
需要订阅 JoVE 才能查看此. 登录或开始免费试用。
Method Article
使用GAL4-UAS系统在冈比亚按蚊的功能遗传学中
摘要
二分GAL4-UAS系统是一种多功能工具,用于以受控的时空方式修饰基因表达,从而允许在 冈比亚按蚊中进行功能性遗传分析。使用该系统所描述的程序是半标准化的克隆策略,性别化和蛹荧光蛋白标志物的筛选以及胚胎固定。
摘要
二分GAL4-UAS系统是功能性遗传分析的多功能且功能强大的工具。该系统的本质是穿越以组织特异性方式表达酵母转录因子GAL4的转基因"驱动"系,转基因"响应者"系携带候选基因/RNA干扰构建体,其表达由结合GAL4的上游激活序列(UAS)控制。在随后的后代中,基因或沉默构建体因此以规定的时空方式表达,从而能够测定所得的表型并推断基因功能。二元系统使实验方法能够灵活地筛选由多种组织特异性模式的转基因表达产生的表型,即使引起严重的适应性成本。我们针对非洲主要疟疾病媒冈 比亚按蚊调整了这一系统。
在本文中,我们提供了GAL4-UAS分析期间使用的一些常用过程。我们描述了已经生成的 An. gambiae GAL4-UAS谱系,以及用于上调和RNAi敲低的新响应器结构的克隆。我们指定了蚊子蛹的分步指南,以建立遗传杂交,其中还包括筛选后代以跟踪标记驾驶员和响应者插入的荧光基因标记的遗传。我们还提出了一个清除 冈比亚锥胚胎 以研究胚胎发育的协议。最后,我们介绍了该方法的潜在适应性,通过CRISPR / Cas9插入靶基因下游的GAL4来产生驱动系。
引言
二分GAL4-UAS系统是昆虫模式生物果蝇中基因功能表征的主力1,2,3。为了使用GAL4-UAS系统,在调节序列的控制下表达酵母转录因子GAL4的转基因驱动系与携带目的基因或RNA干扰(RNAi)构建体的响应系交叉,该响应系由GAL4识别的上游激活序列(UAS)控制。该杂交的后代以由控制GAL4表达的启动子决定的时空模式表达感兴趣的转基因(图1)。可以评估驾驶员 - 响应者杂交后代显示的表型,以阐明候选基因的功能。虽然D. melanogaster已被用于检查来自其他生物的基因4,5,6,7,但GAL4-UAS系统现已适应用于具有医学和农业重要性的昆虫,以提供对感兴趣的物种的直接分析8,9,10,11,12,13,14。
在非洲疟疾蚊子冈比亚按蚊中,GAL4-UAS系统首先通过细胞系共转染9进行测试。对多种结构在不同的成对组合中测定了效率,发现当与GAL4驱动器面板一起使用时,14个串联重复的UAS补充了一个小的人造内含子(UAS-14i),显示出最宽的活化电位。为了证明体内功能,然后使用这些构建体通过PiggyBac转化8创建两个单独的转基因冈比亚锥系:一个由中肠特异性启动子驱动的携带GAL4的驱动系,以及一个在UAS序列调节下含有荧光素酶和增强型黄色荧光蛋白(eYFP)基因的响应系。肠道特异性荧光素酶活性和后代荧光表明该系统在按蚊中是有效的。从那时起,已经创建了驱动系,以接近无处不在的模式10表达对载体能力和杀虫剂耐药性很重要的其他组织中表达转基因。还产生了许多UAS系,以测定被认为参与代谢和封存介导的杀虫剂耐药性,角质层碳氢化合物合成以及荧光标记不同细胞和组织类型的基因(表1)。对于响应者系,现在通过ΦC31催化的重组盒交换17,18进行转基因的位点导向整合,以固定UAS调节基因的基因组上下文。通过这种方式,转基因表达在基因组插入位置方面被归一化,从而可以更准确地比较不同候选基因的表型效应。
迄今为止创造的响应细胞系被设计为在升高的水平上表达转基因或通过RNA干扰(RNAi)减少基因表达。 通常,cDNA克隆被融合到UAS序列中以产生合适的表达质粒,但是假设它们对于克隆来说不太大,完整的基因组序列也是可行的。为了产生沉默构建体,我们使用三种不同的方法来获得合适的串联倒置序列,这些序列形成刺激RNAi的发夹dsRNA。这些包括融合PCR,不对称PCR和发夹构建体的商业合成。每种方法的共同点是在倒置序列之间包含一个内含子序列,以提供克隆稳定性。已经开发出可插入目的基因/RNAi构建体的应答质粒15。这些质粒还携带RMCE所需的ΦC31 attB 位点(在Adolfi随附的JoVE论文中详细描述了RCME技术)。本文包含了在选择插入这些质粒之一进行过表达时所需的重要步骤的实验方案。此外,还描述了并说明了用于RNAi发夹构建体创建的两种方案。
在创建新品系时,鉴定稀有转基因个体对于繁殖以建立和维持转基因菌落至关重要。最重要的是,对于GAL4-UAS系统,有必要区分响应者和驾驶员线,以建立交叉并识别携带两种转基因的个体后代。这是通过使用与驱动和应答盒相连的不同显性可选择标记基因来实现的。最常见的是荧光标记基因,使用滤光片(例如,eYFP,eCFP,dsRed)可以清楚地区分。重要的是,标记物以已知且可靠的时空模式表达,因为这可以更容易地识别异常和污染。荧光标记基因表达通常由合成的 3xP3 启动子调节,其在 冈比亚锥发育 的所有阶段引起眼睛和腹侧神经节特异性表达19。由 3xP3 控制的荧光标记物包含在本文描述的所有转化质粒中。本文详细介绍了用于筛选荧光 冈比亚蛹 GAL4-UAS线的常用方法的方案。
GAL4-UAS系统的关键要素之一是必须跨越差分标记的驾驶员和响应者线。为此,每条线的雄性和雌性必须在交配前分开。成虫很容易通过视觉来区分,然而,为了建立遗传杂交,在成虫出现之前将性别分开是明智的,以确保没有发生交配。雄性和雌性 冈比亚蛹 之间的一般大小差异变化太大,无法成为有效和可靠的性别确定方法20。相反,外生殖器的明显形态差异为 冈比亚羚的性行为提供了可靠的基础。在本文中,我们描述了一种可靠的方法,用于对 冈比亚蛹 进行,以建立适当的杂交。

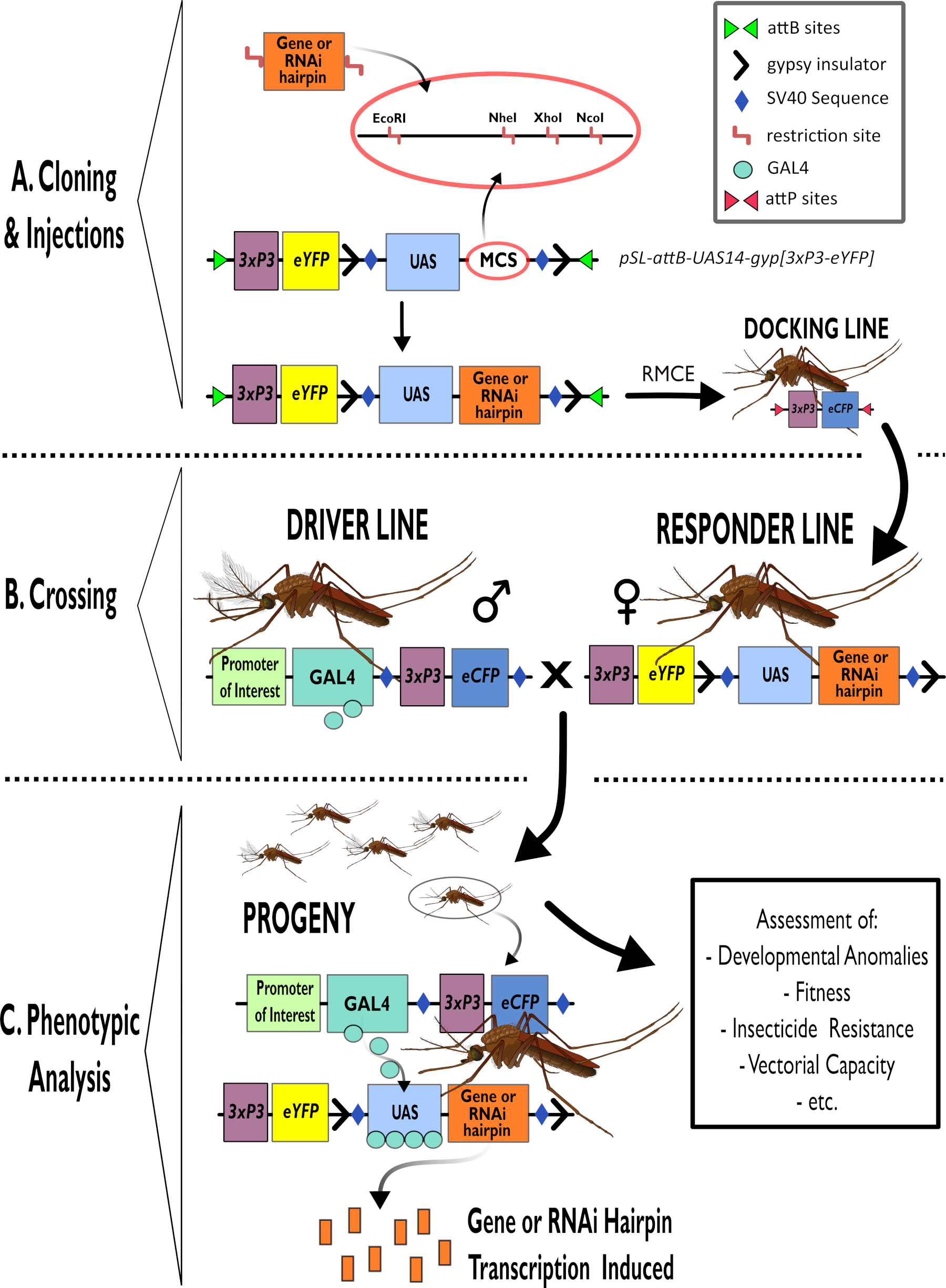
图1 -在冈比亚按蚊中使用二分GAL4-UAS系统的过程示意图。 (A)描述了示例载体(pSL-attB-UAS14-gyp[3xp3-eYFP])的主要成分,详细说明了多个克隆位点中可用于插入目标基因的发夹构建体或编码序列的可用限制位点(EcoRI,NheI,XhoI和NcoI)。还描绘了对接线的结构。(B)交叉步骤图示了来自驱动线的雄性(由感兴趣的启动子携带GAL4驱动并由3xP3启动子驱动的eCFP)和来自响应线的雌性(携带由UAS启动子控制的兴趣基因或发夹构建体和由3xP3启动子控制的eYFP标记)。(C)GAL4驱动B中杂交后代中目的基因表达的图表表示,以及评估的一些典型表型的列表。缩写:多克隆位点(MCS),重组酶介导的盒交换(RMCE),上游激活序列(UAS),增强型黄色荧光蛋白(eYFP),增强型青色荧光蛋白(eCFP)。 请点击此处查看此图的放大版本。
{kind=link}
正是交叉的使用提供了GAL4-UAS系统的二分性质,与更线性的方法相比,它具有明显的优势。例如,可以评估的驱动和响应系组合比必须为每个启动子/基因组合生成和维护新的转基因系时可行得多。更重要的是,它允许分析当其表达受到干扰时产生致命或无菌表型的基因,这些基因在线性系统中难以创建/维持。这种致命的表型可以在所有发育阶段表现出来,这取决于基因功能和时空表达,但最常在胚胎发育过程中观察到。可视化蚊子胚胎发育需要清除覆盖卵子的不透明绒毛膜。按照Trpiš(1970)21 和Kaiser等人(2014)22中描述的方法,我们描述了我们用于固定胚胎的方案,同时保持结构完整性,并漂白以清除允许微观可视化和成像的内脏。
研究方案
1. 无人机系统结构的设计和建造
- 候选基因表达载体的设计和组装
- 确定用于候选基因上调的序列。
- 从感兴趣的菌株中对cDNA / gDNA进行测序,并将其与已发表的序列进行比较,以验证其身份并确定潜在的SNP和限制性位点,以进行诊断消化。
- 确保用于基因扩增的前向引物覆盖天然Kozak序列,并在适当的情况下启动密码子。起始密码子上游约10 bp结合的引物将包含Kozak序列。
- 在大多数情况下,在从反向引物扩增的片段中包括终止密码子。使用所描述的质粒载体中提供的3'终止序列,或从候选基因基因组序列中扩增。
- 如果需要,订购具有特定密码子偏倚的商业序列。
- 使用标准亚克隆程序将基因盒插入UAS质粒载体中,例如pSL-attB-UAS14-gyp[3xP3-eYFP]15(图1),用于上调和RNAi构建。
- 生产使用ΦC31重组介导的盒式交换产生的转基因蚊子10,17,18,23。
- 确定用于候选基因上调的序列。
- RNAi 发夹构建体的创建:使用不对称 PCR 进行单步扩增15,24
- 使用Livak方法从携带所需候选基因的成年雌性冈比亚羚中提取基因组DNA(gDNA)25。
- 设计前向引物,使其与目标外显子结合,在指向相邻内含子的所需片段的5'处。设计桥底的3'端,使其与前一个外显子的末端结合,以放大内含子。5'端与紧随内含子之后的目标外显子的小片段互补。
- 运行非对称PCR反应,如 Xiao (2006)24(图2)中所述。
- 将纯化的PCR产物克隆到携带UAS启动子的合适载体中(例如,pSL-attB-UAS14-gyp[3xP3-eYFP]15)。
注:多个克隆位点中适合于pSL-attB-UAS14-gyp[3xP3-eYFP]克隆的酶15和所需的后续步骤如图1所示。 单酶消化是必不可少的,因为只添加了一个限制性位点。质粒的去磷酸化将提高克隆效率。
- 使用Livak方法从携带所需候选基因的成年雌性冈比亚羚中提取基因组DNA(gDNA)25。
- RNAi发夹结构的构建:cDNA和gDNA15的融合PCR
- 使用Livak方法从携带所需候选基因的成年雌性冈比亚羚中提取基因组DNA(gDNA)25。
- 在PCR反应中包括gDNA,以同时扩增外显子和内含子序列的靶区(图2)。
- 设计正向引物的3'端与反向靶外显子序列结合,向靶内含子序列扩增,5'端携带限制性位点,以促进克隆。
- 设计反向引物(1)以结合内含子的5'端,5'端悬垂携带相邻外显子正向序列的第一个碱基。该悬垂用于融合聚合酶链反应。
- 纯化所需的反应产物。
- 提取RNA,使用DNA酶去除DNA,并按照制造商的协议从携带所需候选基因的成年雌性冈比亚羚羊身上制备cDNA。
- 在PCR反应中使用cDNA仅扩增外显子的靶区(图2)。
- 设计前向引物(2),使3'端结合在互补靶标外显子序列的3'端,引物的5'端携带用于克隆的限制性位点。
注:1.3.1.2的正向引物可在第二反应中再次使用。然而,这将意味着单一酶消化是必不可少的。使用具有不同限制位点的第二个正向引物将允许双重消化,从而可以提高克隆效率。 - 设计反向引物(2) - 3'端与相邻外显子的5'端结合,放大目标外显子。5'端与内含子前链的3'端结合。该悬垂用于融合聚合酶链反应。
- 纯化所需的反应产物。
- 设计前向引物(2),使3'端结合在互补靶标外显子序列的3'端,引物的5'端携带用于克隆的限制性位点。
- 包括步骤1.3.1和1.3.2的产物,作为使用标准浓度和正向引物1和2进行熔融PCR反应的模板。纯化所需产品。
- 消化纯化产物以产生用于克隆的悬垂。克隆到UAS启动子下游的合适载体中。用于pSL-attB-UAS14-gyp[3xP3-eYFP]克隆的适当酶15和所需的后续步骤如图1所示。
- 使用Livak方法从携带所需候选基因的成年雌性冈比亚羚中提取基因组DNA(gDNA)25。

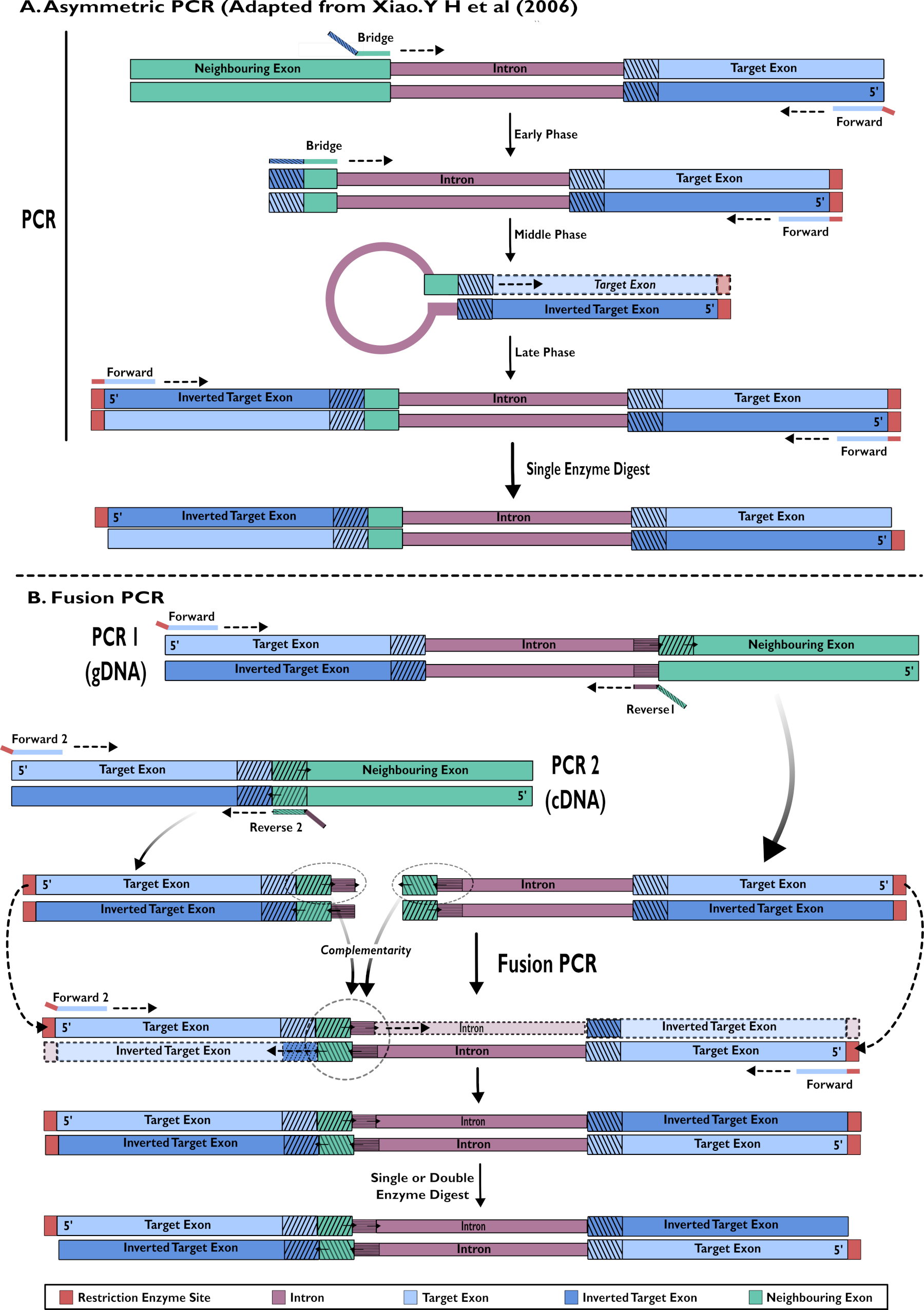
图2 -通过两种方法创建用于插入pSL-attB-UAS14-gyp[3xP3-eYFP]的RNAi构建体的示意图:(A)单步不对称PCR(改编自Xiao。Y H等人(2006)和(B)多步融合PCR。请点击此处查看此图的放大版本。
{kind=link}
2. 冈比亚蛹筛查
- 收集蛹以进行显微镜表征
注意:在这些协议中,水是指补充了0.01%池塘盐的蒸馏水。- 使用标准协议(例如MR426)的后部冈比亚蚊子进入蛹期。
注意:在整个过程中注意不要伤害蛹。 - 将蛹收集到适合与立体显微镜一起使用的透明扁平培养皿上(例如,100 x 15 mm塑料培养皿,避免边缘)。
注意:为了收集蛹,我们使用3毫升塑料巴斯德移液器,从末端切开约10毫米,以扩大末端并防止对蚊子造成伤害。筛查和性爱可以在个人身上完成,但是,这是非常缓慢的。建议对 50 - 200 只蛹的群体进行筛选和(可能的群体规模受所用菜的大小限制,并受个人喜好限制)。如果正在筛选大量蛹,则可以通过首先将蛹排列在大约4至5个深的线中,并将目标蛹移出该线来提高效率。 - 使用巴斯德移液器,小心地从蛹周围除去几乎所有的水。在蛹周围留出足够的水,使它们有效地不动,但可以用细刷子轻松移动。如果它们变得难以移动,那就加更多的水。
注意:当去除足够的水时,蛹将侧卧,允许眼睛的可视化以进行荧光检测和识别二态生殖器(图4DE)。
注意:确保蛹不会干燥。如果只留下非常少量的水,它会随着显微镜灯的热量和在蛹池之间分开时进一步减少。在此过程中,有时必须使用3 mL巴斯德移液器将额外的水添加到所需的组中。
- 使用标准协议(例如MR426)的后部冈比亚蚊子进入蛹期。
- 蛹中荧光标记物的鉴定
注意:使用低倍率立体镜允许宽视场筛选,分拣可以在倒置的复合显微镜上进行,但必须单独完成。- 在筛选荧光标记物时,首先了解预期的表达和遗传模式至关重要。请考虑以下事项:
- 颜色:确定要可视化表达式的筛选器。
- 时空表达模式:了解你期望在什么生命阶段看到表达的位置和阶段。
- 不同表型的比例:确定人口中应携带感兴趣的标记物的百分比。
- 在黑暗中进行荧光筛选,因为即使是低光也会干扰荧光的分辨率。但是,当其他操作需要光线时,请在立体镜旁边使用灯。
注意:在关灯之前,请确保荧光立体镜周围的工作空间是清晰的。 - 打开荧光灯泡,在制造商建议的时期(通常为10-15分钟)保持温暖。在荧光立体镜上选择所需的滤光片,并检查是否有一束彩色可见光指向载物台板的中心。如果这不可见或非常微弱,荧光灯泡可能尚未完全加热,快门关闭,或者显微镜光学元件未很好地对齐。
- 使用白光,将蛹居中置于视野中并使其对焦。在不同滤光片之间切换时,可能需要根据荧光强度更改此放大倍率。
- 使用精细细节画笔确保所检查的蛹不会重叠。
- 关闭立体镜的白光,并使用精细焦点使携带感兴趣表型的蛹区域成为焦点。荧光图案应该是可见的。图3提供了3xP3启动子控制荧光的示例。
- 使用最低的放大倍率,在该放大倍率下,可以可靠地将预期的荧光表型与没有荧光的个体区分开来。
- 对于具有明亮荧光的菌株,如果荧光信号仍然清晰可辨,则在筛选时也使用低强度明场光。
- 完成初级筛选后,快速扫描其他过滤器下的人群以检测潜在的污染。
注意:确保分拣的蛹组之间有明确的距离,以防止蛹移动污染。请注意,随着蛹的性别化,群体的大小会发生变化,并且在放大镜下观察时,距离可能会显得更大。当游泳池不在视野范围内时要特别小心。
- 在筛选荧光标记物时,首先了解预期的表达和遗传模式至关重要。请考虑以下事项:

图3 - 由3xP3启动子(A)eYFP,(B)dsRed和(C)eCFP驱动的荧光标记物的冈比亚按蚊蛹。放大倍率:A=16X,B,C=20X。
-
蛹
- 收集蛹。除去多余的水分,但提供足够的水分,使肛门桨部分稍微远离生殖器,以帮助可视化和形态学表征(图4D,E)。
- 如果任何蛹/e不在一侧,请使用精细的细节画笔轻轻转动蛹并移动肛门桨,以便识别外生殖器。
- 基于独特的外生殖器分离蛹;雄性有一根长管,从最后的背节挤出,大约是肛门桨长度的一半(图4B)。雌性蛹的外生殖器相当短且分叉(图4A)。
注意:有时,如果4龄幼虫外骨骼仍然附着或外生殖器受损(图4C),则更难以确定性别。当蛹的性别不清楚时,最好丢弃它。如果要保留个体,应允许蛹孤立出现,并使用成虫形态特征确定其性别。如果它的生殖器受损,个体可能无法成功交配。 - 在盘子的另一端为每种性别制作一个游泳池,使用精细的细节画笔将识别的蛹移到盘子上。标记两个池将聚集的盘子的底部,以便以后识别它们。
- 当同时需要性别和荧光筛选时,请先进行荧光筛选,因为它是两者中较快的过程。

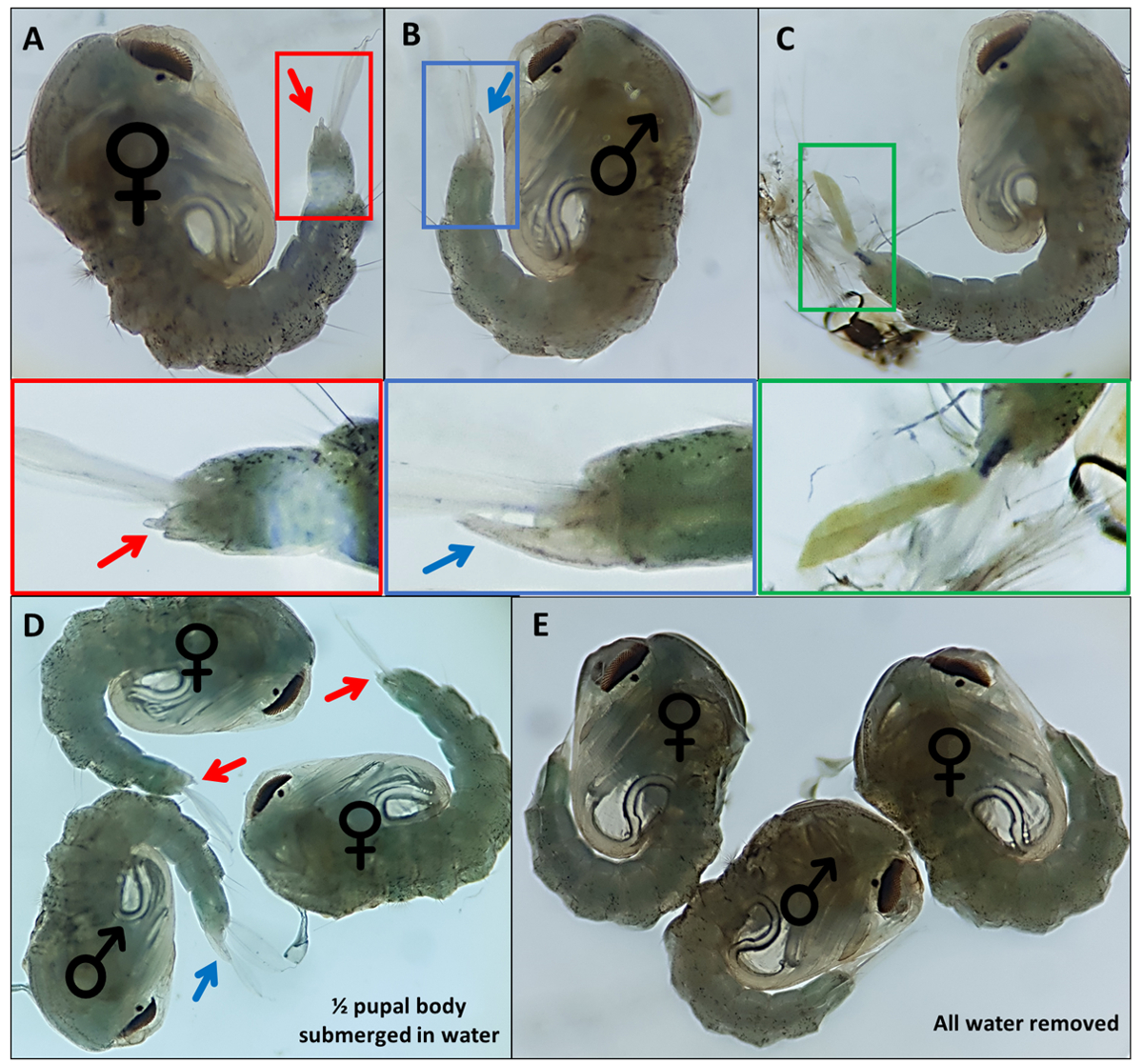
图4 -性别按蚊冈比亚蛹。个体蛹表明(A)雌性(B)雄性和(C)个体的外生殖器,由于幼虫外骨骼的不完全脱离而无法轻易识别。下面的放大图像突出显示了外生殖器。蛹与♀(雌性)和♂(雄性)表示蛹的外生殖器,(D)〜50%的蛹浸没在水中,并且(E)除去所有水,突出了外生殖器易于可视化的差异。放大倍率:A、B、C=40x、D、E=30x。请点击此处查看此图的放大图。
{kind=link}
- 成人性别确认
- 在证明非常低的错误率之前,在出现后通过成虫形态确认蛹的性别。将有性蛹分成10组或更少组,在透明的20 mL管中加入几毫升水,用棉絮球密封,标有预期的性别,并允许过夜。
注意:由于成年人在第二天早上被转移,因此没有必要为新兴成年人提供食物。 - 第二天使用形态特征确认新生成虫的性别。
- 如果雌性集合中存在任何雄性,请丢弃雌性,以防已经发生交配。
- 如果雄性集合中存在任何雌性,请移除雌性并保持雄叉。
- 在证明非常低的错误率之前,在出现后通过成虫形态确认蛹的性别。将有性蛹分成10组或更少组,在透明的20 mL管中加入几毫升水,用棉絮球密封,标有预期的性别,并允许过夜。
- 设置 GAL4-UAS 系统交叉
- 在步骤2.4中,将所需数量的雄性和雌性成虫从管中吸入以标准方式设置的笼子或小桶中,用于桫椤养殖。
注意:在转移过程中,请注意不要损坏成人。 - 使用大约50只雌性,雄性数量相等,而后代需要约2000只成年。
注意:如果要多次喂食十字架以产生多个批次,则可以在30cm x 30cm x 30cm的笼子中设置每个性别最多200个批次。当只有少量雌性(<20)可用于杂交时,我们增加约4倍的雄性数量以增加成功交配的可能性。 - 按照标准规程26,将杂交的雌鸟和后后代血液喂养到适当的阶段,以进行表型评估(例如,杀虫剂耐药性、载体能力和健身成本测定)。
- 在转基因表达可能产生母体效应的地方,设置驱动和应答者系的相互交叉并测定预期的表型。
注意:使用"杂合子"或驾驶员和响应者系的混合群体进行杂交,产生具有4种可能基因型中的每一种的后代。这提供了野生型,仅UAS和仅GAL4对照,以及用于分析表型的转杂合子GAL4-UAS。如果纯合子群体杂交,则设置额外的杂交以提供适当的对照以比较表型。应如上图对后代进行筛查,分离携带两种或仅携带其中一种标记物以及阴性的后代,以进行表型评估。
- 在步骤2.4中,将所需数量的雄性和雌性成虫从管中吸入以标准方式设置的笼子或小桶中,用于桫椤养殖。
- 从通过携带替代荧光标记物的RCME产生的品系中建立纯合子群体
注意:至关重要的是,两条系的荧光标记物都存在于相同的基因组位置,并且它们完全可区分。- 在筛查后,设置大约200名成年人的父母杂交,其中一条线的男性和另一条线的女性具有相同数量的差异标记,以选择显示正确荧光和性别的个体,如上所述。大约一周后,使用既定的协议为十字架喂血26。
- 使用标准方案将F1后代饲养到蛹,并如前所述收集蛹。
- 筛选荧光选择携带两个亲本标志物(转杂合子)的人。与这些蛹建立F1交叉。
- 一周后,按照标准方案将F1雌性和后后代血液喂养到蛹期。
- 筛选F2蛹,选择那些只显示其中一个标记的蛹。这些将是纯合子的插入。每次插入时只有25%的后代是纯合子,因此请确保饲养足够的后代以提供库存笼(400-500)。
注意:经杂合子后代的选择必须完全严格,否则该过程会受到污染,并且可能无法实现完全纯合。仔细检查为F1杂交赛选择的所有后代。
- 在筛查后,设置大约200名成年人的父母杂交,其中一条线的男性和另一条线的女性具有相同数量的差异标记,以选择显示正确荧光和性别的个体,如上所述。大约一周后,使用既定的协议为十字架喂血26。
3. 冈比亚胚胎清除方案
-
血液喂养和维护
- 尾部冈比亚蚊遵循标准规程(例如MR4)对成虫。
- 为5-7天大的成年女性提供血液喂养,确保大多数完全充血。
注意:在整个方案中,快速工作对于确保鸡蛋不燥至关重要。
-
诱导产卵
- 喂血后3天通过诱导产卵收集卵。
- 组装产卵室。
- 用水将产卵罐装满约5毫米的深度。将锅连接到50 mL聚丙烯管的一端,该管以前用钢锯切割,以便两端都打开。(我们使用塑料盘作为锅(图5);但是,可以使用管子的原始盖子代替)。
- 用材料(软管/紧身衣)或用松紧带固定的乳胶手套部分覆盖切割聚丙烯管的另一端,以便可以引入成虫但无法逃脱(图5)。存在其他替代产卵室设计,可以使用26。
- 小心地将10-15名女性(在步骤3.1.2中喂养的血液)引入产卵室。盖上产卵室以产生黑暗并离开20分钟。
注意:产卵后,避免移动产卵罐,以防止卵搁浅和干燥。 - 小心地将50 mL聚丙烯管从产卵罐中取出,同时确保不会释放蚊子。白色的卵子应该是可见的。检查是否已为被禁止的目的奠定了足够的资金。如有必要,请重复上述步骤。
- 盖上锅盖(用于防尘),让鸡蛋成熟到感兴趣的发育阶段。
- 使用精细的细节画笔从锅中捡起鸡蛋,并将它们放在40毫米2挖掘的玻璃块中的水中。

图5 - 拆解产卵室(A)以突出显示组件和(B)组装的示例。 请点击此处查看此图的放大版本。
{kind=link}
-
胚胎固定
注意:由于使用甲醛,请在通风橱中执行所有固定步骤(步骤3.3)。- 准备FAA解决方案,如Kaiser等人(2014)22中所述。FAA由3.6 M甲醛,0.87M乙酸和8.5M无水乙醇组成,体积与蒸馏水(dH2O)组成。
- 对于10 mL FAA,将2.68 mL的13.42 M甲醛,4.96 mL的17.14 M乙醇和0.5 mL的17.4 M乙酸与1.86 mL蒸馏的H2O混合在一起。
- 用微量移液管小心地从玻璃块中取出水,并将鸡蛋盖在500μLFAA中,并在室温下在轨道振荡器上轻轻振荡(约25 RPM)30分钟。此时不可见颜色变化。
- 用蒸馏水彻底冲洗鸡蛋。进行15次冲洗,以去除甲醛的所有痕迹。使用1000μL微量移液器,一次添加然后除去1mL的dH 2 O,确保在此过程中不会损坏卵子。
- 将冲洗产生的废水储存在指定的甲醛丢弃容器中,以便根据安全指南进行处理。
- 此时,固定的鸡蛋可以在4°C的水中储存过夜,以保持水分。
- 准备FAA解决方案,如Kaiser等人(2014)22中所述。FAA由3.6 M甲醛,0.87M乙酸和8.5M无水乙醇组成,体积与蒸馏水(dH2O)组成。
-
胚胎漂白
注意:在通风橱中执行所有漂白步骤(步骤4),因为次氯酸钠和乙酸结合时可能会释放氯气。- 制备漂白溶液(Trpiš溶液 - 在Trpiš(1970)21中描述并根据Kaiser等人(2014)22进行修改)。Trpiš溶液是0.59M次氯酸钠和0.35M乙酸溶解在蒸馏H 2O中。
- 对于 10 mL Trpiš 溶胶的卵,将 2.68 mL 2.2 M 次氯酸钠和 0.2 mL 17.4 M 乙酸与 7.12 mL 蒸馏 H2 O 混合。
注:Trpiš溶液可以在密封的玻璃容器中储存至少3个月,并保存在安全的化学橱柜中。溶液在储存后可能需要涡旋,并且应始终在通风橱中打开,以防氯气释放。
- 对于 10 mL Trpiš 溶胶的卵,将 2.68 mL 2.2 M 次氯酸钠和 0.2 mL 17.4 M 乙酸与 7.12 mL 蒸馏 H2 O 混合。
- 用1 mL Trpiš溶液覆盖固定的鸡蛋,并在室温下孵育30分钟。卵在孵化约5分钟后开始形成苍白斑块,最终在清除后达到乳白色。
- 如步骤3.3.3中冲洗鸡蛋以除去Trpiš溶液。
- 将废水储存在指定的废物容器中,并与多余的水一起排入下水道。
- 制备漂白溶液(Trpiš溶液 - 在Trpiš(1970)21中描述并根据Kaiser等人(2014)22进行修改)。Trpiš溶液是0.59M次氯酸钠和0.35M乙酸溶解在蒸馏H 2O中。
-
存储
- 储存在500μL的dH 2 O中,并在2-8°C之间保持几天。在观察和成像质量之前,请小心地除去大部分水,但要通过在手表玻璃中留下少量水来避免鸡蛋干燥。这不会干扰拍摄鸡蛋。可以将单个卵子放在显微镜载玻片上,以进行更高放大倍率的成像。
结果
eYFP,dsRed和eCFP的3xP3表达提供了可靠,易于区分的个体,这些个体具有在冈比亚蛹的眼睛和腹侧神经节中产生表达的标记基因(图3)。 图 4 突出显示了在用于的男性和女性外生殖器中观察到的差异形态以及无法识别的蛹的示例。从蛹中去除所有水分会增加性交难度,因为肛门桨模糊了生殖器的可视化(图4D,E<...
讨论
了解蚊子的基因功能对于开发控制按蚊和影响疟疾传播的新方法至关重要。所描述的GAL4-UAS系统是一种多功能且功能强大的系统,用于候选基因的功能分析,迄今为止,我们已使用该系统来检查杀虫剂抗性17和角质层碳氢化合物生产的遗传基础15,23,以及荧光标记不同的蚊子细胞群8.到目前为止,已经通过限制?...
披露声明
作者没有什么可透露的。
致谢
我们非常感谢LSTM和IVCC(Adriana Adolfi),BBSRC(新研究员奖(AL),MRC(BCP:MR / P016197 / 1博士生奖学金),Wellcome(Henry Wellcome爵士博士后奖学金LG:215894 / Z / 19 / Z)的资助,这些资金已将Gal4UAS分析纳入提案中。
材料
| Name | Company | Catalog Number | Comments |
| 100 x 15 mm plastic Petri dish | SLS | 2175546 | Pack of 10 |
| 1000 µL Gilson Pipette | Gilson | F144059P | |
| 20/25 mL Universal Tubes | Starlab | E1412-3020 | Pack of 400 |
| 3 mL Pasteur Pipettes | SLS | G612398 | Greiner Pasteur pipette 3 mL sterile individually wrapped |
| 50 mL Falcon Tubes | Fisher Scientific | 11512303 | |
| Absolute Ethanol | Fisher Scientific | BP2818-500 | 500 mL |
| Acetic Acid | SLS | 45726-1L-F | 1 L |
| Cages | SLS | E6099 | 30x30x30 with screen port |
| Fine Paint Brushes | Amazon | UKDPB66 | KOLAMOON 9 Pieces Detail Painting Brush Set Miniture Brushes for Watercolor, Acrylic Painting, Oil Painting (Wine Red) |
| Fish food | Amazon | Tetra Min Fish Food, Complete Food for All Tropical Fish for Health, Colour and Vitality, 10 L | |
| Formaldehyde Solution | Sigma Aldrich | F8775 | |
| Mouth Aspirator | John Hock | 612 | |
| Pond Salt | Amazon | Blagdon Guardian Pond Tonic Salt, for Fish Health, Water Quality, General Tonic, pH Buffer, 9.08 kg, treats 9,092 L | |
| Pupae Pots | Cater4you | SP8OZ | 250 pots with lids |
| Small Plastic Buckets | Amazon | 2.5 L White Plastic Pail Complete with White Lid (Pack of 10) | |
| Sodium Hypochlorite | Fisher Scientific | S25552 |
参考文献
- Brand, A. H., Perimon, N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development. 118 (2), 401-415 (1993).
- Duffy, J. B. GAL4 system in drosophila: A fly geneticist's swiss army knife. Journal of Genetics and Development. 34 (1-2), 1-15 (2002).
- Dow, J. A. . ELS. , (2012).
- Edi, C. V., et al. CYP6 P450 Enzymes and ACE-1 Duplication Produce Extreme and Multiple Insecticide Resistance in the Malaria Mosquito Anopheles gambiae. PLoS Genetics. 10 (3), 1004236 (2014).
- Daborn, P. J., et al. Using Drosophila melanogaster to validate metabolism-based insecticide resistance from insect pests. Insect Biochemistry and Molecular Biology. 42 (12), 918-924 (2012).
- Riveron, J. M., et al. Genome-wide transcription and functional analyses reveal heterogeneous molecular mechanisms driving pyrethroids resistance in the major malaria vector Anopheles funestus across Africa. Genes Genomes Genetics. 7 (6), 1819-1832 (2017).
- Riveron, J. M., et al. A single mutation in the GSTe2 gene allows tracking of metabolically based insecticide resistance in a major malaria vector. Genome Biology. 15 (2), (2014).
- Lynd, A., Lycett, G. J. Development of the Bi-Partite Gal4-UAS System in the African Malaria Mosquito, Anopheles gambiae. PLoS ONE. 7 (2), 31552 (2012).
- Lynd, A., Lycett, G. J. Optimization of the Gal4-UAS system in an Anopheles gambiae cell line. Insect Molecular Biology. 20 (5), 599-608 (2011).
- Adolfi, A., Pondeville, E., Lynd, A., Bourgouin, C., Lycett, G. J. Multi-tissue GAL4-mediated gene expression in all Anopheles gambiae life stages using an endogenous polyubiquitin promoter. Insect Biochemistry and Molecular Biology. 96, 1-9 (2018).
- Kokoza, V. A., Raikhel, A. A. Targeted gene expression in the transgenic Aedes aegypti using the binary Gal4-UAS system. Insect Biochemistry and Molecular Biology. 41, 637-644 (2011).
- O'Brochta, D. A., Pilitt, K. L., Harrell, R. A., Aluvihare, C., Alford, R. T. Gal4-based Enhancer-Trapping in the Malaria Mosquito Anopheles stephensi. Genes Genomes Genetics. 2, 21305-21315 (2012).
- Zhao, B., et al. Regulation of the Gut-Specific Carboxypeptidase: A Study Using the Binary Gal4/UAS System in the Mosquito Aedes Aegypti. Insect Biochemistry and Molecular Biology. 54, 1-10 (2014).
- Imamura, M., et al. Targeted Gene Expression Using the GAL4/UAS System in the Silkworm Bombyx mori. Genetics. 165 (3), 1329-1340 (2003).
- Lynd, A., et al. Development of a functional genetic tool for Anopheles gambiae oenocyte characterisation: application to cuticular hydrocarbon synthesis. bioRxiv. , (2019).
- Pondeville, E., et al. Hemocyte-targeted gene expression in the female malaria mosquito using the hemolectin promoter from Drosophila. Insect Biochemistry and Molecular Biology. 120, 103339 (2020).
- Adolfi, A., et al. Functional genetic validation of key genes conferring insecticide resistance in the major African malaria vector, Anopheles gambiae. Proceedings of the National Academy of Sciences of the United States of America. 116 (51), 25764-25772 (2019).
- Pondeville, E., et al. Efficient integrase-mediated site-specific germline transformation of Anopheles gambiae. Nature Protocols. 9 (7), 1698-1712 (2014).
- Horn, C., Schmid, B. G. M., Pogoda, F. S., Wimmer, E. A. Fluorescent transformation markers for insect transgenesis. Insect Biochemistry and Molecular Biology. 32, 1221-1235 (2002).
- Clements, A. . A. Biology of Mosquitoes, Volume 1: Development, Nutrition and Reproduction. 1, (1992).
- Trpiš, M. A new bleaching and decalcifying method for general use in zoology. Canadian Journal of Zoology. 48, 892-893 (1970).
- Kaiser, M. L., Duncan, F. D., Brooke, B. D. Embryonic Development and Rates of Metabolic Activity in Early and Late Hatching Eggs of the Major Malaria Vector Anopheles gambiae. PLoS ONE. 9 (12), 114381 (2014).
- Grigoraki, L., Grau-Bové, X., Yates, H. C., Lycett, G. J., Ranson, H. Isolation and transcriptomic analysis of Anopheles gambiae oenocytes enables the delineation of hydrocarbon biosynthesis. eLife. 9, 58019 (2020).
- Xiao, Y. -. H., Yin, M. -. H., Hou, L., Pei, Y. Direct amplification of intron-containing hairpin RNA construct from genomic DNA. BioTechniques. 41 (5), 548-552 (2006).
- Livak, K. J. Organization and Mapping of a Sequence on the Drosophila melanogaster X and Y Chromosomes That Is Transcribed during Spermatogenesis. Genetics. 107 (4), 611-634 (1984).
- MR4, CDC, NEI & beiResources. . The MR4 Methods in Anopheles Research Laboratory Manual. 5th Edition. , (2015).
- Sik Lee, Y., Carthew, R. W. Making a better RNAi vector for Drosophila: use of intron spacers. Methods. 30 (4), 322-329 (2003).
- Cha-aim, K., Hoshida, H., Fukunaga, T., Akada, R., Peccoud, J. . Gene Synthesis: Methods and Protocols. , 97-110 (2012).
- Cavener, D. R. Comparison of the consensus sequence flanking translational start sites in Drosophila and vertebrates. Nucleic Acids Research. 15 (4), 1353-1361 (1987).
- Wang, Y., Wang, F., Wang, R., Zhao, P., Xia, Q. 2A self-cleaving peptide-based multi-gene expression system in the silkworm Bombyx mori. Scientific Reports. 5, (2015).
- Galizi, R., et al. A synthetic sex ratio distortion system for the control of the human malaria mosquito. Nature Communications. 5, 3977 (2014).
- Kondo, S., et al. Neurochemical organisation of the Drosophila Brain Visualised by Endogenously Tagged Neurotransmitter Receptors. Cell Reports. 30 (1), 284-297 (2020).
- Lee, P. -. T., et al. A gene-specific T2A-GAL4 library for Drosophila. eLife. 7, 35574 (2018).
- Marois, E., et al. High-throughput sorting of mosquito larvae for laboratory studies and for future vector control interventions. Malaria Journal. 11, 302 (2012).
- Crawford, J. E., et al. Efficient production of male Wolbachia-infected Aedes aegypti mosquitoes enables large-scale suppression of wild populations. Nature Biotechnology. 38 (4), 482-492 (2020).
- Goltsev, Y., et al. Developmental and evolutionary basis for drought tolerance of the Anopheles gambiae embryo. Developmental Biology. 330 (2), 462-470 (2009).
- Rezende, G. L., et al. Embryonic desiccation resistance in Aedes aegypti: presumptive role of the chitinized Serosal Cuticle. BMC Developmental Biology. 8 (1), 82 (2008).
- Vargas, H. C. M., Farnesi, L. C., Martins, A. J., Valle, D., Rezende, G. L. Serosal cuticle formation and distinct degrees of desiccation resistance in embryos of the mosquito vectors Aedes aegypti, Anopheles aquasalis and Culex quinquefasciatus. Journal of Insect Physiology. 62, 54-60 (2014).
- Chang, C. -. H., et al. The non-canonical Notch signaling is essential for the control of fertility in Aedes aegypti. PLOS Neglected Tropical Diseases. 12 (3), 0006307 (2018).
- Clemons, A., Flannery, E., Kast, K., Severson, D., Duman-Scheel, M. Immunohistochemical Analysis of Protein Expression during Aedes aegypti Development. Spring Harbor Protocols. 10, 1-4 (2010).
- Juhn, J., James, A. A. Hybridization in situ of Salivary Glands, Ovaries and Embryos of Vector Mosquitoes. Journal of Visualized Experiments. , e3709 (2012).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。