
Method Article
小鼠基因组工程使用Designer核酸酶
摘要
设计师核酸酶如锌指核酸酶(ZFN)和转录活化因子样效应核酸酶(TALENS)可用于通过触发两个非同源末端连接(NHEJ)和同源重组(HR)途径来修改小鼠早期胚胎的基因组。这些进展使快速生成小鼠的精确的遗传修饰。
摘要
携带位点特异性基因组的修饰(敲除,敲入)转基因小鼠中是极为重要的解剖复杂的生物系统,以及用于模拟人类疾病和测试的治疗策略。在使用设计者核酸酶如锌指核酸酶(ZFN),转录激活因子样效应核酸酶(TALENS),以及集群定期interspaced短回文重复序列(CRISPR)/ CRISPR相关(CAS)系统9对站点的最新进展特定基因工程打开,而不需要依赖于胚胎干(ES)细胞技术在几乎所有的实验室物种进行快速靶向的基因组修饰的可能性。一个基因组编辑实验,通常开始鉴定目的基因其次是建筑定制的DNA结合结构域的直接核酸酶活性的研究者定义的基因组位点内的设计师核酸酶靶位点。设计师核酸质粒在体外</ em>的转录生成的mRNA为受精小鼠卵母细胞显微注射。在这里,我们提供了一个协议,用于通过直接注射TALEN mRNA的实现有针对性的基因组改造成受精小鼠卵母细胞。
引言
小鼠是目前产生转基因动物模型最流行的平台。多功能工具箱的小鼠胚胎1-3的基因工程已经由基于核酸的设计师如锌指核酸酶(ZFN)的基因组编辑方法最近延长4-6,转录激活因子样效应物核酸酶(TALEN)7,8,和群集定期interspaced短回文重复序列(CRISPR)/ CRISPR相关性(CAS)9系统9。 ZFN和TALEN函数作为对两个定制设计的基于蛋白质的DNA结合结构域(锌指蛋白的阵列和重复变量二-残基(倒车视像),分别)分别耦合到所述的FokⅠ核酸内切酶是10-12的。相反,Cas9介导的DNA切割的特异性是通过反式激活CRISPR的RNA(crRNA和tracrRNA,它也可以合并成一个单一的嵌合RNA分子称为导RNA)提供11,其作用在同一个复CRISPR蛋白质。
塔伦斯与定义的倒车视像装置的顺序可以迅速构建了与众多的装配策略,从13-17到选择个别实验者。 CRISPR/Cas9承诺甚至更少的劳动密集型代设计师核酸酶,但导的RNA-DNA的特异性结合仍然没有彻底解决18,19。定制的ZFN的产生迄今被限制在专门的学术实验室和商业供应商如Sangamo生物科学和适马的CompoZr服务。
一般情况下,基因组与设计师核酸编辑的目的是在定义的基因位点,随后吸引非同源末端连接(NHEJ)或同源重组(HR)DNA修复机器10,12引入双链断裂(DSB)。一个DSB的NHEJ介导的维修往往会导致在接近维修部位引入的插入和删除的。因此NHEJ修复ç一个被利用为敲除靶基因通过导入基因的蛋白质编码序列4,7,9中的移码突变的功能。或者,定义添加或更换遗传信息可以通过提供一种DNA供体连同设计者核酸酶来实现。 DNA供体包括由具有同源性靶基因座的两侧区域调查设计的DNA序列,从而作为对DSB修复模板,通过人力资源。两种质粒5,6,20和单链寡核苷酸8,9,21已被成功地用作供体。既不NHEJ-去HR介导的基因组编辑需要引入的选择标记的插入小鼠胚胎,这使得这些策略特别适合于不妨碍总体遗传结构中的核苷酸序列产生小的改变的基因组中。
在这个协议中,我们描述了基因组编辑所有必要程序小鼠胚胎塔兰使用。这些包括:1)识别一个TALEN目标站点22,2)施工TALENS由金门克隆13,3) 在体外合成TALEN mRNA表达,4)微注射TALEN mRNA向受精小鼠卵母细胞中,5)外科手术中用于胚胎移植的和6)在创办人动物TALEN诱导的诱变分析。我们专注于TALEN基因显微注射和奠基人的NHEJ引起的插入/缺失的筛查。为此目的,我们已经生成双官能TALEN结构,允许在哺乳动物细胞中都表达时,如质粒和体外合成TALEN mRNA的显微注射入小鼠胚胎转染。这些结构包括截断TALE骨干23融合到异源二聚体的FokⅠ为最佳的基因组编辑在哺乳动物细胞中域24,25。这个协议也可以用于其他设计师核酸注射或设计师核酸酶结合采用注射和捐助结构(DNA捐赠者的设计已经在良好的技术出版物被描述的Wefers 等 26,27)。
道德声明
所有动物实验均按照苏黎世州的州级兽医办公室的指导方针和规定执行。
研究方案
1。 TALEN目标站点的识别
- 参观TAL效应器核酸Targeter 2.0网站(http://tale-nt.cac.cornell.edu),然后选择“TALEN Targeter”。
- 进入靶基因的序列。如果pCAG-T7-TALEN表达构造,正在使用( 图1C)选择“Miller 等人 ,2011年”,以预测可以与最优间隔长度有针对性的目标站点下的“使用预设架构”(15-20 bp)和TALE倒车视像装置(15-20)为这个TALEN架构的数字。
- 选择“NN”下的“G代以”(鸟苷-NH专用倒车视像装置也可以,但尚未广泛测试TALEN组装)。
- 可选:其他的设置和选项,请按照网站上的指示,其中提供的链接。
- 与潜在目标地点的文本文件生成的,它可以是导入以方便查看电子表格程序。选择TALEN对(s)的金门组装。
- 可选:序列的小鼠品系,将被用于显微注射,以检测可能的单核苷酸多态性(SNP)在公共数据库中可能不入账及利息的基因组区域可能阻止核酸结合。
- 可选:请参阅表3为额外的设计师核酸酶相关的网上资源。
- 需要ZFN和TALEN建设最质粒可以从Addgene以下方式获得:
http://www.addgene.org/special-collections/
在表2中所提供Addgene ID的金门TALEN试剂盒和哺乳动物TALEN表达构建体。
另外,特定的功能模块,如锌指结构域可以从基因合成服务供应商订购。
2。金门TALEN大会
本节描述使用由塞马克等 13全长TALENS使用两个后续金门克隆步骤( 图1)构造一个发表塔兰协议的装配。这种方法允许任意数量的12和31之间的倒车视像装置进入最后的表达结构的结合。装配协议已经适应设计用于产生的mRNA( 图1C),该表达以下的卵母细胞显微注射的高活性TALENS目的载体。请同时参阅金门TALEN套件的在线协议,用于建立和维护质粒文库( http://www.addgene.org/TALeffector/goldengateV2/ )。
- 第1天-金门反应#1 -差饷物业估价署组装阵列的成PFUS载体
- 可选:对于每个TALEN对设计的P中rotocol 1,进入的两个靶位点(包括初始T,5'→3')的DNA序列插入TALEN_Voytas_Pipetting电子表格生成每个金门反应的移液方案(也输入用于组件的所有质粒的浓度)。该电子表格还提供了可用于测序结果的排列在第2.5.2所有倒车视像装置的预期DNA序列。
- 对于每一个独特的TALEN设计方案1:
如果TALEN长度为12-21(标准),选择重复变量二残基(倒车视像装置)1-10和目的载体pFUS_A。选择剩余的倒车视像装置和pFUS_B目的载体, 例如 ,用于一个TALEN 15倒车视像装置(包括最后复读,这将被添加到最后的组装第2.3.3节中,选择倒车视像装置11-14和目的载体pFUS_B4( 图1A)。
如果TALEN长度为22-31,使用1-10倒车视像装置和目标已经构造函数pFUS_A30A。接倒车视像装置11-20和目的载体pFUS_A30B。选择剩余的倒车视像装置和pFUS_Bdestination载体, 例如用于TALEN与24接倒车视像装置倒车视像装置21-23和目的载体pFUS_B3。 - 成立金门反应#1为每个PFUS +差饷物业估价署分别组合, 即 1-10 + pFUS_A和剩余倒车视像装置+各自pFUS_B载体或TALEN超过21倒车视像装置,1-10 + pFUS_A30A,11-20 pFUS_A30B,和其余的倒车视像装置+各pFUS_B载体。用150纳克每RVD矢量,150纳克PFUS载体,1μl的BsaI,1微升T4 DNA连接酶,2微升10×T4 DNA连接酶缓冲液中,与H 2 O至20μl总反应体积。 (使用的每一轮的金门组件T4连接酶缓冲液的新鲜等分建议以来的T4连接酶缓冲液反复解冻/冷冻可以降低反应效率。)
- 将金门反应在热循环仪。
程序:
37℃5分钟
16℃下10分钟
10次
50℃5分钟
80℃5分钟 - 加入1μl10 mM的ATP和1微升质粒核酸安全对每个组合,在37℃下1小时。这种治疗方法可消除可能由在转化的细菌体内同源重组克隆到目的载体线性不完整的连接产物。
- 转化大肠(可以在这里和在随后的转化中使用电感受或化学感受态大肠杆菌有利于α-互补如XL1-蓝或DH5α) 大肠杆菌与个别连接反应。
- 对壮观霉素的细菌平板(50微克/毫升)板用X-gal和IPTG(40微克/毫升每个)为蓝色/白色菌落的选择。
- 第2天-正确PFUS -倒车视像装置组装的确认
- 通过菌落PCR引物pCR8_F1和pCR8_R1( 见表1引物序列)屏幕1-3白从各板块的殖民地。正确PFUS,倒车视像装置组件通常显示对应的克隆所有倒车视像装置( 如周围1.1 kb的10倒车视像装置)的总长度和一个带一个“梯子”小不那么突出条带( 图1B)。
PCR程序:
95℃3分钟
95℃30秒
55℃30秒
72℃1分钟45秒
30-35个循环
72℃10分钟 - 使用证实克隆启动过夜培养(2-5毫升的LB用50微克/毫升大观霉素)。
- 通过菌落PCR引物pCR8_F1和pCR8_R1( 见表1引物序列)屏幕1-3白从各板块的殖民地。正确PFUS,倒车视像装置组件通常显示对应的克隆所有倒车视像装置( 如周围1.1 kb的10倒车视像装置)的总长度和一个带一个“梯子”小不那么突出条带( 图1B)。
- 第3天-金门反应#2 -差饷物业估价署阵列成TALEN表达载体
- 执行“小量制备”来隔离PFUS-RVD组件(取决于倒车视像任pFUS_A和pFUS_B或pFUS_A30A,pFUS_A30B和pFUS_B数)。
- 可选:用引物pCR8_F1,pCR8_R1( 见表1引物序列)序列个别PFUS载体。测序也可于f执行INAL TALEN构造(第2.5.2节);然而,对于更长的TALENS完整读取所有倒车视像的使用Sanger测序可能是不可能的。
- 成立金门反应#2,每单TALEN。 150纳克每PFUS载体,根据差饷物业估价署序列的设计和为l EFT TALEN使用75毫微克150纳克PLR-HD,PLR-NG,PLR-NI,PLR-NN(最后的“半重复”)的pCAG-T7-TALEN-ELD-目的地和合适TALEN用75纳克pCAG-T7-TALEN-KKR公司目标(或反之亦然)。加入1μlEsp3I,1微升T4 DNA连接酶,2微升10×T4 DNA连接酶缓冲液,H 2 O至20μl总反应体积。
- 将金门反应在热循环仪。
程序:
37℃5分钟
16℃下10分钟
10个周期
37℃15分钟
80℃5分钟 - 使用从第2.3.4节反应转化大肠杆菌大肠杆菌 。
- 第4天 - 正确TALEN大会确认
- 从每块板通过菌落PCR用引物TAL_F1和TAL_R2屏幕1-3个白色菌落( 见表1为引物序列)。正确组装TALENS表明PCR产物,其长度对应于结合( 图1D,该频带是有时难以检测,而“阶梯效应”表示成功的组件的健壮的指标)倒车视像的总数。
PCR程序:
95℃3分钟
95℃30秒
55℃30秒
72℃3分钟
30-35个循环
72℃10分钟 - 确认使用正确的克隆,开始过夜细菌培养(2-5毫升的LB 100微克/毫升氨苄青霉素)。
- 从每块板通过菌落PCR用引物TAL_F1和TAL_R2屏幕1-3个白色菌落( 见表1为引物序列)。正确组装TALENS表明PCR产物,其长度对应于结合( 图1D,该频带是有时难以检测,而“阶梯效应”表示成功的组件的健壮的指标)倒车视像的总数。
- 第5天-正确TALEN大会确认
- 执行“小量制备”来隔离pCAG-T7-TALEN质粒。
- 如果没有在本质上进行测序CTION 2.3.2,使用引物TAL_Seq_5-1和TAL_R2( 见表1引物序列),以确定正确的装配差饷物业估价署在全长TALEN。
3。核酸酶mRNA的合成
- DNA模板的生成
- 制备高品质MIDI或pCAG-T7-TALEN质粒mRNA合成maxipreps。
- 使用该优先裂解下游并在靠近核酸酶终止密码子(用于pCAG-T7-TALEN矢量用SacI位 )限制性内切酶线性化10微克的TALEN或ZFN mRNA合成质粒。 mRNA合成的质粒通常包含T7或SP6噬菌体的启动子的核酸编码序列的上游。
- 运行在0.7-1%琼脂糖凝胶200-500纳克消化的质粒来检查完全消化。
- 从质粒除去盐通过用醋酸0.1体积钠和3倍体积的乙醇1小时,在室温温度场中沉淀该DNA消化Ë。通过离心分离沉淀的DNA在14,000×g离心或多个10分钟,用200μl70%乙醇洗涤沉淀,旋另外5分钟,除去乙醇,干燥空气中的无RNA酶的水适当体积的沉淀重悬。将线性化质粒的纯化也可以使用基于列的系统, 例如的QIAquick PCR纯化试剂盒。
- 确定线性模板的浓度,并使用1微克到设置在体外转录。
- mRNA的合成和多聚腺苷酸化
- 用于体外转录的pCAG-T7-TALEN的质粒用T7 mMESSAGEmMACHINE超试剂盒。设置每个TALEN在冰上转录反应:20微升用无核酸酶水,10微升的T7 2×NTP / ARCA,2微升10×T7反应缓冲液,1微克的DNA模板,2微升的T7酶混合物。混合反应孵育1-2小时,在37°C。
- 使用完整的20微升转录化反应混合物中,以设置在冰上的多聚腺苷酸化反应:36微升不含核酸酶的水,20微升5×EPAP缓冲液,10微升的25mM氯化锰,10微升10毫摩尔ATP。混合和(,左,右分别核酸酶)移走2.5微升的反应混合物作为对照样品L1和R1。加入4微升的E-PAP酶孵育45-60分钟的反应在37°C删除另一个2.5微升反应混合物作为对照样品L2和R2。
- mRNA纯化
- 使用NucAway离心柱的mRNA纯化(更换缓冲液和去除未掺入的核苷酸)。点击列定居干凝胶在列的底部。水合物柱用650微升的无RNA酶的显微注射缓冲液(1毫摩尔Tris-CL,0.1 EDTA,pH 7.5)中。帽,涡流,挖掘出气泡,水合物在室温下5-15分钟。
- 将列在集合管和旋在750×g离心,4℃,2分钟以除去过量的间质液。丢弃收集管和地点柱中加入1.5ml洗脱管子。
- 适用于完整的反应混合物从第3.2.2节以750×g离心,4℃,2分钟柱和旋转。核酸酶的mRNA现在将溶解在注射缓冲液中。除去2.5微升纯化样品L3和R3。
- 直到注射等分试样制备店的mRNA在-80°C。
- mRNA的凝胶电泳
- 清洁凝胶室用10%SDS溶液或RNaseZAP去除RNA酶污染。
- 准备在1xTBE运行缓冲液在1%琼脂糖凝胶上。
- 混合各RNA样品L1/R1,L2/R2,L3/R3用3倍体积的NorthernMax甲醛负载染料孵育15分钟,在65℃下
- 负载样品和RNA的大小梯子( 如 RNA千年大小标记)上的凝胶并且在1x TBE凝胶上运行在10伏/厘米,直到加载染料到达凝胶的末端。
- 使用SYBR绿液(Invitrogen)中30〜60分钟在搅拌下于室温染色凝胶。
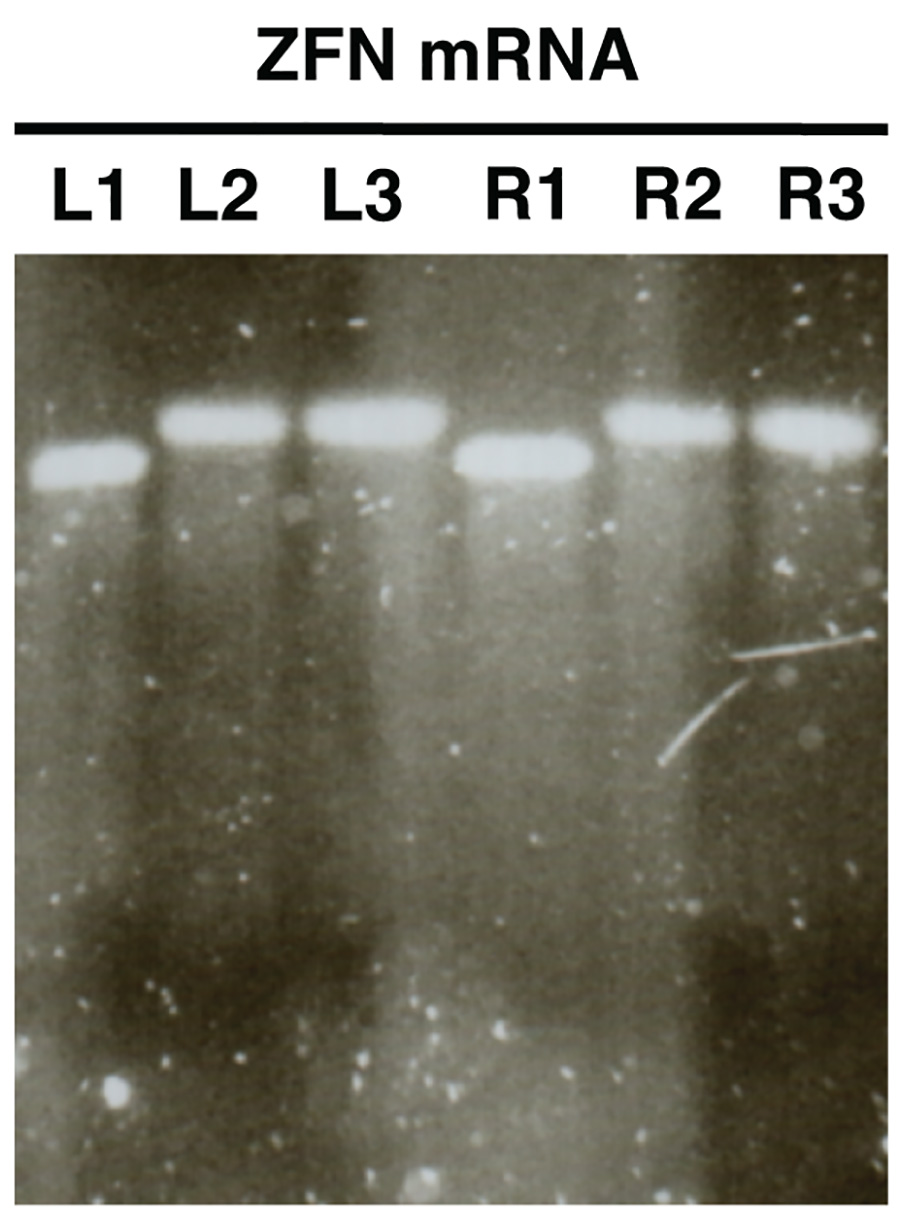
- 图片凝胶和L2/R2和L3/R3比较L1/R1的大小。样品L2/R2和L3/R3应该显示出增加的大小的相对L1/R1频段指示( 图2)成功的多聚腺苷酸化。
- 使用分光光度计测定mRNA的浓度。
- 通过将左核酸酶和核酸酶权利mRNA的以1:1的比例制备的mRNA等份为显微注射。我们建议准备用等分通过显微注射缓冲液稀释的总浓度为200纳克/微升(每个核酸100纳克/微升)。店铺mRNA的等分试样在-80℃下
4。胚胎分离及显微注射
- 胚胎分离 。该协议已经成功地与C57BL/6J和BDF1品系用s和能最有可能适应于显微注射实验,如FVB或CBF1常用的其他菌株。将小鼠在温度和湿度控制设施与食物和水随意容纳下一个12小时,12小时明暗周期。
- Superovulate供体雌性增加胚胎的产率。女16例(4周龄)腹腔(IP)注射5国际单位孕马血清促性腺激素(PMSG),其次为腹腔注射5国际单位绒毛膜促性腺激素(hCG)48小时后超排。
- 交配16女超排向16岁配种(2-8个月)注射hCG后立即男性。
- 使用经批准安乐死协议,如CO 2吸入牺牲女性。
- 恢复受精的卵母细胞。卵母细胞从输卵管采集的第二天早晨,然后用3-5分钟的治疗在0.1%牛透明质酸酶溶解在M2培养基从任何剩余的卵丘细胞释放。取决于配合性能和所使用的菌株的胚胎产率可能有所不同。 16只C57BL/6J雌性通常会产生150-250,而BDF1产量300-400受精的卵母细胞,分别。
- 胚胎显微注射
- 注入核酸酶mRNA的表达。原核阶段的卵母细胞注入通常在午后。细胞质mRNA显微注射是矿物油下进行,在M2培养基使用倒置显微镜配备有利用Nomarski DIC用20X物镜和与胚胎显微操作器,以及一个注射装置。我们建议起TALEN mRNA的注射剂在浓度为10纳克/微升(20纳克/微升总数)。
- 吸出的卵母细胞以保持毛细管和中胚避免与原核接触的细胞质注入核酸酶的mRNA。注射应浅,靠近质膜。注射量应保持在低水平和显微注射针应该是机智hdrawn在细胞质腹胀的第一个迹象。显微注射系列将通常包含100-300卵母细胞。通常情况下,胚胎的至少80-90%应该生存的注入不立即溶解。
- 以下显微注射,将胚胎进行1小时的M16(Sigma)的培养基中,在37℃和5%CO 2和存活的胚胎转移到假孕养母。
5。手术胚胎移植
- 准备假 孕的胚胎收件人(养母) 一天的mRNA显微注射前 。成熟女性(3-6个月),一个强大的远交品系,如CD-1,是非常适合的角色。通过交配的雌性的前一天与任何手术或基因输精管结扎男性28诱发假孕。只有0.5 DPC女性有明显的交配栓作为胚胎收件人。在未使用的女性后约3凌晨的假孕消失KS让他们反复使用。
- 腹腔注射氯胺酮麻醉的0.5 DPC养母女性,甲苯噻嗪(120毫克/公斤和16毫克/公斤)。这一提法担保〜30分钟的手术耐受时间是绰绰有余的5-10分钟的预计运行时间。
- 定位在其腹部麻醉动物在温暖的表面和与合适的消毒剂,如70%乙醇或氯己定和醇混合物消毒切口的面积。
- 切割在动物背部皮肤,用无菌剪刀打开腹腔。
- 可视化和通过拉动连接在卵巢脂肪垫的外部化子宫角。保持湿润的器官用温暖的0.9%NaCl溶液。请记住,手术部位可能要搭着用无菌敷料或剃掉,以符合当地兽医的指导方针。
- 通过裁剪卵巢脂肪垫与斗牛犬钳固定生殖器官。
- 轻轻抓住输卵管与制表师镊子壶腹的上游,并使用30 G注射针来创建一个小孔在输卵管壁。
- 退出针并重新插入含有胚胎成孔允许轻轻吹毛细管持有人的喉舌安置在壶腹部胚胎的薄玻璃毛细管。撤回毛细管一旦胚沉积在壶腹和替换生殖器官背部在体腔中。
- 关闭腹腔一系列无菌缝合线(普理灵6-0)的。
- 使用1-2伤口剪辑(Autoclip9毫米)取决于开口的尺寸接近皮肤。
- 下面的操作中返回的动物,以他们的家笼和监督他们,直到麻醉的效果穿脱。
- 为了尽量减少压力,房子的小鼠稳定的社会群体(III 2-4只/笼型),只要有可能。
- 适用于手术后镇痛在Dafalgan的表格S加入到饮用水(对乙酰氨基酚200毫克/公斤体重)的手术后3天。
6。创办人通过PCR和T7核酸内切酶或限制性内切酶分析
- 设计引物以扩增周围的核酸结合位点200-700碱基之间的区域。的距离,正向引物-核酸间隔区和无核酸酶的间隔区-反向引物应具有足够的不同,以允许检测的消化产物两个单独的条带在琼脂糖凝胶上(参见图3和图4的例子)。
- 运行的PCR优化的条件。检查PCR扩增子大小的琼脂糖凝胶。
- 可选:净化PCR产物, 例如用QIAquick PCR纯化试剂盒从PCR混合物中除去盐和核苷酸。不需要许多限制性内切酶是活性辅以适当的限制性内切酶缓冲液和纯化的PCR样品中。我们也有事业有成在QiagenTaq PCR缓冲液LLY使用T7核酸内切酶辅以NEBuffer 2。
- T7核酸内切酶检测。混合17微升的PCR产物与2μlNEBuffer 2和运行异源双链的形成( 图3b)程序在一个热循环:
95℃2分钟
95°C至85°C(-2°C /秒)
85°C至25°C(-0.1°C /秒)
4℃保持。
加入1μlT7核酸内切酶的每个样品,孵育在37℃下20分钟。 (验船师检验(Transgenomic公司),它依赖于CEL 1核酸酶,也可以在这里使用。请参阅制造商的说明。) - PCR产物的限制性消化。混合17微升的PCR产物与2微升10×限制性酶缓冲液和1μl限制性酶。消化在适当的温度下进行1小时或更长时间。
- T7核酸内切酶检测。混合17微升的PCR产物与2μlNEBuffer 2和运行异源双链的形成( 图3b)程序在一个热循环:
- DNA上样染料添加到样品,并以2%的琼脂糖凝胶上电泳,以检测不同的消化图案为野生型动物和奠基人携带突变的等位基因。参见图3和4为预期的结果。
- 克隆PCR产物的创始人阳性突变的等位基因的Sanger测序( 例如,通过TA克隆到pGEM-T Easyor直接使用新一代测序测序PCR产物的混合物。
结果
我们构建了由塞马克等人 13,允许TALENS在哺乳动物细胞中的表达以及在体外的mRNA合成的T7噬菌体的启动子( 图1C)所发表的金GateTALEN组件兼容的目标质粒。这些质粒携带的异源二聚体的FokⅠ结构域(ELD或KKR突变)已经显示减少的脱靶效应相对的FokⅠ同源二聚体,并提高相对于第一代的FokⅠ的异二聚体25,29裂解活性。金门装配反应#1和#2通常是非常高效的,每一个白色菌落,当通过菌落PCR进行分析,显示了倒车视像克隆( 图1B和1D)的特定数目的预期模式。
在体外合成mRNA的凝胶分析( 图2)要表达的单辨的色带,很少或没有拖影,每样品分析。应该有样品L1/R1和L2/R2,L3/R3,这表明成功的多聚腺苷酸化之间有着明显的尺寸变化。
创办人动物可以进行筛选,使用基因分型的PCR后跟T7核酸内切酶消化( 图3)或限制性消化用酶NHEJ诱发突变的等位基因的核酸酶对的间隔区域内的切割与野生型序列( 图4)。在T7核酸内切酶测定法是适用于任何种类的突变,而不论核酸酶对注射的间隔区域内的特定基因组序列;但是,它在检测到异源双链PCR产物DNA链之间只有不匹配。因此,在创办人有两个相同的等位基因突变的罕见事件,PCR产物不会显示任何T7酶切图谱。这样的区别是,然而,总是可能的,当一个特定的限制性位点被定位在与由NHEJ被淘汰的核酸间隔区域内诱导的插入/缺失( 图5)。这里,未消化的频段表明突变的存在,并没有任何的消化产物强烈暗示创办人携带突变的靶基因的两个等位基因(标有在图4b中的星号)。

图1。差饷物业估价署数组到PFUS载体金门克隆TALEN倒车视像装置的成异源二聚体pCAG-T7-目标向量A)大会。这里,例如,示出为一TALEN对与个别海绵阵列包括15个倒车视像和17倒车视像(分别为海绵阵列长于21倒车视像,3 PFUS - 倒车视像组件是必需的,未示出)。箭头表示引物,PFUS特定菌落PCR反应,B)的PCR产物从正确的PFUS组件扩增通常显示一个波段对应的所有克隆倒车视像装置( 如周围1.1 kb的10倒车视像装置)和一个“阶梯”小不那么突出乐队,由于差饷物业估价署阵列的重复性质。 的C总长度)总装PFUS-RVD阵列和包含最近一次重复(PLR)与FokIELD和FokIKKR变种,分别为异源二聚体TALEN表达载体质粒。该TALEN骨干(注解为N和C)通过类似于Miller 等人 23出版该架构的CAG(CMV早期增强子元件/鸡β-肌动蛋白)的启动子可以确保在转染的哺乳动物细胞中的高表达水平,而在T7噬菌体启动子允许在体外合成mRNA(用SacI位的核酸酶终止密码子下游的载体线性化)D)菌落PCR使用在C箭头所示的引物),可以识别正确组装TALENS的。全lengtħPCR产物往往不那么突出,而“阶梯效应”代表成功装配的强劲指标。 点击这里查看大图 。
{kind=link}

图2。 。核酸酶的mRNA 在使用琼脂糖凝胶电泳体外合成的质量控制的ZFN的mRNA被示为一个例子(L,左ZFN,R,右ZFN)。样品L1/R1显示的mRNA前多聚腺苷酸化,样品L2/R2秀聚腺苷酸化的mRNA和L3/R3显示纯化的多聚腺苷酸化的mRNA。 点击这里查看大图 。
{kind=link}
ALT =“图3”FO:内容宽度=“6英寸”FO:SRC =“/ files/ftp_upload/50930/50930fig3highres.jpg”SRC =“/ files/ftp_upload/50930/50930fig3.jpg”宽度=“600” />
图3。用于识别携带靶基因座的核酸酶诱导的突变创始人动物的T7核酸内切酶测定法的例子。 A)TALEN对被设计在鼠标朊蛋白基因(PRNP的编码区域内切割,TALEN靶序列可根据要求提供)。使用正向引物(F)位于110 bp的上游和反向引物(R)的250 bp处TALEN切割位点。 乙的下游)将PCR产物随后进行异源双链的形成和T7核酸内切酶消化产生的PCR产物。℃)单一创始人的目标基因组区域内TALEN诱导的诱变是通过一个全长的PCR产物与250的消化产物和110基点的存在揭示。50930/50930fig3highres.jpg“目标=”_blank“>点击这里查看大图。

图4。用于识别携带靶基因座的核酸酶诱导的突变。ZFN特异性针对小鼠ROSA26轨迹29靶内的内含子1中的XbaI限制性位点创始人动物的PCR产物的限制性酶切消化的实施例A)的创始人通过使用基因分型的PCR筛选正向引物(F)位于500bp的上游和反向引物(R)的250 bp的切割位点。 乙的下游)扩增产物用XbaI消化揭示酶切图谱表明小鼠双等位基因突变(标有星号),单等位基因突变(消化,未消化的乐队, 如动物2)老鼠与潜在双等位基因修改和野生型小鼠(完全消化, 如动物21)。C)测序显示多达3个(动物24)不同的插入/缺失。 点击这里查看大图 。
{kind=link}
| 引物的名称 | 序列5'至3' |
| pCR8_F1 | ttgatgcctggcagttccct |
| pCR8_R1 | cgaaccgaacaggcttatgt |
| TAL_F1 | ttggcgtcggcaaacagtgg |
| TAL_R2 | ggcgacgaggtggtcgttgg |
| TAL_Seq_5-1 | catcgcgcaatgcactgac |
表1中。用于金门TALEN组件中菌落的PCR和测序引物序列协议。
| 质粒/收藏 | 贡献者 | Addgene编号 | 评论 |
| 金门TALEN和TAL效应Kit 2.0 | Voytas实验室 | 1000000024 | 包含金门TALEN组装所需的所有质粒 |
| pCAG-T7-TALEN -KKR/ELD目的向量 | Pelczar实验室 | 40131,40132 | 附加在哺乳动物细胞和体外 mRNA合成质粒TALEN表达 |
表2。需要金门TALEN装配质粒和质粒集合可以从Addgene(获得 www.addgene.org )。
| 在线资源 | 评论 |
| http://tale-nt.cac.cornell.edu | TALEN设计; TALEN脱靶预测 |
| http://zifit.partners.org/ZiFiT/ | TALEN,开放的ZFN科达ZFN,CRISPR/Cas9设计 |
| http://www.genome-engineering.org | TALEN,CRISPR/Cas9设计; CRISPR/Cas9脱靶预测 |
| http://baolab.bme.gatech.edu/Research/BioinformaticTools/assembleTALSequences.html | TALEN的装配序列确认测序结果 |
| http://pgfe.umassmed.edu/ZFPmo dularsearchV2.html | 模块化装配ZFN设计 |
| www.genomecenter.ucdavis.edu/s egallab / segallabsoftware | 模块化装配ZFN设计 |
表3。网上资源设计的ZFN,TALEN和CRISPR/Cas9。
讨论
设计师核酸酶驱动的基因组编辑方法已显著延长物种适合于它们各自的基因组10,12的靶向修饰的范围内。在小鼠中,基因靶向ES细胞一直是一个标准的技术超过二十年;然而,它已被证明难以适应从品种较其它鼠标胚胎干细胞,虽然出现了一些最近成功在大鼠胚胎干细胞。即使有“现成的,现成的”基因靶向小鼠ES细胞由财团如EUCOMM,KOMP,或NorCOMM 3的基因组编辑由ZFN和TALEN提供更高的精度和有关的修改,可以将频谱灵活性提供克隆的可用性引入到小鼠基因组中。携带核酸酶介导的基因突变动物的创始人似乎是高度的种系主管4-6,20,21,这并不总是对嵌合体从囊胚注射ES细胞的起源的情况。因此,在DES的某些情况下显微注射igner核酸酶可导致显著更快的一代新型鼠标线有针对性的基因组修改。
成功代基因敲除小鼠注射ZFN和TALEN取决于对注入对核酸酶的活性有很大影响。 TALENS已被证明具有高的成功率在目标范围广泛的基因在许多生物体中;然而,最近的研究表明,TALEN结合是胞嘧啶甲基30,31敏感,因此,新产生的核酸对,例如TALENS克隆到pCAG-T7载体,可瞬时转染到鼠细胞系,例如NIH-3T3或神经-2a中,它模仿了小鼠胚胎的染色质状态到一定程度。在这里,核酸酶活性可以用T7核酸内切酶分析或PCR产物的限制性酶切如前mRNA的合成和显微注射在第5节所述来估计。我们建议在尊重兴趣的基因组区域的测序的ive细胞系和用于显微注射实验的小鼠品系。
在小鼠受精卵,不同TALEN或ZFN对将最佳状态工作在不同的mRNA浓度,因此,显微注射核酸酶mRNA的最佳工作浓度可能必须通过实验确定。根据不同的核酸酶对,浓度太低会导致无解理,而过高会导致胚胎致死。取决于核酸对,我们已使用的总mRNA浓度低至2纳克/微升和高达200微克/微升过成功。这些影响是难以从在细胞培养物和核酸浓度最佳为胚胎存活和目标轨迹的变形速率需要根据经验确定的实验来预测。
高活性的ZFN或TALEN可切割其靶序列以外的显微注射胚胎的单细胞阶段,从而导致复杂的诱变的图案ð镶嵌在创始人。我们和其他4已经在一个单一的创始人( 图5C)观察到三个或更多个不同的突变等位基因。因此,建立从这些创办新的鼠标线时,后代应仔细测序的有利突变的存在,因为消化测定提供证据仅一个未定义的突变的存在筛选。
之一的批评经常对ZFN和TALEN系统表示这样的可能性,即这些核酸酶也能够裂解在类似于目标位点的基因组中存在别处序列。这样的脱靶效应已被观察到与初代试剂使用的同型二聚体的FokⅠ域和异源二聚体构建体被设计为减轻脱靶效应25。潜在的脱靶部位可以预测到一定程度,在硅片 32,33和通过PCR和测序进行筛选。一个明显的优势ØF使用的ZFN和TALENS用于产生小鼠而不是细胞系是除去脱靶突变通过执行几个回交至所选择的野生型菌株未链接到所需的基因组修饰的可能性。供了大量的建小鼠,从核酸靶向基因座产生和在计算机芯片预测的脱靶PCR产物的下一代深度测序分析基因座可能提供一个替代的定性和定量的读出到PCR产物的消化测定。
在此协议中描述的辅助生殖技术用于显微注射实验,如C57BL/6J或B6D2F1标准的小鼠品系进行了优化。产地不同,如远交品系小鼠,原则上可以用于基因组的编辑方法和可能提供更合适的遗传背景,具体的研究问题。如超数排卵辅助生殖技术的性能可以prediCTED为一些菌株34-36的,但可能需要为了进一步优化非标准菌株获得的胚胎的足够数量的核酸显微注射。
此外ZFN和TALEN,新设计师核酸酶如RNA引导的CRISPR/Cas9系统9,37,38现在已经被引入的基因组编辑应用程序。所有方法用于显微注射和此处描述的创始人动物的分析也适用于CRISPR/Cas9和基因组编辑的将来模式。
披露声明
作者宣称没有利益冲突。
致谢
我们要感谢莫妮卡Tarnowska的,科妮莉亚阿尔布雷希特和埃娃Skoczylas优秀的技术援助。这项研究是由SNF Sinergia授予CRSI33-125073于PP。
材料
| Name | Company | Catalog Number | Comments |
| BsaI | NEB | R0535S or L | |
| Esp3I | Thermo Scientific | ER0451 | |
| T4 Ligase | NEB | M0202S or L | |
| Spectinomycin | Sigma | S0692-1ML | |
| Ampicillin | Sigma | A0166 | |
| X-Gal | Sigma | B4252 | |
| IPTG | Sigma | I6758 | |
| Plasmid-Safe nuclease | Epicentre | E3101K | |
| QIAprep Spin Miniprep Kit | Qiagen | 27106 | |
| QIAGEN Plasmid Midi Kit | Qiagen | 12143 | |
| QIAquick PCR Purification Kit | Qiagen | 28106 | |
| mMESSAGE mMACHINE T7 Ultra Kit | Invitrogen | AM1345 | |
| NucAway Spin Columns | Invitrogen | AM10070 | |
| RNaseZAP | Sigma | R2020-250ML | |
| NorthernMax Formaldyde Load Dye | Invitrogen | AM8552 | |
| RNA Millennium Markers | Invitrogen | AM7150 | |
| 10x TBE buffer | Thermo Scientific | B52 | |
| T7 endonuclease | NEB | M0302S or L | |
| pGEM-T EasyVector System I | Promega | A3600 | |
| SYBR Green I Nucleic Acid Gel Stain | Invitrogen | S-7563 | |
| pregnant mare's serum gonadotrophin (PMSG) | Sigma | G4877 | |
| human chorionic gonadotropin (hCG) | Sigma | CG5 | |
| M2 embryo culture medium | Sigma | M7167 | |
| M16 embryo culture medium | Sigma | M7292 | |
| Mineral oil, embryo tested | Sigma | M8410 | |
| Ketamine | CentraVet | Ket 201 | |
| Xylazine | Sigma Aldrich | 46995 | |
| Equipment/Tools | |||
| Inverted microscope with Nomarski DIC optics (for example Nikon Eclipse TE200) | Nikon | ||
| Micromanipulator units (for example Narishige, NT88NF) | Narishige | ||
| Embryo holding capillaries | Sutter Instruments | B100-75-10 | |
| Embryo injection capillaries | Narishige | GD-1 | |
| Capillary puller (for example Sutter P97) | Sutter Instruments | ||
| Microforge (for example Narishige MF-900) | Narishige | ||
| Walton skin scissors | FST | 14077-10 | |
| Surgical scissors | FST | 14041-10 | |
| Surgical probe | FST | 10140-03 | |
| Reflex wound clip system (9 mm) | FST | 12031-09 | |
| Reflex wound clips (9 mm) | FST | 12032-09 | |
| Dumont fine forceps 5 | FST | 11254-20 | |
| Moria curved forceps | FST | 11370-31 | |
| Moria fine forceps | FST | 11399-80 | |
| Dietrich bulldog clamp | FST | 18038-45 | |
| C57BL/6J mice | Jackson Labs | strain code 000664 | |
| CD-1 mice | Charles River | strain code 000664 | |
参考文献
- Capecchi, M. R. Gene targeting in mice: functional analysis of the mammalian genome for the twenty-first century. Nat. Rev. Genetics. 6, 507-512 (2005).
- Johansson, T., et al. Building a zoo of mice for genetic analyses: a comprehensive protocol for the rapid generation of BAC transgenic mice. Genesis. 48, 264-280 (2010).
- Skarnes, W. C., et al. A conditional knockout resource for the genome-wide study of mouse gene function. Nature. 474, 337-342 (2011).
- Carbery, I. D., et al. Targeted genome modification in mice using zinc-finger nucleases. Genetics. 186, 451-459 (2010).
- Meyer, M., de Angelis, M. H., Wurst, W., Kuhn, R. Gene targeting by homologous recombination in mouse zygotes mediated by zinc-finger nucleases. Proc. Natl. Acad. Sci. U.S.A. 107, 15022-15026 (2010).
- Cui, X., et al. Targeted integration in rat and mouse embryos with zinc-finger nucleases. Nat. Biotechnol. 29, 64-67 (2011).
- Sung, Y. H., et al. Knockout mice created by TALEN-mediated gene targeting. Nat. Biotechnol. 31, 23-24 (2013).
- Wefers, B., et al. Direct production of mouse disease models by embryo microinjection of TALENs and oligodeoxynucleotides. Proc. Natl. Acad. Sci. U.S.A. 110, 3782-3787 (2013).
- Wang, H., et al. One-Step generation of mice carrying mutations in multiple genes by CRISPR/Cas-mediated genome engineering. Cell. 153, 910-918 (2013).
- Urnov, F. D., Rebar, E. J., Holmes, M. C., Zhang, H. S., Gregory, P. D. Genome editing with engineered zinc finger nucleases. Nat. Rev. Genetics. 11, 636-646 (2010).
- ZFN, T. A. L. E. N. CRISPR/Cas-based methods for genome engineering. Trends Biotechnol. 31, 397-405 (2013).
- Joung, J. K., Sander, J. D. TALENs: a widely applicable technology for targeted genome editing. Nat. Rev. Mol. Cell Biol. 14, 49-55 (2013).
- Cermak, T., et al. Efficient design and assembly of custom TALEN and other TAL effector-based constructs for DNA targeting. Nucleic Acids Res. 39, (2011).
- Reyon, D., et al. FLASH assembly of TALENs for high-throughput genome editing. Nat. Biotechnol. 30, 460-465 (2012).
- Sanjana, N. E., et al. A transcription activator-like effector toolbox for genome engineering. Nat. Protoc. 7, 171-192 (2012).
- Kim, Y., et al. A library of TAL effector nucleases spanning the human genome. Nat. Biotechnol. 31, 251-258 (2013).
- Schmid-Burgk, J. L., Schmidt, T., Kaiser, V., Honing, K., Hornung, V. A ligation-independent cloning technique for high-throughput assembly of transcription activator-like effector genes. Nat. Biotechnol. 31, 76-81 (2013).
- Hsu, P. D., et al. DNA targeting specificity of RNA-guided Cas9 nucleases. Nat. Biotechnol. 31, 827-832 (2013).
- Fu, Y., et al. High-frequency off-target mutagenesis induced by CRISPR-Cas nucleases in human cells. Nat. Biotechnol. 31, 822-826 (2013).
- Hermann, M., et al. Evaluation of OPEN zinc finger nucleases for direct gene targeting of the ROSA26 locus in mouse embryos. PLoS ONE. 7, (2012).
- Meyer, M., Ortiz, O., Hrabe de Angelis, M., Wurst, W., Kuhn, R. Modeling disease mutations by gene targeting in one-cell mouse embryos. Proc. Natl. Acad. Sci. U.S.A. 109, 9354-9359 (2012).
- Doyle, E. L., et al. TAL Effector-Nucleotide Targeter (TALE-NT) 2.0: tools for TAL effector design and target prediction. Nucleic Acids Res. 40, 117-122 (2012).
- Miller, J. C., et al. A TALE nuclease architecture for efficient genome editing. Nat. Biotechnol. 29, 143-148 (2011).
- Guo, J., Gaj, T., Barbas, C. F. Directed evolution of an enhanced and highly efficient FokI cleavage domain for zinc finger nucleases. J. Mol. Biol. 400, 96-107 (2010).
- Doyon, Y., et al. Enhancing zinc-finger-nuclease activity with improved obligate heterodimeric architectures. Nat. Methods. 8, 74-79 (2011).
- Wefers, B., et al. . Current Protocols in Mouse Biology. , (2011).
- Wefers, B., Wurst, W., Kühn, R. . Current Protocols in Mouse Biology. , (2011).
- Haueter, S., et al. Genetic vasectomy-overexpression of Prm1-EGFP fusion protein in elongating spermatids causes dominant male sterility in mice. Genesis. 48, 151-160 (2010).
- Perez-Pinera, P., Ousterout, D. G., Brown, M. T., Gersbach, C. A. Gene targeting to the ROSA26 locus directed by engineered zinc finger nucleases. Nucleic Acids Res. 40, 3741-3752 (2012).
- Bultmann, S., et al. Targeted transcriptional activation of silent oct4 pluripotency gene by combining designer TALEs and inhibition of epigenetic modifiers. Nucleic Acids Res. 40, 5368-5377 (2012).
- Valton, J., et al. Overcoming TALE DNA binding domain sensitivity to cytosine methylation. J. Biol. Chem. 287, 38427-38432 (2012).
- Cradick, T. J., Ambrosini, G., Iseli, C., Bucher, P., McCaffrey, A. P. ZFN-Site searches genomes for zinc finger nuclease target sites and off-target sites. BMC Bioinform. 12, (2011).
- Doyle, E. L., et al. TAL Effector-Nucleotide Targeter (TALE-NT) 2.0: tools for TAL effector design and target prediction. Nucleic Acids Res. 40, 117-122 (2012).
- Byers, S. L., Payson, S. J., Taft, R. A. Performance of ten inbred mouse strains following assisted reproductive technologies (ARTs). Theriogenology. 65, 1716-1726 (2006).
- Luo, C., et al. Superovulation strategies for 6 commonly used mouse strains. J. Am. Assoc. Lab. Animal Sci. 50, 471-478 (2011).
- Pease, S., Saunders, T. L. . International Society for Transgenic Technologies. Advanced protocols for animal transgenesis : an ISTT manual. , (2011).
- Cong, L., et al. Multiplex genome engineering using CRISPR/Cas systems. Science. 339, 819-823 (2013).
- Mali, P., et al. RNA-Guided human genome engineering via Cas9. Science. 339, 823-826 (2013).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。