado por la fuerza y el movimiento browniano, lo que conduce a una caída de fuerza a cero y una disminución del ruido de la señal (Figura S2B). En caso de que esto suceda, la potencia del láser se puede aumentar para tener una trampa más fuerte, la amplitud de la trayectoria trapezoidal que empuja la cuenta hacia el núcleo se puede reducir, o la posición inicial de la microperla atrapada se puede establecer más lejos del núcleo.
La célula se mueve durante la estimulación
Si las células no están suficientemente unidas, la trampa de gradiente óptico moverá las células mientras realiza la rutina de indentación intracelular, de modo que las fuerzas y la mecánica subyacente del núcleo sean artefactuales. Para evitar el desplazamiento de toda la célula, recomendamos aumentar la concentración de moléculas de adhesión celular en la superficie, por ejemplo, ConA.
Compensación de impulso inicial
Si no se dispone de una rutina inicial de compensación de momento en la plataforma de OT (paso 6.5), es necesario corregir una señal de referencia artificial e independiente de la fuerza. Esto es visible como una pendiente en la curva de fuerza incluso sin cuentas atrapadas (Figura S1E). Para hacer la corrección, la misma trayectoria debe realizarse sin una cuenta, fuera de la celda exactamente en la misma posición. Para ello, aleje la celda de la trampa utilizando el control de escenario. Como referencia, el desplazamiento de fuerza cambia 5 pN a través del FOV a 200 mW en nuestro sistema; por lo tanto, se vuelve insignificante para trayectorias cortas. Alternativamente, se puede utilizar una etapa de piezoescaneo para mover las células de la muestra, dejando constante la posición del láser.
Pasos críticos del protocolo presentado
Las microesferas deben inyectarse en la etapa correcta de 1 célula para garantizar la distribución máxima sobre el embrión. Las perlas no deben ser fluorescentes para que no se filtre luz en los canales fluorescentes utilizados para la obtención de imágenes. Por ejemplo, incluso las cuentas rojas fluorescentes típicas son claramente visibles en el canal azul utilizado para obtener imágenes del núcleo celular después de la tinción de Hoechst debido a su brillo (excitación: 405 nm; emisión: 445 nm). La fijación estable de la célula al sustrato es fundamental para evitar el desplazamiento lateral durante la rutina de hendidura. Si la célula se mueve durante la rutina, las fuerzas se subestiman. Si esto sucede con frecuencia, optimice el protocolo de conexión. Para las células de cultivo de tejidos, otras proteínas de adhesión celular, como la fibronectina, el colágeno o la poli-L-lisina conducen a una unión satisfactoria (observaciones no publicadas). Durante el confinamiento, las celdas son sometidas a un estrés mecánico repentino y severo. Esto puede causar daño a las células y, con frecuencia, el experimentador se encontrará con células rotas si el procedimiento no se lleva a cabo con cuidado. Además, si la altura de confinamiento es demasiado pequeña, todas las células sufrirán roturas de envoltura nuclear o daños irreversibles. Para mitigar estos, baje el cubrebocas superior más lentamente y / o aumente el espacio entre el deslizamiento de cubierta.
Limitaciones de la técnica y sugerencias para superarlas
Una clara limitación de la técnica es la penetración de la luz láser en secciones profundas del tejido, lo que conduce a aberraciones y atrapamientos inestables. Por lo tanto, un límite inferior de profundidad de penetración depende de la claridad de la muestra, la corrección de aberración que se puede emplear73 y la potencia láser aplicada. Debe tenerse en cuenta que una mayor potencia láser conduce a la excitación térmica de la muestra en las cercanías de la microesfera. Sin embargo, el calentamiento de la muestra originado por el punto láser de longitud de onda de 1064 nm se minimiza para evitar un estrés plausible relacionado con el calor en nuestras muestras biológicas74.
Otra limitación es la fuerza máxima que se puede medir. A pesar de que la detección directa de momento de luz permite mediciones de fuerza mucho más allá del régimen de respuesta lineal de la trampa óptica40,41, la fuerza máxima aplicada es del orden de unos pocos cientos de picoNewtons. Esto está limitado por la potencia del láser y el umbral de daño consecuente del material biológico blando y las diferencias del índice de refracción, que normalmente no son mayores de 0.1 o 0.344. Se han propuesto varios métodos para aumentar el límite de detección de fuerza, por ejemplo, utilizando luz estructurada75, microesferas recubiertas antirreflectantes76, partículas de alto índice de refracción77 o puntos cuánticos altamente dopados78.
Los OT se pueden utilizar para mediciones de posición a escala nanométrica a través de interferometría BFP, de modo que la posición de la cuenta dentro de la trampa es Δx = β Sx, donde Sx es la señal de voltaje del sensor, y β [μm / V] se puede calibrar sobre la marcha siguiendo diferentes protocolos35,54. Para un sensor de fuerza óptica, se puede demostrar que el factor de conversión invariante de voltaje a fuerza α [pN / V] se relaciona directamente con β y la rigidez de la trampa, k [pN / μm], a través de α = kβ 37) En experimentos con desplazamientos de cuentas que son demasiado pequeños para ser detectados a partir de imágenes ópticas, esta estrategia se puede utilizar para complementar las mediciones de fuerza con la detección de posición pequeña. Un ejemplo es la aplicación de las rutinas experimentales presentadas aquí sobre núcleos muy rígidos, para los cuales las fuerzas a potencias láser razonables (200-500 mW) no son suficientes para inducir valores de indentación lo suficientemente grandes. En ese caso, la perla debe ponerse en contacto con el núcleo y la rigidez de atrapamiento debe calibrarse antes de la medición (paso 8.6). La sangría d del núcleo en función de la fuerza puede determinarse indirectamente como:
d = xtrap - F/k
donde xtrap es la posición de la trampa. A diferencia del factor de momento de luz invariante α [pN/V], el factor β [μm/V] debe calibrarse antes de cada experimento, ya que depende de muchas variables locales que determinan la dinámica de atrapamiento, como el tamaño de partícula, el tamaño del punto de trampa óptica y los índices de refracción relativos.
Divulgaciones
Los autores no tienen nada que revelar.
Agradecimientos
MK agradece el apoyo financiero del Ministerio de Economía y Competitividad a través del Plan Nacional (PGC2018-097882-A-I00), FEDER (EQC2018-005048-P), programa Severo Ochoa para Centros de Excelencia en I+D (CEX2019-000910-S; RYC-2016-21062), de la Fundació Privada Cellex, Fundació Mir-Puig, y de la Generalitat de Catalunya a través del programa CERCA e Research (2017 SGR 1012), además de la financiación a través de ERC (MechanoSystems) y HFSP (CDA00023/2018). V.R. agradece el apoyo del Ministerio de Ciencia e Innovación a la asociación EMBL, el Centro de Excelencia Severo Ochoa, el Plan Nacional del MINECO (BFU2017-86296-P, PID2020-117011GB-I00) y la Generalitat de Catalunya (CERCA). V.V. reconoce el apoyo del Programa de Doctorado ICFOstepstone financiado por el programa de investigación e innovación Horizonte 2020 de la Unión Europea en el marco del acuerdo de subvención Marie Skłodowska-Curie 665884. Agradecemos a Arnau Farré por la lectura crítica del manuscrito; Maria Marsal por la ayuda con la imagen y el montaje del embrión de 24 hpf y; Senda Jiménez-Delgado para apoyo con micronyecciones de pez cebra.
Materiales
| Name | Company | Catalog Number | Comments |
| #1.5 22 mm cover glasses | Ted Pella | 260148 | |
| #1.5 22x60 mm Coverglasses | Ted Pella | 260152 | |
| #1.5H glass bottom dishes | Willco | GWST-5040 | |
| 10-um beads | Supelco | 72986 | |
| 1-mm glass capillaries | Harvard Apparatus | 30-0020 GC100F-15 | |
| 1-um polystyrene microbeads | Sigma | 89904 | |
| 1-um red-fluorescent beads | ThermoFisher | F8816 | |
| Agar | ThermoFisher | 16500500 | |
| Aqcuisition cameras sCMOS | Andor | Sona-4BV11-UNI | |
| Auxiliary camera (Figure 3, AUX) | Blackfly, FLIR | BFS-U3-200S6M-C | |
| Calbryte 520 | AAT Bioquest | 520 AM | |
| Centrifuge | Eppendorf | 5453000011 | |
| Concanavalin A | Sigma | C5275 | |
| DMEM | Sigma | D2906 | |
| DNA-Hoechst | ThermoFisher | 33342 | |
| Double scotch tape | Biesse Adesivi | ||
| E3 | 5 mM NaCl. 0.17 mM KCl. 0.33 mM CaCl2. 0.33 mM MgSO4 | ||
| Eclipse Ti2 | Nikon | ||
| Forceps | Fine Science Tools | 11252-20 | |
| GPI-GFP | |||
| H2A-mCh | |||
| Image acquisition software | Fusion-Andor | ||
| Immersion Oil | Cargille | Type B: 16484 | |
| IR protection googles | Thorlabs | LG1 | |
| Lap2b-eGFP | |||
| Micro loader pipette | Eppendorf | GELoader | |
| Microinjector | World Precision Instruments | SYS-PV820 | |
| MicroManager 2.0 | |||
| Micromiter slide | ID5243 GXMGRAT-5 5mm/100 divisions | ||
| Mineral oil | Sigma | M3616 | |
| Motorized stage | ASI | ||
| Needle puller | Sutter instrument Co. | Model P-97 | |
| Optical tweezers platform | Impetux Optics | Sensocell | |
| OTs software (LightAce) | Impetux Optics | ||
| PDMS | Sigma | Sylgard 184 | |
| PDMS Curing agent | Sigma | Sylgard 184 | |
| Post processing software (Matlab) | Mathworks | ||
| RNAse free water | Thermofisher | AM9937 | |
| Short-pass dichroic mirror (Figure 3, IR-F) | Semrock | FF01-950/SP-25 | |
| Spin-coater | Specialty Coating Systems | Spincoat G3P-8 | |
| Spinning-disk confocal microscope | Andor | DragonFly 502 | |
| Stereomicroscope | Leica M80 | ||
| Triangular microinjection mold | Adaptive Science Tools | TU1 | |
| Universal oven | Memmert | UNB 200 | |
| Water immersion objective | Nikon | MRD07602 |
Referencias
- Chan, C. J., Heisenberg, C. P., Hiiragi, T. Coordination of Morphogenesis and Cell-Fate Specification in Development. Current Biology. 27 (18), 1024-1035 (2017).
- Heller, E., Fuchs, E. Tissue patterning and cellular mechanics. Journal of Cell Biology. 211 (2), 219-231 (2015).
- Heisenberg, C. P., Bellaïche, Y. Forces in tissue morphogenesis and patterning. Cell. 153 (5), 948-962 (2013).
- Petridou, N. I., Spiró, Z., Heisenberg, C. P. Multiscale force sensing in development. Nature Cell Biology. 19 (6), 581-588 (2017).
- Krieg, M., et al. Tensile forces govern germ-layer organization in zebrafish. Nature Cell Biology. 10 (4), 429-436 (2008).
- Ruprecht, V., et al. Cortical contractility triggers a stochastic switch to fast amoeboid cell motility. Cell. 160 (4), 673-685 (2015).
- Shellard, A., Mayor, R. Supracellular migration - Beyond collective cell migration. Journal of Cell Science. 132 (8), (2019).
- Mongera, A., et al. A fluid-to-solid jamming transition underlies vertebrate body axis elongation. Nature. 561 (7723), 401-405 (2018).
- Atia, L., et al. Geometric constraints during epithelial jamming. Nature Physics. 14 (6), 613-620 (2018).
- Turlier, H., Maître, J. -. L. Mechanics of tissue compaction. Seminars in Cell & Developmental Biology. 47-48, 110-117 (2015).
- Ladoux, B., Mège, R. M. Mechanobiology of collective cell behaviours. Nature Reviews Molecular Cell Biology. 18 (12), 743-757 (2017).
- Venturini, V., et al. The nucleus measures shape changes for cellular proprioception to control dynamic cell behavior. Science. 370 (6514), (2020).
- Charras, G., Sahai, E. Physical influences of the extracellular environment on cell migration. Nature Reviews Molecular Cell Biology. 15 (12), 813-824 (2014).
- Kirby, T. J., Lammerding, J. Emerging views of the nucleus as a cellular mechanosensor. Nature Cell Biology. 20 (4), 373-381 (2018).
- Lee, H. P., et al. The nuclear piston activates mechanosensitive ion channels to generate cell migration paths in confining microenvironments. Science Advances. 7 (2), (2021).
- Friedl, P., Wolf, K., Lammerding, J. Nuclear mechanics during cell migration. Current Opinion in Cell Biology. 23 (1), 55-64 (2011).
- Versaevel, M., Riaz, M., Grevesse, T., Gabriele, S. Cell confinement: Putting the squeeze on the nucleus. Soft Matter. 9 (29), 6665-6676 (2013).
- Zuela-Sopilniak, N., et al. Measuring nucleus mechanics within a living multicellular organism: Physical decoupling and attenuated recovery rate are physiological protective mechanisms of the cell nucleus under high mechanical load. Molecular Biology of the Cell. 31 (17), 1943-1950 (2020).
- Kim, D. H., Wirtz, D. Cytoskeletal tension induces the polarized architecture of the nucleus. Biomaterials. 48, 161-172 (2015).
- Lomakin, A. J., et al. The nucleus acts as a ruler tailoring cell responses to spatial constraints. Science. 370 (6514), (2020).
- Hampoelz, B., et al. Microtubule-induced nuclear envelope fluctuations control chromatin dynamics in Drosophila embryos. Development. 138 (16), 3377-3386 (2011).
- Heo, S. J., et al. Differentiation alters stem cell nuclear architecture, mechanics, and mechano-sensitivity. eLife. 5, 1-21 (2016).
- Cosgrove, B. D., et al. Nuclear envelope wrinkling predicts mesenchymal progenitor cell mechano-response in 2D and 3D microenvironments. Biomaterials. 270, 120662 (2021).
- Liu, H., et al. In situ mechanical characterization of the cell nucleus by atomic force microscopy. ACS Nano. 8 (4), 3821-3828 (2014).
- Hobson, C. M., et al. Correlating nuclear morphology and external force with combined atomic force microscopy and light sheet imaging separates roles of chromatin and lamin A/C in nuclear mechanics. Molecular Biology of the Cell. 31 (16), 1788-1801 (2020).
- Pajerowski, J. D., Dahl, K. N., Zhong, F. L., Sammak, P. J., Discher, D. E. Physical plasticity of the nucleus in stem cell differentiation. Proceedings of the National Academy of Sciences of the United States of America. 104 (40), 15619-15624 (2007).
- Rowat, A. C., Lammerding, J., Ipsen, J. H. Mechanical properties of the cell nucleus and the effect of emerin deficiency. Biophysical Journal. 91 (12), 4649-4664 (2006).
- Davidson, P. M., et al. High-throughput microfluidic micropipette aspiration device to probe time-scale dependent nuclear mechanics in intact cells. Lab on a Chip. 19 (21), 3652-3663 (2019).
- Lombardi, M., Zwerger, M., Lammerding, J. Biophysical assays to probe the mechanical properties of the interphase cell nucleus: Substrate strain application and microneedle manipulation. Journal of Visualized Experiments: JoVE. (55), (2011).
- Luo, T., Mohan, K., Iglesias, P. A., Robinson, D. N. Molecular mechanisms of cellular mechanosensing. Nature Materials. 12 (11), 1064-1071 (2013).
- Dahl, K. N., Engler, A. J., Pajerowski, J. D., Discher, D. E. Power-law rheology of isolated nuclei with deformation mapping of nuclear substructures. Biophysical Journal. 89 (4), 2855-2864 (2005).
- Guilluy, C., et al. Isolated nuclei adapt to force and reveal a mechanotransduction pathway in the nucleus. Nature Cell Biology. 16 (4), 376-381 (2014).
- Bustamante, C. J., Wang, M. D. Optical tweezers in single-molecule biophysics. Nature Reviews Methods Primers. , 1-29 (2021).
- Svoboda, K., Block, S. M. Force and velocity measured for single kinesin molecules. Cell. 77 (5), 773-784 (1994).
- Berg-Sørensen, K., Flyvbjerg, H. Power spectrum analysis for optical tweezers. Review of Scientific Instruments. 75 (3), 594-612 (2004).
- Smith, S. B., Cui, Y., Bustamante, C. Optical-trap force transducer that operates by direct measurement of light momentum. Methods in Enzymology. 361 (1994), 134-162 (2003).
- Farré, A., Montes-Usategui, M. A force detection technique for single-beam optical traps based on direct measurement of light momentum changes. Optics Express. 18 (11), 11955 (2010).
- Català, F., Marsà, F., Montes-Usategui, M., Farré, A., Martín-Badosa, E. Extending calibration-free force measurements to optically-trapped rod-shaped samples. Scientific Reports. 7, 1-10 (2017).
- Bui, A. A. M., et al. Calibration of force detection for arbitrarily shaped particles in optical tweezers. Scientific Reports. 8 (1), 1-12 (2018).
- Farré, A., Marsà, F., Montes-Usategui, M. Beyond the hookean spring model: Direct measurement of optical forces through light momentum changes. Methods in Molecular Biology. 1486, (2017).
- Farré, A., Marsà, F., Montes-Usategui, M. Optimized back-focal-plane interferometry directly measures forces of optically trapped particles. Optics Express. 20 (11), 12270 (2012).
- Jun, Y., Tripathy, S. K., Narayanareddy, B. R. J., Mattson-Hoss, M. K., Gross, S. P. Calibration of optical tweezers for in vivo force measurements: How do different approaches compare. Biophysical Journal. 107 (6), 1474-1484 (2014).
- Mas, J., Farré, A., Sancho-Parramon, J., Martín-Badosa, E., Montes-Usategui, M. Force measurements with optical tweezers inside living cells. Optical Trapping and Optical Micromanipulation XI. 9164, (2014).
- Schürmann, M., Scholze, J., Müller, P., Guck, J., Chan, C. J. Cell nuclei have lower refractive index and mass density than cytoplasm. Journal of Biophotonics. 9 (10), 1068-1076 (2016).
- Rosen, J. N., Sweeney, M. F., Mably, J. D. Microinjection of zebrafish embryos to analyze gene function. Journal of Visualized Experiments: JoVE. (25), (2009).
- Westerfield, M. . The Zebrafish Book. A Guide for the Laboratory Use of Zebrafish. (Danio rerio), 5th Edition. , (2007).
- Schubert, R., et al. Assay for characterizing the recovery of vertebrate cells for adhesion measurements by single-cell force spectroscopy. FEBS Letters. 588 (19), 3639-3648 (2014).
- Koschwanez, J. H., Carlson, R. H., Meldrum, D. R. Thin PDMS films using long spin times or tert-butyl alcohol as a solvent. PLoS One. 4 (2), 2-6 (2009).
- Das, R., et al. Mechanical stretch inhibition sensitizes proprioceptors to compressive stresses. bioRxiv. , (2021).
- Chardès, C., Clement, R., Blanc, O., Lenne, P. F. Probing cell mechanics with bead-free optical tweezers in the drosophila embryo. Journal of Visualized Experiments: JoVE. (141), (2018).
- Staunton, J. R., Blehm, B., Devine, A., Tanner, K. In situ calibration of position detection in an optical trap for active microrheology in viscous materials. Optics Express. 25 (3), 1746 (2017).
- Bola, R., Treptow, D., Marzoa, A., Montes-Usategui, M., Martin-Badosa, E. Acousto-holographic optical tweezers. Optics Letters. 45 (10), 2938-2941 (2020).
- Thalhammer, G., Obmascher, L., Ritsch-Marte, M. Direct measurement of axial optical forces. Optics Express. 23 (5), 6112 (2015).
- Vermeulen, K. C., et al. Calibrating bead displacements in optical tweezers using acousto-optic deflectors. Review of Scientific Instruments. 77 (1), 1-6 (2006).
- Dzementsei, A., Barooji, Y. F., Ober, E. A., Oddershede, L. B. Foregut organ progenitors and their niche display distinct viscoelastic properties in vivo during early morphogenesis stages. bioRxiv. , 1-35 (2021).
- Behrndt, M., et al. Forces driving epithelial spreading in zebrafish gastrulation. Science. 338 (6104), 257-260 (2012).
- Krieg, M., et al. Atomic force microscopy-based mechanobiology. Nature Reviews Physics. , (2018).
- A-Hassan, E., et al. Relative microelastic mapping of living cells by atomic force microscopy. Biophysical Journal. 74 (3), 1564-1578 (1998).
- Khalilgharibi, N., et al. Stress relaxation in epithelial monolayers is controlled by the actomyosin cortex. Nature Physics. 15, (2019).
- Crick, S. L., Yin, F. C. Assessing micromechanical properties of cells with atomic force microscopy: importance of the contact point. Biomechanics and Modeling in Mechanobiology. 6 (3), 199-210 (2007).
- Hurst, S., Vos, B. E., Betz, T. Intracellular softening and fluidification reveals a mechanical switch of cytoskeletal material contributions during division. bioRxiv. , 425761 (2021).
- Kaplan, J. L., Bonfanti, A., Kabla, A. RHEOS.jl - A Julia package for rheology data analysis. arXiv. 4, 1-5 (2020).
- Bonfanti, A., Kaplan, J. L., Charras, G., Kabla, A. Fractional viscoelastic models for power-law materials. Soft Matter. 16 (26), 6002-6020 (2020).
- Rivas-Barbosa, R., Escobedo-Sánchez, M. A., Tassieri, M., Laurati, M. i-Rheo: determining the linear viscoelastic moduli of colloidal dispersions from step-stress measurements. Physical Chemistry Chemical Physics: PCCP. 22 (7), 3839-3848 (2020).
- Tassieri, M., et al. i-Rheo: Measuring the materials' linear viscoelastic properties "in a step". Journal of Rheology. 60 (4), 649-660 (2016).
- Zhong, M. C., Wei, X. B., Zhou, J. H., Wang, Z. Q., Li, Y. M. Trapping red blood cells in living animals using optical tweezers. Nature Communications. 4, 1767-1768 (2013).
- Harlepp, S., Thalmann, F., Follain, G., Goetz, J. G. Hemodynamic forces can be accurately measured in vivo with optical tweezers. Molecular Biology of the Cell. 28 (23), 3252-3260 (2017).
- Bayoudh, S., Mehta, M., Rubinsztein-Dunlop, H., Heckenberg, N. R., Critchley, C. Micromanipulation of chloroplasts using optical tweezers. Journal of Microscopy. 203 (2), 214-222 (2001).
- Favre-Bulle, I. A., Stilgoe, A. B., Rubinsztein-Dunlop, H., Scott, E. K. Optical trapping of otoliths drives vestibular behaviours in larval zebrafish. Nature Communications. 8 (1), 630 (2017).
- Bambardekar, K., Clément, R., Blanc, O., Chardès, C., Lenne, P. F. Direct laser manipulation reveals the mechanics of cell contacts in vivo. Proceedings of the National Academy of Sciences of the United States of America. 112 (5), 1416-1421 (2015).
- Ferro, V., Chuai, M., McGloin, D., Weijer, C. J. Measurement of junctional tension in epithelial cells at the onset of primitive streak formation in the chick embryo via non-destructive optical manipulation. Development (Cambridge). 147 (3), (2020).
- Hörner, F., et al. Holographic optical tweezers-based in vivo manipulations in zebrafish embryos. Journal of Biophotonics. 10 (11), 1492-1501 (2017).
- Zhong, M. -. C., Wang, Z. -. Q., Li, Y. -. M. Aberration compensation for optical trapping of cells within living mice. Applied Optics. 56 (7), 1972 (2017).
- Català, F., Marsà, F., Montes-Usategui, M., Farré, A., Martín-Badosa, E. Influence of experimental parameters on the laser heating of an optical trap. Scientific Reports. 7 (1), 1-9 (2017).
- Taylor, M. A., Waleed, M., Stilgoe, A. B., Rubinsztein-Dunlop, H., Bowen, W. P. Enhanced optical trapping via structured scattering. Nature Photonics. 9 (10), 669-673 (2015).
- Bormuth, V., et al. Optical trapping of coated microspheres. Optics Express. 16 (18), 13831-13844 (2008).
- Sudhakar, S., et al. Germanium nanospheres for ultraresolution picotensiometry of kinesin motors. Science. 371 (6530), (2021).
- Shan, X., et al. Optical tweezers beyond refractive index mismatch using highly doped upconversion nanoparticles. Nature Nanotechnology. 16 (5), 531-537 (2021).
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoExplorar más artículos
This article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados