
È necessario avere un abbonamento a JoVE per visualizzare questo. Accedi o inizia la tua prova gratuita.
Method Article
Generazione di transgenici e knockout in specie di Strongyloides mediante microiniezione
In questo articolo
Riepilogo
Il toolkit genomico funzionale per i nematodi parassiti Strongyloides stercoralis e Strongyloides ratti comprende transgenesi, mutagenesi mediata da CRISPR/Cas9 e RNAi. Questo protocollo dimostrerà come utilizzare la microiniezione intragonennale per introdurre transgeni e componenti CRISPR in S. stercoralis e S. ratti.
Abstract
Il genere Strongyloides è costituito da più specie di nematodi penetranti nella pelle con diverse gamme di ospiti, tra cui Strongyloides stercoralis e Strongyloides ratti. S. stercoralis è un nematode umano-parassitario, penetrante nella pelle che infetta circa 610 milioni di persone, mentre il parassita del ratto S. ratti è strettamente correlato a S. stercoralis ed è spesso usato come modello di laboratorio per S. stercoralis. Sia S. stercoralis che S. ratti sono facilmente suscettibili alla generazione di transgenici e knockout attraverso la tecnica di somministrazione esogena di acido nucleico della microiniezione intragonadiale e, come tali, sono emersi come sistemi modello per altri elminti parassiti che non sono ancora suscettibili a questa tecnica.
Gli adulti parassiti di Strongyloides abitano l'intestino tenue del loro ospite e rilasciano la progenie nell'ambiente attraverso le feci. Una volta nell'ambiente, le larve si sviluppano in adulti che vivono liberi, che vivono nelle feci e producono progenie che deve trovare e invadere un nuovo ospite. Questa generazione ambientale è unica per la specie Strongyloides e abbastanza simile nella morfologia al modello di nematode a vita libera Caenorhabditis elegans che le tecniche sviluppate per C. elegans possono essere adattate per l'uso con questi nematodi parassiti, compresa la microiniezione intragonadiale. Utilizzando la microiniezione intragonadiale, un'ampia varietà di transgeni può essere introdotta in Strongyloides. I componenti CRISPR/Cas9 possono anche essere microiniettati per creare larve mutanti di Strongyloides . Qui, viene descritta la tecnica di microiniezione intragonennale in Strongyloides, compresa la preparazione di adulti liberi, la procedura di iniezione e la selezione della progenie transgenica. Sono incluse immagini di larve transgeniche di Strongyloides create utilizzando la mutagenesi CRISPR/Cas9. Lo scopo di questo documento è quello di consentire ad altri ricercatori di utilizzare la microiniezione per creare Strongyloides transgenici e mutanti.
Introduzione
Strongyloides stercoralis è stato a lungo trascurato come un importante agente patogeno umano rispetto agli anchilostomi più ampiamente riconosciuti e al nematode Ascaris lumbricoides1. Precedenti studi sul carico di vermi spesso hanno gravemente sottovalutato la prevalenza di S. stercoralis a causa della bassa sensibilità dei metodi diagnostici comuni per S. stercoralis2. Negli ultimi anni, studi epidemiologici basati su strumenti diagnostici migliorati hanno stimato che la vera prevalenza delle infezioni da S. stercoralis è molto più alta di quanto precedentemente riportato, circa 610 milioni di persone in tutto il mondo2.
Sia S. stercoralis che altre specie di Strongyloides, tra cui il parassita di ratto strettamente correlato e il comune modello di laboratorio S. ratti, hanno un ciclo di vita insolito che è vantaggioso per gli studi genomici sperimentali perché consiste di generazioni sia parassitarie che libere viventi (ambientali)3 (Figura 1). In particolare, sia S. stercoralis che S. ratti possono attraversare un'unica generazione di vita libera. La generazione a vita libera è costituita da larve post-parassitarie che si sviluppano in maschi e femmine adulti che vivono liberi; tutta la progenie degli adulti che vivono liberi si sviluppa in larve infettive, che devono infettare un ospite per continuare il ciclo di vita. Inoltre, questa generazione ambientale o di vita libera può essere manipolata sperimentalmente in laboratorio. Poiché gli adulti Strongyloides a vita libera e gli adulti C. elegans condividono una morfologia simile, tecniche come la microiniezione intragonennale originariamente sviluppate per C. elegans possono essere adattate per l'uso con Strongyloides 4,5 adulti a vita libera. Mentre il DNA viene generalmente introdotto nelle femmine adulte che vivono liberamente, sia i maschi che le femmine di Strongyloides possono essere microiniettati6. Pertanto, sono disponibili strumenti genomici funzionali per interrogare molti aspetti della biologia di Strongyloides. Altri nematodi parassiti mancano di una generazione di vita libera e, di conseguenza, non sono così facilmente suscettibili alle tecniche genomiche funzionali3.

Figura 1: Il ciclo di vita di Strongyloides stercoralis. Le femmine parassitarie di S. stercoralis abitano l'intestino tenue dei loro ospiti mammiferi (umani, primati non umani, cani). Le femmine parassitarie si riproducono per partenogenesi e depongono le uova all'interno dell'intestino tenue. Le uova si schiudono mentre sono ancora all'interno dell'ospite in larve post-parassitarie, che vengono poi passate nell'ambiente con le feci. Se le larve post-parassitarie sono maschi, si sviluppano in maschi adulti che vivono liberamente. Se le larve post-parassitarie sono femmine, possono svilupparsi in femmine adulte a vita libera (sviluppo indiretto) o larve infettive di terzo stadio (iL3s; sviluppo diretto). I maschi e le femmine che vivono liberi si riproducono sessualmente per creare progenie che sono costretti a diventare iL3. In determinate condizioni, S. stercoralis può anche subire l'autoinfezione, in cui alcune delle larve post-parassitarie rimangono all'interno dell'intestino ospite piuttosto che passare nell'ambiente nelle feci. Queste larve possono svilupparsi in larve autoinfettive (L3a) all'interno dell'ospite, penetrare attraverso la parete intestinale, migrare attraverso il corpo e alla fine tornare all'intestino per diventare adulti riproduttivi. Il ciclo di vita di S. ratti è simile, tranne che S. ratti infetta i ratti e non ha un ciclo autoinfettivo. La generazione ambientale è la chiave per l'utilizzo delle specie di Strongyloides per studi genetici. Le femmine adulte a vita libera (P0) possono essere microintetate; la loro progenie, che diventerà tutta iL3, sono i potenziali transgenici F1. Questa figura è stata modificata da Castelletto et al. 3. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}
S. stercoralis condivide molti aspetti della sua biologia con altri nematodi gastrointestinali parassiti umani, tra cui l'invasione dell'ospite e la modulazione immunitaria dell'ospite. Ad esempio, gli anchilostomi umani-parassiti nei generi Necator e Ancylostoma infettano anche la penetrazione della pelle, navigano in modo simile attraverso il corpo e, infine, risiedono come adulti parassiti nell'intestino tenue7. Pertanto, molti nematodi gastrointestinali probabilmente utilizzano comportamenti sensoriali comuni e tecniche di evasione immunitaria. Di conseguenza, le conoscenze raccolte da Strongyloides completeranno i risultati in altri nematodi meno trattabili geneticamente e porteranno a una comprensione più completa di questi parassiti complessi e importanti.
Questo protocollo di microiniezione delinea il metodo per introdurre il DNA nelle femmine adulte viventi libere di Strongyloides per produrre progenie transgenica e mutante. Vengono descritti i requisiti di mantenimento del ceppo, compresi i tempi di sviluppo dei vermi adulti per le microiniezioni e la raccolta della progenie transgenica. Sono inclusi protocolli e una dimostrazione della tecnica completa di microiniezione, insieme ai protocolli per la coltura e lo screening della progenie transgenica, insieme a un elenco di tutte le attrezzature e i materiali di consumo necessari.
Protocollo
NOTA: I gerbilli erano usati per passare S. stercoralis, e i ratti erano usati per passare S. ratti. Tutte le procedure sono state approvate dall'UCLA Office of Animal Research Oversight (Protocollo n. 2011-060-21A), che aderisce agli standard AAALAC e alla Guida per la cura e l'uso degli animali da laboratorio. Le seguenti attività devono essere completate almeno un giorno prima della microiniezione: coltura di vermi, preparazione di microiniezione, creazione di costrutti per il mix di microiniezione e diffusione di batteri (E. coli HB101) su piastre di 6 cm Nematode Growth Media (NGM)8. Le femmine che vivono libere richiedono un minimo di 24 ore di raccolta post-fecale a 25 °C per svilupparsi in giovani adulti prima di poter essere microiniettate. I cuscinetti per microiniezione devono essere completamente asciutti. Le piastre batteriche devono asciugarsi e stabilire un piccolo prato.
1. Preparazione di vetrini per microiniezione: almeno un giorno prima dell'iniezione
NOTA: i vermi sono montati su coverslip di microiniezione con tamponi di agar asciutti per iniezione.
- Impostare un blocco di calore a 90 °C.
- Aggiungere 5 ml di ddH2O, quindi 100 mg di agarosio in un tubo di vetro borosilicato.
- Riscaldare la miscela di agarosio nel tubo su una fiamma fino a quando l'agarosio non si è sciolto.
- Posizionare il tubo in un blocco termico impostato a 90 °C per mantenere l'agarosio allo stato liquido.
- Far cadere ~ 180 μL della soluzione di agarosio su una copertina usando un pipetto Pasteur di vetro o un pipetto con una punta di plastica. Lascia immediatamente cadere un secondo coverslip sulla parte superiore per appiattire l'agarosio in un cuscinetto sottile.
- Dopo 5-10 s, rimuovere il coperchio superiore facendo scorrere i due a parte. Determinare su quale diapositiva si trova il tampone di agar e appoggiarlo a faccia in su.
- Selezionare un piccolo frammento di vetro da una slitta rotta e premerlo delicatamente nell'agar vicino al bordo superiore del pad usando una pinza (Figura supplementare S1).
- Continuare a fare microiniezioni con la soluzione di agarosio.
- Asciugare i cuscinetti di agarosio durante la notte sul banco o in un forno. Conservare nella scatola coverslip.
NOTA: I tamponi di agarosio possono essere utilizzati per un massimo di 2 mesi, ma vengono utilizzati solo per una corsa di iniezione.
2. Coltivare Strongyloides per ottenere vermi per microiniezione: 1 - 2 giorni prima dell'iniezione
NOTA: Un protocollo di mantenimento del ceppo può essere trovato nel materiale supplementare, che include una descrizione dettagliata di come infettare gerbilli e ratti con nematodi e raccogliere nematodi dalle feci di animali infetti.
- Due giorni prima del giorno dell'iniezione, posizionare gli animali infetti 9,10 in gabbie di raccolta durante la notte.
- La mattina dopo, raccogli le feci infestate e prepara piatti di carbone fecale 9,10.
- Posizionare una piastra a 25 °C per 24 ore per consentire ai vermi a vita libera di svilupparsi in giovani adulti.
- La notte prima del giorno dell'iniezione, mettere gli animali ospiti non infetti in gabbie di raccolta.
- Il giorno dell'iniezione, raccogliere feci non infestate per la coltivazione post-iniezione.
3. Preparazione della miscela di microiniezione: prima o il giorno dell'iniezione
NOTA: La miscela di microiniezione è costituita dai plasmidi di interesse diluiti alla concentrazione desiderata in soluzione salina tamponata da verme (BU) (50 mM Na2HPO4, 22 mM KH2PO4, 70 mM NaCl)11.
- Determinare la concentrazione delle scorte di plasmidi e la concentrazione desiderata nella miscela di microiniezione (Tabella 1).
| Mix di microiniezione: costrutto reporter | |||
| Componente | Concentrazione delle scorte | Importo | Concentrazione finale |
| pMLC30 gpa-3::gfp | 300 ng/μL | 1,7 μL | 50 ng/μL |
| Bu | Na | 8,3 μL | Na |
| totale | 10 μL | 50 ng/μL | |
| Miscela di microiniezione: mutagenesi CRISPR/Cas9 | |||
| Componente | Concentrazione delle scorte | Importo | Concentrazione finale |
| pMLC47 tax-4 sgRNA | 300 ng/μL | 2,7 μL | 80 ng/μL |
| pEY11 Ss-tax-4 Plasmide HDR | 400 ng/μL | 2,0 μL | 80 ng/μL |
| pPV540 strCas9 plasmide | 350 ng/μL | 1,1 μL | 40 ng/μL |
| Bu | Na | 4,2 μL | Na |
| totale | 10 μL | 200 ng/μL | |
| Mix di microiniezione: integrazione piggyBac | |||
| Componente | Concentrazione delle scorte | Importo | Concentrazione finale |
| pMLC30 gpa3::gfp | 300 ng/μL | 2,0 μL | 60 ng/μL |
| pPV402 plasmide di trasposasi | 450 ng/μL | 0,9 μL | 40 ng/μL |
| Bu | Na | 7,1 μL | Na |
| totale | 10 μL | 100 ng/μL | |
Tabella 1: Esempi di miscele di microiniezione. I plasmidi e le concentrazioni per tre esempi di microiniezione si mescolano: uno per un costruttore reporter gpa-3::GFP 10, uno per l'interruzione mediata da CRISPR/Cas9 del locus Ss-tax-4 14,15 e uno per l'integrazione mediata da piggyBac di un costrutto Ss-gpa-3::GFP 13,17,18. strCas9 denota il gene Cas9 ottimizzato per il codone Strongyloides. Le concentrazioni finali elencate sono comunemente utilizzate nelle miscele di microiniezione Strongyloides.
- Diluire i plasmidi in BU ad un volume totale di 10-20 μL.
- Ruotare la miscela attraverso una colonna filtrante a 5.000 × g per 1-2 minuti.
- Utilizzare immediatamente la miscela di microiniezione o conservarla a -20 °C per un uso futuro.
4. Raccogliere Strongyloides giovani adulti per microiniezione: mattina del giorno dell'iniezione
- Installare l'apparato di Baermann con 1 piastra fecale-carbone di Strongyloides giovani adulti (Figura 2).
NOTA: La piastra fecale-carbone può contenere alcune larve infettive. I dispositivi di protezione individuale sono costituiti da un cappotto da laboratorio, guanti e protezione per gli occhi. Nessuna pelle deve essere esposta tra il guanto e la manica del camice da laboratorio.

Figura 2: L'apparato di Baermann utilizzato per raccogliere vermi parassiti dalle colture10. Il contenuto di una piastra fecale-carbone è posto nella parte superiore di una colonna di acqua calda. I vermi migrano nell'acqua e si raccolgono sul fondo dell'imbuto. (A) Per installare l'apparato Baermann, il supporto per l'imbuto Baermann è fissato al banco con un morsetto a C. Un tubo di gomma attaccato all'estremità dell'imbuto viene chiuso con morsetti a pizzico e un secchio di cattura viene posizionato sotto il tubo per gocciolare. L'acqua calda viene aggiunta all'imbuto di vetro. (B) Il supporto dell'anello di plastica per la miscela fecale-carbone viene quindi rivestito con 3 pezzi di tessuti di laboratorio (a sinistra). Un bastone di legno o un depressore della lingua (al centro) viene utilizzato per trasferire il contenuto di una piastra fecale-carbone (a destra) nel supporto dell'anello di plastica. (C) Un primo piano del fondo del supporto dell'anello di plastica per la miscela fecale-carbone, che mostra il doppio strato di tulle di nylon che riveste il fondo del supporto. (D) Il supporto fecale-carbone viene quindi posizionato sulla parte superiore dell'imbuto di vetro. (E) Il tessuto di laboratorio viene inumidito con acqua e chiuso sopra la miscela fecale-carbone. Più acqua calda viene aggiunta per immergere principalmente il carbone fecale. (F) L'impostazione completa di Baermann, con la coltura fecale-carbone sommersa sotto l'acqua calda. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}
- Installare un imbuto di vetro con tubi di raccolta in gomma su un supporto ad anello utilizzando un O-ring e fissarlo con un morsetto. Chiudere il tubo di raccolta con 2 morsetti a pizzico (Figura 2A).
- Metti un secchio di cattura sotto l'imbuto per catturare le gocce.
- Aggiungere acqua tiepida (circa 40 °C) all'imbuto a 5 cm sotto il bordo. Verificare che il sistema non perda.
- Fodera il supporto Baermann, un setaccio composto da 2 anelli di plastica con 2 strati di rete in tulle di nylon fissati tra di loro, con 3 pezzi sovrapposti di tessuto da laboratorio. Aggiungere la miscela fecale-carbone al supporto Baermann (Figura 2B,C).
- Posizionare il supporto Baermann con la miscela fecale-carbone nell'imbuto. Piegare i tessuti attorno alla miscela fecale-carbone e aggiungere abbastanza acqua per immergere la maggior parte del carbone fecale. Non riempire oltre 2 cm dal bordo dell'imbuto (Figura 2D,E).
- Coprire l'imbuto con un coperchio in plastica di Petri per contenere l'odore. Etichettare l'imbuto in base alle esigenze (Figura 2F).
- Attendere da 30 minuti a 1 ora per raccogliere i vermi dall'apparato di Baermann.
- Tenere un tubo centrifugo da 50 ml sotto il tubo di gomma nella parte inferiore dell'imbuto. Aprire con attenzione i morsetti sul fondo per erogare 30-40 ml di acqua contenente vermi nel tubo da 50 ml.
- Trasferire 15 mL dell'acqua Baermann contenente i vermi in un tubo centrifugo da 15 mL. Ruotare il tubo della centrifuga da 15 ml per 1 minuto a ~ 750 × g (lento). In alternativa, lasciare che i vermi si depositino per gravità per 10-15 minuti.
- Rimuovere il surnatante a ~ 2 ml e scartare il surnatante in un contenitore liquido di scarto con iodio per uccidere eventuali vermi.
- Aggiungere altra acqua Baermann al tubo di raccolta da 15 ml e ripetere la rotazione. Rimuovere il surnatante a ~2 mL ed eliminare come nel passaggio 4.11.
- Ripetere i passaggi 4.11 e 4.12 fino a quando tutti i vermi non vengono raccolti nel tubo della centrifuga da 15 ml. Dopo la rotazione finale, rimuovere quanta più acqua possibile.
- Ispezionare il pellet di vermi (40-100 μL) sul fondo del tubo. Se non sono visibili vermi, attendere altre 1-2 ore e provare a raccogliere più vermi dall'apparato di Baermann.
- Trasferire i vermi in quanta meno acqua possibile in una piastra NGM al 2% di 6 cm con un prato di E. coli HB101. Utilizzare questa piastra come piastra sorgente per la microiniezione.
- Scartare la miscela fecale-carbone trattandola con iodio diluito (una diluizione del 50% dello iodio di Lugol in acqua), avvolgendola in pellicola di plastica per catturare gocce e mettendola in un contenitore per rifiuti a rischio biologico.
- Aggiungere 10 ml di iodio diluito al secchio di raccolta e scaricare l'acqua in eccesso dal Baermann in esso.
- Lavare i componenti riutilizzabili (l'imbuto, il secchio di raccolta, il supporto in plastica con tulle, il coperchio di plastica e i morsetti) con candeggina al 10% e risciacquare abbondantemente.
5. Tirare e caricare gli aghi per microiniezione: poco prima dell'iniezione
- Preparare gli aghi per microiniezione tirando tubi capillari di vetro usando un estrattore di aghi.
NOTA: le impostazioni di esempio per un estrattore di aghi commerciale dotato di un filamento di platino/iridio da 3 mm sono Calore = 810-820, Pull = 800-820, micrometro = 2,5. - Visualizza i suggerimenti sotto un microscopio di dissezione. Se gli aghi hanno la forma desiderata (Figura 3A-F), tirare 4-6 aghi (2-3 tubi capillari). Per ottenere la corretta forma dell'ago, modificare le impostazioni in base alle esigenze: regolare le impostazioni Heat o Pull di 10 e tirare nuovi aghi fino a quando la forma del cono e dell'albero non sono più appropriati.

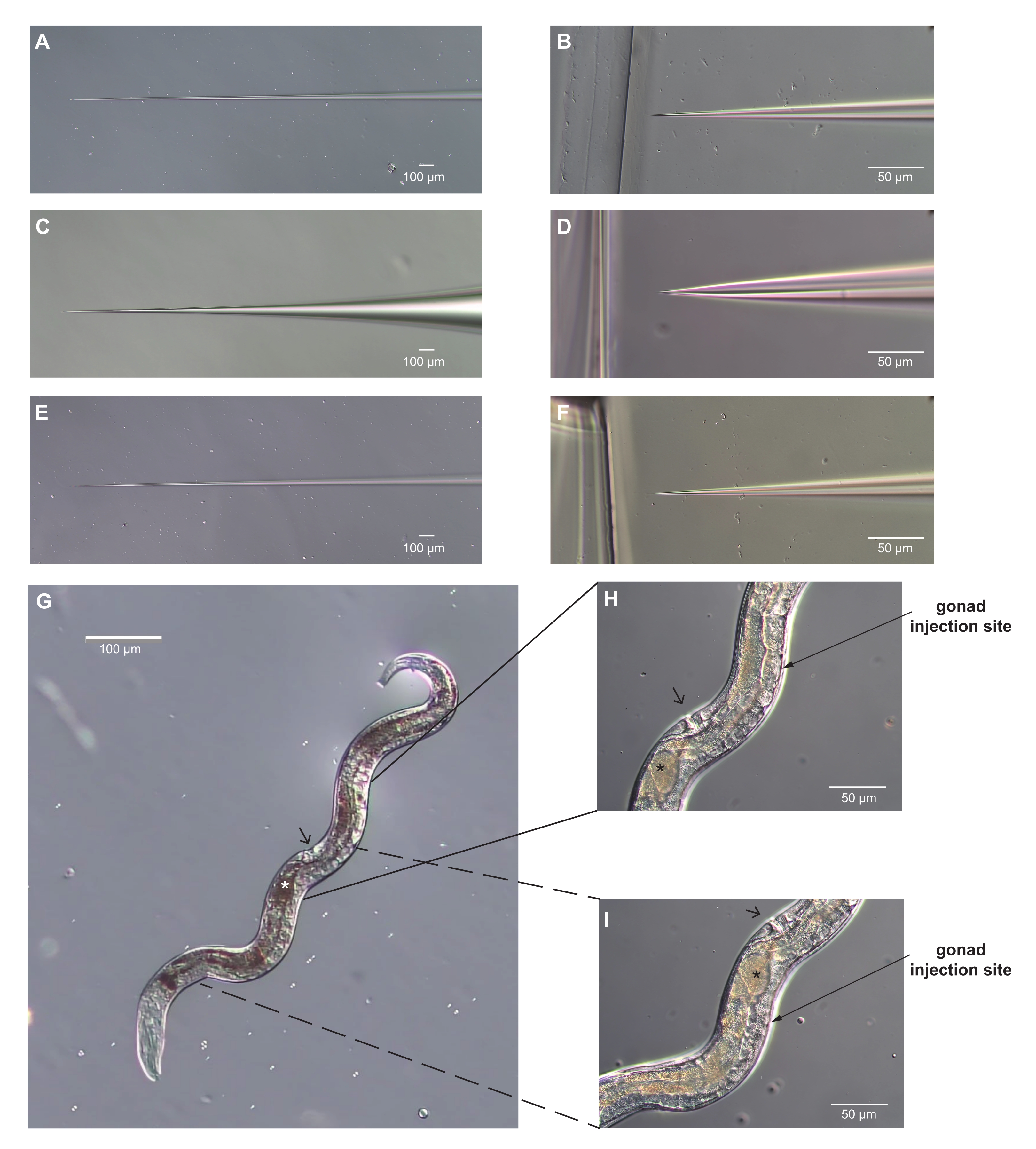
Figura 3: Identificati aghi per microiniezione e una femmina adulta di Strongyloides stercoralis con siti ottimali per la microiniezione. (A-F) Immagini di aghi per microiniezione. (A-B) Il cono dell'albero (A) e la punta (B) di un ago che è correttamente modellato per la microiniezione. La punta è abbastanza affilata da perforare la cuticola e abbastanza stretta da non causare danni eccessivi. (C-D) Il cono dell'albero (C) e la punta (D) di un ago per microiniezione che è modellato in modo errato per la microiniezione. La punta è troppo smussata e larga e causerà danni eccessivi al verme. (E-F) Il cono dell'albero (E) e la punta (F) di un ago che rischia di essere troppo lungo e sottile per funzionare per la microiniezione. La punta in F è molto simile alla punta in D. Tuttavia, l'albero è più stretto e troppo flessibile per perforare efficacemente la cuticola. Inoltre, aghi molto sottili si intasano facilmente. (G) Un'immagine dell'intero verme correttamente posizionato per la microiniezione, supponendo che l'ago arrivi da destra. Anterior è in basso e a sinistra; la vulva è indicata dalla punta della freccia. La gonade è visibile lungo il lato destro della femmina. Questa femmina ha un solo uovo nel suo utero (indicato dall'asterisco). (H, I) Viste ingrandite dei siti di microiniezione. L'angolo della freccia si avvicina all'angolo dell'ago per iniezione. La vulva può essere utilizzata come punto di riferimento; è sul lato opposto del verme dalle braccia della gonade. Le braccia della curva della gonade attorno all'intestino e le estremità con i nuclei divisori sono opposte alla vulva. (H) Il braccio posteriore della gonade; (I) il braccio anteriore. Uno o entrambi i bracci possono essere iniettati. Per H, I, le convenzioni sono come in G. Barre di scala = 50 μm (B, D, F, H, I); 100 μm (A, C, E, G). Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}
- Conservare gli aghi tirati in una capsula di Petri di plastica da 15 cm con un pezzo di nastro arrotolato per fissare gli aghi ed evitare l'accumulo di polvere sulle punte.
- Posizionare una goccia di 0,7 μL della miscela di microiniezione sull'estremità aperta dell'albero. Appendere l'ago perpendicolarmente a uno scaffale usando un pezzo di nastro arrotolato per riempire l'albero affusolato con la miscela entro 10 minuti. Preparare 2 aghi alla volta nel caso in cui il primo non funzioni.
6. Preparare il microscopio e rompere l'ago
NOTA: La microiniezione utilizza un microscopio invertito con obiettivi 5x e 40x dotato di una configurazione microiniettore per controllare il movimento dell'ago. Il microscopio invertito deve essere posizionato su un tavolo pesante o su un tavolo d'aria antivibrante per ridurre il rumore vibrazionale. Il portaago del microiniettore è collegato al gas azoto che applica la pressione necessaria per erogare la miscela di microiniezione. Un microscopio di dissezione più piccolo nelle vicinanze viene utilizzato per trasferire i vermi.
- Impostare la pressione del serbatoio del gas su ~ 40-60 psi per rompere l'ago e su ~ 30-50 psi per la microiniezione, a seconda del flusso del liquido.
- Sul microscopio di dissezione, coprire il frammento di vetro sul coperchio del pad di microiniezione con olio di alocarbonio utilizzando un plettro a vite senza fine di platino standard.
- Posizionare il coperchio del pad di microiniezione sul telescopio di microiniezione e individuare il frammento di vetro coperto di olio. Allineare il frammento di vetro in modo tale che un bordo sia perpendicolare alla direzione dell'ago per fungere da superficie utilizzata per rompere l'ago.
- Verificare che l'ago non abbia bolle o detriti nell'albero conico utilizzando il microscopio di dissezione. Quindi, fissare l'ago 1-1,5 cm nel supporto pressurizzato.
- Posizionare la punta dell'ago al centro del campo visivo del microscopio a occhio. Quindi, sotto basso ingrandimento, posizionare la punta dell'ago nel campo visivo, perpendicolare al lato del frammento di vetro.
- Passare all'alta potenza e allineare la punta dell'ago con il bordo del vetro, vicino ma senza toccarlo.
NOTA: Una volta tirati, gli aghi vengono fusi chiusi. - Per rompere la punta dell'ago per consentire il flusso del liquido, picchiettarlo delicatamente sul lato del pezzo di vetro mentre si applica una pressione continua dal gas (Figura supplementare S1). Una volta che il liquido inizia a fluire, controllare la forma della punta e assicurarsi che sia affilata con liquido che scorre facilmente.
NOTA: se il liquido scorre troppo velocemente o l'estremità è troppo smussata, i vermi saranno danneggiati durante la microiniezione (Video 1 e Figura 3A-F). - Quando il liquido scorre bene dall'ago, spostare il vetrino di microiniezione nell'ambito di dissezione e posizionare gocce di olio di alocarburi da 1-2 μL sul tampone di agar per il posizionamento dei vermi.
- Trasferire 20-30 Strongyloides giovani adulti in una piastra NGM al 2% senza batteri per almeno 5 minuti per rimuovere i batteri superficiali in eccesso e selezionare singoli vermi per la microiniezione. Aggiungere più vermi alla piastra NGM secondo necessità durante l'iniezione.
7. Microiniezione di Strongyloides
- Usa una piccola quantità di olio di alocarburi su un verme per selezionare una giovane femmina adulta di Strongyloides con 1-4 uova nella sua gonade dalla piastra NGM al 2% senza batteri.
- Trasferire il verme in una piccola goccia di olio sul cuscinetto di agar. Usando il worm pick, posiziona delicatamente il worm in modo che non sia arrotolato e la gonade sia visibile e di facile accesso. Notare la direzione della gonade (Figura 3G).
- Posizionare il verme nel campo visivo del microscopio a microiniezione. Assicurarsi che la gonade si trovi sullo stesso lato dell'ago e posizionata in modo che l'ago entri in contatto con la gonade con una leggera angolazione (Figura 3H,I).
- Portare la punta dell'ago sul lato del verme nello stesso piano focale. Mirare al braccio gondico vicino al centro del verme. Utilizzare il microiniettore per inserire delicatamente l'ago nella gonade (Video 2).
- Applicare immediatamente una pressione sull'ago per riempire delicatamente l'intero braccio gondico con la soluzione di DNA. Determinare ad occhio quando è stato iniettato abbastanza liquido (Video 2).
NOTA: potrebbero essere necessari fino a 2 s per riempire la gonade. - Rimuovere l'ago e controllare per determinare che la ferita si chiuda.
NOTA: il verme è troppo danneggiato per produrre progenie se la gonade sporge attraverso la parete del corpo (Video supplementare S1). - Ripeti con l'altro braccio della gonade se è visibile.
- Al termine dell'iniezione, verificare rapidamente che l'ago non sia intasato applicando una pressione con la punta dell'ago sul tampone di agar. Trasferire il vetrino con il verme iniettato al microscopio di dissezione.
- Per recuperare il verme iniettato, prima posiziona alcune gocce di BU sul verme per farlo galleggiare fuori dal cuscinetto di agar.
- Raccogli una piccola quantità di batteri HB101 su un plettro di vermi. Toccare il verme con i batteri aderenti sul plettro del verme per rimuoverlo dal liquido.
- Trasferire delicatamente il verme nella piastra di recupero , una piastra NGM al 2% contenente un prato HB101.
NOTA: il worm dovrebbe iniziare a eseguire la scansione in pochi minuti. - Dopo che alcune femmine sono state iniettate, aggiungere alcuni maschi non iniettati dalla piastra di origine.
NOTA: un minimo di un maschio per cinque femmine è una buona linea di base; è preferito un eccesso di maschi. - Ripeti tutti i passaggi fino a quando non sono state iniettate abbastanza femmine per l'esperimento.
- Lasciare gli adulti sulla piastra di recupero per almeno 1 ora dopo l'iniezione per consentire ai vermi di recuperare e accoppiarsi.
8. Recupero e coltura di Strongyloides iniettati
- Raccogli le feci durante la notte da animali ospiti non infetti, utilizzando lo stesso protocollo degli animali infetti.
- Mescolare le feci non infestate con una piccola quantità di carbone (feci al rapporto carbone di circa 2 a 1 per queste piastre).
- Versare una piccola quantità di miscela fecale-carbone in una capsula di Petri di 6 cm rivestita con carta da filtro umida. Assicurarsi che il mix non tocchi il coperchio del piatto.
- Inondare la piastra di recupero con BU. Utilizzando un pipetto impostato a 3 μL, trasferire i vermi alle feci nella piastra fecale-carbone. Posiziona i vermi direttamente sulle feci, non sul carbone.
- Verificare che gli adulti siano sulla piastra fecale-carbone usando un cannocchiale di dissezione.
- Per coltivare i vermi, posizionare la piastra in una camera umidificata, cioè una scatola di plastica con un coperchio aderente rivestito con asciugamani di carta umidi.
NOTA: Dopo 2 giorni, ci sarà un mix di stadi larvali. Dopo 5 giorni, la maggior parte delle larve si sarà sviluppata in iL3s; alcune larve più giovani rimarranno. Dopo 7 giorni, tutte le larve dovrebbero essere iL3s.
9. Raccolta e screening delle larve F 1 per recuperare transgenici/knockout
- Utilizzando una configurazione di Baermann, raccogliere le larve dalle piastre di coltivazione di carbone fecale su piccola scala post-iniezione. Per ottenere il maggior numero possibile di larve, attendere almeno 2 ore prima di recuperare i vermi dall'apparato di Baermann.
- Concentrare le larve in un tubo di centrifuga da 15 ml come nei passaggi 4.10-4.14 e trasferire le larve in un piccolo vetro per orologi con BU.
- Se le larve verranno utilizzate per esperimenti comportamentali, utilizzare piastre NGM al 2% con un prato spesso di HB101 per lo screening.
- Trasferire 20-30 larve al prato HB101.
NOTA: I batteri rallenteranno il movimento delle larve. - Sotto un microscopio di dissezione a fluorescenza, identificare le larve che esprimono il transgene di interesse. Usa un worm pick per selezionare le larve transgeniche e spostarle su un piccolo vetro per orologi con BU.
- Utilizzare una nuova piastra HB101 per schermare un altro piccolo lotto di larve. Quando sono state raccolte abbastanza larve per uso sperimentale, trattare le piastre HB101 e i vermi in eccesso con iodio diluito (50% iodio di Lugol diluito in acqua) e scartarli come rifiuti a rischio biologico. In alternativa, uccidere i vermi in eccesso usando un detergente concentrato per canili contenente cloruri di alchil benzil ammonio.
- Usa immediatamente i vermi o lasciali in un vetro da orologio poco profondo in una piccola quantità di BU durante la notte.
NOTA: i vermi possono diventare ipossici se il liquido è troppo profondo. È possibile che lasciare le larve nella BU durante la notte possa influenzare determinati comportamenti; pertanto, utilizzare le larve per esperimenti comportamentali entro 6 ore.
- Trasferire 20-30 larve al prato HB101.
- Se le larve saranno utilizzate per la microscopia e non per saggi comportamentali, immobilizzare i vermi con paralisi della nicotina in modo reversibile per lo screening.
- Usando una lama di rasoio, segna una griglia sul fondo di plastica di una piastra di chemiotassi da10 cm 12 per rendere più facile tenere traccia della posizione dei vermi sulla piastra.
- Lascia cadere ~ 3 μL di larve in BU in un quadrato sulla griglia. Riempi tutti i quadrati necessari. Non usare quelli vicino ai bordi della piastra, poiché le larve potrebbero strisciare ai lati della piastra.
- Aggiungere 15-20 μL gocce di nicotina all'1% in acqua alle gocce di verme.
NOTA: Dopo 4 minuti, i vermi saranno paralizzati. - Schermare i vermi usando un microscopio di dissezione a fluorescenza.
- Usa un worm pick per trasferire le larve transgeniche in un piccolo vetro per orologi con 1-2 ml di BU.
NOTA: Le larve saranno paralizzate per diverse ore e possono essere facilmente montate su vetrini per microscopio per microscopia. Se lasciato durante la notte in BU, gli iL3 si riprenderanno e potranno essere utilizzati per alcuni test o infezione dell'ospite nei mammiferi. Tuttavia, la paralisi della nicotina e l'incubazione notturna nella BU possono influenzare determinati comportamenti.
Risultati
Se l'esperimento ha avuto successo, le larve F1 esprimeranno il fenotipo transgenico e/o mutante di interesse (Figura 4). Tuttavia, i tassi di trasformazione sono molto variabili e dipendono dai costrutti, dalla salute dei vermi, dalle condizioni di coltura post-iniezione e dall'abilità dello sperimentatore. In generale, un esperimento di successo produrrà >15 F1 larve per femmina iniettata e un tasso di trasformazione di >3% per i marcatori fluorescenti. Se il numero...
Discussione
Questo protocollo di microiniezione descrive in dettaglio i metodi per introdurre costrutti per la transgenesi e la mutagenesi mediata da CRISPR / Cas9 in S. stercoralis e S. ratti. Sia per S. stercoralis che per S. ratti, la sopravvivenza post-iniezione e il tasso di transgenesi o mutagenesi sono soggetti a diverse variabili che possono essere messe a punto.
La prima considerazione critica per una transgenesi di successo è come sono costruiti i transgeni ...
Divulgazioni
Gli autori non dichiarano conflitti di interesse.
Riconoscimenti
pPV540 e pPV402 sono stati regali gentili del Dr. James Lok dell'Università della Pennsylvania. Ringraziamo Astra Bryant per i commenti utili sul manoscritto. Questo lavoro è stato finanziato da un Burroughs-Wellcome Fund Investigators in the Pathogenesis of Disease Award, un Howard Hughes Medical Institute Faculty Scholar Award e National Institutes of Health R01 DC017959 (E.A.H.).
Materiali
| Name | Company | Catalog Number | Comments |
| (−)-Nicotine, ≥99% (GC), liquid | Sigma-Aldrich | N3876-5ML | nicotine for paralyzing worms |
| 3" iron C-clamp, 3" x 2" (capacity x depth) | VWR | 470121-790 | C-clamp to secure setup to bench top |
| Agarose LE | Phenix | RBA-500 | agarose for slides |
| Bone char, 4 lb pail, 10 x 28 mesh | Ebonex | n/a | charcoal for fecal-charcoal cultures |
| Bone char, granules, 10 x 28 mesh | Reade | bonechar10x28 | charcoal for fecal-cultures (alternative to the above) |
| Coarse micromanipulator | Narishige | MMN-1 | coarse micromanipulator |
| Corning Costar Spin-X centrifuge tube filters | Fisher | 07-200-385 | microfilter column |
| Cover glass, 48 x 60 mm, No. 1 thickness | Brain Research Lab | 4860-1 | coverslips (48 x 60 mm) |
| Deep Petri dishes, heavy version with 6 vents, 100 mm diameter | VWR | 82050-918 | 10 cm Petri dishes (for fecal-charcoal cultures) |
| Eisco retort base w/ rod | Fisher | 12-000-101 | stand for Baermann apparatus |
| Eppendorf FemtoJet microinjector microloader tips | VWR | 89009-310 | for filling microinjection needles |
| Fisherbrand absorbent underpads | Fisher | 14-206-62 | bench paper (for prepping) |
| Fisherbrand Cast-Iron Rings | Fisher | 14-050CQ | Baermann o-ring |
| Fisherbrand tri-cornered polypropylene beakers | Fisher | 14-955-111F | Plastic beaker (for mixing) |
| Fisherbrand tri-cornered polypropylene beakers | Fisher | 14-955-111F | Plastic beaker (for catch bucket/water bucket) |
| Fisherbrand tri-cornered polypropylene beakers | Fisher | 14-955-111F | Plastic beaker (x2) (to make holder) |
| Gorilla epoxie in syringe | McMaster-Carr | 7541A51 | glue (to attach tubing) |
| Halocarbon oil 700 | Sigma-Aldrich | H8898-50ML | halocarbon oil |
| High-temperature silicone rubber tubing for food and beverage, 1/2" ID, 5/8" OD | McMaster-Carr | 3038K24 | tubing (for funnel) |
| KIMAX funnels, long stem, 60° Angle, Kimble Chase | VWR | 89001-414 | Baermann funnel |
| Kimberly-Clark Professional Kimtech Science benchtop protectors | Fisher | 15-235-101 | bench paper (for prepping) |
| Leica stereomicroscope with fluorescence | Leica | M165 FC | GFP stereomicroscope for identifying and sorting transgenic worms |
| microINJECTOR brass straight arm needle-holder | Tritech | MINJ-4 | microinjection needle holder |
| microINJECTOR system | Tritech | MINJ-1 | microinjection system |
| Mongolian Gerbils | Charles River Laboratories | 213-Mongolian Gerbil | gerbils for maintenance of S. stercoralis, male 4-6 weeks |
| Nasco Whirl-Pak easy-to-close bags, 18 oz | VWR | 11216-776 | Whirl-Pak sample bags |
| Nylon tulle (mesh) | Jo-Ann Fabrics | zprd_14061949a | nylon mesh for Baermann holder |
| Platinum wire, 36 Gauge, per inch | Thomas Scientific | 1233S72 | platinum/iridium wire for worm picks |
| Puritan tongue depressors, 152 mm (L) x 17.5 mm (W) | VWR | 62505-007 | wood sticks (for mixing samples) |
| QIAprep Spin Miniprep Kit (250) | QIAGEN | 27106 | QIAGEN miniprep kit |
| Rats-Long Evans | Envigo | 140 HsdBlu:LE Long Evans | rats for maintenance of S. ratti, female 4-6 weeks |
| Rats-Sprague Dawley | Envigo | 002 Hsd:Sprague Dawley SD | rats for maintenance of S. ratti, female 4-6 weeks |
| Really Useful Boxes translucent storage boxes with lids, 1.6 L capacity, 7-5/8" x 5-5/16" x 4-5/16" | Office Depot | 452369 | plastic boxes for humidified chamber |
| Shepherd techboard, 8 x 16.5 inches | Newco | 999589 | techboard |
| Stainless steel raised wire floor | Ancare | R20SSRWF | wire cage bottoms |
| StalkMarket compostable cutlery spoons, 6", white, pack of 1,000 | Office Depot | 9587303 | spoons |
| Stender dish, stacking type, 37 x 25 mm | Carolina (Science) | 741012 | watch glasses (small, round) |
| Stereomicroscope | Motic | K-400 LED | dissecting prep scope |
| Storage tote, color clear/white, outside height 4-7/8 in, outside length 13-5/8 in, Sterilite | Grainger | 53GN16 | plastic boxes for humidified chamber |
| Sutter P-30 micropipette puller | Sutter | P-30/P | needle puller with platinum/iridium filament |
| Syracuse watch glasses | Fisher | S34826 | watch glasses (large, round) |
| Thermo Scientific Castaloy fixed-angle clamps | Fisher | 05-769-2Q | funnel clamps (2x) |
| Three-axis hanging joystick oil hydrolic micromanipulator | Narishige | MM0-4 | fine micromanipulator |
| United Mohr pinchcock clamps | Fisher | S99422 | Pinch clamps (2x) |
| Vented, sharp-edge Petri dishes (60 mm diameter) | Tritech Research | T3308P | 6 cm Petri dishes (for small-scale fecal-charcoal cultures) |
| VWR light-duty tissue wipers | VWR | 82003-820 | lining for Baermann holder |
| watch glass, square, 1-5/8 in | Carolina (Science) | 742300 | watch glasses (small, square) |
| Whatman qualitative grade plain circles, grade 1, 5.5 cm diameter | Fisher | 09-805B | filter paper (for 6 cm Petri dishes) |
| Whatman qualitative grade plain circles, grade 1, 9 cm diameter | Fisher | 09-805D | filter paper (for 10 cm Petri dishes) |
| World Precision Instrument borosilicate glass capillary, 1.2 mm x 4 in | Fisher | 50-821-813 | glass capillaries for microinjection needles |
| X-Acto Knives, No. 1 Knife With No. 11 Blade | Office Depot | 238816 | X-Acto knives without blades to hold worm picks |
| Zeiss AxioObserver A1 | Zeiss | n/a | inverted microscope |
Riferimenti
- Krolewiecki, A. J., et al. A public health response against Strongyloides stercoralis: time to look at soil-transmitted helminthiasis in full. PLoS Neglected Tropical Diseases. 7 (5), 2165 (2013).
- Buonfrate, D., et al. The global prevalence of Strongyloides stercoralis infection. Pathogens. 9 (6), 468 (2020).
- Castelletto, M. L., Gang, S. S., Hallem, E. A. Recent advances in functional genomics for parasitic nematodes of mammals. Journal of Experimental Biology. 223, 206482 (2020).
- Evans, T. C., et al. Transformation and microinjection. WormBook. , (2006).
- Lok, J. B., Unnasch, T. R., et al. Transgenesis in animal parasitic nematodes: Strongyloides spp. and Brugia spp. WormBook. , (2013).
- Shao, H. G., Li, X. S., Lok, J. B. Heritable genetic transformation of Strongyloides stercoralis by microinjection of plasmid DNA constructs into the male germline. International Journal for Parasitology. 47 (9), 511-515 (2017).
- Schafer, T. W., Skopic, A. Parasites of the small intestine. Current Gastroenterology Reports. 8 (4), 312-320 (2006).
- Stiernagle, T. Maintenance of C. elegans. The C. elegans Research Community, WormBook. , (2006).
- Gang, S. S., et al. Targeted mutagenesis in a human-parasitic nematode. PLoS Pathogens. 13 (10), 1006675 (2017).
- Lok, J. B. Strongyloides stercoralis: a model for translational research on parasitic nematode biology. The C. elegans Research Community, WormBook. , (2007).
- Hawdon, J. M., Schad, G. A. Long-term storage of hookworm infective larvae in buffered saline solution maintains larval responsiveness to host signals. Proceedings of the Helminthological Society of Washington (USA). 58 (1), 140-142 (1991).
- Bargmann, C. I., Hartwieg, E., Horvitz, H. R. Odorant-selective genes and neurons mediate olfaction in C. elegans. Cell. 74 (3), 515-527 (1993).
- Junio, A. B., et al. Strongyloides stercoralis: cell- and tissue-specific transgene expression and co-transformation with vector constructs incorporating a common multifunctional 3' UTR. Experimental Parasitology. 118 (2), 253-265 (2008).
- Gang, S. S., et al. Chemosensory mechanisms of host seeking and infectivity in skin-penetrating nematodes. Proceedings of the National Academy of Sciences of the United States of America. 117 (30), 17913-17923 (2020).
- Bryant, A. S., et al. A critical role for thermosensation in host seeking by skin-penetrating nematodes. Current Biology. 28 (14), 2338-2347 (2018).
- Lok, J. B. Nucleic acid transfection and transgenesis in parasitic nematodes. Parasitology. 139 (5), 574-588 (2012).
- Shao, H., et al. Transposon-mediated chromosomal integration of transgenes in the parasitic nematode Strongyloides ratti and establishment of stable transgenic lines. PLoS Pathogens. 8 (8), 1002871 (2012).
- Lok, J. piggyBac: a vehicle for integrative DNA transformation of parasitic nematodes. Mobile Genetic Elements. 3 (2), 24417 (2013).
- Li, X., et al. Successful transgenesis of the parasitic nematode Strongyloides stercoralis requires endogenous non-coding control elements. International Journal for Parasitology. 36 (6), 671-679 (2006).
- Bryant, A. S., Hallem, E. A. The Wild Worm Codon Adapter: a web tool for automated codon adaptation of transgenes for expression in non-Caenorhabditis nematodes. G3. 3 (7), (2021).
- Crane, M., et al. In vivo measurements reveal a single 5'-intron is sufficient to increase protein expression level in Caenorhabditis elegans. Scientific Reports. 9 (1), 9192 (2019).
- Han, Z., et al. Improving transgenesis efficiency and CRISPR-associated tools through codon optimization and native intron addition in Pristionchus nematodes. Genetics. 216 (4), 947-956 (2020).
- Adams, S., Pathak, P., Shao, H., Lok, J. B., Pires-daSilva, A. Liposome-based transfection enhances RNAi and CRISPR-mediated mutagenesis in non-model nematode systems. Scientific Reports. 9 (1), 483 (2019).
- Dulovic, A., Puller, V., Streit, A. Optimizing culture conditions for free-living stages of the nematode parasite Strongyloides ratti. Experimental Parasitology. 168, 25-30 (2016).
- Harvey, S. C., Gemmill, A. W., Read, A. F., Viney, M. E. The control of morph development in the parasitic nematode Strongyloides ratti. Proceedings of the Royal Society B: Biological Sciences. 267 (1457), 2057-2063 (2000).
- Kim, A., Pyykko, I. Size matters: versatile use of PiggyBac transposons as a genetic manipulation tool. Molecular and Cellular Biochemistry. 354 (1-2), 301-309 (2011).
- Lok, J. B., Shao, H., Massey, H. C., Li, X. Transgenesis in Strongyloides and related parasitic nematodes: historical perspectives, current functional genomic applications and progress towards gene disruption and editing. Parasitology. 144 (3), 327-342 (2017).
- Farboud, B., Meyer, B. J. Dramatic enhancement of genome editing by CRISPR/Cas9 through improved guide RNA design. Genetics. 199 (4), 959-971 (2015).
- Cheong, M. C., et al. Identification of a nuclear receptor/coactivator developmental signaling pathway in the nematode parasite Strongyloides stercoralis. Proceedings of the National Academy of Sciences of the United States of America. 118 (8), 2021864118 (2021).
- Nolan, T. J., Megyeri, Z., Bhopale, V. M., Schad, G. A. Strongyloides stercoralis: the first rodent model for uncomplicated and hyperinfective strongyloidiasis, the Mongolian gerbil (Meriones unguiculatus). Journal of Infectious Diseases. 168 (6), 1479-1484 (1993).
- Li, X., et al. Transgenesis in the parasitic nematode Strongyloides ratti. Molecular and Biochemical Parasitology. 179 (2), 114-119 (2011).
- Viney, M. E. Exploiting the life cycle of Strongyloides ratti. Parasitology Today. 15 (6), 231-235 (1999).
- Stoltzfus, J. D., Massey, H. C., Nolan, T. J., Griffith, S. D., Lok, J. B. Strongyloides stercoralis age-1: a potential regulator of infective larval development in a parasitic nematode. PLoS ONE. 7 (6), 38587 (2012).
- Castelletto, M. L., Massey, H. C., Lok, J. B. Morphogenesis of Strongyloides stercoralis infective larvae requires the DAF-16 ortholog FKTF-1. PLoS Pathogens. 5 (4), 1000370 (2009).
- Douglas, B., et al. Transgenic expression of a T cell epitope in Strongyloides ratti reveals that helminth-specific CD4+ T cells constitute both Th2 and Treg populations. PLoS Pathogens. 17 (7), 1009709 (2021).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneEsplora altri articoli
This article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati