
Method Article
אפיון פונקציונלי של מסוג הטבעת E3 ליגטיות בתוך מבחנה ובפלאנטה
In This Article
Summary
מטרת כתב היד היא להציג מיתאר למחקרים הביוכימיים והפונקציונליים המקיפים את הליסים של הטבעת. צינור מרובה-צעדים זה, עם פרוטוקולים מפורטים, מאמת פעילות אנזימטית של החלבון שנבדק ומדגים כיצד לקשר את הפעילות לפעולה.
Abstract
אוביקווינציה, כשינוי פוסט-ממדי של חלבונים, ממלאת תפקיד רגולטורי חשוב הומאוסטזיס של תאים איקריוטית. הקובץ המצורף הקוולנטי של 76 חומצת אמינו מכפילי משתנה לחלבון מטרה, תלוי באורך ובטופולוגיה של שרשרת הpolyubiquitin, יכול לגרום לתוצאות שונות החל מירידה בחלבון לשינויים בלוקליזציה ו/או פעילות של חלבון שונה. שלושה אנזימים ברצף לזרז את תהליך אוביקווינציה: E1 אוביקוויב-הפעלת אנזים, E2 אוביקוויב האנזים מעלה, ו E3 אוביקוויב ligase. E3 אוביקוויב ליגאז קובע ספציפיות מצע, ולכן, מייצג נושא לימוד מעניין מאוד. כאן אנו מציגים גישה מקיפה כדי ללמוד את הקשר בין הפעילות האנזימטית והפונקציה של מערכת הטבעת E3 אוביקוויב ligase. פרוטוקול ארבעה צעדים זה מתאר 1) כיצד ליצור מוטציה לקויה של E3 ליגאז באמצעות מוטסיס בבימויו של אתר המיועד בתחום הטבעת הקיימת; 2 – 3) כיצד לבחון את פעילות אוביקווינציה הן במבחנה והן בפלטה; 4) כיצד לקשר בין הניתוחים הביוכימיים הללו למשמעות הביולוגית של החלבון שנבדק. הדור של מוטציה לקויה E3 ligase כי עדיין אינטראקציה עם המצע שלה, אבל כבר לא אוביקוויטים זה עבור השפלה מקלה על בדיקות של אינטראקציות אנזימים-מצע ב vivo. יתר על כן, המוטציה בתחום טבעת שימור לעתים קרובות מקבל שלילי הפנוטיפים שליליים שיכולים להיות מנוצל ללימודי הסתרה תפקודית כגישה חלופית לגישת RNA-הפרעה. השיטות שלנו היו ממוטבת כדי לחקור את התפקיד הביולוגי של הצמח טפילים נמטודות RHA1B, אשר ממלאים את מערכת אוביקווינציה מארח בתאי הצמח כדי לקדם פרזיטיזם. באמצעות שינוי קל של מערכת הביטויים הvivo, ניתן להחיל פרוטוקול זה על הניתוח של כל E3 מסוג RING ללא קשר למקורותיו.
Introduction
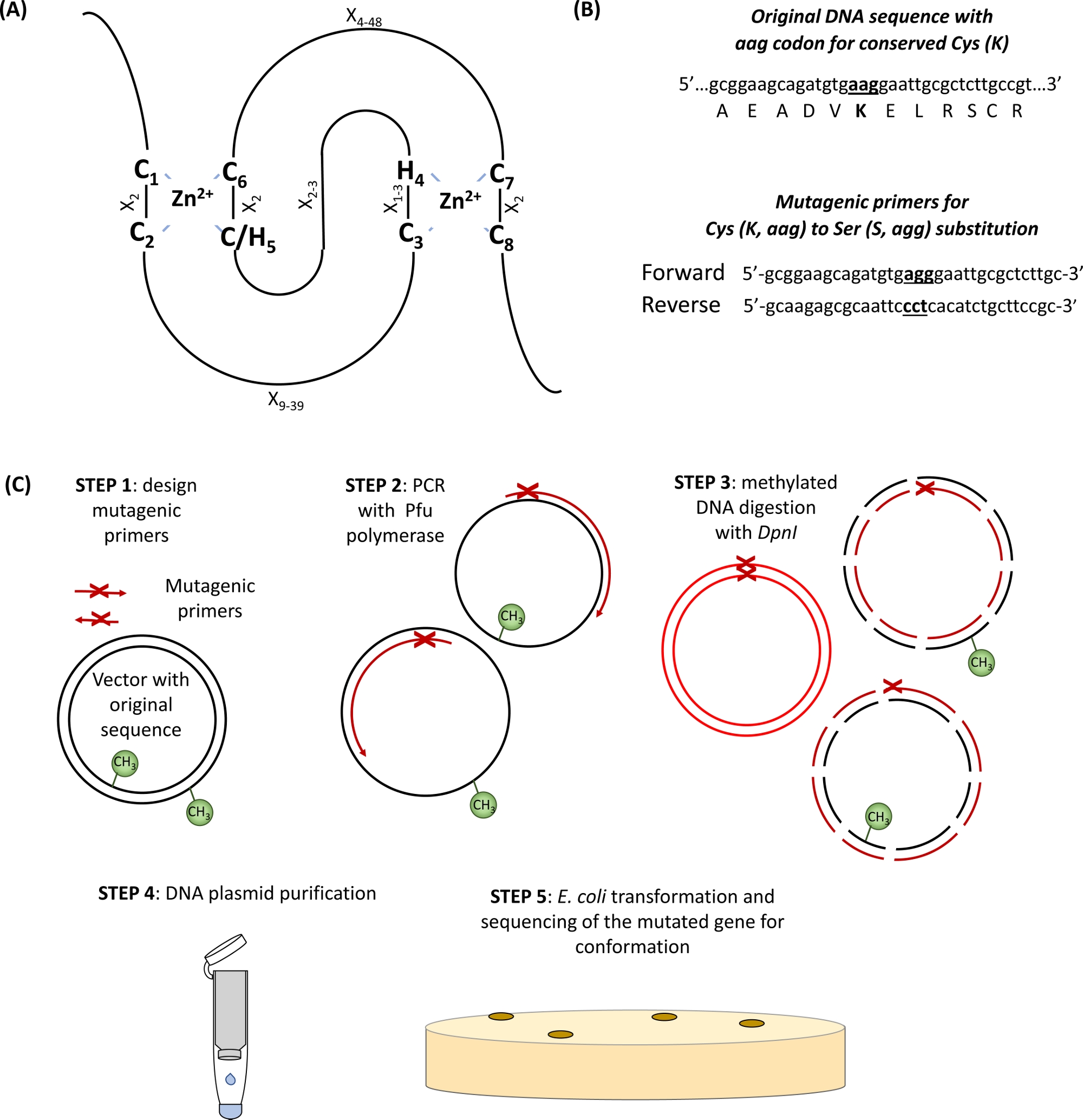
הרוב המכריע של E3 אוביקווילין ליגטיות שייך לטבעת (ר' לימעניין מעניין N ew)-סוג חלבונים. מתחם טבעת האצבע זוהה במקור על ידי Freemont et al. 1 ומבחינה פונקציונלית מתוארת כתחום אינטראקציה חלבונים בחלבון-מדיה2. האצבע הקנונית היא סוג מיוחד של תחום התאמת האבץ המוגדר כרצף הסכמה של שמונה Cys שימור (C) ו שלו (H) רווח במיוחד על ידי שאריות חומצות אמינו אחרות (X), C-X2-c-x9 – 39-c-x1 – 3-H-x2 – 3-C/h-x2-c-x4 – 48-c-x2-c. שני zn2 + יונים מיוצב על ידי הליבה c ו-h שאריות דרך ייחודי "צולבות סוגר" טופולוגיה עם c1/c2 ו c/H5/c6 תיאום הראשון zn2 + יון ואילו c3/H4 ו-c7/c8 לאגד את השני (איור 1A)3,4. בהתאם לנוכחות של C או H באתר החמישי Zn2 +-תיאום, הוגדרו שני מחלקות משניים קאנוני של חלבוני טבעת-אצבע: C3HC4 ו-C3H2C3 (טבעת-HC וטבעת-H2, בהתאמה). מאחר שתחום הטבעת של E3 אוביקוויב ליגאז מדיה את האינטראקציה בין E2 מעלה אנזימים ומצעים, מוטציה של C חיוניים אלה שאריות H הוכח לשבש את פעילות ליגאז5. חמישה מחלקות שכיחות פחות משותפות של טבעת E3 ליגסים תוארו (RING-v, טבעת-C2, טבעת-D, טבעת-S/T, ו-RING-G)6. ליגטיות מסוג RING מאפשר לחלק את הטבעות לאנזימים פשוטים ומורכבים של E3. יחידות ה-יחידת הפשוטות של הטבעת מכילות הן את אתר זיהוי המצע והן את קבוצת המחשבים של טבעת איגוד E2. לעומת זאת, מורכב המצע של מערך היחידות הרב-ממדי של היחידה, או איגוד המדיה של E2-אוביקוויב למכלול E3. מתחם RING שאריות שארית (s) המשמש כאתר המצורף אוביקוויב מצורף העיקרי עבור אוביקווינציה העצמית יכול להיות גם חשוב עבור פעילות E3 ליגאז.
לא כל החלבונים המכילים טבעת מתפקדים כ-E3 ליגסים. לפיכך, החיזוי הביומטי של תחום הטבעת-אצבע והיכולת של אוביקוויטיות בחלבון התלוי ב-E2, חייב להיות מאומת ביולוגית ומקושרת לתפקיד הביולוגי של החלבון הנבדק. כאן, אנו מתארים פרוטוקול צעד אחר צעד לתאר כיצד לזהות ולאפיין באופן פונקציונלי את הפעילות האנזימטית של הטבעת מסוג הצלצול השני, הן בתוך מבחנה והן בפלנטה, באמצעות גישת מוטזיס מכוונת באתר. התוצאות של הנציג מצינור זה מוצגות עבור מסוג הטבעת E3 ליגאז RHA1B. RHA1B הוא חלבון אפקטור המיוצר על ידי הצמח טפיל טפילים נמטודות globodera לידה כדי לדכא את החסינות הצמח לתמרן את המבנה של תאי השורש של הצמח. כדי להגן על עצמם מפני הפלישה הפתוגן/טפיל, הצמחים התפתחו נוקלאוטיד מחייב התחום ו leucine-עשיר לחזור (NB-LRR) סוג החיסון לזהות את נוכחותו של פתוגן או טפיל, כתוצאה מכך, לפתח את התגובה רגישות יתר (HR), שהיא צורה של מוות מהיר מקומי התאים המתרחשים באתר אחד כזה קולטן החיסון הוא תפוחי אדמה Gpa2 חלבון המאלף התנגדות לכמה בודד של G. pallida ידה (אוכלוסיות שדות D383 ו D372)7.
באמצעות הפרוטוקולים המוצגים, זה כבר נמצא לאחרונה כי RHA1B מפריעה עם איתות החיסונית הצמח באופן התלוי E3 על ידי מיקוד הצמח Gpa2 immunoreceptor עבור אוביקווינציה והשפלה8.
Protocol
1. מוטגנזה בבימויו של האתר (איור 1)
- לזהות את ה-Cys השומרו ואת חומצות האמינו שלו בתחום הטבעת (איור 1א) ועיצוב התחל בנושא ההחלפה החלפת הריבית מוקף 15 זוגות בסיסים משני צדדיו של אתר המוטציה (איור 1B).
- להציג את המוטציה הרצויה על ידי ה-PCR מבוססי הגברה של הפלסטלינה מחסה את הגן של הריבית באמצעות מוטציות התחל ובאיכות גבוהה DNA פולימראז המכיל Pfu ב 50 μl של נפח התגובה של PCR הכולל כפי שמוצג בטבלה 1 ובטבלה 2 על פי פרוטוקול של היצרן.
- לעכל את esהמנשכיה קולי-נגזר הורים מתילף ו-DNA semi-מתילבן על ידי הוספת 3 μl של האנזים הגבלת dpni ישירות לתגובת ה-PCR (שלב 1.2) ו דגירה ב 37 ° צ' עבור 2 h.
הערה: מתילציה היא שינוי חלבון פוסט-שניוני, המתווסף לפלסטלינה המופקת ומבודדת מחיידקים. לפיכך, עותקים חדשים של מתילציה שנוצרו באמצעות PCR, ולכן העותקים החדשים יישארו ללא שינוי במהלך הטיפול Dpni . - לטהר את הפלמידים המוטגנים באמצעות ערכת החילוץ DNA מסחרי המבוסס על טכנולוגיית העמודה ספין ו-DNA עם 50 μL של מים.
- המרה DH5α E. coli תאים כימיים המוסמכת עם 0.5 μl של ה-DNA התאושש מוטייזאת ה-דנ א בהתאם לפרוטוקול של היצרן. בקצרה, כאשר התאים המוסמכים ב-DNA על קרח במשך 30 דקות, לאחר מכן חום-הלם אותם 20 s ב 42 ° c, והמקום צינורות שוב על הקרח עבור 2 דקות. דגירה התאים עם 500 μL של מדיה LB ב 37 ° צ' עבור 1 h ב 250 rpm ולאחר מכן להפיץ אותם על pates סלקטיבי.
- לאמת את המוטציה הרצויה על ידי Sanger רצף הפלמידים DNA מבודדים E. coli.
2. טיהור חלבון רקומביננטי ובתוך שיטת הפריה חוץ גופית
- שיבוט הטבעת סוג פראי וגנים מוטציה טבעת של עניין לתוך וקטור pMAL-c2 (בצע את הפרוטוקול של היצרן; שולחן 3) כדי למזג את הגנים האלה עם תג האפיפ MBP המאפשר טיהור צעד אחד באמצעות עמילאבד שרף. הצג את המבנים המתקבלים לתוך BL21 זן E. coli כפי שמתואר בשלב 1.5.
- לגדל את החיידק E. COLI BL21 מחסה את המבנה הרצוי 50 ML ליברות בינוני נוזלי ב 37 ° צ' עבור 2 – 3 h עד שהוא מגיע לשלב לוגריתמי (OD600 של 0.4 – 0.6).
- הוסף IPTG לריכוז הסופי של 0.1 – 1 מ"מ כדי לגרום את הביטוי של MBP-מתויג רקומביננטי חלבון של ריבית ו-מודתיאת התרבות E. coli עבור 2 – 3 h ב 28 ° c. מניחים את התרבות על הקרח לאחר הדגירה.
הערה: בצע שלבים 2.4 – 2.13 על קרח כדי להגן על חלבונים מפני השפלה. - כדי לבדוק את יעילות אינדוקציה, לאסוף 1.5 mL של תאים המושרה, לסובב אותם ב 13,000 x g עבור 2 דקות, להסיר את supernatant, ולהשעות מחדש את התאים 20 μl של 2X sds-עמוד טעינה מאגר (24 mM טריס-HCl pH 6.8, 0.8% sds, 10% (v/v) גליצרול, 4 מ"מ dtt, 0.04% (w/v) בbromophenol כחול).
- מרתיחים את הדגימות עבור 5 דקות ולהפעיל אותם על 10% SDS-דף ג'ל. כדי להעריך חזותית הצטברות של חלבון MBP-fusion (משקל מולקולרי של חלבון הריבית + 42.5 kDa MBP), כתם ג'ל עבור 20 דקות על ידי המתולפת עם מאגר צביעת Coomassie (50% מתנול, 10% חומצה אצטית, 0.1% Coomassie כחול) ו להכתים לילה עם מאגר מכתים (20% מתנול, 10% חומצה אצטית).
- לקצור את התאים הנותרים E. coli על ידי צנטריפוגה ב 1,350 x g עבור 6 דקות, למחוק את supernatant, ולהשעות מחדש את הגלולה תא עם 5 מ ל של מאגר עמודה (20 mm טריס HCl, 200 מ"מ הנאל, 1 מ"מ edta, מעכב פרוטאז חיידקי).
הערה: זהו מקום טוב לעצור את הפרוטוקול בן לילה. ניתן לאחסן את התאים הקפואים עד שבוע אחד ב-20 ° c. - לשבור את התאים E. coli על ידי הצבת הצינור המכיל את החיידקים באמבט מים קרח והחלה 10 sonication מחזורים: 10 s sonication ב 30% המגבר ואחריו 20 s הפסקות.
- מדגם צנטריפוגה ב 13,000 x g ב -4 ° c עבור 10 דקות ולשמור את הסופרנטנט (תמצית גולמי).
- הכן 500 μL של עמילאבד שרף בצינור 15 מ ל. לשטוף את השרף על ידי הוספת 10 מ ל של מאגר העמודה קר ו תפרידו ב 1,800 x g, 4 ° צ' עבור 5 דקות. . תעשה את זה 2x
- הוסף 5 מ ל של תמצית גולמי לתוך הצינור עם שמני את השרף ואת הדגירה לילה ב 4 ° c.
- צנטריפוגה ב 1,800 x g ב 4 ° צ' עבור 5 דקות ולמחוק את supernatant.
- הוסף 10 מ ל של מאגר הטור כדי הגלולה שרף ו דגירה עבור 20 דקות. ואז צנטריפוגה ב 1,800 x g ב 4 ° c עבור 5 דקות. חזור על שלב זה 2x.
- לולאה חלבון היתוך עם 0.5 mL של מאגר עמודה המכיל 10 מ"מ מז על ידי הדגירה של מדגם 2 h ב 4 ° c. צנטריפוגה ב 1,800 x g ב 4 ° צ' עבור 5 דקות ולאסוף את החלבון החומק. חזור על שלב 2x זה.
- Dialyze 1 מ ל של שבר החלבון נגד ה-PBS הקרה. חלבון להשתמש בצינורות חד שימוש (10 – 20 μL) כדי למנוע להקפיא-להפשיר ולאחסן ב-80 ° צ' עד הצורך.
- למדוד את ריכוז החלבון באמצעות שיטת ברדפורד9.
- הגדר את התגובה האוביליקיתיים בהיקף כולל של 30 μL על-ידי ערבוב 40 ng של E1 (למשל, AtUBA1), 100 ng של E2 (למשל, AtUBC8, SlUBC1/4/6/7/12/13/17/20/22/27/32), 1 μg מיקרומטר-טבעת סוג חלבון, ו 2 μg דגל-רוב (או הא-והוא) ב מאגר אוביקווינציה (50 mM טריס-HCl pH 7.5, 2 מ"מ ATP, 5 מ"מ MgCl2, 30 מ"מ קראטין פוספט, 50 Μg/mL קריאטין פוספהוקינאז). מודג את התערובת ב 30 ° צלזיוס עבור 2 h.
הערה: הפוך את המאגר ל-20x, ואחסן אותו עד 6 חודשים ב-20 ° c בתוספת של שימוש בודד. קראטין פוספהוקינאז מאבד בקלות את פעילותה האנזימטית כאשר המאגר מופר ומוקפא שוב ושוב. - לסיים את התגובה על ידי ערבוב 30 μL דגימות עם 7.5 μL של 5x SDS-עמוד טעינת מאגר (60 mM טריס-HCl pH 6.8, 2% SDS, 25% (v/v) גליצרול, 10 מ"מ DTT, 0.1% (w/v) ברומאופנול כחול) ו לרתיחה עבור 5 דקות.
- הפרד את החלבונים עם 7.5% SDS-פוליאקרילמיד ג'ל אלקטרופורזה (SDS-PAGE), לאחר מכן להעביר על קרום PDVF, ולגלות אוביקווינציה על ידי בלוק המערבי באמצעות האנטי דגל (או anti-HA).
- כתם את קרום PVDF עם כחול Coomassie כדי לאמת את הטעינה שווה של בדיקות MBP-RING-סוג-החלבון.
3. Agrobacterium-מתווך חלבון ארעי ביטוי בעלי טבק benthamiana בתוך שיטת אוביטרנציה של התוכנית
- פס המתאים Agrobacterium tumefaciens זנים נושאת את אפירופה-גן מתויג של עניין (למשל, HA-RHA1B, HA-RHA1BC135S, ha-RHA1BK146R, ha-רוב) וקטור ריק כפקד על בינונית LB המכיל את האנטיביוטיקה הבחירה המתאימה.
- לאחר 2 ימים של צמיחה ב 28 ° צ', להרים מושבות יחיד ולצמוח אותם בינוני נוזלי LB עם אנטיביוטיקה המתאים 28 ° צ'/250 סל ד עבור עוד 24 h.
- העברת 100 μL של תרבות agrobacterial כדי 3 mL טרי ליברות עם אנטיביוטיקה המתאים מקרין את התרבות עבור תוספת 4 – 6 h ב 28 ° צ' עם סיבוב (250 rpm) לשלב הצמיחה המעריכית המאוחרת.
- לסובב את התאים agrobacterial ב 1,800 x g עבור 6 דקות, למחוק את supernatant, ולהשהות מחדש את התאים עם 3 מ ל של מאגר לשטוף (50 mm MES-pH 5.6, 28 מ"מ גלוקוז, 2 מ"מ מילימטר2הפו4). חזור על שלב 2x זה.
- לאחר השטיפה השנייה, השהה מחדש את התאים במאגר אינדוקציה (50 mM MES 5.6, 28 מ"מ גלוקוז, 2 מ"מ מילימטר2PO4, 200 μm acetosyringone, 37 mm4Cl, 5 מ"מ mgso4. 7H2o, 4 מ"מ kcl, 18 μm Feso4. 7h2o, 2 מ"מ cacl 2) דגירה את התאים עם מאגר אינדוקציה עבור נוספים 10 – 12 h ב 28 ° c.
הערה: Acetosyringone מעוררת העברת T-DNA. - צנטריפוגה את התאים ב 1,800 x g עבור 6 דקות, למחוק את supernatant, ולהשעות מחדש את התאים עם 2 מ ל של מאגר הסתננות (10 מ"מ MES-pH 5.5, 200 μm acetosyringone).
הערה: אם Agrobacteria צבירה לאחר דגירה עם מאגר אינדוקציה, לתת תאים צבורים לשקוע בתחתית הצינור על ידי השארת אותו על הספסל כמה דקות, ולהעביר את ההשעיה Agrobacterium ברור לצינור חדש לפני שתמשיך עם שלב 3.6. - למדוד את הריכוז של חיידקים באמצעות OD600 ערך (הצפיפות האופטית בספיגה של 600 ננומטר). התאם את ערכי OD600 לאלה הרצויים.
הערה: בדרך כלל הערך של OD600 מתאים ל-0.2-0.4 המתאים ביותר לביטוי כתם יחיד. אם שילוב של זנים agrobacterial שונים מוחל, סך OD600 ערכים של זנים agrobacterial לא יעלה על 1. - האגרוהסתנן בת 4-שבועות N. בית-החזה עוזב בעדינות ומכה אותם במחט, ואחריו מזריק יד אגרוטריום עם מזרק ללא מחט. הקף את אזור העלים החדרו עם הסמן (בדרך כלל 1 – 2 ס מ קוטר).
- לאסוף את רקמות עלה החדרו 36 h לאחר הסתננות. לטחון את הרקמה לאבקה. משובחת עם חנקן נוזלי
- אבקת רקמה מחדש עם 300 μL של מאגר חילוץ חלבון (50 mM טריס-HCl pH 7.5, 150 מ"מ, 5 מ"מ EDTA, 2 מ"מ DTT, 10% גליצרול, 1% polyvinyl, 1 מ"מ PMSF, הצמח מעכב פרוטאז המפעל) ו צנטריפוגה ב 15,000 x g עבור 15 דקות ב 4 ° c.
- . העבר את הסופרנאנט לשפופרת חדשה הוסף מאגר העמסה 5x SDS-PAGE לריכוז סופי של 1x ולרתיחה עבור 5 דקות.
- הפרדת חלבונים גולמיים על 10% SDS-דף ג ' לים, העברה אל ממברנות PVDF, ולבדוק עם anti-HA לזהות את התוכנית אוביקווינציה.
4. הקמת הקשר בין פעילות אנזימטית לפונקציה בפלאנטה
הערה: לדוגמה, RHA1B מקדמת השפלה של חלבון עמיד Gpa2 לדכא את מוות התא HR. שלב זה מראה כיצד לוודא שהפעילויות הRHA1B הללו של הפעולה הללו הן תלויות-E3.
- פס המתאים Agrobacterium tumefaciens זני נושאת גנים מתויגים של עניין (בדוגמה זו HA-RHA1B, HA-RHA1BC135S, ha-RHA1BK146R, myc-Gpa2, RBP1) וקטור ריק כפקד. בצע את השלבים 3.1 – 3.8 עבור Agrobacterium הכנה והזרקה על N. בית בעמינה עלים.
- לירידה בחלבון המצע התלוי ב-E3, בצע את הצעדים 3.9 – 3.12 ובצע את הכתמים המערביים באמצעות נוגדנים מתאימים כדי לזהות את הצטברות החלבון בתאי הצמח (למשל, anti-HA ו anti-MYC).
- עבור התגובה התלוית-היתר של ה-E3 (HR) מתווכת מוות תא, לפקח על העלים האגרוחדרו למוות הסימפטומים של תא HR 2 – 4 ימים לאחר הסתננות.
תוצאות
בסעיף זה, תוצאות הנציג מסופקים עבור הפרוטוקול המשמש לבדיקה של יחידת משנה אחת E3 אוביקוויב ליגאז RHA1B כי יש מתחם מסוג הטבעת החזוי של prosite (132 – 176 חומצות אמינו)10. כפי שמוצג באיור 1, על מנת להשיג חלבון מוטציה לקוי של E3, לפחות אחד משמונת המחשבים האחרים שמרו או Hs בתחום הטבעת (איור 1א) צריך להיות מוטסניק (איור 1ב). כך, כצעד הראשון, שתי גרסאות מוטציה של RHA1B, RHA1BC135S (החלפה של cys על ידי Ser ב שימור C3 של מתחם טבעת) ו RHA1BK146R (החלפה של ליס ידי arg בלבד lys הנוכחי ב RHA1B) נוצרו. למרות שיחידות משנה אחת E3 לתווך אוביקוויב העברה של אוביקוויב מחסה E2 למצע במקום לקיים אינטראקציה ישירה עם אוביקוויב, self-אוביקווינציה של E3 ב Lys עשוי להידרש לפעילות אנזימטית המקסימלית שלה.
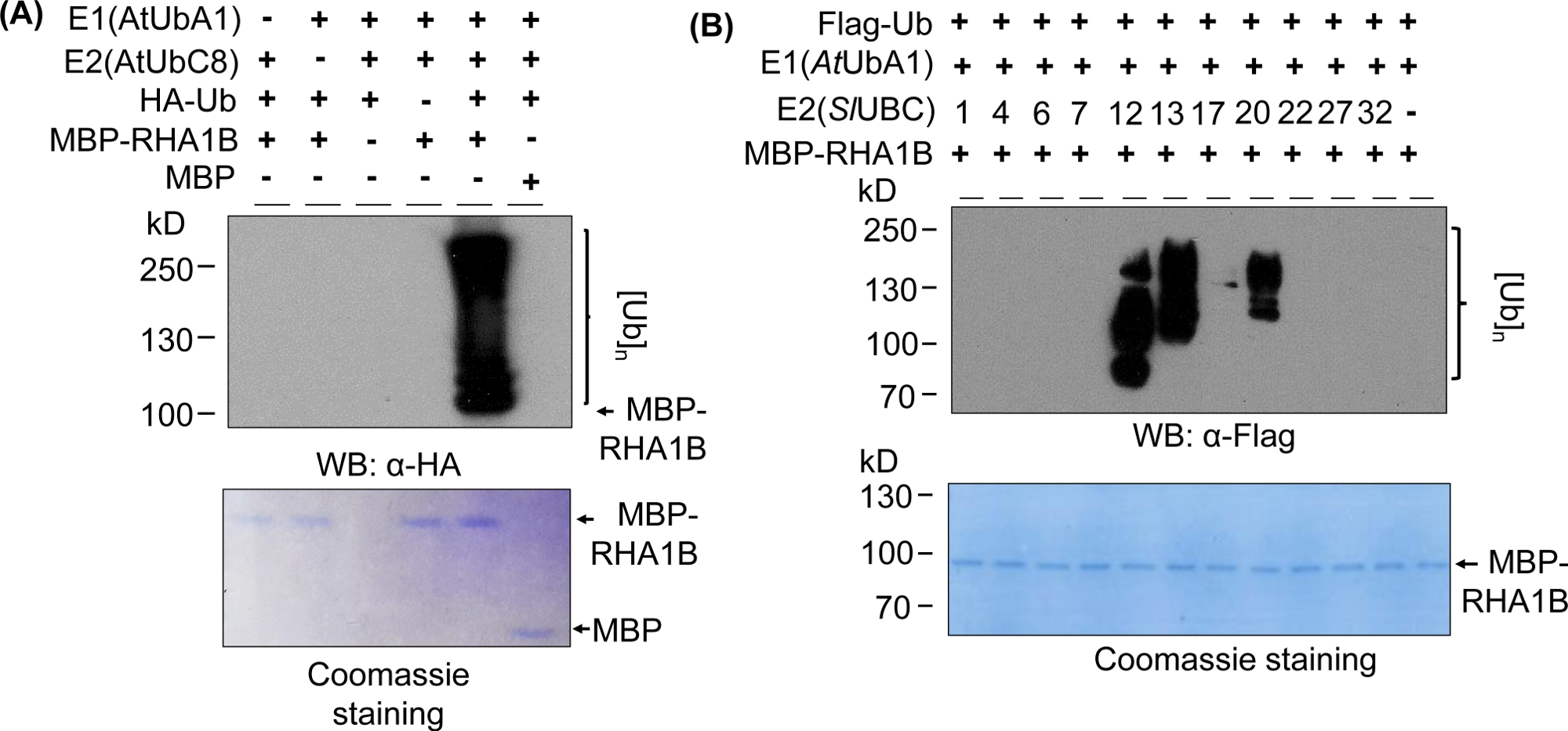
התוצאות המערביות מראה באיור 2מופע חיובי טיפוסי בתוצאה של שיטת הבחנה מחוץ לבית, עם הכתם מרובה פסים החל במשקל המולקולרי של החלבון נבדק (e. g., mpb-התמזגו RHA1B ~ 100 kda) ומתקדם כלפי מעלה. הנוגדן נגד HA זיהה את ה-המתויג ומשולבים לתוך שרשרת פולי אובילנציה של אורכים שונים, יצירת זה מטפל אופייני אוביקוויב הקשורים הסולם כמו. כדי לאמת את התוצאות החיוביות, איור 2A מציג גם את כל הפקדים השליליים החשובים החסרים רכיבים בודדים (E1, E2, רוב, או mbp-RHA1B) או באמצעות mbp כפקד וחסר האות המרוח אוביקווינציה. יתר על כן, Coomassie כחול כתמים של קרום PVDF הראה טעינה שווה של MBP-RHA1B או MBP בכל הפקדים.
איור 2ב מראה כיצד התוצאות מחוץ לגופית אוביקווינציה מגוונות בהתאם לצירוף E2/E3 הספציפי. בדוגמה זו נבדקו 11 E2s שונות שייצגו 10 משפחות שונות של E2. הפעילות המזוהה אוביקווינציה נע ללא אות (ללא מכתם) לכתם מרובה פסים החל משקולות מולקולריות שונות, המציין דפוסי אוביקווינציה שונים.
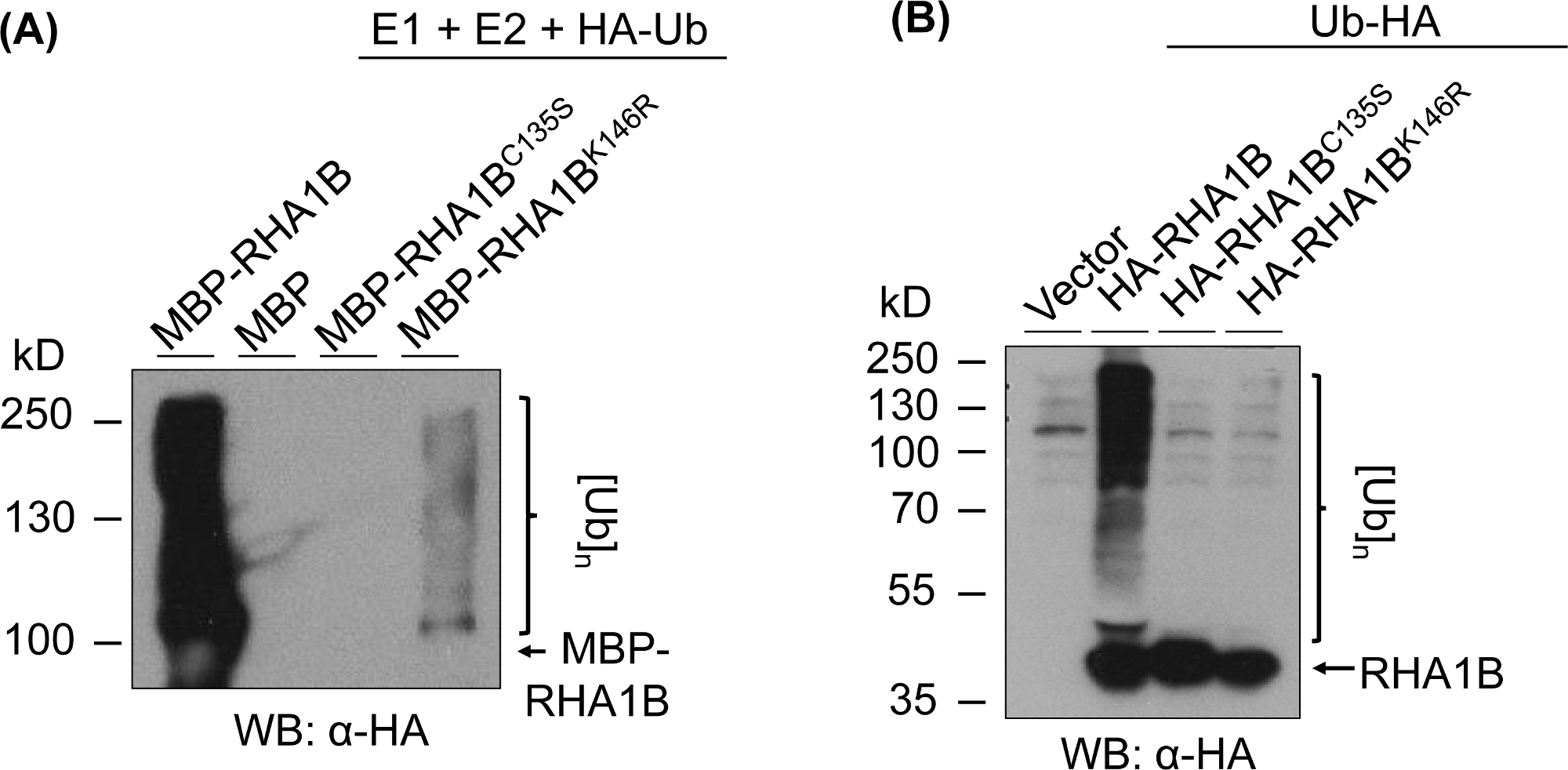
איור 3 מציג את תוצאות הבדיקה אוביקווינציה עבור הגירסאות RING ו-K-מוטציה של חלבון נבדק. העדר פעילות אנזימטית עבור RHA1BC135S נתמכת על-ידי חוסר יכולתה לייצר מיכל מרובה פסים במבחנה (איור 3א) או לקדם את האות הפולי-אוביקוויטי בפלאפטה (איור 3ב'). ראוי לציון כי ביטוי יתר של המתויגים ומתויג ב רצפה משלו נתן ברמה הבזליים אוביקווינציה בכל הדגימות נבדק, כולל שליטה וקטורית, בניגוד לאות אוביקווינציה חזק שהוענקו על ידי הפעילות אנזימטית של סוג פראי RHA1B. יתר על כן, הניתוח על RHA1BK146R מוטציה מציע כי שאריות K146 הוא חיוני גם לפעילות E3 של RHA1B. למרות שהאות העצמי השולי שלנו התגלתה במבחנה (איור 3א), בדיקת הפלטא קבעה כי המוטציה היא E3-לקויה (איור 3ב, רק רקע האות אוביקווינציה זוהה).
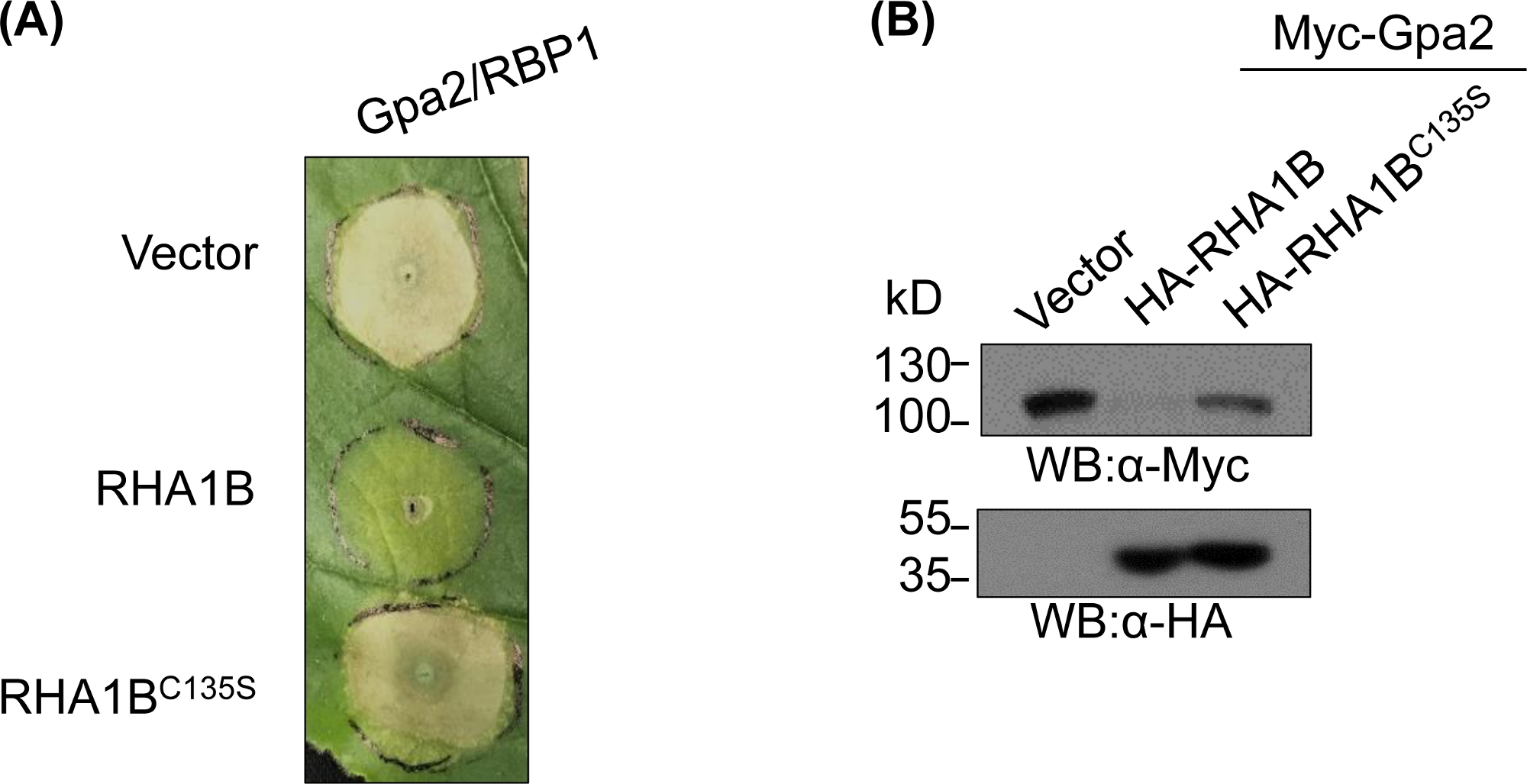
לאחר יצירת וביוכימית אימות המוטציה E3-לקויה, מחקרים פונקציונליים יכול להיות מתוכנן כדי לקבוע את התפקיד הביולוגי E3-הקשורים של הטבעת הנבדק E3 אוביקוויב ligase. במקרה של RHA1B, נמטודות זה אפקטור מדכאת איתות החיסונית של הצמח, כפי שבאה לידי ביטוי בדיכוי של מוות תא משאבי אנוש המופעל על-ידי Gpa2. כפי שהוצג באיור 4A, בניגוד לסוג פראי RHA1B, הפעילות RHA1BC135S מוטציה לא הפריעה למוות תא משאבי אנוש. בהינתן כי התוצאה השכיחה ביותר של החלבון אוביקווינציה היא השפלה מתווכת שלה, מוטציות המתגוררים בתחום הטבעת יכול לשמש גם כדי לאמת את היכולת התלוית E3 להפעיל השפלה של שלהם ישיר ו/או מצעים עקיפים. כך, באופן משמעותי, התוצאות המערביות המערבי באיור 4B לאשר כי Gpa2 לא צברו בנוכחותו של סוג פראי RHA1B אבל RHA1BC135S לא היתה השפעה על הGpa2 יציבות חלבון.

איור 1: ייצוג סכמטי של העיקרון והצעדים המעורבים במוטגנזה בבימויו של האתר. (א) טבעת-CH/H2 התחום עםשימורcys וחומצות האמינו שלו מודגש. (ב) דוגמה לעיצוב מוטאלי. (ג) צעדים של מוטזיס מונחה אתרים. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 2: נציג בתוך שיטת החוץ הגופית אוביקוויל. (א) ג'ל עליוןמציגאת הערך השלילי הכולל את כל הפקדים השליליים, והג התחתון מראה טעינה שווה. (ב) טווח התוצאות הצפויות תלוי באנזימים של E2. דמות זו שונתה מ-Kud ואח '8. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 3: התוצאות של הרשת אוביקווינציה עבור טבעת-ו-K-מוטציות (RHA1BC135S ו RHA1BK146R). (א) בתוצאות מחוץ למבחנה עבור RHA1BC135S ו RHA1BK146R. (ב) בתוצאות שיטת הRHA1BC135S והRHA1BK146R. דמות זו שונתה מ-Kud ואח '8. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 4: מחקר פונקציונלי מייצג לפונקציות ביולוגיות תלויות-E3. דוגמה למחקרים פונקציונליים המציגים את הפונקציה הביולוגית התלוית E3. (א) לדיכוי מוות תאי HR התלוי ב-E3 ו-(ב) השפלה של הצמח הimmunoreceptor Gpa2. דמות זו שונתה מ-Kud ואח '8. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}
| הגדרת ה-PCR | |
| 1 ליטר | פלמיד (~ 100 ng) |
| 1.5 מיקרומטר | F מוטאנגית פריימר (10 μM) |
| 1.5 מיקרומטר | R מוטאנניק פריימר (10 μM) |
| 1 ליטר | dNTPs (10 ממ) |
| 5 מיקרומטר | מאגר (10x) |
| 1 ליטר | אולטרה פפו פולימראז (2.5 U/μl) |
| 39 מיקרומטר | הנריהשני |
| 50 מיקרומטר | סה כ אמצעי אחסון |
שולחן 1: תגובת ה-PCR הוגדרה
| תוכנית התרמוציקלer | |||
| 1 | 95 ° c | שלושים בנות | |
| 2 | 95 ° c | שלושים בנות | |
| 3 | 60 ° c | שלושים בנות | |
| 4 | 72 ° c | 5 דקות | חזור 2-4 30 פעמים |
| מיכל 5 | 72 ° c | 5 דקות | |
שולחן 2: תוכנית ה-PCR התרמוציקלer
| תגובה מוגדרת לRHA1B לדוגמא | ||
| 1.5 מיקרומטר | pMAL-c2:: וקטור MBP באמצעות העיכול עם BamHI וסאלי (60 ng) | |
| 7 מיקרומטר | RHA1B/RHA1BC135S או RHA1BK146R להוסיף מתעכל BamHI וסאלי (25 ng) | |
| 1 ליטר | מאגר T4 ליגאז (10x) | |
| 0.5 מיקרומטר | T4 ליגאז (400 U/μl) | |
| 10 מיקרומטר | סה כ אמצעי אחסון | |
שולחן 3: התגובה הRHA1B מוגדרת לדוגמא.
Discussion
להבהיר את הבסיס הביוכימי והמכונה של סוג הטבעת E3 אוביקוויב יכול לתרום מאוד להבנת המשמעות הביולוגית שלהם בפיתוח, המתח איתות, ותחזוקה של הומאוסטזיס. הפרוטוקול המתואר כאן זוגות גישה של מוטגנזה באמצעות מבחנה ובמחקרים פונקציונליים בפלסטה. על-ידי החדרת החלפת חומצת אמינו בודדת בשקעים הניתנים לשימוש בתחום הטבעת באמצעות מוטזיס ישיר לאתר, ניתן לבדוק את המוטציה המתקבלת על-ידי מוטציה במקביל לחלבון מסוג פראי כדי לקשר בין פעילות אנזימטית לבין פונקציונליות.
חשוב לזהות כראוי את תחום הטבעת, במיוחד את ה-Cys השומרו והשקעים שלו. כלים מקוונים כגון PROSITE ניתן להשתמש כדי לעשות זאת10. כדי לערער את היציבות של תחום הטבעת האחראי לגיוס האנזים E2, Cys מוחלף בדרך כלל עם Ser, שהוא התחליף המבני הקרוב ביותר החסר את היכולת ליצור איגרת חוב דיאטתית המשמשת לתאום אבץ. Lorick ואח ' הראו כי המוטציה בכל אלה שאריות cys קריטיים היה לבטל את הפעילות אוביקוויטציה של היחידה יחידת מסוג טבעת-type5. למרות שאריות cys מסוימים חשובים גם עבור מתחמי מרובת יחידות E3 ליגאז המכילים חלבונים מסוג טבעת, בשל מבנה תלת מימדי דינמי ודינאמי של אלה מתחמי אוביקווינציה ותפקיד שונה של הטבעת-סוג חלבונים, החלפות יחיד של שאריות שימור בתחום הטבעת ב מרובת יחידות E3 ליגאז לא הצליחה ביצירת פנו
לגבי מוטזיס בבימויו של האתר, גילינו כי באמצעות וקטורים קטנים יותר בפלסטלינה ומחזורי הגברה נמוכים הניבו בדרך כלל יעילות גבוהה יותר עבור מוטזיס. האנזים Pfu יכול להיות מוחלף עם אחרים בעלי אמינות גבוהה ו-DNA התהליך גבוהה. יתר על כן, אם הגן של העניין מכיל codons נדירים, נוסף E. coli כתם, רוזטה, ניתן להשתמש כדי להשיג תשואה גבוהה יותר של חלבון רקומביננטי. בנוסף, גם זמן הדגירה וגם הטמפרטורה של האינדוקציה IPTG יכול להיות אופטימיזציה נוספת. טמפרטורות נמוכות מפחיתות את שיעור החלוקה של ה-coli , שעשוי להיות חיובי לביטוי של חלבונים מסוימים. למרות שריכוז גבוה יותר של IPTG יכול לשפר את ביטוי החלבון, הוא גם מעכב את התהליכים של החטיבה E. coli ולא מומלץ.
יחידה יחידת סוג הטבעת E3 ליגסים לא רק לתפקד כפיגום מולקולרי המהווה את המתווך E2-ורוב בסמיכות למצע, אבל גם לעורר את הפעילות העברה אוביקוויב של קנצוני שלהם E2s. יתרה מזאת, בהתחשב בכך ששילוב E2/E3 חשוב לאורך ולזמן הקישור של שרשרת הpolyubiquitin הקובעת את גורלה של מצע משתנה, כל התחשבות בסוג טבעת E3s חייבת לכלול את השותפות האנזימטית שלהם, E2s12. כפי שמוצג באיור 3B, לא כל הE2s נבדק תואמים ל-RHA1B ligase. לכן יש לבצע במקביל לאנזימים בעלי מספר רב של מחלקות e2 המייצגות שיעורי E2 שונים כדי להימנע מתוצאות שליליות כוזבות.
המוצג כאן הוא בתוך מבחנה אנזימטית מתורבת המזהה את היכולת העצמית אוביקווינציה של חלבונים מסוג טבעת בדקה. עם זאת, עם שינויים קטנים, פרוטוקול זה ניתן להתאים בקלות כדי לזהות אוביקווינציה של מצעים. למטרה זו, התערובת מחוץ לגופית אוביקווינציה משלב 2.15 צריך להיות שיושלם עם חלבון רקומביננטי של המצע E3 ליגאז פוטנציאלי (500 ng). בעקבות הדגירה של 2 h ב 30 ° צ', חלבון אוביקוויטי צריך להיות שנתפסו באמצעות 15 μL של מטריצה הזיקה אנטי-HA (אם משתמשים ב-ורוב, או אנטי דגל מטריצה הזיקה אם הדגל-רוב משמש) על ידי עצבנות עבור 2 h ב 4 ° c. לאחר שטיפת חרוזים 4x פעמים עם קר ומבריק לשטוף מאגר (20 mM Tris pH 7.5, 100 מ"מ הנאל, 0.1 mM EDTA, 0.05% רצף 20, 1x PMSF), למחוק את כל אבל 40 μL של המאגר ולעבור לשלב 2.16. האות אוביקווינציה, זוהה על ידי נוגדנים ספציפיים לאפירופה-מתויג ובמצע, בהתאמה, המתעוררים מן המשקל המולקולרי של חלבון המצע, מאשרת את המצע/האנזים ספציפיות.
יתר על כן, זיהוי של מצעים E3 ליגאז ב vivo משויך בדרך כלל עם אתגרים מרובים עקב אינטראקציה במצע האנזים ארעי והשפלה מהירה של חלבון היעד אוביקוויטים. באמצעות מוטציה לקויה E3 ליגאז, אשר עדיין אינטראקציה עם היעד שלה, אבל כבר לא אוביקוויטים זה13, היא חלופה מאוד שימושי כדי לתוספת של מעכבי פרוטאספאל MG132, אשר לא תמיד להתערב מספיק עם הפונקציה של 26 פרוטאסדום.
מאפיין נפוץ של E3 מסוג הטבעת הוא נטייה ליצור ולתפקד כמו הומו-ו/או הטרודיומרס. מעניין, ההחלפה בשקעים המשומרת של תחום הטבעת משויך בדרך כלל לפנוטיפ שלילי דומיננטי שבו מוטציה טבעת סוג E3 ליגאז חוסם הפעילות אנזימטית של סוג פראי יליד חלבון13. כך, ביטוי יתר של מוטציות RING בפלאנטה עשוי להיות גישה חלופית כדי להפיל את הגן E3 ליגאז.
Disclosures
. למחברים אין מה לגלות
Acknowledgements
עבודתנו נעשתה אפשרית על ידי התמיכה הפיננסית מתוך היוזמה החקלאית ומחקר מזון מענק תחרותי (2017-67014-26197; 2017-67014-26591) של המכון הלאומי למזון ולחקלאות, משרד החקלאות-NIFA החווה, מערב תפוחי אדמה . והוא מתמחה בגידולים
Materials
| Name | Company | Catalog Number | Comments |
| Acetic acid | Sigma-Aldrich | A6283 | |
| Acetosyringone | Sigma-Aldrich | D134406 | |
| Amylose resin | NEB | E8021S | |
| ATP | Sigma-Aldrich | A1852 | |
| Bacterial protease inhibitor | Sigma-Aldrich | P8465 | |
| Bromphenol Blue | VWR | 97061-690 | |
| CaCl2 | Sigma-Aldrich | C1016 | |
| Centrifuge | Beckman Coulter | model: Avanti J-25 | |
| Commassie Blue | VWR | 97061-738 | |
| Creatine phosphate | Sigma-Aldrich | P7936 | |
| Creatine phosphokinase | Sigma-Aldrich | C3755 | |
| DNA clean & concentrator Kit | ZYMO RESEARCH | D4029 | |
| DpnI | NEB | R0176S | |
| DTT | Sigma-Aldrich | D0632 | |
| E. coli BL21 | Thermo Fisher Scientific | C600003 | |
| E. coli DH5α competent cells | Thermo Fisher Scientific | 18265017 | |
| EDTA | Sigma-Aldrich | 324504 | |
| FeSO4 7H2O | Sigma-Aldrich | F7002 | |
| FLAG-Ub | BostonBiochem | U-120 | |
| Glucose | VWR | 188 | |
| Glycerol | Sigma-Aldrich | G5516 | |
| HA-Ub | BostonBiochem | U-110 | |
| Heat block | VWR | model: 10153-318 | |
| Incubator | VWR | model: 1525 Digital Incubator | |
| Incubator shaker | Thermo Fisher Scientific | model: MaxQ 4000 | |
| IPTG | Roche | 10724815001 | |
| KCl | Sigma-Aldrich | P9333 | |
| LB Broth | Sigma-Aldrich | L3022 | |
| Liquide nitrogen | university chemistore | ||
| Maltose | Sigma-Aldrich | 63418 | |
| MES | Sigma-Aldrich | M3671 | |
| Methanol | Sigma-Aldrich | 34860 | |
| MgCl2 | Sigma-Aldrich | 63138 | |
| MgSO4 7H2O | Sigma-Aldrich | 63138 | |
| Microcentrifuge | Eppendorf | model: 5424 | |
| Miniprep plasmid purification kit | ZYMO RESEARCH | D4015 | |
| monoclonal anti-FLAG antibody | Sigma-Aldrich | F3165 | |
| monoclonal anti-HA antibody | Sigma-Aldrich | H9658 | |
| monoclonal anti-MYC antibody | Sigma-Aldrich | WH0004609M2 | |
| Mortar | VWR | 89038-144 | |
| NaCl | Sigma-Aldrich | S7653 | |
| NaH2PO4 | Sigma-Aldrich | S8282 | |
| NanoDrop | Thermo Fisher Scientific | model: 2000 Spectrophotometer | |
| Needle | Thermo Fisher Scientific | 14-826-5C | |
| NH4Cl | Sigma-Aldrich | A9434 | |
| PCR machine | Bio-Rad | model: C1000 | |
| Pestle | VWR | 89038-160 | |
| Pfu Ultra | Agilent Technologies | 600380 | |
| Plant protease inhibitor coctail | Sigma-Aldrich | P9599 | |
| pMAL-c2 | NEB | N8076S | |
| PMSF | Sigma-Aldrich | P7626 | |
| Polyvinylpolypyrrolidone | Sigma-Aldrich | P6755 | |
| SDS | Sigma-Aldrich | 1614363 | |
| Sonicator | Qsonica Sonicators | model: Q125 | |
| Syringe | Thermo Fisher Scientific | 22-253-260 | |
| Tris | Sigma-Aldrich | T1503 | |
| T4 ligase | NEB | M0202S |
References
- Freemont, P. S., Hanson, I. M., Trowsdale, J. A novel gysteine-rich sequence motif. Cell. 64, 483-484 (1991).
- Borden, K. L. B. RING fingers and B-boxes: Zinc-binding protein-protein interaction domains. Biochemistry and Cell Biology. 76, 351-358 (1998).
- Barlow, P. N., Luisi, B., Milner, A., Elliott, M., Everett, R. Structure of the C3HC4 Domain by 1H-nuclear Magnetic Resonance Spectroscopy: A New Structural Class of Zinc-finger. Journal of Molecular Biology. 237, 201-211 (1994).
- Borden, K. L. B., et al. The solution structure of the RING finger domain from the acute promyelocytic leukaemia proto-oncoprotein PML. The EMBO Journal. 14, 1532-1541 (1995).
- Lorick, K. L., et al. RING fingers mediate ubiquitin-conjugating enzyme (E2)-dependent ubiquitination. Proceedings of the National Academy of Sciences. 96, 11364-11369 (1999).
- Jiménez-López, D., Muñóz-Belman, F., González-Prieto, J. M., Aguilar-Hernández, V., Guzmán, P. Repertoire of plant RING E3 ubiquitin ligases revisited: New groups counting gene families and single genes. PLoS ONE. 13, 1-28 (2018).
- Sacco, M. A., et al. The Cyst Nematode SPRYSEC Protein RBP-1 Elicits Gpa2- and RanGAP2-Dependent Plant Cell Death. PLoS Pathogens. 5, 1-14 (2009).
- Kud, J., et al. The potato cyst nematode effector RHA1B is a ubiquitin ligase and uses two distinct mechanisms to suppress plant immune signaling. PLoS Pathogens. 15, 1007720 (2019).
- Bradford, M. M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry. 72, 248-254 (1976).
- Sigrist, C. J. A., et al. New and continuing developments at PROSITE. Nucleic Acids Research. 41, 344-347 (2013).
- Dove, K. K., Stieglitz, B., Duncan, E. D., Rittinger, K., Klevit, R. E. Molecular insights into RBR E3 ligase ubiquitin transfer mechanisms. EMBO Reports. 17, 1221-1235 (2016).
- Metzger, M. B., Pruneda, J. N., Klevit, R. E., Weissman, A. M. RING-type E3 ligases: Master manipulators of E2 ubiquitin-conjugating enzymes and ubiquitination. Biochimica et Biophysica Acta - Molecular Cell Research. 1843, 47-60 (2014).
- Xie, Q., et al. SINAT5 promotes ubiquitin-related degradation of NAC1 to attenuate auxin signals. Nature. 419, 167-170 (2002).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved