
Method Article
使用荧光显微镜驱动蛋白-1的货物的识别
摘要
在这里,提出一个协议,以确定驱动蛋白-1的货物。该驱动蛋白1重链(KIF5B)的无电机突变体聚集在细胞质和诱导其货物的聚集。两个聚集体在荧光显微镜下检测。类似的策略可以用于识别其他马达蛋白的货物。
摘要
Fluorescence microscopy is employed to identify Kinesin-1 cargos. Recently, the heavy chain of Kinesin-1 (KIF5B) was shown to transport the nuclear transcription factor c-MYC for proteosomal degradation in the cytoplasm. The method described here involves the study of a motorless KIF5B mutant for fluorescence microscopy. The wild-type and motorless KIF5B proteins are tagged with the fluorescent protein tdTomato. The wild-type tdTomato-KIF5B appears homogenously in the cytoplasm, while the motorless tdTomato-KIF5B mutant forms aggregates in the cytoplasm. Aggregation of the motorless KIF5B mutant induces aggregation of its cargo c-MYC in the cytoplasm. Hence, this method provides a visual means to identify the cargos of Kinesin-1. A similar strategy can be utilized to identify cargos of other motor proteins.
引言
驱动蛋白1是介导其货物1,2-顺行输送马达蛋白。它是驱动蛋白轻链1(KLC1)和驱动蛋白重链的两个亚基(KHCs)的两个亚单位的异四聚体。 KIF5B 1,KHC,包含在其N-末端,其水解ATP和化学能转换为机械能用于沿微管运动的电机领域。其C-末端区包含与KLC1,其与货物相关联的交互二聚化结构域。驱动蛋白-1运输货物,如囊泡,细胞器和mRNA的3,4。最近,KIF5B显示出运送核转录因子c-Myc的对蛋白酶体降解在细胞质5。三种方法(化学抑制剂的siRNA / shRNA的和显性负突变体)被用来抑制驱动蛋白1功能。它们都诱导的c-MYC的聚集在细胞质中。在过去的方法,c-Myc的仅受主导negat香港专业教育学院KIF5B的突变体,但不是由另一相关KIF5A马达的蛋白质,这表明该突变不会对细胞内的部件或一般的效果(如微管中断)对蛋白聚集。 KIF5B的显性负突变体也没有影响另一个转录因子,这表明它不会对转录因子的一般效果。相反,它表明,显性负突变体上施加其货物特异性作用。
使用显性负突变体的是在电动机的蛋白质的领域常见的。驱动蛋白和肌球蛋白的类似无电机突变体以前使用。它们主要是用来展示在他们的货物的亚细胞定位或对细胞功能的6-12突变体的效果。不太重视被提上了突变体和受其影响的货物之间的空间关系。然而,在一些发生率,观察到突变体与其CA到共定位RGOS 6,10。
KIF5B及其相关蛋白质之间的相互作用以前由体内酵母双杂交测定和生化下拉测定法诸如免疫共沉淀和体外下拉实验13-16证实。在这篇文章中,一个额外的视觉方法使用荧光显微镜描述识别KIF5B货物蛋白质。该方法利用充当显性负突变体一种无马达KIF5B突变体。它聚合在细胞质和诱导其货物的聚集。
野生型和无电机KIF5B突变体与荧光蛋白tdTomato 17的标记通过荧光显微镜实现其可视化。加标签KIF5B蛋白可以与融合到与来自KIF5B标签适当地分离的光谱特性的不同荧光蛋白的候选蛋白共表达。该标记的蛋白在活细胞中直接观察在荧光显微镜下。由无电机KIF5B突变候选蛋白质的聚集的诱导将确认该候选蛋白是KIF5B 的体内的货物。此外,tdTomato标记KIF5B蛋白可以单独表达在细胞中,研究了内源性货物蛋白它们的影响。以后,免疫荧光显微镜下进行,其中转染的细胞固定并用抗内源性候选蛋白的特异性抗体染色,接着用荧光染料缀合的适当的二级抗体。在这种情况下,在其生理水平的内源候选蛋白质进行了研究。其他马达蛋白的类似无动力突变体可以准备识别他们的货物。
研究方案
1. tdTomato标记野生型和无电机KIF5B蛋白质的克隆
- 扩增的cDNA为使用引物在表1中的人野生型和无电机KIF5B蛋白质,Taq DNA聚合酶(5单位为100微升),dNTP混合物(2mM的每种脱氧核苷酸)和30个循环的10×缓冲液。每个循环由一个变性步骤(95℃,30秒),退火步骤(45℃,30秒)和延伸步骤(72℃,3分钟)的。
- 提取扩增DNA产物与等体积的苯酚/氯仿(1:1)。
注:苯酚是易燃,可致灼伤。三氯甲烷是危险的。避免与他们直接接触,化学通风柜下使用它们。可替代地,PCR产物可通过各种试剂盒进行纯化。- 自旋18000 XG在微量,在室温下放置1分钟。水溶液转移到新的试管中,用等体积的氯仿萃取。
- 纺在室温下微量离心0.5分钟18000 xg离心。转移水溶液至新管。
- 混合使用的3米乙酸钠十分之一体积和无水乙醇两体积的水溶液中。
- 自旋18000 XG在微量在室温下5分钟。丢弃水溶液。
- 两个体积的75%乙醇洗涤DNA沉淀。丢弃水溶液和空气干燥,在室温下将DNA沉淀5分钟。重悬34微升水DNA沉淀。
- 消化的扩增产物中的40微升用限制性终体积内切酶SalI(10单位)和BamHI(10单位),在37℃下4微升的10X缓冲液两小时。解决在琼脂糖凝胶含溴化乙锭(0.2微克/毫升)的消化产物(在100V 1.0%)。
注意:溴化乙锭是一种强效的诱变剂,并且可以通过皮肤被吸收。因此,为了避免直接或间接W接触是很重要的第i溴化乙锭。 - 按列削减了净化正确的波段。称重含有该DNA片段的琼脂糖凝胶。然后将其溶解在37℃的溶解缓冲液(300微升为0.1克)约20分钟。
- 在室温下添加所得到的溶液,以一列和自旋在微量5秒。丢弃的流量通过。
- 通过重复步骤1.5洗用0.5ml溶解缓冲液的柱。重复步骤1.5 0.75 1ml洗涤缓冲液洗一遍列。旋转2分钟摆脱剩余的洗涤缓冲液的列。
- 通过重复步骤1.5洗脱用50μl水的DNA片段。通过在含有溴化乙锭(0.2微克/毫升)的琼脂糖凝胶(在100V 1.0%)运行它针对DNA大小梯等分试样估计的DNA片段的浓度。
- 先前与日消化ptdTomato-C1载体17(约100纳克)结扎纯化的产品(约100纳克)È相同限制性内切酶在使用T4DNA连接酶(2000单位)和1微升10X连接缓冲液在室温下搅拌16小时10微升。
注意:野生型和无电机突变体分别含有氨基酸2至963以及584至963。先前,较大的无电机KIF5B突变体被用来确定其功能9。用较小的无动力KIF5B突变本研究,以提高其表达水平。
2.免疫荧光显微镜
- 对活细胞或具有等于或低于40X放大率间接荧光成像,种子0.2-0.3百万HeLa细胞在1ml完全培养基[Dulbecco氏改良Eagle培养基(DMEM)中,用10%胎牛血清(FBS)〕中的各孔6孔板。在37℃下生长,用5% 的 CO 2。
- 对于上述40X放大倍率间接免疫荧光研究,洗盖玻片,在无水乙醇短暂(18毫米×18毫米1.5厚)风干他们一个6孔板的孔中,一个生物安全柜内以避免污染。
- 种子0.2-0.3百万个细胞的1ml在六孔板的每个孔中完成的DMEM培养基。
- 转染复合物的制备
- 第二天,一个生物安全柜内,稀0.6微克为在存在或不存在的表达质粒的用于货物的候选蛋白质tdTomato标记的野生型或无电机KIF5B表达质粒的(总)(pTagCFP-c-Myc的5 )在1.6 ml离心管0.1毫升转染培养基。
注:矢量pTagCFP-c-Myc的表达TagCFP标记的c-MYC。 - 添加1.8微升转染试剂按0.1ml转染培养基中另外16 ml离心管。通常情况下,准备稀释转染试剂足够12个样品的主结构。
- 孵育5分钟,在室温下。
- 稀释0.1毫升转染试剂添加到稀释的0.1毫升的DNA。弥反相管的X含量。旋管用于在微量离心5秒。
- 孵育45分钟,在室温。
- 第二天,一个生物安全柜内,稀0.6微克为在存在或不存在的表达质粒的用于货物的候选蛋白质tdTomato标记的野生型或无电机KIF5B表达质粒的(总)(pTagCFP-c-Myc的5 )在1.6 ml离心管0.1毫升转染培养基。
- 细胞洗涤
- 洗接种的细胞用磷酸缓冲盐水(PBS)三次潜伏期间的转染复合物的形成。
- 在各替换接种的细胞的培养基以及用0.8mL预热转染培养基。返回所述板的孵化器在37℃。
- 细胞转染
- 45分钟温育后,增加±毫升的DNA /转染试剂复合物(在步骤2.3中制备)滴加至板的各孔中。
- 轻轻摇动6孔板5秒钟,它们在37℃下放回培养箱之前。
- 6-8小时后,更换完全DMEM培养基的媒介。
- 活细胞荧光成像18
- 孵化一个额外16小时后吨37℃时,DNA嵌入污点的Hoechst 33342加入到培养基中,以1μM的终浓度。
注意:赫斯特33342是一种细胞渗透性,DNA嵌入染料染色细胞核。 - 通过使用40倍物镜的荧光显微镜检查它们的荧光蛋白质的表达之前孵育在孵化10分钟的转染细胞在37℃。过滤套的DAPI,CFP,FITC和Cy3的是(Ex350纳米/ Em460纳米),(Ex436纳米/ Em480纳米),(Ex470纳米/ Em525毫微米)和(Ex545纳米/ Em605纳米),分别。
注意:如果背景是高,由于完整的DMEM培养基的自体荧光,无酚红培养基中。
- 孵化一个额外16小时后吨37℃时,DNA嵌入污点的Hoechst 33342加入到培养基中,以1μM的终浓度。
3.间接免疫荧光研究
- 多聚甲醛溶液的制备
- 称重多聚甲醛粉末(每100毫升PBS中的4克)和将其添加到PBS中。
注意:多聚甲醛是一种易燃固体和潜在的癌症哈萨ð。它会刺激眼睛,呼吸系统和皮肤。因此,为了避免多聚甲醛的接触或吸入它是重要的。 - 添加NaOH,使溶液(150微升10的1N NaOH / 100毫升)中。
- 保持在37溶液至42℃2-3小时,偶尔晃动。
- 通过加入冰醋酸至多聚甲醛粉末后的溶液(约75微升/ 100毫升)溶解pH调整到7.0。
- 称重多聚甲醛粉末(每100毫升PBS中的4克)和将其添加到PBS中。
- 靶蛋白染色
- 在37℃下进一步温育16小时后,由漩六孔板5秒洗在用PBS室温一次转染的细胞。
- 修复细胞用1毫升每孔新鲜配制,4%多聚甲醛溶液中。在室温下孵育30分钟。
- 一旦通过涡旋5秒的6孔板用PBS洗细胞。弃去溶液。
- 孵育所述细胞在室温下将1ml 0.1%的Triton-X 100(在PBS中)处理30分钟。该DETergent的Triton-X 100将透化细胞膜以允许抗体访问它们的细胞内的目标。
- 由旋流六孔板的每个时间5秒清洗细胞用PBS四次。每个丢弃后洗净解决方案。
- 通过摇动4小时,在室温下的PBS溶液中的10%胎牛血清;孵育用1ml的初级抗体的细胞(0.1微克/毫升的c-MYC兔抗体或p53的小鼠抗体)。
- 由旋流六孔板的每个时间5秒,用PBS洗涤四次。每个丢弃后洗净解决方案。
- 通过摇动2小时,在PBS中的10%FBS的在黑暗中于室温;(0.5微克/毫升的Alexa Fluor 488缀合的抗兔或抗小鼠IgG抗体)用1ml荧光染料缀合的次级抗体孵育。
- 由旋流六孔板的每个时间5秒,用PBS洗涤四次。每个丢弃后洗净解决方案。
- 核染色和安装
- 孵育1毫升DNA嵌入染料4',6-二脒基-2-苯基的细胞(DAPI; 0.5微克/毫升)的PBS在室温下,在黑暗中进行10分钟。继续执行步骤3.4被利用无盖眼镜时。
注意:DAPI是DNA嵌入染料,即污迹细胞核。 - 应用10微升安装解决方案到每个显微镜载玻片。挂载每个玻璃盖在载玻片上的安装解决方案。
- 在黑暗中过夜孵育在室温下。
- 密封的玻璃盖用指甲油的边缘。在通风橱内的暗放置过夜以除去指甲油烟雾。
- 孵育1毫升DNA嵌入染料4',6-二脒基-2-苯基的细胞(DAPI; 0.5微克/毫升)的PBS在室温下,在黑暗中进行10分钟。继续执行步骤3.4被利用无盖眼镜时。
- 荧光显微镜19
- 第二天,研究使用40倍物镜荧光显微镜细胞。过滤套的DAPI,CFP,FITC和Cy3的是(Ex350纳米/ Em460纳米),(Ex436纳米/ Em480纳米),(Ex470纳米/ Em525毫微米)和(Ex545纳米/ Em605纳米),分别。
结果
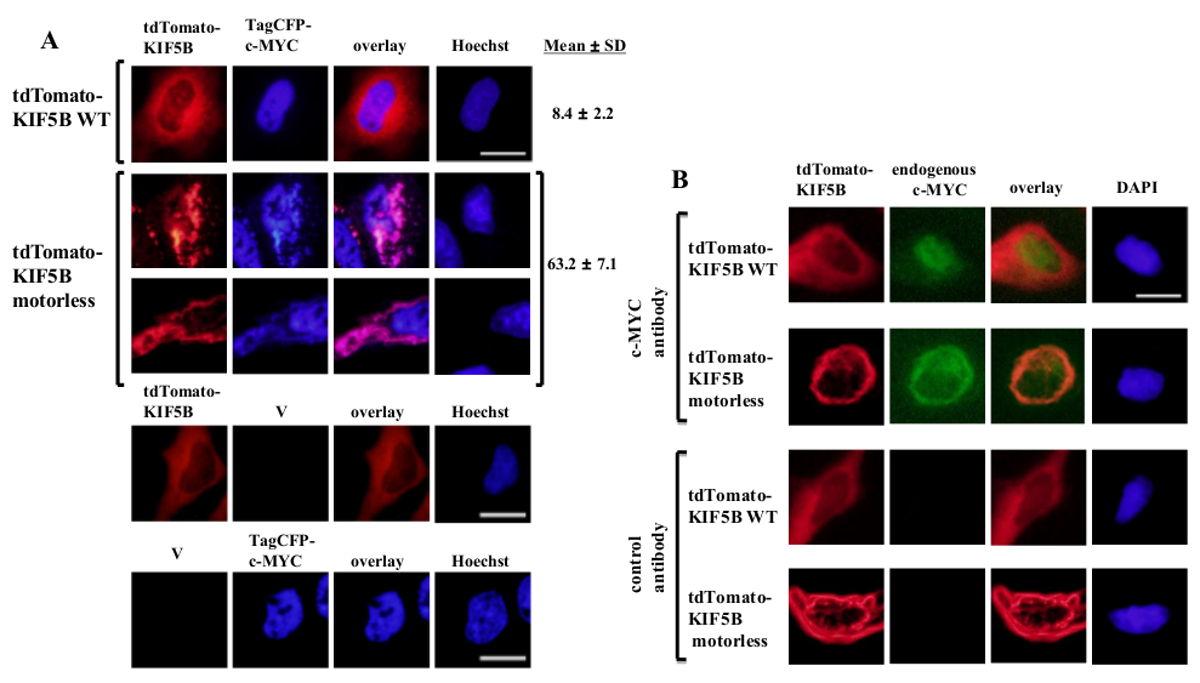
外源性表达的野生型KIF5B在细胞质中均匀出现,而c-myc的主要出现在细胞核中(图1A)。但是,无电机KIF5B突变体形成在细胞质中(图1A)的聚集体。无电机KIF5B聚集诱导的c-MYC的聚集。细胞比例合计将表达TagCFP标记的c-MYC表达野生型KIF5B细胞是很低的。然而,这是当无电机KIF5B被共表达(图1A)显著更高。用C-MYC突变KIF5B观察到的共定位(图1)表明,KIF5B调节的c-Myc的亚细胞定位和c-MYC是驱动蛋白1的货物。在该构建体单独表达阴性对照被包括在图1A的下面板。结果表明:有荧光emissio的显著渗出通过无ñ。无电机KIF5B也诱导内源性c-Myc的(图1B)和转录因子p53的(图2)的聚集,这表明 c-myc和P53都是驱动蛋白-1的货物和KIF5B调节内源性蛋白的亚细胞定位。一起的结果是,驱动蛋白1抑制剂玫瑰红内酯(RBL)20诱导形成高分子量物种的两个c-myc和p53的5,p53是有可能驱动蛋白-1的货物。有趣的是,要注意的是,核转录因子的p53,如c-MYC,也从核输出到细胞质为蛋白酶体降解21,22。转录因子由KIF5B运动似乎是特定因为无电机KIF5B突变体的表达没有影响的亚细胞定位的c-Fos蛋白或使其聚集( 图 3)。上述数据表明,方法具本出版物中使用ð允许驱动蛋白1货物的高特异性识别。类似的策略可以应用到其它动力蛋白为好。

图1:无电机KIF5B突变体的表达诱导在细胞质中的c-Myc聚合 (A)中的HeLa细胞用tdTomato标记的野生型(WT)KIF5B(红色)和TagCFP标记的c-MYC(蓝色)转染。 tdTomato标记WT KIF5B主要出现在细胞质中,而TagCFP标记的c-MYC主要出现在细胞核中。然而,tdTomato标记无电机KIF5B突变体(红色)形成在细胞质丝状或点状的聚集体。引起无电机KIF5B表达的c-MYC的聚集。突变KIF5B和c-MYC共定位在一起(粉红色)。说明CFP-C-MYC(%)的聚集细胞(%)百分比显示。结果示如平均值±SD(N = 3)。与tdTomato标记KIF5B蛋白质或空载体(V)的沿TagCFP标记的c-MYC表达的阴性对照示于下面板。 (B)中类似的结果与内源性c-Myc的(绿色)时表达tdTomato标记WT或无电机KIF5B蛋白(红色)获得。所述tdTomato标记无电机KIF5B突变体形成在细胞质聚集体和诱导的内源c-Myc的聚集。两种聚集体在细胞质中(橙色)共定位在一起。细胞核用染料赫斯特33342或DAPI染色。比例尺; 20微米。 请点击此处查看该图的放大版本。
{kind=link}

图2:无动力KIF5B突变表达诱导在T内源性p53的聚集他细胞质中。在HeLa细胞中,外源tdTomato标记的野生型(WT)KIF5B(红色)主要出现在细胞质中,而内源性p53蛋白(绿色)主要出现在细胞核中。与此相反,tdTomato标记无电机KIF5B突变体(红色)形成在细胞质聚集体。诱导无电机KIF5B的表达p53的聚集,从而导致KIF5B的共定位和p53在细胞质中(黄色)。该实验进行三次。细胞核用染料DAPI染色。比例尺; 20微米。 请点击此处查看该图的放大版本。
{kind=link}

图3:无电机KIF5B突变体的表达不诱导的c-Fos蛋白的聚集在细胞质 tdTomato标记的野生型(WT)KIF5B(红色)出现主要在HeLa细胞的细胞质中,而EGFP-c-fos(绿色)主要出现在细胞核中。与此相反,在tdTomato标记无电机KIF5B突变体(红色)形成在细胞质聚集体。无电机KIF5B的表达并没有诱导c-Fos蛋白的聚集。该实验进行四次。细胞的细胞核与染料DAPI染色。比例尺; 20微米。 请点击此处查看该图的放大版本。
{kind=link}
| 常见的反向引物 | 5'-AGAGGATCCTTACACTTGTTTGCCTCCTC-3' |
| 野生型KIF5B正向引物 | 5'AGAGTCGACGCGGACCTGGCCGAGTGCAACATCAAAGT-3' |
| 无动力KIF5B正向引物 | 5'AGAGTCGACGATGAAGAGTTCACTGTTGC-3' |
表1:用于克隆野生型和无电机KIF5B蛋白的引物序列。
讨论
所描述的方法利用无电机KIF5B突变体,它缺乏沿微管移动的能力的性质,但保留以形成具有野生型KIF5B二聚体和,由此,允许所述四聚体蛋白具有相同货物蛋白质相互作用的能力作为野生型KIF5B。无电机KIF5B,因此,作为一个显性负突变体和形式的错误定位,其货物聚集。这种方法被证明识别驱动蛋白1货物的c-MYC(图1)5。在本文中,相同的无电机KIF5B突变体来鉴定的p53作为驱动蛋白1的另一个潜在的货物( 图2)。这表明,该方法是确定驱动蛋白1的其它货物可行的。此外,该突变体的特异性是由缺乏对阴性对照蛋白c-Fos蛋白(图3)的突变体的效果提供。
在这个协议中,tdTomato标记的野生TYPE或无电机KIF5B蛋白共表达与另一种荧光蛋白标记的候选蛋白质的货物。在这种情况下,活细胞的荧光显微镜和成像中进行。聚集体的形成可通过时间推移成像进行跟踪。可替代地,tdTomato标记蛋白可以单独表达,并在其生理水平的候选货物蛋白是通过使用特异性抗体的间接免疫荧光显微镜观察。荧光蛋白tdTomato被选择用于其亮度和光17。如果背景是由于完全的DMEM培养基自动荧光高,则使用不含酚红介质。
无电机KIF5B突变体形成具有野生型KIF5B化学计量二聚体。因此,它是表达无马达KIF5B突变体的足够量的,以形成与野生型对应二聚体以抑制其功能,并形成聚集体的关键。为了解决这个问题,expr的优化无电机突变体的分裂国家是必需的。它是通过使用含有二聚结构域的小无马达的突变来实现。此外,适当的转染试剂和协议的优化也是必要的。 HeLa细胞的培养用DNA /转染试剂的持续时间在HeLa细胞中被优化为外源蛋白质和细胞活力的表达。孵育持续时间可以具有用于其它细胞系进行优化。
对于4X之间的放大倍数40X的,不需要盖眼镜。细胞可以直接检查在孔中。因此,在这种情况下,该协议是便宜和方便。对于以上40X放大倍数,将细胞生长在盖玻片在孔中和,染色后,被安装在显微镜载片上为下油浸物镜检查。
无电机KIF5B蛋白质的聚集体的观察由细胞质的大小的限制。这是很容易检测的聚集体在许多哺乳动物细胞。然而,这是相对难以观察中的神经元细胞的聚集体时,细胞质的体积大约是细胞核和在突起小。
该协议是用于显示除了在体内酵母双杂交和生化下拉实验13-16 KIF5B及其货物之间的关联。所有这些试验确定不同的身体状况下,联想及其结果可以互相补充。此外,该荧光测定在其它测定的优点在于,它可以显示由KIF5B的货物的亚细胞定位的调节( 图 1和2)。
的协议不局限于KIF5B,可用于识别其它马达蛋白的货物如其他一些驱动蛋白马达 3和一些在货物3的胞内运输中使用的肌球蛋白马达23,24的。这些马达蛋白还包含用于沿微管驱动蛋白电机和肌球蛋白马达微丝,以及用于低聚卷曲螺旋区段移动电机域。他们中的大多数形成二聚体3,23。因此,类似的策略可以通过创建自己的无动力显性负突变体,以确定他们的货物适用于他们。
披露声明
这篇文章的免费接入出版和生产由罗氏公司支付。
致谢
Roche's Publication Grant covered the Free Access publication and production of this article. The author also thanks E. Premkumar Reddy, Richard V. Mettus, Stephen C. Cosenza, Sau Ying Yip and Sol D. Gloria for their support and critical reading of the manuscript.
材料
| Name | Company | Catalog Number | Comments |
| absolute ethanol (200 proof) | Fisher Scientific | BP2818 | |
| DAPI | Sigma-Aldrich | D9542 | |
| Hoechst 33342 | Sigma-Aldrich | B2261 | |
| Opti-MEM-I (transfection medium) | Life Technologies | 51985 | |
| ProLong Diamond Antifade Mountant | Life Technologies | P36961 | |
| formaldehyde, para | Fisher Scientific | O4042-500 | |
| Triton-X100 | Fisher Scientific | BP151-500 | |
| X-tremeGENE 9 DNA Transfection Reagent | Roche | 6365787001 | |
| Taq DNA polymerase | Life Technologies | 10342020 | |
| PCR Grade Nucleotide Mix (dNTP Mix) | Roche | 12111424 | |
| Microscope cover glass | Fisher Scientific | 12-541A | |
| GeneRuler 1 kb Plus DNA ladder | Life Technologies | SM1333 | |
| PureLink Quick Gel Extraction kit | Life Technologies | K210012 | |
| BamHI | New England Biolab | R0136 | |
| SalI | New England Biolab | R0138 | |
| T4 DNA ligase | New England Biolab | M0202T | |
| Ethidium bromide | Thermo Scientific | 17898 | |
| DMEM | Life Technologies | 11995-065 | |
| c-MYC rabbit antibody | Cell Signaling | 5606 | |
| p53 mouse antibody | Santa Cruz | sc-126 | |
| Alexa Fluor 488-conjugated anti-rabbit IgG antibody | Life Technologies | A11008 | |
| Alexa Fluor 488-conjugated anti-mouse IgG antibody | Life Technologies | A11059 | |
| fluorescent microscope | Olympus IX71_Fluoview | ||
| computer software for imaging | cellSens |
参考文献
- Hirokawa, N., Niwa, S., Tanaka, Y. Molecular motors in neurons: transport mechanisms and roles in brain function, development, and disease. Neuron. 68, 610-638 (2010).
- Yu, Y., Feng, Y. M. The role of kinesin family proteins in tumorigenesis and progression: potential biomarkers and molecular targets for cancer therapy. Cancer. 116, 5150-5160 (2010).
- Verhey, K. J., Hammond, J. W. Traffic control: regulation of kinesin motors. Nat Rev Mol Cell Biol. 10, 765-777 (2009).
- Hirokawa, N., Noda, Y., Tanaka, Y., Niwa, S. Kinesin superfamily motor proteins and intracellular transport. Nat Rev Mol Cell Biol. 10, 682-696 (2009).
- Lee, C. M. Transport of c-MYC by Kinesin-1 for proteasomal degradation in the cytoplasm. Biochim Biophys Acta. 1843, 2027-2036 (2014).
- Bittins, C. M., Eichler, T. W., Hammer, J. A., Gerdes, H. H. Dominant-negative myosin Va impairs retrograde but not anterograde axonal transport of large dense core vesicles. Cell Mol Neurobiol. 30, 369-379 (2010).
- Kimura, T., Watanabe, H., Iwamatsu, A., Kaibuchi, K. Tubulin and CRMP-2 complex is transported via Kinesin-1. J Neurochem. 93, 1371-1382 (2005).
- Lindsay, A. J., McCaffrey, M. W. Myosin Va is required for the transport of fragile X mental retardation protein (FMRP) granules. Biol Cell. 106, 57-71 (2014).
- Rivera, J., Chu, P. J., Lewis, T. L., Arnold, D. B. The role of Kif5B in axonal localization of Kv1 K(+) channels. Eur J Neurosci. 25, 136-146 (2007).
- Roland, J. T., Kenworthy, A. K., Peranen, J., Caplan, S., Goldenring, J. R. Myosin Vb interacts with Rab8a on a tubular network containing EHD1 and EHD3. Mol Biol Cell. 18, 2828-2837 (2007).
- Uchida, A., Alami, N. H., Brown, A. Tight functional coupling of kinesin-1A and dynein motors in the bidirectional transport of neurofilaments. Mol Biol Cell. 20, 4997-5006 (2009).
- Zadeh, A. D., et al. Kif5b is an essential forward trafficking motor for the Kv1.5 cardiac potassium channel. J Physiol. 587, 4565-4574 (2009).
- Su, Y. Y., et al. KIF5B promotes the forward transport and axonal function of the voltage-gated sodium channel Nav1.8. J Neurosci. 33, 17884-17896 (2013).
- Lalioti, V. S., Vergarajauregui, S., Tsuchiya, Y., Hernandez-Tiedra, S., Sandoval, I. V. Daxx functions as a scaffold of a protein assembly constituted by GLUT4, JNK1 and KIF5B. . J Cell Physiol. 218, 416-426 (2009).
- Cho, K. I., et al. Association of the kinesin-binding domain of RanBP2 to KIF5B and KIF5C determines mitochondria localization and function. Traffic. 8, 1722-1735 (2007).
- Diefenbach, R. J., Diefenbach, E., Douglas, M. W., Cunningham, A. L. The ribosome receptor, p180, interacts with kinesin heavy chain, KIF5B. Biochem Biophys Res Commun. 319, 987-992 (2004).
- Shaner, N. C., et al. Improved monomeric red, orange and yellow fluorescent proteins derived from Discosoma sp. red fluorescent protein. Nat Biotechnol. 22, 1567-1572 (2004).
- Waters, J. C. Live-cell fluorescence imaging. Methods Cell Biol. 114, 125-150 (2013).
- Sanderson, M. J., Smith, I., Parker, I., Bootman, M. D. Fluorescence microscopy. Cold Spring Harb Protoc. 2014, (2014).
- Hopkins, S. C., Vale, R. D., Kuntz, I. D. Inhibitors of kinesin activity from structure-based computer screening. Biochemistry. 39, 2805-2814 (2000).
- Zhang, Y., Xiong, Y. Control of p53 ubiquitination and nuclear export by MDM2 and ARF. Cell Growth Differ. 12, 175-186 (2001).
- Michael, D., Oren, M. The p53-Mdm2 module and the ubiquitin system. Semin Cancer Biol. 13, 49-58 (2003).
- Kneussel, M., Wagner, W. Myosin motors at neuronal synapses: drivers of membrane transport and actin dynamics. Nat Rev Neurosci. 14, 233-247 (2013).
- Maravillas-Montero, J. L., Santos-Argumedo, L. The myosin family: unconventional roles of actin-dependent molecular motors in immune cells. J Leukoc Biol. 91, 35-46 (2012).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。