
Method Article
Caracterização funcional do RING-Tipo E3 Ubiquitin Ligases In Vitro e In Planta
Neste Artigo
Resumo
O objetivo deste manuscrito é apresentar um esboço para os estudos bioquímicos e funcionais abrangentes das ligaduras de ubiquitina ring-tipo E3. Este pipeline multipasso, com protocolos detalhados, valida uma atividade enzimática da proteína testada e demonstra como vincular a atividade à função.
Resumo
A ubiquitinação, como uma modificação pós-translacional de proteínas, desempenha um papel regulatório importante na homeostase das células eucarióticas. O acessório covalente de 76 modificadores de ubiquitina de aminoácidos a uma proteína alvo, dependendo do comprimento e da topologia da cadeia de poliubiquitina, pode resultar em diferentes resultados que vão desde a degradação de proteínas até mudanças na localização e/ou atividade de proteína modificada. Três enzimas catalisam sequencialmente o processo de ubiquitinação: enzima ativadora de ubiquitina E1, enzima de conjugação de ubiquitina E2 e ligase de ubiquitina E3. A ligase de ubiquitina e3 determina a especificidade do substrato e, portanto, representa um assunto de estudo muito interessante. Aqui apresentamos uma abordagem abrangente para estudar a relação entre a atividade enzimática e a função da ligase ubiquitina ring-tipo E3. Este protocolo de quatro etapas descreve 1) como gerar um mutante deficiente da Ligase E3 através da mutagênese dirigida pelo local, direcionada ao domínio RING conservado; 2-3) como examinar a atividade de ubiquitinação tanto in vitro quanto na planta; 4) como ligar essas análises bioquímicas ao significado biológico da proteína testada. A geração de um mutante deficiente em ligase E3 que ainda interage com seu substrato, mas não o ubiquitina para degradação, facilita o teste de interações com substrato de enzimas in vivo. Além disso, a mutação no domínio ring conservado muitas vezes confere um fenótipo negativo dominante que pode ser utilizado em estudos de nocaute funcional como uma abordagem alternativa a uma abordagem de interferência de RNA. Nossos métodos foram otimizados para investigar o papel biológico do efetuor de nematóide parasitário da planta RHA1B, que sequestra o sistema de ubiquitinação hospedeiro nas células vegetais para promover o parasitismo. Com ligeira modificação do sistema de expressão in vivo, este protocolo pode ser aplicado à análise de qualquer ligase E3 do tipo RING, independentemente de suas origens.
Introdução
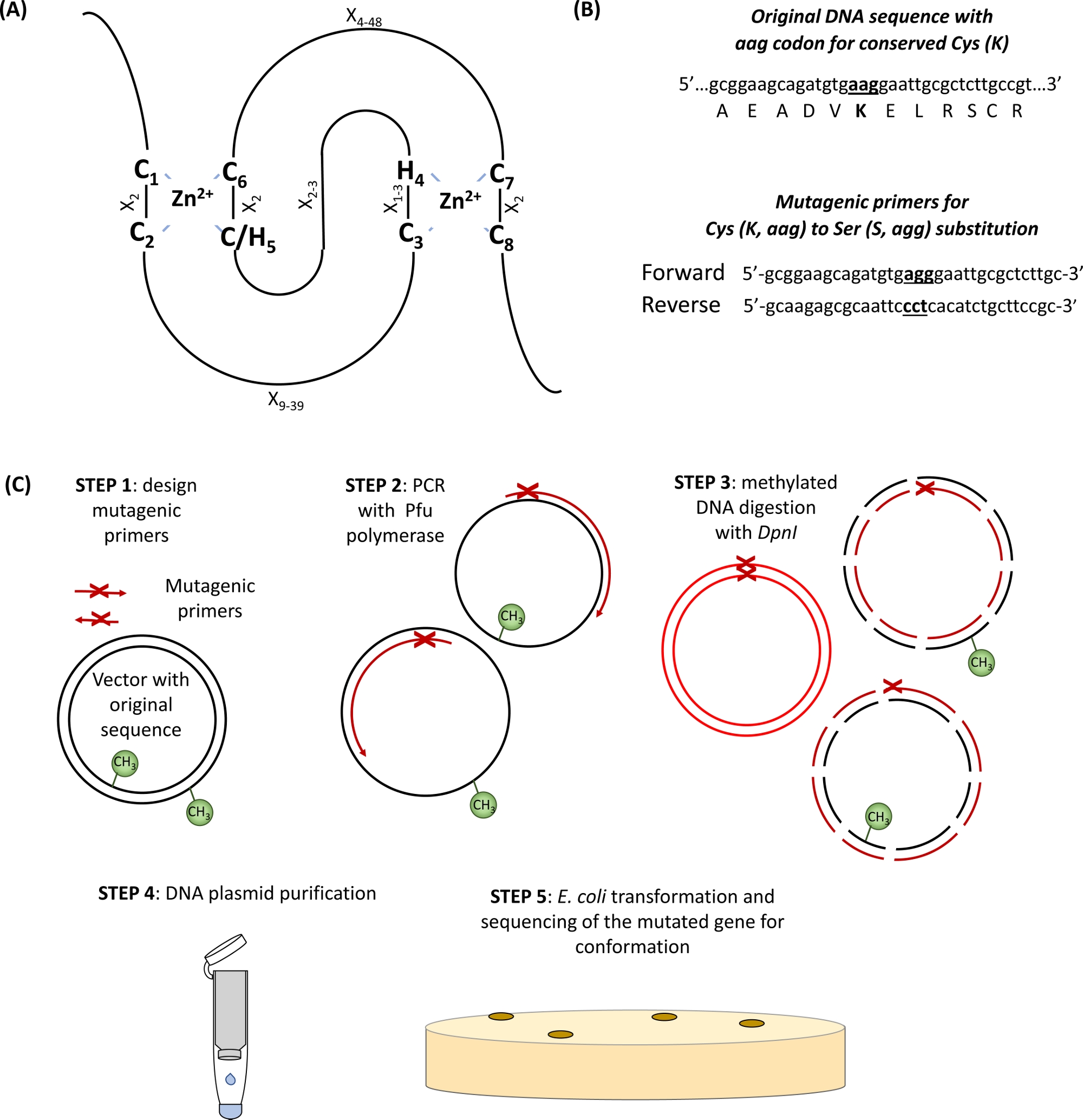
A grande maioria das ligas de ubiquitina E3 pertence ao RING(Really interesting New Gene) proteínas do tipo. O domínio ring-finger foi originalmente identificado por Freemont et al. 1 e funcionalmente descrito como um domínio mediador interação proteína-proteína2. O dedo anelar canônico é um tipo especial de domínio coordenador de zinco definido como uma seqüência de consenso de oito Cs conservados (C) e Seu (H) espaçados especificamente por outros resíduos de aminoácidos (X), C-X2-C-X9-39-C-X1-3-H-X2-3-C/H-X2-C-X 48-C-X2-C. Dois íons Zn2+ são estabilizados pelos resíduos c e h do núcleo através da topologia única "cross-brace" com C1/C2 e C/H5/C6 coordenando o primeiro íon Zn2+, enquanto C3/H4 e C7/C8 ligam o segundo (Figura 1A)3,4. Dependendo da presença de C ou H no quinto zn2+-local de coordenação, duas subclasses canônicas de proteínas ring-finger foram definidas: C3HC4 e C3H2C3 (RING-HC e RING-H2, respectivamente). Como o domínio RING da ligase ubiquitina E3 media a interação entre enzimas conjugantes E2 e substratos, a mutação desses resíduos essenciais de C e H tem sido mostrada para interromper a atividade da ligase5. Foram descritas mais cinco subclasses menos comuns de ligases RING E3 (RING-v, RING-C2, RING-D, RING-S/T e RING-G)6. As ligases de ubiquitina ring-tipo E3 podem ser ainda mais subdivididas em enzimas E3 simples e complexas. As ligases RING E3 da subunidade simples contêm o site de reconhecimento de substratos e o domínio RING de ligação e2. Em contrapartida, o complexo multisubunidade RING-tipo E3 recruta o substrato ou media a ligação do intermediário E2-ubiquitin ao complexo E3. O domínio RING Lys resíduos (s) que serve como um site de fixação de ubiquitina primário (s) para a auto-ubiquitinação também pode ser importante para a atividade da ligase E3.
Nem todas as proteínas contendo ANEL funcionam como ligases E3. Assim, a previsão bioinformática do domínio do dedo ANEL e a capacidade de ubiquitinação proteica dependente de E2 devem ser bioquimicamente validadas e ligadas ao papel biológico da proteína testada. Aqui, descrevemos um protocolo passo a passo descrevendo como detectar e caracterizar funcionalmente a atividade enzimática das ligases de ubiquitina ring-tipo E3, tanto in vitro quanto na planta, através de uma abordagem mutagênese dirigida pelo local. Os resultados representativos deste gasoduto são mostrados para o RING-tipo E3 ligase RHA1B. Rha1B é uma proteína efetora produzida pelo cisto parasita da planta nematode Globodera pallida para suprimir a imunidade vegetal e manipular a morfologia das células de raiz de plantas. Para se protegerem de patógenos/parasitárias, as plantas evoluíram receptores imunológicos do tipo de repetição de nucleotídeos e leucina (NB-LRR) que detectam a presença de um patógeno ou parasita e, como conseqüência, desenvolvem a resposta hipersensível (FC), que é uma forma de morte celular rápida e localizada ocorrendo no local da infecção para deter a colonização de patógenos. Um desses receptores imunológicos é a proteína Gpa2 de batata que confere resistência a alguns isolados de G. pallida (populações de campo D383 e D372)7.
Usando os protocolos apresentados, verificou-se recentemente que o RHA1B interfere com a sinalização imune às plantas de forma dependente da E3, visando o imunoreceptor gpa2 da planta para ubiquitinação e degradação8.
Protocolo
1. Mutagênese dirigida pelo local (Figura 1)
- Identifique os cis e seus aminoácidos conservados no domínio RING(Figura 1A)e primers de design carregando o codon de substituição de interesse ladeado por 15 pares de base em ambos os lados do local da mutação (Figura 1B).
- Introduzir a mutação desejada pela amplificação baseada em PCR do plasmídeo que abriga o gene do interesse usando primers mutagênicos e polimererase de DNA de alta fidelidade contendo Pfu em 50 μL do volume total de reação pcr como mostrado na Tabela 1 e Tabela 2 de acordo com o protocolo do fabricante.
- Dime o DNA metilado e semimetilado de Escherichia coli, derivado da escherichia,adicionando 3 μL de enzima de restrição DpnI diretamente à reação do PCR (passo 1.2) e incubando a 37 °C por 2 h.
NOTA: A metilação é uma modificação de proteína pós-transcricional que é adicionada ao plasmídeo produzido e isolado das bactérias. Novas cópias de plasmídeo gerada por PCR carecem de metilação, portanto, as novas cópias permanecerão intactas durante o tratamento com DpnI. - Purifique os plasmídeos mutagenizados usando um kit de extração de DNA comercial baseado na tecnologia de coluna de spin e elute o DNA com 50 μL de água.
- Transforme células dh5α E. coli quimicamente competentes com 0,5 μL do DNA de plasmídeo mutagenizado recuperado de acordo com o protocolo do fabricante. Em resumo, incubar células competentes com DNA no gelo por 30 min, em seguida, choque-calor-los para 20 s em 42 ° C, e coloque tubos novamente no gelo por 2 min. Células incubadas com 500 μL de mídia LB em 37 ° C para 1 h em 250 rpm e depois espalhá-los em pates seletivos.
- Verifique a mutação desejada por Sanger sequenciando os plasmídeos de DNA isolados de E. coli.
2. Purificação de proteína recombinante e ensaio de ubiquitização in vitro
- Clonar o tipo selvagem RING e genes ring mutantes de interesse no vetor pMAL-c2 (siga o protocolo do fabricante; Tabela 3) para fundir esses genes com a marca epitope MBP que permite a purificação de um passo usando resina de amillose. Introduzir as construções resultantes na cepa E. coli BL21, conforme descrito no passo 1.5.
- Crescer a cepa E. coli BL21 que abriga a construção desejada em 50 mL LB médio líquido em 37 ° C para 2-3 h até atingir a fase logarítica (OD600 de 0,4-0,6).
- Adicione o IPTG a uma concentração final de 0,1-1 mM para induzir a expressão de proteína recombinante de interesse marcada por MBP e incubar a cultura E. coli por 2-3 h a 28 °C. Coloque a cultura no gelo após a incubação.
NOTA: Executar etapas 2.4-2.13 no gelo para proteger as proteínas da degradação. - Para verificar a eficiência da indução, colete 1,5 mL de células induzidas, g-los a 13.000 x g por 2 min, remover o supernatant, e resuspender as células em 20 μL de 2x SDS-PAGE buffer de carregamento (24 mM Tris-HCl pH 6,8, 0,8% SDS, 10% (v/ v) glicerol, 4 mM DTT, 0,04% (w/ v) bromehenol azul).
- Ferva as amostras por 5 min e executá-los em um gel 10% SDS-PAGE. Para avaliar visualmente o acúmulo de proteína de fusão MBP (peso molecular da proteína de interesse + 42,5 kDa MBP), manchar o gel por 20 min agitando com tampão de coloração Coomassie (50% metanol, 10% ácido acético, 0,1% azul Coomassie) e destaining durante a noite com o amortecedor destaining (20% metanol, ácido acético de 10%).
- Colher as células e. coli restantes por centrífuga a 1.350 x g por 6 min, descartar o supernatant, e resuspender a pelota celular com 5 mL de buffer de coluna (20 mM Tris HCl, 200 mM NaCl, 1 mM EDTA, inibidor protease bacteriano).
NOTA: Este é um bom lugar para parar o protocolo durante a noite. As células congeladas podem ser armazenadas até 1 semana a -20 °C. - Quebrar as células De E. coli, colocando o tubo contendo as bactérias em um banho de água gelada e aplicando 10 ciclos de sonication: 10 ssonication em 30% amp seguido por 20 s quebras.
- Amostra de centrífuga a 13.000 x g a 4 °C por 10 min e salvar o supernatant (extrato bruto).
- Prepare 500 μL de resina de amillose em um tubo de 15 mL. Lave a resina adicionando 10 mL de buffer de coluna fria e centrífuga em 1.800 x g,4 °C por 5 min. Faça isso 2x.
- Adicione 5 mL de extrato de petróleo bruto para o tubo com a resina de amillose e incubar durante a noite em 4 °C.
- Centrífuga a 1.800 x g a 4 °C por 5 min e descartar o supernatant.
- Adicione 10 mL de buffer de coluna para a pelota de resina e incubar por 20 min. Em seguida, centrífuga em 1.800 x g a 4 °C para 5 min. Repita este passo 2x.
- Elute a proteína da fusão com 0.5 mL do amortecedor da coluna que contem o maltose de 10 mM incubando uma amostra para 2 h em 4 °C. Centrífuga a 1.800 x g a 4 °C por 5 min e coletar a proteína eluted. Repita este passo 2x.
- Diálise 1 mL da fração de proteína contra o PBS frio. Aliquot proteína em tubos de uso único (10-20 μL) para evitar o congelamento de descongelamento e armazenar a -80 °C até necessário.
- Medir a concentração de proteína usando o ensaio Bradford9.
- Configure a reação de ubiquitinação in vitro em um volume total de 30 μL, misturando 40 ng de E1 (por exemplo, AtUBA1), 100 ng de E2 (por exemplo, AtUBC8, SlUBC1/4/6/7/12/13/17/20/22/27/32), 1 μg MBP-RING tipo proteína, e 2 μg FLAG-Ub (ou HA-Ub) no buffer de ubiquitinação (50 mM Tris-HCl pH 7.5, 2 mM ATP, 5 mM MgCl2, 30 mM creatine fosfato, 50 μg/mL creatina fosfosfosse). Incubar a mistura a 30 °C por 2 h.
NOTA: Pré-fazer 20x ubiquitinação buffer e armazená-lo até 6 meses em -20 ° C em pequenos alíquos para um único uso. A fosfoquinase creatina perde facilmente sua atividade enzimática quando o amortecedor é descongelado e congelado repetidamente. - Encerre a reação misturando as amostras de 30 μL com 7,5 μL de buffer de carregamento 5x SDS-PAGE (60 mM Tris-HCl pH 6.8, 2% SDS, 25% (v/v) glicerol, 10 mM DTT, 0.1% (w/v) bromehenol azul) e ebulição por 5 min.
- Separe as proteínas com eletroforese de gel sds-poliacrilamida de 7,5% (SDS-PAGE), em seguida, transfira para a membrana PDVF e detecte ubiquitinação por borrão ocidental usando o anti-FLAG (ou anti-HA).
- Manchar a membrana PVDF com azul Coomassie para verificar o carregamento igual de proteína testada do tipo MBP-RING.
3. Expressão de proteína transitória mediada pela agrobacteriumem folhas de Nicotiana benthamiana e no ensaio de ubiquitinação da planta
- Raia apropriada Agrobacterium tumefaciens cepas carregando o gene de interesse epitráutopo -marcado (por exemplo, HA-RHA1B, HA-RHA1BC135S, HA-RHA1BK146R, HA-Ub) e vetor vazio como um controle sobre o LB médio contendo os antibióticos de seleção apropriados.
- Após 2 dias de crescimento a 28 °C, pegue colônias individuais e cultivá-las em lb líquido médio com os antibióticos adequados em 28 °C/250 rpm para outro 24 h.
- Transfira 100 μL de cultura agrobacteriana para 3 mL LB fresco com os antibióticos apropriados e incubar a cultura para um adicional de 4-6 h em 28 °C com rotação (250 rpm) para a fase de crescimento exponencial tardio.
- Despine as células agrobacterianas a 1.800 x g por 6 min, descarte o supernatant e resuspenda as células com 3 mL de tampão de lavagem (50 mM MES pH 5.6, 28 mM glicose, 2 mM NaH2PO4). Repita este passo 2x.
- Após a segunda lavagem, suspender as células no buffer de indução (50 mM MES pH 5.6, 28 mM glicose, 2 mM NaH2PO4, 200 μM acetosyringone, 37 mM NH4Cl, 5 mMgSO4.7H2O, 4 mM KCl, 18 μM FeSO4.7H2O, 2 mMCaCl 2Cl 2. Incubar as células com buffer de indução para um adicional de 10-12 h a 28 °C.
NOTA: Acetosyringone induz transferência de T-DNA. - Centrífugaas as células a 1.800 x g por 6 min, descartam o supernatant e resuspendem as células com 2 mL de buffer de infiltração (10 mM MES pH 5.5, 200 μM acetosyringone).
NOTA: Se aagrobactérias agregar após a incubação com buffer de indução, deixe as células agregadas afundar para o fundo do tubo, deixando-o no banco por alguns minutos, e transferir a suspensão agrobacterium claro para um novo tubo antes de prosseguir com a etapa 3.6. - Medir a concentração de bactérias usando o valor od600 (a densidade óptica na absorção de 600 nm). Ajuste os valores do OD600 aos desejados.
NOTA: Normalmente, um valor OD600 entre 0,2-0,4 funciona melhor para uma única expressão mancha agrobacteriana. Se uma combinação de diferentes cepas agrobacterianas for aplicada, o total de600 valores de OD de cepas agrobacterianas não deve exceder 1. - Agroinfiltrado de 4 semanas de idade N. bethamiana sai gentilmente picando-os com uma agulha, seguido por agrobacterium injetando à mão com uma seringa sem a agulha. Circule a área de folha infiltrada com o marcador (geralmente 1-2 cm de diâmetro).
- Recolher os tecidos de folha infiltrados 36 h pós-infiltração. Moer o tecido para um pó fino com nitrogênio líquido.
- Pó de tecido resuspendido com 300 μL de tampão de extração de proteína (50 mM Tris-HCl pH 7.5, 150 mM NaCl, 5 mM EDTA, 2 mM DTT, 10% glicerol, 1% polivinilpolipirrolido, 1 mM PMSF, planta protease inibidor cocktail) e centrífuga em 15,000 x g para 15 min em 4 °C.
- Transfira o supernatant para um novo tubo. Adicione 5x SDS-PAGE buffer de carregamento para uma concentração final de 1x e deixe ferver por 5 min.
- Separe as proteínas brutas em géis 10% SDS-PAGE, transfira para membranas PVDF e sonda com anti-HA para detectar na ubiquitinação da planta.
4. Estabelecer a ligação entre a atividade enzimática e a função na planta
NOTA: Por exemplo, rha1b promove a degradação da proteína resistente Gpa2 para suprimir a morte de células de RH. Esta etapa mostra como verificar se essas atividades virulentas do RHA1B são dependentes da E3.
- Raia apropriada Agrobacterium tumefaciens cepas carregando genes marcados de interesse (neste exemplo HA-RHA1B, HA-RHA1BC135S, HA-RHA1BK146R, myc-Gpa2, RBP1) e vetor vazio como um controle. Siga os passos 3.1-3.8 para a preparação agrobacterium e injeção em folhas de N. bethamiana.
- Para a degradação da proteína de substrato dependente da E3, siga os passos 3,9-3,12 e execute a mancha ocidental usando anticorpos apropriados para detectar o acúmulo de proteínas nas células vegetais (por exemplo, anti-HA e anti-MYC).
- Para a resposta hipersensível dependente da E3 (RH) mediando a inibição da morte celular, monitore as folhas agroinfiltradas para os sintomas de morte de células de RH 2-4 dias após a infiltração.
Resultados
Nesta seção, os resultados representativos são fornecidos para o protocolo utilizado para exame de uma única subunidade E3 ubiquitin ligase RHA1B que tem um domínio do tipo RING-H2 prosite previsto (132-176 aminoácidos)10. Como mostrado na Figura 1,a fim de obter uma proteína mutante deficiente em E3, pelo menos um dos oito Cs ou Hs conservados no domínio RING (Figura 1A)precisa ser mutagenizado ( Figura1B). Assim, como primeiro passo, duas versões mutantes do RHA1B, RHA1BC135S (uma substituição de Cys by Ser no cconservado 3 do domínio RING) e RHA1BK146R (uma substituição de Lys por Arg no único Lys presente no RHA1B) foram geradas. Embora a única subunidade e3 ligases mediar a transferência de ubiquitina da ubiquitina que abriga E2 para o substrato, em vez de interagir diretamente com a ubiquitina, a auto-ubiquitinação da E3 em Lys pode ser necessária para sua atividade enzimática máxima.
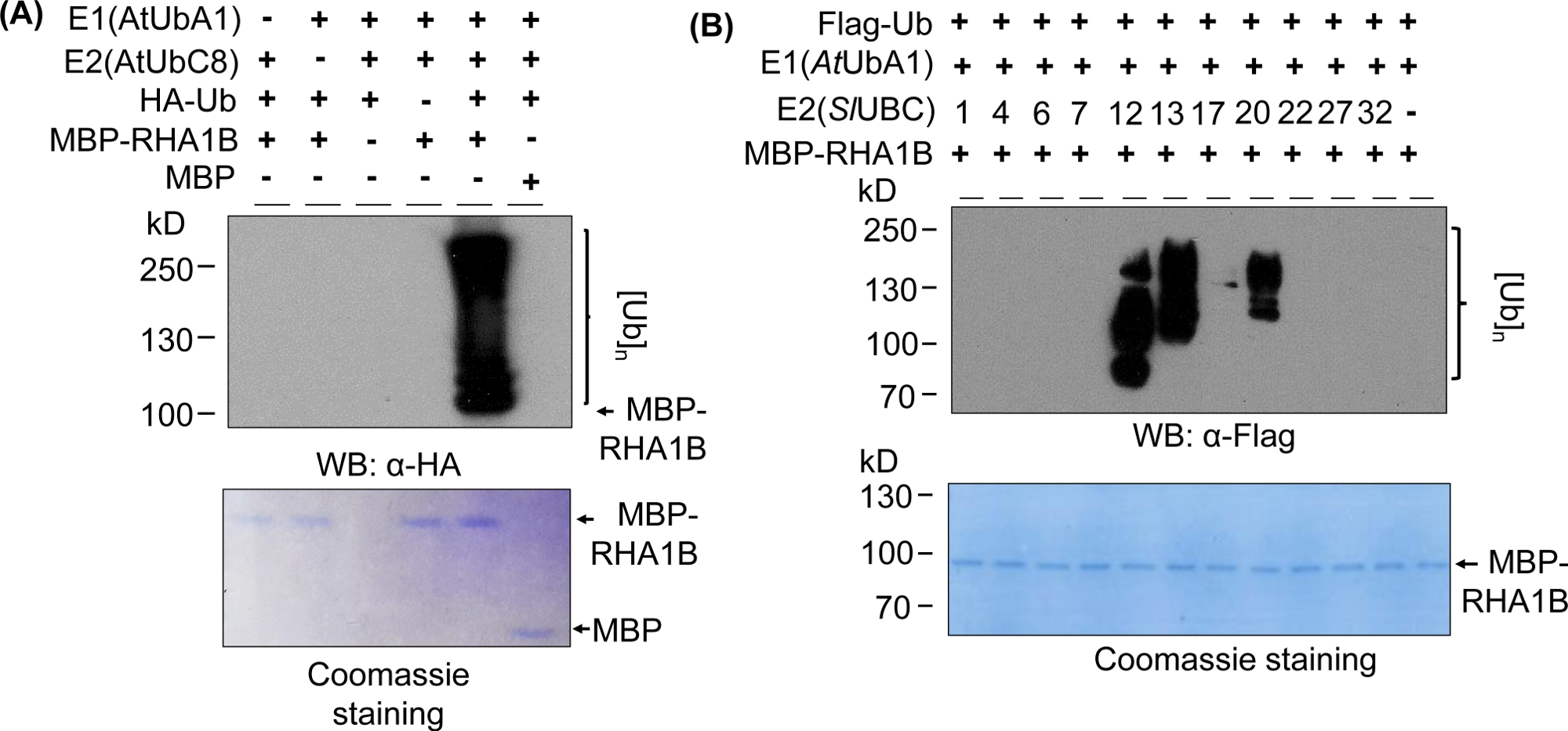
Os resultados de borrão ocidental na Figura 2A mostram um resultado típico positivo de ensaio de onipresença in vitro, com uma mancha multibanding começando com o peso molecular da proteína testada (por exemplo, RHA1B k1B k1B kDa com fusão de MPB) e progredindo para cima. O anticorpo anti-HA reconheceu ub marcado pela HA incorporado na cadeia de poli-onipresença de diferentes comprimentos, criando essa típica mancha de escada associada à ubiquitina. Para validar os resultados positivos, a Figura 2A também apresenta todos os controles negativos importantes que faltam componentes individuais (E1, E2, Ub ou MBP-RHA1B) ou usando o MBP como controle e sem o sinal de ubiquitização manchada. Além disso, a coloração azul Coomassie da membrana PVDF apresentou carregamento igual de MBP-RHA1B ou MBP em todos os controles.
A Figura 2B mostra como os resultados da ubiquitinação in vitro variaram dependendo da combinação específica de E2/E3. Neste exemplo, foram testados 11 E2s diferentes, representando 10 famílias E2 diferentes. A atividade detectada da ubiquitinação variou de nenhum sinal (nenhuma mancha) a uma mancha multibanding que começa em pesos moleculares diferentes, indicando testes padrões diferentes da ubiquitinação.
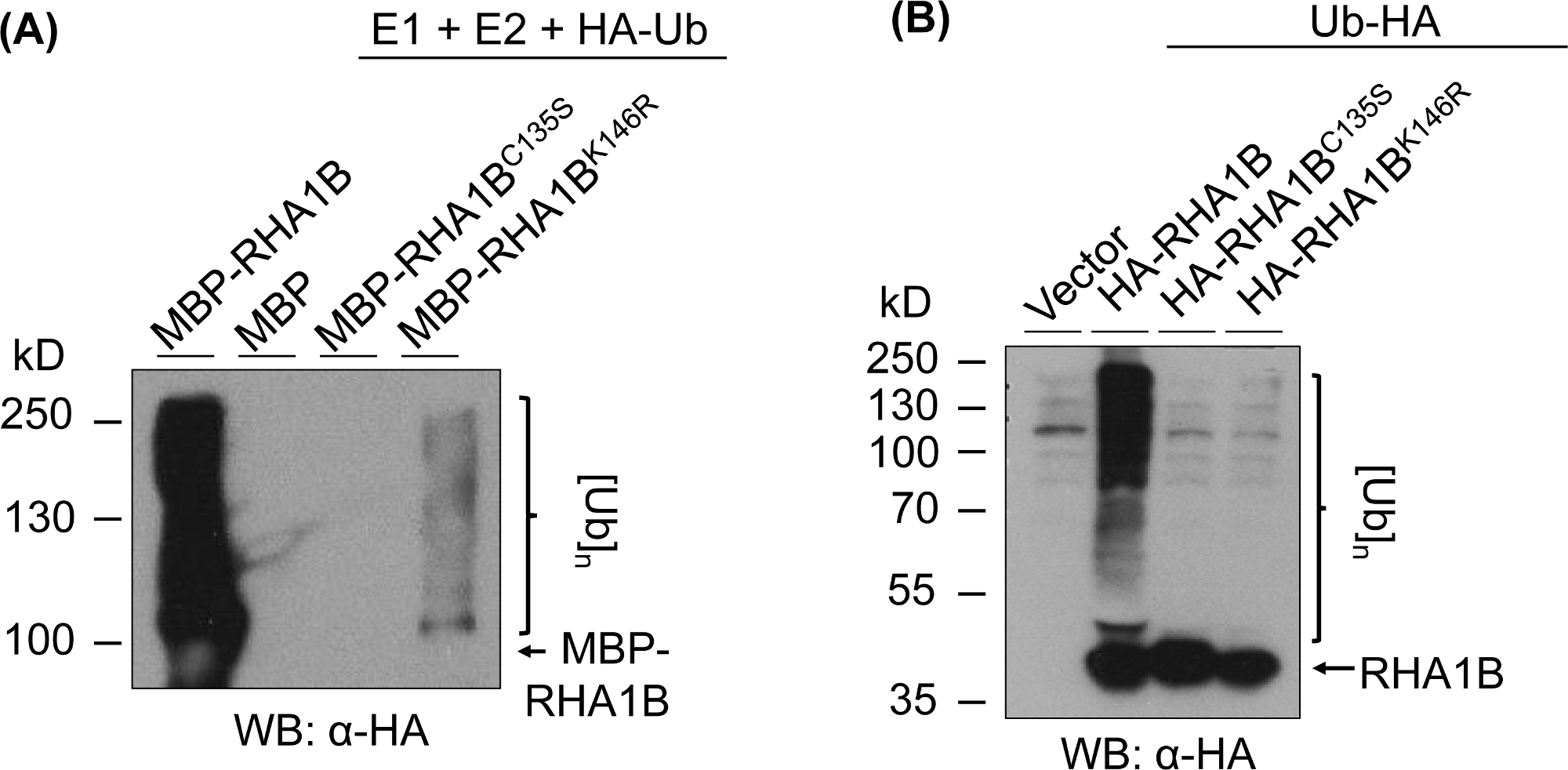
A Figura 3 mostra os resultados do ensaio de ubiquitinação para versões ring- e k-mutante de proteína testada. A falta de atividade enzimática para rha1bc135s é apoiada por sua incapacidade de gerar uma mancha multibanding in vitro (Figura 3A)ou promover o sinal de poli-ubiquitinação na planta (Figura 3B). É notável que a superexpressão do Ub marcado pela HA na planta por si só deu ubiquitinação de nível basal em todas as amostras testadas, incluindo o controle vetorial, em contraste com o forte sinal de ubiquitinação conferido pela atividade enzimática do tipo selvagem RHA1B. Além disso, a análise do mutante RHA1BK146R sugere que o resíduo K146 também é essencial para a atividade E3 do RHA1B. Embora um sinal marginal de auto-onipresença tenha sido detectado in vitro (Figura 3A),o ensaio na planta determinou que o mutante é deficiente em E3 (Figura 3B,apenas sinal de ubiquitinação de fundo detectado).
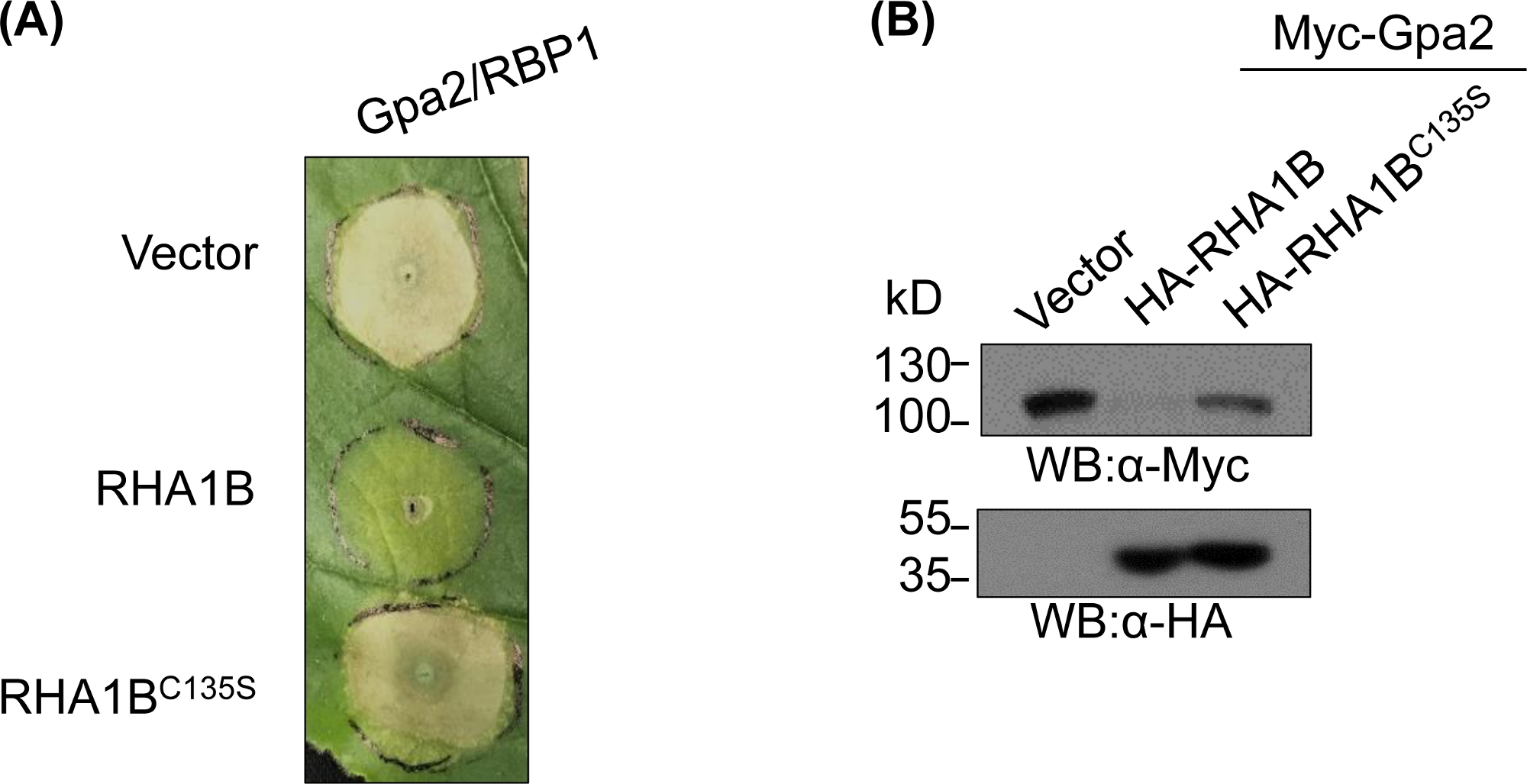
Depois de gerar e bioquimicamente validar o mutante deficiente em E3, estudos funcionais podem ser projetados para determinar o papel biológico associado à E3 da ligase ubiquitina RING testada. No caso do RHA1B, este efetor de nematóide suprime a sinalização imune da planta, como manifestado pela supressão da morte desencadeada por células de RH desencadeada pelo Gpa2. Conforme apresentado na Figura 4A,ao contrário do tipo selvagem RHA1B, o mutante RHA1BC135S sem atividade de ligase E3 não interferiu na morte de células de RH. Dado que o resultado mais comum da ubiquitinação proteica é a sua degradação mediada pela proteasome, mutações que residem no domínio RING também podem ser usadas para verificar uma capacidade dependente da E3 de desencadear degradação de seus substratos diretos e/ou indiretos. Assim, significativamente, os resultados de borrão ocidental na Figura 4B confirmam que a Gpa2 não se acumulou na presença do tipo selvagem RHA1B, mas o RHA1BC135S não teve impacto na estabilidade proteica gpa2.

Figura 1: Representação esquemática do princípio e passos envolvidos na mutagênese dirigida pelo local. (A)DOMÍNIO RING-CH/H2 com cis conservados e seus aminoácidos destacados. (B)Um exemplo de design de primers mutagênicos. (C)Passos de mutagênese dirigido pelo local. Clique aqui para ver uma versão maior deste número.
{kind=link}

Figura 2: Representante in vitro ubiquitinação ensaio. (A)O gel superior mostra o ensaio da ubiquitinação que inclui todos os controles negativos, e o gel inferior mostra o carregamento igual. (B) A gama de resultados esperados, dependendo das enzimas E2. Este número foi modificado a partir de Kud et al8. Clique aqui para ver uma versão maior deste número.
{kind=link}

Figura 3: Resultados do ensaio da Ubiquitination para RING- e K- mutantes (RHA1BC135S e RHA1BK146R). (A) Resultados de ubiquitização in vitro para RHA1BC135S e RHA1BK146R. (B) Em resultados de ensaio de ubiquitinação de planta para RHA1BC135S e RHA1BK146R. Este número foi modificado a partir de Kud et al8. Clique aqui para ver uma versão maior deste número.
{kind=link}

Figura 4: Estudo funcional representativo para funções biológicas dependentes de E3. Um exemplo de estudos funcionais mostrando a função biológica dependente de E3. (A)Supressão de morte de células de RH dependente de E3 e(B)degradação de um imunoreceptor de plantas Gpa2. Este número foi modificado a partir de Kud et al8. Clique aqui para ver uma versão maior deste número.
{kind=link}
| PCR configurado | |
| 1 μL | plasmid (~100 ng) plasmid (~100 ng) |
| 1,5 μL | Primer mutagênico F (10 μM) |
| 1,5 μL | R primer mutagênico (10 μM) |
| 1 μL | dNTPs (10 mM) |
| 5 μL 5 μL | buffer (10x) |
| 1 μL | Polímera Ultra Pfu (2,5 U/μl) |
| 39 μL | ddH 2 O DDH2O |
| 50 μL 50 μL | TOTAL VOLUME |
Tabela 1: Reação pcr configurada
| programa termociclor | |||
| 1 | 95 °C | 30 s | |
| 2 | 95 °C | 30 s | |
| 3 | 60 °C | 30 s | |
| 4 | 72 °C | 5 min | repita 2-4 30 vezes |
| 5 | 72 °C | 5 min | |
Tabela 2: Programa de termocicloparador PCR
| reação de ligadura criada para o exemplo RHA1B | ||
| 1,5 μL | pMAL-c2::MBP vetor linearizado por digestão com BamHI e SalI (60 ng) | |
| 7 μL | RHA1B/RHA1BC135S ou inserção RHA1BK146R digerida com BamHI e SalI (25 ng) | |
| 1 μL | Tampão da ligase T4 (10x) | |
| 0,5 μL 0,5 μL | T4 ligase (400 U/μL) | |
| 10 μL 10 μL | TOTAL VOLUME | |
Tabela 3: Reação de ligação criada para o exemplo RHA1B.
Discussão
Elucidar a base bioquímica e mecanicista das ligases de ubiquitina do tipo RING E3 pode contribuir muito para a nossa compreensão de seu significado biológico no desenvolvimento, sinalização de estresse e manutenção da homeostase. O protocolo descreveu aqui pares uma aproximação do mutagenesis com in vitro e em estudos funcionais da planta. Ao introduzir uma única substituição de aminoácidos nos resíduos conservados do domínio RING através da mutagênese direta do local, o mutante deficiente em E3 resultante pode ser testado em paralelo com proteína selvagem do tipo para ligar a atividade enzimática com a funcionalidade.
É fundamental identificar adequadamente o domínio RING, particularmente seus cis conservados e seus resíduos. Ferramentas on-line como prosite pode ser usado para fazê-lo10. Para desestabilizar o domínio RING responsável pelo recrutamento da enzima E2, cys é normalmente substituído por Ser, que é a sua substituição estrutural mais próxima sem a capacidade de criar uma ligação de dissulfeto usado para a coordenação de zinco. Lorick et al. mostraram que a mutação em qualquer um desses resíduos críticos de Ciso aboliria a atividade de ubiquitinação das ligaduras E3 do tipo RING única subunidade5. Embora alguns resíduos de Cys também sejam importantes para complexos multiunidades de ligase E3 contendo proteínas do tipo RING, devido à estrutura tridimensional multifacetada e dinâmica desses complexos de ubiquitinação e ao papel diferente das proteínas do tipo RING, substituições únicas de resíduos conservados no domínio RING na ligase multiunitária e3 não foram bem-sucedidas na geração de um fenótipo dedeficiente ligase11.
Para a mutagênese dirigida pelo local, descobrimos que o uso de vetores plasmídeos menores e ciclos de amplificação mais baixos geralmente produzia maior eficiência para mutagênese. A enzima Pfu pode ser substituída por qualquer outra polimererase de DNA de alta fidelidade e alta processabilidade. Além disso, se o gene do interesse contem codons raros, uma outra mancha de E. coli, Rosetta, pode ser usada para conseguir um rendimento mais elevado da proteína recombinante. Além disso, tanto o tempo de incubação quanto a temperatura para a indução do IPTG podem ser otimizados. Temperaturas mais baixas reduzem a taxa de divisão de E. coli, o que pode ser favorável à expressão de certas proteínas. Embora uma concentração mais elevada de IPTG poderia melhorar a expressão da proteína, igualmente inibe processos da divisão de E. coli e não é recomendado.
Ligases de anel único subunidade E3 não só funcionam como um andaime molecular que posiciona o intermediário E2-Ub nas proximidades do substrato, mas também estimula a atividade de transferência de ubiquitina de seus E2s cognatos. Além disso, dado que uma combinação E2/E3 é importante para o comprimento e as ligações da cadeia de poliubiquitina que determina o destino de um substrato modificado, qualquer consideração de Ring-tipo E3s deve incluir seus parceiros enzimáticos, E2s12. Como mostrado na Figura 3B, nem todos os E2s testados são compatíveis com a ligase RHA1B. Portanto, ensaios de ubiquitinação in vitro devem ser realizados em paralelo com várias enzimas E2 representando diferentes classes E2 para evitar resultados falsos negativos.
Apresentado aqui é o ensaio enzimático in vitro que detecta a capacidade de auto-ubiquitinação de proteínas testadas do tipo RING. No entanto, com pequenas modificações, este protocolo pode ser facilmente adaptado para detectar onipresença in vitro de substratos. Para este fim, a mistura de ubiquitinação in vitro do passo 2.15 deve ser complementada com a proteína recombinante do potencial substrato de ligase E3 (500 ng). Após uma incubação de 2 h a 30 °C, a proteína ubiquitinada deve ser capturada usando 15 μL de matriz de afinidade anti-HA (se ha-ub for usado, ou matriz de afinidade anti-FLAG se flag-ub é usado) por agitação por 2 h em 4 °C. Depois de lavar as contas 4x vezes com o tampão de lavagem Ub frio (20 mM Tris pH 7.5, 100 mM NaCl, 0.1 mM EDTA, 0.05% Tween 20, 1x PMSF), descartar todos, mas 40 μL do buffer e passar para o passo 2,16. O sinal de onipresença, detectado por anticorpos específicos do Ub e substrato marcados por epítopos, respectivamente, emergindo do peso molecular da proteína do substrato, confirma a especificidade do substrato/enzima.
Além disso, a identificação de substratos de ligase E3 in vivo é geralmente associada a múltiplos desafios devido à interação transitória de substrato de enzimas e à rápida degradação da proteína alvo ubiquitinada. Usar um mutante deficiente em ligase E3, que ainda interage com seu alvo, mas não mais o ubiquitina13,é uma alternativa muito útil para a adição do inibidor proteassômico MG132, que nem sempre interfere suficientemente com a função proteasome 26S.
Uma característica comum das ligaduras E3 do tipo RING é uma tendência a formar e funcionar como homo e/ou heterodimers. Curiosamente, a substituição nos resíduos conservados do domínio RING é geralmente associada a um fenótipo negativo dominante, onde mutado RING-tipo E3 ligase bloqueia a atividade enzimática de uma proteína do tipo selvagem nativo13. Assim, a superexpressão de mutantes RING na planta pode ser uma abordagem alternativa para derrubar o gene da ligase E3.
Divulgações
Os autores não têm nada a divulgar.
Agradecimentos
Nosso trabalho foi possível graças ao apoio financeiro da Bolsa Competitiva da Iniciativa de Agricultura e Pesquisa Alimentar (2017-67014-26197; 2017-67014-26591) do Instituto Nacional de Alimentação e Agricultura do USDA, USDA-NIFA Farm Bill, Northwest Potato Consórcio e Cultura Especializada da ISDA.
Materiais
| Name | Company | Catalog Number | Comments |
| Acetic acid | Sigma-Aldrich | A6283 | |
| Acetosyringone | Sigma-Aldrich | D134406 | |
| Amylose resin | NEB | E8021S | |
| ATP | Sigma-Aldrich | A1852 | |
| Bacterial protease inhibitor | Sigma-Aldrich | P8465 | |
| Bromphenol Blue | VWR | 97061-690 | |
| CaCl2 | Sigma-Aldrich | C1016 | |
| Centrifuge | Beckman Coulter | model: Avanti J-25 | |
| Commassie Blue | VWR | 97061-738 | |
| Creatine phosphate | Sigma-Aldrich | P7936 | |
| Creatine phosphokinase | Sigma-Aldrich | C3755 | |
| DNA clean & concentrator Kit | ZYMO RESEARCH | D4029 | |
| DpnI | NEB | R0176S | |
| DTT | Sigma-Aldrich | D0632 | |
| E. coli BL21 | Thermo Fisher Scientific | C600003 | |
| E. coli DH5α competent cells | Thermo Fisher Scientific | 18265017 | |
| EDTA | Sigma-Aldrich | 324504 | |
| FeSO4 7H2O | Sigma-Aldrich | F7002 | |
| FLAG-Ub | BostonBiochem | U-120 | |
| Glucose | VWR | 188 | |
| Glycerol | Sigma-Aldrich | G5516 | |
| HA-Ub | BostonBiochem | U-110 | |
| Heat block | VWR | model: 10153-318 | |
| Incubator | VWR | model: 1525 Digital Incubator | |
| Incubator shaker | Thermo Fisher Scientific | model: MaxQ 4000 | |
| IPTG | Roche | 10724815001 | |
| KCl | Sigma-Aldrich | P9333 | |
| LB Broth | Sigma-Aldrich | L3022 | |
| Liquide nitrogen | university chemistore | ||
| Maltose | Sigma-Aldrich | 63418 | |
| MES | Sigma-Aldrich | M3671 | |
| Methanol | Sigma-Aldrich | 34860 | |
| MgCl2 | Sigma-Aldrich | 63138 | |
| MgSO4 7H2O | Sigma-Aldrich | 63138 | |
| Microcentrifuge | Eppendorf | model: 5424 | |
| Miniprep plasmid purification kit | ZYMO RESEARCH | D4015 | |
| monoclonal anti-FLAG antibody | Sigma-Aldrich | F3165 | |
| monoclonal anti-HA antibody | Sigma-Aldrich | H9658 | |
| monoclonal anti-MYC antibody | Sigma-Aldrich | WH0004609M2 | |
| Mortar | VWR | 89038-144 | |
| NaCl | Sigma-Aldrich | S7653 | |
| NaH2PO4 | Sigma-Aldrich | S8282 | |
| NanoDrop | Thermo Fisher Scientific | model: 2000 Spectrophotometer | |
| Needle | Thermo Fisher Scientific | 14-826-5C | |
| NH4Cl | Sigma-Aldrich | A9434 | |
| PCR machine | Bio-Rad | model: C1000 | |
| Pestle | VWR | 89038-160 | |
| Pfu Ultra | Agilent Technologies | 600380 | |
| Plant protease inhibitor coctail | Sigma-Aldrich | P9599 | |
| pMAL-c2 | NEB | N8076S | |
| PMSF | Sigma-Aldrich | P7626 | |
| Polyvinylpolypyrrolidone | Sigma-Aldrich | P6755 | |
| SDS | Sigma-Aldrich | 1614363 | |
| Sonicator | Qsonica Sonicators | model: Q125 | |
| Syringe | Thermo Fisher Scientific | 22-253-260 | |
| Tris | Sigma-Aldrich | T1503 | |
| T4 ligase | NEB | M0202S |
Referências
- Freemont, P. S., Hanson, I. M., Trowsdale, J. A novel gysteine-rich sequence motif. Cell. 64, 483-484 (1991).
- Borden, K. L. B. RING fingers and B-boxes: Zinc-binding protein-protein interaction domains. Biochemistry and Cell Biology. 76, 351-358 (1998).
- Barlow, P. N., Luisi, B., Milner, A., Elliott, M., Everett, R. Structure of the C3HC4 Domain by 1H-nuclear Magnetic Resonance Spectroscopy: A New Structural Class of Zinc-finger. Journal of Molecular Biology. 237, 201-211 (1994).
- Borden, K. L. B., et al. The solution structure of the RING finger domain from the acute promyelocytic leukaemia proto-oncoprotein PML. The EMBO Journal. 14, 1532-1541 (1995).
- Lorick, K. L., et al. RING fingers mediate ubiquitin-conjugating enzyme (E2)-dependent ubiquitination. Proceedings of the National Academy of Sciences. 96, 11364-11369 (1999).
- Jiménez-López, D., Muñóz-Belman, F., González-Prieto, J. M., Aguilar-Hernández, V., Guzmán, P. Repertoire of plant RING E3 ubiquitin ligases revisited: New groups counting gene families and single genes. PLoS ONE. 13, 1-28 (2018).
- Sacco, M. A., et al. The Cyst Nematode SPRYSEC Protein RBP-1 Elicits Gpa2- and RanGAP2-Dependent Plant Cell Death. PLoS Pathogens. 5, 1-14 (2009).
- Kud, J., et al. The potato cyst nematode effector RHA1B is a ubiquitin ligase and uses two distinct mechanisms to suppress plant immune signaling. PLoS Pathogens. 15, 1007720 (2019).
- Bradford, M. M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry. 72, 248-254 (1976).
- Sigrist, C. J. A., et al. New and continuing developments at PROSITE. Nucleic Acids Research. 41, 344-347 (2013).
- Dove, K. K., Stieglitz, B., Duncan, E. D., Rittinger, K., Klevit, R. E. Molecular insights into RBR E3 ligase ubiquitin transfer mechanisms. EMBO Reports. 17, 1221-1235 (2016).
- Metzger, M. B., Pruneda, J. N., Klevit, R. E., Weissman, A. M. RING-type E3 ligases: Master manipulators of E2 ubiquitin-conjugating enzymes and ubiquitination. Biochimica et Biophysica Acta - Molecular Cell Research. 1843, 47-60 (2014).
- Xie, Q., et al. SINAT5 promotes ubiquitin-related degradation of NAC1 to attenuate auxin signals. Nature. 419, 167-170 (2002).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados