
JoVE 비디오를 활용하시려면 도서관을 통한 기관 구독이 필요합니다. 전체 비디오를 보시려면 로그인하거나 무료 트라이얼을 시작하세요.
Method Article
Anopheles 감비아에서 기능 유전학을 위한 GAL4-UAS 시스템을 이용하십시오
요약
이 양파르트 갈4-UAS 시스템은 Anopheles 감비아에서 기능성 유전 분석을 허용하는 제어된 스파티오 측량 방식으로 유전자 발현을 수정하기 위한 다목적 도구입니다. 이 시스템을 사용하기 위해 설명된 절차는 형광 단백질 마커 및 배아 고정을 위한 pupae의 반 표준화된 복제 전략, 섹스링 및 검열입니다.
초록
이중 형 GAL4-UAS 시스템은 기능성 유전 분석을위한 다재다능하고 강력한 도구입니다. 시스템의 본질은 조직 특정 방식으로 효모 전사 인자 GAL4를 표현하는 형질전환 '드라이버' 라인을 교차하는 것으로, GAL4를 결합하는 업스트림 활성화 서열(UAS)에 의해 발현이 조절되는 후보 유전자/RNA 간섭 구조를 운반하는 형질형 '응답자' 라인을 갖는다. 이어지는 자손에서, 유전자 또는 침묵 구조는 이렇게 규정된 현면 방식으로 발현되어 결과 표현형을 분석하고 유전자 기능을 유추할 수 있게 한다. 이 바이너리 시스템은 심한 피트니스 비용이 유도되더라도 여러 조직 별 패턴에서 트랜스진 발현에 의해 생성된 표현형을 선별하는 실험적 접근법의 유연성을 가능하게 한다. 우리는 Anopheles 감비아에 대 한이 시스템을 적응, 아프리카에 있는 주요 말라리아 벡터.
이 문서에서는 GAL4-UAS 분석 중에 사용되는 몇 가지 일반적인 절차를 제공합니다. 우리는 이미 생성 된 An. 감비아 GAL4-UAS 라인뿐만 아니라 업 규제 및 RNAi 노크다운을위한 새로운 응답자 구조의 복제를 설명합니다. 우리는 유전 십자가를 확립하기 위하여 모기 강아지의 성화를 위한 단계별 가이드를 지정합니다, 또한 드라이버와 응답자 삽입을 태그하는 형광 유전자 마커의 상속을 따르는 검열 자성을 포함합니다. 우리는 또한 배아 발달을 연구하기 위하여 An. 감비아 어 태아를 지우기 위한 프로토콜을 제시합니다. 마지막으로, 대상 유전자의 GAL4 다운스트림의 CRISPR/Cas9 삽입을 통해 드라이버 라인을 생성하는 방법의 잠재적 인 적응을 소개합니다.
서문
이 양파르트-UAS 시스템은 곤충 모델 유기체 Drosophila melanogaster1,2,3에서 유전자의 기능적 특성화의 주력이다. GAL4-UAS 시스템을 사용하기 위해, 변성 드라이버 라인은, 규제 서열의 통제하에 효모 전사 인자 GAL4를 표현하고, GAL4에 의해 인식되는 업스트림 활성화 서열(UAS)에 의해 제어되는 관심 또는 RNA 간섭(RNAi) 구조의 유전자를 운반하는 응답자 라인과 교차된다. 본 십자가의 자손은 GAL4 발현을 제어하는 프로모터에 의해 지시된 현면 패턴에 대한 관심의 변형을 표현한다(도 1). 운전자-응답자 십자가의 자손에 의해 표시되는 표현형은 후보 유전자의 기능을 해명하기 위하여 평가될 수 있다. D. 멜라노가스터는 다른 유기체로부터 유전자를 검사하는 데 사용되었지만4,5,6,7, GAL4-UAS 시스템은 이제 관심있는 종에 대한 직접 분석을 제공하기 위해 의료 및 농업 중요성의 곤충에 사용하기 위해 적응되어 8,9,9,10,11,12,13,14.
아프리카 말라리아 모기에서, Anopheles 감비아, GAL4-UAS 시스템은 세포주 공동 형질제9에 의해 처음으로 시험되었습니다. 여러 개의 구조는 서로 다른 쌍방향 조합의 효율성을 위해 분석되었으며, 14개의 UAS가 작은 인공 인트론(UAS-14i)으로 보충되어 GAL4 드라이버 패널과 함께 사용할 때 가장 광범위한 활성화 잠재력을 나타낸다는 것을 발견했습니다. 생체 내 기능을 입증하기 위해, 이러한 구조는 PiggyBac 변환8에 의해 두 개의 별도의 형질형 An. 감비아 라인을 만드는 데 사용되었다: midgut 특정 프로모터에 의해 구동 GAL4를 운반하는 드라이버 라인, 그리고 UAS 서열의 규정에 따라 루시파라제및 향상된 황색 형광 단백질 (eYFP) 유전자를 모두 포함하는 응답자 라인. 자손의 직감 특이적 루시파제 활동과 형광은 시스템이 Anopheles에서 효율적이라고 지적했다. 그 이후, 운전자 라인은 이노뉴테15 및 혈구16을 포함한 벡터 용량 및 살충제 저항에 중요한 다른 조직에서 전경과를 발현하고, 유비쿼터스 패턴10에 가깝게 만들어졌다. 수많은 UAS 라인은 또한 대사 및 격리 중재 살충제 저항, 큐티칸 탄화수소 합성 및 형광 태그 다른 세포 및 조직 유형 (표 1)에 관여하는 것으로 생각 되는 분석 유전자에 생성 되었습니다. 응답자 라인의 경우, 이제 ΦC31 촉매 재조합 카세트 교환17,18에 의해 생성되어 UAS 규제 유전자의 게놈 컨텍스트를 수정하여 트랜스진의 현장 지향적 통합이 수행된다. 이러한 방식으로, 유전자 발현은 게놈 삽입 위치에 대해 정상화되어 상이한 후보 유전자의 표현 효과보다 정확한 비교를 가능하게 한다.
현재까지 생성된 응답자 라인은 높은 수준에서 트랜스진을 발현하거나 RNA 간섭(RNAi)을 통해 유전자 발현을 감소시키도록 설계되었습니다. 일반적으로 cDNA 클론은 UAS 서열에 융합되어 적절한 발현 플라스미드를 생성하지만 전체 게놈 서열도 복제에 너무 크지 않다고 가정할 수 있습니다. 침묵 구조를 생성하기 위해, 우리는 RNAi를 자극하는 헤어 핀 dsRNA를 형성하는 적절한 탠덤 반전 시퀀스를 얻기 위해 세 가지 다른 방법을 사용했다. 여기에는 융합 PCR, 비대칭 PCR 및 헤어핀 구조의 상업적 합성이 포함되었습니다. 각 방법에 공통적으로 복제 안정성을 제공하기 위해 반전된 시퀀스 사이에 인트론 시퀀스를 포함시키는 것이 일반적이다. 관심/RNAi 구조유전자를 삽입할 수 있는 응답자 플라스미드가 개발되었다15. 이 플라스미드는 또한 RMCE에 필요한 ΦC31 attB 사이트를 수행합니다 (RCME 기술을 자세히 설명하는 JoVE 종이와 함께 아돌피에 설명). 과발현을 위해 이러한 플라스미드 중 하나에 삽입시퀀스를 선택할 때 필요한 중요한 단계를 다루는 프로토콜이 이 원고에 포함되어 있습니다. 또한 RNAi 헤어핀 구조 생성을 위한 두 가지 프로토콜이 설명되고 설명되어 있습니다.
새로운 라인을 만들 때, 희귀 한 형질 전환 개인의 식별은 설립하고 형질 전환 식민지를 유지하기 위해 번식하는 것이 중요합니다. 가장 중요한 것은 GAL4-UAS 시스템의 경우 응답자와 운전자 라인을 구별하여 교차를 설정하고 두 트랜스유전자를 모두 수행하는 개별 자손을 식별할 필요가 있습니다. 이것은 드라이버와 응답자 카세트에 연결된 다른 지배적인 선택 가능한 마커 유전자를 사용하여 달성됩니다. 가장 일반적으로 이들은 광학 필터를 사용하여 명확하게 구별되는 형광 마커 유전자입니다 (예를 들어, eYFP, eCFP, dsRed). 이 이상과 오염의 식별을 쉽게하기 때문에 마커는 알려진 신뢰할 수있는 실각 측량 패턴으로 표현되는 것이 중요합니다. 형광 마커 유전자 발현은 합성 3xP3 프로모터에 의해 일상적으로 조절되며, 이는 An. 감비아 개발의 모든 단계에서 눈과 복부 신경리아 특이적 발현을 일으킨다. 3xP3 에 의해 제어되는 형광 마커는 이 문서에 기재된 모든 변환 플라스미드에 포함되어 있습니다. 형광을 검사하는 데 사용되는 일반적인 방법을 자세히 설명하는 프로토콜 An. 감비아 에 푸파에 GAL4-UAS 라인이 여기에 포함되어 있습니다.
GAL4-UAS 시스템의 핵심 요소 중 하나는 차별화된 표시된 드라이버 및 응답자 라인을 교차해야 한다는 것입니다. 각 줄에서이 남성과 여성은 짝짓기 전에 분리해야합니다. 성인은 시력에 의해 쉽게 구별 할 수 있습니다, 그러나, 유전 십자가를 확립하기위한 것은 짝짓기가 발생하지 않았는지 확인하기 위해 성인 출현 전에 남녀를 분리하는 것이 합리적이다. 남성과 여성의 일반적인 크기 차이는 너무 가변적이며, 성결정의 효율적이고 신뢰할 수 있는 방법이 될 수 있다20. 대신 외부 생식기에 명확한 형태적 차이는 An. 감비아에서 섹스에 대한 신뢰할 수있는 기초를 제공합니다. 이 기사에서는 적절한 십자가를 세우기 위해 An. 감비아 푸를 섹스하는 신뢰할 수있는 방법을 설명합니다.

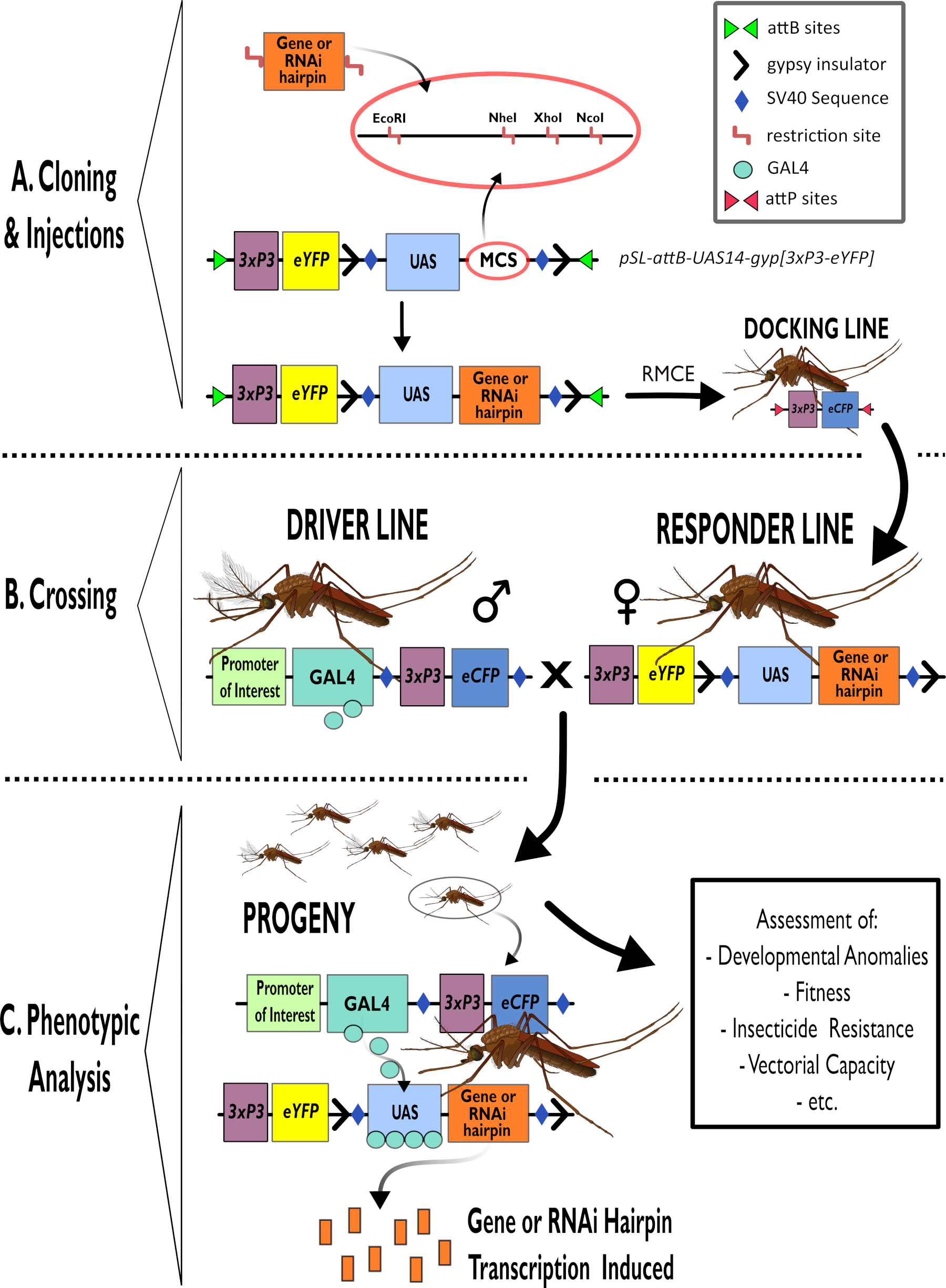
그림 1 - 아노펠스 감비아에서 이중 PARTite GAL4-UAS 시스템을 사용하기 위한 공정의 다이어그램 표현. (A) 예시 벡터(pSL-attB-UAS14-gyp[3xp3-eYFP]의 주요 구성 요소는 사용 가능한 제한 부위(EcoRI, NheI, XhoI 및 NcoI)를 상세히 묘사하며, 이는 관심 있는 유전자에 대한 헤어핀 구조 또는 코딩 서열을 삽입하는 데 사용하기에 적합한 다중 복제 부위 내에서 묘사된다. 도킹 라인의 구조도 묘사됩니다. (B) 교차 단계는 운전자 라인(3xP3 프로모터에 의해 구동되는 관심 및 eCFP의 발기인에 의해 GAL4 드라이버를 운반)과 응답자 라인에서 여성의 사용을 나타내는 것으로 도시된다(UAS 프로모터에 의해 제어되는 관심 또는 헤어핀 구조의 유전자를 운반하고 3xP3 프로모터에 의해 제어되는 eYFP 마커). (C) B에서 십자가의 자손에 대한 관심 유전자의 GAL4 구동 발현과 평가되는 전형적인 표현형의 일부 목록의 다이어그램 표현. 약어: 다중 복제 부위(MCS), 재조합 중재 카세트 교환(RMCE), 업스트림 액티베이터 서열(UAS), 향상된 황색 형광 단백질(eYFP), 향상된 시안 형광 단백질(eCFP). 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
그것은 더 선형 접근 에 비해 뚜렷한 장점을 가지고 GAL4-UAS 시스템의 쌍층 특성을 제공하는 십자가의 사용이다. 예를 들어, 드라이버 와 응답자 라인의 더 많은 조합은 새로운 형질 전환선이 생성되고 각 프로모터/유전자 조합에 대해 유지되어야 하는 경우 실현 가능한 것보다 평가될 수 있습니다. 더 중요한 것은, 선형 시스템에서 생성/유지가 어려운 발현이 위태로빙될 때 치명적또는 멸균 표현형을 생성하는 유전자의 분석을 허용한다. 이러한 치명적인 표현형은 유전자 기능 및 현면 발현에 따라 모든 발달 단계에서 나타날 수 있지만 배아 발달 중에 가장 자주 관찰됩니다. 모기 배아 발달을 시각화하려면 계란을 코팅하는 불투명한 초리온을 비우는 것이 필요합니다. Trpiš (1970)21 및 Kaiser 외.(2014)22에 기술된 방법에 따라, 우리는 구조적 무결성을 유지하면서 배아를 고치는 데 사용하는 프로토콜을 설명하고, 미세한 시각화 및 이미징을 허용하는 엔도토리온을 지우기 위해 표백합니다.
프로토콜
1. UAS 구조물의 설계 및 시공
- 후보 유전자 발현을 위한 벡터의 설계 및 조립
- 후보 유전자 업 규제에 사용할 서열을 결정한다.
- 관심 있는 변형으로부터 cDNA/gDNA를 시퀀싱하고 게시된 서열과 비교하여 신원을 확인하고 진단 다이제스트를 위한 잠재적인 SMP 및 제한 부위를 식별합니다.
- 유전자 증폭에 사용되는 전방 프라이머가 네이티브 Kozak 서열을 커버하고 적절한 경우 코돈을 시작하도록 하십시오. 시작 코돈의 상류에 ~ 10 bp 바인딩 프라이머는 코작 시퀀스를 포함합니다.
- 대부분의 상황에서 역 프라이머로부터 증폭된 단편에 스톱 코든을 포함합니다. 설명된 플라스미드 벡터에 제공된 3' 종단 서열을 사용하거나, 후보 유전자 게놈 서열로부터 증폭한다.
- 원하는 경우 특정 코돈 바이어스로 상업용 시퀀스를 주문하십시오.
- 표준 서브클론 절차를 사용하여 UAS 플라스미드 벡터에 유전자 카세트를 삽입합니다(예: pSL-attB-UAS14-gyp[3xP3-eYFP]15(그림 1) 모두 업규제 및 RNAi 구조에 대해 삽입한다.
- ΦC31 재조합 카세트 교환10,17,18,23을 사용하여 생성 된 형질 전환 모기를 생산합니다.
- 후보 유전자 업 규제에 사용할 서열을 결정한다.
- RNAi 헤어핀 구조 생성: 비대칭 PCR15,24를 이용한 단일 단계 증폭
- 추출물 게놈 DNA (gDNA)는 Livak 방법을 사용하여 원하는 후보 유전자를 운반하는 성인 여성 An. 감비아인으로부터 25.
- 원하는 조각의 5'에서 대상 엑손에 바인딩하는 전진 프라이머를 설계하여 인접한 인트론을 향합니다. 브리지 프라이머의 3'끝을 설계하여 앞의 엑손 끝에 결합하여 인트론을 증폭시합니다. 5'end는 인트론 직후 대상 엑손의 작은 조각에 보완된다.
- 샤오(2006)24(도 2)에 기재된 바와 같이 비대칭 PCR 반응을 실행한다.
- 정제된 PCR 제품을 UAS 프로모터를 운반하는 적합한 벡터로 복제한다(예를 들어, pSL-attB-UAS14-gyp[3xP3-eYFP]15).
참고: pSL-attB-UAS14-gyp[3xP3-eYFP] 복제15 및 필요한 다음 단계에 적합한 다중 복제 부위 내의 효소가 도 1에 표시됩니다. 단일 효소 다이제스트는 하나의 제한 부위만 추가될 때 필수적입니다. 플라스미드의 디포스포릴레이션은 복제 효율을 향상시킵니다.
- 추출물 게놈 DNA (gDNA)는 Livak 방법을 사용하여 원하는 후보 유전자를 운반하는 성인 여성 An. 감비아인으로부터 25.
- RNAi 헤어핀 구조 의 건설 : cDNA와 gDNA15의 융합 PCR
- 추출물 게놈 DNA (gDNA)는 Livak 방법을 사용하여 원하는 후보 유전자를 운반하는 성인 여성 An. 감비아인으로부터 25.
- 엑손 및 인트론 서열의 표적 영역을 함께 증폭시키는 PCR 반응에 gDNA를 포함한다(도 2).
- 전방 프라이머의 3'끝을 설계하여 역표적 엑손 서열에 결합하여 대상 인트론 서열을 향해 증폭하고 5'end는 복제를 용이하게 하기 위해 제한 부위를 운반한다.
- 디자인 리버스 프라이머(1)는 인트론의 5'엔드에 결합하고 5'끝 오버행은 이웃 엑손의 전방 시퀀스의 첫 번째 베이스를 운반한다. 이 오버행은 융합 PCR에 사용됩니다.
- 원하는 반응 제품을 정화합니다.
- RNA를 추출하고, DNase를 사용하여 DNA를 제거하고 제조업체의 프로토콜에 따라 원하는 후보 유전자를 운반하는 성인 여성 An. 감비아에서 cDNA를 준비합니다.
- PCR 반응에서 cDNA를 사용하여 엑슨의 표적 영역만 증폭한다(도 2).
- 설계 포워드 프라이머(2)는 3'end가 보완 대상 엑슨 서열의 3'끝에 바인딩되도록 하고 프라이머의 5'끝이 복제에 사용하기 위한 제한 부위를 전달한다.
참고: 1.3.1.2의 전방 프라이머는 이 두 번째 반응에서 다시 사용할 수 있습니다. 그러나, 이것은 단 하나 효소 다이제스트가 필수적이다는 것을 의미할 것입니다. 다른 제한 사이트와 두 번째 앞으로 프라이머를 사용하면 복제 효율을 높일 수있는 이중 다이제스트를 허용합니다. - 디자인 역프라이머 (2) - 3'엔드는 대상 엑슨을 증폭 이웃 엑슨의 5'끝에 바인딩합니다. 5'끝은 인트론의 3'끝에 결합하여 앞으로 가닥을 앞으로 합니다. 이 오버행은 융합 PCR에 사용됩니다.
- 원하는 반응 제품을 정화합니다.
- 설계 포워드 프라이머(2)는 3'end가 보완 대상 엑슨 서열의 3'끝에 바인딩되도록 하고 프라이머의 5'끝이 복제에 사용하기 위한 제한 부위를 전달한다.
- 단계 1.3.1 및 1.3.2의 제품을 앞으로 프라이머 1 및 2를 사용하여 융합 PCR 반응을 위한 템플릿으로 포함한다. 원하는 제품을 정화합니다.
- 정제된 제품을 소화하여 복제를 위한 오버행을 생성합니다. UAS 프로모터의 적절한 벡터 다운스트림으로 복제합니다. pSL-attB-UAS14-gyp[3xP3-eYFP] 복제15 및 필요한 다음 단계를 위한 적절한 효소가 도 1에 표시된다.
- 추출물 게놈 DNA (gDNA)는 Livak 방법을 사용하여 원하는 후보 유전자를 운반하는 성인 여성 An. 감비아인으로부터 25.

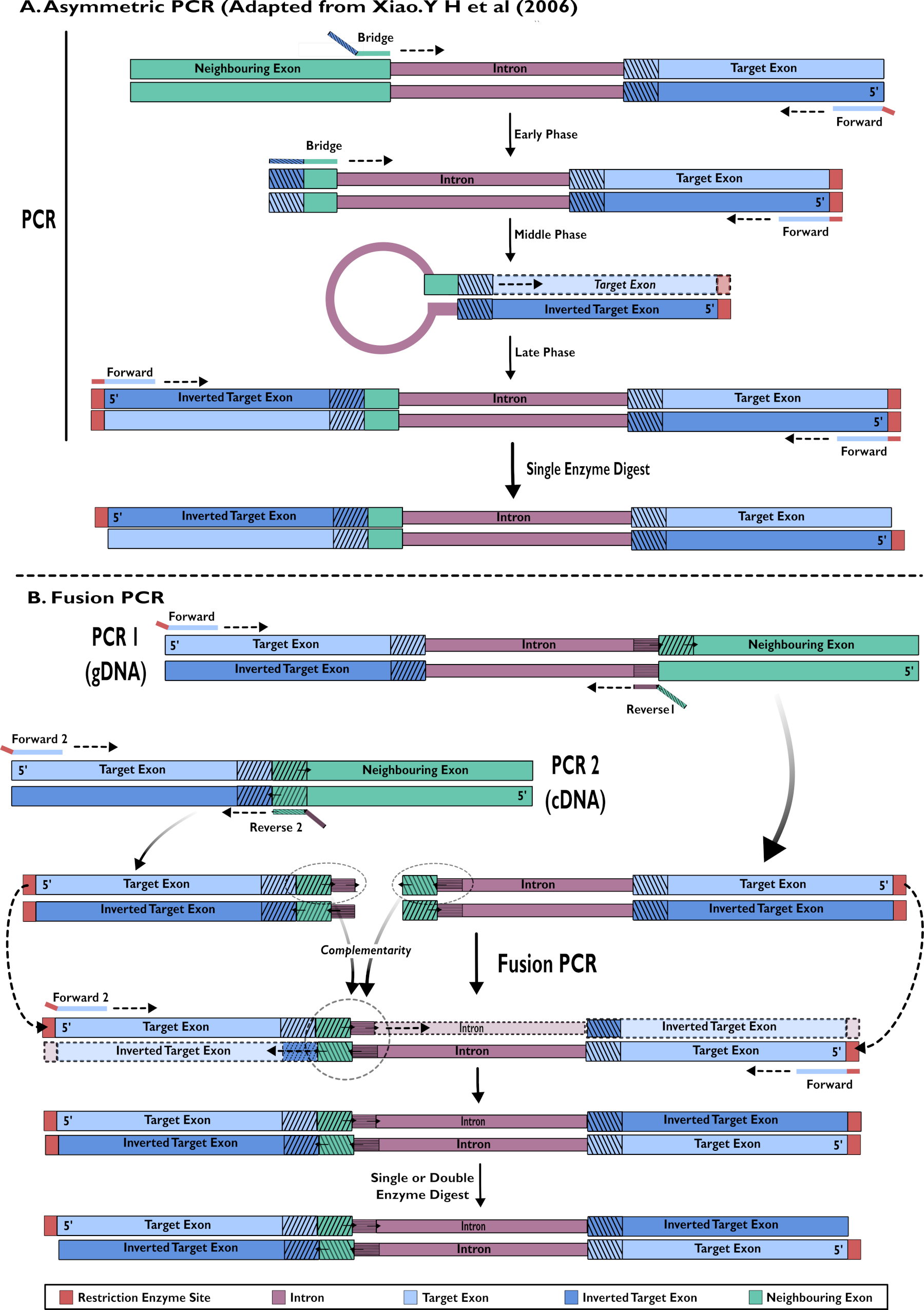
도 2 - pSL-attB-UAS14-gyp[3xP3-eYFP]에 삽입하기 위한 RNAi 구문 생성의 다이어그램 표현:(A) 단일 단계 비대칭 PCR(Xiao에서 적응) Y H 외 (2006) 및 (B) 다중 단계 융합 PCR. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
2. A. 감비아 푸파 스크리닝
- 현미경 특성화를 위한 강아지 컬렉션
참고: 이러한 프로토콜을 통해 물은 0.01%의 연못 소금으로 보충된 증류수를 말합니다.- 후방 표준 프로토콜(예: MR426)을 사용하여 푸팔 단계에 있는 감비아에 모기를 후면합니다.
주의: 이 과정에서 강아지를 다치게 하지 않도록 주의하십시오. - 입체 현미경 (예 : 100 x 15mm 플라스틱 페트리 접시, 가장자리를 피하기)와 함께 사용하기에 적합한 명확한 평평한 접시에 pupae를 수집합니다.
참고 : 강아지를 수집하기 위해 우리는 끝에서 약 10mm 절단 3 mL 플라스틱 파스퇴르 파이펫을 사용하여 끝을 넓히고 모기의 부상을 방지합니다. 스크리닝과 섹스는 개인에 완료 될 수 있습니다, 그러나, 이것은 매우 느립니다. 50-200 pupae의 그룹에 대한 선별 및 섹스를 수행하는 것이 좋습니다 (가능한 그룹의 크기는 사용되는 요리의 크기에 의해 제한되고 개인적인 취향의 대상이됩니다). 많은 수의 선별이 이루어지는 경우 먼저 pupae를 라인 깊숙이 약 4~5개 정렬하고 대상 새끼를 이 라인에서 이동시켜 효율성을 높일 수 있습니다. - 파스퇴르 파이펫을 사용하여 푸파 주변에서 거의 모든 물을 조심스럽게 제거하십시오. pupae 주위에 충분한 물을 남겨 두어 효과적으로 움직이지만 미세한 브러시로 쉽게 이동할 수 있습니다. 이동이 어려워지면 물을 더 추가합니다.
참고: 충분한 물이 제거되면, 강아지는 형광 검출 및 이형성 생식기의 식별을 위해 눈의 시각화를 허용하여 측면에 누워 있습니다 (그림 4DE).
주의: 푸파에가 담정하지 않도록 하십시오. 아주 소량의 물만 방치하면 현미경의 램프와 푸파풀 사이에 분할될 때 더 이상 감소시킬 수 있습니다. 원하는 그룹에 3mL 파스퇴르 파이펫을 사용하여 공정 중에 추가 물을 추가해야 합니다.
- 후방 표준 프로토콜(예: MR426)을 사용하여 푸팔 단계에 있는 감비아에 모기를 후면합니다.
- 푸파에 형광 마커의 식별
참고: 낮은 배율 입체 현미경의 사용은 넓은 필드 스크리닝을 허용, 정렬은 반전 된 화합물 현미경에 수행 할 수 있지만, 개별적으로 수행해야합니다.- 형광 마커를 검사 할 때 발현과 상속의 예상 패턴을 아는 것이 가장 중요합니다. 다음을 고려하십시오.
- 색상: 식을 시각화할 필터(들)를 결정합니다.
- 실자적 표현 패턴: 표현식을 볼 것으로 예상되는 위치와 삶의 단계에서 이해합니다.
- 다른 표현형의 비율: 관심있는 마커를 수행해야 하는 인구의 비율을 설정합니다.
- 저조도에서도 형광의 해결을 방해할 수 있기 때문에 어둠 속에서 형광 성 전염 검사를 실시합니다. 그러나 다른 조작에 조명이 필요할 때 입체 범위 옆에 램프를 사용합니다.
주의: 조명을 끄기 전에 형광 스테레오스코프 주변의 작업 영역이 명확하도록 합니다. - 형광 전구를 켜고 제조업체의 권장 기간 (일반적으로 10-15 분)을 따뜻하게 하십시오. 형광 성입체에 필요한 필터를 선택하고 스테이지 플레이트의 중앙을 향하는 빛의 색깔의 광선이 있는지 확인합니다. 이 보이지 않거나 매우 희미한 경우 형광 전구가 완전히 따뜻하지 않을 수 있습니다, 셔터가 닫히거나, 현미경 광학이 잘 정렬되지 않습니다.
- 백색광을 사용하여, 시야에서 강아지를 중심으로 초점을 맞춥니다. 형광 강도에 따라 다른 필터 사이를 전환할 때 이러한 배율을 변경해야 할 수도 있습니다.
- 미세한 디테일의 페인트 브러시를 사용하여 검사된 새끼가 겹치지 않도록 합니다.
- 스테레오스코프의 백색광을 끄고 미세 한 초점을 사용하여 관심표현형을 들고 있는 강아지의 영역을 집중시다. 형광 패턴을 표시해야합니다. 3xP3 프로모터 제어 형광의 예는 도 3에서 제공됩니다.
- 예상형 형광 표현형이 형광이 없는 개인과 안정적으로 구별될 수 있는 가장 낮은 배율을 사용합니다.
- 밝은 형광을 가진 긴장의 경우 형광 신호가 여전히 명확하게 식별 할 수있는 경우, 스크리닝하는 동안뿐만 아니라 낮은 강도 밝은 필드 빛을 사용합니다.
- 1차 심사를 마쳤을 때, 다른 필터 하에서 인구를 빠르게 스캔하여 잠재적인 오염을 감지합니다.
주의: pupae 운동에 의한 오염을 방지하기 위해 정렬된 강아지 그룹 사이에 명확한 거리가 있는지 확인하십시오. 강아지가 성별되고 배율 아래에서 볼 때 거리가 더 커질 수 있습니다. 풀이 시야 내에 없을 때는 각별한 주의를 기울여야 합니다.
- 형광 마커를 검사 할 때 발현과 상속의 예상 패턴을 아는 것이 가장 중요합니다. 다음을 고려하십시오.

도 3 - 3xP3 프로모터(A) eYFP, (B) dsRed 및 (C) eCFP에 의해 구동되는 형광 마커를 발현하는 아노펠스 감비아에. 배율: A=16X, B, C=20X.
-
섹스 푸파
- 강아지를 수집합니다. 여분의 물을 제거하지만 항문 패들이 생식기에서 약간 부분하여 시각화 및 형태학적 특성화를 돕도록 충분합니다(그림 4D, E).
- 어떤 pupa / e가 그들의 측면에 없는 경우, 부드럽게 강아지를 켜고 외부 생식기를 식별 할 수 있도록 항문 패들을 이동 미세 한 디테일 페인트 브러시를 사용합니다.
- 특유의 외부 생식기에 기초한 별도의 강아지; 남성은 최종 등쪽 세그먼트에서 항문 패들의 길이의 약 절반(그림 4B)에서 압출되는 긴 튜브를 가지고 있습니다. 여성 강아지의 외부 생식기는 상당히 짧고 양면 (그림 4A).
참고 : 때때로, 4 번째 인스타 애벌레 외골격이 부착된 상태로 남아 있거나 외부 생식기가 손상된 경우 (도 4C), 섹스의 자신감 식별이 더 어렵다. 강아지의 섹스가 명확하지 않을 때, 그것을 버리는 것이 가장 좋습니다. 개인이 유지되어야하는 경우, 강아지는 고립에서 등장할 수 있어야하고 성체는 성인 형태 학적 특징을 사용하여 결정해야합니다. 생식기가 손상되면 개인이 성공적으로 짝짓지 않을 수 있습니다. - 요리의 반대쪽 끝에 있는 각 성별에 대한 수영장을 만들어 성별되지 않은 수영장으로 만들고, 섬세한 디테일의 페인트 브러시를 사용하여 접시를 가로질러 식별된 강아지를 이동시십시오. 나중에 두 개의 수영장이 모이는 요리의 밑면에 레이블을 지정합니다.
- 섹스링과 형광 스크리닝이 모두 필요한 경우, 둘 중 더 빠른 과정이기 때문에 먼저 형광 검사를 수행하십시오.

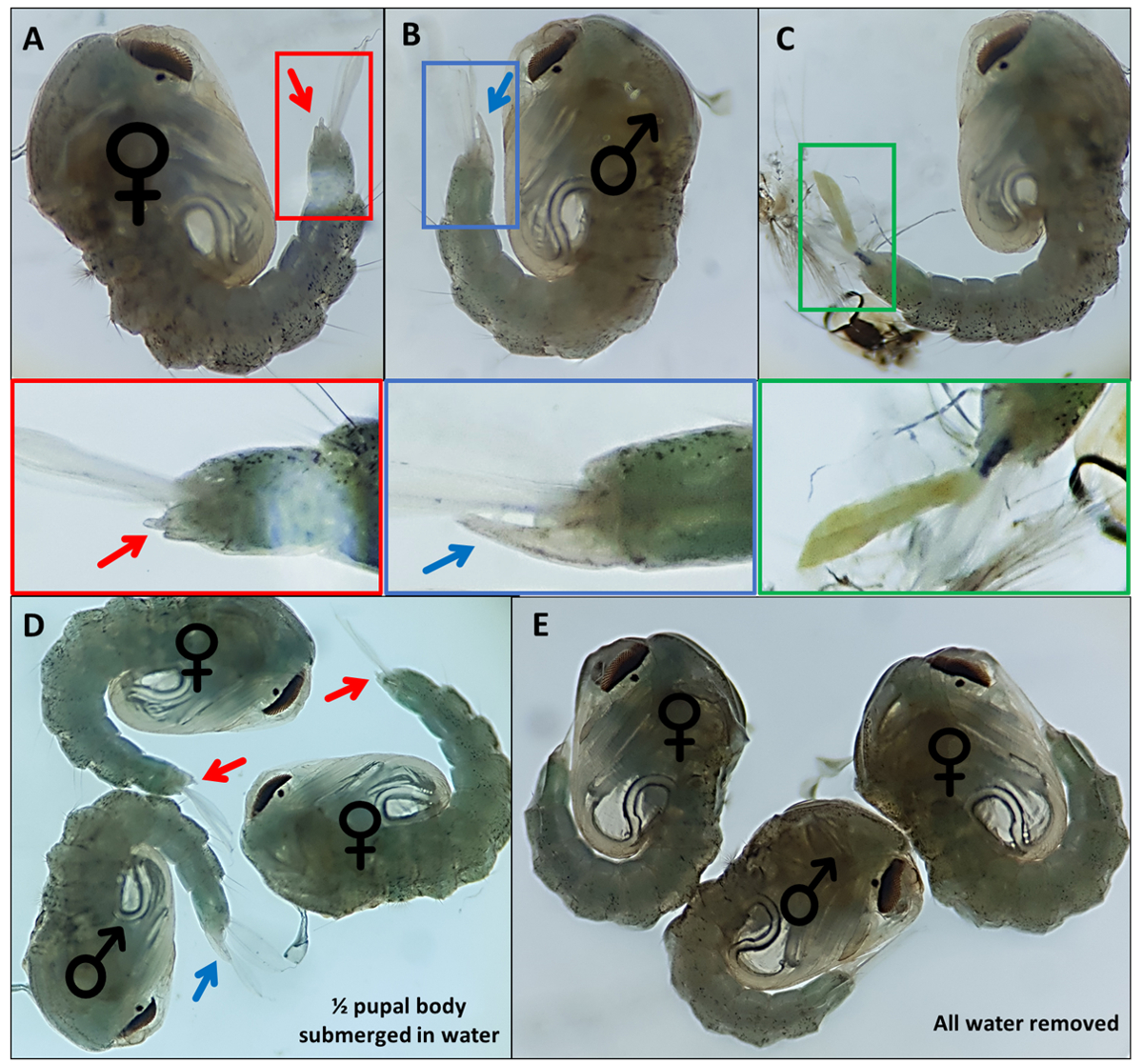
그림 4 - 섹스 아노펠스 감비아에 푸파. (A) 여성(B)의 외성 생식기를 나타내는 개별 푸파는 유충 외골격의 불완전한 분리로 인해 쉽게 식별 할 수없는 개인 (A) 여성 (B) 및 (C) 개인의 외성. 외부 생식기를 강조 아래 확대 된 이미지. 푸파(여성)와 ♂ (수컷)는 ♀ 푸파의 외발성기와 (D) ~50%의 강아지를 물에 담그고 (E) 모든 물을 제거하여 외부 생식기의 시각화의 용이성을 부각시키는 차이를 부각시한다. 배율: A, B, C=40x, D, E=30x. 여기를 클릭하여 이 그림의 더 큰 버전을 확인하십시오.
{kind=link}
- 성인으로 섹스 확인
- 매우 낮은 오류 율이 입증 될 때까지, 출현 후 성인 형태에 의해 pupae 섹스를 확인합니다. 몇 mL의 물로 투명한 20mL 튜브에서 10 mL 이하로 성별된 강아지를 분리하고, 면모 공으로 밀봉하고, 예상 된 섹스로 표시하고 하룻밤 사이에 등장 할 수 있습니다.
참고: 성인이 다음날 아침에 이송되기 때문에 신흥 성인에게 음식을 공급할 필요가 없습니다. - 다음 날 형태학적 특징을 사용하여 등장한 성인의 성별을 확인하십시오.
- 어떤 남성이 여성 컬렉션에 존재하는 경우, 짝짓기가 이미 발생한 경우, 여성을 폐기.
- 남성 컬렉션에 여성이 있는 경우 여성/s를 제거하고 횡단을 위해 남성을 유지하십시오.
- 매우 낮은 오류 율이 입증 될 때까지, 출현 후 성인 형태에 의해 pupae 섹스를 확인합니다. 몇 mL의 물로 투명한 20mL 튜브에서 10 mL 이하로 성별된 강아지를 분리하고, 면모 공으로 밀봉하고, 예상 된 섹스로 표시하고 하룻밤 사이에 등장 할 수 있습니다.
- GAL4-UAS 시스템 교차 설정
- 2.4단계에서 튜브에서 원하는 수의 남성과 여성 성인을 A. 감비아 사육의 표준 방식으로 설정된 케이지 또는 작은 양동이로 흡입합니다.
참고: 이 환승 기간 동안 성인에게 피해를 주지 않도록 주의하십시오. - ~2000 명의 성인이 자손에서 필요한 경우 동일한 수의 수의 암컷을 약 50 명의 여성을 사용하십시오.
참고: 십자가를 여러 번 공급하여 각 성별의 최대 200개 배치를 생성하는 경우 30cm x 30cm x 30cm 케이지에 설치될 수 있습니다. 소수의 여성(<20)만 십자가에 사용할 수 있는 경우, 성공적인 짝짓기의 가능성을 높이기 위해 수컷의 수 ~4배를 추가합니다. - 혈액은 표준 프로토콜26에 따라 교차된 여성과 후면 자손에게 적절한 단계로 공급하여 표면 평가(예를 들어, 살충제 저항, 벡터 용량 및 피트니스 비용 분석)를 수행합니다.
- 트랜스진 발현의 모성 효과 가능성이 있는 경우, 운전자와 응답자 라인의 상호 십자가를 설정하고 분석 예상 표현형을 설정합니다.
참고: '이종화구스'를 사용하거나 운전자와 응답자 라인의 혼합 된 인구를 사용하여 교차, 4 가능한 유전자형 각각의 자성을 생성합니다. 이것은 야생 유형, UAS 만 및 GAL4 만 컨트롤을 제공하며 표현형을 분석할 수있는 transheterozygotes GAL4-UAS를 제공합니다. 동종구균 개체수가 교차하는 경우 추가 십자가를 설정하여 표현형을 비교하는 적절한 컨트롤을 제공합니다. 자손은 표면 평가를 위해 마커뿐만 아니라 네거티브를 모두 또는 만 운반하는 자손을 분리하는 것으로 선별되어야합니다.
- 2.4단계에서 튜브에서 원하는 수의 남성과 여성 성인을 A. 감비아 사육의 표준 방식으로 설정된 케이지 또는 작은 양동이로 흡입합니다.
- 대체 형광 마커를 운반하는 RCME를 통해 생성된 라인에서 균질계 인구 확립
참고: 두 선의 형광 마커가 동일한 게놈 위치에 존재하며 완전히 구별되는 것이 필수적입니다.- 위에서 설명한 바와 같이 올바른 형광과 성별을 표시하는 개인을 선택하기 위해 심사 다음 다른 라인의 한 줄과 여성의 동일한 수의 분별 표시 된 남성의 동등한 수를 가진 약 200 명의 성인의 부모 십자가를 설정합니다. 약 1 주일 후에 혈액공급은 확립된 프로토콜s26을 사용하여 십자가를 공급합니다.
- F1 자손은 표준 프로토콜을 사용하여 강아지를 하고 이전에 설명한 바와 같이 pupae를 수집합니다.
- 양 부모 마커 (transheterozygous)를 모두 운반하는 사람들을 선택하는 형광을위한 화면. 이 강아지와 F1 크로스를 설정합니다.
- 1 주일 후, 혈액은 표준 프로토콜에 따라 pupal 단계에 F1 여성과 후방 자손피드.
- 마커 중 하나만 표시하는 P2 pupae를 선택합니다. 이들은 삽입을 위한 동질이 될 것입니다. 자손의 25%만이 각 삽입에 대해 동질이 되므로 충분한 자손이 사육되어 스톡 케이지(400-500)를 제공해야 합니다.
참고: 트랜게테로지구스 자손의 선택은 프로세스가 오염되고 완전한 동질성이 달성되지 않을 수 있습니다 그렇지 않으면 완전히 엄격해야합니다. F1 크로스에 대해 선택한 모든 자손을 다시 확인합니다.
- 위에서 설명한 바와 같이 올바른 형광과 성별을 표시하는 개인을 선택하기 위해 심사 다음 다른 라인의 한 줄과 여성의 동일한 수의 분별 표시 된 남성의 동등한 수를 가진 약 200 명의 성인의 부모 십자가를 설정합니다. 약 1 주일 후에 혈액공급은 확립된 프로토콜s26을 사용하여 십자가를 공급합니다.
3. A. 감비아아배아 클리어링 프로토콜
-
혈액 공급 및 유지 보수
- 표준 프로토콜(예: MR4)을 따르는 성인에게 Gambiae 모기를 후면합니다.
- 혈액 공급 5-7 일 오래 된 여성 성인, 대부분의 완전히 유혹 보장.
주의: 이 프로토콜 을 통해 신속하게 작동하면 계란이 담정할 수 없도록 하는 것이 필수적입니다.
-
유도 된 계란 누워
- 혈액 먹이 후 3 일 유도 누워를 통해 계란을 수집합니다.
- oviposition 챔버를 조립합니다.
- 약 5mm깊이로 물로 oviposition 냄비를 채웁니다. 50 mL 폴리 프로필렌 튜브의 한쪽 끝에 냄비를 부착, 이전에 두 끝이 열려 있도록 쇠톱으로 잘라. (우리는 냄비에 플라스틱 디스크를 사용 (그림 5); 그러나, 튜브의 원래 뚜껑대신 사용할 수 있습니다).
- 컷 폴리 프로필렌 튜브의 다른 쪽 끝을 재료 (호스 / 스타킹) 또는 탄성 밴드로 고정 라텍스 장갑의 섹션으로 커버, 그래서 성인을 소개 할 수 있지만 탈출 할 수 없습니다 (그림 5). 다른 대체 oviposition 챔버 설계 존재 하 고 사용할 수 있습니다26.
- 조심스럽게 oviposition 챔버에 10-15 여성 (단계 3.1.2에서 먹이 혈액)을 소개합니다. 암흑을 일으키고 20 분 동안 떠나도록 oviposition 챔버를 덮습니다.
주의: 계란이 계란의 좌초와 탈수를 방지하기 위해 누워일단 oviposition 냄비를 이동하지 마십시오. - 모기를 방출하지 않도록 하면서 50mL 폴리 프로필렌 튜브를 oviposition 냄비에서 조심스럽게 분리합니다. 흰 알을 볼 수 있어야 합니다. 규정된 목적을 위해 충분한 것이 배치되었는지 확인합니다. 필요한 경우 반복합니다.
- 냄비를 덮고 (먼지 보호를 위해) 계란이 관심의 발달 단계로 성숙 할 수 있도록합니다.
- 미세한 디테일의 페인트 브러시를 사용하여 냄비에서 계란을 집어 40mm2 굴착 유리 블록에 물에 놓습니다.

그림 5 - 조립된 구성요소및 (B)를 강조하기 위해 해체된 오비포지션 챔버(A)의 예. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
-
배아 고정
주의: 포름알데히드 사용으로 인해 연기 후드에서 모든 고정 단계(3.3단계)를 수행합니다.- Kaiser 외(2014)22에 설명된 바와 같이 FAA 솔루션을 준비하십시오. FAA는 3.6M 포름알데히드, 0.87M 아세트산, 8.5M 절대 에탄올로 이루어져 증류수(dH2O)로 부피까지 구성된다.
- FAA 10mL의 경우 13.42M 포름알데히드2.68mL, 17.14M 에탄올4mL, 17.4m 아세트산 0.5mL, 증류H2O 1.86mL의 1.86mL를 결합하여 단단히 밀봉된 유리 용기에 보관할 수 있습니다.
- 유리 블록에서 마이크로피펫으로 물을 조심스럽게 제거하고 FAA의 500 μL에서 계란을 덮고 실온에서 궤도 셰이커에서 부드럽게 진동 (~25 RPM)을 30 분 동안 진동시. 이 시점에서 색상 변경이 표시되지 않습니다.
- 증류수로 계란을 완전히 헹구세요. 포름알데히드의 모든 흔적을 제거하기 위해 15 번 헹도 을 수행합니다. 1000 μL 마이크로파이프를 사용하여 한 번에 1mL의 dH2O를 추가한 다음 제거하여 알을 손상시키지 않도록 합니다.
- 안전 지침에 따라 폐기를 위해 지정된 포름알데히드 폐기 용기에 헹구는 폐수를 저장합니다.
- 이 시점에서 고정 된 계란은 수분을 유지하기 위해 물에 하룻밤 4 °C에서 저장 할 수 있습니다.
- Kaiser 외(2014)22에 설명된 바와 같이 FAA 솔루션을 준비하십시오. FAA는 3.6M 포름알데히드, 0.87M 아세트산, 8.5M 절대 에탄올로 이루어져 증류수(dH2O)로 부피까지 구성된다.
-
배아 표백
주의: 하이포염 나트륨과 아세트산이 결합될 때 염소 가스의 잠재적 방출로 인해 연기 후드에서 모든 표백 단계(4단계)를 수행합니다.- 표백 용액준비(Trpiš 솔루션 - Trpiš(1970)21에 설명되어 있으며 카이저 외(2014)22)에 따라 수정됩니다. Trpiš 용액은 0.59 M 나트륨 하이포염및 0.35 M 아세트산이 증류된 H2O에 용해된다.
- Trpiš 용액의 10mL ovolume의 경우 2.2M 의 나트륨 hypochlorite의 2.68 mL과 17.4 M 아세트산의 0.2 mL을 증류 된 H2O의 7.12 mL로 결합하십시오.
참고: Trpiš 용액은 단단히 밀봉된 유리 용기에 3개월 이상 보관하고 안전한 화학 찬장에 보관할 수 있습니다. 용액은 저장 후 소용돌이해야 할 수 있으며 염소 가스가 방출될 경우 항상 연기 후드에 열어야 합니다.
- Trpiš 용액의 10mL ovolume의 경우 2.2M 의 나트륨 hypochlorite의 2.68 mL과 17.4 M 아세트산의 0.2 mL을 증류 된 H2O의 7.12 mL로 결합하십시오.
- 고정 된 계란을 Trpiš 용액 1 mL로 덮고 실온에서 30 분 동안 배양하십시오. 계란은 잠복의 약 5 분 후에 창백한 패치를 개발하기 시작하고, 결국 한 번 지워진 유백색에 도달합니다.
- 트르피시 용액을 제거하기 위해 3.3.3 단계와 같이 계란을 헹구세요.
- 지정된 폐수에 폐수를 저장하고 배수구 아래로 여분의 물로 폐기하십시오.
- 표백 용액준비(Trpiš 솔루션 - Trpiš(1970)21에 설명되어 있으며 카이저 외(2014)22)에 따라 수정됩니다. Trpiš 용액은 0.59 M 나트륨 하이포염및 0.35 M 아세트산이 증류된 H2O에 용해된다.
-
보관
- dH2O의 500 μL에 저장하고 며칠 동안 2-8 °C 사이에 보관하십시오. 질량에 보고 이미징하기 전에 물 대부분을 주의 깊게 제거하지만 시계 유리에 소량의 물을 남겨 계란의 건조를 피하십시오. 이것은 계란 촬영을 방해하지 않습니다. 개별 계란은 더 높은 배율 화상 진찰을 위한 현미경 슬라이드에 놓일 수 있습니다.
결과
eYFP, dsRed 및 eCFP의 3xP3 발현은 안감비아푸에의 눈과 복부 신경리아에서 발현을 생성하는 마커 유전자를 소유한 개인의 신뢰할 수 있고 쉽게 구별할 수 있는 식별을 제공한다(도 3). 성별에 사용되는 남성 및 여성 외성 생식기에서 관찰되는 차동 형태와 식별할 수 없는 pupae의 예는 도 4에서 강조된다. 항문 패들이 생식기의 시각화를 모호하?...
토론
모기 유전자 기능을 이해하는 것은 Anopheles를 통제하고 말라리아 전송에 영향을 미치기 위하여 새로운 접근을 개발하는 것이 중요합니다. 설명된 GAL4-UAS 시스템은 후보 유전자의 기능적 분석을 위한 다재다능하고 강력한 시스템으로 현재까지 우리는 살충제 저항17 및 큐티큘러 탄화수소 생산의 유전적 기초를 검사하고, 형광태?...
공개
저자는 공개 할 것이 없습니다.
감사의 말
우리는 감사로 LSTM과 IVCC (아드리아나 아돌피), BBSRC (새로운 조사자 상 (AL), MRC (BCP에 박사 학생 교직 박사 학생: MR / P016197/1), 웰컴 (경 헨리 웰컴 박사 후 박사 과정 펠로우십 LG: 215894/ Z/ 19/Z) 그 제안에 Gal4UAS 분석을 통합했습니다.
자료
| Name | Company | Catalog Number | Comments |
| 100 x 15 mm plastic Petri dish | SLS | 2175546 | Pack of 10 |
| 1000 µL Gilson Pipette | Gilson | F144059P | |
| 20/25 mL Universal Tubes | Starlab | E1412-3020 | Pack of 400 |
| 3 mL Pasteur Pipettes | SLS | G612398 | Greiner Pasteur pipette 3 mL sterile individually wrapped |
| 50 mL Falcon Tubes | Fisher Scientific | 11512303 | |
| Absolute Ethanol | Fisher Scientific | BP2818-500 | 500 mL |
| Acetic Acid | SLS | 45726-1L-F | 1 L |
| Cages | SLS | E6099 | 30x30x30 with screen port |
| Fine Paint Brushes | Amazon | UKDPB66 | KOLAMOON 9 Pieces Detail Painting Brush Set Miniture Brushes for Watercolor, Acrylic Painting, Oil Painting (Wine Red) |
| Fish food | Amazon | Tetra Min Fish Food, Complete Food for All Tropical Fish for Health, Colour and Vitality, 10 L | |
| Formaldehyde Solution | Sigma Aldrich | F8775 | |
| Mouth Aspirator | John Hock | 612 | |
| Pond Salt | Amazon | Blagdon Guardian Pond Tonic Salt, for Fish Health, Water Quality, General Tonic, pH Buffer, 9.08 kg, treats 9,092 L | |
| Pupae Pots | Cater4you | SP8OZ | 250 pots with lids |
| Small Plastic Buckets | Amazon | 2.5 L White Plastic Pail Complete with White Lid (Pack of 10) | |
| Sodium Hypochlorite | Fisher Scientific | S25552 |
참고문헌
- Brand, A. H., Perimon, N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development. 118 (2), 401-415 (1993).
- Duffy, J. B. GAL4 system in drosophila: A fly geneticist's swiss army knife. Journal of Genetics and Development. 34 (1-2), 1-15 (2002).
- Dow, J. A. . ELS. , (2012).
- Edi, C. V., et al. CYP6 P450 Enzymes and ACE-1 Duplication Produce Extreme and Multiple Insecticide Resistance in the Malaria Mosquito Anopheles gambiae. PLoS Genetics. 10 (3), 1004236 (2014).
- Daborn, P. J., et al. Using Drosophila melanogaster to validate metabolism-based insecticide resistance from insect pests. Insect Biochemistry and Molecular Biology. 42 (12), 918-924 (2012).
- Riveron, J. M., et al. Genome-wide transcription and functional analyses reveal heterogeneous molecular mechanisms driving pyrethroids resistance in the major malaria vector Anopheles funestus across Africa. Genes Genomes Genetics. 7 (6), 1819-1832 (2017).
- Riveron, J. M., et al. A single mutation in the GSTe2 gene allows tracking of metabolically based insecticide resistance in a major malaria vector. Genome Biology. 15 (2), (2014).
- Lynd, A., Lycett, G. J. Development of the Bi-Partite Gal4-UAS System in the African Malaria Mosquito, Anopheles gambiae. PLoS ONE. 7 (2), 31552 (2012).
- Lynd, A., Lycett, G. J. Optimization of the Gal4-UAS system in an Anopheles gambiae cell line. Insect Molecular Biology. 20 (5), 599-608 (2011).
- Adolfi, A., Pondeville, E., Lynd, A., Bourgouin, C., Lycett, G. J. Multi-tissue GAL4-mediated gene expression in all Anopheles gambiae life stages using an endogenous polyubiquitin promoter. Insect Biochemistry and Molecular Biology. 96, 1-9 (2018).
- Kokoza, V. A., Raikhel, A. A. Targeted gene expression in the transgenic Aedes aegypti using the binary Gal4-UAS system. Insect Biochemistry and Molecular Biology. 41, 637-644 (2011).
- O'Brochta, D. A., Pilitt, K. L., Harrell, R. A., Aluvihare, C., Alford, R. T. Gal4-based Enhancer-Trapping in the Malaria Mosquito Anopheles stephensi. Genes Genomes Genetics. 2, 21305-21315 (2012).
- Zhao, B., et al. Regulation of the Gut-Specific Carboxypeptidase: A Study Using the Binary Gal4/UAS System in the Mosquito Aedes Aegypti. Insect Biochemistry and Molecular Biology. 54, 1-10 (2014).
- Imamura, M., et al. Targeted Gene Expression Using the GAL4/UAS System in the Silkworm Bombyx mori. Genetics. 165 (3), 1329-1340 (2003).
- Lynd, A., et al. Development of a functional genetic tool for Anopheles gambiae oenocyte characterisation: application to cuticular hydrocarbon synthesis. bioRxiv. , (2019).
- Pondeville, E., et al. Hemocyte-targeted gene expression in the female malaria mosquito using the hemolectin promoter from Drosophila. Insect Biochemistry and Molecular Biology. 120, 103339 (2020).
- Adolfi, A., et al. Functional genetic validation of key genes conferring insecticide resistance in the major African malaria vector, Anopheles gambiae. Proceedings of the National Academy of Sciences of the United States of America. 116 (51), 25764-25772 (2019).
- Pondeville, E., et al. Efficient integrase-mediated site-specific germline transformation of Anopheles gambiae. Nature Protocols. 9 (7), 1698-1712 (2014).
- Horn, C., Schmid, B. G. M., Pogoda, F. S., Wimmer, E. A. Fluorescent transformation markers for insect transgenesis. Insect Biochemistry and Molecular Biology. 32, 1221-1235 (2002).
- Clements, A. . A. Biology of Mosquitoes, Volume 1: Development, Nutrition and Reproduction. 1, (1992).
- Trpiš, M. A new bleaching and decalcifying method for general use in zoology. Canadian Journal of Zoology. 48, 892-893 (1970).
- Kaiser, M. L., Duncan, F. D., Brooke, B. D. Embryonic Development and Rates of Metabolic Activity in Early and Late Hatching Eggs of the Major Malaria Vector Anopheles gambiae. PLoS ONE. 9 (12), 114381 (2014).
- Grigoraki, L., Grau-Bové, X., Yates, H. C., Lycett, G. J., Ranson, H. Isolation and transcriptomic analysis of Anopheles gambiae oenocytes enables the delineation of hydrocarbon biosynthesis. eLife. 9, 58019 (2020).
- Xiao, Y. -. H., Yin, M. -. H., Hou, L., Pei, Y. Direct amplification of intron-containing hairpin RNA construct from genomic DNA. BioTechniques. 41 (5), 548-552 (2006).
- Livak, K. J. Organization and Mapping of a Sequence on the Drosophila melanogaster X and Y Chromosomes That Is Transcribed during Spermatogenesis. Genetics. 107 (4), 611-634 (1984).
- MR4, CDC, NEI & beiResources. . The MR4 Methods in Anopheles Research Laboratory Manual. 5th Edition. , (2015).
- Sik Lee, Y., Carthew, R. W. Making a better RNAi vector for Drosophila: use of intron spacers. Methods. 30 (4), 322-329 (2003).
- Cha-aim, K., Hoshida, H., Fukunaga, T., Akada, R., Peccoud, J. . Gene Synthesis: Methods and Protocols. , 97-110 (2012).
- Cavener, D. R. Comparison of the consensus sequence flanking translational start sites in Drosophila and vertebrates. Nucleic Acids Research. 15 (4), 1353-1361 (1987).
- Wang, Y., Wang, F., Wang, R., Zhao, P., Xia, Q. 2A self-cleaving peptide-based multi-gene expression system in the silkworm Bombyx mori. Scientific Reports. 5, (2015).
- Galizi, R., et al. A synthetic sex ratio distortion system for the control of the human malaria mosquito. Nature Communications. 5, 3977 (2014).
- Kondo, S., et al. Neurochemical organisation of the Drosophila Brain Visualised by Endogenously Tagged Neurotransmitter Receptors. Cell Reports. 30 (1), 284-297 (2020).
- Lee, P. -. T., et al. A gene-specific T2A-GAL4 library for Drosophila. eLife. 7, 35574 (2018).
- Marois, E., et al. High-throughput sorting of mosquito larvae for laboratory studies and for future vector control interventions. Malaria Journal. 11, 302 (2012).
- Crawford, J. E., et al. Efficient production of male Wolbachia-infected Aedes aegypti mosquitoes enables large-scale suppression of wild populations. Nature Biotechnology. 38 (4), 482-492 (2020).
- Goltsev, Y., et al. Developmental and evolutionary basis for drought tolerance of the Anopheles gambiae embryo. Developmental Biology. 330 (2), 462-470 (2009).
- Rezende, G. L., et al. Embryonic desiccation resistance in Aedes aegypti: presumptive role of the chitinized Serosal Cuticle. BMC Developmental Biology. 8 (1), 82 (2008).
- Vargas, H. C. M., Farnesi, L. C., Martins, A. J., Valle, D., Rezende, G. L. Serosal cuticle formation and distinct degrees of desiccation resistance in embryos of the mosquito vectors Aedes aegypti, Anopheles aquasalis and Culex quinquefasciatus. Journal of Insect Physiology. 62, 54-60 (2014).
- Chang, C. -. H., et al. The non-canonical Notch signaling is essential for the control of fertility in Aedes aegypti. PLOS Neglected Tropical Diseases. 12 (3), 0006307 (2018).
- Clemons, A., Flannery, E., Kast, K., Severson, D., Duman-Scheel, M. Immunohistochemical Analysis of Protein Expression during Aedes aegypti Development. Spring Harbor Protocols. 10, 1-4 (2010).
- Juhn, J., James, A. A. Hybridization in situ of Salivary Glands, Ovaries and Embryos of Vector Mosquitoes. Journal of Visualized Experiments. , e3709 (2012).
재인쇄 및 허가
JoVE'article의 텍스트 или 그림을 다시 사용하시려면 허가 살펴보기
허가 살펴보기This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. 판권 소유