
Un abonnement à JoVE est nécessaire pour voir ce contenu. Connectez-vous ou commencez votre essai gratuit.
Method Article
Génération de transgéniques et de Knockouts chez les espèces de strongyloïdes par microinjection
Dans cet article
Résumé
La boîte à outils génomique fonctionnelle pour les nématodes parasites Strongyloides stercoralis et Strongyloides ratti comprend la transgénèse, la mutagénèse médiée par CRISPR / Cas9 et l’ARNi. Ce protocole montrera comment utiliser la microinjection intragonadienne pour introduire des transgènes et des composants CRISPR dans S. stercoralis et S. ratti.
Résumé
Le genre Strongyloides se compose de plusieurs espèces de nématodes pénétrant dans la peau avec différentes gammes d’hôtes, y compris Strongyloides stercoralis et Strongyloides ratti. S. stercoralis est un nématode parasite humain, pénétrant dans la peau, qui infecte environ 610 millions de personnes, tandis que le parasite du rat S. ratti est étroitement lié à S. stercoralis et est souvent utilisé comme modèle de laboratoire pour S. stercoralis. S. stercoralis et S. ratti se prêtent facilement à la génération de transgéniques et de knockouts grâce à la technique exogène d’administration d’acides nucléiques de la microinjection intragonadale et, en tant que tels, sont devenus des systèmes modèles pour d’autres helminthes parasites qui ne se prêtent pas encore à cette technique.
Les adultes parasites Strongyloides habitent l’intestin grêle de leur hôte et libèrent de la progéniture dans l’environnement via les matières fécales. Une fois dans l’environnement, les larves se développent en adultes libres, qui vivent dans les excréments et produisent une progéniture qui doit trouver et envahir un nouvel hôte. Cette génération environnementale est unique à l’espèce Strongyloides et sa morphologie est suffisamment similaire à celle du nématode vivant librement Caenorhabditis elegans pour que les techniques développées pour C. elegans puissent être adaptées pour être utilisées avec ces nématodes parasites, y compris la microinjection intragonadaire. En utilisant la microinjection intragonadienne, une grande variété de transgènes peuvent être introduits dans Strongyloides. Les composants CRISPR/Cas9 peuvent également être microinjectés pour créer des larves mutantes de Strongyloides . Ici, la technique de microinjection intragonadale en Strongyloides, y compris la préparation d’adultes libres, la procédure d’injection et la sélection de la progéniture transgénique, est décrite. Des images de larves transgéniques de Strongyloides créées à l’aide de la mutagénèse CRISPR/Cas9 sont incluses. L’objectif de cet article est de permettre à d’autres chercheurs d’utiliser la microinjection pour créer des Strongyloides transgéniques et mutants.
Introduction
Strongyloides stercoralis a longtemps été négligé comme un agent pathogène humain important par rapport aux ankylostomes plus largement reconnus et au ver rond Ascaris lumbricoides1. Des études antérieures sur la charge de vers ont souvent gravement sous-estimé la prévalence de S. stercoralis en raison de la faible sensibilité des méthodes de diagnostic courantes pour S. stercoralis2. Au cours des dernières années, des études épidémiologiques basées sur des outils de diagnostic améliorés ont estimé que la prévalence réelle des infections à S. stercoralis est beaucoup plus élevée que ce qui avait été rapporté précédemment, soit environ 610 millions de personnes dans le monde2.
S. stercoralis et d’autres espèces de Strongyloides, y compris le parasite de rat étroitement apparenté et le modèle de laboratoire commun S. ratti, ont un cycle de vie inhabituel qui est avantageux pour les études génomiques expérimentales car il se compose à la fois de générations parasites et libres (environnementales)3 (Figure 1). Plus précisément, S. stercoralis et S. ratti peuvent traverser une seule génération de vie libre. La génération de la vie libre se compose de larves post-parasites qui se développent en mâles et femelles adultes libres; toutes les descendants des adultes libres se développent en larves infectieuses, qui doivent infecter un hôte pour poursuivre le cycle de vie. De plus, cette génération environnementale ou libre peut être manipulée expérimentalement en laboratoire. Étant donné que les adultes Strongyloides vivant en liberté et les adultes de C. elegans partagent une morphologie similaire, des techniques telles que la microinjection intragonadale qui ont été développées à l’origine pour C. elegans peuvent être adaptées pour être utilisées avec des Strongyloides 4,5 adultes vivant en liberté. Alors que l’ADN est généralement introduit chez les femelles adultes libres, les mâles et les femelles de Strongyloides peuvent être microinjectés6. Ainsi, des outils génomiques fonctionnels sont disponibles pour interroger de nombreux aspects de la biologie des Strongyloides. D’autres nématodes parasites n’ont pas de génération libre et, par conséquent, ne se prêtent pas aussi facilement aux techniques génomiques fonctionnelles3.

Figure 1 : Cycle de vie de Strongyloides stercoralis. Les femelles parasites de S. stercoralis habitent l’intestin grêle de leurs hôtes mammifères (humains, primates non humains, chiens). Les femelles parasites se reproduisent par parthénogenèse et pondent des œufs dans l’intestin grêle. Les œufs éclosent encore à l’intérieur de l’hôte en larves post-parasites, qui sont ensuite passées dans l’environnement avec des matières fécales. Si les larves post-parasites sont des mâles, elles se développent en mâles adultes libres. Si les larves postparasitaires sont des femelles, elles peuvent se développer en femelles adultes libres (développement indirect) ou en larves infectieuses de troisième stade (iL3s; développement direct). Les mâles et les femelles libres se reproduisent sexuellement pour créer une progéniture qui est contrainte de devenir des iL3. Dans certaines conditions, S. stercoralis peut également subir une auto-infection, dans laquelle certaines des larves postparasitaires restent à l’intérieur de l’intestin hôte plutôt que de passer dans l’environnement dans les matières fécales. Ces larves peuvent se développer en larves auto-infectieuses (L3a) à l’intérieur de l’hôte, pénétrer à travers la paroi intestinale, migrer à travers le corps et éventuellement retourner dans l’intestin pour devenir des adultes reproducteurs. Le cycle de vie de S. ratti est similaire, sauf que S. ratti infecte les rats et n’a pas de cycle auto-infectieux. La génération environnementale est essentielle à l’utilisation des espèces de Strongyloides pour les études génétiques. Les femelles adultes libres (P0) peuvent être microinjectées; leur progéniture, qui deviendront toutes des iL3, sont les transgéniques potentiels F1. Ce chiffre a été modifié à partir de Castelletto et al. 3. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
S. stercoralis partage de nombreux aspects de sa biologie avec d’autres nématodes gastro-intestinaux parasitaires humains, y compris l’invasion de l’hôte et la modulation immunitaire de l’hôte. Par exemple, les ankylostomes parasitaires humains des genres Necator et Ancylostoma infectent également par pénétration cutanée, naviguent de manière similaire dans le corps et résident finalement comme des adultes parasites dans l’intestin grêle7. Ainsi, de nombreux nématodes gastro-intestinaux utilisent probablement des comportements sensoriels courants et des techniques d’évasion immunitaire. En conséquence, les connaissances glanées auprès de Strongyloides compléteront les découvertes chez d’autres nématodes moins génétiquement traitables et conduiront à une compréhension plus complète de ces parasites complexes et importants.
Ce protocole de micro-injection décrit la méthode d’introduction de l’ADN dans les femelles adultes strongyloïdes vivant librement pour fabriquer une progéniture transgénique et mutante. Les exigences en matière d’entretien de la souche, y compris le calendrier de développement des vers adultes pour les microinjections et la collecte de la progéniture transgénique, sont décrites. Des protocoles et une démonstration de la technique complète de micro-injection, ainsi que des protocoles pour la culture et le dépistage de la progéniture transgénique, sont inclus, ainsi qu’une liste de tous les équipements et consommables nécessaires.
Protocole
NOTE: Les gerbilles ont été utilisées pour passer S. stercoralis, et les rats ont été utilisés pour passer S. ratti. Toutes les procédures ont été approuvées par le Bureau de surveillance de la recherche animale de l’UCLA (Protocole n ° 2011-060-21A), qui adhère aux normes AAALAC et au Guide pour le soin et l’utilisation des animaux de laboratoire. Les tâches suivantes doivent être effectuées au moins un jour avant la microinjection : culture de vers, préparation de tampons de micro-injection, création de constructions pour le mélange de micro-injection et propagation de bactéries (E. coli HB101) sur des plaques de milieu de croissance des nématodes (NGM) de 6 cm8. Les femelles libres ont besoin d’un minimum de 24 heures après la collecte fécale à 25 °C pour devenir de jeunes adultes avant de pouvoir être microinjectées. Les tampons de micro-injection doivent être complètement secs. Les plaques bactériennes doivent sécher et établir une petite pelouse.
1. Préparation des lames de micro-injection: au moins un jour avant l’injection
REMARQUE: Les vers sont montés sur des couvercles de micro-injection avec des coussinets de gélose sèche pour injection.
- Réglez un bloc chauffant à 90 °C.
- Ajouter 5 mL de ddH2O, puis 100 mg d’agarose dans un tube en verre borosilicate.
- Chauffer le mélange d’agarose dans le tube au-dessus d’une flamme jusqu’à ce que l’agarose soit dissoute.
- Placez le tube dans un bloc chauffant réglé à 90 °C pour maintenir l’agarose à l’état liquide.
- Déposer ~180 μL de la solution d’agarose sur un couvercle à l’aide d’une pipette Pasteur en verre ou d’une pipette avec une pointe en plastique. Laissez immédiatement tomber un deuxième couvercle sur le dessus pour aplatir l’agarose en un tampon mince.
- Après 5-10 s, retirez le couvercle supérieur en faisant glisser les deux. Déterminez sur quelle glissière se trouve le coussinet de gélose et posez-le face vers le haut.
- Sélectionnez un minuscule éclat de verre dans un couvercle cassé et appuyez doucement dans la gélose près du bord supérieur du tampon à l’aide d’une pince (figure supplémentaire S1).
- Continuez à fabriquer des tampons de micro-injection avec la solution d’agarose.
- Séchez les tampons d’agarose pendant la nuit sur le banc ou dans un four. Conserver dans la boîte à couvercle.
REMARQUE: Les tampons d’agarose peuvent être utilisés jusqu’à 2 mois, mais ne sont utilisés que pour une seule injection.
2. Cultiver des strongyloïdes pour obtenir des vers pour la microinjection: 1 à 2 jours avant l’injection
REMARQUE: Un protocole d’entretien de la souche peut être trouvé dans le matériel supplémentaire, qui comprend une description détaillée de la façon d’infecter les gerbilles et les rats avec des nématodes et de récolter des nématodes dans les excréments d’animaux infectés.
- Deux jours avant le jour de l’injection, placez les animaux infectés 9,10 dans des cages de collecte pendant la nuit.
- Le lendemain matin, ramassez les excréments infestés et faites des assiettes fécales-charbonde bois 9,10.
- Placez une plaque à 25 °C pendant 24 h pour permettre aux vers libres de se développer en jeunes adultes.
- La veille du jour de l’injection, placez les animaux hôtes non infectés dans des cages de collecte.
- Le jour de l’injection, collectez les matières fécales non infestées pour la culture post-injection.
3. Préparation du mélange de micro-injection : avant ou le jour de l’injection
REMARQUE: Le mélange de microinjection est constitué des plasmides d’intérêt dilués à la concentration souhaitée dans une solution saline tamponnée par ver (BU) (50 mM Na2HPO4, 22 mM KH2PO4, 70 mM NaCl)11.
- Déterminer la concentration des stocks de plasmides et la concentration souhaitée dans le mélange de microinjection (tableau 1).
| Mélange de micro-injection : construction de rapporteur | |||
| Composant | Concentration des stocks | Quantité | Concentration finale |
| pMLC30 gpa-3::gfp | 300 ng/μL | 1,7 μL | 50 ng/μL |
| BU | Na | 8,3 μL | Na |
| total | 10 μL | 50 ng/μL | |
| Mélange de micro-injection : mutagénèse CRISPR/Cas9 | |||
| Composant | Concentration des stocks | Quantité | Concentration finale |
| pMLC47 tax-4 sgRNA | 300 ng/μL | 2,7 μL | 80 ng/μL |
| pEY11 Plasmide Ss-tax-4 HDR | 400 ng/μL | 2,0 μL | 80 ng/μL |
| Plasmide pPV540 strCas9 | 350 ng/μL | 1,1 μL | 40 ng/μL |
| BU | Na | 4,2 μL | Na |
| total | 10 μL | 200 ng/μL | |
| Mélange de micro-injection : intégration piggyBac | |||
| Composant | Concentration des stocks | Quantité | Concentration finale |
| pMLC30 gpa3::gfp | 300 ng/μL | 2,0 μL | 60 ng/μL |
| Plasmide de transposase pPV402 | 450 ng/μL | 0,9 μL | 40 ng/μL |
| BU | Na | 7,1 μL | Na |
| total | 10 μL | 100 ng/μL | |
Tableau 1 : Exemples de mélanges de micro-injection. Les plasmides et les concentrations de trois exemples de mélanges de microinjection : un pour une construction de rapporteur gpa-3::GFP 10, un pour une perturbation médiée par CRISPR/Cas9 du locus Ss-tax-4 14,15 et une pour l’intégration médiée par piggyBac d’une construction Ss-gpa-3::GFP 13,17,18. strCas9 désigne le gène Cas9 optimisé pour le codon Strongyloides. Les concentrations finales énumérées sont couramment utilisées dans les mélanges de microinjection strongyloides.
- Diluer les plasmides dans BU à un volume total de 10-20 μL.
- Faire tourner le mélange à travers une colonne filtrante à 5 000 × g pendant 1 à 2 min.
- Utilisez immédiatement le mélange de micro-injection ou conservez-le à -20 °C pour une utilisation future.
4. Recueillir les Strongyloides jeunes adultes pour la microinjection: matin du jour de l’injection
- Installez l’appareil Baermann avec 1 plaque fécale-charbon de bois de jeunes strongyloïdes adultes (Figure 2).
REMARQUE: La plaque de charbon fécal peut contenir des larves infectieuses. L’équipement de protection individuelle se compose d’une blouse de laboratoire, de gants et d’une protection oculaire. Aucune peau ne doit être exposée entre le gant et la manche de la blouse de laboratoire.

Figure 2 : L’appareil de Baermann utilisé pour collecter les vers parasites des cultures10. Le contenu d’une plaque fécale-charbon de bois est placé au sommet d’une colonne d’eau tiède. Les vers migrent dans l’eau et s’accumulent au fond de l’entonnoir. (A) Pour installer l’appareil Baermann, le support de l’entonnoir Baermann est fixé au banc avec une pince en C. Un tube en caoutchouc attaché à l’extrémité de l’entonnoir est fermé avec des pinces à pincer et un seau de capture est placé sous le tube pour les gouttes. De l’eau tiède est ajoutée à l’entonnoir en verre. (B) Le support d’anneau en plastique pour le mélange fécal-charbon de bois est ensuite tapissé de 3 morceaux de tissus de laboratoire (à gauche). Un bâton en bois ou un dépresseur de langue (au milieu) est utilisé pour transférer le contenu d’une plaque de charbon fécal (à droite) dans le support de l’anneau en plastique. (C) Un gros plan du fond du porte-anneau en plastique pour le mélange fécal-charbon de bois, montrant la double couche de tulle de nylon tapissant le fond du support. (D) Le porte-charbon fécal est ensuite placé sur le dessus de l’entonnoir en verre. (E) Le tissu de laboratoire est humidifié avec de l’eau et fermé sur le mélange fécal-charbon de bois. Plus d’eau chaude est ajoutée pour submerger principalement le charbon fécal. (F) L’installation complète de Baermann, avec la culture fécale-charbon immergée sous l’eau chaude. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
- Installez un entonnoir en verre avec un tube de collecte en caoutchouc sur un support d’anneau à l’aide d’un joint torique et fixez-le avec une pince. Fermez le tube de collecte à l’aide de 2 pinces à pincement (Figure 2A).
- Placez un seau de capture sous l’entonnoir pour attraper les gouttes.
- Ajouter de l’eau tiède (environ 40 °C) à l’entonnoir à 5 cm sous le rebord. Vérifiez que le système ne fuit pas.
- Tapissez le support Baermann, un tamis fabriqué à partir de 2 anneaux en plastique avec 2 couches de filet en tulle de nylon fixé entre eux, avec 3 morceaux de tissu de laboratoire qui se chevauchent. Ajouter le mélange fécal-charbon de bois au support Baermann (Figure 2B,C).
- Placez le support Baermann avec le mélange fécal-charbon de bois dans l’entonnoir. Pliez les tissus autour du mélange fécal-charbon de bois et ajoutez suffisamment d’eau pour immerger la majeure partie du charbon fécal. Ne pas remplir à plus de 2 cm du bord de l’entonnoir (Figure 2D,E).
- Recouvrez l’entonnoir avec un couvercle de boîte de Petri en plastique de 15 cm pour contenir l’odeur. Étiquetez l’entonnoir au besoin (Figure 2F).
- Attendez 30 min à 1 h pour récupérer les vers de l’appareil Baermann.
- Tenez un tube de centrifugeuse de 50 mL sous le tube en caoutchouc au bas de l’entonnoir. Ouvrez soigneusement les pinces au fond pour distribuer 30 à 40 mL d’eau contenant des vers dans le tube de 50 mL.
- Transférer 15 mL d’eau Baermann contenant les vers dans un tube de centrifugeuse de 15 mL. Faire tourner le tube de centrifugeuse de 15 mL pendant 1 min à ~750 × g (lentement). Alternativement, laissez les vers se déposer par gravité pendant 10-15 min.
- Retirez le surnageant à ~2 mL et jetez le surnageant dans un récipient de déchets liquides avec de l’iode pour tuer les vers.
- Ajoutez plus d’eau Baermann au tube de collecte de 15 mL et répétez le spin. Retirez le surnageant à ~2 mL et jetez-le comme à l’étape 4.11.
- Répétez les étapes 4.11 et 4.12 jusqu’à ce que tous les vers soient collectés dans le tube de centrifugeuse de 15 mL. Après la dernière rotation, retirez autant d’eau que possible.
- Inspectez la pastille de vers (40-100 μL) au fond du tube. Si aucun ver n’est visible, attendez encore 1-2 h et essayez de collecter plus de vers de l’appareil Baermann.
- Transférer les vers dans le moins d’eau possible sur une plaque NGM de 6 cm 2% avec une pelouse d’E. coli HB101. Utilisez cette plaque comme plaque source pour la microinjection.
- Jetez le mélange fécal-charbon de bois en le traitant avec de l’iode dilué (une dilution de 50% de l’iode de Lugol dans l’eau), en l’enveloppant dans un film plastique pour attraper les gouttes et en le plaçant dans un conteneur à déchets biodangereux.
- Ajouter 10 mL d’iode dilué dans le seau de capture et y évacuer l’excès d’eau du Baermann.
- Lavez les composants réutilisables (l’entonnoir, le seau à prises, le support en plastique avec du tulle, le couvercle en plastique et les pinces) avec 10% d’eau de Javel et rincez abondamment.
5. Tirer et charger les aiguilles de micro-injection: juste avant l’injection
- Préparez les aiguilles de micro-injection en tirant des tubes capillaires en verre à l’aide d’un extracteur d’aiguilles.
REMARQUE: Les exemples de réglages pour un extracteur d’aiguilles commercial équipé d’un filament de platine / iridium de 3 mm sont Heat = 810-820, Pull = 800-820, micromètre = 2,5. - Visualisez les pointes sous un microscope à dissection. Si les aiguilles ont la forme désirée (Figure 3A-F), tirez sur 4 à 6 aiguilles (2 à 3 tubes capillaires). Pour obtenir la bonne forme d’aiguille, modifiez les réglages au besoin: ajustez les réglages Heat ou Pull de 10 et tirez de nouvelles aiguilles jusqu’à ce que la forme de la conicité et de l’arbre soit plus appropriée.

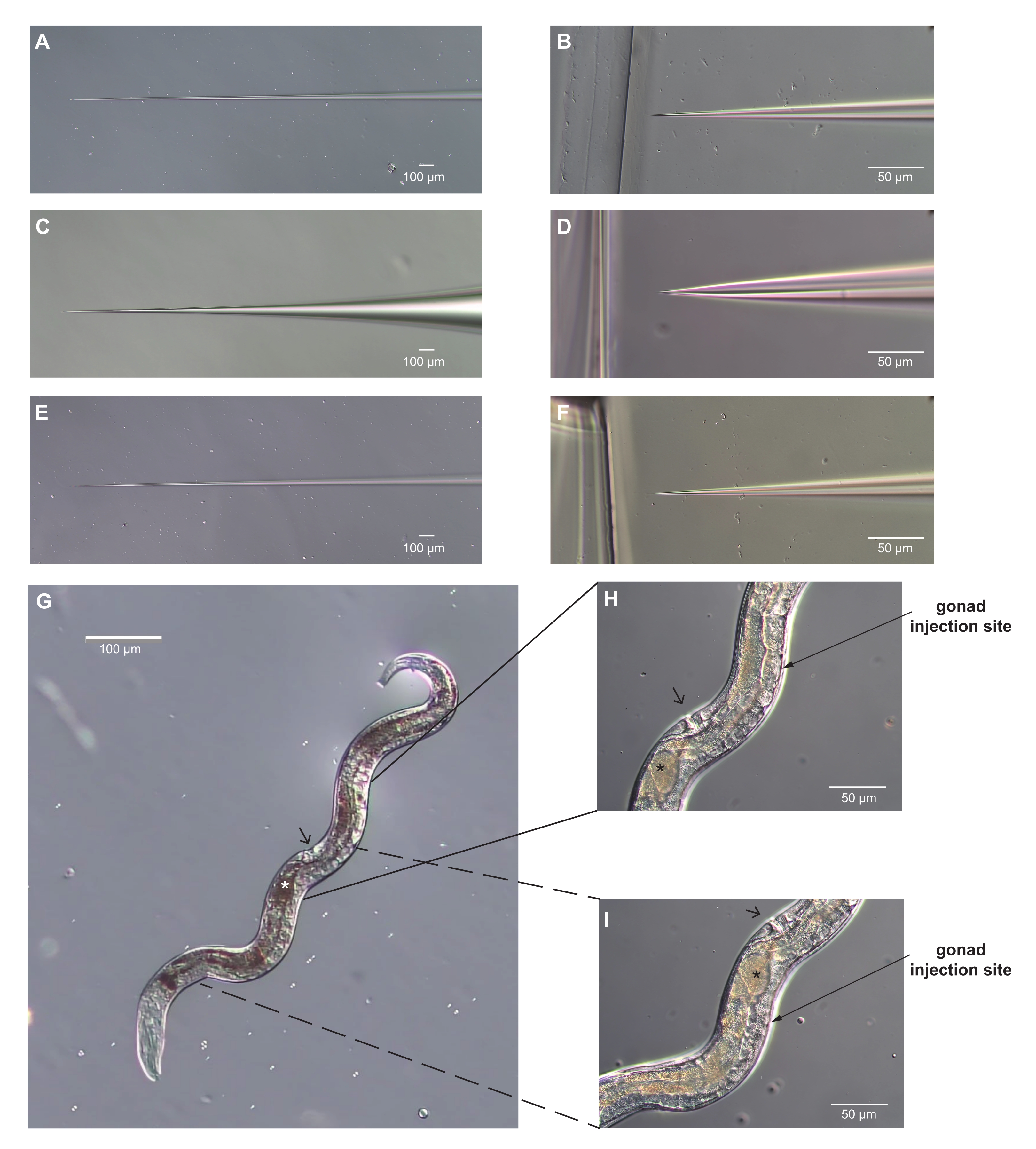
Figure 3 : Aiguilles de micro-injection et femelle adulte Strongyloides stercoralis avec des sites optimaux pour la microinjection identifiés. (A-F) Images d’aiguilles de microinjection. (A-B) La tige conique (A) et la pointe (B) d’une aiguille correctement formée pour la micro-injection. La pointe est assez pointue pour percer la cuticule et assez étroite pour ne pas causer de dommages excessifs. (C-D) La conicité de l’arbre (C) et la pointe (D) d’une aiguille de micro-injection mal formée pour la micro-injection. La pointe est trop émoussée et large, et causera des dommages excessifs au ver. (E-F) La tige conique (E) et la pointe (F) d’une aiguille qui est susceptible d’être trop longue et mince pour fonctionner pour la microinjection. La pointe en F est très similaire à la pointe en D. Cependant, la tige est plus étroite et trop flexible pour percer efficacement la cuticule. De plus, les aiguilles très minces se bouchent facilement. (G) Une image du ver entier correctement positionné pour la micro-injection, en supposant que l’aiguille arrive de la droite. L’antérieur est vers le bas et vers la gauche; la vulve est indiquée par la pointe de flèche. La gonade est visible le long du côté droit de la femelle. Cette femelle n’a qu’un seul ovule dans son utérus (indiqué par l’astérisque). (H, I) Vues agrandies des sites de micro-injection. L’angle de la flèche se rapproche de l’angle de l’aiguille d’injection. La vulve peut être utilisée comme point de repère; il se trouve du côté opposé du ver des bras de la gonade. Les bras de la gonade se courbent autour de l’intestin et les extrémités avec les noyaux en division sont opposées à la vulve. H) Le bras postérieur de la gonade; (I) le bras antérieur. L’un ou l’autre ou les deux bras peuvent être injectés. Pour H, I, les conventions sont comme dans G. Barres d’échelle = 50 μm (B, D, F, H, I); 100 μm (A, C, E, G). Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
- Rangez les aiguilles tirées dans une boîte de Petri en plastique de 15 cm avec un morceau de ruban roulé pour fixer les aiguilles et éviter l’accumulation de poussière sur les pointes.
- Placez une goutte de 0,7 μL du mélange de micro-injection sur l’extrémité ouverte de l’arbre. Suspendez l’aiguille perpendiculairement à une étagère à l’aide d’un morceau de ruban roulé pour remplir l’arbre conique avec le mélange dans les 10 minutes. Préparez 2 aiguilles à la fois au cas où la première ne fonctionnerait pas.
6. Préparer le microscope et casser l’aiguille
REMARQUE: La micro-injection utilise un microscope inversé avec des objectifs 5x et 40x équipé d’une configuration de micro-injecteur pour contrôler le mouvement de l’aiguille. Le microscope inversé doit être placé sur une table lourde ou une table d’air anti-vibration pour réduire le bruit vibratoire. Le porte-aiguille du micro-injecteur est relié à l’azote gazeux qui applique la pression nécessaire pour fournir le mélange de microinjection. Un microscope à dissection plus petit à proximité est utilisé pour transférer les vers.
- Réglez la pression du réservoir de gaz à ~ 40-60 psi pour casser l’aiguille et à ~ 30-50 psi pour la micro-injection, en fonction du débit de liquide.
- Sur le microscope à dissection, recouvrez le tesson de verre sur le couvercle du tampon de micro-injection avec de l’huile d’halocarbure à l’aide d’un pic de ver de platine standard.
- Placez le couvercle du tampon de micro-injection sur la lunette de micro-injection et localisez le tesson de verre recouvert d’huile. Alignez le tesson de verre de manière à ce qu’un bord soit perpendiculaire à la direction de l’aiguille pour servir de surface utilisée pour casser l’aiguille.
- Vérifiez que l’aiguille ne contient pas de bulles ou de débris dans l’arbre conique à l’aide du microscope à dissection. Ensuite, fixez l’aiguille de 1 à 1,5 cm dans le support sous pression.
- Placez la pointe de l’aiguille au centre du champ de vision du microscope à l’œil. Ensuite, sous faible grossissement, positionnez la pointe de l’aiguille dans le champ de vision, perpendiculairement au côté du tesson de verre.
- Passez à haute puissance et alignez la pointe de l’aiguille avec le bord du verre, près mais sans le toucher.
REMARQUE: Lorsqu’elles sont tirées, les aiguilles sont fusionnées fermées. - Pour casser l’extrémité de l’aiguille afin de permettre l’écoulement du liquide, tapotez-la doucement sur le côté du morceau de verre tout en appliquant une pression continue du gaz (figure supplémentaire S1). Une fois que le liquide commence à couler, vérifiez la forme de la pointe et assurez-vous qu’elle est tranchante avec un liquide qui s’écoule facilement.
REMARQUE: Si le liquide s’écoule trop vite ou si l’extrémité est trop émoussée, les vers seront endommagés lors de la microinjection (vidéo 1 et figure 3A-F). - Lorsque le liquide s’écoule bien de l’aiguille, déplacez la lame de micro-injection vers la lunette de dissection et placez des gouttes d’huile d’halocarbure de 1 à 2 μL sur le tampon de gélose pour le placement des vers.
- Transférer 20 à 30 jeunes adultes Strongyloides dans une plaque de NGM à 2% sans bactéries pendant au moins 5 minutes pour éliminer l’excès de bactéries de surface et sélectionner des vers uniques pour la microinjection. Ajoutez plus de vers à la plaque NGM au besoin lors de l’injection.
7. Microinjection de Strongyloides
- Utilisez une petite quantité d’huile d’halocarbure sur un pic à vers pour sélectionner une jeune femelle adulte Strongyloides avec 1 à 4 œufs dans sa gonade à partir de la plaque NGM à 2% sans bactéries.
- Transférez le ver dans une petite goutte d’huile sur le tampon d’agar. À l’aide du pic à ver, positionnez doucement le ver de sorte qu’il ne soit pas enroulé et que la gonade soit visible et facile d’accès. Notez la direction de la gonade (Figure 3G).
- Positionnez le ver dans le champ de vision du microscope à micro-injection. Assurez-vous que la gonade est du même côté que l’aiguille et positionnée de manière à ce que l’aiguille entre en contact avec la gonade à un léger angle (Figure 3H,I).
- Amenez la pointe de l’aiguille sur le côté du ver dans le même plan focal. Visez le bras de la gonade près du milieu du ver. Utilisez le micro-injecteur pour insérer doucement l’aiguille dans la gonade (Vidéo 2).
- Appliquez immédiatement une pression sur l’aiguille pour remplir doucement tout le bras de la gonade avec la solution d’ADN. Déterminer à l’œil nu quand suffisamment de liquide a été injecté (Vidéo 2).
REMARQUE: Il peut prendre jusqu’à 2 s pour remplir la gonade. - Retirez l’aiguille et vérifiez que la plaie se ferme.
REMARQUE: Le ver est trop endommagé pour produire une progéniture si la gonade dépasse à travers la paroi du corps (vidéo supplémentaire S1). - Répétez avec l’autre bras de la gonade si elle est visible.
- Lorsque vous avez terminé l’injection, vérifiez rapidement que l’aiguille n’est pas obstruée en appliquant une pression avec la pointe de l’aiguille sur le tampon de gélose. Transférez la lame avec le ver injecté au microscope à dissection.
- Pour récupérer le ver injecté, placez d’abord quelques gouttes de BU sur le ver pour le faire flotter hors du tampon de gélose.
- Recueillez une petite quantité de bactéries HB101 sur un pic à vers. Touchez le ver avec les bactéries adhérentes sur le pic à vers pour l’éliminer du liquide.
- Transférez doucement le ver sur la plaque de récupération , une plaque NGM à 2% contenant une pelouse HB101.
REMARQUE : Le ver devrait commencer à explorer en quelques minutes. - Après l’injection de quelques femelles, ajoutez des mâles non injectés à partir de la plaque source.
REMARQUE: Un minimum d’un homme pour cinq femmes est une bonne base de référence; un excès de mâles est préféré. - Répétez toutes les étapes jusqu’à ce que suffisamment de femelles aient été injectées pour l’expérience.
- Laissez les adultes sur la plaque de récupération pendant au moins 1 h après l’injection pour permettre aux vers de récupérer et de s’accoupler.
8. Récupération et culture des Strongyloides injectés
- Recueillir les excréments pendant la nuit sur des animaux hôtes non infectés, en utilisant le même protocole que pour les animaux infectés.
- Mélanger les matières fécales non infestées avec une petite quantité de charbon de bois (rapport matières fécales/charbon de bois d’environ 2 pour 1 pour ces assiettes).
- Versez une petite quantité du mélange fécal-charbon de bois dans une boîte de Petri de 6 cm tapissée de papier filtre humide. Assurez-vous que le mélange ne touche pas le couvercle du plat.
- Inonder la plaque de récupération avec BU. À l’aide d’une pipette fixée à 3 μL, transférer les vers dans les matières fécales dans la plaque de charbon fécal. Placez les vers directement sur les matières fécales, pas sur le charbon de bois.
- Vérifiez que les adultes sont sur la plaque de charbon fécal à l’aide d’une lunette de dissection.
- Pour cultiver les vers, placez la plaque dans une chambre humidifiée, c’est-à-dire une boîte en plastique avec un couvercle bien ajusté doublé d’essuie-tout humides.
REMARQUE: Après 2 jours, il y aura un mélange de stades larvaires. Après 5 jours, la plupart des larves se seront développées en iL3; il restera quelques larves plus jeunes. Après 7 jours, toutes les larves doivent être iL3s.
9. Collecte et dépistage des larves F 1 pour récupérer les transgéniques/knockouts
- À l’aide d’une configuration Baermann, collectez les larves dans les plaques de culture de charbon fécal à petite échelle post-injection. Pour obtenir autant de larves que possible, attendez au moins 2 heures avant de récupérer les vers de l’appareil de Baermann.
- Concentrez les larves dans un tube de centrifugeuse de 15 mL comme aux étapes 4.10-4.14 et transférez les larves dans un petit verre de montre avec BU.
- Si les larves seront utilisées pour des expériences comportementales, utilisez 2% de plaques NGM avec une pelouse épaisse de HB101 pour le dépistage.
- Transférer 20 à 30 larves sur la pelouse HB101.
REMARQUE: Les bactéries ralentiront le mouvement des larves. - Sous un microscope à dissection à fluorescence, identifiez les larves exprimant le transgène d’intérêt. Utilisez un pic à vers pour sélectionner les larves transgéniques et déplacez-les vers un petit verre de montre avec BU.
- Utilisez une nouvelle plaque HB101 pour filtrer un autre petit lot de larves. Lorsque suffisamment de larves ont été collectées pour une utilisation expérimentale, traitez les plaques HB101 et les vers en excès avec de l’iode dilué (50% d’iode de Lugol dilué dans l’eau) et jetez-les comme déchets à risque biologique. Alternativement, tuez les vers en excès en utilisant un nettoyant de chenil concentré contenant des chlorures d’alkylbenzyle et d’ammonium.
- Utilisez les vers immédiatement ou laissez-les dans un verre de montre peu profond dans une petite quantité de BU pendant la nuit.
REMARQUE: Les vers peuvent devenir hypoxiques si le liquide est trop profond. Il est possible que laisser les larves dans bu pendant la nuit puisse affecter certains comportements; par conséquent, utilisez des larves pour des expériences comportementales dans les 6 heures.
- Transférer 20 à 30 larves sur la pelouse HB101.
- Si les larves sont utilisées pour la microscopie et non pour des tests comportementaux, immobilisez les vers par paralysie à la nicotine de manière réversible pour le dépistage.
- À l’aide d’une lame de rasoir, marquez une grille sur le fond en plastique d’une plaque de chimiotaxiede 10 cm 12 pour faciliter le suivi de l’emplacement des vers sur la plaque.
- Déposer ~ 3 μL de larves dans BU dans un carré sur la grille. Remplissez autant de carrés que nécessaire. N’utilisez pas ceux près des bords de la plaque, car les larves peuvent ramper sur les côtés de la plaque.
- Ajouter 15-20 μL gouttes de nicotine à 1% dans l’eau aux gouttes de ver.
REMARQUE: Après 4 minutes, les vers seront paralysés. - Filtrez les vers à l’aide d’un microscope à dissection à fluorescence.
- Utilisez un pic à vers pour transférer les larves transgéniques dans un petit verre de montre avec 1 à 2 mL de BU.
REMARQUE: Les larves seront paralysées pendant plusieurs heures et peuvent être facilement montées sur des lames de microscope pour la microscopie. S’ils sont laissés pendant la nuit dans la BU, les iL3 se rétabliront et peuvent être utilisés pour certains tests ou infections de l’hôte chez les mammifères. Cependant, la paralysie nicotinique et l’incubation nocturne dans la BU peuvent affecter certains comportements.
Résultats
Si l’expérience est couronnée de succès, les larves F1 exprimeront le phénotype transgénique et/ou mutant d’intérêt (Figure 4). Cependant, les taux de transformation sont très variables et dépendent des constructions, de la santé des vers, des conditions de culture post-injection et de l’habileté de l’expérimentateur. En général, une expérience réussie donnera >15 larves F1 par femelle injectée et un taux de transformation de >3% pour les marque...
Discussion
Ce protocole de microinjection détaille les méthodes d’introduction de constructions pour la transgénèse et la mutagénèse médiée par CRISPR/Cas9 chez S. stercoralis et S. ratti. Pour S. stercoralis et S. ratti, la survie post-injection et le taux de transgénèse ou de mutagénèse sont soumis à plusieurs variables qui peuvent être affinées.
La première considération critique pour une transgénèse réussie est la façon dont les transgènes ...
Déclarations de divulgation
Les auteurs ne déclarent aucun conflit d’intérêts.
Remerciements
pPV540 et pPV402 étaient de bons cadeaux du Dr James Lok de l’Université de Pennsylvanie. Nous remercions Astra Bryant pour ses commentaires utiles sur le manuscrit. Ce travail a été financé par un Burroughs-Wellcome Fund Investigators in the Pathogenesis of Disease Award, un Howard Hughes Medical Institute Faculty Scholar Award et National Institutes of Health R01 DC017959 (E.A.H.).
matériels
| Name | Company | Catalog Number | Comments |
| (−)-Nicotine, ≥99% (GC), liquid | Sigma-Aldrich | N3876-5ML | nicotine for paralyzing worms |
| 3" iron C-clamp, 3" x 2" (capacity x depth) | VWR | 470121-790 | C-clamp to secure setup to bench top |
| Agarose LE | Phenix | RBA-500 | agarose for slides |
| Bone char, 4 lb pail, 10 x 28 mesh | Ebonex | n/a | charcoal for fecal-charcoal cultures |
| Bone char, granules, 10 x 28 mesh | Reade | bonechar10x28 | charcoal for fecal-cultures (alternative to the above) |
| Coarse micromanipulator | Narishige | MMN-1 | coarse micromanipulator |
| Corning Costar Spin-X centrifuge tube filters | Fisher | 07-200-385 | microfilter column |
| Cover glass, 48 x 60 mm, No. 1 thickness | Brain Research Lab | 4860-1 | coverslips (48 x 60 mm) |
| Deep Petri dishes, heavy version with 6 vents, 100 mm diameter | VWR | 82050-918 | 10 cm Petri dishes (for fecal-charcoal cultures) |
| Eisco retort base w/ rod | Fisher | 12-000-101 | stand for Baermann apparatus |
| Eppendorf FemtoJet microinjector microloader tips | VWR | 89009-310 | for filling microinjection needles |
| Fisherbrand absorbent underpads | Fisher | 14-206-62 | bench paper (for prepping) |
| Fisherbrand Cast-Iron Rings | Fisher | 14-050CQ | Baermann o-ring |
| Fisherbrand tri-cornered polypropylene beakers | Fisher | 14-955-111F | Plastic beaker (for mixing) |
| Fisherbrand tri-cornered polypropylene beakers | Fisher | 14-955-111F | Plastic beaker (for catch bucket/water bucket) |
| Fisherbrand tri-cornered polypropylene beakers | Fisher | 14-955-111F | Plastic beaker (x2) (to make holder) |
| Gorilla epoxie in syringe | McMaster-Carr | 7541A51 | glue (to attach tubing) |
| Halocarbon oil 700 | Sigma-Aldrich | H8898-50ML | halocarbon oil |
| High-temperature silicone rubber tubing for food and beverage, 1/2" ID, 5/8" OD | McMaster-Carr | 3038K24 | tubing (for funnel) |
| KIMAX funnels, long stem, 60° Angle, Kimble Chase | VWR | 89001-414 | Baermann funnel |
| Kimberly-Clark Professional Kimtech Science benchtop protectors | Fisher | 15-235-101 | bench paper (for prepping) |
| Leica stereomicroscope with fluorescence | Leica | M165 FC | GFP stereomicroscope for identifying and sorting transgenic worms |
| microINJECTOR brass straight arm needle-holder | Tritech | MINJ-4 | microinjection needle holder |
| microINJECTOR system | Tritech | MINJ-1 | microinjection system |
| Mongolian Gerbils | Charles River Laboratories | 213-Mongolian Gerbil | gerbils for maintenance of S. stercoralis, male 4-6 weeks |
| Nasco Whirl-Pak easy-to-close bags, 18 oz | VWR | 11216-776 | Whirl-Pak sample bags |
| Nylon tulle (mesh) | Jo-Ann Fabrics | zprd_14061949a | nylon mesh for Baermann holder |
| Platinum wire, 36 Gauge, per inch | Thomas Scientific | 1233S72 | platinum/iridium wire for worm picks |
| Puritan tongue depressors, 152 mm (L) x 17.5 mm (W) | VWR | 62505-007 | wood sticks (for mixing samples) |
| QIAprep Spin Miniprep Kit (250) | QIAGEN | 27106 | QIAGEN miniprep kit |
| Rats-Long Evans | Envigo | 140 HsdBlu:LE Long Evans | rats for maintenance of S. ratti, female 4-6 weeks |
| Rats-Sprague Dawley | Envigo | 002 Hsd:Sprague Dawley SD | rats for maintenance of S. ratti, female 4-6 weeks |
| Really Useful Boxes translucent storage boxes with lids, 1.6 L capacity, 7-5/8" x 5-5/16" x 4-5/16" | Office Depot | 452369 | plastic boxes for humidified chamber |
| Shepherd techboard, 8 x 16.5 inches | Newco | 999589 | techboard |
| Stainless steel raised wire floor | Ancare | R20SSRWF | wire cage bottoms |
| StalkMarket compostable cutlery spoons, 6", white, pack of 1,000 | Office Depot | 9587303 | spoons |
| Stender dish, stacking type, 37 x 25 mm | Carolina (Science) | 741012 | watch glasses (small, round) |
| Stereomicroscope | Motic | K-400 LED | dissecting prep scope |
| Storage tote, color clear/white, outside height 4-7/8 in, outside length 13-5/8 in, Sterilite | Grainger | 53GN16 | plastic boxes for humidified chamber |
| Sutter P-30 micropipette puller | Sutter | P-30/P | needle puller with platinum/iridium filament |
| Syracuse watch glasses | Fisher | S34826 | watch glasses (large, round) |
| Thermo Scientific Castaloy fixed-angle clamps | Fisher | 05-769-2Q | funnel clamps (2x) |
| Three-axis hanging joystick oil hydrolic micromanipulator | Narishige | MM0-4 | fine micromanipulator |
| United Mohr pinchcock clamps | Fisher | S99422 | Pinch clamps (2x) |
| Vented, sharp-edge Petri dishes (60 mm diameter) | Tritech Research | T3308P | 6 cm Petri dishes (for small-scale fecal-charcoal cultures) |
| VWR light-duty tissue wipers | VWR | 82003-820 | lining for Baermann holder |
| watch glass, square, 1-5/8 in | Carolina (Science) | 742300 | watch glasses (small, square) |
| Whatman qualitative grade plain circles, grade 1, 5.5 cm diameter | Fisher | 09-805B | filter paper (for 6 cm Petri dishes) |
| Whatman qualitative grade plain circles, grade 1, 9 cm diameter | Fisher | 09-805D | filter paper (for 10 cm Petri dishes) |
| World Precision Instrument borosilicate glass capillary, 1.2 mm x 4 in | Fisher | 50-821-813 | glass capillaries for microinjection needles |
| X-Acto Knives, No. 1 Knife With No. 11 Blade | Office Depot | 238816 | X-Acto knives without blades to hold worm picks |
| Zeiss AxioObserver A1 | Zeiss | n/a | inverted microscope |
Références
- Krolewiecki, A. J., et al. A public health response against Strongyloides stercoralis: time to look at soil-transmitted helminthiasis in full. PLoS Neglected Tropical Diseases. 7 (5), 2165 (2013).
- Buonfrate, D., et al. The global prevalence of Strongyloides stercoralis infection. Pathogens. 9 (6), 468 (2020).
- Castelletto, M. L., Gang, S. S., Hallem, E. A. Recent advances in functional genomics for parasitic nematodes of mammals. Journal of Experimental Biology. 223, 206482 (2020).
- Evans, T. C., et al. Transformation and microinjection. WormBook. , (2006).
- Lok, J. B., Unnasch, T. R., et al. Transgenesis in animal parasitic nematodes: Strongyloides spp. and Brugia spp. WormBook. , (2013).
- Shao, H. G., Li, X. S., Lok, J. B. Heritable genetic transformation of Strongyloides stercoralis by microinjection of plasmid DNA constructs into the male germline. International Journal for Parasitology. 47 (9), 511-515 (2017).
- Schafer, T. W., Skopic, A. Parasites of the small intestine. Current Gastroenterology Reports. 8 (4), 312-320 (2006).
- Stiernagle, T. Maintenance of C. elegans. The C. elegans Research Community, WormBook. , (2006).
- Gang, S. S., et al. Targeted mutagenesis in a human-parasitic nematode. PLoS Pathogens. 13 (10), 1006675 (2017).
- Lok, J. B. Strongyloides stercoralis: a model for translational research on parasitic nematode biology. The C. elegans Research Community, WormBook. , (2007).
- Hawdon, J. M., Schad, G. A. Long-term storage of hookworm infective larvae in buffered saline solution maintains larval responsiveness to host signals. Proceedings of the Helminthological Society of Washington (USA). 58 (1), 140-142 (1991).
- Bargmann, C. I., Hartwieg, E., Horvitz, H. R. Odorant-selective genes and neurons mediate olfaction in C. elegans. Cell. 74 (3), 515-527 (1993).
- Junio, A. B., et al. Strongyloides stercoralis: cell- and tissue-specific transgene expression and co-transformation with vector constructs incorporating a common multifunctional 3' UTR. Experimental Parasitology. 118 (2), 253-265 (2008).
- Gang, S. S., et al. Chemosensory mechanisms of host seeking and infectivity in skin-penetrating nematodes. Proceedings of the National Academy of Sciences of the United States of America. 117 (30), 17913-17923 (2020).
- Bryant, A. S., et al. A critical role for thermosensation in host seeking by skin-penetrating nematodes. Current Biology. 28 (14), 2338-2347 (2018).
- Lok, J. B. Nucleic acid transfection and transgenesis in parasitic nematodes. Parasitology. 139 (5), 574-588 (2012).
- Shao, H., et al. Transposon-mediated chromosomal integration of transgenes in the parasitic nematode Strongyloides ratti and establishment of stable transgenic lines. PLoS Pathogens. 8 (8), 1002871 (2012).
- Lok, J. piggyBac: a vehicle for integrative DNA transformation of parasitic nematodes. Mobile Genetic Elements. 3 (2), 24417 (2013).
- Li, X., et al. Successful transgenesis of the parasitic nematode Strongyloides stercoralis requires endogenous non-coding control elements. International Journal for Parasitology. 36 (6), 671-679 (2006).
- Bryant, A. S., Hallem, E. A. The Wild Worm Codon Adapter: a web tool for automated codon adaptation of transgenes for expression in non-Caenorhabditis nematodes. G3. 3 (7), (2021).
- Crane, M., et al. In vivo measurements reveal a single 5'-intron is sufficient to increase protein expression level in Caenorhabditis elegans. Scientific Reports. 9 (1), 9192 (2019).
- Han, Z., et al. Improving transgenesis efficiency and CRISPR-associated tools through codon optimization and native intron addition in Pristionchus nematodes. Genetics. 216 (4), 947-956 (2020).
- Adams, S., Pathak, P., Shao, H., Lok, J. B., Pires-daSilva, A. Liposome-based transfection enhances RNAi and CRISPR-mediated mutagenesis in non-model nematode systems. Scientific Reports. 9 (1), 483 (2019).
- Dulovic, A., Puller, V., Streit, A. Optimizing culture conditions for free-living stages of the nematode parasite Strongyloides ratti. Experimental Parasitology. 168, 25-30 (2016).
- Harvey, S. C., Gemmill, A. W., Read, A. F., Viney, M. E. The control of morph development in the parasitic nematode Strongyloides ratti. Proceedings of the Royal Society B: Biological Sciences. 267 (1457), 2057-2063 (2000).
- Kim, A., Pyykko, I. Size matters: versatile use of PiggyBac transposons as a genetic manipulation tool. Molecular and Cellular Biochemistry. 354 (1-2), 301-309 (2011).
- Lok, J. B., Shao, H., Massey, H. C., Li, X. Transgenesis in Strongyloides and related parasitic nematodes: historical perspectives, current functional genomic applications and progress towards gene disruption and editing. Parasitology. 144 (3), 327-342 (2017).
- Farboud, B., Meyer, B. J. Dramatic enhancement of genome editing by CRISPR/Cas9 through improved guide RNA design. Genetics. 199 (4), 959-971 (2015).
- Cheong, M. C., et al. Identification of a nuclear receptor/coactivator developmental signaling pathway in the nematode parasite Strongyloides stercoralis. Proceedings of the National Academy of Sciences of the United States of America. 118 (8), 2021864118 (2021).
- Nolan, T. J., Megyeri, Z., Bhopale, V. M., Schad, G. A. Strongyloides stercoralis: the first rodent model for uncomplicated and hyperinfective strongyloidiasis, the Mongolian gerbil (Meriones unguiculatus). Journal of Infectious Diseases. 168 (6), 1479-1484 (1993).
- Li, X., et al. Transgenesis in the parasitic nematode Strongyloides ratti. Molecular and Biochemical Parasitology. 179 (2), 114-119 (2011).
- Viney, M. E. Exploiting the life cycle of Strongyloides ratti. Parasitology Today. 15 (6), 231-235 (1999).
- Stoltzfus, J. D., Massey, H. C., Nolan, T. J., Griffith, S. D., Lok, J. B. Strongyloides stercoralis age-1: a potential regulator of infective larval development in a parasitic nematode. PLoS ONE. 7 (6), 38587 (2012).
- Castelletto, M. L., Massey, H. C., Lok, J. B. Morphogenesis of Strongyloides stercoralis infective larvae requires the DAF-16 ortholog FKTF-1. PLoS Pathogens. 5 (4), 1000370 (2009).
- Douglas, B., et al. Transgenic expression of a T cell epitope in Strongyloides ratti reveals that helminth-specific CD4+ T cells constitute both Th2 and Treg populations. PLoS Pathogens. 17 (7), 1009709 (2021).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationExplorer plus d’articles
This article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.