
Method Article
RING-E3型无二体气体在Vitro和Planta中的功能特性
摘要
本手稿的目的是为 RING 型 E3 泛性白化酶的综合生化和功能研究提供大纲。此多步骤管道,具有详细的协议,验证被测试蛋白质的酶活性,并演示如何将活动与功能联系起来。
摘要
泛化作为蛋白质的翻译后修饰,在真核细胞的平衡中起着重要的调节作用。76种氨基酸泛性基辛修饰剂与靶蛋白的共价附着,取决于多聚基丁链的长度和拓扑结构,可导致从蛋白质降解到改性蛋白质定位和/或活性变化等不同结果。三种酶依次催化泛化过程:E1泛基质激活酶、E2泛性联因结合酶和E3泛基丁结合酶。E3泛基辛结合酶决定基质特异性,因此代表一个非常有趣的研究课题。在这里,我们提出了一个全面的方法,研究酶活性和功能之间的关系的RING型E3泛性联结酶。此四步协议描述了 1) 如何通过针对保守的 RING 域的站点定向诱变生成 E3 连带缺乏突变体;2⁄3) 如何检查体外和植物体中的泛化活性;4) 如何将这些生化分析与被测试蛋白质的生物意义联系起来。生成E3型缺乏附联物突变体,该突变体仍与其基质相互作用,但不再用于降解,有助于测试体内的酶-基质相互作用。此外,保守的RING域中的突变通常具有占主导地位的负表型,可用于功能敲除研究,作为RNA干扰方法的替代方法。我们的方法经过优化,以研究植物寄生线虫效应器RHA1B的生物作用,它劫持了植物细胞中的宿主泛化系统,以促进寄生。通过对体内表达系统稍作修改,该协议可应用于任何RING型E3连体的分析,无论其来源如何。
引言
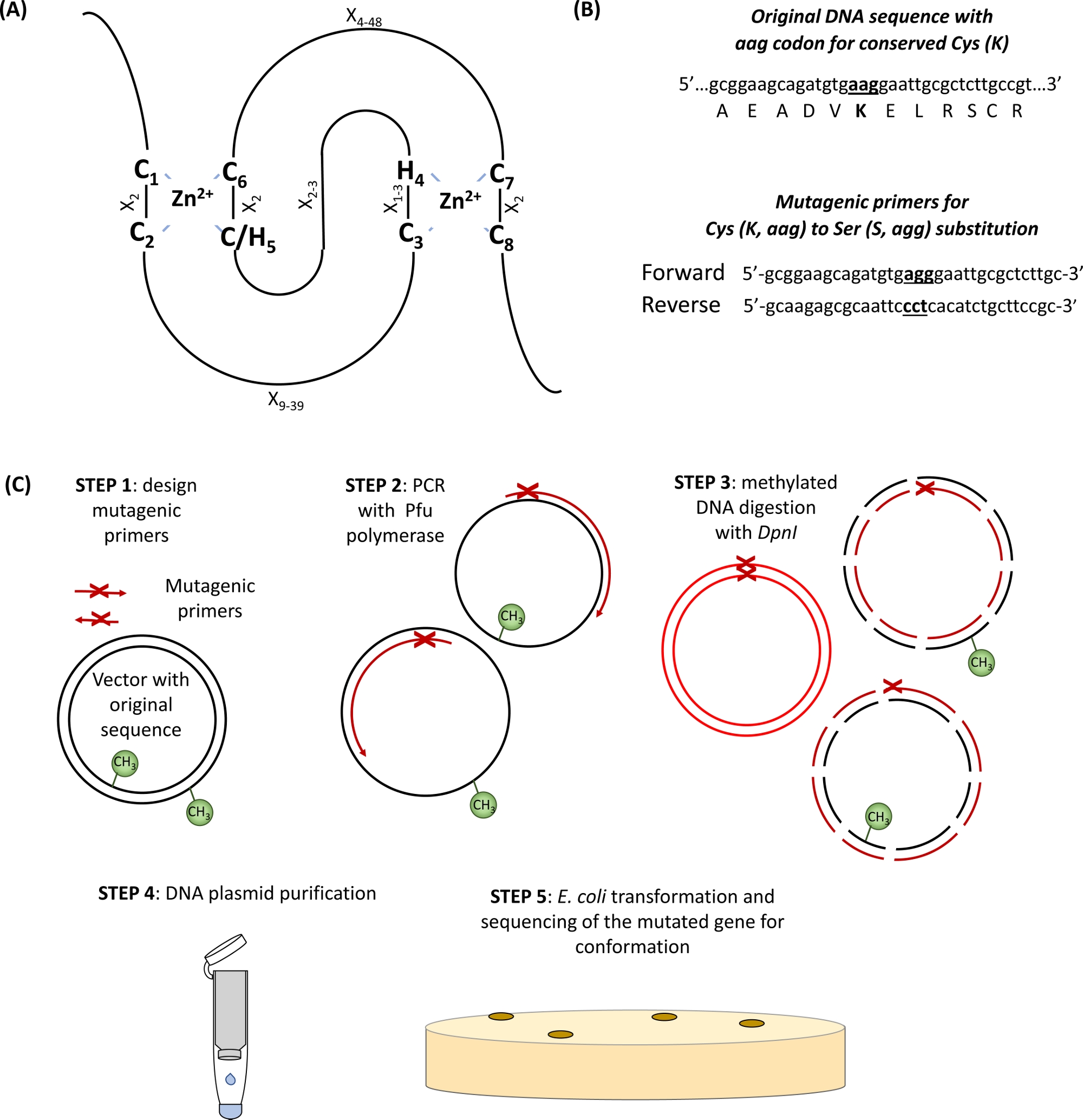
绝大多数的E3泛基蛋白属于RING(R eally Iinin ine n e)型蛋白质。环指域最初是由弗里蒙特等人识别的。1和功能描述为域中介蛋白-蛋白质相互作用2。规范的环指是一种特殊类型的锌协调域,定义为由 8 个保存的 Cys (C) 和他的 (H) 组成的共识序列,具体由其他氨基酸残留物 (X)、 C-X2-C-X9+39-C-X1+3-H-X2+3-C/H-X2-C-X4+48-C-X2 -C-C-X 2 -C-X - C-X 2 -C-C-X 2-C-X。两个 Zn2+离子通过独特的"交叉支撑"拓扑通过独特的"交叉支撑"拓扑稳定,C1/C2和 C/H5/C6协调第一个 Zn2+离子,而 C3/H4和 C7/C8绑定第二个 (图 1A)3,4。根据第五个Zn2+协调位点中C或H的存在,定义了环指蛋白的两个规范子类:C3HC4和C3H2C3(环-HC和环-H2)。由于E3泛素结合酶的环环域介导E2结合酶和基质之间的相互作用,这些必需的C和H残留物的突变已被证明会破坏结合酶活性5。还描述了另外五个不太常见的环 E3 利气亚类 (RING-v、 环-C2、 环-D、环-S/T 和环-G)6。RING型E3泛基体酶可进一步细分为简单和复杂的E3酶。简单的单亚单元环 E3 利气包含基板识别位点和 E2 绑定环域。相比之下,多子单元 RING 型 E3 复合体要么招募基板,要么将 E2-泛基质中间体与 E3 复合体进行中介结合。作为自泛化的主要泛泛物附着位点(s)的环域Lys残留物对于E3附着酶活动可能也很重要。
并非所有含环蛋白都作为E3利气蛋白发挥作用。因此,对环指域的生物信息预测和E2依赖性蛋白质的泛化能力必须经过生物化学验证,并链接到被测蛋白质的生物作用。在这里,我们描述了一个分步协议,概述如何通过位点定向诱变方法,在体外和植株中检测环型E3泛性利类气体的酶活性并表征功能。显示此管道的代表性结果,用于环型 E3 连带 RHA1B。RHA1B是植物寄生囊肿线虫球菌皮脂产生的一种效应蛋白,用于抑制植物免疫力,操纵植物根细胞的形态。为了保护自己免受病原体/寄生虫入侵,植物已经进化出核苷酸结合领域和富含亮氨酸的重复(NB-LRR)型免疫受体,检测病原体或寄生虫的存在,并因此产生高敏反应(HR),这是一种在感染地点发生的快速和局部细胞死亡,以阻止病原体的殖民化。其中一种免疫受体是马铃薯Gpa2蛋白,它能抵抗G.pallida(田间种群D383和D372)的一些分离物7。
使用提出的协议,最近发现RHA1B通过靶向植物Gpa2免疫受体进行泛化和降解8,以E3依赖性的方式干扰植物免疫信号。
研究方案
1. 现场导向的诱变 (图 1)
- 识别 RING 域中保存的 Cys 及其氨基酸(图 1A)和设计引物,在突变位点两侧携带 15 个碱基对,并带有感兴趣的替代子(图 1B)。
- 根据制造商的协议,在总PCR反应体积的50μL中,通过PCR对含有Pfu的扩热基质质基进行基于PCR的扩增,从而引入所需的突变。
- 通过在PCR反应中直接加入3μL的DpnI限制性酶(步骤1.2),并在37°C孵育2小时,消化大肠杆菌衍生的亲甲基化和半甲基化DNA。
注:甲基化是一种转录后蛋白质修饰,添加到产生和从细菌中分离出来的质粒中。PCR生成的质粒的新副本缺乏甲基化,因此,在DpnI治疗期间,新拷贝将保持不变。 - 使用基于旋转柱技术的商业DNA提取试剂盒纯化诱变质粒,并用50μL的水洗脱DNA。
- 根据制造商的协议,使用0.5 μL的诱变质粒DNA改造DH5+大肠杆菌化学能力细胞。简而言之,用DNA在冰上孵育合格细胞30分钟,然后在42°C下热休克20秒,在冰上再次放置管2分钟。 在37°C下用500μL的LB培养基孵育细胞1小时,在250rpm时将其传播。
- 通过桑格测序从大肠杆菌中分离的DNA质粒来验证所需的突变。
2. 重组蛋白纯化和体外泛化测定
- 将野生类型 RING 和突变的环状基因克隆到 pMAL-c2 载体中(遵循制造商的协议;表 3)将这些基因与MBP表位标记融合,允许使用米糖树脂进行一步纯化。如步骤 1.5 所述,将生成的结构引入大肠杆菌BL21 菌株中。
- 在37°C 的 50 mL LB 液体介质中生长大肠杆菌菌株 BL21,持续 2~3 小时,直到达到对数相(OD600的 0.4±0.6)。
- 将IPTG添加到0.1~1 mM的最终浓度中,以诱导感兴趣的MBP标记重组蛋白的表达,并在28°C下孵育大肠杆菌培养物2⁄3小时。孵化后将培养在冰上。
注:在冰上执行步骤2.4~2.13,保护蛋白质不降解。 - 要检查诱导效率,收集1.5 mL的诱导细胞, 在 13,000 x g下旋转 2 分钟,去除上清液,并在 20 μL 的 2x SDS-PAGE 加载缓冲液(24 mM Tris-HCl pH 6.8, 0.8% SDS, 10% (v/v) 甘油, 4 mM DTT, 0.04% (w/v) 溴酚蓝色) 中重新悬浮细胞。
- 将样品煮沸 5 分钟,然后用 10% SDS-PAGE 凝胶进行。要直观地评估 MBP 融合蛋白的积累(感兴趣的蛋白质的分子量 = 42.5 kDa MBP),使用 Coomassie 染色缓冲液搅拌,将凝胶染色 20 分钟(50% 甲醇、10% 醋酸、0.1% Coomassie 蓝色),并一夜之间与脱色缓冲液(20%甲醇,10%醋酸)。
- 在1,350 x g下离心6分钟,丢弃上清液,用5 mL的柱缓冲液(20 mM Tris HCl,200 mM NaCl,1mM EDTA,细菌蛋白酶抑制剂)重新悬浮细胞颗粒,收获剩余的大肠杆菌细胞。
注意:这是一个在一夜之间停止协议的好地方。冷冻细胞可在-20°C下储存长达1周。 - 将含有细菌的管子放入冰水浴中,并应用10个声波循环:在30%的安培下10s的声波,然后是20s的休息,从而分解大肠杆菌细胞。
- 在4°C下13,000 x g的离心样品10分钟,并保存上清液(粗提取物)。
- 在 15 mL 管中制备 500 μL 的糖脂。通过加入10 mL的冷柱缓冲液和在1,800 x g下离心,4°C 5分钟,清洗树脂。做这个2次。
- 将5 mL的粗提取物加入其中,加入其中的糖脂,并在4°C下孵育过夜。
- 在4°C下在1,800 x g下离心5分钟,并丢弃上清液。
- 在树脂颗粒中加入10 mL的柱缓冲液,孵育20分钟。然后在4°C下以1,800 x g离心5分钟。重复此步骤2倍。
- 通过在4°C下孵育样品2小时,用含有10 mM麦芽糖的0.5 mL柱缓冲液洗脱融合蛋白。在4°C下在1,800 x g下离心5分钟,并收集洗脱蛋白。重复此步骤 2 倍。
- 对冷PBS的蛋白质分量进行1 mL的透析。将蛋白添加到一次性管中(10~20 μL),避免冻融,并储存在-80°C,直到需要为止。
- 使用布拉德福德测定方法9测量蛋白质浓度。
- 通过混合 40 ng 的 E1(例如 AtUBA1)、E2 的 100 纳克(例如 AtUBC8、SlUBC1/4/6/7/12/13/17/20/22/22/27/32)、1 μg MBP-RING 型蛋白质,在总体积为 30 μL 的体外泛化反应中设置外体泛化反应 和 2 μg FLAG-Ub (或 HA-Ub) 在泛化缓冲液 (50 mM Tris-HCl pH 7.5, 2 mM MgCl2, 30 mM 磷酸肌酸, 50 μg/mL 肌酸磷酸酶).在30°C孵育混合物2小时。
注:预制20x泛化缓冲液,并将其储存在-20°C的小等分中,用于一次性使用。当缓冲液解冻和反复冷冻时,肌酸磷酸激酶很容易失去酶活性。 - 通过将 30 μL 样品与 7.5 μL 的 5x SDS-PAGE 加载缓冲液(60 mM Tris-HCl pH 6.8,2% SDS、25%(v/v) 甘油、10 mM DTT、0.1%(w/v) 溴酚蓝)混合并沸腾 5 分钟,从而终止反应。
- 用7.5%SDS-聚丙烯酰胺凝胶电泳(SDS-PAGE)分离蛋白质,然后转移到PDVF膜上,并使用抗FLAG(或抗HA)检测渗透。
- 用库马西蓝染色PVDF膜,以验证经测试的MBP-RING型蛋白质的同等载荷。
3. 在尼科蒂亚纳弯曲的叶和植物中,农业细菌介导的瞬态蛋白表达
- 携带表位标记相关基因(例如,HA-RHA1B、HA-RHA1BC135S、HA-RHA1BK146R、HA-Ub)和空载体的带状的带状的表位标记基因的带状的带状的带状的带状农业细菌菌株,以及含有适当选择抗生素的LB介质上的空载体。
- 在28°C下生长2天后,拿起单菌群,在LB液体介质中用适当的抗生素在28°C/250rpm下再生长24小时。
- 使用适当的抗生素将 100 μL 的农业细菌培养物转移到 3 mL 新鲜 LB,并在 28°C 下在 28°C 下孵育培养物,在旋转(250 rpm)至晚期指数生长阶段。
- 以1,800 x g旋转农业细菌细胞6分钟,丢弃上清液,用3mL洗涤缓冲液(50m MES pH 5.6,28 mM葡萄糖,2 mM NaH2PO4)重新悬浮细胞。重复此步骤 2 倍。
- 第二次洗涤后,重新悬浮诱导缓冲液中的细胞(50 mM MES pH 5.6,28 mM MM 葡萄糖,2 mM NaH2PO 4,200μM 醋酸酮,37 mM NH4Cl,5 mM MgSO4.7H2O,4 mM KCl,18 μM FeSO4.7H2O,2 mM CaCl2。在28°C下,用诱导缓冲液孵育细胞10~12小时。
注: 醋酸酮诱导T-DNA转移。 - 在1,800 x g下将细胞离心6分钟,丢弃上清液,用2 mL渗透缓冲液(10 mM MES pH 5.5,200 μM醋酮)重新悬浮细胞。
注:如果Agro细菌在用诱导缓冲液孵育后聚集,则让聚合的细胞沉入管底,将其留在长凳上几分钟,并将明确的农业细菌悬浮液转移到新管中,然后再进行步骤3.6。 - 使用 OD600值(600 nm 吸收时的光密度)测量细菌的浓度。将 OD600值调整为所需的值。
注: 通常 0.2-0.4 之间的 OD600值最适合单个农用细菌染色表达。如果应用了不同农业细菌菌株的组合,则农业细菌菌株的总 OD600值不应超过 1。 - 农用Infiltrate4周大的N.Bethamiana叶,用针轻轻刺破它们,然后用没有针头的注射器用手注射Agrobac。用标记圈入渗入的叶子区域(通常直径为 1⁄2 厘米)。
- 收集渗透叶组织36小时后渗透。用液氮将组织研磨为细粉末。
- 具有 300 μL 蛋白质提取缓冲液(50 mM Tris-HCl pH 7.5,150 mM NaCl,5 mM EDTA,2 mM DTT,10% 甘油,1% 多聚苯丙酮,1 mM PMSF,植物蛋白酶抑制剂鸡尾酒)和离心机在 15,000 x g下,在 4 °C 下 15 分钟。
- 将上清液转移到新管中。将 5x SDS-PAGE 加载缓冲液添加到 1x 的最终浓度,煮 5 分钟。
- 在10%SDS-PAGE凝胶上分离粗蛋白,转移到PVDF膜上,用抗HA探针检测植物中无所不在。
4. 建立植物酶活性与功能之间的联系
注:例如,RHA1B促进耐药蛋白Gpa2的降解,以抑制HR细胞死亡。此步骤演示如何验证 RHA1B 的这些毒性活动是否与 E3 相关。
- 带标记感兴趣的基因(在本例中HA-RHA1B、HA-RHA1B C135S、HA-RHA1BK146R、myc-Gpa2、RBP1)和空载体作为对照的适宜的农用细菌菌株。按照步骤3.1~3.8进行农业细菌制备和注射N.贝萨米亚纳叶。
- 对于E3依赖性基质蛋白降解,按照步骤3.9~3.12执行西方印迹,使用适当的抗体检测植物细胞中的蛋白质积累(例如抗HA和抗MYC)。
- 对于E3依赖性高敏反应(HR)介导细胞死亡抑制,监测农浸叶的HR细胞死亡症状2-4天后渗透。
结果
本节中提供了用于检查单个亚单位 E3 泛性酶 RHA1B 的协议的代表性结果,该协议具有 PROSITE 预测环-H2 类型域 (132–176 氨基酸)10。如图1所示,为了获得 E3 缺乏突变蛋白,在 RING 域(图 1A)中的 8 个保存的 Cs 或 Hs 中,至少有一个需要诱变(图 1B)。因此,作为第一步,生成了 RHA1B、RHA1BC135S(在环圈域的保守 C3中由 Ser 替换 Cys)和 RHA1BK146R(在 RHA1B 中唯一的 Lys 中由 Arg 替换的 Lys)的两个突变版本。虽然单亚单位E3利气介导泛性泛基质从泛化物转移至基质,而不是与泛化物直接相互作用,但其最大酶活性可能需要在Lys中自我泛化。
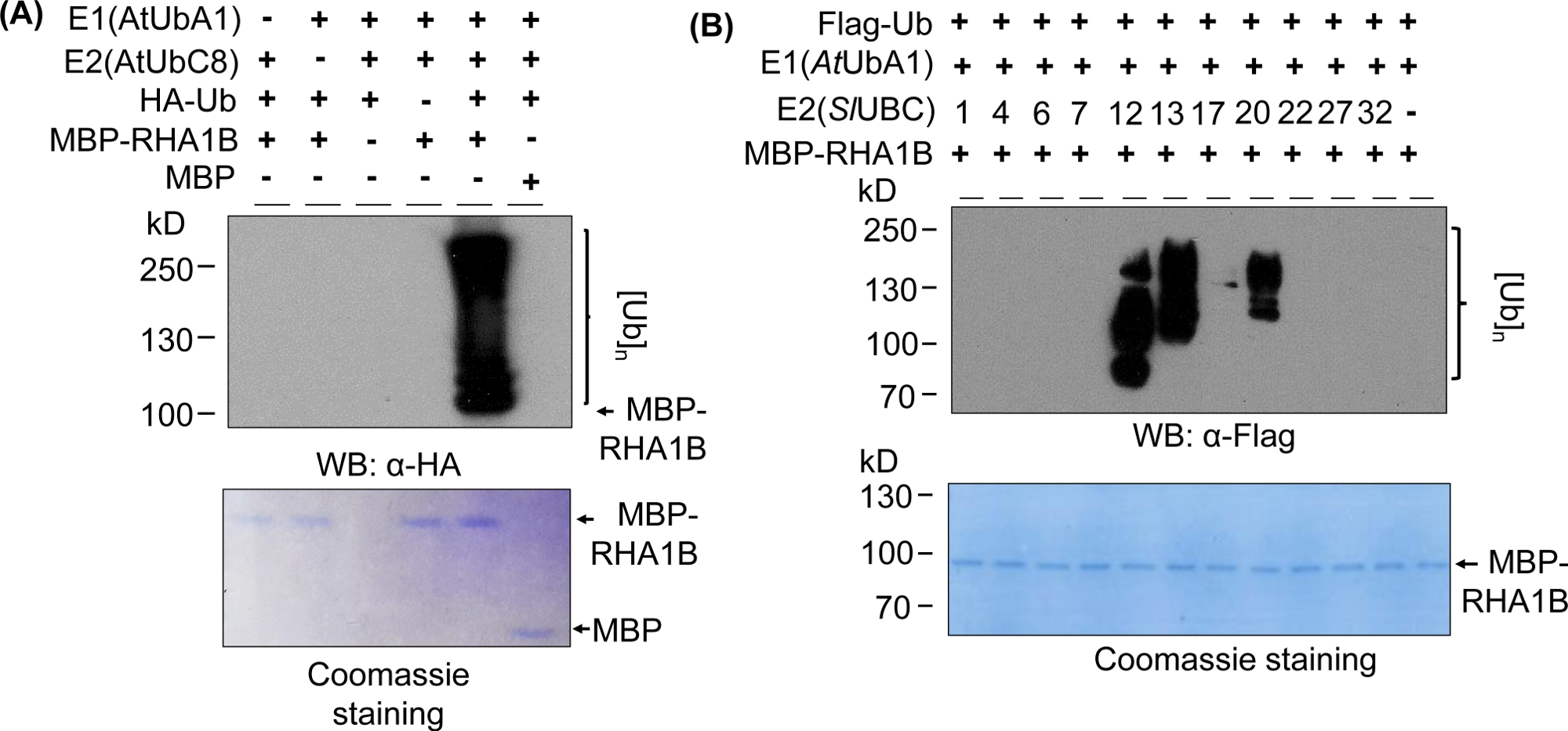
图2A中的西方印迹结果显示了典型的体外泛化检测结果,从被测蛋白质的分子量(例如MPB熔融RHA1B =100 kDa)开始,多带涂片开始,并呈上升趋势。抗HA抗体识别HA标记Ub纳入不同长度的多泛化链,形成这种典型的泛性与阶梯状的涂片。为了验证阳性结果,图 2A还显示了缺少单个组件(E1、E2、Ub 或 MBP-RHA1B)或使用 MBP 作为控制和缺乏涂抹泛化信号的所有重要负控制。此外,PVDF膜的Coomassie蓝色染色在所有对照中均显示MBP-RHA1B或MBP的负载。
图 2B显示了体外泛化结果如何因特定的 E2/E3 组合而异。在此示例中,测试了代表 10 个不同 E2 族的 11 个不同的 E2。检测到的泛化活性范围从无信号(无涂片)到从不同分子量开始的多带涂片,表明不同的泛化模式。
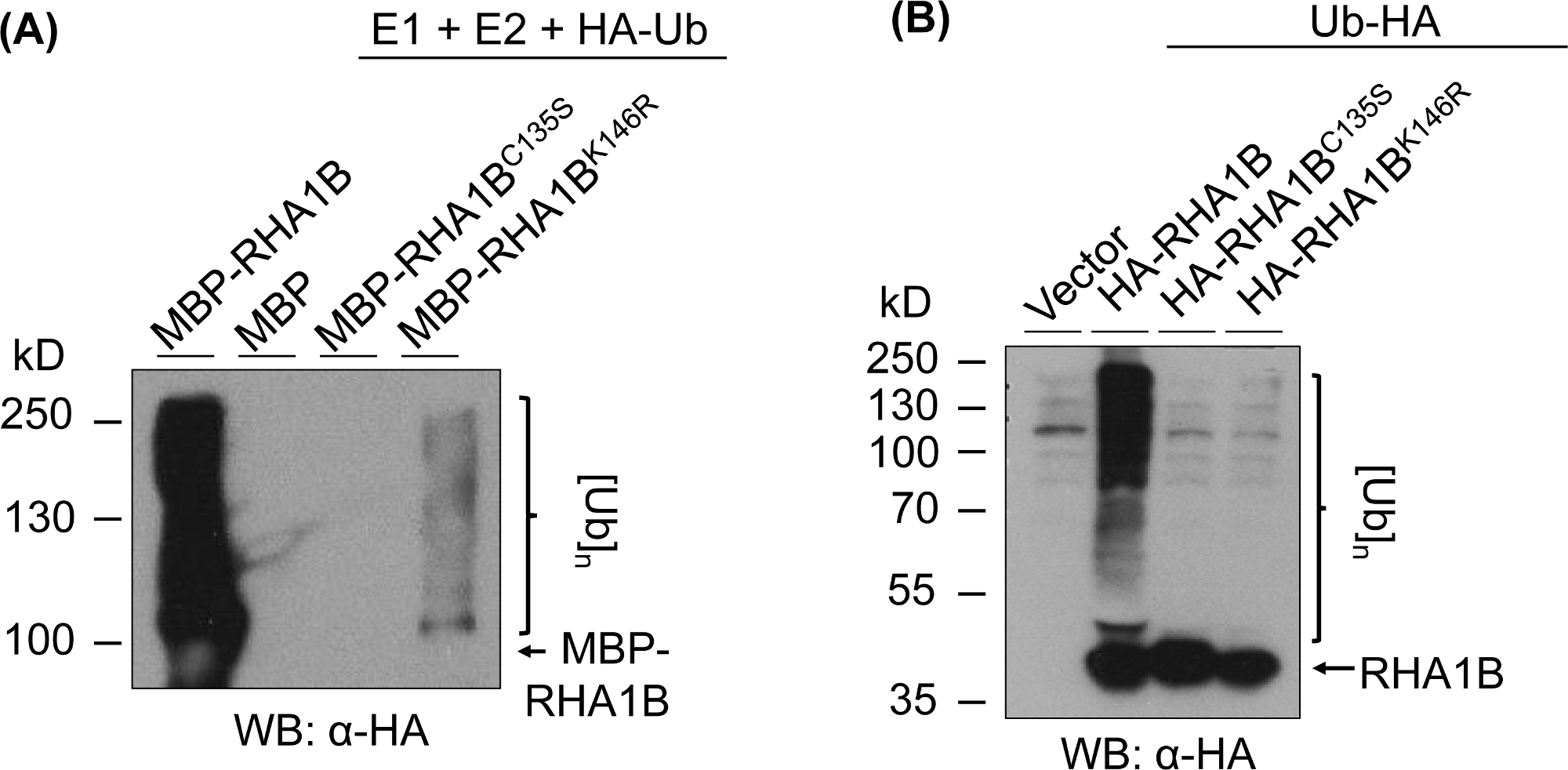
图3显示了已测试蛋白质的RING和K突变版本的泛化测定结果。RHA1BC135S缺乏酶活性是支持它无法在体外生成多带涂片(图 3A)或促进植物中的多泛化信号(图 3B)。值得注意的是,与野生RHA1B型的酶活性所赋予的强烈泛化信号相反,在植物中HA标记Ub的过度表达使所有测试样本(包括病媒对照)的基础水平均无所不在。此外,对RHA1BK146R突变物的分析表明,K146残留物对RHA1B的E3活性也至关重要。虽然在体外检测到边际自泛化信号(图3A),但植株测定突变体为E3缺陷(图3B,仅检测到背景泛化信号)。
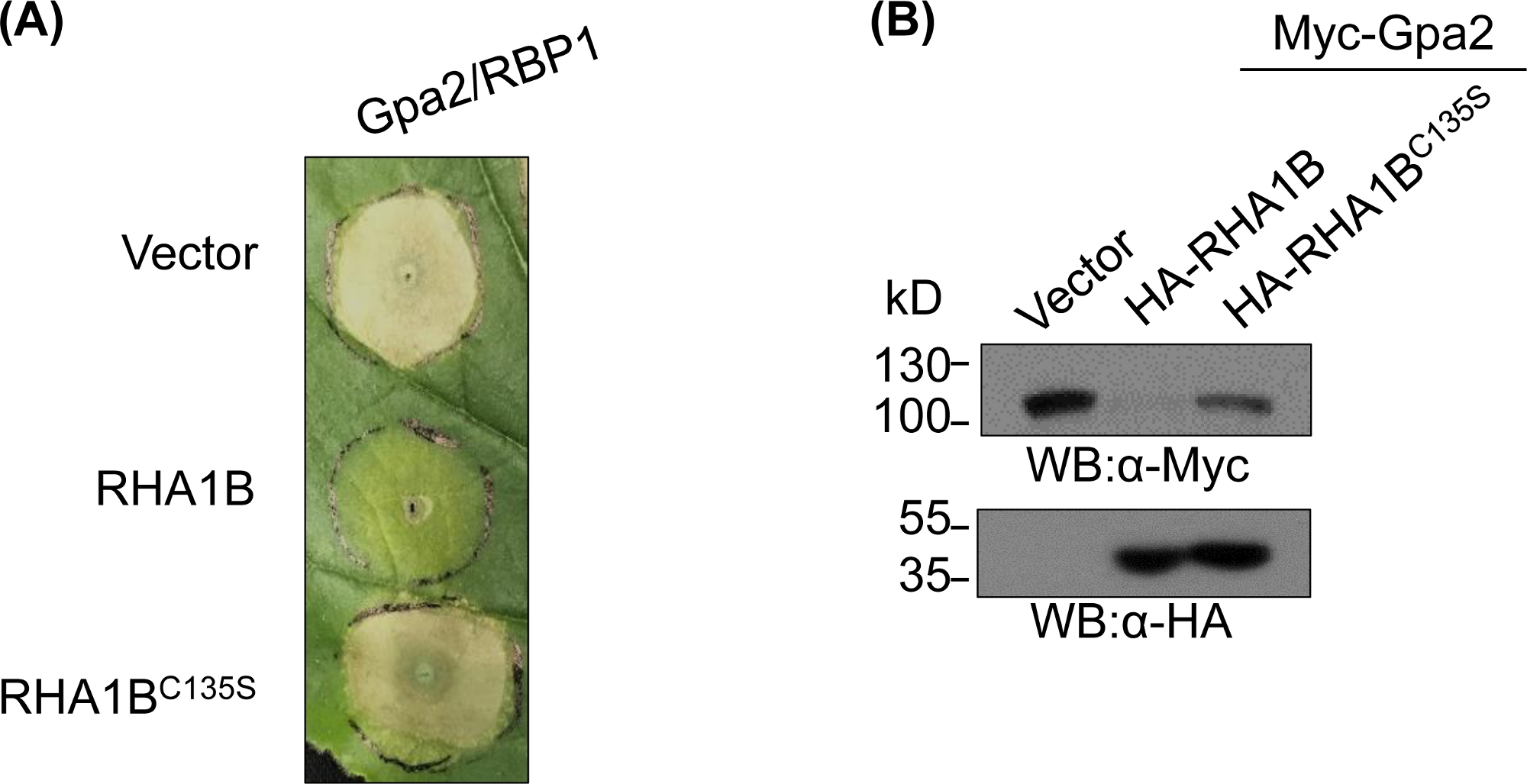
在生成和生化验证E3缺乏突变体后,功能研究可以设计以确定经测试的RING E3泛素联结酶的E3相关生物作用。在RHA1B的情况下,这种线虫效应器抑制植物免疫信号,表现为抑制Gpa2触发的HR细胞死亡。如图4A所示,与野生型RHA1B不同,缺乏E3连体活性的RHA1BC135S突变体不会干扰HR细胞死亡。鉴于蛋白质普遍渗透的最常见结果是其蛋白酶体介导的降解,驻留在RING域中的突变也可用于验证E3依赖性的能力,以触发其直接和/或间接基质的降解。因此,显著地,图4B中的西方印迹结果证实Gpa2在野生RHA1B存在的情况下没有积累,但RHA1BC135S对Gpa2蛋白稳定性没有影响。

图1:站点定向诱变所涉及的原理和步骤的原理表示。(A) 环-CH/H2 域,具有保存的 Cys 及其氨基酸。(B) 诱变引物设计的例子.(C) 站点定向诱变的步骤。请点击此处查看此图的较大版本。
{kind=link}

图2:代表性体外泛化测定。(A) 顶部凝胶显示泛化测定,包括所有负控制,而底部凝胶显示相等的负载。(B) 根据E2酶的预期结果范围。这个数字已由库德等人8修改。请点击此处查看此图的较大版本。
{kind=link}

图3:环和K突变体(RHA1BC135S和RHA1BK146R)的泛化测定结果。(A) RHA1BC135S和 RHA1BK146R的体外泛化结果。(B) 在植物中 RHA1BC135S和 RHA1BK146R的泛化测定结果。这个数字已由库德等人8修改。请点击此处查看此图的较大版本。
{kind=link}

图4:E3依赖生物功能的代表性功能研究。显示E3依赖性生物功能的功能研究示例。(A) E3依赖性HR细胞抑制和 (B) 植物免疫受体Gpa2的降解.这个数字已由库德等人8修改。请点击此处查看此图的较大版本。
{kind=link}
| PCR 设置 | |
| 1 μL | 质粒 (±100 ng) |
| 1.5 μL | F 诱变底漆 (10 μM) |
| 1.5 μL | R 诱变引物 (10 μM) |
| 1 μL | dNTP (10 mM) |
| 5 μL | 缓冲器 (10x) |
| 1 μL | 超普氟酶(2.5 U/μl) |
| 39 μL | ddH2O |
| 50 μL | 总容量 |
表1:PCR反应设置
| 热循环器程序 | |||
| 1 | 95 °C | 30 s | |
| 2 | 95 °C | 30 s | |
| 3 | 60 °C | 30 s | |
| 4 | 72 °C | 5分钟 | 重复 2-4 30 次 |
| 5 | 72 °C | 5分钟 | |
表2:PCR热循环器程序
| 为 RHA1B 示例设置结扎反应 | ||
| 1.5 μL | pMAL-c2:MBP 线性化矢量通过 BamHI 和 SalI (60 ng) 进行消化 | |
| 7 μL | RHA1B/RHA1BC135S 或 RHA1BK146R 刀片随 BamHI 和 SalI (25 ng) 一起消化 | |
| 1 μL | T4 连形缓冲液 (10x) | |
| 0.5 μL | T4 连带 (400 U/μL) | |
| 10 μL | 总容量 | |
表3:为RHA1B示例设置结扎反应。
讨论
阐明RING型E3泛基的生化和力学基础,有助于我们理解其在发育、应力信号和平衡维持中的生物学意义。这里描述的方案将诱变方法与体外和植物功能研究结合。通过在环域的保存残留物中引入单一氨基酸替代,通过位点直接诱变,可以同时测试产生的E3缺乏突变体与野生型蛋白质,将酶活性与功能联系起来。
正确识别环域至关重要,尤其是其保存的 Cys 及其残留物。在线工具,如PROSITE可用于做到这一点10。为了破坏负责招募E2酶的RING域的稳定,Cys通常被Ser取代,这是其最接近的结构替代物,缺乏创建用于锌协调的二硫化物键的能力。Lorick等人表明,这些关键Cys残留物中的突变将消除单亚单位RING型E3利气5的泛化活性。虽然一些Cys残留物对于含有RING型蛋白质的多单元E3连体复合物也很重要,但由于这些泛化复合物的多面和动态的三维结构以及RING型蛋白的不同作用,在多单元E3连体中,环域中保存的残留物的单一替代尚未成功产生一个缺乏环状的表型11。
对于位点导向的诱变,我们发现使用更小的质粒载体和较低的扩增周期通常能提高诱变效率。Pfu酶可以替代任何其他高保真和高工艺性DNA聚合酶。此外,如果感兴趣的基因含有稀有的科顿,另一种大肠杆菌染色,罗塞塔,可用于实现重组蛋白的更高产量。此外,还可以进一步优化IPTG诱导的孵育时间和温度。较低的温度降低大肠杆菌分裂率,这可能有利于某些蛋白质的表达。虽然IPTG浓度较高可以改善蛋白质表达,但它也抑制大肠杆菌分裂过程,不建议这样做。
单亚单位环型E3利气不仅作为分子支架的作用,将E2-Ub中间体定位在靠近基板的位置,而且还刺激其共性E2的泛基体转移活性。此外,鉴于E2/E3组合对于决定改性基板命运的多聚二苯链的长度和连接非常重要,因此对环型E3的任何考虑都必须包括其酶伙伴E2s12。如图3B所示,并非所有经过测试的E2都与RHA1B连带相容。因此,体外泛化检测应与代表不同E2类的多个E2酶并行进行,以避免误阴性结果。
这里介绍的是体外酶测定,检测已测试的RING型蛋白质的自我渗透能力。然而,通过小的修改,该协议可以很容易地适应检测基质的体外普及。为此,步骤2.15的体外泛化混合物应补充潜在E3结合基质(500 ng)的重组蛋白。在30°C孵育2小时后,应使用15μL的抗HA亲和基质基质(如果使用HA-Ub,如果使用FLAG-Ub)通过搅拌在4°C下搅拌2小时捕获泛化蛋白。使用冷 Ub 洗涤缓冲液(20 mM Tris pH 7.5、100 mM NaCl、0.1 mM EDTA、0.05% 补间 20、1x PMSF)洗涤珠子 4 次后,丢弃缓冲液的 40 μL,并移动到步骤 2.16。由分别由表位标记Ub和基质的抗体检测的泛化信号,分别从基质蛋白的分子量中产生,确认基质/酶特异性。
此外,由于瞬态酶-基质相互作用和泛化靶蛋白的快速降解,体内E3结合基质的鉴定通常与多重挑战相关。使用E3连体缺乏突变体,它仍然与其目标相互作用,但不再泛化它13,是一个非常有用的替代添加蛋白酶抑制剂MG132,并不总是充分干扰26S蛋白酶体功能。
RING型E3气的一个常见特征是形成和发挥作用作为同质和/或异体。有趣的是,在环域的保守残留物的替代通常与一个显性负表型相关,其中突变的RING型E3连体块酶活性的原生野生类型蛋白质13。因此,在植物中过度表达环突变体可能是敲出E3连体基因的替代方法。
披露声明
作者没有什么可透露的。
致谢
我们的工作是由美国农业部国家粮食和农业研究所、美国农业部-NIFA农场法案、西北马铃薯协会提供的财政支持(2017-67014-26197;2017-67014-26591)提供的。联盟,和ISDA特种作物。
材料
| Name | Company | Catalog Number | Comments |
| Acetic acid | Sigma-Aldrich | A6283 | |
| Acetosyringone | Sigma-Aldrich | D134406 | |
| Amylose resin | NEB | E8021S | |
| ATP | Sigma-Aldrich | A1852 | |
| Bacterial protease inhibitor | Sigma-Aldrich | P8465 | |
| Bromphenol Blue | VWR | 97061-690 | |
| CaCl2 | Sigma-Aldrich | C1016 | |
| Centrifuge | Beckman Coulter | model: Avanti J-25 | |
| Commassie Blue | VWR | 97061-738 | |
| Creatine phosphate | Sigma-Aldrich | P7936 | |
| Creatine phosphokinase | Sigma-Aldrich | C3755 | |
| DNA clean & concentrator Kit | ZYMO RESEARCH | D4029 | |
| DpnI | NEB | R0176S | |
| DTT | Sigma-Aldrich | D0632 | |
| E. coli BL21 | Thermo Fisher Scientific | C600003 | |
| E. coli DH5α competent cells | Thermo Fisher Scientific | 18265017 | |
| EDTA | Sigma-Aldrich | 324504 | |
| FeSO4 7H2O | Sigma-Aldrich | F7002 | |
| FLAG-Ub | BostonBiochem | U-120 | |
| Glucose | VWR | 188 | |
| Glycerol | Sigma-Aldrich | G5516 | |
| HA-Ub | BostonBiochem | U-110 | |
| Heat block | VWR | model: 10153-318 | |
| Incubator | VWR | model: 1525 Digital Incubator | |
| Incubator shaker | Thermo Fisher Scientific | model: MaxQ 4000 | |
| IPTG | Roche | 10724815001 | |
| KCl | Sigma-Aldrich | P9333 | |
| LB Broth | Sigma-Aldrich | L3022 | |
| Liquide nitrogen | university chemistore | ||
| Maltose | Sigma-Aldrich | 63418 | |
| MES | Sigma-Aldrich | M3671 | |
| Methanol | Sigma-Aldrich | 34860 | |
| MgCl2 | Sigma-Aldrich | 63138 | |
| MgSO4 7H2O | Sigma-Aldrich | 63138 | |
| Microcentrifuge | Eppendorf | model: 5424 | |
| Miniprep plasmid purification kit | ZYMO RESEARCH | D4015 | |
| monoclonal anti-FLAG antibody | Sigma-Aldrich | F3165 | |
| monoclonal anti-HA antibody | Sigma-Aldrich | H9658 | |
| monoclonal anti-MYC antibody | Sigma-Aldrich | WH0004609M2 | |
| Mortar | VWR | 89038-144 | |
| NaCl | Sigma-Aldrich | S7653 | |
| NaH2PO4 | Sigma-Aldrich | S8282 | |
| NanoDrop | Thermo Fisher Scientific | model: 2000 Spectrophotometer | |
| Needle | Thermo Fisher Scientific | 14-826-5C | |
| NH4Cl | Sigma-Aldrich | A9434 | |
| PCR machine | Bio-Rad | model: C1000 | |
| Pestle | VWR | 89038-160 | |
| Pfu Ultra | Agilent Technologies | 600380 | |
| Plant protease inhibitor coctail | Sigma-Aldrich | P9599 | |
| pMAL-c2 | NEB | N8076S | |
| PMSF | Sigma-Aldrich | P7626 | |
| Polyvinylpolypyrrolidone | Sigma-Aldrich | P6755 | |
| SDS | Sigma-Aldrich | 1614363 | |
| Sonicator | Qsonica Sonicators | model: Q125 | |
| Syringe | Thermo Fisher Scientific | 22-253-260 | |
| Tris | Sigma-Aldrich | T1503 | |
| T4 ligase | NEB | M0202S |
参考文献
- Freemont, P. S., Hanson, I. M., Trowsdale, J. A novel gysteine-rich sequence motif. Cell. 64, 483-484 (1991).
- Borden, K. L. B. RING fingers and B-boxes: Zinc-binding protein-protein interaction domains. Biochemistry and Cell Biology. 76, 351-358 (1998).
- Barlow, P. N., Luisi, B., Milner, A., Elliott, M., Everett, R. Structure of the C3HC4 Domain by 1H-nuclear Magnetic Resonance Spectroscopy: A New Structural Class of Zinc-finger. Journal of Molecular Biology. 237, 201-211 (1994).
- Borden, K. L. B., et al. The solution structure of the RING finger domain from the acute promyelocytic leukaemia proto-oncoprotein PML. The EMBO Journal. 14, 1532-1541 (1995).
- Lorick, K. L., et al. RING fingers mediate ubiquitin-conjugating enzyme (E2)-dependent ubiquitination. Proceedings of the National Academy of Sciences. 96, 11364-11369 (1999).
- Jiménez-López, D., Muñóz-Belman, F., González-Prieto, J. M., Aguilar-Hernández, V., Guzmán, P. Repertoire of plant RING E3 ubiquitin ligases revisited: New groups counting gene families and single genes. PLoS ONE. 13, 1-28 (2018).
- Sacco, M. A., et al. The Cyst Nematode SPRYSEC Protein RBP-1 Elicits Gpa2- and RanGAP2-Dependent Plant Cell Death. PLoS Pathogens. 5, 1-14 (2009).
- Kud, J., et al. The potato cyst nematode effector RHA1B is a ubiquitin ligase and uses two distinct mechanisms to suppress plant immune signaling. PLoS Pathogens. 15, 1007720 (2019).
- Bradford, M. M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry. 72, 248-254 (1976).
- Sigrist, C. J. A., et al. New and continuing developments at PROSITE. Nucleic Acids Research. 41, 344-347 (2013).
- Dove, K. K., Stieglitz, B., Duncan, E. D., Rittinger, K., Klevit, R. E. Molecular insights into RBR E3 ligase ubiquitin transfer mechanisms. EMBO Reports. 17, 1221-1235 (2016).
- Metzger, M. B., Pruneda, J. N., Klevit, R. E., Weissman, A. M. RING-type E3 ligases: Master manipulators of E2 ubiquitin-conjugating enzymes and ubiquitination. Biochimica et Biophysica Acta - Molecular Cell Research. 1843, 47-60 (2014).
- Xie, Q., et al. SINAT5 promotes ubiquitin-related degradation of NAC1 to attenuate auxin signals. Nature. 419, 167-170 (2002).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。