
Method Article
تصور بؤر الحمض النووي المفردة التي تقطعت بها السبل في المرحلة G1 من دورة الخلية
In This Article
Summary
يقدم البروتوكول التالي الكشف عن بؤر الحمض النووي التي تقطعت بها السبل في المرحلة G1 من دورة الخلية باستخدام مزامنة دورة الخلية متبوعة بتلطيخ RPA2 المناعي.
Abstract
خصص الحمض النووي مسارات إصلاح خلوية قادرة على التعامل مع الآفات التي يمكن أن تنشأ من مصادر داخلية و / أو خارجية. يتطلب إصلاح الحمض النووي التعاون بين العديد من البروتينات ، المسؤولة عن تغطية مجموعة واسعة من المهام من التعرف على وجود آفة الحمض النووي والإشارة إليها إلى إصلاحها جسديا. خلال هذه العملية ، غالبا ما يتم إنشاء مسارات الحمض النووي أحادي الشريط (ssDNA) ، والتي يتم ملؤها في النهاية بواسطة بوليميراز الحمض النووي. طبيعة مسارات ssDNA هذه (من حيث الطول والعدد) ، جنبا إلى جنب مع البلمرة التي تم تجنيدها لملء هذه الفجوات ، هي مسار إصلاح محدد. يمكن أن يساعدنا تصور مسارات ssDNA هذه في فهم الديناميات المعقدة لآليات إصلاح الحمض النووي.
يوفر هذا البروتوكول طريقة مفصلة لإعداد الخلايا المتزامنة G1 لقياس تكوين بؤر ssDNA عند الإجهاد السمي الجيني. باستخدام نهج التألق المناعي سهل الاستخدام ، نتصور ssDNA عن طريق تلطيخ RPA2 ، وهو أحد مكونات مركب بروتين النسخ المتماثل غير المتجانس A (RPA). ترتبط تقنية RPA2 بوسيطات ssDNA التي تنشأ عند الإجهاد السمي الجيني أو النسخ المتماثل وتثبيطها للتحكم في إصلاح الحمض النووي وتنشيط نقطة فحص تلف الحمض النووي. يستخدم تلطيخ 5-Ethynyl-2'-deoxyuridine (EdU) لتصور تكرار الحمض النووي لاستبعاد أي خلايا طور S. يوفر هذا البروتوكول نهجا بديلا للمقايسات التقليدية القائمة على 5-برومو-2'-ديوكسييوريدين (BrdU) غير المسخ وهو أكثر ملاءمة للكشف عن بؤر ssDNA خارج المرحلة S.
Introduction
للحفاظ على الحياة ، تقوم الخلايا باستمرار بمسح وإصلاح الحمض النووي للحفاظ على سلامتها الجينومية. قد تتراكم الخلايا أنواعا مختلفة من تلف الحمض النووي بسبب كل من المصادر الداخلية (على سبيل المثال ، الأكسدة ، الألكلة ، نزع الأمين ، أخطاء النسخ المتماثل) والخارجية (مثل الأشعة فوق البنفسجية ، التشعيع المؤين) لضغوط الحمض النووي. يؤدي الفشل في إصلاح هذه الآفات إما إلى موت الخلايا المبرمج أو توقف دورة الخلية أو الشيخوخة ويمكن أن يؤدي إلى الأمراض1. يمكن معالجة آفات الحمض النووي من خلال أي من مسارات إصلاح الحمض النووي الرئيسية التالية: DR (إصلاح الانعكاس المباشر) ، والذي يصلح بشكل أساسي قواعد الألكيلات2 ؛ BER (إصلاح استئصال القاعدة) ، والذي يستهدف أخطاء قواعد الحمض النووي غير الضخمة وفواصل الحمض النووي أحادية الشريط (SSBs) 3 ؛ NER (إصلاح استئصال النوكليوتيدات) تصحيح آفات الحمض النووي الضخمة المشوهة للحلزون4 ؛ MMR (إصلاح عدم التطابق) يستهدف بشكل أساسي عدم تطابق الحمض النووي ، وحلقات الإدراج / الحذف (IDLs) ، وبعض الأضرار الأساسية5 ؛ NHEJ (الانضمام النهائي غير المتماثل) و HRR (إصلاح إعادة التركيب المتماثل) كلاهما نشط في فواصل الحمض النووي المزدوجة التي تقطعت بها السبل (DSBs)6 ؛ و TLS (تخليق الآفة) ، وهي آلية تجاوز آفة الحمض النووي7. على الرغم من أن هذه المسارات لها خصائص ركيزة مميزة ، إلا أن هناك بعض التداخلات بينها لضمان التكرار للإصلاح الفعال. يعد فهم عمل مسارات إصلاح الحمض النووي المختلفة في مراحل دورة الخلية المختلفة أمرا بالغ الأهمية لأن عوامل إصلاح الحمض النووي هذه يمكن أن تكون بمثابة أهداف أساسية للنهج العلاجية لعلاج السرطان والشيخوخة والاضطرابات العصبية 8,9.
يتم إنشاء الحمض النووي أحادي الشريط (ssDNA) طوال دورة الخلية بسبب إصلاح آفات الحمض النووي الناتجة عن كل من العوامل الضارة بالحمض النووي الداخلية والخارجية. عند الإجهاد السام الجيني ، يتم إنشاء ssDNA بكثرة في المرحلتين S و G2 حيث يكون ل HRR و MMR أعلى نشاط لهما وعندما تتوقف آلية النسخ المتماثل أو تنهار عند مواجهة آفات الحمض النووي6،10،11. مسارات إصلاح الحمض النووي الأخرى (على سبيل المثال ، NHEJ / الانضمام النهائي بوساطة microhomology (MMEJ) / التلدين أحادي الشريط [SSA]) تولد أيضا ssDNA أثناء إصلاح DSB12. عادة ما تنشأ مسارات ssDNA هذه من استئصال الحمض النووي ، الذي يتم تنفيذه بواسطة exonucleases مثل EXO1 و DNA2 و CtIP أثناء HR و MMR ، والنوكليازات الداخلية مثل XPF و XPG أثناء NER ، أو من خلال العمل المشترك ل POLB و FEN1 خلال BER4،13،14،15،16،17،18،19. نظرا لعمل آلية النسخ المتماثل ، يتم إنشاء مسارات ssDNA أيضا عندما تقوم حلزونات الحمض النووي بفك الحمض النووي أمام البلمرة المتماثلة المرتبطة بPCNA 20. في المقابل ، في المرحلة G1 ، يقلل نقص HRR وتكرار الحمض النووي والنشاط المحدود ل MMR من مدى مسارات ssDNA المتولدة وبالتالي يكون اكتشاف10،11،21 أكثر صعوبة.
مسارات ssDNA الخلوية هي هياكل حساسة للغاية يجب حمايتها لتجنب تكوين DSBs. يتم تحقيق ذلك عن طريق طلاء مسارات ssDNA باستخدام RPA. RPA عبارة عن مركب بروتين غير متجانس وفير يتكون من وحدات فرعية متعددة (RPA1 و RPA2 و RPA3 ، يشار إليها أيضا باسم RPA70 و RPA32 و RPA14 ، على التوالي) ، والتي يتم التعبير عنها في كل مكان طوال دورة الخلية22. تحتوي كل وحدة فرعية RPA على مجال ربط الحمض النووي (DBD) ، قادر على التفاعل مع 4-6 نيوكليوتيدات ، وتشكل الوحدات الفرعية المدمجة نواة تشذيب مستقرة. إجمالا ، يرتبط RPA بحوالي 20-30 نيوكليوتيدات مع تقارب شبه نانوي23,24.
تستخدم الطرق التقليدية الفحص المجهري المناعي (IF) لتصور بؤر ssDNA عن طريق وضع علامات على 5-bromo-2'-deoxyuridine (BrdU) المدمجة في الحمض النووي الجينومي باستخدام الأجسام المضادة BrdU25. يعتمد هذا النهج على حقيقة أن الأجسام المضادة BrdU يمكنها فقط اكتشاف BrdU في ssDNA25 المكشوف. على الرغم من أن هذا النهج واضح ومباشر ، إلا أنه يعرض أيضا بعض القيود. على سبيل المثال ، تتم معالجة الخلايا مسبقا لدمج BrdU قبل بدء التجربة ، وهو أمر يستغرق وقتا طويلا ويمكن أن يتداخل مع المستجيبات النهائية. لذلك ، يقتصر اكتشاف ssDNA المستند إلى BrdU على تكرار الخلايا ولا يمكن استخدامه للخلايا الهادئة. هذا يستثني تطبيق هذه الطريقة لدراسة إصلاح الحمض النووي في الخلايا غير المتكاثرة على الرغم من أهميتها في العديد من الأمراض مثل السرطان والتنكس العصبي 5,26. بالإضافة إلى ذلك ، نظرا لأن هياكل BrdU و EdU متشابهة جدا ، فإن معظم الأجسام المضادة BrdU تظهر تفاعلا متقاطعا تجاه EdU ، والذي يجب مراعاته عند استهداف تجارب وضع العلامات المزدوجة27. تم استخدام تلطيخ RPA سابقا لإظهار بؤر ssDNA بشكل رئيسي في خلايا المرحلة S. ومع ذلك ، فقد استخدمته بعض الأوراق بنجاح خارج المرحلة S28،29،30،31،32،33،34،35. يستخدم البروتوكول التالي خصائص RPA بكفاءة ، مما يسمح بتصور بؤر ssDNA بعد تلف الحمض النووي في المرحلة G1 من دورة الخلية (على الرغم من أنه يمكن استخدامه في جميع مراحل دورة الخلية).
Protocol
1. صيانة الخلايا الظهارية الصبغية الصباغية للشبكية المخلدة hTERT (RPE1)
- الحفاظ على خطوط خلايا RPE1 في وسط النسر المعدل (DMEM) من Dulbecco المكمل بمصل بقري جنيني معطل حراريا بنسبة 10٪ (Hi-FBS) و 100 ميكروغرام / مل من البنسلين-الستربتومايسين (يشار إليه باسم وسط الاستزراع من الآن فصاعدا) في حاضنة مرطبة مع 5٪ CO2 عند 37 درجة مئوية. للزراعة الروتينية ، تنمو خلايا RPE1 في طبق معالج بزراعة الأنسجة 15 سم وتنقسم عند الوصول إلى التقاء 80-90٪ (~ 16-18 × 106 خلايا لكل طبق 15 سم).
- عند الانقسام ، قم بإزالة الوسط وشطف الخلايا ب 10 مل من 1x محلول ملحي مخزن بالفوسفات (PBS).
- أضف 3 مل من 0.05٪ Trypsin-EDTA لتغطية سطح الطبق بالكامل. الحفاظ على الخلايا في 37 درجة مئوية مع التربسين حتى تنفصل.
- بعد التربسين، يعاد تعليق الخلايا بوسط الاستزراع وتدويرها عند 150 × جم لمدة 5 دقائق في درجة حرارة الغرفة (RT، 22-25 درجة مئوية). قم بإزالة المادة الطافية وأعد تعليق الخلايا برفق في 10 مل من وسط الاستزراع.
- البذور 1.6-1.8 × 106 خلايا في طبق جديد 15 سم (~ 1 مل من تعليق الخلية).
ملاحظة: يجب أن تتم جميع أعمال زراعة الأنسجة تحت مستويات أمان BSL-2. يعتمد وقت حضانة التربسين على التقاء الخلايا. عادة ، تستغرق العملية 2-3 دقائق لإكمالها للحصول على لوحة متقاربة بنسبة 90٪. يجب فحص الخلايا بحثا عن تلوث الميكوبلازما على أساس منتظم باستخدام مجموعات متاحة تجاريا (انظر الأمثلة في جدول المواد).
2. siRNA يهدم الجين محل الاهتمام (GOI)
- بذرة 1.0 × 106 خلايا RPE1 في صفيحة معالجة بزراعة الأنسجة 10 سم مع 10 مل من وسط الاستزراع في اليوم السابق للنقل.
- في يوم النقل ، مجمع siRNA. بالنسبة للوحة 10 سم ، استخدم تركيزا نهائيا قدره 20 نانومتر siRPA2 و 12 ميكرولتر من كاشف النقل القائم على الدهون في 500 ميكرولتر من وسط نقل المصل المنخفض. امزج جميع المكونات برفق عن طريق تحريك الأنبوب واحتضانه في RT (22-25 درجة مئوية) لمدة 5 دقائق.
- أضف خليط siRNA المعقد إلى الخلايا بالتنقيط واحتضان الخلايا باستخدام siRNA لمدة 48 ساعة.
3. مزامنة خلايا RPE1 في المرحلة G0
- التربسين خلايا RPE1 من الخطوة 2.3 كما هو موضح في القسم 1 (~ 2 × 106 خلايا).
- نقل تعليق الخلية إلى أنابيب الطرد المركزي 15 مل والطرد المركزي في 150 × غرام ، RT (22-25 درجة مئوية) لمدة 5 دقائق.
- قم بإزالة المادة الطافية وإعادة تعليق الخلايا في 12 مل من برنامج تلفزيوني. أجهزة الطرد المركزي الخلايا في 150 × غرام في RT (22-25 درجة مئوية) لمدة 5 دقائق. كرر إزالة المادة الطافية والطرد المركزي مرتين.
- أعد تعليق الخلايا في 10 مل من DMEM الخالي من المصل المكمل ب 100 ميكروغرام / مل من البنسلين - الستربتومايسين ، 1 مليمتر بيروفات الصوديوم ، 15 مللي متر HEPES ، ووضعها في طبق زراعة الأنسجة 10 سم.
ملاحظة: إذا كانت الخلايا تميل إلى التكتل ، فأعد تعليقها في 1 مل فقط من DMEM الخالي من المصل وماصة لأعلى ولأسفل 5x باستخدام طرف P1000 لإزاحة الكتل قبل تخفيف التعليق حتى الحجم النهائي 10 مل. - بعد 24 ساعة من تجويع المصل ، أدخل الجولة الثانية من الإسكات باستخدام نفس الإجراء كما هو موضح في القسم 2 عن طريق إضافة siRNA المعقد إلى الخلايا المتعطشة للمصل.
- احتفظ بخلايا RPE1 في DMEM الخالي من المصل لمدة 72 ساعة قبل الانتقال إلى إصدار G1.
4. طلاء Coverslip وإطلاق الخلايا في المرحلة G1
- تعقيم الملقط بنسبة 70٪ من الإيثانول ووضع غطاء زجاجي واحد (قطره 12 مم وسمك # 1.5 [0.17 مم]) في بئر من 24 بئرا.
- قم بتخفيف مصفوفة طلاء الفيترونيكتين باستخدام برنامج تلفزيوني للحصول على تركيز نهائي قدره 10 ميكروغرام / مل. أضف 500 ميكرولتر من محلول الفيترونيكتين في كل بئر يحتوي على أغطية الغطاء واحتضانها لمدة 1 ساعة في RT.
- قم بإزالة محلول الطلاء وغسل أغطية الغطاء ب 1 مل من PBS.
- افصل خلايا RPE1 المتعطشة للمصل عن الصفيحة المعالجة بالأنسجة المزروعة بطول 10 سم باستخدام 1 مل من 0.05٪ تربسين بعد غسل PBS لمدة دقيقة واحدة عند 37 درجة مئوية.
ملاحظة: تنفصل الخلايا بشكل أسرع بعد تجويع المصل. توخي الحذر عند غسل الخلايا مع برنامج تلفزيوني واستخدام أوقات التربسين القصيرة. - لتعطيل التربسين ، أعد تعليق خلايا RPE1 في إجمالي 6 مل من وسط الاستزراع. قم بإزالة التربسين المعطل عن طريق تدوير الخلايا باستخدام 150 × جم عند RT (22-25 درجة مئوية) لمدة 5 دقائق.
- أعد تعليق الخلايا في 1 مل من وسط الزراعة وقم بقياس رقم الخلية.
- بذرة 4 × 104 خلايا RPE1 على غطاء الغطاء المطلي في ما مجموعه 500 ميكرولتر من وسط الاستزراع.
ملاحظة: تأكد من أن صلاحية الخلية أعلى من 90٪ قبل المتابعة إلى خطوات المصب. يمكن تقييم صلاحية الخلية بسرعة عن طريق تلطيخ التريبان الأزرق أثناء خطوة عد الخلايا. - بعد 6 ساعات من طلاء الخلايا في وسط الاستزراع ، ستكون الخلايا التي تم إطلاقها G0 في مرحلة G1 المبكرة. قم بإجراء تجارب في G1 في هذه النافذة من 6 إلى 12 ساعة قبل أن تبدأ الخلايا في دخول المرحلة S.
- قبل إدخال تلف الحمض النووي ، قم بنبض الخلايا ب 10 ميكرومتر 5-إيثينيل -2'-ديوكسوريدين (EdU) لمدة 30 دقيقة عند 37 درجة مئوية ، مخففة في وسط الاستزراع.
- قم بإزالة الوسط الذي يحتوي على EdU وطارد الخلايا باستخدام 10 ميكرومتر ثيميدين لمدة 10 دقائق عند 37 درجة مئوية لمنع دمج EdU المتبقي أثناء تحريض تلف الحمض النووي.
- إزالة الوسط مع ثيميدين وعلاج الخلايا مع 250 μMH 2O2 لمدة 1 ساعة ، مخففة في وسط الاستزراع.
5. تلطيخ التألق المناعي ل ssDNA
- اغسل الخلايا مرة واحدة باستخدام 1 مل من RT (22-25 درجة مئوية) PBS لإزالة مكونات الوسط والمصل.
ملاحظة: كن لطيفا عند غسل الخلايا لتجنب الانفصال والتجفيف. لا تقم بمعالجة العديد من الآبار في نفس الوقت. - ما قبل الاستخراج: احتضان الخلايا المغسولة في 1 مل من محلول استخراج CSK (الجدول 1) لمدة 5 دقائق في RT (22-25 درجة مئوية).
ملاحظة: يزيل الاستخراج المسبق ل CSK جميع البروتينات غير المرتبطة بالكروماتين ، بما في ذلك RPA2 القابل للذوبان.
تنبيه: Triton X-100 ضار إذا تم ابتلاعه وقد يسبب تهيج الجلد وتلف العين. - قم بإزالة المخزن المؤقت CSK من الخلايا وإصلاحه مباشرة عن طريق إضافة 0.5 مل من محلول بارافورمالدهايد 3.6٪ (في PBS) يحتوي على 0.05٪ Triton X-100 لمدة 10 دقائق عند RT (22-25 درجة مئوية).
تنبيه: من المهم تحضير 3.6٪ PFA من 32٪ PFA طازجة. قد يسبب بارافورمالدهيد تلفا خطيرا للعين وتهيجا للجلد وتهيجا في الجهاز التنفسي. - اغسل الخلايا مرة واحدة باستخدام 1 مل من PBS تحتوي على 0.05٪ Triton X-100 لإزالة PFA.
- مزيد من نفاذية الخلايا باستخدام 1 مل من PBS تحتوي على 0.5٪ Triton X-100 لمدة 15 دقيقة عند RT (22-25 درجة مئوية).
- تفاعل EdU click-IT لتصور الخلايا المكررة (المرحلة S)
- قم بإزالة محلول النفاذية وغسل الخلايا 2x باستخدام 1 مل من المخزن المؤقت للحظر (الجدول 1).
تنبيه: قد يسبب ألبومين مصل الأبقار (BSA) تهيج الجهاز التنفسي. - أضف 1 مل من المخزن المؤقت المانع (الجدول 1) وهز اللوحة المحتوية على غطاء برفق لمدة 10 دقائق عند RT (22-25 درجة مئوية).
- قم بإزالة المخزن المؤقت المانع وأضف 500 ميكرولتر من كوكتيل تفاعل النقر الذي يحتوي على picolyl azide 647 (الجدول 1). احتضان أغطية الغطاء لمدة 30 دقيقة في RT (22-25 درجة مئوية) باستخدام هزاز لطيف وقم بإجراء حضانات في اتجاه مجرى النهر في الظلام.
ملاحظة: عند استخدام الأجسام المضادة BrdU ، استخدم ضعف الكمية (1 مل) والوقت (60 دقيقة) لتفاعل النقر على النحو الموصى به من قبل الشركة المصنعة للتأكد من أن التفاعل مشبع وأن EdU المدمج موسوم. هذا يحد من التفاعل المتبادل للأجسام المضادة BrdU27.
- قم بإزالة محلول النفاذية وغسل الخلايا 2x باستخدام 1 مل من المخزن المؤقت للحظر (الجدول 1).
- قم بإزالة خليط تفاعل النقر واغسل الخلايا 2x باستخدام PBS مع 0.05٪ Triton X-100 لمدة 10 دقائق عند RT (22-25 درجة مئوية) (الشكل 1 والشكل 2).
- أضف 1 مل من المخزن المؤقت المانع واحتضانه في RT (22-25 درجة مئوية) لمدة 30 دقيقة. بدلا من ذلك ، احتفظ بالخلايا في المخزن المؤقت عند 4 درجات مئوية طوال الليل.
- ضع الجسم المضاد الأساسي (فأر مضاد ل RPA2 ، تخفيف 1: 1000) لمدة ساعتين عند RT (22-25 درجة مئوية) في 250-500 ميكرولتر من المخزن المؤقت المانع مع التأرجح اللطيف.
- اغسل الخلايا 2x باستخدام برنامج تلفزيوني يحتوي على 0.05٪ Triton X-100 لإزالة معظم محلول الأجسام المضادة بسرعة.
- استمر في غسل الخلايا لمدة 3 × 10 دقائق مع حاجز مانع عند RT (22-25 درجة مئوية).
- تطبيق الجسم المضاد الثانوي (مضاد للفئران Alexa-488 ، 1: 1000 تخفيف) في 250-500 ميكرولتر من المخزن المؤقت المانع عند RT (22-25 درجة مئوية) لمدة 2 ساعة مع هزاز لطيف.
- اغسل الخلايا بمخزن مؤقت مانع 2x لإزالة معظم الأجسام المضادة الثانوية بسرعة. استمر في غسل الخلايا لمدة 3 × 10 دقائق باستخدام PBS الذي يحتوي على 0.05٪ Triton X-100 عند RT (22-25 درجة مئوية).
- لموازنة النوى، اغسل الخلايا مرة واحدة باستخدام برنامج تلفزيوني يحتوي على 0.05٪ Triton X-100 و 1 ميكروغرام / مل 4',6-دياميدينو-2-فينيليندول (DAPI) لمدة 10 دقائق عند RT (22-25 درجة مئوية). اغسل الخلايا مرة واحدة باستخدام PBS لمدة 5 دقائق في RT (22-25 درجة مئوية).
- قم بتركيب زجاج الغطاء على شرائح المجهر باستخدام 10 ميكرولتر من وسيط التثبيت / غطاء الغطاء. اغمس الغطاء في الماء المقطر قبل تركيبه للتخلص من أي بلورات ملحية. تخيل الشرائح في اليوم التالي وقم بتخزينها في درجة حرارة 4 درجات مئوية لأسابيع (الشكل 3).
6. الحصول على الصور والقياس الكمي
- لالتقاط الصور ، استخدم أي مجهر فوق فلوري متاح مزود بمجموعات مرشحات روتينية لتصوير قنوات DAPI و FITC و Cy5 بتكبير 60-63x على الأقل وفتحة رقمية عالية وأهداف زيتية لتصور البؤر النووية.
ملاحظة: الإثارة المثلى DAPI هي ~ 359 نانومتر ؛ الإثارة اليكسا 488 هي ~ 488 نانومتر ؛ بينما الإثارة Alexa 647 هي ~ 647 نانومتر. - لتحليل الصور، افتح ملفات الصور في Fiji/ImageJ.
- اصنع أقنعة نووية باستخدام تلطيخ DAPI (الشكل 4A-F والفيديو التكميلي S1).
- افتح صورة DAPI.
- حدد العملية | تحسين التباين وتعيين بكسل مشبعة إلى 0.35.
- انقر فوق عملية | ثنائي | قم بالتحويل إلى قناع. اختر ثنائي | املأ الثقوب وانقر على تحليل | تحليل الجسيمات. اضبط الحجم على 10 إلى ما لا نهاية.
- في مدير عائد الاستثمار ، انقر فوق إظهار الكل.
- العثور على بؤر RPA2 في النواة (الشكل 4G ، H والفيديو التكميلي S1)
- افتح صورة RPA2.
- اختر العملية | ابحث عن ماكسيما. قم بتعيين البروز إلى قيمة تبرز بؤر RPA2 (بين 500 و750) ، وتفصلها عن الخلفية.
- أخيرا ، انقر فوق الزر "قياس " في مدير عائد الاستثمار.
- احسب العدد الإجمالي لبؤر ssDNA النووية بقسمة القيمة في عمود RawinDen على 255 (القيمة القصوى لكثافة البكسل في كل بؤرة).
- إجراء التحليل الإحصائي باستخدام أداة البرامج الإحصائية المفضلة.
ملاحظة: استبعاد كافة الخلايا الإيجابية EdU وأقنعة DAPI مجزأة بشكل غير صحيح من التحليل.
- اصنع أقنعة نووية باستخدام تلطيخ DAPI (الشكل 4A-F والفيديو التكميلي S1).
النتائج
للتغلب على قيود اكتشاف ssDNA في G1 ، استخدمنا RPA2 ، مما يعزز خصوصية وشدة اكتشاف بؤر ssDNA35. لتحقيق مزامنة دقيقة للخلايا ، استخدمنا خلايا RPE1 التي يمكن تجويعها للأمصال بكفاءة ومزامنتها في مرحلة G0. يمكن بعد ذلك حثهم على العودة إلى دورة الخلية عن طريق إضافة المصل بعد الحرمان من المصل. لتأكيد كفاءة التزامن ، قمنا بتسمية الخلايا ب EdU ومحتوى الحمض النووي الخاص بها ببروبيديوم-يوديد. كما جمعنا نتائج نوعية وكمية من خلال قياس التدفق الخلوي (الشكل التكميلي S1A). تظهر المخططات النقطية أنه بعد 72 ساعة من تجويع المصل ، ~ 98٪ من الخلايا في مرحلة G0. بعد إضافة الوسائط المحتوية على المصل لمدة 6 ساعات ، تعود الخلايا إلى دورة الخلية (كما يتضح من الزيادة في مستويات p27 في الشكل 1A) ، مع وجود ~ 97٪ من الخلايا في G1 ، بينما تحتوي على خلايا <1٪ فقط في المرحلة S ، <2٪ خلايا في المرحلة G2 (الشكل التكميلي S1A). بعد 20-28 ساعة من إضافة المصل إلى الخلايا ، فإنها تمر تدريجيا عبر المرحلة S ، كما هو موضح في مخططات قياس التدفق الخلوي (الشكل التكميلي S1A). يعطي بروتوكول مزامنة الخلايا هذا ~ 97٪ من سكان G1 النقي (6 ساعات بعد إضافة المصل بعد 72 ساعة من تجويع المصل). لمزيد من التحقق من كفاءة التزامن ، قمنا بمقارنة تعبير علامات دورة الخلية بعد إطلاق المصل باستخدام النشاف الغربي (الشكل 1A والشكل التكميلي S1B) وبالتوازي مع ذلك ، أجرينا مقايسة دمج EdU لتصور تكرار الحمض النووي. يسلط تلطيخ EdU الضوء أيضا على كفاءة التزامن وعدم تكرار الحمض النووي في مرحلة G1 (الشكل 1B ، C).
تعتمد الطرق التقليدية للكشف عن ssDNA في خلايا الثدييات على اكتشاف BrdU في ssDNA. يوضح الشكل 2A أنه عند علاج H2O2 و neocarzinostatin (NCS) ، كانت بؤر BrdU قابلة للاكتشاف فقط في خلايا الطور S ، بينما لم يتم اكتشاف بؤر ssDNA في خلايا الطور غير S. أظهر تلطيخ الأجسام المضادة BrdU أيضا تلطيخا ملحوظا للخلفية الأساسية يمكن اكتشافه في جميع النوى ، بغض النظر عن مرحلة دورة الخلية أو العلاجات المطبقة. باستخدام بروتوكول النقر EdU الموضح هنا ، لم نتمكن من اكتشاف التوطين المشترك لبؤر EdU و BrdU ، وهو ما يتضح في العينات غير المعالجة من الشكل 2A. لاستبعاد أي إشارة BrdU نشأت من التفاعل المتقاطع تماما ، تجنبنا وضع العلامات على EdU واستخدمنا بدلا من ذلك cyclin A2 كعلامة S-G2. ومع ذلك ، لم يسمح تلطيخ cyclin A2 بالاستخراج المسبق ل CSK ، وفي ظل هذه الحالة ، لم نر أي بؤر BrdU ، حتى بعد الإجهاد السمي الجيني (الشكل التكميلي S2A). هذا يسلط الضوء على حقيقة أن الاستخراج المسبق ل CSK ضروري لتلطيخ ssDNA القائم على BrdU. كعنصر تحكم ، اختبرنا تلطيخ الأجسام المضادة BrdU في ظل ظروف تغيير طبيعة الطبيعة. هذا يفتح الحمض النووي لفضح BrdU المدمج ، والذي يكشف أن BrdU تم دمجه بشكل موحد (الشكل التكميلي S2B).
في المقابل ، يظهر تلطيخ RPA2 تكوين بؤر تعتمد على NCS و H2O 2 ليس فقط في المرحلة S ولكن أيضا في مراحل دورة الخلية الأخرى (الشكل 2B). كعنصر تحكم ، عالجنا أيضا الخلايا باستخدام HU ، والذي يتسبب فقط في تراكم ssDNA في الخلايا التي تخضع للتكاثر. كما هو متوقع ، اكتشفنا فقط زيادة في الإشارة عند علاج HU بالجسم المضاد RPA2 في الخلايا الإيجابية EdU ، مما يسلط الضوء على خصوصية هذا النهج. يمكن للجسم المضاد RPA2 أيضا اكتشاف تكوين ssDNA الذي يحدث بشكل طبيعي أثناء النسخ المتماثل في غياب الإجهاد السمي الجيني الخارجي (الشكل 2B). دفعتنا الطبيعة الحساسة للغاية للجسم المضاد RPA2 إلى محاولة استخدامه في مرحلة G1 حيث فشل تلطيخ BrdU التقليدي في اكتشاف أي إشارة عند الإجهاد السمي الجيني (الشكل التكميلي S2C). يوضح الشكل 3 أ أن تكوين بؤر ssDNA عند علاج H2O2 كان قابلا للاكتشاف عند استخدام جسم مضاد ل RPA2 ، حتى في G1. كانت هناك زيادة كبيرة في عدد بؤر RPA2 في هذه النوى عند معالجة H2O2 (الشكل 3B). كانت هذه البؤر خاصة ب RPA2 حيث أدى إسكات RPA2 إلى إلغاء إشارة IF (الشكل 3A ، B). يوضح الشكل 3C والشكل التكميلي S1C كفاءة إسكات RPA2 في هذه الخلايا. بالمقارنة مع الطرق التقليدية ، فإن الكشف المستند إلى RPA2 ل ssDNA حساس للغاية ، وبالتالي يمكن توسيع تطبيقه ليشمل خلايا الطور G1.

الشكل 1: كفاءة تزامن خلايا RPE1 بعد تجويع المصل. (أ) تظهر البقع المناعية مستويات البروتين المشار إليها في خلايا RPE1 غير المتزامنة و G1 و S المتزامنة. (B) تظهر الصور التمثيلية خلايا RPE1 متزامنة غير متزامنة ، G1 ، و S المرحلة التي تعرضت ل 10 μM EdU لمدة 30 دقيقة قبل التثبيت وتصورها بواسطة تفاعل Click-IT. تم استخدام DAPI لمقاومة الحمض النووي النووي. أشرطة المقياس = 50 ميكرومتر. (C) يوضح الرسم البياني النسبة المئوية للخلايا الموجبة ل EdU على إجمالي عدد الخلايا التي تم تقييمها بواسطة DAPI. يمثل شريط الخطأ الخطأ القياسي للمتوسط ، وكانت الأرقام التي تم تحليلها للنوى كما يلي: AS n = 219 ، G1 n = 630 ، S n = 437. الاختصارات: RPE1 = الخلايا الظهارية الصباغية الصبغية الشبكية الخلدة hTERT. AS = غير متزامن ؛ EdU = 5-إيثينيل -2'-ديوكسي يوريدين. DAPI = 4 '، 6-دياميدينو -2-فينيليندول. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
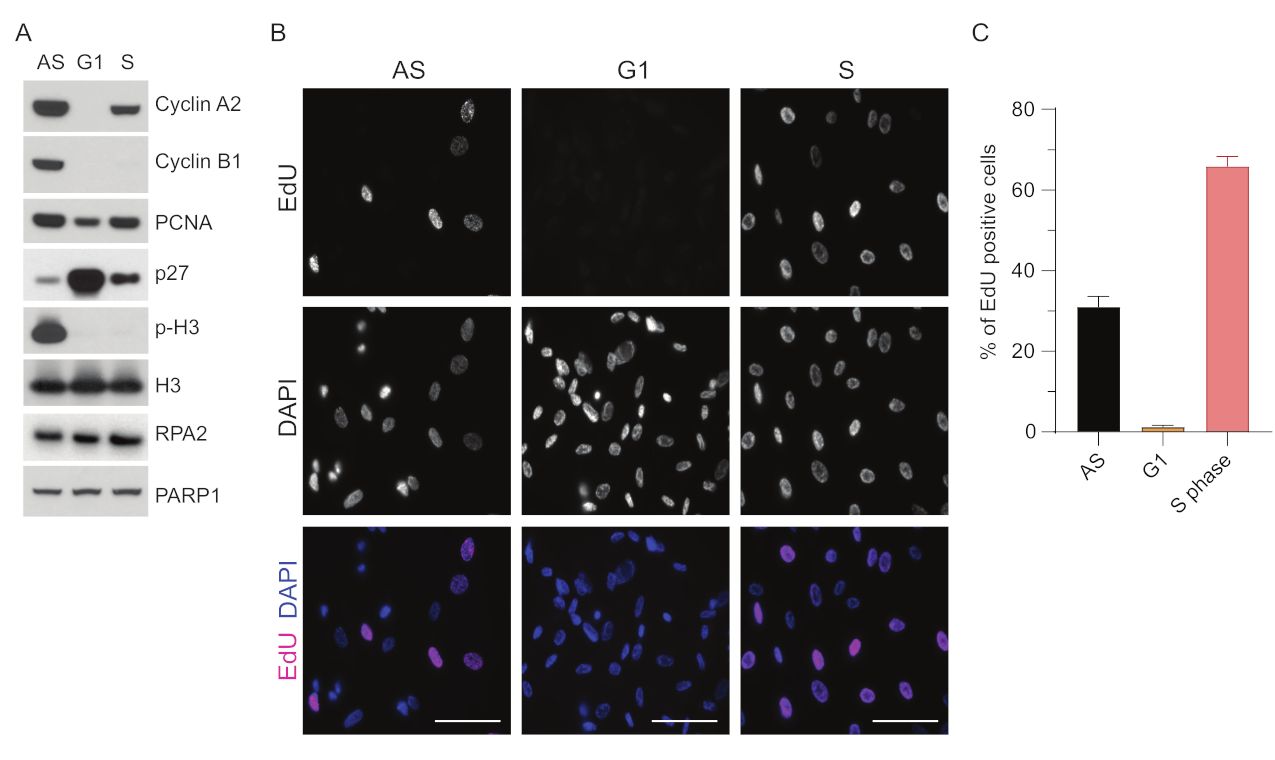{kind=link}

الشكل 2: اكتشاف ssDNA إما باستخدام الجسم المضاد BrdU أو الجسم المضاد RPA2 عند تلف الحمض النووي. (أ) توضح الصور التمثيلية بؤر ssDNA باستخدام αBrdU (الأخضر) ، ويتم تمييز خلايا الطور S بواسطة EdU (أرجواني) ، وتم استخدام DAPI لمقاومة الحمض النووي النووي (الأزرق). تم الاحتفاظ بخلايا RPE1 في 10 ميكرومتر BrdU لمدة 48 ساعة قبل أي علاج إضافي. بعد 48 ساعة ، تم نبض الخلايا ب 10 ميكرومتر EdU لمدة 30 دقيقة تليها معالجة H2O2 (250 μM) لمدة 1 ساعة أو نيوكارزينوستاتين (0.5 ميكروغرام / مل) لمدة 4 ساعات. تم إصلاح الخلايا بعد الاستخراج المسبق ل CSK. يشير الخط الأبيض المتقطع إلى حدود كل نواة. شريط المقياس = 5 ميكرومتر. اللوحات الموجودة على اليمين عبارة عن صور مكبرة لنوى الطور S أو غير S المشار إليها. (B) توضح الصور التمثيلية بؤر ssDNA باستخدام الجسم المضاد αRPA2 (الأخضر). يتم تمييز خلايا الطور S بواسطة EdU (أرجواني) ، وتم استخدام DAPI لمقاومة الحمض النووي النووي (الأزرق). تم نبض خلايا RPE1 ب 10 ميكرومتر EdU لمدة 30 دقيقة متبوعة إما ب 1 ساعةH 2O2 (250 μM) ، أو 4 ساعات من هيدروكسي يوريا (2 mM) ، أو 4 ساعات من NCS (0.5 ميكروغرام / مل). تم إصلاح الخلايا بعد الاستخراج المسبق ل CSK. يشير الخط الأبيض المتقطع إلى حدود كل نواة. شريط المقياس = 10 ميكرومتر. اللوحات الموجودة على اليمين عبارة عن صور مكبرة لنوى الطور S أو غير S المشار إليها. الاختصارات: ssDNA = الحمض النووي أحادي الشريط ؛ BrdU = 5-برومو-2'-ديوكسي يوريدين. DAPI = 4 '، 6-دياميدينو -2-فينيليندول ؛ RPE1 = الخلايا الظهارية الصباغية الشبكية الخالدة hTERT ؛ EdU = 5-إيثينيل -2'-ديوكسي يوريدين. NCS = نيوكارزينوستاتين. HU = هيدروكسي يوريا. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
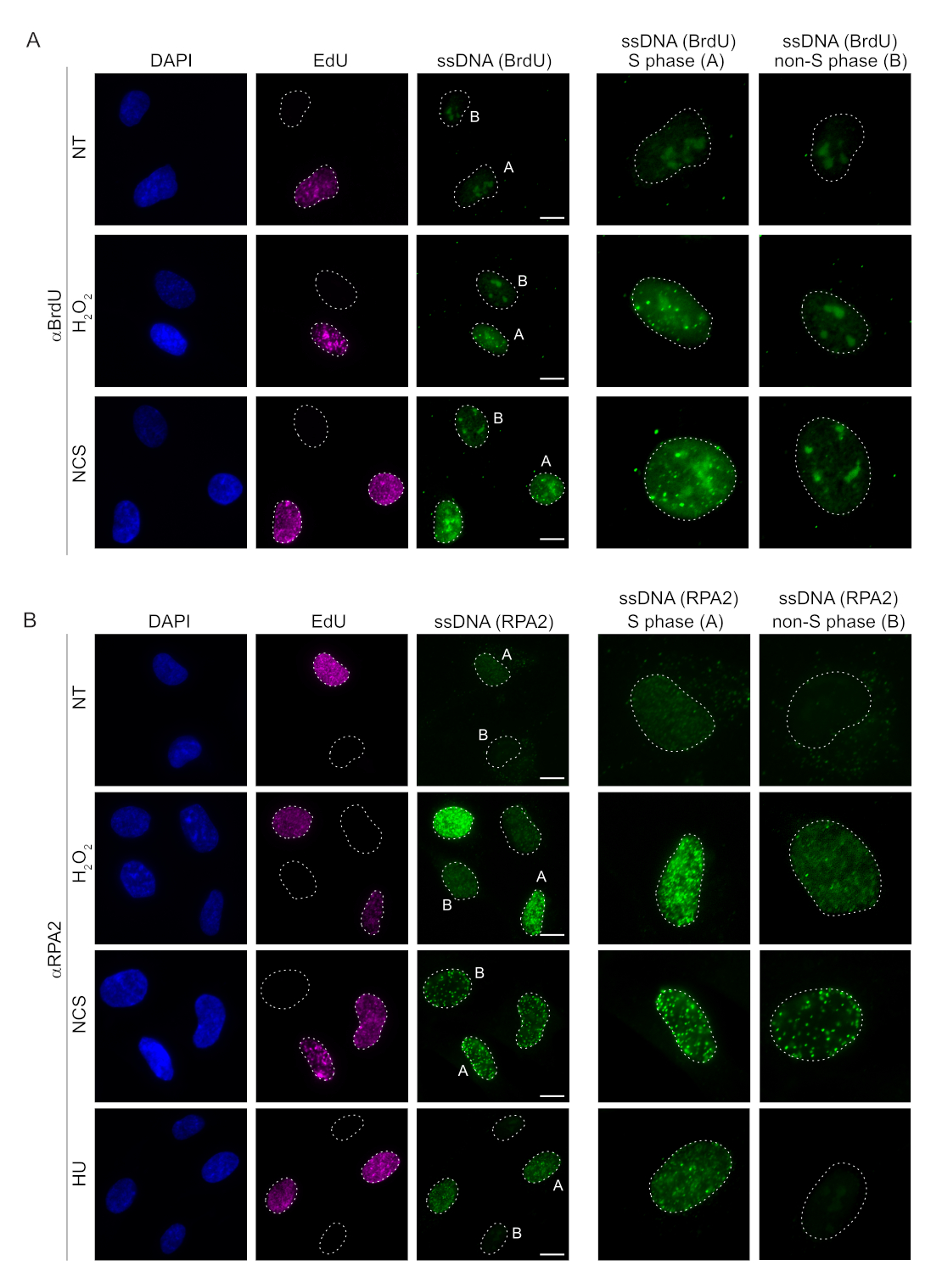{kind=link}

الشكل 3: الكشف عن بؤر ssDNA في المرحلة G1 باستخدام الجسم المضاد RPA2. (أ) تم نقل خلايا RPE1 إما باستخدام siRNAs التي تستهدف RPA2 أو عنصر تحكم siRNA غير مستهدف ، ثم تمت مزامنتها لاحقا في G1 وتم تسميتها بالنبض مع 10 μM EdU لمدة 30 دقيقة قبل معالجتها ب H2O2 (250 μM) لمدة 1 ساعة عند الإشارة إليها. تم استخدام DAPI لمقاومة الحمض النووي النووي. تم إصلاح الخلايا بعد الاستخراج المسبق ل CSK. يشير الخط الأبيض المتقطع إلى حدود كل نواة. شريط المقياس = 5 ميكرومتر. (ب) أجريت قياسات عدد بؤر / نواة RPA2 من تجربتين مستقلتين. تم النظر فقط في الخلايا السلبية EdU أثناء التحليل. تمثل الخطوط القيمة المتوسطة على المؤامرات. تم إجراء اختبار ANOVA غير المعلمية (Kruskal-Wallis) للتحليل الإحصائي. تشير النجوم إلى P < 0.0001. كان عدد النوى الذي تم تحليله كما يلي: siNT no H2O2 n = 513 ، siNT H2O2 n = 603 ، siRPA2 no H2O2 n = 266 ، siRPA2 H2O2 n = 536. (ج) تظهر كفاءة ضربة قاضية للحمض النووي الريبوزي الريبوزي الرسول (siRNA) في النشاف المناعي. الاختصارات: siNT = التحكم في siRNA غير المستهدف ؛ BrdU = 5-برومو-2'-ديوكسي يوريدين. DAPI = 4 '، 6-دياميدينو -2-فينيليندول ؛ RPE1 = الخلايا الظهارية الصباغية الشبكية الخالدة hTERT ؛ EdU = 5-إيثينيل -2'-ديوكسي يوريدين. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
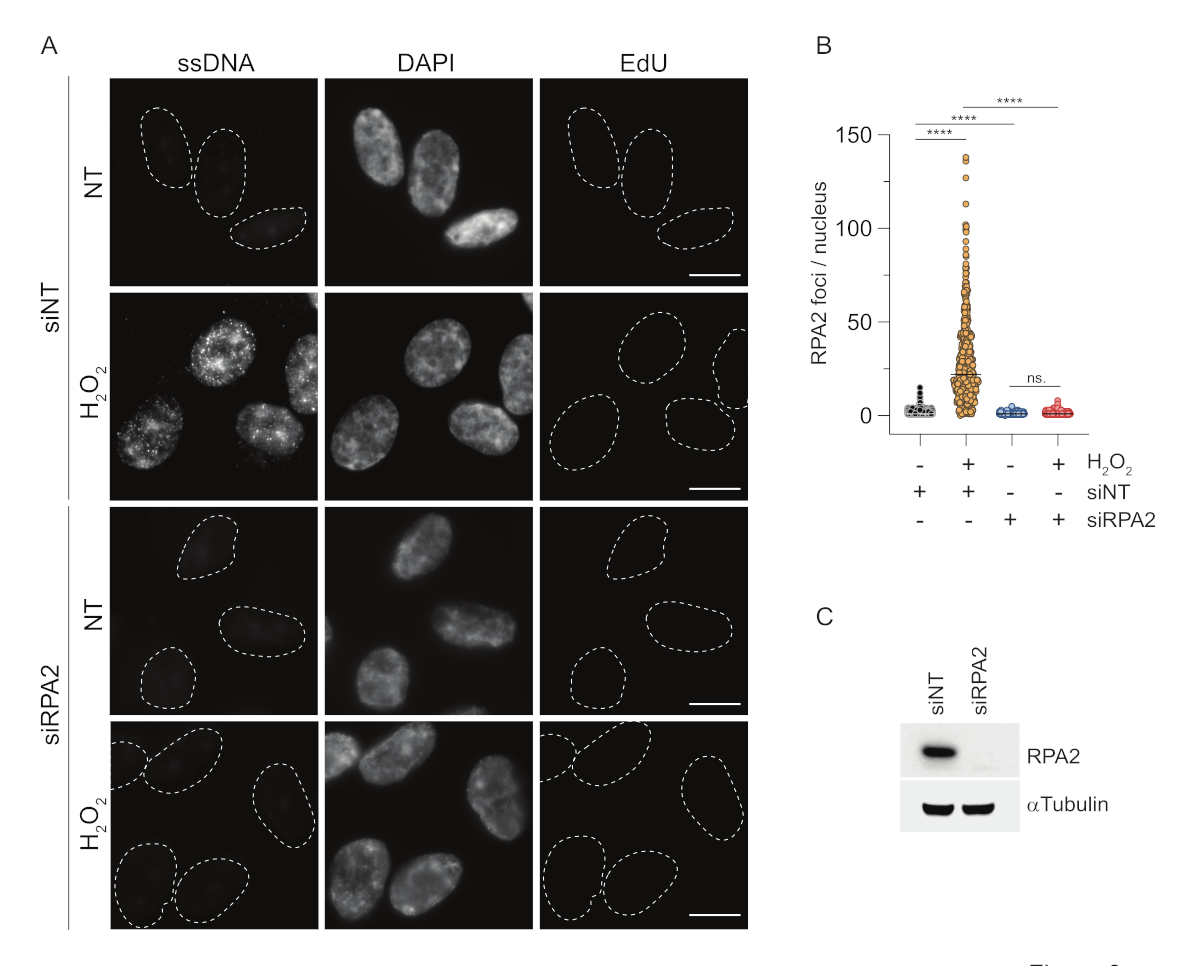{kind=link}

الشكل 4: القياس الكمي لبؤر ssDNA باستخدام فيجي. خطوات مفصلة في فيجي توضح كيفية تقييم أعداد بؤر RPA2 في النواة. (أ-ه) إنشاء قناع نووي باستخدام قناة DAPI. (ف-ح) عتبة لتحديد بؤر ssDNA النووية الفردية من إشارة الخلفية. الاختصارات: ssDNA = الحمض النووي أحادي الشريط ؛ DAPI = 4 '، 6-دياميدينو -2-فينيليندول. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}
| المخزن المؤقت للهيكل الخلوي (CSK) | |
| أنابيب درجة الحموضة 7.0 | 10 مللي متر |
| كلوريد الصوديوم | 100 مللي متر |
| EDTA درجة الحموضة 8 | 1 مللي متر |
| مغ سي سي ال2 | 3 مللي متر |
| د- السكروز | 300 مللي متر |
| تريتون X-100 | 0.20% |
| كوكتيل مثبطات الفوسفاتيز | 1 قرص لكل 10 مل |
| كوكتيل مثبطات الأنزيم البروتيني | 1 قرص لكل 10 مل |
| مخفف في ddH2O | غير متوفر |
| غسل العازلة | |
| تريتون X-100 | 0.05% |
| مخفف في برنامج تلفزيوني | غير متوفر |
| العازلة نفاذية | |
| تريتون X-100 | 0.50% |
| مخفف في برنامج تلفزيوني | غير متوفر |
| حل التثبيت | |
| بارافورمالدهيد | 3.60% |
| تريتون X-100 | 0.05% |
| مخفف في برنامج تلفزيوني | غير متوفر |
| حظر المخزن المؤقت | |
| ألبومين مصل الأبقار (BSA) | 5% |
| تريتون X-100 | 0.10% |
| مخفف في برنامج تلفزيوني | غير متوفر |
| كوكتيل رد فعل Click-iT Plus | |
| 1x المخزن المؤقت لتفاعل Click-iT | 435 مل |
| حل اليكسا فلور PCA | 5 مل |
| CuSO4-النحاس واقية بريمكس | 10 مل |
| 1x انقر فوق iT العازلة المضافة | 50 مل |
| الحجم الكلي | 500 مل |
الجدول 1: تكوين المخازن المؤقتة المستخدمة في هذا البروتوكول.
الشكل التكميلي S1. (أ) تمت مزامنة خلايا RPE1 مع المرحلة G0 باستخدام تجويع المصل لمدة 72 ساعة ثم تم إطلاقها لاحقا في مراحل دورة الخلية المختلفة عن طريق إعادة إدخال المصل. تظهر المخططات النقطية الخلايا في مراحل G0 / G1 أو S أو G2 / M ، حيث تشير الساعات إلى الوقت بعد إعادة إضافة المصل بعد تجويع المصل. يوضح الرسم البياني الموجود على اليمين النسبة المئوية لخلايا G0 / G1 و S و G2 / M في كل حالة. وأجري تحليل نظام مراقبة الأصول الميدانية باستخدام مجموعة أدوات تكاثر الخلايا المتاحة تجاريا باستخدام وحدات المعالجة الإلكترونية ويوديد البروبيديوم وفقا لتوصيات الجهة الصانعة. (ب) مسح اللطخة الغربية غير المقصوصة للشكل 1. تظهر الأرقام علامات الوزن الجزيئي بالكيلو دالتون. تم استخدام PARP1 كعنصر تحكم في التحميل وتم تحميله على الجل الذي تم تطويره أيضا ضد CCNA2 و p27 (تم تجريده أيضا من أجل PCNA) و pH3 (S10) (تم تجريده من H3) عن طريق تقطيع الغشاء. تم تحميل CCNB1 و RPA2 على هلام منفصل ، باستخدام نفس الكمية من البروتين المحللة لضمان المقارنة. (ج) مسح اللطخة الغربية غير المقصوصة للشكل 3. تظهر الأرقام علامات الوزن الجزيئي بالكيلو دالتون. اختصار: EdU = 5-إيثينيل -2'-ديوكسي يوريدين. الرجاء الضغط هنا لتنزيل هذا الملف.
الشكل التكميلي S2: (أ) صور تمثيلية توضح بؤر ssDNA باستخدام الجسم المضاد BrdU (أخضر)؛ يتم تمييز خلايا الطور S بواسطة cyclin A2 (أحمر) ؛ و DAPI تم استخدامه لمقاومة الحمض النووي النووي (الأزرق). تم الاحتفاظ بخلايا RPE1 في 10 ميكرومتر BrdU لمدة 48 ساعة قبل مزيد من العلاج. بعد 48 ساعة ، عولجت الخلايا ب H2O2 (250 μM) لمدة 1 ساعة أو نيوكارزينوستاتين (0.5 ميكروغرام / مل) لمدة 4 ساعات قبل التثبيت. يشير الخط الأبيض المتقطع إلى حدود كل نواة. شريط المقياس = 5 ميكرومتر. (B) تلطيخ BrdU لخلايا RPE1 مع وبدون تغيير طبيعة الحالة. تمت معالجة خلايا RPE1 غير المتزامنة مسبقا باستخدام 10 ميكرومتر BrdU لمدة 48 ساعة. شريط المقياس = 10 ميكرومتر. (ج) أجريت قياسات عدد بؤر / نواة BrdU من تجربتين مستقلتين في خلايا RPE1 المتزامنة G1. تم النظر فقط في الخلايا السلبية EdU أثناء التحليل. تمثل الخطوط القيمة المتوسطة على المؤامرات. تم إجراء اختبار ANOVA غير المعلمية (Kruskal-Wallis) للتحليل الإحصائي. يشير الحرف "ns" إلى اختلاف غير مهم. كان عدد النوى التي تم تحليلها كما يلي: NT n = 52 ، NCS n = 105 ، H2O2 n = 82. الاختصارات: siNT = التحكم في siRNA غير المستهدف ؛ BrdU = 5-برومو-2'-ديوكسي يوريدين. DAPI = 4 '، 6-دياميدينو -2-فينيليندول ؛ RPE1 = الخلايا الظهارية الصباغية الشبكية الخالدة hTERT ؛ NCS = نيوكارزينوستاتين. الرجاء الضغط هنا لتنزيل هذا الملف.
الفيديو التكميلي S1: تسجيل شاشة لتحليل بؤر RPA2 المستندة إلى فيجي. الرجاء الضغط هنا لتنزيل هذا الملف.
Discussion
يعد الحفاظ على ثقافة خلايا صحية خالية من الميكوبلازما أمرا بالغ الأهمية لجميع التجارب الموضحة أعلاه. خلايا RPE1 لها ارتباط قوي بالأدوات البلاستيكية المعالجة بزراعة الأنسجة تحت وسائط الاستزراع العادية. ومع ذلك ، فإن خصائصها الملزمة تتضاءل بشكل كبير عند الاحتفاظ بها في ظروف خالية من المصل. بالإضافة إلى ذلك ، لالتقاط صور عالية الدقة لبؤر ssDNA تحت المجهر ، يجب طلاء الخلايا على زجاج غطاء بسمك 0.17 مم ، وهو غير محب للماء بما يكفي لدعم الارتباط المناسب لخلايا RPE1. بدون خلايا مسطحة بشكل صحيح وموزعة بالتساوي ، من الصعب للغاية تصور بؤر ssDNA الفردية. لذلك ، من الأهمية بمكان اختيار مادة الطلاء المناسبة (على سبيل المثال ، فيترونيكتين) وترك وقت كاف (6-12 ساعة) للخلايا لتنتشر وتلتصق بعد إطلاقها في مرحلة G1.
يتمثل جزء صعب من البروتوكول في الحصول على خلايا RPE1 متزامنة متجانسة G1. وهذا يتطلب خطوتين حاسمتين. أولا ، من أجل تجويع المصل الفعال ، يجب أن تكون الخلايا تريبسين ، وتغسل جيدا باستخدام برنامج تلفزيوني ، وتزرع مباشرة على أطباق زراعة الأنسجة الجديدة باستخدام وسائط خالية من المصل. غسل الخلايا مباشرة في أطباق زراعة الأنسجة لإزالة المصل لن يؤدي إلى مزامنة G0 فعالة. ثانيا ، عند إطلاق الخلايا في مرحلة G1 ، يجب أن تكون الخلايا التربسين مرة أخرى وبذرها على لوحات زراعة الأنسجة الطازجة. وبالمثل ، فإن مجرد تغيير الوسط وإضافة وسيط استزراع يحتوي على المصل إلى الخلايا لن يؤدي إلى إدخال G1 متزامن. بالإضافة إلى ذلك ، من أجل الدخول الصحيح ل G1 ، يجب أن تكون كثافة البذر للخلايا الموجودة على زجاج الغطاء المطلي عند مستويات التقاء معينة. في حين أن مزامنة الخلايا المثالية غير قابلة للتحقيق بشكل عام ، فإن بروتوكول المزامنة هذا الموصوف هنا يعطي ~ 97٪ من السكان G1 النقي. كثافة البذر الموصى بها ل RPE1 على غطاء قطره 12 مم هي ~ 4 × 104 للحصول على مجال رؤية متجانس للتصوير ، مع التقاء 70٪ تقريبا. تتسبب كثافة البذر العالية في انفصال الخلايا و "تقشيرها" بعد استخراج CSK وستؤدي إلى إشارة خلفية أعلى أثناء التقاط الصورة.
لتقليل أي إشارة خلفية وتحقيق نسبة إشارة إلى ضوضاء مواتية ، من الضروري إجراء غسيل شامل بعد حضانة الأجسام المضادة الأولية والثانوية. نظرا لأنه يجب تطبيق العديد من خطوات الغسيل ، فمن الضروري أيضا منع جفاف البئر أثناء كل خطوة غسيل. نقوم بتقليل هذه القطعة الأثرية من خلال تطبيق ما لا يقل عن 0.05٪ Triton X-100 في جميع خطوات الغسيل والحضانة. بمجرد جفاف الآبار ، أظهرت الخلايا نسبة إشارة إلى ضوضاء متغيرة. هذا يؤدي إلى نمط يشبه الفسيفساء تحت المجهر ويمكن أن يتداخل مع التقييم. يمكن أن يساعد الحصول على صورة Z-stack جنبا إلى جنب مع إزالة الالتفاف في التقاط البؤر في مستويات بؤرية مختلفة لتحسين التحليل.
تعتمد الطرق التقليدية على اكتشاف BrdU المدمج في ظل ظروف غير مسخة. ومع ذلك ، تعتمد هذه الطرق على المعالجة المسبقة للخلايا بجرعات عالية من BrdU لمدة 1-2 أيام على الأقل (أو وقت يعادل دورة خلية كاملة في خط الخلية المستخدم) لضمان دمج جينومي موحد. بشكل غير مرغوب فيه ، يمكن أن يتسبب دمج BrdU الشامل في تداخل دورة الخلية36. لمعالجة هذه القيود ، تستخدم هذه الطريقة RPA2 الداخلي للكشف عن بؤر ssDNA. لا يتطلب هذا النهج دمج BrdU المدفوع بالتكرار ، بل يمكن استخدامه أيضا في خلايا ما بعد الانقسام. نظرا لعدم الحاجة إلى دمج BrdU على نطاق واسع ، فإن هذا يوفر الوقت ويقلل من التعقيد التجريبي. باستخدام تلطيخ RPA2 لتصور ssDNA ، يمكننا استخدام 2′-deoxy-5-ethynyluridine (EdU) وكيمياء النقر لتمييز تكرار الحمض النووي مع تجنب التفاعل المتبادل المحتمل للأجسام المضادة BrdU ضد EdU27،37،38. يجب توخي الحذر بشكل خاص لإخفاء EdU المدمج بشكل صحيح أثناء تفاعل النقر حتى لا تتفاعل الأجسام المضادة BrdU مع EdU27,39.
أخيرا ، تتمثل إحدى الفوائد المهمة لاستخدام RPA2 بدلا من BrdU في وجود نسبة إشارة إلى ضوضاء فائقة عند مقارنتها بتلطيخ BrdU خارج مرحلة S. وجدنا أن تلطيخ BrdU غير المتنوع وقدرته على تصور ssDNA يقتصر على المرحلة S حتى في الخلايا المكررة (الشكل 2). يرتبط الجسم المضاد BrdU فقط ب BrdU المكشوف بدرجة كافية في امتدادات ssDNA. قد يؤدي ارتباط بروتينات الإصلاح ، بما في ذلك RPA2 ، بامتدادات ssDNA إلى قمع أو إعاقة التعرض الكافي ل BrdU في ssDNA. وجدنا أيضا أن الاستخراج المسبق ل CSK ضروري لتصور ssDNA باستخدام الجسم المضاد BrdU. هذا ممكن لأن مسارات ssDNA لا يمكن الوصول إليها للجسم المضاد دون إزالة مكونات البروتين المرتبطة قليلا منها.
ومع ذلك ، هناك بعض القيود المرتبطة بهذا البروتوكول. يتمثل أحد قيود استخدام RPA2 للكشف عن ssDNA في الحاجة إلى تحسين خطوة ما قبل الاستخراج CSK. يجب غسل RPA2 الزائد غير المنضم بعيدا عن الحمض النووي قبل تثبيت الخلايا. من ناحية ، يؤدي الاستخراج الناقص إلى خلفية عالية بسبب جزء بروتين RPA2 غير المرتبط ب ssDNA. من ناحية أخرى ، سيؤدي الإفراط في الاستخراج إلى فقدان الإشارة. بالنسبة للكشف عن BrdU ، هذا ليس متغيرا لأن BrdU مدمج بثبات في الحمض النووي ولا يمكن غسله عن طريق الاستخراج المسبق. لذلك ، يجب مراعاة وقت الاستخراج المسبق ل CSK ، وكمية Triton X-100 في المخزن المؤقت ، والحجم ، ودرجة الحرارة التي يتم عندها إجراء الاستخراج المسبق. يحد الاستخراج المسبق ل CSK أيضا من استخدام حجم النواة لتمييز خلايا G0 / G1 عن خلايا S / G2.
بالإضافة إلى ذلك ، لا يمكننا استبعاد احتمال أن تكون بعض الإشارات التي تأتي من RPA2 ناتجة عن ارتباطها بتفاعلات البروتين الأخرى المرتبطة بالكروماتين. يجب على المرء أيضا مراعاة خصوصية الأنواع للجسم المضاد RPA2. يمكن للجسم المضاد المستخدم في هذا البروتوكول التعرف على الإنسان والفأر والجرذ والهامستر RPA2. هناك قيد آخر لهذا النهج وهو أنه لا يمكن أن تكون جميع خطوط الخلايا متعطشة للمصل لمزامنة G0. يمكن لمعظم خطوط الخلايا السرطانية تجاوز نقاط تفتيش دورة الخلية والتكاثر حتى في الوسائط المحرومة من المصل. على الرغم من أن تجويع المصل مفيد ، لأنه لا يسبب تلف الحمض النووي ، يجب على المرء أن يراقب بعناية كفاءة مزامنة الخلايا للتأكد من تحقيق إثراء مرحلة دورة الخلية المناسبة. بالنسبة للخلايا التي لا تستجيب للحرمان من المصل ، يجب مراعاة طرق مزامنة الخلايا الأخرى (على سبيل المثال ، التخلص من الانقسام ، أو تثبيط CDK1 لإيقاف G2 ، أو التقنيات غير الغازية مثل التطهير بالطرد المركزي). طريقة أخرى ممكنة هي استخدام التصوير عالي المحتوى لقياس محتوى EdU والحمض النووي النووي لتوصيف دورة الخلية للخلايا غير المتزامنة31. يجب على المرء أن ينظر في الآثار المترتبة على استخدام طرق التزامن البديلة لمنع التداخل مع التحليل النهائي. على سبيل المثال ، سيؤدي استخدام كتلة الثيميدين المزدوجة أو أفيديكولين ، التي تستخدم غالبا في الأدبيات ، إلى إجهاد النسخ المتماثل وتلف الحمض النووي40.
لا يزال التحقيق في آليات إصلاح الحمض النووي نقطة محورية للمناقشة في مجالات السرطان وبيولوجيا الخلية. يقدم البروتوكول المقدم هنا نهجا قيما لإعداد الخلايا ، مما يتيح التصور والتحليل الكمي ل ssDNA عند التعرض للعوامل الضارة بالحمض النووي. والجدير بالذكر أن هذا البروتوكول يسلط الضوء على استخدام بروتين ربط ssDNA ، RPA2 ، مما يدل على خصوصيته العالية لتصور كميات صغيرة من بؤر ssDNA مع تجنب التفاعل المتبادل غير المرغوب فيه في جميع مراحل دورة الخلية. يمنح استخدام RPA2 العديد من المزايا ، خاصة للباحثين الذين يهدفون إلى تحليل الخلايا في المرحلة G1 من دورة الخلية. يأخذ هذا البروتوكول في الاعتبار العديد من القيود ويعالج المخاوف المتعلقة بتداخل الإشارة وضوضاء الخلفية غير المرغوب فيها والتفاعل المتبادل عند استخدام تلطيخ RPA2 أو BrdU لاكتشاف ssDNA.
Disclosures
ليس لدى المؤلفين مصالح متنافسة يعلنونها.
Acknowledgements
يشكر المؤلفون ميشيل باغانو على دعمه وأفكاره المفيدة ، وآشلي تشوي وشارون كايساري على قراءة المخطوطة بشكل نقدي ، وجيفري إسترادا وفيلما دياز على دعمهم المستمر. تم دعم هذا العمل من خلال ملحق التنوع لمنحة المعاهد الوطنية للصحة GM136250.
Materials
| Name | Company | Catalog Number | Comments |
| Alpha-tubulin antibody | Sigma-Aldrich | T6074 | primary antibody (1:5,000) |
| Axio Observer Inverted Microscope | Zeiss | na | microscope |
| Bis-Tris Plus Mini Protein Gels, 4-12% | Invitrogen | NW04127BOX | Western Blot |
| Bovine Serum Albumin | Jackson ImmunoResearch | 001-000-162 | blocking |
| BrdU (5-Bromo-2'-deoxyuridine) | Sigma-Aldrich | B5002-100MG | nucleotide analogue |
| BrdU antibody BU1/75 | Abcam | ab6326 | primary antibody (1:500) |
| CellAdhere Dilution Buffer | Stemcell Technologies | 07183 | coating reagent |
| Click-iT Plus EdU Flow Cytometry Assay Kits | Invitrogen | C10632 | flow cytomery |
| Click-iT Plus EdU Cell Proliferation Kit for Imaging, Alexa Fluor 647 dye | Thermo Fisher Scientific | C10640 | click-reaction kit |
| cOmplete ULTRA Protease inhibitor tablets | Sigma-Aldrich | 5892791001 | reagent |
| Countess 3 Automated cell counter | Thermo Scientific | AMQAX2000 | cell counter |
| Coverslip | neuVitro | GG12PRE | tissue culture |
| Cyclin A2 antibody | Santa Cruz Biotechnology | sc-271682 | primary antibody (1:1,000) for IF and WB |
| Cyclin B1 antibody | Santa Cruz Biotechnology | sc-245 | primary antibody (1:5,000) |
| Dimethyl sulfoxide (DMSO) | Sigma-Aldrich | D2650-100ML | vehicle control |
| DMEM, high glucose, with HEPES | Gibco | 12430051 | cell culture medium for RPE cells |
| DPBS, no calcium, no magnesium | Gibco | 14190144 | the PBS used throughout the protocol |
| D-Sucrose | Thermo Fisher Scientific | bp220-1 | reagent |
| Eclipse Ti2 Series Epifluorescent Microscope | Nikon | na | microscope |
| EdU (5-Ethynyl-2'-deoxyuridine) | Thermo Fisher Scientific | C10637 | nucleotide analogue |
| Falcon 24-well plate | Corning | 351147 | tissue culture |
| Falcon Cell Culture Dishes 100 mm | Corning | 353003 | tissue culture |
| Fetal Bovine Serum, heat inactivated | Gibco | 16140071 | media supplement |
| Fiji (ImageJ) | NIH | version 1.54f | software and algorithms |
| FxCycle PI/RNase Staining Solution | Invitrogen | F10797 | PI staining |
| Goat anti-mouse IgG (H+L) Highly Cross-Adsorbed Secondary Antibody, Alexa Fluor Plus 555 | Thermo Fisher Scientific | A21422 | secondary antibody (1:1,000) |
| Goat anti-rat IgG (H+L) Highly Cross-Adsorbed Secondary Antibody, Alexa Fluor Plus 488 | Thermo Fisher Scientific | A48262 | secondary antibody (1:1,000) |
| Histone H3 antibody | Abcam | ab1791 | primary antibody (1:10,000) |
| hTERT RPE1 | ATCC | CRL-3216 | cell line |
| Hydrochloric acid | Sigma-Aldrich | H1758-100ML | reagent |
| Hydrogen peroxide 30% soultion | Sigma-Aldrich | H1009-100ML | reagent |
| Hydroxyurea,98% powder | Sigma-Aldrich | H8627-5G | reagent |
| Invitrogen Ultra Pure 0.5 M EDTA pH 8.0 | Thermo Fisher Scientific | 15-575-020 | reagent |
| Lipfectamine RNAiMAX Transfection Reagent | Invitrogen | 13778150 | transfection reagent |
| Magnesium chloride solution 1 M | Sigma-Aldrich | M1028-100ML | reagent |
| MycoFluor | Thermo Fisher | M7006 | Mycoplasma Detection Kit |
| Neocarzinostatin from Streptomyces carzinostaticus | Sigma-Aldrich | N9162-100UG | reagent |
| NuPage MES SDS Running Buffer (20x) | Invitrogen | NP0002 | Western Blot |
| onTARGETplus Human RPA2 siRNA | Dharmacon | L-017058-01-0005 | siRNA |
| p27 antibody | BD Biosciences | 610241 | primary antibody (1:1,000) |
| Paraformaldehyde aqueous solution (32%) | Electron Microscopy Sciences | 50-980-494 | fixative |
| PARP1 antibody | Cell Signaling Technology | 9542S | primary antibody (1:1,000) |
| PCNA antibody | Cell Signaling Technology | 13110S | primary antibody (1:2,000) |
| Penicillin-Streptomycin | Gibco | 15140163 | media supplement |
| pH3 antibody | Cell Signaling Technology | 3377S | primary antibody (1:2,000) |
| PhosSTOP phosphatase inhibitor tablets | Sigma-Aldrich | 4906837001 | reagent |
| PIPES Buffer 0.5 M solution, pH 7.0 | Bioworld | 41620034-1 | reagent |
| Precision Plus Protein Kaleidoscope Prestained Protein Standards | Bio-Rad | 1610395 | Western Blot |
| Prism | GraphPad | version 10 | statistical analysis and graph |
| ProLong Diamond Antifade Mountant | Thermo Scientific | P36961 | mounting media |
| Reduced serum media (Opti-MEM) | Gibco | 31985070 | used for transfection |
| Rpa32/rpa2 antibody (mouse) | EMD Millipore | NA19L | primary antibody (1:1,000) for WB |
| Rpa32/rpa2 antibody (rat) | Cell Signaling Technology | 2208S | primary antibody (1:1,000) for IF |
| Sodium Chloride solution (5 M) | Sigma-Aldrich | S5150 | reagent |
| Sodium Pyruvate (100 mM) | Gibco | 11360070 | media supplement |
| Sodium tetraborate decahydrate | Sigma-Aldrich | B3535-500G | reagent |
| Thermo Scientific Pierce DAPI Nuclear Counterstain | Thermo Scientific | 62248 | nucleic acid stain |
| Thymidine,powder | Sigma-Aldrich | T1985-1G | reagent |
| Triton X-100 aqueous solution (10%) | Sigma-Aldrich | 11332481001 | detergent |
| Trypsin-EDTA (0.5%), no phenol red | Gibco | 1540054 | cell dissociation agent |
| Vitronectin XF | Stemcell Technologies | 07180 | coating reagent |
| ZE5 Cell Analyzer | Bio-Rad | na | flow cytomery |
References
- Hakem, R. DNA-damage repair; the good, the bad, and ugly. EMBO J. 27 (4), 589-605 (2008).
- Gutierrez, R., O'Connor, T. R. DNA direct reversal repair and alkylating agent drug resistance. Cancer Drug Resist. 4 (2), 414-423 (2021).
- Krokan, H. E., Bjoras, M. Base excision repair. Cold Spring Harb Perspect Biol. 5 (4), a012583 (2013).
- Marteijn, J. A., Lans, H., Vermeulen, W., Hoeijmakers, J. H. Understanding nucleotide excision repair and its roles in cancer and ageing. Nat Rev Mol Cell Biol. 15 (7), 465-481 (2014).
- Li, G. M. Mechanisms and functions of DNA mismatch repair. Cell Res. 18 (1), 85-98 (2008).
- Hustedt, N., Durocher, D. The control of DNA repair by the cell cycle. Nat Cell Biol. 19 (1), 1-9 (2016).
- Yang, W., Gao, Y. Translesion and repair DNA polymerases: diverse structure and mechanism. Annu Rev Biochem. 87, 239-261 (2018).
- Bhat, D. S., et al. Therapeutic disruption of RAD52-ssDNA complexation via novel drug-like inhibitors. NAR Cancer. 5 (2), zcad018 (2023).
- Gupta, P., Saha, B., Chattopadhyay, S., Patro, B. S. Pharmacological targeting of differential DNA repair, radio-sensitizes WRN-deficient cancer cells in vitro and in vivo. Biochem Pharmacol. 186, 114450 (2021).
- Pena-Diaz, J., et al. Noncanonical mismatch repair as a source of genomic instability in human cells. Mol Cell. 47 (5), 669-680 (2012).
- Schroering, A. G., Edelbrock, M. A., Richards, T. J., Williams, K. J. The cell cycle and DNA mismatch repair. Exp Cell Res. 313 (2), 292-304 (2007).
- Scully, R., Panday, A., Elango, R., Willis, N. A. DNA double-strand break repair-pathway choice in somatic mammalian cells. Nat Rev Mol Cell Biol. 20 (11), 698-714 (2019).
- Escribano-Diaz, C., et al. A cell cycle-dependent regulatory circuit composed of 53BP1-RIF1 and BRCA1-CtIP controls DNA repair pathway choice. Mol Cell. 49 (5), 872-883 (2013).
- Genschel, J., Modrich, P. Mechanism of 5'-directed excision in human mismatch repair. Mol Cell. 12 (5), 1077-1086 (2003).
- Hu, J., et al. Nucleotide excision repair in human cells: fate of the excised oligonucleotide carrying DNA damage in vivo. J Biol Chem. 288 (29), 20918-20926 (2013).
- Huertas, P., Jackson, S. P. Human CtIP mediates cell cycle control of DNA end resection and double strand break repair. J Biol Chem. 284 (14), 9558-9565 (2009).
- Keijzers, G., et al. Human exonuclease 1 (EXO1) regulatory functions in DNA replication with putative roles in cancer. Int J Mol Sci. 20 (1), 74 (2018).
- Symington, L. S. End resection at double-strand breaks: mechanism and regulation. Cold Spring Harb Perspect Biol. 6 (8), a016436 (2014).
- Liu, Y., et al. DNA polymerase beta and flap endonuclease 1 enzymatic specificities sustain DNA synthesis for long patch base excision repair. J Biol Chem. 280 (5), 3665-3674 (2005).
- Wold, M. S., Kelly, T. Purification and characterization of replication protein A, a cellular protein required for in vitro replication of simian virus 40 DNA. Proc Natl Acad Sci U S A. 85 (8), 2523-2527 (1988).
- Wienholz, F., Vermeulen, W., Marteijn, J. A. Amplification of unscheduled DNA synthesis signal enables fluorescence-based single cell quantification of transcription-coupled nucleotide excision repair. Nucleic Acids Res. 45 (9), e68 (2017).
- Wold, M. S. Replication protein A: a heterotrimeric, single-stranded DNA-binding protein required for eukaryotic DNA metabolism. Annu Rev Biochem. 66, 61-92 (1997).
- Chen, R., Wold, M. S. Replication protein A: single-stranded DNA's first responder: dynamic DNA-interactions allow replication protein A to direct single-strand DNA intermediates into different pathways for synthesis or repair. Bioessays. 36 (12), 1156-1161 (2014).
- Kang, Y., et al. Alteration of replication protein A binding mode on single-stranded DNA by NSMF potentiates RPA phosphorylation by ATR kinase. Nucleic Acids Res. 51 (15), 7936-7950 (2023).
- Kilgas, S., Kiltie, A. E., Ramadan, K. Immunofluorescence microscopy-based detection of ssDNA foci by BrdU in mammalian cells. STAR Protoc. 2 (4), 100978 (2021).
- Madabhushi, R., Pan, L., Tsai, L. H. DNA damage and its links to neurodegeneration. Neuron. 83 (2), 266-282 (2014).
- Liboska, R., Ligasova, A., Strunin, D., Rosenberg, I., Koberna, K. Most anti-BrdU antibodies react with 2'-deoxy-5-ethynyluridine -- the method for the effective suppression of this cross-reactivity. PLoS One. 7 (12), e51679 (2012).
- Biehs, R., et al. DNA double-strand break resection occurs during non-homologous end joining in G1 but is distinct from resection during homologous recombination. Mol Cell. 65 (4), 671-684.e5 (2017).
- Cruz-Garcia, A., Lopez-Saavedra, A., Huertas, P. BRCA1 accelerates CtIP-mediated DNA-end resection. Cell Rep. 9 (2), 451-459 (2014).
- Ercilla, A., et al. Physiological tolerance to ssDNA enables strand uncoupling during DNA replication. Cell Rep. 30 (7), 2416-2429.e7 (2020).
- Lezaja, A., et al. RPA shields inherited DNA lesions for post-mitotic DNA synthesis. Nat Commun. 12 (1), 3827 (2021).
- Mukherjee, B., Tomimatsu, N., Burma, S. Immunofluorescence-based methods to monitor DNA end resection. Methods Mol Biol. 1292, 67-75 (2015).
- Ochs, F., et al. 53BP1 fosters fidelity of homology-directed DNA repair. Nat Struct Mol Biol. 23 (8), 714-721 (2016).
- Raderschall, E., Golub, E. I., Haaf, T. Nuclear foci of mammalian recombination proteins are located at single-stranded DNA regions formed after DNA damage. Proc Natl Acad Sci U S A. 96 (5), 1921-1926 (1999).
- Forment, J. V., Walker, R. V., Jackson, S. P. A high-throughput, flow cytometry-based method to quantify DNA-end resection in mammalian cells. Cytometry A. 81 (10), 922-928 (2012).
- Mistrik, M., et al. Cells and stripes: A novel quantitative photo-manipulation technique. Sci Rep. 6, 19567 (2016).
- Aten, J. A., Bakker, P. J., Stap, J., Boschman, G. A., Veenhof, C. H. DNA double labelling with IdUrd and CldUrd for spatial and temporal analysis of cell proliferation and DNA replication. Histochem J. 24 (5), 251-259 (1992).
- Podgorny, O., Peunova, N., Park, J. H., Enikolopov, G. Triple S-phase labeling of dividing stem cells. Stem Cell Reports. 10 (2), 615-626 (2018).
- Cappella, P., Gasparri, F., Pulici, M., Moll, J. Cell proliferation method: click chemistry based on BrdU coupling for multiplex antibody staining. Curr Protoc Cytom. Chapter 7, (2008).
- Ligasova, A., Koberna, K. Strengths and weaknesses of cell synchronization protocols based on inhibition of DNA synthesis. Int J Mol Sci. 22 (19), 10759 (2021).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved