
Method Article
Funktionelle Charakterisierung von RING-Typ E3 Ubiquitin Ligases In Vitro und In Planta
In diesem Artikel
Zusammenfassung
Ziel dieses Manuskripts ist es, einen Überblick über die umfassenden biochemischen und funktionellen Studien der RING-Typ E3 Ubiquitin Ligasen zu präsentieren. Diese mehrstufige Pipeline mit detaillierten Protokollen validiert eine enzymatische Aktivität des getesteten Proteins und zeigt, wie die Aktivität mit der Funktion verknüpft werden kann.
Zusammenfassung
Die Ubiquitination als posttranslationale Modifikation von Proteinen spielt eine wichtige regulatorische Rolle bei der Homöostase eukaryotischer Zellen. Die kovalente Anhaftung von 76 Aminosäure-Ubiquitin-Modifikatoren an ein Zielprotein, abhängig von der Länge und Topologie der Polyubiquitin-Kette, kann zu unterschiedlichen Ergebnissen führen, die von Proteinabbau bis hin zu Veränderungen in der Lokalisierung und/oder Aktivität modifizierter Proteine reichen. Drei Enzyme katalysieren den Ubiquitinationsprozess sequenziell: E1-Ubiquitin-aktivierendes Enzym, E2-Ubiquitin-konjuzierendes Enzym und E3-Ubiquitinligase. E3 Ubiquitin Ligase bestimmt die Substratspezifität und stellt daher ein sehr interessantes Studienfach dar. Hier stellen wir einen umfassenden Ansatz vor, um die Beziehung zwischen der enzymatischen Aktivität und Funktion des RING-Typs E3 ubiquitin ligase zu untersuchen. Dieses vierstufige Protokoll beschreibt 1) wie ein E3-Ligase-Mangel-Mutant durch standortgesteuerte Mutagenese erzeugt wird, die auf die konservierte RING-Domäne ausgerichtet ist; 2–3) wie die Ubiquitationsaktivität sowohl in vitro als auch in planta untersucht werden kann; 4) wie diese biochemischen Analysen mit der biologischen Signifikanz des getesteten Proteins verknüpft werden können. Die Erzeugung eines E3-Ligase-mangelhaften Mutanten, der immer noch mit seinem Substrat interagiert, es aber nicht mehr zum Abbau ubiquitiniert, erleichtert die Prüfung von Enzym-Substrat-Wechselwirkungen in vivo. Darüber hinaus verleiht die Mutation in der konservierten RING-Domäne oft einen dominanten negativen Phänotyp, der in funktionellen Knockout-Studien als alternativer Ansatz für einen RNA-Interferenzansatz genutzt werden kann. Unsere Methoden wurden optimiert, um die biologische Rolle des pflanzenparasiten Nematodeneffektors RHA1B zu untersuchen, der das Wirts-Ubiquitinationssystem in Pflanzenzellen entführt, um Denparasitismus zu fördern. Mit geringfügiger Modifikation des In-vivo-Expressionssystems kann dieses Protokoll unabhängig von seiner Herkunft auf die Analyse jeder RING-Typ E3-Ligase angewendet werden.
Einleitung
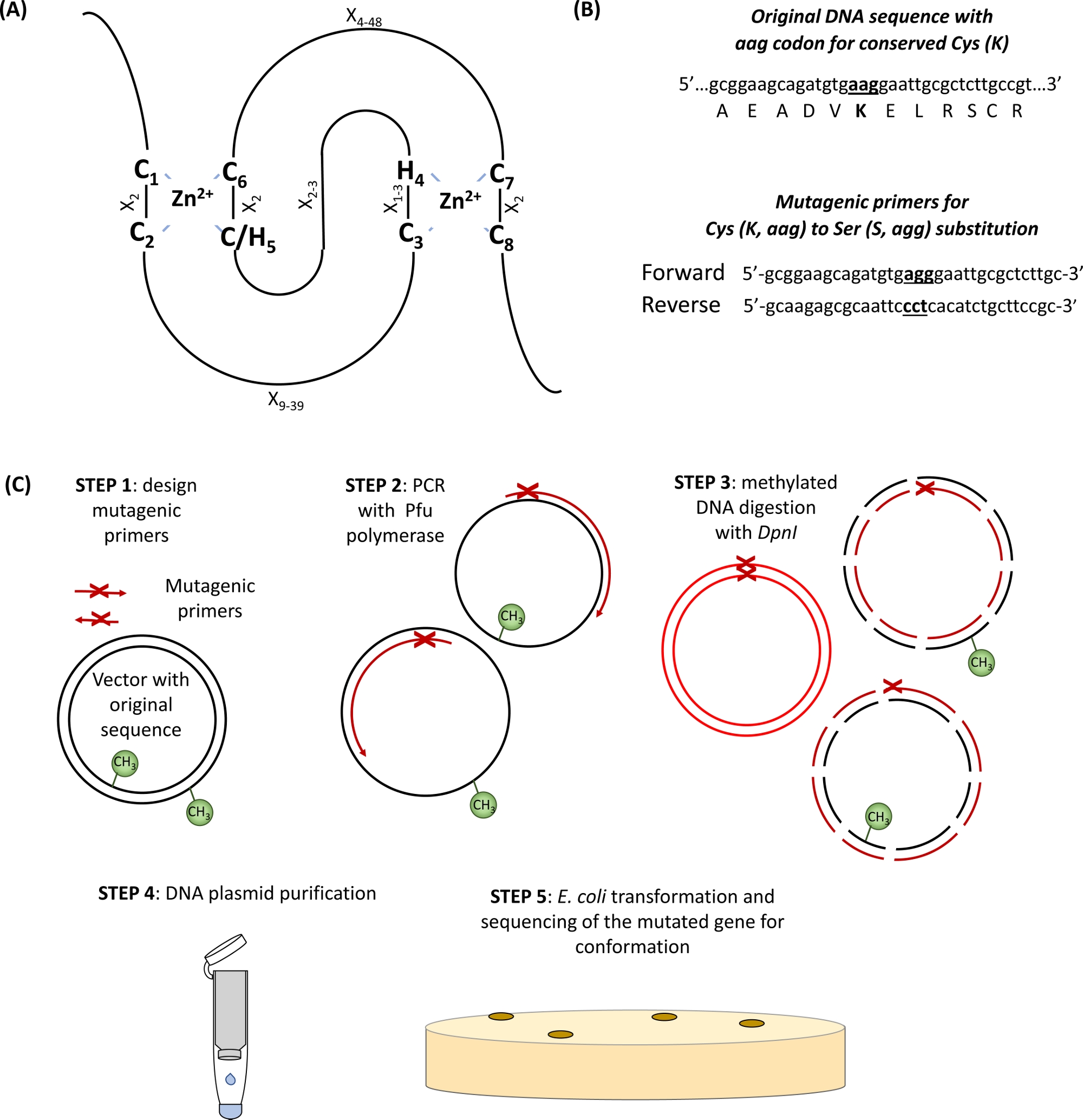
Die überwiegende Mehrheit der E3-Ubiquitinligasen gehört zu RING (Really Interesting New Gene)-Typ Proteine. Die RING-Finger-Domain wurde ursprünglich von Freemont et al.identifiziert. 1 und funktionell als Domäne beschrieben, die Protein-Protein-Interaktionvermittelt 2. Der kanonische RING-Finger ist eine spezielle Art von Zink-Koordinierungsdomäne, definiert als Konsenssequenz von acht konservierten Cys (C) und His (H), die speziell durch andere Aminosäurerückstände (X), C-X2-C-X9-39-C-X1–3-H-X2–3-C/H-X2-C-X4–48-C-X2-C- und -C- Zwei Zn2+-Ionen werden durch Kern-C- und H-Rückstände durch einzigartige "Cross-Brace"-Topologie mit C1/C2 und C/H5/C6 stabilisiert, die das erste Zn2+ Ionen koordinieren, während C3/H4 und C7/C8 die zweite binden (Abbildung 1A)3,4. Je nach Vorhandensein von C oder H in der fünften Zn2+-Koordinationsstelle wurden zwei kanonische Unterklassen von RING-Fingerproteinen definiert: C3HC4 und C3H2C3 (RING-HC bzw. RING-H2). Da die RING-Domäne von E3-Ubiquitinligase die Wechselwirkung zwischen E2-konjugierenden Enzymen und Substraten vermittelt, hat sich gezeigt, dass die Mutation dieser essentiellen C- und H-Rückstände die Ligaseaktivität stört5. Weitere fünf weniger häufige Unterklassen von RING E3-Ligasen wurden beschrieben (RING-v, RING-C2, RING-D, RING-S/T und RING-G)6. Die RING-Typ E3 Ubiquitin Ligasen können weiter in einfache und komplexe E3-Enzyme unterteilt werden. Die einfache Einzeleinheit RING E3 Ligases enthält sowohl die Substraterkennungsstelle als auch die E2-bindende RING-Domäne. Im Gegensatz dazu ist der Multisubunit RING-Typ E3-Komplex entweder das Substrat des Renousesubstrats oder vermittelt die Bindung des E2-Ubiquitin-Zwischenprodukts an den E3-Komplex. Die RING-Domäne Lys-Residuen, die als primäre Ubiquitin-Anhang-Site(n) zur Selbstallgegenwärtigkeit dienen, könnte auch für die E3-Ligase-Aktivität wichtig sein.
Nicht alle RING-haltigen Proteine funktionieren als E3-Ligasen. Daher müssen die bioinformatische Vorhersage der RING-Finger-Domäne und die Fähigkeit zur E2-abhängigen Protein-Ubiquitination biochemisch validiert und mit der biologischen Rolle des getesteten Proteins verknüpft werden. Hier beschreiben wir ein Schritt-für-Schritt-Protokoll, in dem beschrieben wird, wie die enzymatische Aktivität von RING-Typ E3-Ubiquitinligasen sowohl in vitro als auch in planta durch einen standortgesteuerten Mutagenese-Ansatz erkannt und funktional charakterisiert werden kann. Die repräsentativen Ergebnisse dieser Pipeline werden für den RING-Typ E3 ligase RHA1B angezeigt. RHA1B ist ein Effektorprotein, das von der pflanzlichen parasitären Zystennematode Globodera pallida produziert wird, um die Pflanzenimmunität zu unterdrücken und die Morphologie von Pflanzenwurzelzellen zu manipulieren. Um sich vor einer Invasion von Erregern/Parasiten zu schützen, haben Pflanzen Nukleotid-bindende Domäne und Leucin-reiche Wiederholung (NB-LRR) Typ Immunrezeptoren entwickelt, die das Vorhandensein eines Erregers oder Parasiten erkennen und als Folge die überempfindliche Reaktion (HR) entwickeln, die eine Form des schnellen und lokalisierten Zellsterbens ist, das an der Infektionsstelle auftritt, um die Besiedlung von Krankheitserregern zu stoppen. Ein solcher Immunrezeptor ist das Kartoffel-Gpa2-Protein, das resistenzen gegen einige Isolate von G. pallida (Feldpopulationen D383 und D372)7verleiht.
Mit den vorgestellten Protokollen, Es wurde vor kurzem festgestellt, dass RHA1B mit pflanzenimmunsignalisierung in einer E3-abhängigen Weise durch die Ausrichtung auf die Pflanze Gpa2 Immunrezeptor für Ubiquitination und Abbau8stört.
Protokoll
1. Standortgesteuerte Mutagenese (Abbildung 1)
- Identifizieren Sie die konservierten Cys und Seine Aminosäuren in der RING-Domäne (Abbildung 1A) und Entwerfenprimer, die das Substitutionscodon von Interesse tragen, flankiert von 15 Basenpaaren auf beiden Seiten der Mutationsstelle (Abbildung 1B).
- Einführung der gewünschten Mutation durch PCR-basierte Amplifikation des Plasmids, das das gen von Interesse enthält, mit mutagenen Primern und hoch-fidelity DNA-Polymerase, die Pfu enthalten, in 50 l des gesamten PCR-Reaktionsvolumens gemäß Tabelle 1 und Tabelle 2 gemäß dem Herstellerprotokoll einführen.
- Verdauen Sie die Escherichia coli-abgeleitete elterliche methylierte und halbmethylierte DNA, indem Sie 3 L dpnI-Restriktionsenzym direkt zur PCR-Reaktion (Schritt 1.2) und inkubieren bei 37 °C für 2 h hinzufügen.
HINWEIS: Methylierung ist eine posttranskriptielle Proteinmodifikation, die dem plasmid produziertund und von Bakterien isoliert wird. Neue Kopien von PCR-generierten Plasmid mangel Methylierung, daher werden die neuen Kopien während der DpnI-Behandlung intakt bleiben. - Reinigen Sie die mutagenisierten Plasmide mit einem kommerziellen DNA-Extraktionskit, das auf Spin-Säulen-Technologie basiert, und löschen Sie die DNA mit 50 l Wasser.
- Transformieren Sie dh5- E. coli chemisch kompetente Zellen mit 0,5 l der zurückgewonnenen mutagenisierten Plasmid-DNA gemäß dem Herstellerprotokoll. Kurz gesagt, inkubieren Sie kompetente Zellen mit DNA auf Eis für 30 min, dann Hitze-Schock sie für 20 s bei 42 °C, und legen Sie Röhren wieder auf Eis für 2 min. Inkubieren Zellen mit 500 l LB-Medien bei 37 °C für 1 h bei 250 U/min und dann auf selektiven Paten verteilen.
- Überprüfen Sie die gewünschte Mutation, indem Sanger die von E. coliisolierten DNA-Plasmide sequenziert.
2. Rekombinante Proteinreinigung und In-vitro-Ubiquitinationstest
- Klonen Sie den wilden Typ RING und mutierte RING-Gene von Interesse in den pMAL-c2-Vektor (folgen Sie dem Herstellerprotokoll; Tabelle 3) diese Gene mit dem MBP-Epitop-Tag zu verschmelzen, das eine einstufige Reinigung mit Amyloseharz ermöglicht. Führen Sie die resultierenden Konstrukte in den E. coli BL21-Stamm ein, wie in Schritt 1.5 beschrieben.
- Den E. coli-Stamm BL21 mit dem gewünschten Konstrukt in 50 ml LB flüssigem Medium bei 37 °C für 2–3 h anbauen, bis er die logarithmische Phase erreicht (OD600 von 0,4–0,6).
- Fügen Sie IPTG zu einer Endkonzentration von 0,1–1 mM hinzu, um die Expression von MBP-markiertem rekombinantem Protein zu induzieren und die E. coli-Kultur für 2–3 h bei 28 °C zu inkubieren. Legen Sie die Kultur nach der Inkubation auf Eis.
HINWEIS: Führen Sie die Schritte 2.4–2.13 auf Eis aus, um Proteine vor Dementierung zu schützen. - Um die Induktionseffizienz zu überprüfen, sammeln Sie 1,5 ml induzierter Zellen, Drehen Sie sie bei 13.000 x g für 2 min, entfernen Sie den Überstand, und setzen Sie die Zellen in 20 l 2x SDS-PAGE Ladepuffer (24 mM Tris-HCl pH 6,8, 0,8% SDS, 10% (v/v) Glycerin, 4 mM DTT, 0,04% (w/v) Bromphenol blau).
- Kochen Sie die Proben für 5 min und führen Sie sie auf einem 10% SDS-PAGE Gel. Um die Ansammlung von MBP-Fusionsprotein (Molekulargewicht des von Interesse + 42,5 kDa MBP) zu bewerten, färben Sie das Gel 20 min lang, indem Sie mit Coomassie-Färbungspuffer (50% Methanol, 10% Essigsäure, 0,1% Coomassie blau) und den Entsalzungspuffer (20% Methanol, 10% Essigsäure).
- Die restlichen E. coli-Zellen durch Zentrifugieren bei 1.350 x g für 6 min ernten, den Überstand entsorgen und das Zellpellet mit 5 ml Säulenpuffer (20 mM Tris HCl, 200 mM NaCl, 1 mM EDTA, bakteriellem Proteaseinhibitor) wieder aufsetzen.
HINWEIS: Dies ist ein guter Ort, um das Protokoll über Nacht zu stoppen. Die gefrorenen Zellen können bis zu 1 Woche bei -20 °C gelagert werden. - Brechen Sie E. coli-Zellen auf, indem Sie die Röhre, die die Bakterien enthält, in ein Eiswasserbad legen und 10 Beschallungszyklen anwenden: 10 s Beschallung bei 30% Ampere, gefolgt von 20 s Pausen.
- Zentrifugenprobe bei 13.000 x g bei 4 °C für 10 min und den Überstand (Rohextrakt) speichern.
- Bereiten Sie 500 l Amyloseharz in einem 15 ml-Rohr vor. Waschen Sie das Harz, indem Sie 10 ml Kaltsäulenpuffer und Zentrifugieren bei 1.800 x g,4 °C für 5 min hinzufügen. Tun Sie dies 2x.
- 5 ml Rohextrakt mit dem Amyloseharz in die Röhre geben und über Nacht bei 4 °C inkubieren.
- Zentrifugieren Sie bei 1.800 x g bei 4 °C für 5 min und entsorgen Sie den Überstand.
- 10 ml Säulenpuffer in das Harzpellet geben und 20 min brüten. Dann Zentrifuge bei 1.800 x g bei 4 °C für 5 min. Wiederholen Sie diesen Schritt 2x.
- Das Fusionsprotein mit 0,5 ml Säulenpuffer mit 10 mM Maltose durch Inkubation einer Probe für 2 h bei 4 °C elute. Zentrifugieren Sie bei 1.800 x g bei 4 °C für 5 min und sammeln Sie das eluierte Protein. Wiederholen Sie diesen Schritt 2x.
- Dialyse 1 ml der Proteinfraktion gegen die kalte PBS. Aliquot-Protein in Einweg-Röhren (10–20 l), um ein Einfrieren zu vermeiden und bei -80 °C zu lagern, bis es benötigt wird.
- Messen Sie die Proteinkonzentration mit dem Bradford-Assay9.
- Richten Sie die In-vitro-Ubiquitinationsreaktion in einem Gesamtvolumen von 30 l ein, indem Sie 40 ng E1 (z.B. AtUBA1), 100 ng E2 (z. B. AtUBC8, SlUBC1/4/6/7/12/13/17/20/22/27/32), 1 g MBP-RING-Protein, und 2 g FLAG-Ub (oder HA-Ub) im Ubiquitinationspuffer (50 mM Tris-HCl pH 7,5, 2 mM ATP, 5 mM MgCl2, 30 mM Kreatinphosphat, 50 g/ml Kreatinphosphokinase). Inkubieren Sie die Mischung bei 30 °C für 2 h.
HINWEIS: 20x Ubiquitinationspuffer vorbereiten und bis zu 6 Monate bei -20 °C in kleinen Aliquots für eine einzige Verwendung lagern. Die Kreatinphosphokinase verliert leicht ihre enzymatische Aktivität, wenn der Puffer wiederholt aufgetaut und eingefroren wird. - Beenden Sie die Reaktion, indem Sie die 30 L-Proben mit 7,5 l 5x SDS-PAGE-Ladepuffer (60 mM Tris-HCl pH 6,8, 2% SDS, 25% (v/v) Glycerin, 10 mM DTT, 0,1% (w/v) Bromphenolblau) und 5 min kochen.
- Trennen Sie die Proteine mit 7,5% SDS-Polyacrylamid-Gelelektrophorese (SDS-PAGE), übertragen Sie sie dann auf die PDVF-Membran und detektieren Sie die Ubiquitination durch Western Blotting mit der Anti-FLAG (oder Anti-HA).
- Färben Sie die PVDF-Membran mit Coomassie blue, um die gleiche Belastung des getesteten MBP-RING-Proteins zu überprüfen.
3. Agrobacterium-vermittelte transiente Proteinexpression in Nicotiana Benthamiana Blättern und in planta ubiquitination assay
- Streifen geeignete Agrobacterium tumefaciens Stämme, die das epitopgetaggte Gen von Interesse (z.B. HA-RHA1B, HA-RHA1BC135S, HA-RHA1BK146R, HA-Ub) und leeren Vektor als Kontrolle auf dem LB-Medium mit den entsprechenden Selektionsantibiotika tragen.
- Nach 2 Tagen Wachstum bei 28 °C einzelne Kolonien aufnehmen und in LB flüssigem Medium mit den entsprechenden Antibiotika bei 28 °C/250 Rpm für weitere 24 h anbauen.
- 100 l agrobakterielle Kultur mit den entsprechenden Antibiotika auf 3 ml frischeLB übertragen und die Kultur für weitere 4–6 h bei 28 °C mit Rotation (250 Rpm) in die späte exponentielle Wachstumsphase eintauchen.
- Spin-down agrobakterielle Zellen bei 1.800 x g für 6 min, entsorgen Sie den Überstand, und resuspendieren Zellen mit 3 ml Waschpuffer (50 mM MES pH 5,6, 28 mM Glukose, 2 mM NaH2PO4). Wiederholen Sie diesen Schritt 2x.
- Nach der zweiten Wäsche die Zellen im Induktionspuffer (50 mM MES pH 5,6, 28 mM Glukose, 2 mM NaH2PO4, 200 m Acetosyringon, 37 mM NH4Cl, 5 mM MgSO4.7H2O, 4 mM KCl, 18 'M FeSO4.7H2O, 2 mM CaCl2) wieder aufsetzen. Inkubieren Sie die Zellen mit Induktionspuffer für weitere 10–12 h bei 28 °C.
HINWEIS: Acetosyringon induziert T-DNA-Transfer. - Zentrifugieren Sie die Zellen bei 1.800 x g für 6 min, entsorgen Sie den Überstand und setzen Sie die Zellen mit 2 ml Infiltrationspuffer (10 mM MES pH 5,5, 200 M Acetosyringon) wieder aus.
HINWEIS: Wenn Agrobakterien nach der Inkubation mit Induktionspuffer aggregieren, lassen Sie die aggregierten Zellen auf den Boden des Rohres sinken, indem Sie es für ein paar Minuten auf der Bank lassen, und übertragen Sie die klare Agrobacterium Suspension in ein neues Rohr, bevor Sie mit Schritt 3.6 fortfahren. - Messen Sie die Konzentration von Bakterien mit dem OD600-Wert (die optische Dichte bei Absorption von 600 nm). Passen Sie die OD600-Werte an die gewünschten werte an.
HINWEIS: Normalerweise eignet sich ein OD600-Wert zwischen 0,2 und 0,4 am besten für eine einzelne agrobakterielle Fleckenexpression. Wenn eine Kombination verschiedener agrobakterieller Stämme angewendet wird, sollten die gesamten OD600-Werte von Agrobakterienstämmen 1 nicht überschreiten. - Agroinfiltrate 4 Wochen alte N. bethamiana Blätter durch sanftes Stechen mit einer Nadel, gefolgt von Hand injizieren Agrobacterium mit einer Spritze ohne die Nadel. Kreisen Sie den infiltrierten Blattbereich mit dem Marker (normalerweise 1–2 cm Durchmesser).
- Sammeln Sie die infiltrierten Blattgewebe 36 h nach der Infiltration. Schleifen Sie das Gewebe zu einem feinen Pulver mit flüssigem Stickstoff.
- Resuspendiertes Gewebepulver mit 300 l Proteinextraktionspuffer (50 mM Tris-HCl pH 7,5, 150 mM NaCl, 5 mM EDTA, 2 mM DTT, 10% Glycerin, 1% Polyvinylpyrrolidon, 1 mM PMSF, Pflanzenproteasehemmer-Cocktail) und Zentrifuge bei 15.000 x g für 15 min bei 4 °C.
- Übertragen Sie den Überstand auf eine neue Röhre. 5x SDS-PAGE Ladepuffer zu einer Endkonzentration von 1x hinzufügen und 5 min kochen.
- Trennen Sie Rohproteine auf 10% SDS-PAGE Gele, übertragen Sie auf PVDF-Membranen, und Sonde mit Anti-HA in Planta-Ubiquitination zu erkennen.
4. Herstellung der Verbindung zwischen enzymatischer Aktivität und Funktion in Planta
HINWEIS: RHA1B fördert beispielsweise den Abbau des resistenten Proteins Gpa2, um den Tod von HR-Zellen zu unterdrücken. In diesem Schritt wird gezeigt, wie Sie überprüfen können, ob diese virulenten Aktivitäten von RHA1B E3-abhängig sind.
- Streifen geeignete Agrobacterium tumefaciens Stämme tragen getaggte Gene von Interesse (in diesem Beispiel HA-RHA1B, HA-RHA1BC135S, HA-RHA1BK146R, myc-Gpa2, RBP1) und leeren Vektor als Steuerung. Befolgen Sie die Schritte 3.1–3.8 für die Agrobacterium-Präparation und Injektion auf N. bethamiana Blätter.
- Für den E3-abhängigen Substratproteinabbau folgen Sie den Schritten 3.9–3.12 und führen Sie die Western-Blotting mit geeigneten Antikörpern durch, um die Proteinakkumulation in Pflanzenzellen (z. B. Anti-HA und Anti-MYC) zu detektieren.
- Für die E3-abhängige überempfindliche Reaktion (HR) vermittelte Zelltodhemmung, überwachen Sie die agroinfiltrierten Blätter auf HR-Zell-Todessymptome 2–4 Tage nach der Infiltration.
Ergebnisse
In diesem Abschnitt werden repräsentative Ergebnisse für das Protokoll zur Verfügung gestellt, das für die Untersuchung einer einzelnen Untereinheit E3 Ubiquitin ligase RHA1B verwendet wird, die eine PROSITE-vorhergesagte RING-H2-Domäne (132–176 Aminosäuren)10hat. Wie in Abbildung 1dargestellt, muss mindestens eines der acht konservierten Cs oder Hs in der RING-Domäne(Abbildung 1A) mutagenisiert werden (Abbildung 1B). So wurden in einem ersten Schritt zwei mutierte Versionen von RHA1B, RHA1BC135S (eine Substitution von Cys durch Ser in der konservierten C3 der RING-Domäne) und RHA1BK146R (eine Substitution von Lys durch Arg in der einzigen lys in RHA1B) erzeugt. Obwohl einzelne Untereinheit E3-Ligisten den Ubiquitin-Transfer von Ubiquitin, das E2 beherbergt, auf das Substrat vermitteln, anstatt direkt mit Ubiquitin zu interagieren, könnte eine Selbst-Ubiquitination von E3 bei Lys für seine maximale enzymatische Aktivität erforderlich sein.
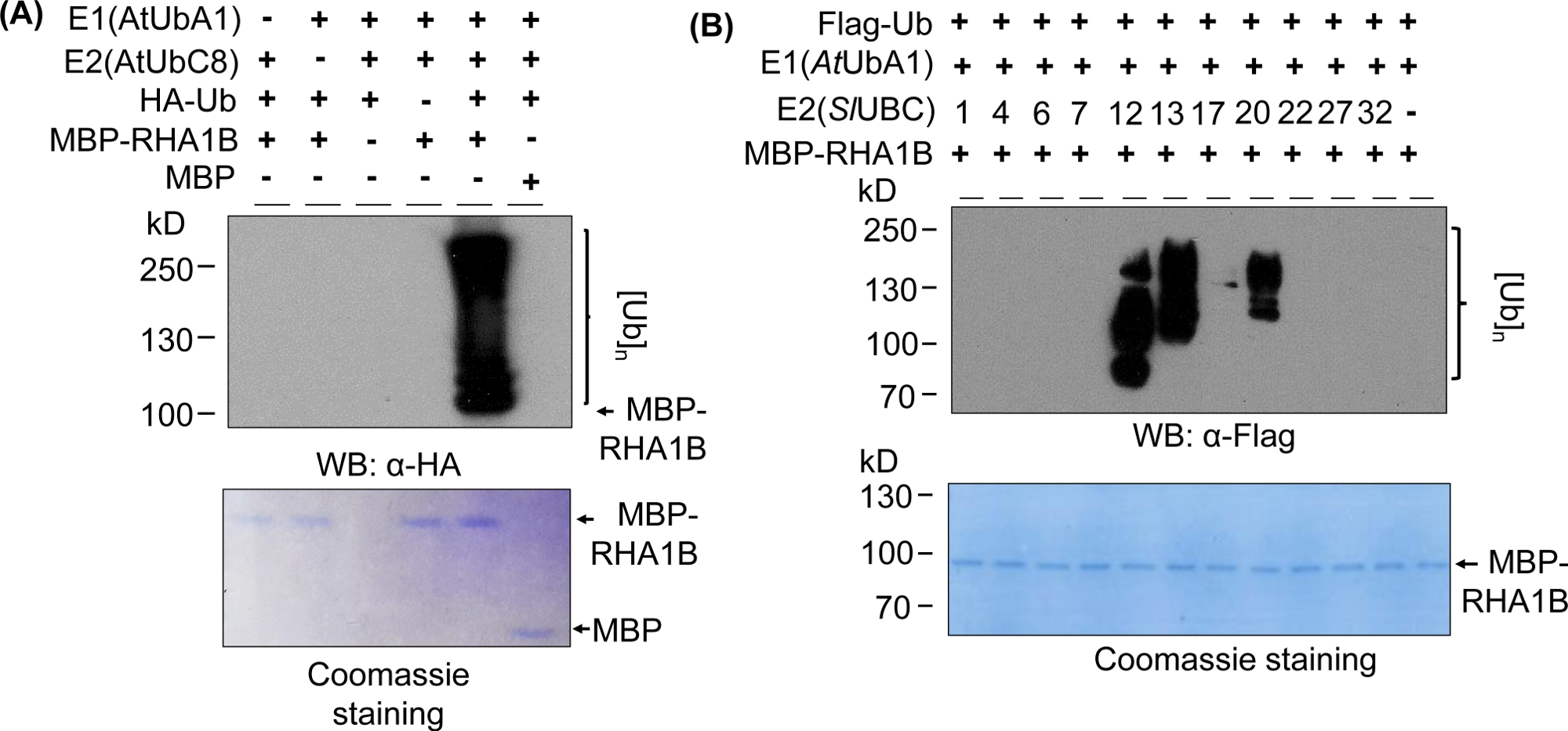
Die Western Blotting-Ergebnisse in Abbildung 2A zeigen ein typisches positives In-vitro-Ubiquitinations-Assay-Ergebnis, mit einem Multibanding-Abstrich, der beim Molekulargewicht des getesteten Proteins beginnt (z. B. MPB-fused RHA1B -100 kDa) und nach oben voranschreitet. Der Anti-HA-Antikörper erkannte HA-markierte Ub in die Poly-Ubiquitinationskette unterschiedlicher Längen integriert, wodurch dieser typische Ubiquitin-assoziierte Leiter-ähnliche Abstrich entsteht. Um die positiven Ergebnisse zu validieren, zeigt Abbildung 2A auch alle wichtigen Negativkontrollen, die einzelne Komponenten (E1, E2, Ub oder MBP-RHA1B) oder MBP als Steuerung und fehlendes verschmiertes Ubiquitinationssignal vermissen. Darüber hinaus zeigte die Coomassie-Blaufärbung der PVDF-Membran in allen Steuerungen die gleiche Belastung von MBP-RHA1B oder MBP.
Abbildung 2B zeigt, wie die Ergebnisse der In-vitro-Ubiquitination je nach spezifischer E2/E3-Kombination variierten. In diesem Beispiel wurden 11 verschiedene E2 getestet, die 10 verschiedene E2-Familien darstellen. Die nachgewiesene Ubiquitinationsaktivität reichte von keinem Signal (kein Abstrich) bis hin zu einem Multibanding-Abstrich, der bei unterschiedlichen Molekulargewichten begann, was auf unterschiedliche Ubiquitinationsmuster hindeutet.
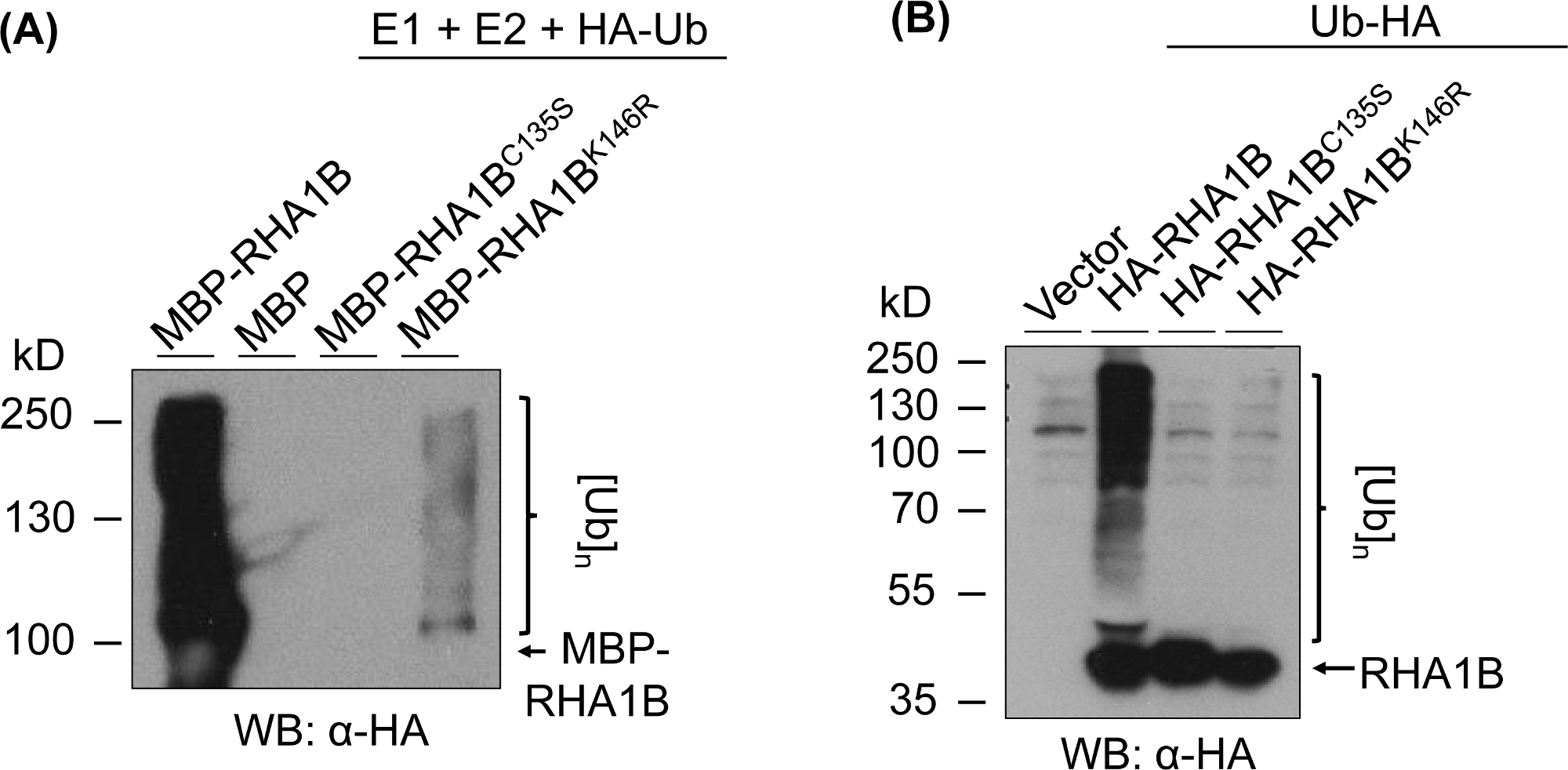
Abbildung 3 zeigt die Ergebnisse des Ubiquitinationstests für RING- und K-mutante Versionen des getesteten Proteins. Der Mangel an enzymatischer Aktivität für RHA1BC135S wird durch seine Unfähigkeit unterstützt, entweder einen Multibanding-Abstrich in vitro zu erzeugen (Abbildung 3A) oder poly-Ubiquitinationssignal in Planta zu fördern (Abbildung 3B). Es ist bemerkenswert, dass die Überexpression von HA-markierten Ub in Planta allein gab Basalebene Ubiquitination in allen getesteten Proben, einschließlich der Vektorkontrolle, im Gegensatz zu der starken Ubiquitinationssignal durch die enzymatische Aktivität des wilden Typs RHA1B übertragen. Darüber hinaus legt die Analyse der RHA1BK146R-Mutation nahe, dass der K146-Rückstand auch für die E3-Aktivität von RHA1B von wesentlicher Bedeutung ist. Obwohl ein marginales Selbstallgegenwärtigationssignal in vitro nachgewiesen wurde (Abbildung 3A), stellte der in planta assay fest, dass die Mutante E3-mangelhaft ist(Abbildung 3B, nur Hintergrund-Ubiquitinationssignal erkannt).
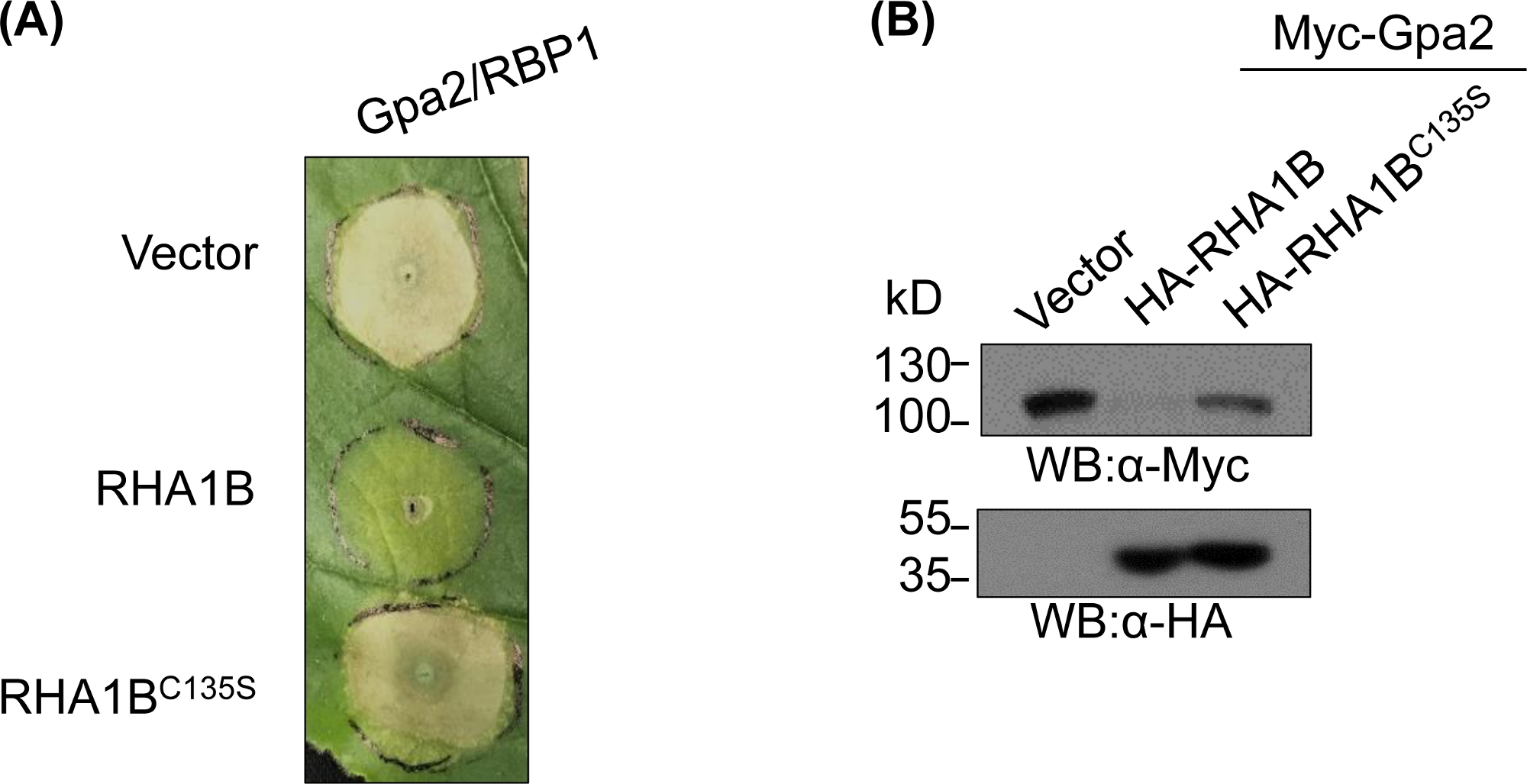
Nach der Erzeugung und biochemischen Validierung des E3-mangelhaften Mutanten können funktionelle Studien entwickelt werden, um die E3-assoziierte biologische Rolle der getesteten RING E3 Ubiquitin ligase zu bestimmen. Im Falle von RHA1B unterdrückt dieser Nematodeneffektor die pflanzenimmune Signalisierung, wie sie sich durch unterdrückung des Gpa2-ausgelösten HR-Zelltodes manifestiert. Wie in Abbildung 4Adargestellt, störte die RHA1BC135S Mutante ohne E3-Ligase-Aktivität im Gegensatz zum Wildtyp RHA1B den Tod der HR-Zellen nicht. Da das häufigste Ergebnis der Proteinubiquitination der proteasome-vermittelte Abbau ist, können Mutationen, die sich in der RING-Domäne befinden, auch verwendet werden, um eine E3-abhängige Fähigkeit zu überprüfen, den Abbau ihrer direkten und/oder indirekten Substrate auszulösen. Somit bestätigen signifikant die Ergebnisse der westlichen Blotting in Abbildung 4B, dass sich Gpa2 nicht in Gegenwart von wildem RHA1B ansammelte, aber RHA1BC135S keinen Einfluss auf die Gpa2-Proteinstabilität hatte.

Abbildung 1: Schematische Darstellung des Prinzips und der Schritte bei der standortgesteuerten Mutagenese. (A) RING-CH/H2-Domain mit konservierten Cys und Seinen Aminosäuren hervorgehoben. (B) Ein Beispiel für mutagene Primer Design. (C) Schritte der ortsgesteuerten Mutagenese. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}

Abbildung 2: Repräsentativer In-vitro-Ubiquitinationstest. (A) Top-Gel zeigt Ubiquitinationstest einschließlich aller Negativkontrollen, und das untere Gel zeigt die gleiche Belastung. (B) Der Bereich der erwarteten Ergebnisse abhängig von E2-Enzymen. Diese Zahl wurde von Kud et al8geändert. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}

Abbildung 3: Ubiquitinationstest ergebnisse für RING- und K-Mutanten (RHA1BC135S und RHA1BK146R). (A) In-vitro-Ubiquitinationsergebnisse für RHA1BC135S und RHA1BK146R. (B) In planta ubiquitination assay Ergebnisse für RHA1BC135S und RHA1BK146R. Diese Zahl wurde von Kud et al8geändert. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}

Abbildung 4: Repräsentative funktionelle Studie für E3-abhängige biologische Funktionen. Ein Beispiel für funktionelle Studien, die die E3-abhängige biologische Funktion zeigen. (A) E3-abhängige HR-Zelltodunterdrückung und (B) Abbau eines Pflanzenimmunrezeptors Gpa2. Diese Zahl wurde von Kud et al8geändert. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}
| PCR-Einrichtung | |
| 1 L | Plasmid (ca. 100 ng) |
| 1,5 l | F mutagene Grundierung (10 m) |
| 1,5 l | R mutagene Grundierung (10 m) |
| 1 L | dNTPs (10 mM) |
| 5 l | Puffer (10x) |
| 1 L | Ultra-Pfu-Polymerase (2,5 U/l) |
| 39 l | ddH2O |
| 50 l | GESAMTVOLUMEN |
Tabelle 1: PcR-Reaktion eingerichtet
| thermocycler Programm | |||
| 1 | 95 °C | 30 s | |
| 2 | 95 °C | 30 s | |
| 3 | 60 °C | 30 s | |
| 4 | 72 °C | 5 Min. | 2-4 30-mal wiederholen |
| 5 | 72 °C | 5 Min. | |
Tabelle 2: PCR-Thermocycler-Programm
| Ligationsreaktion für das RHA1B-Beispiel eingerichtet | ||
| 1,5 l | pMAL-c2::MBP linearisierter Vektor durch Verdauung mit BamHI und SalI (60 ng) | |
| 7 l | RHA1B/RHA1BC135S oder RHA1BK146R Einsatz mit BamHI und SalI (25 ng) verdaut | |
| 1 L | T4 Ligase Puffer (10x) | |
| 0,5 l | T4 Ligase (400 U/L) | |
| 10 l | GESAMTVOLUMEN | |
Tabelle 3: Ligationsreaktion für das RHA1B-Beispiel eingerichtet.
Diskussion
Die Erdeutlichung der biochemischen und mechanistischen Basis des RING Typs E3 Ubiquitin Ligasen kann wesentlich zu unserem Verständnis ihrer biologischen Bedeutung in der Entwicklung, Stresssignalisierung und Aufrechterhaltung der Homöostase beitragen. Das hier beschriebene Protokoll verbindet einen Mutageneseansatz mit in vitro und in planta-funktionellen Studien. Durch die Einführung einer einzigen Aminosäuresubstitution in die konservierten Rückstände der RING-Domäne durch standortdirekte Mutagenese kann der resultierende E3-mangelhafte Mutant parallel zu Wild-Protein getestet werden, um enzymatische Aktivität mit Funktionalität zu verknüpfen.
Es ist wichtig, die RING-Domäne richtig zu identifizieren, insbesondere ihre konservierten Cys und seine Rückstände. Online-Tools wie PROSITE können dazu10verwendet werden. Um die RING-Domäne zu destabilisieren, die für die Rekrutierung des E2-Enzyms verantwortlich ist, wird Cys normalerweise durch Ser ersetzt, was sein nächster struktureller Ersatz ist, der nicht in der Lage ist, eine Disulfidbindung zu erzeugen, die für die Zinkkoordination verwendet wird. Lorick et al. zeigten, dass die Mutation in einem dieser kritischen Cys-Rückstände die Ubiquitinationsaktivität der einzelnen Untereinheit RING-Typ E3 Ligases5abschaffen würde. Obwohl einige Cys-Rückstände auch für Mehrunit-E3-Ligasekomplexe wichtig sind, die RING-Proteine enthalten, ist es aufgrund der facettenreichen und dynamischen dreidimensionalen Struktur dieser Ubiquitinationskomplexe und der unterschiedlichen Rolle von RING-Proteinen nicht gelungen, einen Ligase-defiziden-Proteintyp11zu erzeugen.
Für die standortgesteuerte Mutagenese fanden wir heraus, dass die Verwendung kleinerer Plasmidvektoren und niedrigerer Amplifikationszyklen in der Regel zu einer höheren Effizienz bei der Mutagenese führte. Das Pfu-Enzym kann durch jede andere DNA-Polymerase mit hoher Genauigkeit und hoher Prozessivität ersetzt werden. Darüber hinaus kann, wenn das Gen von Interesse seltene Codons enthält, ein weiterer E. coli-Fleck, Rosetta, verwendet werden, um eine höhere Ausbeute des rekombinanten Proteins zu erzielen. Darüber hinaus können sowohl inkubationszeit als auch Temperatur für IPTG-Induktion weiter optimiert werden. Niedrigere Temperaturen verringern die E. coli-Teilungsrate, die für die Expression bestimmter Proteine günstig sein könnte. Obwohl eine höhere Konzentration von IPTG die Proteinexpression verbessern könnte, hemmt sie auch E. coli-Teilungsprozesse und wird nicht empfohlen.
Einzelne Untereinheit RING Typ E3 Ligases fungieren nicht nur als molekulares Gerüst, das das E2-Ub-Zwischenprodukt in unmittelbarer Nähe zum Substrat positioniert, sondern stimulieren auch die Ubiquitin-Transferaktivität ihrer Cognate E2s. Da eine E2/E3-Kombination für die Länge und die Verbindungen der Polyubiquitinkette, die das Schicksal eines modifizierten Substrats bestimmt, wichtig ist, muss jede Berücksichtigung von RING-Typ E3s ihre enzymatischen Partner E2s12umfassen. Wie in Abbildung 3Bdargestellt, sind nicht alle getesteten E2 mit der RHA1B-Ligase kompatibel. Daher sollten In-vitro-Ubiquitinationstests parallel zu mehreren E2-Enzymen durchgeführt werden, die verschiedene E2-Klassen darstellen, um falsche negative Ergebnisse zu vermeiden.
Hier wird der in vitro enzymatische Assay vorgestellt, der die Selbstallgegenwärtigisierungsfähigkeit von getesteten RING-Proteinen erkennt. Mit kleinen Modifikationen kann dieses Protokoll jedoch leicht angepasst werden, um die In-vitro-Ubiquitination von Substraten zu erkennen. Zu diesem Zweck sollte die In-vitro-Ubiquitinationsmischung aus Schritt 2.15 mit dem rekombinanten Protein des potentiellen E3-Ligasesubstrats (500 ng) ergänzt werden. Nach einer 2 h Inkubation bei 30 °C sollte ubiquitiniertes Protein mit 15 L Anti-HA-Affinitätsmatrix (wenn HA-Ub verwendet wird, oder Anti-FLAG-Affinitätsmatrix, wenn FLAG-Ub verwendet wird) durch Rühren für 2 h bei 4 °C eingefangen werden. Nach dem Waschen der Perlen 4x mal mit dem kalten Ub Waschpuffer (20 mM Tris pH 7,5, 100 mM NaCl, 0,1 mM EDTA, 0,05% Tween 20, 1x PMSF), alle bis auf 40 l des Puffers entsorgen und auf Schritt 2,16 bewegen. Das Ubiquitinationssignal, das von Antikörpern nachgewiesen wird, die spezifisch für das epitopmarkierte Ub bzw. Substrat sind und aus dem Molekulargewicht des Substratproteins hervorgehen, bestätigt die Substrat-/Enzymspezifität.
Darüber hinaus ist die Identifizierung von E3-Ligase-Substraten in vivo in der Regel mit mehreren Herausforderungen aufgrund einer transienten Enzym-Substrat-Interaktion und einem schnellen Abbau des ubiquitinierten Zielproteins verbunden. Die Verwendung eines E3-Ligase-Mangelmutanten, der immer noch mit seinem Ziel interagiert, es aber nicht mehr ubiquitiniert13, ist eine sehr nützliche Alternative zur Zugabe von proteasomalem Inhibitor MG132, der die 26S-Proteasomfunktion nicht immer ausreichend beeinträchtigt.
Ein gemeinsames Merkmal von RING-Typ E3 Ligasen ist die Tendenz, sich als Homo- und/oder Heterodimere zu bilden und zu funktionieren. Interessanterweise ist die Substitution in den konservierten Rückständen der RING-Domäne in der Regel mit einem dominanten negativen Phänotyp verbunden, bei dem mutierte RING-Typ E3 Ligase enzymatische Aktivität eines nativen Wildtypproteins13blockiert. Daher kann die Überexpression von RING-Mutanten in Planta ein alternativer Ansatz sein, um das E3-Ligase-Gen auszuschalten.
Offenlegungen
Die Autoren haben nichts zu verraten.
Danksagungen
Unsere Arbeit wurde durch die finanzielle Unterstützung der Landwirtschafts- und Ernährungsforschungsinitiative (2017-67014-26197; 2017-67014-26591) des USDA National Institute of Food and Agriculture, USDA-NIFA Farm Bill, Northwest Potato ermöglicht. Konsortium und ISDA Specialty Crop.
Materialien
| Name | Company | Catalog Number | Comments |
| Acetic acid | Sigma-Aldrich | A6283 | |
| Acetosyringone | Sigma-Aldrich | D134406 | |
| Amylose resin | NEB | E8021S | |
| ATP | Sigma-Aldrich | A1852 | |
| Bacterial protease inhibitor | Sigma-Aldrich | P8465 | |
| Bromphenol Blue | VWR | 97061-690 | |
| CaCl2 | Sigma-Aldrich | C1016 | |
| Centrifuge | Beckman Coulter | model: Avanti J-25 | |
| Commassie Blue | VWR | 97061-738 | |
| Creatine phosphate | Sigma-Aldrich | P7936 | |
| Creatine phosphokinase | Sigma-Aldrich | C3755 | |
| DNA clean & concentrator Kit | ZYMO RESEARCH | D4029 | |
| DpnI | NEB | R0176S | |
| DTT | Sigma-Aldrich | D0632 | |
| E. coli BL21 | Thermo Fisher Scientific | C600003 | |
| E. coli DH5α competent cells | Thermo Fisher Scientific | 18265017 | |
| EDTA | Sigma-Aldrich | 324504 | |
| FeSO4 7H2O | Sigma-Aldrich | F7002 | |
| FLAG-Ub | BostonBiochem | U-120 | |
| Glucose | VWR | 188 | |
| Glycerol | Sigma-Aldrich | G5516 | |
| HA-Ub | BostonBiochem | U-110 | |
| Heat block | VWR | model: 10153-318 | |
| Incubator | VWR | model: 1525 Digital Incubator | |
| Incubator shaker | Thermo Fisher Scientific | model: MaxQ 4000 | |
| IPTG | Roche | 10724815001 | |
| KCl | Sigma-Aldrich | P9333 | |
| LB Broth | Sigma-Aldrich | L3022 | |
| Liquide nitrogen | university chemistore | ||
| Maltose | Sigma-Aldrich | 63418 | |
| MES | Sigma-Aldrich | M3671 | |
| Methanol | Sigma-Aldrich | 34860 | |
| MgCl2 | Sigma-Aldrich | 63138 | |
| MgSO4 7H2O | Sigma-Aldrich | 63138 | |
| Microcentrifuge | Eppendorf | model: 5424 | |
| Miniprep plasmid purification kit | ZYMO RESEARCH | D4015 | |
| monoclonal anti-FLAG antibody | Sigma-Aldrich | F3165 | |
| monoclonal anti-HA antibody | Sigma-Aldrich | H9658 | |
| monoclonal anti-MYC antibody | Sigma-Aldrich | WH0004609M2 | |
| Mortar | VWR | 89038-144 | |
| NaCl | Sigma-Aldrich | S7653 | |
| NaH2PO4 | Sigma-Aldrich | S8282 | |
| NanoDrop | Thermo Fisher Scientific | model: 2000 Spectrophotometer | |
| Needle | Thermo Fisher Scientific | 14-826-5C | |
| NH4Cl | Sigma-Aldrich | A9434 | |
| PCR machine | Bio-Rad | model: C1000 | |
| Pestle | VWR | 89038-160 | |
| Pfu Ultra | Agilent Technologies | 600380 | |
| Plant protease inhibitor coctail | Sigma-Aldrich | P9599 | |
| pMAL-c2 | NEB | N8076S | |
| PMSF | Sigma-Aldrich | P7626 | |
| Polyvinylpolypyrrolidone | Sigma-Aldrich | P6755 | |
| SDS | Sigma-Aldrich | 1614363 | |
| Sonicator | Qsonica Sonicators | model: Q125 | |
| Syringe | Thermo Fisher Scientific | 22-253-260 | |
| Tris | Sigma-Aldrich | T1503 | |
| T4 ligase | NEB | M0202S |
Referenzen
- Freemont, P. S., Hanson, I. M., Trowsdale, J. A novel gysteine-rich sequence motif. Cell. 64, 483-484 (1991).
- Borden, K. L. B. RING fingers and B-boxes: Zinc-binding protein-protein interaction domains. Biochemistry and Cell Biology. 76, 351-358 (1998).
- Barlow, P. N., Luisi, B., Milner, A., Elliott, M., Everett, R. Structure of the C3HC4 Domain by 1H-nuclear Magnetic Resonance Spectroscopy: A New Structural Class of Zinc-finger. Journal of Molecular Biology. 237, 201-211 (1994).
- Borden, K. L. B., et al. The solution structure of the RING finger domain from the acute promyelocytic leukaemia proto-oncoprotein PML. The EMBO Journal. 14, 1532-1541 (1995).
- Lorick, K. L., et al. RING fingers mediate ubiquitin-conjugating enzyme (E2)-dependent ubiquitination. Proceedings of the National Academy of Sciences. 96, 11364-11369 (1999).
- Jiménez-López, D., Muñóz-Belman, F., González-Prieto, J. M., Aguilar-Hernández, V., Guzmán, P. Repertoire of plant RING E3 ubiquitin ligases revisited: New groups counting gene families and single genes. PLoS ONE. 13, 1-28 (2018).
- Sacco, M. A., et al. The Cyst Nematode SPRYSEC Protein RBP-1 Elicits Gpa2- and RanGAP2-Dependent Plant Cell Death. PLoS Pathogens. 5, 1-14 (2009).
- Kud, J., et al. The potato cyst nematode effector RHA1B is a ubiquitin ligase and uses two distinct mechanisms to suppress plant immune signaling. PLoS Pathogens. 15, 1007720 (2019).
- Bradford, M. M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry. 72, 248-254 (1976).
- Sigrist, C. J. A., et al. New and continuing developments at PROSITE. Nucleic Acids Research. 41, 344-347 (2013).
- Dove, K. K., Stieglitz, B., Duncan, E. D., Rittinger, K., Klevit, R. E. Molecular insights into RBR E3 ligase ubiquitin transfer mechanisms. EMBO Reports. 17, 1221-1235 (2016).
- Metzger, M. B., Pruneda, J. N., Klevit, R. E., Weissman, A. M. RING-type E3 ligases: Master manipulators of E2 ubiquitin-conjugating enzymes and ubiquitination. Biochimica et Biophysica Acta - Molecular Cell Research. 1843, 47-60 (2014).
- Xie, Q., et al. SINAT5 promotes ubiquitin-related degradation of NAC1 to attenuate auxin signals. Nature. 419, 167-170 (2002).
Nachdrucke und Genehmigungen
Genehmigung beantragen, um den Text oder die Abbildungen dieses JoVE-Artikels zu verwenden
Genehmigung beantragenThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Alle Rechte vorbehalten