
Method Article
تحديد كينيسين-1 الشحنات عن طريق الميكروسكوب الإسفار
In This Article
Summary
هنا، ويرد على بروتوكول لتحديد كينيسين-1 الشحنات. ومتحولة بلا محركات من السلسلة الثقيلة كينيسين-1 (KIF5B) المجاميع في السيتوبلازم ويدفع تجميع الشحنات به. تم الكشف عن كل من المجاميع تحت المجهر مضان. يمكن استخدام استراتيجية مماثلة لتحديد الشحنات من البروتينات السيارات الأخرى.
Abstract
Fluorescence microscopy is employed to identify Kinesin-1 cargos. Recently, the heavy chain of Kinesin-1 (KIF5B) was shown to transport the nuclear transcription factor c-MYC for proteosomal degradation in the cytoplasm. The method described here involves the study of a motorless KIF5B mutant for fluorescence microscopy. The wild-type and motorless KIF5B proteins are tagged with the fluorescent protein tdTomato. The wild-type tdTomato-KIF5B appears homogenously in the cytoplasm, while the motorless tdTomato-KIF5B mutant forms aggregates in the cytoplasm. Aggregation of the motorless KIF5B mutant induces aggregation of its cargo c-MYC in the cytoplasm. Hence, this method provides a visual means to identify the cargos of Kinesin-1. A similar strategy can be utilized to identify cargos of other motor proteins.
Introduction
كينيسين-1 هو بروتين المحرك الذي يتوسط النقل تقدمي من الشحنات في 1،2. وهو heterotetramer اثنين من مفارز كينيسين ضوء سلسلة 1 (KLC1) واثنين من مفارز من سلاسل كينيسين الثقيلة (KHCs). KIF5B 1، شركة المملكة القابضة، ويحتوي على مجال السيارات في دورته N-المحطة، التي hydrolyzes اعبي التنس المحترفين وتحويل الطاقة الكيميائية إلى طاقة ميكانيكية لحركة على طول الأنابيب الدقيقة. تحتوي على منطقة C-محطة المجال dimerization التي تتفاعل مع KLC1، من شأنها أن تربط مع الشحنات. كينيسين-1 ينقل الشحنات مثل الحويصلات، العضيات ومن mRNAs 3،4. في الآونة الأخيرة، وقد تبين KIF5B لنقل عامل النسخ النووي ج-MYC لتدهور proteosomal في السيتوبلازم 5. استخدمت ثلاث منهجيات (المانع الكيميائية، سيرنا / shRNA ومتحولة السلبي المهيمن) لمنع كينيسين-1 وظيفة. أنهم جميعا الناجم عن تجميع ج-MYC في السيتوبلازم. لمنهجية الماضية، وج-MYC تتأثر فقط من قبل negat المهيمنإيف متحولة من KIF5B، ولكن ليس هذا البروتين KIF5A محرك آخر ذي صلة، مما يوحي بأن متحولة لا تمارس الآثار العامة على مكونات الخلايا (مثل اضطراب أنيبيب) أو على تجميع البروتين. متحولة السلبي السائد للKIF5B كما لم يؤثر عامل النسخ أخرى، مما يشير إلى أنها لا تمارس الآثار العامة على عوامل النسخ. بدلا من ذلك، فإنه يشير إلى أن متحولة السلبي السائد يمارس آثار محددة على الشحنات لها.
استخدام المسوخ السلبية السائدة هو شائع في مجال البروتينات الحركية. استخدمت المسوخ بلا محركات مماثلة من kinesins وmyosins سابقا. كانت تستخدم أساسا لإثبات آثار المسوخ على تعريب التحت خلوية من الشحنات أو على الوظائف الخلوية 6-12. وقد وضع تركيز أقل على العلاقة المكانية بين المسوخ والشحنات المتضررين منها. ومع ذلك، في بعض حالات، لوحظت المسوخ للمشاركة في توطين مع كاليفورنيا الخاصةrgos 6،10.
التفاعل بين KIF5B والبروتينات المرتبطة بها والتي سبق أن أكدتها في الجسم الحي الخميرة فحص هجين اثنين والكيمياء الحيوية المقايسات المنسدلة مثل شارك في مناعي وفحوصات في المختبر المنسدلة 13-16. في هذه المقالة، وصفت طريقة استخدام البصرية المجهر إضافية مضان لتحديد البروتينات البضائع KIF5B. الطريقة يجعل من استخدام متحولة KIF5B بلا محركات التي تعمل بمثابة متحولة السلبي السائد. أنه يجمع في السيتوبلازم ويؤدي تجميع الشحنات به.
وضع علامات على النوع البري وبلا محركات متحولة KIF5B مع بروتين فلوري tdTomato 17 تمكن التصور من خلال الفحص المجهري مضان. البروتينات KIF5B الموسومة يمكن أن يشترك في التعبير عنها بروتين مرشح تنصهر لبروتين فلوري مختلف مع الخصائص الطيفية فصل مناسب من العلامة KIF5B. ويلاحظ البروتينات الموسومة مباشرة في الخلايا الحيةتحت المجهر مضان. سوف تحريض تجميع البروتين مرشح متحولة KIF5B بلا محركات تأكيد أن البروتين المرشح هو البضائع في الجسم الحي من KIF5B. وعلاوة على ذلك، فإن البروتينات KIF5B الموسومة tdTomato يمكن التعبير عن وحده في الخلايا لدراسة تأثيرها على البروتينات البضائع المحلية. وفي وقت لاحق، ويجري الفحص المجهري المناعي التي يتم إصلاحها الخلايا transfected وملطخة الأجسام المضادة المحددة ضد البروتين المرشح الذاتية، تليها الضد الثانوية المناسب مترافق مع صبغة الفلورسنت. في هذه الحالة، يتم دراسة البروتين المرشح الذاتية على مستوى الفسيولوجية. المسوخ بلا محركات مماثلة من البروتينات السيارات الأخرى يمكن أن تكون على استعداد لتحديد الشحنات الخاصة بهم.
Protocol
1. الاستنساخ من نوع البرية الموسومة tdTomato وبلا محركات KIF5B البروتينات
- تضخيم cDNAs لالبرية من نوع البشري والبروتينات KIF5B بلا محركات باستخدام بادئات في الجدول رقم 1، طق الحمض النووي بوليميريز (5 وحدات لمدة 100 ميكرولتر)، مزيج dNTP (2 مم لكل deoxynucleotide) وعازلة 10X لها لمدة 30 دورات. وتتكون كل دورة من خطوة تمسخ (95 درجة مئوية لمدة 30 ثانية)، خطوة الصلب (45 درجة مئوية لمدة 30 ثانية) وخطوة التمديد (72 درجة مئوية لمدة 3 دقائق).
- استخراج المنتج تضخيم الحمض النووي مع حجم مساو من الفينول / الكلوروفورم (1: 1).
ملاحظة: الفينول هو قابل للاشتعال ويمكن أن يسبب حروقا. الكلوروفورم خطرة. تجنب الاتصال المباشر معهم واستخدامها تحت غطاء الدخان الكيميائية. بدلا من ذلك، منتجات PCR يمكن تنقيته من قبل مختلف مجموعات.- تدور في 18000 x ج في microcentrifuge في درجة حرارة الغرفة لمدة 1 دقيقة. نقل محلول مائي لأنبوب جديد واستخراج مع حجم مساو من كلوروفورم.
- غزلفي 18000 x ج في microcentrifuge في درجة حرارة الغرفة لمدة 0.5 دقيقة. نقل محلول مائي لأنبوب جديد.
- مزيج من محلول مائي مع المجلد العاشر واحدة من 3 M نا-خلات ومجلدين من الايثانول المطلق.
- تدور في 18000 x ج في microcentrifuge في درجة حرارة الغرفة لمدة 5 دقائق. تجاهل محلول مائي.
- غسل بيليه الحمض النووي مع مجلدين من 75٪ من الإيثانول. تجاهل محلول مائي والهواء الجاف بيليه الحمض النووي في درجة حرارة الغرفة لمدة 5 دقائق. Resuspend بيليه في الحمض النووي في 34 ميكرولتر المياه.
- هضم منتجات تضخيم في الحجم النهائي من 40 ميكرولتر مع تقييد الانزيمات سالي (10 وحدة) وBamHI (10 وحدة) و 4 ميكرولتر من العازلة 10X بهم عند 37 درجة مئوية لمدة ساعة. حل المنتجات هضمها في agarose هلام (1.0٪ عند 100 V) التي تحتوي على إيثيديوم بروميد (0.2 ميكروغرام / مل).
ملاحظة: بروميد إيثيديوم هو المغير قوية ويمكن استيعابها من خلال الجلد. وبالتالي، فمن المهم تجنب الاتصال المباشر أو غير المباشر ثإيث إيثيديوم بروميد. - قطع العصابات الصحيحة لتنقية قبل الأعمدة. تزن agarose هلام يحتوي على جزء الحمض النووي. ثم حله في المخزن المؤقت الإذابة (300 ميكرولتر عن 0.1 غ) عند 37 درجة مئوية لمدة 20 دقيقة.
- إضافة محلول أدى إلى عمود وتدور في microcentrifuge في درجة حرارة الغرفة لمدة 5 ثوان. تجاهل التدفق من خلال.
- غسل العمود مع العازلة الإذابة 0.5 مل من تكرار الخطوة 1.5. غسل العمود مرة أخرى مع 0.75 مل العازلة غسل بتكرار الخطوة 1.5. تدور العمود لمدة 2 دقيقة للتخلص من غسل العازلة المتبقية.
- أزل جزء الحمض النووي مع 50 ميكرولتر المياه بتكرار الخطوة 1.5. تقدير تركيز جزء الحمض النووي عن طريق تشغيل قسامة منه ضد السلم حجم الحمض النووي في agarose هلام (1.0٪ عند 100 V) التي تحتوي على إيثيديوم بروميد (0.2 ميكروغرام / مل).
- Ligate منتجات النقاء (حوالي 100 نانوغرام) مع ناقل ptdTomato-C1 17 (حوالي 100 نانوغرام) يهضم سابقا مع الالانزيمات ه التقييد نفسه في 10 ميكرولتر باستخدام T4 DNA يغاز (2000 وحدة) و 1 ميكرولتر العازلة 10X ربط في درجة حرارة الغرفة لمدة 16 ساعة.
ملاحظة: من النوع البري ومتحولة بلا محركات يحتوي على الأحماض الأمينية 2-963 و584-963، على التوالي. سابقا، كانت تستخدم أكبر متحولة KIF5B بلا محركات لتحديد وظيفتها 9. تم استخدام KIF5B متحولة بلا محركات أصغر للدراسات الحالية لزيادة مستويات التعبير عنها.
2. المناعي المجهري
- لحية خلية أو التصوير مضان غير المباشرة مع التكبير عند أو أقل من 40X، البذور 0،2-0٬300٬000 خلايا هيلا في 1 مل المتوسطة كاملة [Dulbecco لتعديل النسر متوسطة (DMEM) مع 10٪ مصل بقري جنيني (FBS)] في كل بئر من ل ستة جيدا لوحة. ينمو بمعدل 37 درجة مئوية مع 5٪ CO 2.
- للدراسات غير المباشرة المناعي مع تكبير 40X فوق، وغسل نظارات الغطاء (18 ملم × 18 ملم؛ 1.5 سمك) في فترة وجيزة الايثانول المطلق والهواء الجاف لهم فيآبار لوحة ستة جيدا، داخل مجلس الوزراء للسلامة الأحيائية لتجنب التلوث.
- البذور 0،2-0٬300٬000 الخلايا في 1 مل يكمل DMEM المتوسطة في كل بئر من لوحة ستة جيدا.
- إعداد مجمع ترنسفكأيشن
- في اليوم التالي، داخل مجلس الوزراء السلامة البيولوجية، تمييع 0.6 ميكروغرام (الكل) من البلازميد التعبير عن الموسومة tdTomato من نوع البرية أو KIF5B بلا محركات في وجود أو عدم وجود التعبير البلازميد لبروتين مرشح البضائع (pTagCFP-ج-MYC 5 ) مع 0.1 مل المتوسطة ترنسفكأيشن في أنبوب 1.6 مل microcentrifuge ل.
ملاحظة: ناقلات pTagCFP-ج-MYC تعبر عن الموسومة TagCFP-ج-MYC. - إضافة 1.8 ميكرولتر من كاشف ترنسفكأيشن إلى 0.1 مل ترنسفكأيشن المتوسطة في أنبوب 1.6 مل microcentrifuge لآخر. عادة، وإعداد مزيج الرئيسي المخفف كاشف ترنسفكأيشن يكفي لمدة 12 عينات.
- احتضان لمدة 5 دقائق، في درجة حرارة الغرفة.
- إضافة المخفف 0.1 مل كاشف ترنسفكأيشن إلى 0.1 مل مخففة السلطات الوطنية المعينة. ميلس محتويات كتبها قلب الأنابيب. تدور الأنابيب لمدة 5 ثانية في microcentrifuge.
- احتضان لمدة 45 دقيقة، في درجة حرارة الغرفة.
- في اليوم التالي، داخل مجلس الوزراء السلامة البيولوجية، تمييع 0.6 ميكروغرام (الكل) من البلازميد التعبير عن الموسومة tdTomato من نوع البرية أو KIF5B بلا محركات في وجود أو عدم وجود التعبير البلازميد لبروتين مرشح البضائع (pTagCFP-ج-MYC 5 ) مع 0.1 مل المتوسطة ترنسفكأيشن في أنبوب 1.6 مل microcentrifuge ل.
- غسل الخلايا
- غسل الخلايا المصنفة ثلاث مرات مع الفوسفات مخزنة المالحة (PBS) خلال فترة الحضانة لتشكيل مجمع ترنسفكأيشن.
- استبدال المتوسطة من الخلايا المصنفة في كل جيدا مع 0.8 مل قبل تحسنت ترنسفكأيشن المتوسطة. العودة لوحات لالحاضنة عند 37 درجة مئوية.
- ترنسفكأيشن من الخلايا
- بعد مرور فترة الحضانة من 45 دقيقة، إضافة 0.2 مل من مجمع كاشف الحمض النووي / ترنسفكأيشن (المعد في الخطوة 2.3) قطرة قطرة إلى كل بئر من لوحة.
- موسيقى الروك لوحة 6 جيدا برفق لمدة 5 ثوان، قبل أن يتم إعادتها إلى حاضنة عند 37 درجة مئوية.
- بعد 6-8 ساعة، استبدال المتوسطة مع المتوسط DMEM كاملة.
- لخلية حية الإسفار التصوير 18
- بعد 16 ساعة إضافية من الحضانة لر 37 درجة مئوية، إضافة صمة عار الحمض النووي الإقحام هويشت 33342 إلى وسيلة لتركيز النهائي من 1 ميكرومتر.
ملاحظة: هويشت 33342 هو خلايا قابلة للاختراق، صبغ الحمض النووي الإقحام أن البقع نواة الخلية. - احتضان خلايا transfected لمدة 10 دقيقة في الحاضنة عند 37 درجة مئوية قبل دراستها للتعبير عن البروتينات الفلورية بواسطة المجهر الفلورسنت باستخدام الهدف 40X. مجموعات تصفية دابي، CFP، FITC وو Cy3 هي (Ex350 نانومتر / Em460 نانومتر)، (Ex436 نانومتر / Em480 نانومتر)، (Ex470 نانومتر / Em525 نانومتر) و (Ex545 نانومتر / Em605 نانومتر)، على التوالي.
ملاحظة: إذا كانت خلفية عالية نظرا لصناعة السيارات في مضان من المتوسط DMEM كاملة، يتم استخدام وسيط وبدون أحمر الفينول.
- بعد 16 ساعة إضافية من الحضانة لر 37 درجة مئوية، إضافة صمة عار الحمض النووي الإقحام هويشت 33342 إلى وسيلة لتركيز النهائي من 1 ميكرومتر.
3. للدراسات المناعي غير المباشرة
- إعداد امتصاص العرق الحل
- وزن مسحوق امتصاص العرق (4 غ لكل 100 مل PBS) وإضافته إلى برنامج تلفزيوني.
ملاحظة: لامتصاص العرق هو قابل للاشتعال الصلبة وهزار سرطان محتملد. يهيج العينين والجهاز التنفسي والجلد. وبالتالي، فمن المهم تجنب الاتصال مع أو استنشاق امتصاص العرق. - إضافة هيدروكسيد الصوديوم إلى محلول (150 ميكرولتر 10 N هيدروكسيد الصوديوم / 100 مل).
- إبقاء الحل في 37-42 درجة مئوية لمدة 2-3 ساعة مع الهز في بعض الأحيان.
- ضبط الرقم الهيدروجيني إلى 7.0 بإضافة حمض الخليك الجليدي إلى حل (حوالي 75 ميكرولتر / 100 مل) بعد مسحوق امتصاص العرق يذوب.
- وزن مسحوق امتصاص العرق (4 غ لكل 100 مل PBS) وإضافته إلى برنامج تلفزيوني.
- الهدف البروتينات تلطيخ
- وبعد مزيد من الحضانة لمدة 16 ساعة على 37 درجة مئوية، وغسل الخلايا transfected في درجة حرارة الغرفة مرة واحدة مع برنامج تلفزيوني من قبل يحوم لوحة ستة جيدا لمدة 5 ثوان.
- إصلاح الخلايا مع 1 مل من الطازجة، و 4٪ حل لامتصاص العرق لكل بئر. يحضن في درجة حرارة الغرفة لمدة 30 دقيقة.
- غسل الخلايا مع برنامج تلفزيوني مرة واحدة من قبل يحوم لمدة 5 ثانية لوحة ستة جيدا. تجاهل الحل.
- احتضان الخلايا مع 1 مل من 0.1٪ تريتون X-100 (في برنامج تلفزيوني) في درجة حرارة الغرفة لمدة 30 دقيقة. وديتergent تريتون-X 100 سوف permeabilize غشاء الخلية للسماح بالوصول من الأجسام المضادة لأهدافها داخل الخلايا.
- غسل الخلايا مع برنامج تلفزيوني أربع مرات من قبل يحوم لوحة ستة جيدا لمدة 5 ثانية في كل مرة. تجاهل الحل بعد كل غسل.
- احتضان الخلايا مع 1 مل من الأجسام المضادة الأولية (ج MYC أرنب الأجسام المضادة أو الأجسام المضادة البروتين p53 الماوس؛ 0.1 ميكروغرام / مل) في FBS 10٪ في حل برنامج تلفزيوني في درجة حرارة الغرفة قبل هزاز لمدة 4 ساعة.
- يغسل مع برنامج تلفزيوني أربع مرات من قبل يحوم لوحة ستة جيدا لمدة 5 ثانية في كل مرة. تجاهل الحل بعد كل غسل.
- احتضان مع 1 مل من الأجسام المضادة الثانوية صبغ مترافق فلوري (اليكسا فلور 488 مترافق المضادة للأرنب أو المضادة للماوس مفتش الأضداد؛ 0.5 ميكروغرام / مل) في 10٪ FBS في برنامج تلفزيوني في درجة حرارة الغرفة في الظلام بواسطة هزاز لمدة 2 ساعة.
- يغسل مع برنامج تلفزيوني أربع مرات من قبل يحوم لوحة ستة جيدا لمدة 5 ثانية في كل مرة. تجاهل الحل بعد كل غسل.
- تلطيخ النووية وتركيب
- احتضان الخلايا مع 1 مل من الحمض النووي الإقحام صبغ 4 "، 6 diamidino-2-phenylindole (دابي؛ 0.5 ميكروغرام / مل) في برنامج تلفزيوني في درجة حرارة الغرفة، في الظلام لمدة 10 دقيقة. انتقل إلى الخطوة 3.4 عندما يتم استخدام أي نظارات غطاء.
- تطبيق 10 ميكرولتر من محلول متزايدة على كل شريحة المجهر. جبل كل غطاء زجاجي على حل متزايدة على شريحة المجهر.
- يحضن في درجة حرارة الغرفة في ليلة وضحاها الظلام.
- ختم حواف من النظارات غطاء بطلاء الأظافر. ضع في الظلام داخل غطاء الدخان بين عشية وضحاها لإزالة مسمار البولندية الأبخرة.
ملاحظة: دابي هو الحمض النووي الإقحام الصبغة التي البقع نواة الخلية. - مضان المجهر 19
- وفي اليوم التالي، فحص الخلايا بواسطة المجهر الفلورسنت باستخدام الهدف 40X. مجموعات تصفية دابي، CFP، FITC وو Cy3 هي (Ex350 نانومتر / Em460 نانومتر)، (Ex436 نانومتر / Em480 نانومتر)، (Ex470 نانومتر / Em525 نانومتر) و (Ex545 نانومتر / Em605 نانومتر)، على التوالي.
النتائج
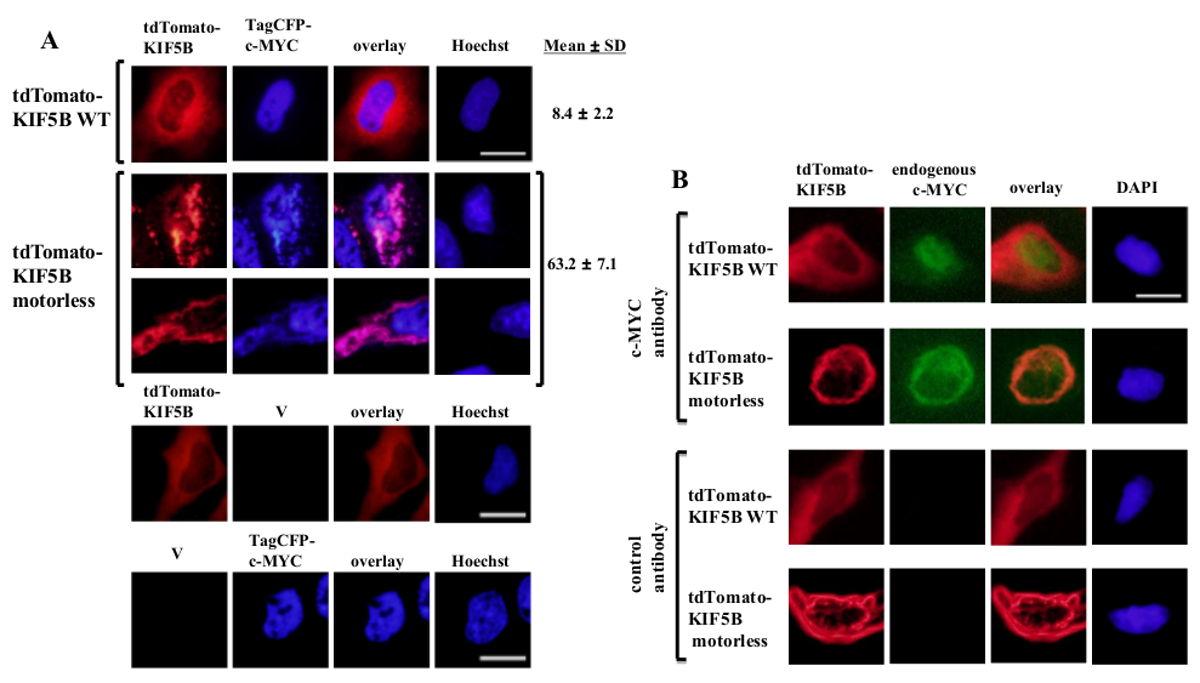
وأعرب خارجيا من النوع البري KIF5B ظهر متجانس في السيتوبلازم، في حين بدا ج-MYC أساسا في النواة (الشكل 1A). ومع ذلك، شكلت KIF5B متحولة بلا محركات المجاميع في السيتوبلازم (الشكل 1A). تجميع للKIF5B بلا محركات يسببها تجميع ج-MYC. النسبة المئوية للخلايا معربا عن تجميع الكلمات الدلالية TagCFP-ج-MYC في الخلايا معربا عن النوع البري KIF5B كانت منخفضة. ومع ذلك، كان أعلى بكثير عندما كان يشارك في التعبير عن KIF5B بلا محركات (الشكل 1A). لاحظ شارك في توطين KIF5B متحولة مع ج-MYC (الشكل 1) تشير إلى أن KIF5B ينظم توطين التحت خلوية من ج-MYC وج-MYC هو شحنة من كينيسين-1. وشملت الضوابط السلبية التي أبديت يبني وحده في اللوحة السفلى من الشكل 1A. وأظهرت النتائج أنه لا يوجد كبير تنزف من خلال من emissio مضانن. كما أقنع KIF5B بلا محركات تجميع والذاتية ج-MYC (الشكل 1B) والبروتين p53 عامل النسخ (الشكل 2)، مشيرا إلى أن ج-MYC والبروتين p53 كلاهما الشحنات من كينيسين-1 وKIF5B ينظم توطين التحت خلوية من كل من البروتينات الذاتية. جنبا إلى جنب مع النتائج التي المانع كينيسين-1 ارتفع اكتون البنغال (ربل) تشكيل 20 الحث من الأنواع عالية الوزن الجزيئي لكلا ج-MYC والبروتين p53 5، من المرجح أن يكون البروتين p53 شحنة من كينيسين-1. ومن المثير للاهتمام أن نلاحظ أن النسخ عامل البروتين p53 النووي، مثل ج-MYC، ويتم تصدير أيضا من النواة إلى السيتوبلازم لتدهور proteasomal 21،22. تبدو الحركة من عوامل النسخ التي كتبها KIF5B أن تكون محددة لأن التعبير عن متحولة KIF5B بلا محركات لم تؤثر على توطين التحت خلوية ج-فو أو يؤدي إلى تجميع (الشكل 3). وتوضح البيانات الواردة أعلاه أن methoد المستخدمة في هذا المنشور يسمح لتحديد عالية التحديد من كينيسين-1 الشحنات. ويمكن تطبيق استراتيجية مماثلة لالبروتينات السيارات الأخرى كذلك.

الشكل 1: التعبير عن متحولة KIF5B بلا محركات الحث تجميع ج-MYC في السيتوبلازم و transfected (A) خلايا هيلا مع المعلمة tdTomato من نوع البرية (WT) KIF5B (الأحمر) والموسومة TagCFP-ج-MYC (الأزرق). ظهر الموسومة tdTomato-WT KIF5B أساسا في السيتوبلازم، في حين TagCFP الموسومة ج-MYC ظهرت أساسا في النواة. ومع ذلك، فإن الكلمات الدلالية tdTomato-متحولة KIF5B بلا محركات (الحمراء) تشكيل المجاميع الخيطية أو منقط في السيتوبلازم. التعبير عن KIF5B بلا محركات يسببها تجميع ج-MYC. وKIF5B متحولة وج-MYC المشارك المحلية معا (الوردي). وتظهر النسب المئوية للخلايا (٪) مما يدل مجاميع من CFP-ج-MYC (٪). وتظهر النتائج على النحويعني ± SD (N = 3). وترد ضوابط السلبية مع التعبير عن البروتينات KIF5B الموسومة tdTomato أو المعلمة TagCFP-ج-MYC جنبا إلى جنب مع ناقلات فارغة (V) في اللوحة السفلى. تم الحصول على (ب) نتائج مماثلة مع الذاتية ج-MYC (الأخضر) عندما أبديت الموسومة tdTomato-WT أو البروتينات KIF5B بلا محركات (الحمراء). وKIF5B متحولة بلا محركات الموسومة tdTomato-تشكيل المجاميع في السيتوبلازم والتي يسببها تراكم من الذاتية ج-MYC. كلا النوعين من المجاميع شارك مترجمة معا في السيتوبلازم (برتقالي). كانت ملطخة النوى مع صبغة هويشت 33342 أو دابي. شريط الحجم؛ 20 ميكرون. الرجاء انقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}

الشكل 2: التعبير عن متحولة KIF5B بلا محركات الحث تجميع البروتين p53 الذاتية في تيانه السيتوبلازم. وفي خلايا هيلا، خارجية المنشأ الموسومة tdTomato البرية من نوع (WT) KIF5B (الحمراء) ظهرت أساسا في السيتوبلازم، في حين أن البروتين p53 الذاتية (الأخضر) ظهرت أساسا في النواة. في المقابل، فإن الكلمات الدلالية tdTomato-متحولة KIF5B بلا محركات (الحمراء) تشكيل المجاميع في السيتوبلازم. التعبير عن KIF5B بلا محركات يسببها تجميع البروتين p53، مما أدى إلى شارك في توطين KIF5B والبروتين p53 في السيتوبلازم (الصفراء). وأجريت التجربة ثلاث مرات. كانت ملطخة النوى مع دابي صبغ. شريط الحجم؛ 20 ميكرون. الرجاء انقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}

الرقم 3: التعبير عن متحولة KIF5B بلا محركات لا يسبب تجميع ج-فو في السيتوبلازم tdTomato الموسومة البرية من نوع (WT) KIF5B (الحمراء) بدا أساسا في سيتوبلازم خلايا هيلا، في حين EGFP-ج-فو (الأخضر) ظهرت أساسا في النواة. على العكس من ذلك، الموسومة tdTomato-متحولة KIF5B بلا محركات (الحمراء) تشكيل المجاميع في السيتوبلازم. لم تعبير عن KIF5B بلا محركات لا لحث على تجميع ج-فو. وأجريت التجربة أربع مرات. كانت ملطخة نوى الخلايا مع دابي صبغ. شريط الحجم؛ 20 ميكرون. الرجاء انقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}
| التمهيدي عكس الشائع | 5'-AGAGGATCCTTACACTTGTTTGCCTCCTC-3 " |
| من النوع البري KIF5B التمهيدي إلى الأمام | 5'- AGAGTCGACGCGGACCTGGCCGAGTGCAACATCAAAGT-3 " |
| KIF5B بلا محركات التمهيدي إلى الأمام | 5'- AGAGTCGACGATGAAGAGTTCACTGTTGC-3 " |
: المحافظة على together.within الصفحات = "1"> الجدول 1: تسلسل التمهيدي لاستنساخ البرية من نوع والبروتينات KIF5B بلا محركات.
Discussion
وصف الأسلوب يستخدم خصائص متحولة KIF5B بلا محركات، التي تفتقر إلى القدرة على التحرك على طول الأنابيب الدقيقة، لكنه يحتفظ القدرة على تشكيل dimers مع من النوع البري KIF5B، وبالتالي السماح للبروتين رباعي القسيمات للتفاعل مع حمولة البروتينات نفسها كما البرية من نوع KIF5B. KIF5B بلا محركات، وبالتالي، بمثابة متحولة السلبي السائد وأشكال mislocalized المجاميع مع الشحنات لها. وقد ثبت هذا الأسلوب لتحديد كينيسين-1 البضائع ج-MYC (الشكل 1) 5. في هذه المقالة، تم استخدام نفس متحولة KIF5B بلا محركات لتحديد البروتين p53 كما البضائع أخرى محتملة من كينيسين-1 (الشكل 2). هذا يدل على أن المنهجية الممكنة لتحديد الشحنات الأخرى من كينيسين-1. وعلاوة على ذلك، يتم توفير خصوصية متحولة بسبب عدم وجود تأثير متحولة على البروتين تحكم سلبي ج-فو (الشكل 3).
في هذا البروتوكول، وtdTomato الموسومة البرية تايالمؤسسة العامة أو coexpressed بلا محركات بروتين KIF5B مع الفلورسنت الموسومة البروتين البروتين البضائع مرشح آخر. في هذه الحالة، يتم تنفيذ الخلية الحية مضان المجهر والتصوير. تشكيل المجاميع يمكن تتبع من قبل الوقت الفاصل بين التصوير. بدلا من ذلك، يتم التعبير عن بروتين الموسومة tdTomato وحده، وهو تصور البروتين البضائع مرشح على المستويات الفسيولوجية بواسطة المجهر المناعي غير المباشر باستخدام أجسام مضادة محددة. يتم اختيار tdTomato البروتين الفلوري لمعانه وصمود 17. إذا كانت الخلفية عالية نظرا لصناعة السيارات في مضان من المتوسط DMEM كاملة، يتم استخدام وسيط وبدون أحمر الفينول.
وKIF5B متحولة بلا محركات يشكل dimers مع من النوع البري KIF5B stoichiometrically. لذا، فمن الأهمية بمكان للتعبير عن كمية كافية من المسخ KIF5B بلا محركات لتشكيل dimers مع البرية من نوع النظير لتثبيط وظيفة ولتشكيل المجاميع. لمعالجة هذه المسألة، والتحسين من EXPRession من متحولة بلا محركات أمر ضروري. ويتحقق ذلك عن طريق استخدام متحولة بلا محركات صغير يحتوي المجال dimerization. وبالإضافة إلى ذلك، والتحسين من كاشف ترنسفكأيشن المناسب وبروتوكول ضروري أيضا. وكان محسن مدة حضانة المرض من خلايا هيلا مع كاشف الحمض النووي / ترنسفكأيشن في خلايا هيلا للتعبير عن البروتينات الخارجية وبقاء الخلية. قد يكون من الضروري الأمثل لخطوط الخلايا الأخرى مدة الحضانة.
لتكبير بين 4X إلى 40X، ليس هناك حاجة لنظارات غطاء. الخلايا يمكن فحص مباشرة في الآبار. لذلك، في هذه الحالة، بروتوكول غير مكلفة ومريحة. لتكبير فوق 40X، وتزرع الخلايا على نظارات غطاء في الآبار، وبعد التلوين، هي التي شنت على الشرائح المجهر لفحصها تحت الأهداف النفط الغمر.
مراقبة مجاميع من البروتين KIF5B بلا محركات محدود بسبب حجم السيتوبلازم. فمن السهل للكشف عنالمجاميع في العديد من خلايا الثدييات. ومع ذلك، فإنه من الصعب نسبيا لمراقبة المجاميع في الخلايا العصبية عندما يكون حجم السيتوبلازم صغير حول النواة وفي neurites التي.
يتم استخدام البروتوكول لإظهار العلاقة بين KIF5B والشحنات لها بالإضافة إلى المجراة الخميرة اثنين الهجين والكيمياء الحيوية المقايسات المنسدلة 13-16. كل هذه المقايسات تحدد الجمعية في ظل ظروف مادية مختلفة، ويمكن نتائجها يكمل كل منهما الآخر. وعلاوة على ذلك، والاستفادة من هذا الاختبار مضان خلال فحوصات أخرى هي أنه يمكن أن تظهر تنظيم توطين التحت خلوية من الشحنات التي كتبها KIF5B (الشكلان 1 و 2).
البروتوكول لا يقتصر على KIF5B، ويمكن استخدامها لتحديد الشحنات من البروتينات السيارات الأخرى، مثل بعض المحركات كينيسين أخرى 3 و بعض المحركات الميوسين 23،24 المستخدمة في نقل الخلايا من الشحنات 3. تحتوي هذه البروتينات المحرك أيضا المجالات الحركية للحركة على طول الأنابيب الدقيقة لمحركات كينيسين وخيوط الأكتين للمحركات الميوسين، وشرائح ملفوف لفائف لoligomerization. معظمهم من تشكيل homodimers 3،23. لذلك، يمكن تطبيق استراتيجية مماثلة لهم من خلال خلق بلا محركات المسوخ السلبية المهيمنة لتحديد الشحنات الخاصة بهم.
Disclosures
ويدفع مجانا نشر وصول وإنتاج هذه المادة لشركة روش.
Acknowledgements
Roche's Publication Grant covered the Free Access publication and production of this article. The author also thanks E. Premkumar Reddy, Richard V. Mettus, Stephen C. Cosenza, Sau Ying Yip and Sol D. Gloria for their support and critical reading of the manuscript.
Materials
| Name | Company | Catalog Number | Comments |
| absolute ethanol (200 proof) | Fisher Scientific | BP2818 | |
| DAPI | Sigma-Aldrich | D9542 | |
| Hoechst 33342 | Sigma-Aldrich | B2261 | |
| Opti-MEM-I (transfection medium) | Life Technologies | 51985 | |
| ProLong Diamond Antifade Mountant | Life Technologies | P36961 | |
| formaldehyde, para | Fisher Scientific | O4042-500 | |
| Triton-X100 | Fisher Scientific | BP151-500 | |
| X-tremeGENE 9 DNA Transfection Reagent | Roche | 6365787001 | |
| Taq DNA polymerase | Life Technologies | 10342020 | |
| PCR Grade Nucleotide Mix (dNTP Mix) | Roche | 12111424 | |
| Microscope cover glass | Fisher Scientific | 12-541A | |
| GeneRuler 1 kb Plus DNA ladder | Life Technologies | SM1333 | |
| PureLink Quick Gel Extraction kit | Life Technologies | K210012 | |
| BamHI | New England Biolab | R0136 | |
| SalI | New England Biolab | R0138 | |
| T4 DNA ligase | New England Biolab | M0202T | |
| Ethidium bromide | Thermo Scientific | 17898 | |
| DMEM | Life Technologies | 11995-065 | |
| c-MYC rabbit antibody | Cell Signaling | 5606 | |
| p53 mouse antibody | Santa Cruz | sc-126 | |
| Alexa Fluor 488-conjugated anti-rabbit IgG antibody | Life Technologies | A11008 | |
| Alexa Fluor 488-conjugated anti-mouse IgG antibody | Life Technologies | A11059 | |
| fluorescent microscope | Olympus IX71_Fluoview | ||
| computer software for imaging | cellSens |
References
- Hirokawa, N., Niwa, S., Tanaka, Y. Molecular motors in neurons: transport mechanisms and roles in brain function, development, and disease. Neuron. 68, 610-638 (2010).
- Yu, Y., Feng, Y. M. The role of kinesin family proteins in tumorigenesis and progression: potential biomarkers and molecular targets for cancer therapy. Cancer. 116, 5150-5160 (2010).
- Verhey, K. J., Hammond, J. W. Traffic control: regulation of kinesin motors. Nat Rev Mol Cell Biol. 10, 765-777 (2009).
- Hirokawa, N., Noda, Y., Tanaka, Y., Niwa, S. Kinesin superfamily motor proteins and intracellular transport. Nat Rev Mol Cell Biol. 10, 682-696 (2009).
- Lee, C. M. Transport of c-MYC by Kinesin-1 for proteasomal degradation in the cytoplasm. Biochim Biophys Acta. 1843, 2027-2036 (2014).
- Bittins, C. M., Eichler, T. W., Hammer, J. A., Gerdes, H. H. Dominant-negative myosin Va impairs retrograde but not anterograde axonal transport of large dense core vesicles. Cell Mol Neurobiol. 30, 369-379 (2010).
- Kimura, T., Watanabe, H., Iwamatsu, A., Kaibuchi, K. Tubulin and CRMP-2 complex is transported via Kinesin-1. J Neurochem. 93, 1371-1382 (2005).
- Lindsay, A. J., McCaffrey, M. W. Myosin Va is required for the transport of fragile X mental retardation protein (FMRP) granules. Biol Cell. 106, 57-71 (2014).
- Rivera, J., Chu, P. J., Lewis, T. L., Arnold, D. B. The role of Kif5B in axonal localization of Kv1 K(+) channels. Eur J Neurosci. 25, 136-146 (2007).
- Roland, J. T., Kenworthy, A. K., Peranen, J., Caplan, S., Goldenring, J. R. Myosin Vb interacts with Rab8a on a tubular network containing EHD1 and EHD3. Mol Biol Cell. 18, 2828-2837 (2007).
- Uchida, A., Alami, N. H., Brown, A. Tight functional coupling of kinesin-1A and dynein motors in the bidirectional transport of neurofilaments. Mol Biol Cell. 20, 4997-5006 (2009).
- Zadeh, A. D., et al. Kif5b is an essential forward trafficking motor for the Kv1.5 cardiac potassium channel. J Physiol. 587, 4565-4574 (2009).
- Su, Y. Y., et al. KIF5B promotes the forward transport and axonal function of the voltage-gated sodium channel Nav1.8. J Neurosci. 33, 17884-17896 (2013).
- Lalioti, V. S., Vergarajauregui, S., Tsuchiya, Y., Hernandez-Tiedra, S., Sandoval, I. V. Daxx functions as a scaffold of a protein assembly constituted by GLUT4, JNK1 and KIF5B. . J Cell Physiol. 218, 416-426 (2009).
- Cho, K. I., et al. Association of the kinesin-binding domain of RanBP2 to KIF5B and KIF5C determines mitochondria localization and function. Traffic. 8, 1722-1735 (2007).
- Diefenbach, R. J., Diefenbach, E., Douglas, M. W., Cunningham, A. L. The ribosome receptor, p180, interacts with kinesin heavy chain, KIF5B. Biochem Biophys Res Commun. 319, 987-992 (2004).
- Shaner, N. C., et al. Improved monomeric red, orange and yellow fluorescent proteins derived from Discosoma sp. red fluorescent protein. Nat Biotechnol. 22, 1567-1572 (2004).
- Waters, J. C. Live-cell fluorescence imaging. Methods Cell Biol. 114, 125-150 (2013).
- Sanderson, M. J., Smith, I., Parker, I., Bootman, M. D. Fluorescence microscopy. Cold Spring Harb Protoc. 2014, (2014).
- Hopkins, S. C., Vale, R. D., Kuntz, I. D. Inhibitors of kinesin activity from structure-based computer screening. Biochemistry. 39, 2805-2814 (2000).
- Zhang, Y., Xiong, Y. Control of p53 ubiquitination and nuclear export by MDM2 and ARF. Cell Growth Differ. 12, 175-186 (2001).
- Michael, D., Oren, M. The p53-Mdm2 module and the ubiquitin system. Semin Cancer Biol. 13, 49-58 (2003).
- Kneussel, M., Wagner, W. Myosin motors at neuronal synapses: drivers of membrane transport and actin dynamics. Nat Rev Neurosci. 14, 233-247 (2013).
- Maravillas-Montero, J. L., Santos-Argumedo, L. The myosin family: unconventional roles of actin-dependent molecular motors in immune cells. J Leukoc Biol. 91, 35-46 (2012).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved