
Method Article
Caractérisation fonctionnelle des ligases d'ubiquitine RING-Type E3 In Vitro et In Planta
Dans cet article
Résumé
L'objectif de ce manuscrit est de présenter un aperçu des études biochimiques et fonctionnelles complètes des ligases d'ubiquitine E3 de type RING. Ce pipeline en plusieurs étapes, avec des protocoles détaillés, valide une activité enzymatique de la protéine testée et démontre comment lier l'activité à la fonction.
Résumé
L'ubiquitination, en tant que modification post-traductionnelle des protéines, joue un rôle réglementaire important dans l'homéostasie des cellules eucaryotes. L'attachement covalent de 76 modificateurs d'ubiquitine d'acide aminé à une protéine cible, selon la longueur et la topologie de la chaîne de polyubiquitine, peut avoir comme conséquence différents résultats s'étendant de la dégradation de protéine aux changements dans la localisation et/ou l'activité des protéine modifiées. Trois enzymes catalysent séquentiellement le processus d'ubiquitination : l'enzyme ubiquiitin-activant E1, l'enzyme e2 ubiquiitin-conjuguée, et la ligase d'ubiquitine d'E3. E3 ubiquitine ligase détermine la spécificité du substrat et, par conséquent, représente un sujet d'étude très intéressant. Ici, nous présentons une approche globale pour étudier la relation entre l'activité enzymatique et la fonction de la ligase d'ubiquitine E3 de type RING. Ce protocole en quatre étapes décrit 1) comment générer un mutant déficient e3 ligase par mutagénèse dirigée par site ciblée sur le domaine RING conservé; 2-3) comment examiner l'activité d'ubiquitination à la fois in vitro et dans le planta; 4) comment lier ces analyses biochimiques à la signification biologique de la protéine testée. La génération d'un mutant déficient e3 qui interagit encore avec son substrat mais ne l'ubiquite plus pour la dégradation facilite l'essai des interactions enzyme-substrat in vivo. En outre, la mutation dans le domaine RING conservé confère souvent un phénotype négatif dominant qui peut être utilisé dans les études fonctionnelles knock-out comme une approche alternative à une approche d'interférence de l'ARN. Nos méthodes ont été optimisées pour étudier le rôle biologique de l'effecteur de nématode parasite des plantes RHA1B, qui détourne le système d'ubiquitination hôte dans les cellules végétales pour promouvoir le parasitisme. Avec une légère modification du système d'expression in vivo, ce protocole peut être appliqué à l'analyse de toute ligase E3 de type RING, quelles que soient ses origines.
Introduction
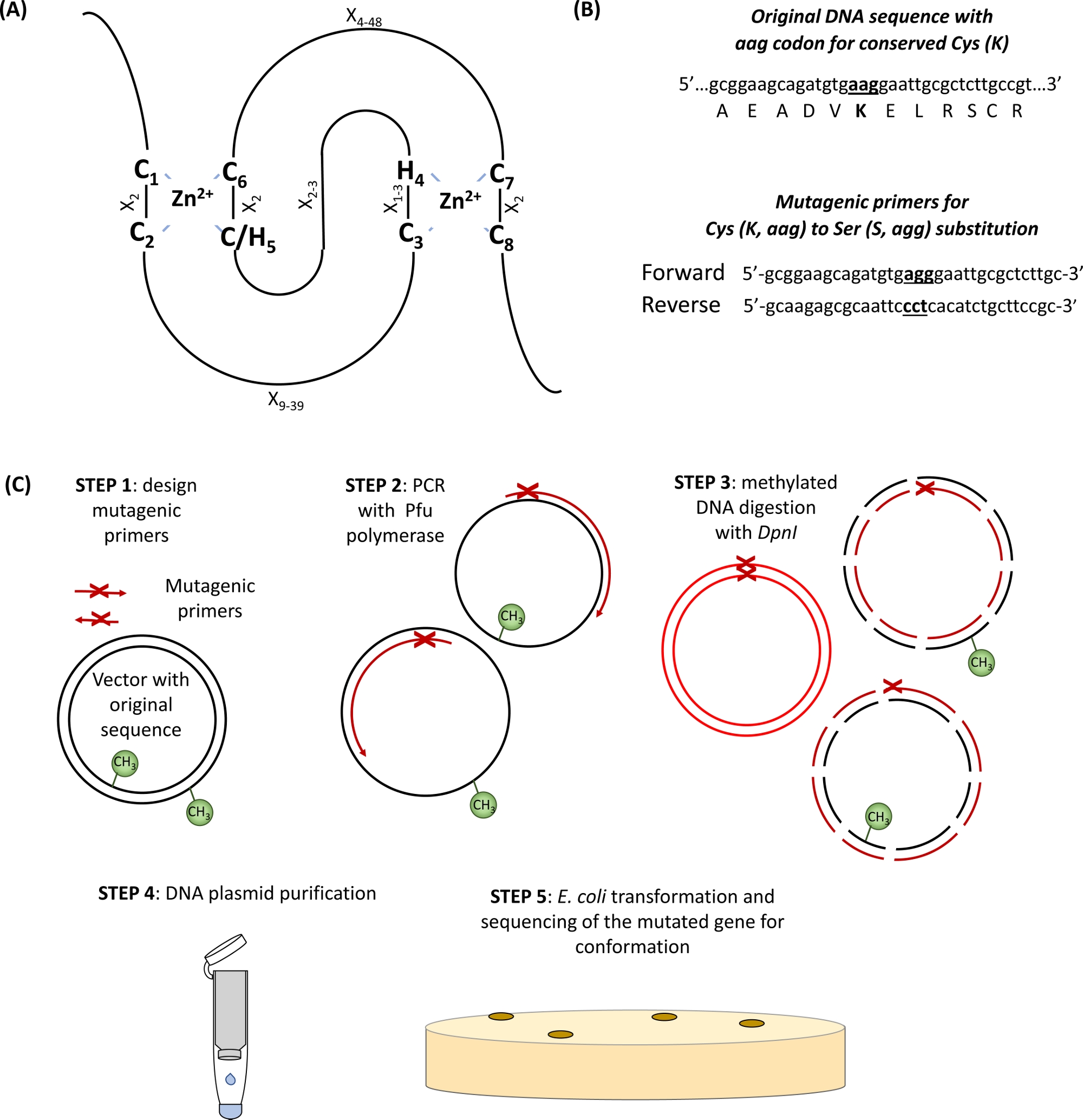
La grande majorité des ligases d'ubiquitine E3 appartiennent à RING (Really Interesting New Gene) -type protéines. Le domaine RING-doigt a été identifié à l'origine par Freemont et al. 1 et fonctionnellement décrit comme un domaine de médiation des protéines-protéines interaction2. Le doigt canonique RING est un type spécial de domaine de coordination du zinc défini comme une séquence consensuelle de huit Cys conservés (C) et Son (H) spécifiquement espacé par d'autres résidus d'acides aminés (X), C-X2-C-X9-39-C-X1-3-H-X2-3-C/H-X2-C-X4-48-C-X2-C. Deux ions Zn2 sont stabilisés par les résidus cœur C et H grâce à une topologie unique « cross-brace » avec C1/C2 et C/H5/C6 coordonnant le premier ion Zn2, tandis que C3/H4 et C7/C8 lient la seconde (Figure 1A)3,4. Selon la présence de C ou de H dans le cinquième site de coordination Zn2,deux sous-classes canoniques de protéines RING-doigt ont été définies : C3HC4 et C3H2C3 (RING-HC et RING-H2, respectivement). Parce que le domaine RING de l'E3 ubiquitine ligase médiateur de l'interaction entre les enzymes et les substrats conjugués E2, la mutation de ces résidus essentiels C et H a été montré pour perturber l'activité ligase5. Cinq autres sous-classes moins courantes de ligases RING E3 ont été décrites (RING-v, RING-C2, RING-D, RING-S/T et RING-G)6. Les ligases d'ubiquitine E3 de type RING peuvent être subdivisées en enzymes E3 simples et complexes. Les simples ligases RING E3 de sous-unité unique contiennent à la fois le site de reconnaissance du substrat et le domaine RING reliure E2. En revanche, le complexe E3 multisubunit de type RING est soit le substrat de la recrue, soit la liaison intermédiaire e2-ubiquitin au complexe E3. Le domaine RING Lys résidus (s) qui sert de site d'attachement ubiquitine primaire (s) pour l'auto-ubiquitination pourrait également être important pour l'activité de ligase E3.
Toutes les protéines contenant du RING ne fonctionnent pas comme des ligases E3. Ainsi, la prédiction bioinformatique du domaine RING-doigt et la capacité d'ubiquitination des protéines dépendantes de l'E2 doivent être validées biochimiquement et liées au rôle biologique de la protéine testée. Ici, nous décrivons un protocole étape par étape décrivant comment détecter et caractériser fonctionnellement l'activité enzymatique des ligases d'ubiquitine E3 de type RING, in vitro et dans le planta, par une approche mutagenesis dirigée par le site. Les résultats représentatifs de ce pipeline sont indiqués pour le RHA1B de ligase E3 de type RING. RHA1B est une protéine effectrice produite par le nématode de kyste parasite des plantes Globodera pallida pour supprimer l'immunité des plantes et manipuler la morphologie des cellules racinaires végétales. Pour se protéger contre l'invasion d'agents pathogènes/parasites, les plantes ont évolué dans le domaine liant les nucléotides et les récepteurs immunitaires de type répéter riche en leucine (NB-LRR) qui détectent la présence d'un agent pathogène ou d'un parasite et, par conséquent, développent la réponse hypersensible (HR), qui est une forme de mort cellulaire rapide et localisée sur le site de l'infection pour arrêter la colonisation des agents pathogènes. L'un de ces récepteurs immunitaires est la protéine Gpa2 de la pomme de terre qui confère une résistance à certains isolats de G. pallida (populations de champ D383 et D372)7.
En utilisant les protocoles présentés, il a été récemment constaté que RHA1B interfère avec la signalisation immunitaire des plantes d'une manière E3-dépendante en ciblant la plante Gpa2 immunorécepteur pour l'ubiquitination et la dégradation8.
Protocole
1. Mutagénèse dirigée par le site (Figure 1)
- Identifier les Cys et ses acides aminés conservés dans le domaine RING (Figure 1A) et concevoir des amorces portant le codon de substitution d'intérêt flanqué de 15 paires de base de chaque côté du site de mutation (Figure 1B).
- Introduire la mutation souhaitée par l'amplification à base de PCR du plasmide hébergeant le gène d'intérêt à l'aide d'amorces mutagènes et de polymérase d'ADN haute fidélité contenant du Pfu dans 50 'L du volume total de réaction PCR comme indiqué dans le tableau 1 et le tableau 2 selon le protocole du fabricant.
- Diger l'Escherichia coli-dérivéparent méthylé et semi-méthylé de l'ADN en ajoutant 3 L d'enzyme de restriction DpnI directement à la réaction PCR (étape 1.2) et l'incubation à 37 oC pendant 2 h.
REMARQUE : La méthylation est une modification de protéine posttranscriptional qui est ajoutée au plasmide produit et isolée des bactéries. Les nouvelles copies de plasmide PCR-généré s'absenter ont, par conséquent, les nouvelles copies resteront intactes pendant le traitement de DpnI. - Purifez les plasmides mutagérisés à l'aide d'un kit commercial d'extraction d'ADN basé sur la technologie des colonnes de spin et élichez l'ADN avec 50 l'eau.
- Transformez les cellules chimiquement compétentes de DH5MD E. coli avec 0,5 L de l'ADN plasmide mutagérisé récupéré selon le protocole du fabricant. En bref, incuber des cellules compétentes avec de l'ADN sur la glace pendant 30 min, puis les choquer à la chaleur pendant 20 s à 42 oC, et placer les tubes à nouveau sur la glace pendant 2 min. Incubate les cellules avec 500 l de bac LB à 37 oC pour 1 h à 250 tr/min, puis les étalez sur des pâtés sélectifs.
- Vérifier la mutation souhaitée par le séquençage de Sanger des plasmides d'ADN isolés de E. coli.
2. Purification recombinante des protéines et ubiquitination in vitro
- Cloner le type sauvage RING et les gènes RING mutés d'intérêt dans le vecteur pMAL-c2 (suivez le protocole du fabricant; Tableau 3) pour fusionner ces gènes avec l'étiquette d'épitope MBP qui permet une purification en une seule étape à l'aide de résine d'amylose. Introduire les constructions résultantes dans la souche E. coli BL21 décrite à l'étape 1.5.
- Cultivez la souche E. coli BL21 abritant la construction désirée dans un milieu liquide lb de 50 ml à 37 oC pendant 2 à 3 h jusqu'à ce qu'elle atteigne la phase logarithmique (OD600 de 0,4 à 0,6).
- Ajoutez iPTG à une concentration finale de 0,1 à 1 mM pour induire l'expression d'une protéine recombinante étiquetée MBP d'intérêt et incuber la culture E. coli pendant 2 à 3 h à 28 oC. Placer la culture sur la glace après l'incubation.
REMARQUE : Effectuez des étapes 2.4-2.13 sur la glace pour protéger les protéines contre la dégradation. - Pour vérifier l'efficacité de l'induction, recueillir 1,5 ml de cellules induites, les faire tourner vers le bas à 13 000 x g pendant 2 min, retirer le supernatant et resuspendre les cellules en 20 l de 2 x SDS-PAGE (24 mM Tris-HCl pH 6,8, 0,8 % SDS, 10 % (v/v) glycérol, 4 mM DTT, 0,04 % (w/v) bromopheno bluel).
- Faire bouillir les échantillons pendant 5 min et les faire fonctionner sur un gel SDS-PAGE de 10 %. Pour évaluer visuellement l'accumulation de protéines MBP-fusion (poids moléculaire de la protéine d'intérêt - 42,5 kDa MBP), tacher le gel pendant 20 min en agitant avec le tampon de coloration DeCoomassie (50% de méthanol, 10% d'acide acétique, 0,1% de bleu Coomassie) et en se desatteignant du jour au lendemain avec tampon de détacité (20% de méthanol, 10% d'acide acétique).
- Récoltez les cellules E. coli restantes par centrifugation à 1 350 x g pendant 6 min, jetez le supernatant et suspendez la pastille cellulaire avec 5 ml de tampon de colonne (20 mM Tris HCl, 200 mM NaCl, 1 mM EDTA, inhibiteur de la protéase bactérienne).
REMARQUE: C'est un bon endroit pour arrêter le protocole pendant la nuit. Les cellules congelées peuvent être stockées jusqu'à 1 semaine à -20 oC. - Décomposez les cellules E. coli en plaçant le tube contenant les bactéries dans un bain d'eau glacée et en appliquant 10 cycles de sonication : 10 s sonication à 30% d'ampli suivi de 20 s pauses.
- Échantillon de centrifugeuse à 13 000 x g à 4 oC pendant 10 min et enregistrer le supernatant (extrait brut).
- Préparer 500 l de résine d'amylose dans un tube de 15 ml. Laver la résine en ajoutant 10 ml de tampon de colonne froide et en centrifuge à 1 800 x g, 4 oC pendant 5 min. Faites-le 2x.
- Ajouter 5 ml d'extrait de brut dans le tube avec la résine d'amylose et incuber toute la nuit à 4 oC.
- Centrifugeuse à 1 800 x g à 4 oC pendant 5 min et jetez le supernatant.
- Ajouter 10 ml de tampon de colonne à la granule de résine et couver pendant 20 min. Puis centrifugeuse à 1 800 x g à 4 oC pendant 5 min. Répétez cette étape 2x.
- Éluter la protéine de fusion avec 0,5 ml de tampon de colonne contenant 10 mM de maltose en couve un échantillon pendant 2 h à 4 oC. Centrifugeuse à 1 800 x g à 4 oC pendant 5 min et recueillir la protéine élucée. Répétez cette étape 2x.
- Dialyser 1 ml de la fraction protéique contre le PBS froid. Protéine Aliquot en tubes à usage unique (10 à 20 l) pour éviter le gel-dégel et stocker à -80 oC jusqu'à ce que nécessaire.
- Mesurer la concentration protéique à l'aide de l'assayBradford 9.
- Configurez la réaction d'ubiquitination in vitro dans un volume total de 30 L en mélangeant 40 ng d'E1 (p. ex. AtUBA1), 100 ng d'E2 (p. ex., AtUBC8, SlUBC1/4/6/7/12/13/17/20/22/27/32), 1 g de protéine de type MBP,et 2 g FLAG-Ub (ou HA-Ub) dans le tampon d'ubiquitination (50 mM Tris-HCl pH 7,5, 2 mM ATP, 5 mM MgCl2, 30 mM de phosphate de créatine, 50 g/mL de phosphokinase créatine). Incuber le mélange à 30 oC pendant 2 h.
REMARQUE : Préfaites 20x tampon d'ubiquitination et stockez-le jusqu'à 6 mois à -20 oC dans de petits aliquots pour un usage unique. La créatine phosphokinase perd facilement son activité enzymatique lorsque le tampon est décongelé et congelé à plusieurs reprises. - Terminez la réaction en mélangeant les échantillons de 30 l avec 7,5 l de tampon de chargement SDS-PAGE 5x (60 mM Tris-HCl pH 6,8, 2 % SDS, 25 % (v/v) glycérol, 10 mM DTT, 0,1 % (w/v) bleu bromenol de phénophilol) et ébullition pendant 5 min.
- Séparez les protéines avec 7,5% D'électrophoresis de gel SDS-polyacrylamide (SDS-PAGE), puis transférez-les sur la membrane PDVF, et détectez l'ubiquitination par le ballonnement occidental à l'aide de l'anti-FLAG (ou anti-HA).
- Tainer la membrane PVDF avec le bleu Coomassie pour vérifier le chargement égal de la protéine testée de type MBP-RING.
3. Agrobacterium-expression de protéine transitoire médiée dans les feuilles de Nicotiana benthamiana et dans l'évaluation d'ubiquitination de planta
- Stries appropriées Agrobacterium tumefaciens souches portant le gène d'intérêt épitope-marqué (par exemple, HA-RHA1B, HA-RHA1BC135S, HA-RHA1BK146R, HA-Ub) et vecteur vide comme un contrôle sur le milieu LB contenant les antibiotiques de sélection appropriés.
- Après 2 jours de croissance à 28 oC, prenez des colonies simples et cultivez-les dans le milieu liquide LB avec les antibiotiques appropriés à 28 oC/250 tr/h pendant 24 h.
- Transférer 100 l de culture agrobactérienne à 3 ml de LB frais avec les antibiotiques appropriés et incuber la culture pendant 4 à 6 h supplémentaires à 28 oC avec rotation (250 tr/min) jusqu'à la phase de croissance exponentielle tardive.
- Faire tourner les cellules agrobactériennes à 1 800 x g pendant 6 min, jeter le supernatant et resuspendre les cellules avec 3 ml de tampon de lavage (50 mM MES pH 5,6, 28 mM de glucose, 2 mM NaH2PO4). Répétez cette étape 2x.
- Après le deuxième lavage, resuspendre les cellules dans le tampon d'induction (50 mM MES pH 5,6, 28 mM de glucose, 2 mM NaH2PO4, 200 M acetosyringone, 37 mM NH4Cl, 5 mM MgSO4.7H2O, 4 mM KCl, 18 MM FeSO4.7H2O, 2 mM CaCl2). Incuber les cellules avec un tampon d'induction pendant 10 à 12 h supplémentaires à 28 oC.
REMARQUE : L'acétosyringone induit le transfert de T-ADN. - Centrifuger les cellules à 1 800 x g pendant 6 min, jeter le supernatant et resuspendre les cellules avec 2 ml de tampon d'infiltration (10 mM MES pH 5,5, 200 m acetosyringone).
REMARQUE : Si les agrobactéries s'agrégent après l'incubation avec un tampon d'induction, laissez les cellules agrégées s'enfoncer au fond du tube en le laissant sur le banc pendant quelques minutes, et transférez la suspension Claire d'Agrobacterium dans un nouveau tube avant de procéder à l'étape 3.6. - Mesurer la concentration de bactéries à l'aide de la valeur OD600 (la densité optique à l'absorption de 600 nm). Ajustez les valeurs OD600 aux valeurs souhaitées.
REMARQUE : Habituellement, une valeur OD600 entre 0,2 et 0,4 fonctionne mieux pour une seule expression de tache agrobactérienne. Si une combinaison de différentes souches agrobactériennes est appliquée, les valeurs totales oD600 des souches agrobactériennes ne doivent pas dépasser 1. - Agroindin 4-week-old N. bethamiana leaves by gently pricking them with a needle, followed by hand-injecting Agrobacterium with a syringe without the needle. Encerclez la zone infiltrée de la feuille avec le marqueur (habituellement 1 à 2 cm de diamètre).
- Recueillir les tissus foliaires infiltrés 36 h après l'infiltration. Broyer le tissu en poudre fine avec de l'azote liquide.
- Poudre de tissu rechargée avec 300 l de tampon d'extraction de protéines (50 mM Tris-HCl pH 7.5, 150 mM NaCl, 5 mM EDTA, 2 mM DTT, 10% glycérol, 1% polyvinylpolypyrrolidone, 1 mM PMSF, cocktail inhibiteur de la protéase végétale) et centrifugeuse à 15 000 x g pendant 15 min à 4 oC.
- Transférer le supernatant dans un nouveau tube. Ajouter 5x tampon de chargement SDS-PAGE à une concentration finale de 1x et faire bouillir pendant 5 min.
- Séparer les protéines brutes sur les gels SDS-PAGE de 10 %, les transférer sur les membranes PVDF et sonder les anti-HA pour détecter l'ubiquitination planta.
4. Établir le lien entre l'activité enzymatique et la fonction dans les planta
REMARQUE : Par exemple, RHA1B favorise la dégradation de la protéine résistante Gpa2 pour supprimer la mort des cellules HR. Cette étape montre comment vérifier que ces activités virulentes de RHA1B sont e3-dépendants.
- Streak approprié Agrobacterium tumefaciens souches portant des gènes marqués d'intérêt (dans cet exemple HA-RHA1B, HA-RHA1BC135S, HA-RHA1BK146R, myc-Gpa2, RBP1) et vecteur vide comme un contrôle. Suivez les étapes 3.1-3.8 pour la préparation et l'injection d'Agrobacterium sur les feuilles de N. bethamiana.
- Pour la dégradation des protéines du substrat dépendante de l'E3, suivez les étapes 3.9-3.12 et effectuez le ballonnement occidental à l'aide d'anticorps appropriés pour détecter l'accumulation de protéines dans les cellules végétales (p. ex., anti-HA et anti-MYC).
- Pour l'inhibition de la mort cellulaire par médiation de la réponse hypersensible dépendante de l'E3 (HR), surveillez les feuilles infiltrées pour les symptômes de mort des cellules HR 2 à 4 jours après l'infiltration.
Résultats
Dans cette section, les résultats représentatifs sont fournis pour le protocole utilisé pour l'examen d'une seule sous-unité E3 ubiquitin ligase RHA1B qui a un domaine de type RING-H2 PROSITE-prévu (132-176 acides aminés)10. Comme le montre la figure 1, afin d'obtenir une protéine mutante déficiente e3, au moins l'un des huit C ou H conservés dans le domaine RING (Figure 1A) doit être mutagé (Figure 1B). Ainsi, dans un premier temps, deux versions mutantes de RHA1B, RHA1BC135S (une substitution de Cys par Ser dans le C3 conservé du domaine RING) et RHA1BK146R (une substitution de Lys par Arg dans le seul Lys présent dans RHA1B) ont été générées. Bien que les ligases E3 de subunit unique mediate ubiquitin transfert de l'ubiquitine hébergeant E2 au substrat plutôt que d'interagir directement avec l'ubiquiitin, l'auto-ubiquitination de l'E3 à Lys pourrait être nécessaire pour son activité enzymatique maximale.
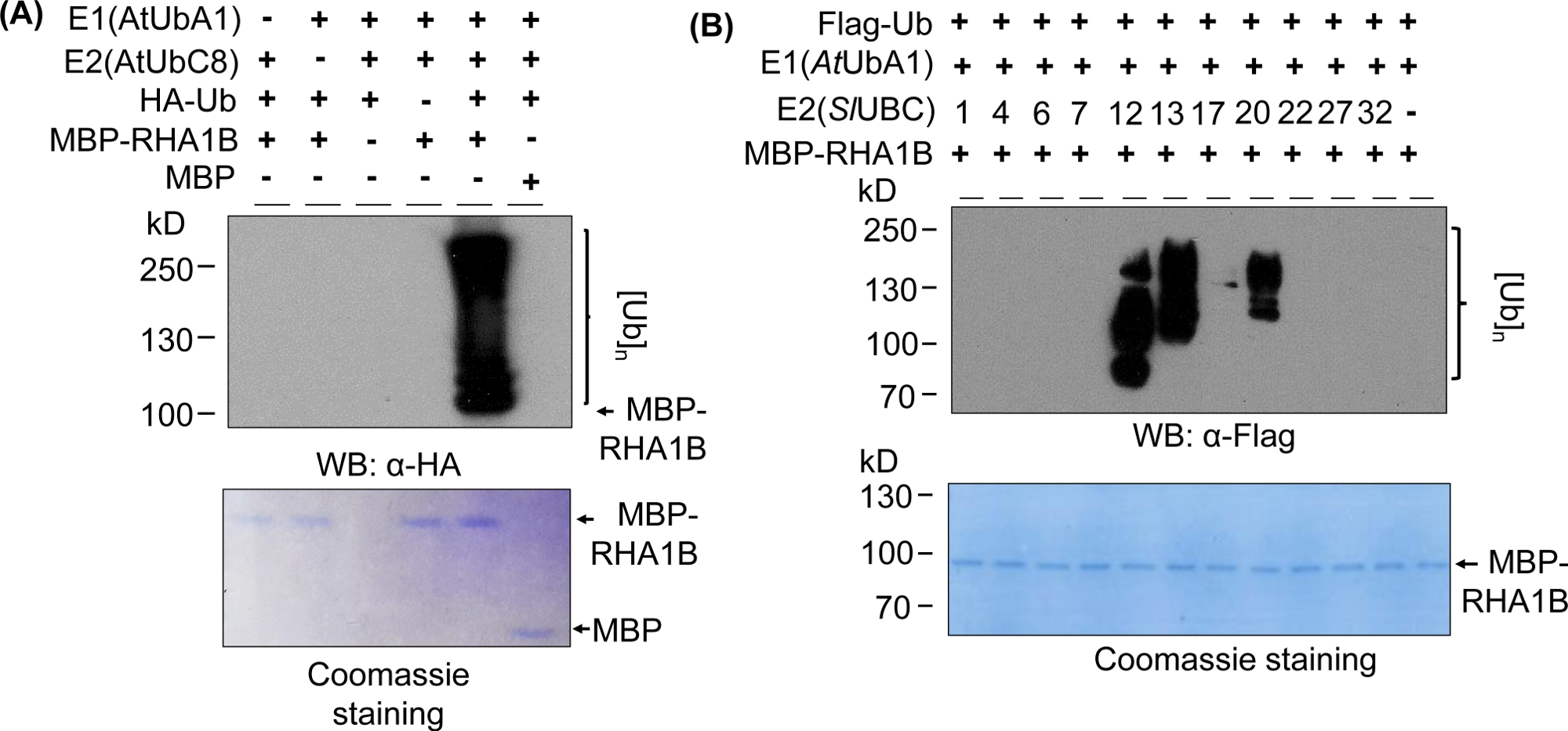
Les résultats de l'analyse de l'Ouest dans la figure 2A montrent un résultat typique positif d'analyse d'ubiquitination in vitro, avec un frottis multibande commençant au poids moléculaire de la protéine testée (par exemple, MPB-fused RHA1B '100 kDa) et progressant vers le haut. L'anticorps anti-HA a reconnu l'Ub étiqueté HA incorporé dans la chaîne de poly-ubiquitination de différentes longueurs, créant ce frottis typique ubiquitine-associé à l'échelle-comme. Pour valider les résultats positifs, la figure 2A présente également tous les contrôles négatifs importants manquant des composants individuels (E1, E2, Ub ou MBP-RHA1B) ou utilisant MBP comme contrôle et manquant du signal d'ubiquitination barbouillé. En outre, la coloration bleue de Coomassie de la membrane PVDF a montré la charge égale de MBP-RHA1B ou De MBP dans tous les contrôles.
La figure 2B montre comment les résultats de l'ubiquitination in vitro variaient selon la combinaison E2/E3 spécifique. Dans cet exemple, 11 E2 différents représentant 10 familles E2 différentes ont été testés. L'activité d'ubiquitination détectée s'est étendue de l'absence de signal (pas de frottis) à un frottis multibande à partir de différents poids moléculaires, ce qui indique différents modèles d'ubiquitination.
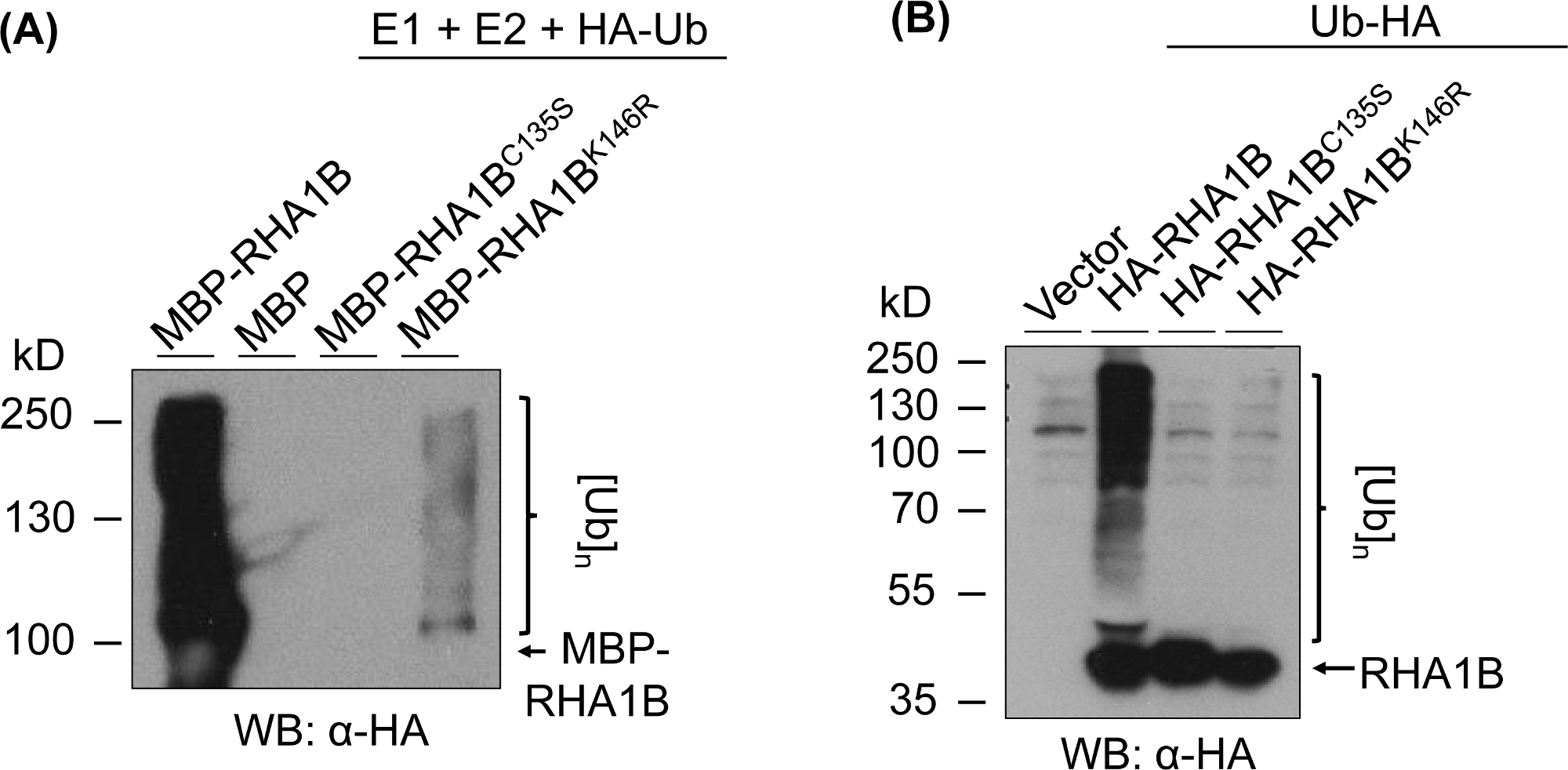
La figure 3 montre les résultats de l'analyse de l'ubiquitination pour les versions RING et K-mutant de protéines testées. Le manque d'activité enzymatique pour RHA1BC135S est étayé par son incapacité soit à générer un frottis multibandein (Figure 3A) ou à promouvoir le signal de poly-ubiquitination dans planta (Figure 3B). Il est à noter que la surexpression de l'Ub en planta étiqueté ha sur son propre a donné l'ubiquitination de niveau basal dans tous les échantillons testés, y compris la commande vectorielle, contrairement au signal d'ubiquitination fort conféré par l'activité enzymatique de type sauvage RHA1B. En outre, l'analyse sur le mutant RHA1BK146R suggère que le résidu K146 est également essentiel pour l'activité E3 de RHA1B. Bien qu'un signal marginal d'auto-ubiquitination ait été détecté in vitro (figure 3A), l'analyse in planta a déterminé que le mutant est E3-déficient(figure 3B, seul signal d'ubiquitination de fond détecté).
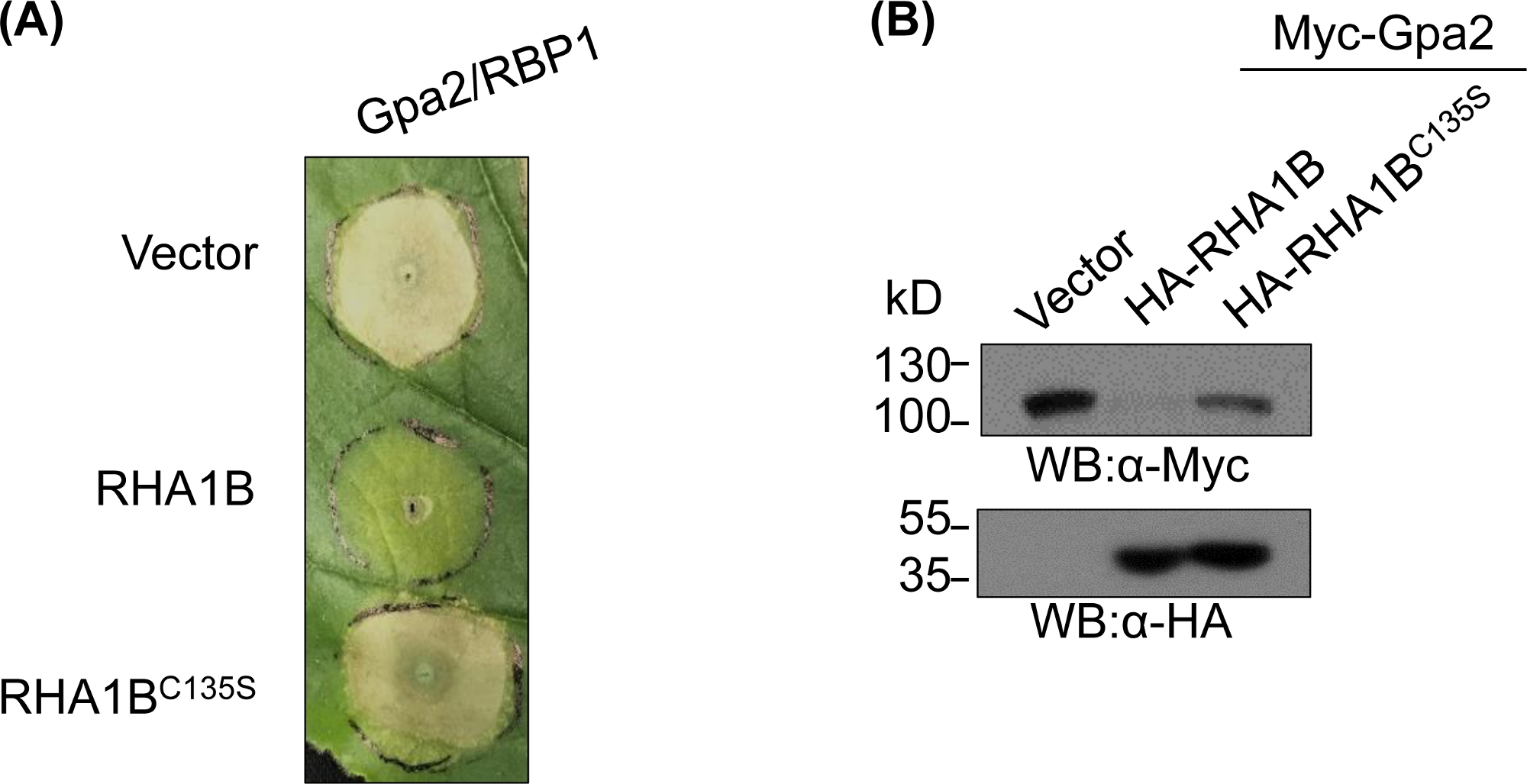
Après avoir généré et validé biochimiquement le mutant Déficient E3, des études fonctionnelles peuvent être conçues pour déterminer le rôle biologique e3 associé à la ligase d'ubiquitine RING E3 testée. Dans le cas de RHA1B, cet effecteur de nématode supprime la signalisation immunitaire de plante, comme manifesté par la suppression de la mort de cellule DE RH Gpa2-déclenchée. Comme présenté dans la figure 4A, contrairement au type sauvage RHA1B, le mutant RHA1BC135S manquant d'activité de ligase E3 n'a pas interféré avec la mort des cellules RH. Étant donné que le résultat le plus commun de l'ubiquitination des protéines est sa dégradation protéasome-négociée, les mutations résidant dans le domaine RING peuvent également être utilisées pour vérifier une capacité E3-dépendante de déclencher la dégradation de leurs substrats directs et/ou indirects. Ainsi, de façon significative, les résultats de l'ouest de la figure 4B confirment que le Gpa2 ne s'est pas accumulé en présence de type sauvage RHA1B, mais le RHA1BC135S n'a eu aucun impact sur la stabilité des protéines Gpa2.

Figure 1 : Représentation schématique du principe et des étapes de la mutagénèse dirigée par le site. (A) Domaine RING-CH/H2 avec Cys conservés et ses acides aminés mis en évidence. (B) Un exemple de conception d'amorces mutagènes. (C) Étapes de la mutagénèse dirigée par le site. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 2 : Exemple d'ubiquitination in vitro représentatif. (A) Le gel supérieur montre l'exemple d'ubiquitination, y compris tous les contrôles négatifs, et le gel inférieur montre une charge égale. (B) La gamme des résultats attendus en fonction des enzymes E2. Ce chiffre a été modifié à partir de Kud et al8. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 3 : Résultats d'analyse d'ubiquitination pour les mutants RING et K (RHA1BC135S et RHA1BK146R). (A) Résultats d'ubiquitination in vitro pour RHA1BC135S et RHA1BK146R. (B) Dans les résultats d'analyse d'ubiquitination planta pour RHA1BC135S et RHA1BK146R. Ce chiffre a été modifié à partir de Kud et al8. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 4 : Étude fonctionnelle représentative pour les fonctions biologiques dépendantes de l'E3. Un exemple d'études fonctionnelles montrant la fonction biologique E3-dépendante. (A) E3-dépendant suppression de la mort cellulaire HR et (B) dégradation d'une plante immunorécepteur Gpa2. Ce chiffre a été modifié à partir de Kud et al8. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
{kind=link}
| Mise en place de PCR | |
| 1 l | plasmide (100 ng) |
| 1,5 l | Apprêt mutagène F (10 M) |
| 1,5 l | Amorce mutagène R (10 M) |
| 1 l | dNTPs (10 mM) |
| 5 ll | tampon (10x) |
| 1 l | Polymère ultra-pfu (2,5 U/l) |
| 39 l | ddH2O |
| 50 l | TOTAL VOLUME |
Tableau 1 : Mise en place de la réaction PCR
| programme thermocycler | |||
| 1 | 95 oC | 30 s | |
| 2 | 95 oC | 30 s | |
| 3 | 60 oC | 30 s | |
| 4 | 72 oC | 5 min | répéter 2-4 30 fois |
| 5 | 72 oC | 5 min | |
Tableau 2 : Programme de thermocycler PCR
| réaction de ligature mise en place pour l'exemple RHA1B | ||
| 1,5 l | pMAL-c2::MBP vector linéaire par digestion avec BamHI et SalI (60 ng) | |
| 7 l | INSERT RHA1B/RHA1BC135S ou RHA1BK146R digéré avec BamHI et SalI (25 ng) | |
| 1 l | Tampon de ligase T4 (10x) | |
| 0,5 l | T4 ligase (400 U/L) | |
| 10 l | TOTAL VOLUME | |
Tableau 3 : Réaction de ligation mise en place pour l'exemple RHA1B.
Discussion
L'élucidation de la base biochimique et mécaniste des ligases d'ubiquitine de type E3 de RING peut contribuer considérablement à notre compréhension de leur signification biologique dans le développement, la signalisation de stress, et le maintien de l'homéostasie. Le protocole décrit ici couple une approche mutagénèse avec in vitro et dans les études fonctionnelles planta. En introduisant une substitution d'acide aminé unique dans les résidus conservés du domaine RING par mutagénèse directe du site, le mutant E3-déficient résultant peut être testé en parallèle avec la protéine de type sauvage pour lier l'activité enzymatique avec la fonctionnalité.
Il est essentiel d'identifier correctement le domaine RING, en particulier ses Cys conservés et ses résidus. Les outils en ligne tels que PROSITE peuvent être utilisés pour le faire10. Pour déstabiliser le domaine RING responsable du recrutement de l'enzyme E2, Cys est normalement remplacé par Ser, qui est son remplacement structurel le plus proche n'ayant pas la capacité de créer une liaison disulfure utilisée pour la coordination du zinc. Lorick et coll. ont montré que la mutation dans l'un de ces résidus critiques de Cys abolirait l'activité d'ubiquitination des ligases E3 de type RING de type ring de la sous-unité unique5. Bien que certains résidus de Cys soient également importants pour les complexes multiunit e3 ligase contenant des protéines de type RING, en raison de la structure tridimensionnelle multiforme et dynamique de ces complexes d'ubiquitination et du rôle différent des protéines de type RING, les substitutions uniques de résidus conservés dans le domaine RING en ligase E3 multiunitaire n'ont pas réussi à générer un phénotype déficient en ligase11.
Pour la mutagénèse dirigée par le site, nous avons constaté que l'utilisation de petits vecteurs plasmides et de cycles d'amplification plus faibles donnait généralement une plus grande efficacité pour la mutagénèse. L'enzyme Pfu peut être remplacée par n'importe quelle autre polymérase d'ADN haute fidélité et à haute teneur en processivité. En outre, si le gène d'intérêt contient des codons rares, une autre tache d'E. coli, Rosetta, peut être utilisée pour obtenir un rendement plus élevé de la protéine recombinante. En outre, le temps d'incubation et la température pour l'induction IPTG peuvent être encore optimisés. Des températures plus basses réduisent le taux de division d'E. coli, ce qui pourrait être favorable à l'expression de certaines protéines. Bien qu'une concentration plus élevée d'IPTG puisse améliorer l'expression des protéines, elle inhibe également les processus de division d'E. coli et n'est pas recommandée.
Les ligases E3 de type RING sous-unité unique fonctionnent non seulement comme un échafaudage moléculaire qui positionne l'intermédiaire E2-Ub à proximité du substrat, mais stimulent également l'activité de transfert d'ubiquitine de leurs E2 cognates. En outre, étant donné qu'une combinaison E2/E3 est importante pour la longueur et les liens de la chaîne de polyubiquitin qui détermine le sort d'un substrat modifié, toute considération de RING-type E3s doit inclure leurs partenaires enzymatiques, E2s12. Comme le montre la figure 3B, tous les E2 testés ne sont pas compatibles avec la ligase RHA1B. Par conséquent, les essais d'ubiquitination in vitro doivent être effectués en parallèle avec plusieurs enzymes E2 représentant différentes classes E2 afin d'éviter les faux résultats négatifs.
Présenté ici est l'essai enzymatique in vitro qui détecte la capacité d'auto-ubiquitination des protéines testées de type RING. Cependant, avec de petites modifications, ce protocole peut être facilement adapté pour détecter l'ubiquitination in vitro des substrats. À cette fin, le mélange d'ubiquitination in vitro de l'étape 2.15 devrait être complété avec la protéine recombinante du substrat potentiel de ligase E3 (500 ng). À la suite d'une incubation de 2 h à 30 oC, la protéine ubiquitinated doit être capturée à l'aide de 15 l de matrice d'affinité anti-HA (si HA-Ub est utilisé, ou matrice d'affinité anti-FLAG si FLAG-Ub est utilisé) par agitation pendant 2 h à 4 oC. Après avoir lavé les perles 4 fois avec le tampon de lavage Ub froid (20 mM Tris pH 7,5, 100 mM NaCl, 0,1 mM EDTA, 0,05% Tween 20, 1x PMSF), jeter tous sauf 40 l de la mémoire tampon et passer à l'étape 2,16. Le signal d'ubiquitination, détecté par des anticorps spécifiques à l'Ub et au substrat étiquetés épitopères, respectivement, émergeant du poids moléculaire de la protéine de substrat, confirme la spécificité du substrat/enzyme.
En outre, l'identification des substrats de ligase E3 in vivo est habituellement associée à de multiples défis dus à l'interaction transitoire enzyme-substrat et à la dégradation rapide de la protéine cible ubiquitinated. L'utilisation d'un mutant déficient E3 ligase, qui interagit toujours avec sa cible, mais ne l'ubiquitine plus13, est une alternative très utile à l'ajout de l'inhibiteur protéasomal MG132, qui n'interfère pas toujours suffisamment avec la fonction protéasome 26S.
Une caractéristique commune des ligases E3 de type RING est une tendance à se former et à fonctionner en tant qu'homo- et/ou hétérodistes. Fait intéressant, la substitution dans les résidus conservés du domaine RING est généralement associée à un phénotype négatif dominant où muté RING-type E3 ligase bloque l'activité enzymatique d'une protéine de type sauvage indigène13. Ainsi, la surexpression des mutants RING dans planta peut être une approche alternative à assommer le gène de ligase E3.
Déclarations de divulgation
Les auteurs n'ont rien à révéler.
Remerciements
Notre travail a été rendu possible grâce au soutien financier de la subvention concurrentielle de l'Initiative pour l'agriculture et la recherche alimentaire (2017-67014-26197; 2017-67014-26591) de l'INSTITUT national de l'alimentation et de l'agriculture de l'USDA, du projet de loi agricole de l'USDA-NIFA, de la pomme de terre du Nord-Ouest culture spécialisée ISDA.
matériels
| Name | Company | Catalog Number | Comments |
| Acetic acid | Sigma-Aldrich | A6283 | |
| Acetosyringone | Sigma-Aldrich | D134406 | |
| Amylose resin | NEB | E8021S | |
| ATP | Sigma-Aldrich | A1852 | |
| Bacterial protease inhibitor | Sigma-Aldrich | P8465 | |
| Bromphenol Blue | VWR | 97061-690 | |
| CaCl2 | Sigma-Aldrich | C1016 | |
| Centrifuge | Beckman Coulter | model: Avanti J-25 | |
| Commassie Blue | VWR | 97061-738 | |
| Creatine phosphate | Sigma-Aldrich | P7936 | |
| Creatine phosphokinase | Sigma-Aldrich | C3755 | |
| DNA clean & concentrator Kit | ZYMO RESEARCH | D4029 | |
| DpnI | NEB | R0176S | |
| DTT | Sigma-Aldrich | D0632 | |
| E. coli BL21 | Thermo Fisher Scientific | C600003 | |
| E. coli DH5α competent cells | Thermo Fisher Scientific | 18265017 | |
| EDTA | Sigma-Aldrich | 324504 | |
| FeSO4 7H2O | Sigma-Aldrich | F7002 | |
| FLAG-Ub | BostonBiochem | U-120 | |
| Glucose | VWR | 188 | |
| Glycerol | Sigma-Aldrich | G5516 | |
| HA-Ub | BostonBiochem | U-110 | |
| Heat block | VWR | model: 10153-318 | |
| Incubator | VWR | model: 1525 Digital Incubator | |
| Incubator shaker | Thermo Fisher Scientific | model: MaxQ 4000 | |
| IPTG | Roche | 10724815001 | |
| KCl | Sigma-Aldrich | P9333 | |
| LB Broth | Sigma-Aldrich | L3022 | |
| Liquide nitrogen | university chemistore | ||
| Maltose | Sigma-Aldrich | 63418 | |
| MES | Sigma-Aldrich | M3671 | |
| Methanol | Sigma-Aldrich | 34860 | |
| MgCl2 | Sigma-Aldrich | 63138 | |
| MgSO4 7H2O | Sigma-Aldrich | 63138 | |
| Microcentrifuge | Eppendorf | model: 5424 | |
| Miniprep plasmid purification kit | ZYMO RESEARCH | D4015 | |
| monoclonal anti-FLAG antibody | Sigma-Aldrich | F3165 | |
| monoclonal anti-HA antibody | Sigma-Aldrich | H9658 | |
| monoclonal anti-MYC antibody | Sigma-Aldrich | WH0004609M2 | |
| Mortar | VWR | 89038-144 | |
| NaCl | Sigma-Aldrich | S7653 | |
| NaH2PO4 | Sigma-Aldrich | S8282 | |
| NanoDrop | Thermo Fisher Scientific | model: 2000 Spectrophotometer | |
| Needle | Thermo Fisher Scientific | 14-826-5C | |
| NH4Cl | Sigma-Aldrich | A9434 | |
| PCR machine | Bio-Rad | model: C1000 | |
| Pestle | VWR | 89038-160 | |
| Pfu Ultra | Agilent Technologies | 600380 | |
| Plant protease inhibitor coctail | Sigma-Aldrich | P9599 | |
| pMAL-c2 | NEB | N8076S | |
| PMSF | Sigma-Aldrich | P7626 | |
| Polyvinylpolypyrrolidone | Sigma-Aldrich | P6755 | |
| SDS | Sigma-Aldrich | 1614363 | |
| Sonicator | Qsonica Sonicators | model: Q125 | |
| Syringe | Thermo Fisher Scientific | 22-253-260 | |
| Tris | Sigma-Aldrich | T1503 | |
| T4 ligase | NEB | M0202S |
Références
- Freemont, P. S., Hanson, I. M., Trowsdale, J. A novel gysteine-rich sequence motif. Cell. 64, 483-484 (1991).
- Borden, K. L. B. RING fingers and B-boxes: Zinc-binding protein-protein interaction domains. Biochemistry and Cell Biology. 76, 351-358 (1998).
- Barlow, P. N., Luisi, B., Milner, A., Elliott, M., Everett, R. Structure of the C3HC4 Domain by 1H-nuclear Magnetic Resonance Spectroscopy: A New Structural Class of Zinc-finger. Journal of Molecular Biology. 237, 201-211 (1994).
- Borden, K. L. B., et al. The solution structure of the RING finger domain from the acute promyelocytic leukaemia proto-oncoprotein PML. The EMBO Journal. 14, 1532-1541 (1995).
- Lorick, K. L., et al. RING fingers mediate ubiquitin-conjugating enzyme (E2)-dependent ubiquitination. Proceedings of the National Academy of Sciences. 96, 11364-11369 (1999).
- Jiménez-López, D., Muñóz-Belman, F., González-Prieto, J. M., Aguilar-Hernández, V., Guzmán, P. Repertoire of plant RING E3 ubiquitin ligases revisited: New groups counting gene families and single genes. PLoS ONE. 13, 1-28 (2018).
- Sacco, M. A., et al. The Cyst Nematode SPRYSEC Protein RBP-1 Elicits Gpa2- and RanGAP2-Dependent Plant Cell Death. PLoS Pathogens. 5, 1-14 (2009).
- Kud, J., et al. The potato cyst nematode effector RHA1B is a ubiquitin ligase and uses two distinct mechanisms to suppress plant immune signaling. PLoS Pathogens. 15, 1007720(2019).
- Bradford, M. M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry. 72, 248-254 (1976).
- Sigrist, C. J. A., et al. New and continuing developments at PROSITE. Nucleic Acids Research. 41, 344-347 (2013).
- Dove, K. K., Stieglitz, B., Duncan, E. D., Rittinger, K., Klevit, R. E. Molecular insights into RBR E3 ligase ubiquitin transfer mechanisms. EMBO Reports. 17, 1221-1235 (2016).
- Metzger, M. B., Pruneda, J. N., Klevit, R. E., Weissman, A. M. RING-type E3 ligases: Master manipulators of E2 ubiquitin-conjugating enzymes and ubiquitination. Biochimica et Biophysica Acta - Molecular Cell Research. 1843, 47-60 (2014).
- Xie, Q., et al. SINAT5 promotes ubiquitin-related degradation of NAC1 to attenuate auxin signals. Nature. 419, 167-170 (2002).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.