
需要订阅 JoVE 才能查看此. 登录或开始免费试用。
Method Article
通过显微注射在类 圆线虫 物种中产生转基因和敲除
摘要
寄生线虫 类圆线 虫粪类和大 鼠类圆线 虫的功能基因组工具包包括转基因、CRISPR/Cas9 介导的诱变和 RNAi。该协议将演示如何使用性腺内显微注射将转基因和CRISPR成分引入 粪类猪笼草 和 鼠李糖杆菌中。
摘要
类圆线虫属由多种具有不同宿主范围的穿透皮肤线虫组成,包括粪类圆线虫和大鼠类圆线虫。S. stercoralis是一种人类寄生虫,穿透皮肤的线虫,感染约6.1亿人,而大鼠寄生虫S. ratti与粪类S. stercoralis密切相关,通常用作粪类菌丝虫的实验室模型。S. stercoralis和S. ratti都很容易通过性内显微注射的外源核酸递送技术产生转基因和敲除,因此,已经成为其他尚未适应该技术的寄生蠕虫的模型系统。
寄生 类圆线虫 成虫栖息在宿主的小肠中,并通过粪便将后代释放到环境中。一旦进入环境,幼虫就会发育成自由生活的成虫,它们生活在粪便中并产生必须找到并入侵新宿主的后代。这种环境生成是 类圆线虫 物种所独有的,在形态上与自由生活线虫 秀丽隐杆线虫 模型足够相似,为 秀丽隐杆线虫 开发的技术可以适应与这些寄生线虫一起使用,包括性腺内显微注射。使用性腺内显微注射,可以将各种各样的转基因引入类 圆线虫中。CRISPR / Cas9组分也可以显微注射以产生突变 的圆线虫 幼虫。这里,描述了性腺内显微注射到 类圆线虫的技术, 包括自由生活成人的制备,注射程序和转基因后代的选择。包括使用CRISPR / Cas9诱变产生的转基因 类圆线虫 幼虫的图像。本文的目的是使其他研究人员能够使用显微注射来产生转基因和突变 的类圆线虫。
引言
与更广泛认可的钩虫和蛔虫蛔虫相比,类粪类圆线虫长期以来一直被忽视为一种重要的人类病原体。以前对蠕虫负担的研究往往严重低估了粪类链球菌的患病率,因为粪类疟原虫的常见诊断方法的敏感性较低 2.近年来,基于改进的诊断工具的流行病学研究估计,粪类菌感染的真实患病率远高于先前报告的,全世界约有6.1亿人2。
S. stercoralis和其他类圆线虫物种,包括密切相关的大鼠寄生虫和常见的实验室模型S. ratti,都有一个不寻常的生命周期,对实验基因组研究有利,因为它由寄生和自由生活(环境)第3代组成(图1)。具体来说,S. stercoralis和S. ratti都可以通过单一的自由生活世代循环。自由生活的一代由后寄生虫幼虫组成,这些幼虫发育成自由生活的成年雄性和雌性;自由生活的成虫的所有后代都发育成感染性幼虫,必须感染宿主才能继续生命周期。此外,这种环境或自由生活的一代可以在实验室中通过实验操纵。由于自由生活的类圆线虫成虫和秀丽隐杆线虫成虫具有相似的形态,因此最初为秀丽隐杆线虫开发的性腺内显微注射等技术可以适用于自由生活的成年类圆线虫4,5。虽然DNA通常被引入自由生活的成年雌性中,但类圆线虫的雄性和雌性都可以被显微注射6。因此,功能基因组工具可用于询问类圆线虫生物学的许多方面。其他寄生线虫缺乏自由生活的一代,因此,不容易适应功能基因组技术3。

图1:类圆线虫的粪类动物生命周期。 粪类孢子虫寄生雌性栖息在其哺乳动物宿主(人类,非人类灵长类动物,狗)的小肠中。寄生雌性通过单性生殖繁殖,并在小肠内产卵。卵在宿主体内孵化成后寄生幼虫,然后与粪便一起进入环境。如果寄生后幼虫是雄性,它们会发育成自由生活的成年雄性。如果寄生后幼虫是雌性,它们可以发育成自由生活的成年雌性(间接发育)或第三阶段感染性幼虫(iL3s;直接发育)。自由生活的雄性和雌性有性繁殖以产生被限制成为iL3的后代。在某些情况下,粪类杀伤性链球菌也可以进行自身感染,其中一些后寄生幼虫留在宿主肠道内,而不是通过粪便进入环境。这些幼虫可以在宿主体内发育成自身感染性幼虫(L3a),穿透肠壁,通过身体迁移,最终返回肠道成为生殖成虫。S. ratti的生命周期是相似的,除了S. ratti感染大鼠并且没有自身感染周期。环境生成是使用类圆线虫物种进行遗传研究的关键。自由生活的成年雌性(P0)可以显微注射;它们的后代,全部将成为iL3s,是潜在的F1转基因。这个数字是从Castelletto等人修改而来的。3. 请按此浏览此图的大图。
{kind=link}
S. stercoralis 与其他胃肠道人寄生虫线虫共享其生物学的许多方面,包括宿主入侵和宿主免疫调节。例如, Necator 属和 Ancylostoma 属的人寄生虫钩虫也通过皮肤渗透感染,以类似的方式穿过身体,最终作为寄生成虫居住在小肠7中。因此,许多胃肠道线虫可能使用常见的感觉行为和免疫规避技术。因此,从 类圆线虫 中收集的知识将补充其他遗传上不太易处理的线虫的发现,并导致对这些复杂而重要的寄生虫有更完整的了解。
该显微注射方案概述了将DNA引入圆 线虫 自由生活成年雌性以制造转基因和突变后代的方法。描述了菌株维持要求,包括用于显微注射的成虫的发育时间和转基因后代的收集。包括实验方案和完整的显微注射技术演示,以及培养和筛选转基因后代的方案,以及所有必要设备和消耗品的清单。
Access restricted. Please log in or start a trial to view this content.
研究方案
注意:沙鼠用于通过 S. stercoralis,大鼠用于通过 S. ratti。所有程序均由加州大学洛杉矶分校动物研究监督办公室(协议编号2011-060-21A)批准,该办公室符合AAALAC标准和实验室动物护理和使用指南。以下任务必须在显微注射前至少一天完成:蠕虫培养,制备显微注射垫,为显微注射混合物创建结构,以及将细菌(大肠杆菌 HB101)扩散到6厘米线虫生长培养基(NGM)平板8上。自由生活的雌性需要在25°C下收集粪便后至少24小时才能发育成年轻人,然后才能进行显微注射。显微注射垫必须完全干燥。细菌板必须干燥并建立小草坪。
1.显微注射载玻片的制备:注射前至少一天
注意:蠕虫安装在带有干燥琼脂垫的显微注射盖玻片上。
- 将加热块设置为90°C。
- 加入5mL ddH2O,然后将100mg琼脂糖加入硼硅酸盐玻璃管中。
- 在火焰上加热管中的琼脂糖混合物,直到琼脂糖溶解。
- 将管置于90°C的加热块中,以保持琼脂糖处于液态。
- 使用玻璃巴斯德移液管或带有塑料吸头的移液管将约180μL琼脂糖溶液滴到盖玻片上。立即在上面放上第二个盖玻片,将琼脂糖压平成薄垫。
- 5-10秒后,通过将顶部盖玻片分开来取下两个盖玻片。确定琼脂垫在哪个滑块上,并正面朝上放置。
- 从破碎的盖玻片中选择一小块玻璃碎片,然后用镊子轻轻将其按入垫子顶部边缘附近的琼脂中(补充图S1)。
- 继续用琼脂糖溶液制作显微注射垫。
- 在工作台上或在烤箱中将琼脂糖垫干燥过夜。存放在盖玻片盒中。
注意:琼脂糖垫可以使用长达2个月,但仅用于一次注射。
2.培养 类圆线虫 以获得用于显微注射的蠕虫:注射前1 - 2天
注意:菌株维持方案可以在 补充材料中找到,其中包括如何用线虫感染沙鼠和大鼠以及从受感染动物的粪便中收获线虫的详细说明。
- 注射前两天,将受感染的动物9,10 放在收集笼中过夜。
- 第二天早上,收集受感染的粪便并制作粪便炭板9,10。
- 将板在25°C下放置24小时,以使自由生活的蠕虫发育成年轻人。
- 注射日的前一天晚上,将未感染的宿主动物放在收集笼中。
- 在注射当天,收集未感染的粪便进行注射后培养。
3.制作显微注射混合物:注射前或注射当天
注意:显微注射混合物由在蠕虫缓冲盐水(BU)(50 mM Na 2 HPO 4,22mM KH 2PO4,70 mM NaCl)11中稀释至所需浓度的感兴趣质粒组成。
- 确定质粒储备的浓度和显微注射混合物中的所需浓度(表1)。
| 显微注射组合:报告器构造 | |||
| 元件 | 库存集中度 | 量 | 最终浓度 |
| pMLC30 gpa-3::gfp | 300 纳克/微升 | 1.7 微升 | 50 纳克/微升 |
| 埠 | 那 | 8.3 微升 | 那 |
| 总 | 10 微升 | 50 纳克/微升 | |
| 显微注射混合:CRISPR/Cas9 诱变 | |||
| 元件 | 库存集中度 | 量 | 最终浓度 |
| pMLC47 tax-4 sgRNA | 300 纳克/微升 | 2.7 微升 | 80 纳克/微升 |
| pEY11 Ss-tax-4 HDR 质粒 | 400 纳克/微升 | 2.0 微升 | 80 纳克/微升 |
| pPV540 strCas9 质粒 | 350 纳克/μL | 1.1 微升 | 40 纳克/微升 |
| 埠 | 那 | 4.2 微升 | 那 |
| 总 | 10 微升 | 200 纳克/微升 | |
| 显微注射组合:小猪Bac集成 | |||
| 元件 | 库存集中度 | 量 | 最终浓度 |
| pMLC30 gpa3::gfp | 300 纳克/微升 | 2.0 微升 | 60 纳克/微升 |
| pPV402 转座酶质粒 | 450 纳克/μL | 0.9 微升 | 40 纳克/微升 |
| 埠 | 那 | 7.1 微升 | 那 |
| 总 | 10 微升 | 100 纳克/微升 | |
表1:显微注射混合物的示例。 三个示例显微注射混合物的质粒和浓度:一个用于 gpa-3::GFP 报告器构建10,一个用于CRISPR / Cas9介导的 Ss-tax-4 位点14,15的破坏,一个用于小猪Bac介导的 Ss-gpa-3::GFP 构建13,17,18的整合。 strCas9 表示 类圆线虫 密码子优化的Cas9基因。列出的最终浓度通常用于类 圆线虫 显微注射混合物。
- 将BU中的质粒稀释至总体积为10-20μL。
- 将混合物通过5,000× g 的过滤柱旋转1-2分钟。
- 立即使用显微注射混合物或将其储存在-20°C以备将来使用。
4.收集年轻成人 类圆线虫 进行显微注射:注射当天的早晨
- 用1个年轻成年 圆线虫 的粪便炭板设置Baermann装置(图2)。
注意:粪便炭板可能包含一些感染性幼虫。个人防护装备包括实验室外套、手套和护目镜。手套和实验室外套套套套之间不应暴露任何皮肤。

图2:用于从培养物中收集寄生虫的Baermann装置10。 粪便炭板的内容物放置在温水柱的顶部。蠕虫迁移到水中并聚集在漏斗的底部。(A)为了设置贝尔曼装置,用C型夹钳将贝尔曼漏斗的支架夹在工作台上。用夹钳将连接到漏斗末端的橡胶管封闭,并在管子下方放置一个收集桶以进行滴注。将温水添加到玻璃漏斗中。(B)然后将用于粪便 - 木炭混合物的塑料环支架衬里3片实验室组织(左)。木棍或压舌器(中间)用于将粪便炭板(右)的内容物转移到塑料环支架中。(C)塑料环支架底部的特写,用于粪便 - 木炭混合物,显示支架底部的双层尼龙薄纱衬里。(D)然后将粪便炭架放置在玻璃漏斗的顶部。(E)用水蘸湿实验室组织,并封闭在粪便 - 木炭混合物上。添加更多的温水,以大部分淹没粪便 - 木炭。(F)完整的贝尔曼设置,将粪便 - 木炭培养物浸没在温水中。 请点击此处查看此图的大图。
{kind=link}
- 使用O形圈在环形支架上安装带有橡胶收集管的玻璃漏斗,并用夹具固定。用2个夹钳关闭收集管(图2A)。
- 在漏斗下方放置一个捕获桶以捕获滴水。
- 将温水(约40°C)加入漏斗中,使其在边缘下方5厘米处。验证系统是否未泄漏。
- 将贝尔曼支架排成一行,这是一个由2个塑料环制成的筛子,它们之间固定着2层尼龙薄纱网,并带有3个重叠的实验室组织。将粪便 - 木炭混合物加入Baermann支架(图2B,C)。
- 将贝尔曼支架与粪便 - 木炭混合物一起放入漏斗中。折叠粪便 - 木炭混合物周围的组织,并加入足够的水以淹没大部分粪便 - 木炭。不要在距离漏斗边缘2厘米以上的地方填充(图2D,E)。
- 在漏斗顶部加一个15厘米的塑料培养皿盖,以容纳气味。根据需要标记漏斗(图2F)。
- 等待30分钟至1小时,从Baermann设备中收集蠕虫。
- 将一个 50 mL 离心管放在漏斗底部的橡胶管下。小心地打开底部的夹子,将 30-40 mL 含有蠕虫的水分配到 50 mL 试管中。
- 将 15 mL 含有蠕虫的 Baermann 水转移到 15 mL 离心管中。以〜750× g (缓慢)旋转15mL离心管1分钟。或者,让蠕虫重力沉降10-15分钟。
- 将上清液除去至~2mL,并将上清液丢弃到装有碘的废液容器中以杀死任何蠕虫。
- 向 15 mL 收集管中加入更多 Baermann 水并重复旋转。将上清液除去至~2mL,并如步骤4.11所述丢弃。
- 重复步骤 4.11 和 4.12,直到所有蠕虫都收集在 15 mL 离心管中。最后旋转后,尽可能多地除去水。
- 检查管底部的蠕虫颗粒(40-100μL)。如果没有可见的蠕虫,请再等待1-2小时,并尝试从Baermann设备中收集更多蠕虫。
- 将蠕虫在尽可能少的水中转移到带有 大肠杆菌 HB101草坪的6厘米2%NGM板上。使用此板作为显微注射的 源板 。
- 丢弃粪便 - 木炭混合物,方法是用稀释的碘(Lugol碘在水中稀释50%)处理,将其包裹在塑料薄膜中以吸收滴水,并将其放入生物危害废物容器中。
- 向收集桶中加入 10 mL 稀释的碘,并将 Baermann 中多余的水排入其中。
- 用10%漂白剂清洗可重复使用的组件(漏斗,收集桶,带薄纱的塑料支架,塑料盖和夹具)并彻底冲洗。
5.拉动和装载显微注射针:注射前
- 通过使用拉针器拉动玻璃毛细管来制备显微注射针。
注:配备 3 mm 铂/铱灯丝的商用拉针机的示例设置为 Heat = 810-820,Pull = 800-820,micrometer = 2.5。 - 在解剖显微镜下查看尖端。如果针具有所需的形状(图3A-F),请拉4-6针(2-3毛细管)。要获得正确的针形,请根据需要更改设置:将"加热"或"拉动"设置调整10,并拉动新针,直到锥度和轴的形状更合适。

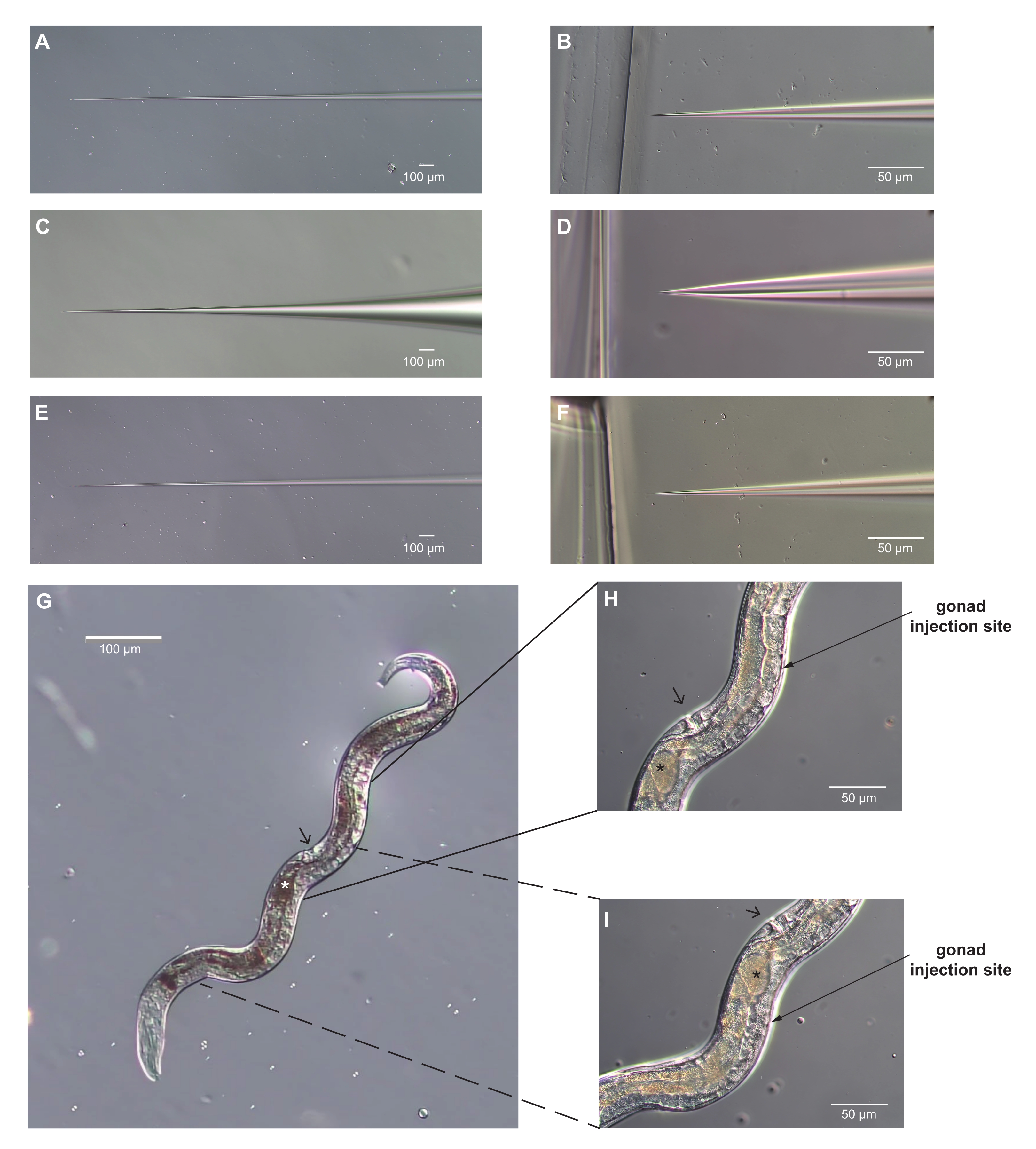
图3:显微注射针和类圆线虫胸骨疣成年雌性,具有显微注射的最佳部位。(A-F)显微注射针的图像。(A-B)针的轴锥度(A)和针尖(B),其形状正确用于显微注射。尖端足够锋利,可以刺穿角质层,足够窄,不会造成过度损伤。(C-D)显微注射针的轴锥度(C)和尖端(D),其形状不正确,无法进行显微注射。尖端太钝,太宽,会对蠕虫造成过度伤害。(E-F)针的轴锥度(E)和尖端(F),可能太长而细长而无法用于显微注射。F 中的尖端与 D 中的尖端非常相似。然而,轴更窄,太灵活,无法有效地刺穿角质层。此外,非常细长的针头很容易堵塞。(G)正确定位用于显微注射的整个蠕虫的图像,假设针头是从右侧进入的。前部向下和向左;外阴由箭头表示。性腺沿着雌性的右侧可见。这只雌性的子宫里只有一个卵子(用星号表示)。(H, I)显微注射部位的放大视图。箭头的角度近似于注射针的角度。外阴可以用作地标;它位于性腺手臂蠕虫的另一侧。性腺的手臂围绕肠道弯曲,末端与分裂的核相反。(H) 性腺的后臂;(一)前臂。可以注射任何一个或两个手臂。对于 H,I,约定与 G 中一样。比例尺= 50 μm (B, D, F, H, I);100 μm (A, C, E, G).请点击此处查看此图的大图。
{kind=link}
- 将拉动的针头存放在15厘米的塑料培养皿中,并用一块卷起的胶带固定针头并避免灰尘积聚在尖端上。
- 将0.7μL显微注射混合物滴在轴的开口端。使用卷起的胶带将针垂直于架子上,在10分钟内用混合物填充锥形轴。一次准备2针,以防第一根针不起作用。
6.准备显微镜并断针
注:显微注射使用带有5倍和40倍物镜的倒置显微镜,并配有显微注射器装置来控制针的运动。倒置显微镜应放置在厚重的工作台或防振空气工作台上,以减少振动噪音。显微注射器针座连接到氮气,氮气施加输送显微注射混合物所需的压力。附近较小的解剖显微镜用于转移蠕虫。
- 根据液体流量,将气罐压力设置为 ~40-60 psi(用于断针)和 ~30-50 psi(用于显微注射)。
- 在解剖显微镜上,使用标准铂蠕虫镐用卤烃油覆盖显微注射垫盖玻片上的玻璃碎片。
- 将显微注射垫盖玻片放在显微注射镜上,并找到沾满油的玻璃碎片。对齐玻璃碎片,使边缘垂直于针的方向,以用作用于折断针的表面。
- 使用解剖显微镜验证针在锥形轴中没有气泡或碎屑。然后,将针头固定在加压支架中1-1.5厘米。
- 用眼睛将针尖放在显微镜视场的中心。然后在低放大倍率下,将针尖定位在视场上,垂直于玻璃碎片的一侧。
- 切换到高功率,并将针尖与玻璃边缘对齐,靠近但不接触它。
注意:拉动时,针头熔合闭合。 - 要打破针尖以允许液体流动,请在玻璃片的侧面轻轻敲击它,同时施加来自气体的连续压力(补充图S1)。一旦液体开始流动,检查尖端的形状,并确保它与容易流动的液体一起尖锐。
注:如果液体流动过快或末端太钝,蠕虫在显微注射过程中会损坏(视频1和图3A-F)。 - 当液体从针头流出时,将显微注射载玻片移动到解剖范围,并在琼脂垫上滴下1-2μL卤烃油以放置蠕虫。
- 将20-30个年轻成年 类圆线虫 转移到没有细菌的2%NGM平板上至少5分钟,以除去多余的表面细菌并选择单个蠕虫进行显微注射。注射时根据需要向NGM板添加更多蠕虫。
7. 显微注射 类圆线虫
- 在蠕虫镐上使用少量卤代碳油,从2%NGM平板中选择一个类 圆线虫 的年轻成年雌性,她的性腺中有1-4个卵,没有细菌。
- 将蠕虫转移到琼脂垫上的一小滴油中。使用蠕虫镐,轻轻定位蠕虫,使其不卷曲,性腺可见且易于接近。注意性腺的方向(图3G)。
- 将蠕虫定位在显微注射显微镜的视野中。确保性腺与针头位于同一侧,并定位,以便针头以轻微的角度接触性腺(图3H,I)。
- 将针尖放在同一焦平面内蠕虫的一侧。瞄准靠近蠕虫中间的性腺手臂。使用显微注射器将针头轻轻插入性腺(视频2)。
- 立即对针头施加压力,用DNA溶液轻轻填充整个性腺手臂。用眼睛确定何时注射了足够的液体(视频2)。
注意:填充性腺可能需要长达2秒的时间。 - 取下针头并检查以确定伤口是否闭合。
注意:如果性腺突出体壁,蠕虫受损太严重,无法产生后代(补充视频S1)。 - 如果性腺的另一只手臂可见,则重复该动作。
- 注射完成后,通过在琼脂垫上用针尖施加压力来快速验证针头是否堵塞。将带有注射蠕虫的载玻片转移到解剖显微镜上。
- 为了恢复注射的蠕虫,首先在蠕虫上滴几滴BU,使其漂浮在琼脂垫上。
- 在蠕虫镐上收集少量HB101细菌。用蠕虫镐上的粘附细菌触摸蠕虫,将其从液体中取出。
- 轻轻地将蠕虫转移到回收板上, 回收 板是含有HB101草坪的2%NGM板。
注意:蠕虫应在几分钟内开始爬网。 - 注射一些雌性后,从源板中加入一些未注射的雄性。
注意:五名女性中至少一名男性是一个很好的基线;男性过多是首选。 - 重复所有步骤,直到为实验注射了足够的雌性。
- 注射后将成虫留在恢复板上至少1小时,以使蠕虫恢复和交配。
8. 注射类圆线虫的回收和培养
- 从未感染的宿主动物那里收集粪便过夜,使用与受感染动物相同的方案。
- 将未感染的粪便与少量木炭混合(这些板的粪便与木炭的比例约为2:1)。
- 将少量粪便 - 木炭混合物倒入衬有湿滤纸的6厘米培养皿中。确保混合物不会接触盘子的盖子。
- 使用设置为3μL的移液管将蠕虫转移到粪便 - 木炭板中的粪便中。将蠕虫直接放在粪便上,而不是木炭上。
- 使用解剖镜验证成虫是否在粪便炭板上。
- 为了培养蠕虫,将板放在加湿室中, 即带有紧密盖子的塑料盒,内衬湿纸巾。
注意:2天后,将有混合的幼虫阶段。5天后,大多数幼虫将发育成iL3;一些年轻的幼虫将保留下来。7天后,所有幼虫应为iL3s。
9. 收集和筛选F 1 幼虫以回收转基因/敲除
- 使用Baermann装置,从注射后小规模粪便 - 木炭培养板中收集幼虫。为了获得尽可能多的幼虫,请等待至少2小时,然后再从Baermann设备中恢复蠕虫。
- 按照步骤4.10-4.14将幼虫浓缩在15 mL离心管中,并将幼虫转移到装有BU的小手表玻璃杯中。
- 如果幼虫将用于行为实验,请使用2%NGM板和HB101厚厚的草坪进行筛选。
- 将20-30只幼虫转移到HB101草坪上。
注意:细菌会减缓幼虫的运动。 - 在荧光解剖显微镜下,识别表达目标转基因的幼虫。使用蠕虫镐选择转基因幼虫,并将它们移动到带有BU的小手表玻璃上。
- 使用新的HB101平板筛选另一小批幼虫。当收集了足够的幼虫用于实验用途时,用稀释的碘(50%的Lugol碘稀释在水中)处理HB101板和多余的蠕虫,并将它们作为生物危害废物丢弃。或者,使用含有烷基苄基氯化铵的浓缩犬舍清洁剂杀死多余的蠕虫。
- 立即使用蠕虫或将它们放在浅表玻璃中,在少量BU中过夜。
注意:如果液体太深,蠕虫可能会变得缺氧。将幼虫留在BU中过夜可能会影响某些行为;因此,在6小时内使用幼虫进行行为实验。
- 将20-30只幼虫转移到HB101草坪上。
- 如果幼虫将用于显微镜检查而不是行为测定,则通过尼古丁麻痹可逆地固定蠕虫以进行筛选。
- 使用剃须刀片,在10厘米趋化性板12 的塑料底部划出网格,以便更容易跟踪板上蠕虫的位置。
- 将〜3μL幼虫滴入BU中的正方形中。根据需要填充任意数量的正方形。不要使用靠近板边缘的那些,因为幼虫可能会爬到板的侧面。
- 在水中加入15-20μL滴1%尼古丁到蠕虫滴剂中。
注意:4分钟后,蠕虫将瘫痪。 - 使用荧光解剖显微镜筛选蠕虫。
- 使用蠕虫镐将转基因幼虫转移到装有1-2 mL BU的小手表玻璃杯中。
注意:幼虫将瘫痪数小时,并且可以轻松安装在显微镜载玻片上进行显微镜检查。如果在BU中过夜,iL3s将恢复,可用于某些测定或哺乳动物宿主感染。然而,尼古丁麻痹和BU中的过夜孵育可能会影响某些行为。
Access restricted. Please log in or start a trial to view this content.
结果
如果实验成功,F1 幼虫将表达感兴趣的转基因和/或突变表型(图4)。然而,转化率变化很大,取决于结构,蠕虫的健康状况,注射后培养条件和实验者的技能。一般来说,成功的实验将产生每只注射雌性>15 F 1 幼虫,荧光标记物的转化率为>3%。如果活的后代总数平均少于10只幼虫/雌性,那么该结构可能是有毒的,并且转化的幼虫无法存活。发现大量荧光卵?...
Access restricted. Please log in or start a trial to view this content.
讨论
该显微注射方案详细介绍了将转基因结构和CRISPR / Cas9介导的诱变引入 粪类链球菌 和 鼠李糖杆菌的方法。 对于粪 类葡萄球菌 和 鼠李松,注射后存活率和转基因或诱变率受制于几个可以微调的变量。
成功转基因的第一个关键考虑因素是如何构建质粒转基因。以前的研究发现,类 圆线虫 中外源转基因的表达需要使用 类圆线虫 5'启动子...
Access restricted. Please log in or start a trial to view this content.
披露声明
作者声明没有利益冲突。
致谢
pPV540和pPV402是宾夕法尼亚大学James Lok博士的善意礼物。我们感谢阿斯特拉·布莱恩特对手稿的有益评论。这项工作由Burroughs-Wellcome Fund Researchs in the Pathogenesis of Disease Award,Howard Hughes Medical Institute教师学者奖和美国国立卫生研究院R01 DC017959(E.A.H.)资助。
Access restricted. Please log in or start a trial to view this content.
材料
| Name | Company | Catalog Number | Comments |
| (−)-Nicotine, ≥99% (GC), liquid | Sigma-Aldrich | N3876-5ML | nicotine for paralyzing worms |
| 3" iron C-clamp, 3" x 2" (capacity x depth) | VWR | 470121-790 | C-clamp to secure setup to bench top |
| Agarose LE | Phenix | RBA-500 | agarose for slides |
| Bone char, 4 lb pail, 10 x 28 mesh | Ebonex | n/a | charcoal for fecal-charcoal cultures |
| Bone char, granules, 10 x 28 mesh | Reade | bonechar10x28 | charcoal for fecal-cultures (alternative to the above) |
| Coarse micromanipulator | Narishige | MMN-1 | coarse micromanipulator |
| Corning Costar Spin-X centrifuge tube filters | Fisher | 07-200-385 | microfilter column |
| Cover glass, 48 x 60 mm, No. 1 thickness | Brain Research Lab | 4860-1 | coverslips (48 x 60 mm) |
| Deep Petri dishes, heavy version with 6 vents, 100 mm diameter | VWR | 82050-918 | 10 cm Petri dishes (for fecal-charcoal cultures) |
| Eisco retort base w/ rod | Fisher | 12-000-101 | stand for Baermann apparatus |
| Eppendorf FemtoJet microinjector microloader tips | VWR | 89009-310 | for filling microinjection needles |
| Fisherbrand absorbent underpads | Fisher | 14-206-62 | bench paper (for prepping) |
| Fisherbrand Cast-Iron Rings | Fisher | 14-050CQ | Baermann o-ring |
| Fisherbrand tri-cornered polypropylene beakers | Fisher | 14-955-111F | Plastic beaker (for mixing) |
| Fisherbrand tri-cornered polypropylene beakers | Fisher | 14-955-111F | Plastic beaker (for catch bucket/water bucket) |
| Fisherbrand tri-cornered polypropylene beakers | Fisher | 14-955-111F | Plastic beaker (x2) (to make holder) |
| Gorilla epoxie in syringe | McMaster-Carr | 7541A51 | glue (to attach tubing) |
| Halocarbon oil 700 | Sigma-Aldrich | H8898-50ML | halocarbon oil |
| High-temperature silicone rubber tubing for food and beverage, 1/2" ID, 5/8" OD | McMaster-Carr | 3038K24 | tubing (for funnel) |
| KIMAX funnels, long stem, 60° Angle, Kimble Chase | VWR | 89001-414 | Baermann funnel |
| Kimberly-Clark Professional Kimtech Science benchtop protectors | Fisher | 15-235-101 | bench paper (for prepping) |
| Leica stereomicroscope with fluorescence | Leica | M165 FC | GFP stereomicroscope for identifying and sorting transgenic worms |
| microINJECTOR brass straight arm needle-holder | Tritech | MINJ-4 | microinjection needle holder |
| microINJECTOR system | Tritech | MINJ-1 | microinjection system |
| Mongolian Gerbils | Charles River Laboratories | 213-Mongolian Gerbil | gerbils for maintenance of S. stercoralis, male 4-6 weeks |
| Nasco Whirl-Pak easy-to-close bags, 18 oz | VWR | 11216-776 | Whirl-Pak sample bags |
| Nylon tulle (mesh) | Jo-Ann Fabrics | zprd_14061949a | nylon mesh for Baermann holder |
| Platinum wire, 36 Gauge, per inch | Thomas Scientific | 1233S72 | platinum/iridium wire for worm picks |
| Puritan tongue depressors, 152 mm (L) x 17.5 mm (W) | VWR | 62505-007 | wood sticks (for mixing samples) |
| QIAprep Spin Miniprep Kit (250) | QIAGEN | 27106 | QIAGEN miniprep kit |
| Rats-Long Evans | Envigo | 140 HsdBlu:LE Long Evans | rats for maintenance of S. ratti, female 4-6 weeks |
| Rats-Sprague Dawley | Envigo | 002 Hsd:Sprague Dawley SD | rats for maintenance of S. ratti, female 4-6 weeks |
| Really Useful Boxes translucent storage boxes with lids, 1.6 L capacity, 7-5/8" x 5-5/16" x 4-5/16" | Office Depot | 452369 | plastic boxes for humidified chamber |
| Shepherd techboard, 8 x 16.5 inches | Newco | 999589 | techboard |
| Stainless steel raised wire floor | Ancare | R20SSRWF | wire cage bottoms |
| StalkMarket compostable cutlery spoons, 6", white, pack of 1,000 | Office Depot | 9587303 | spoons |
| Stender dish, stacking type, 37 x 25 mm | Carolina (Science) | 741012 | watch glasses (small, round) |
| Stereomicroscope | Motic | K-400 LED | dissecting prep scope |
| Storage tote, color clear/white, outside height 4-7/8 in, outside length 13-5/8 in, Sterilite | Grainger | 53GN16 | plastic boxes for humidified chamber |
| Sutter P-30 micropipette puller | Sutter | P-30/P | needle puller with platinum/iridium filament |
| Syracuse watch glasses | Fisher | S34826 | watch glasses (large, round) |
| Thermo Scientific Castaloy fixed-angle clamps | Fisher | 05-769-2Q | funnel clamps (2x) |
| Three-axis hanging joystick oil hydrolic micromanipulator | Narishige | MM0-4 | fine micromanipulator |
| United Mohr pinchcock clamps | Fisher | S99422 | Pinch clamps (2x) |
| Vented, sharp-edge Petri dishes (60 mm diameter) | Tritech Research | T3308P | 6 cm Petri dishes (for small-scale fecal-charcoal cultures) |
| VWR light-duty tissue wipers | VWR | 82003-820 | lining for Baermann holder |
| watch glass, square, 1-5/8 in | Carolina (Science) | 742300 | watch glasses (small, square) |
| Whatman qualitative grade plain circles, grade 1, 5.5 cm diameter | Fisher | 09-805B | filter paper (for 6 cm Petri dishes) |
| Whatman qualitative grade plain circles, grade 1, 9 cm diameter | Fisher | 09-805D | filter paper (for 10 cm Petri dishes) |
| World Precision Instrument borosilicate glass capillary, 1.2 mm x 4 in | Fisher | 50-821-813 | glass capillaries for microinjection needles |
| X-Acto Knives, No. 1 Knife With No. 11 Blade | Office Depot | 238816 | X-Acto knives without blades to hold worm picks |
| Zeiss AxioObserver A1 | Zeiss | n/a | inverted microscope |
参考文献
- Krolewiecki, A. J., et al. A public health response against Strongyloides stercoralis: time to look at soil-transmitted helminthiasis in full. PLoS Neglected Tropical Diseases. 7 (5), 2165(2013).
- Buonfrate, D., et al. The global prevalence of Strongyloides stercoralis infection. Pathogens. 9 (6), 468(2020).
- Castelletto, M. L., Gang, S. S., Hallem, E. A. Recent advances in functional genomics for parasitic nematodes of mammals. Journal of Experimental Biology. 223, Pt Suppl 1 206482(2020).
- Evans, T. C., et al. Transformation and microinjection. WormBook. , ed. The C. elegans Research Communnity (2006).
- Lok, J. B., Unnasch, T. R., et al. Transgenesis in animal parasitic nematodes: Strongyloides spp. and Brugia spp. WormBook. , ed. The C. elegans Research Communnity (2013).
- Shao, H. G., Li, X. S., Lok, J. B. Heritable genetic transformation of Strongyloides stercoralis by microinjection of plasmid DNA constructs into the male germline. International Journal for Parasitology. 47 (9), 511-515 (2017).
- Schafer, T. W., Skopic, A. Parasites of the small intestine. Current Gastroenterology Reports. 8 (4), 312-320 (2006).
- Stiernagle, T. Maintenance of C. elegans. The C. elegans Research Community, WormBook. , (2006).
- Gang, S. S., et al. Targeted mutagenesis in a human-parasitic nematode. PLoS Pathogens. 13 (10), 1006675(2017).
- Lok, J. B. Strongyloides stercoralis: a model for translational research on parasitic nematode biology. The C. elegans Research Community, WormBook. , (2007).
- Hawdon, J. M., Schad, G. A. Long-term storage of hookworm infective larvae in buffered saline solution maintains larval responsiveness to host signals. Proceedings of the Helminthological Society of Washington (USA). 58 (1), 140-142 (1991).
- Bargmann, C. I., Hartwieg, E., Horvitz, H. R. Odorant-selective genes and neurons mediate olfaction in C. elegans. Cell. 74 (3), 515-527 (1993).
- Junio, A. B., et al. Strongyloides stercoralis: cell- and tissue-specific transgene expression and co-transformation with vector constructs incorporating a common multifunctional 3' UTR. Experimental Parasitology. 118 (2), 253-265 (2008).
- Gang, S. S., et al. Chemosensory mechanisms of host seeking and infectivity in skin-penetrating nematodes. Proceedings of the National Academy of Sciences of the United States of America. 117 (30), 17913-17923 (2020).
- Bryant, A. S., et al. A critical role for thermosensation in host seeking by skin-penetrating nematodes. Current Biology. 28 (14), 2338-2347 (2018).
- Lok, J. B. Nucleic acid transfection and transgenesis in parasitic nematodes. Parasitology. 139 (5), 574-588 (2012).
- Shao, H., et al. Transposon-mediated chromosomal integration of transgenes in the parasitic nematode Strongyloides ratti and establishment of stable transgenic lines. PLoS Pathogens. 8 (8), 1002871(2012).
- Lok, J. piggyBac: a vehicle for integrative DNA transformation of parasitic nematodes. Mobile Genetic Elements. 3 (2), 24417(2013).
- Li, X., et al. Successful transgenesis of the parasitic nematode Strongyloides stercoralis requires endogenous non-coding control elements. International Journal for Parasitology. 36 (6), 671-679 (2006).
- Bryant, A. S., Hallem, E. A. The Wild Worm Codon Adapter: a web tool for automated codon adaptation of transgenes for expression in non-Caenorhabditis nematodes. G3. 3 (7), (2021).
- Crane, M., et al. In vivo measurements reveal a single 5'-intron is sufficient to increase protein expression level in Caenorhabditis elegans. Scientific Reports. 9 (1), 9192(2019).
- Han, Z., et al. Improving transgenesis efficiency and CRISPR-associated tools through codon optimization and native intron addition in Pristionchus nematodes. Genetics. 216 (4), 947-956 (2020).
- Adams, S., Pathak, P., Shao, H., Lok, J. B., Pires-daSilva, A. Liposome-based transfection enhances RNAi and CRISPR-mediated mutagenesis in non-model nematode systems. Scientific Reports. 9 (1), 483(2019).
- Dulovic, A., Puller, V., Streit, A. Optimizing culture conditions for free-living stages of the nematode parasite Strongyloides ratti. Experimental Parasitology. 168, 25-30 (2016).
- Harvey, S. C., Gemmill, A. W., Read, A. F., Viney, M. E. The control of morph development in the parasitic nematode Strongyloides ratti. Proceedings of the Royal Society B: Biological Sciences. 267 (1457), 2057-2063 (2000).
- Kim, A., Pyykko, I. Size matters: versatile use of PiggyBac transposons as a genetic manipulation tool. Molecular and Cellular Biochemistry. 354 (1-2), 301-309 (2011).
- Lok, J. B., Shao, H., Massey, H. C., Li, X. Transgenesis in Strongyloides and related parasitic nematodes: historical perspectives, current functional genomic applications and progress towards gene disruption and editing. Parasitology. 144 (3), 327-342 (2017).
- Farboud, B., Meyer, B. J. Dramatic enhancement of genome editing by CRISPR/Cas9 through improved guide RNA design. Genetics. 199 (4), 959-971 (2015).
- Cheong, M. C., et al. Identification of a nuclear receptor/coactivator developmental signaling pathway in the nematode parasite Strongyloides stercoralis. Proceedings of the National Academy of Sciences of the United States of America. 118 (8), 2021864118(2021).
- Nolan, T. J., Megyeri, Z., Bhopale, V. M., Schad, G. A. Strongyloides stercoralis: the first rodent model for uncomplicated and hyperinfective strongyloidiasis, the Mongolian gerbil (Meriones unguiculatus). Journal of Infectious Diseases. 168 (6), 1479-1484 (1993).
- Li, X., et al. Transgenesis in the parasitic nematode Strongyloides ratti. Molecular and Biochemical Parasitology. 179 (2), 114-119 (2011).
- Viney, M. E. Exploiting the life cycle of Strongyloides ratti. Parasitology Today. 15 (6), 231-235 (1999).
- Stoltzfus, J. D., Massey, H. C., Nolan, T. J., Griffith, S. D., Lok, J. B. Strongyloides stercoralis age-1: a potential regulator of infective larval development in a parasitic nematode. PLoS ONE. 7 (6), 38587(2012).
- Castelletto, M. L., Massey, H. C., Lok, J. B. Morphogenesis of Strongyloides stercoralis infective larvae requires the DAF-16 ortholog FKTF-1. PLoS Pathogens. 5 (4), 1000370(2009).
- Douglas, B., et al. Transgenic expression of a T cell epitope in Strongyloides ratti reveals that helminth-specific CD4+ T cells constitute both Th2 and Treg populations. PLoS Pathogens. 17 (7), 1009709(2021).
Access restricted. Please log in or start a trial to view this content.
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。