
Для просмотра этого контента требуется подписка на Jove Войдите в систему или начните бесплатную пробную версию.
Method Article
Генерация трансгенов и нокаутов у видов стронгилоидов путем микроинъекции
В этой статье
Резюме
Функциональный геномный инструментарий для паразитических нематод Strongyloides stercoralis и Strongyloides ratti включает трансгенез, CRISPR/Cas9-опосредованный мутагенез и RNAi. Этот протокол продемонстрирует, как использовать внутригонадную микроинъекцию для введения трансгенов и компонентов CRISPR в S. stercoralis и S. ratti.
Аннотация
Род Strongyloides состоит из нескольких видов проникающих в кожу нематод с различными диапазонами хозяев, включая Strongyloides stercoralis и Strongyloides ratti. S. stercoralis является паразитической, проникающей в кожу нематодой, которая заражает около 610 миллионов человек, в то время как крысиный паразит S. ratti тесно связан с S. stercoralis и часто используется в качестве лабораторной модели для S. stercoralis. И S. stercoralis , и S. ratti легко поддаются генерации трансгенов и нокаутов с помощью метода доставки экзогенных нуклеиновых кислот интрагонадной микроинъекции и, как таковые, появились в качестве модельных систем для других паразитических гельминтов, которые еще не поддаются этому методу.
Паразитические strongyloides взрослые населяют тонкую кишку своего хозяина и выпускают потомство в окружающую среду через фекалии. Попав в окружающую среду, личинки развиваются в свободноживущих взрослых особей, которые живут в фекалиях и производят потомство, которое должно найти и вторгнуться в нового хозяина. Это поколение окружающей среды уникально для видов Strongyloides и достаточно похоже по морфологии на модель свободноживущей нематоды Caenorhabditis elegans , что методы, разработанные для C. elegans , могут быть адаптированы для использования с этими паразитическими нематодами, включая внутригонадную микроинъекцию. Используя внутригонадную микроинъекцию, в стронгилоиды можно вводить широкий спектр трансгенов. Компоненты CRISPR/Cas9 также могут быть микроинъектированы для создания мутантных личинок Strongyloides . Здесь описана методика интрагонадной микроинъекции в стронгилоиды, включая подготовку свободноживущих взрослых, процедуру инъекций и отбор трансгенного потомства. Включены изображения трансгенных личинок Strongyloides , созданных с использованием мутагенеза CRISPR/Cas9. Цель этой статьи состоит в том, чтобы позволить другим исследователям использовать микроинъекцию для создания трансгенных и мутантных стронгилоидов.
Введение
Strongyloides stercoralis долгое время игнорировался как важный патоген человека по сравнению с более широко признанными анкилостомами и круглым червем Ascaris lumbricoides1. Предыдущие исследования глистной нагрузки часто сильно недооценивали распространенность S. stercoralis из-за низкой чувствительности общих методов диагностики S. stercoralis2. В последние годы эпидемиологические исследования, основанные на улучшенных диагностических инструментах, показали, что истинная распространенность инфекций S. stercoralis намного выше, чем сообщалось ранее, примерно 610 миллионов человек во всем мире2.
Как S. stercoralis, так и другие виды Strongyloides, включая близкородственного крысиного паразита и общую лабораторную модель S. ratti, имеют необычный жизненный цикл, который выгоден для экспериментальных геномных исследований, поскольку он состоит как из паразитических, так и из свободноживущих (экологических) поколений3 (рисунок 1). В частности, как S. stercoralis, так и S. ratti могут циклически проходить через одно свободноживущее поколение. Свободноживущее поколение состоит из постпаразитарных личинок, которые развиваются в свободноживущих взрослых самцов и самок; все потомство свободноживущих взрослых особей развивается в инфекционных личинок, которые должны заразить хозяина, чтобы продолжить жизненный цикл. Кроме того, этим экологическим или свободноживущим поколением можно экспериментально манипулировать в лаборатории. Поскольку свободноживущие взрослые strongyloides и взрослые особи C. elegans имеют схожую морфологию, такие методы, как интрагонадная микроинъекция, которые были первоначально разработаны для C. elegans, могут быть адаптированы для использования со свободноживущими взрослыми Strongyloides 4,5. В то время как ДНК обычно вводится в свободно живущих взрослых самок, как самцы, так и самки стронгилоидов могут быть микроинъективированы6. Таким образом, функциональные геномные инструменты доступны для изучения многих аспектов биологии стронгилоидов. Другие паразитические нематоды не имеют свободно живущего поколения и, как следствие, не так легко поддаются функциональным геномным методам3.

Рисунок 1: Жизненный цикл Strongyloides stercoralis. Паразитические самки S. stercoralis населяют тонкую кишку своих млекопитающих-хозяев (людей, нечеловеческих приматов, собак). Паразитические самки размножаются путем партеногенеза и откладывают яйца в тонком кишечнике. Яйца вылупляются, все еще находясь внутри хозяина, в постпаразитарные личинки, которые затем попадают в окружающую среду с фекалиями. Если постпаразитарные личинки являются самцами, они развиваются в свободноживущих взрослых самцов. Если постпаразитарные личинки являются самками, они могут либо развиться в свободноживущих взрослых самок (косвенное развитие), либо в личинок третьей стадии инфекции (iL3s; прямое развитие). Свободно живущие самцы и самки размножаются половым путем, чтобы создать потомство, которое ограничено, чтобы стать iL3s. При определенных условиях S. stercoralis также может подвергаться аутоинфекции, при которой часть постпаразитарных личинок остается внутри кишечника хозяина, а не попадает в окружающую среду с калом. Эти личинки могут развиваться в аутоинфективные личинки (L3a) внутри хозяина, проникать через стенку кишечника, мигрировать по организму и в конечном итоге возвращаться в кишечник, чтобы стать репродуктивными взрослыми. Жизненный цикл S. ratti аналогичен, за исключением того, что S. ratti заражает крыс и не имеет аутоинфекционного цикла. Генерация окружающей среды является ключом к использованию видов Strongyloides для генетических исследований. Свободно живущие взрослые самки (P0) могут быть микроинъективированы; их потомство, которое станет iL3s, является потенциальным трансгеномF1. Эта цифра была изменена по сравнению с Castelletto et al. 3. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
S. stercoralis разделяет многие аспекты своей биологии с другими желудочно-кишечными паразитическими нематодами, включая инвазию хозяина и иммунную модуляцию хозяина. Например, человеко-паразитические анкилостомы в родах Necator и Ancylostoma также заражаются при проникновении в кожу, аналогичным образом перемещаются по телу и в конечном итоге живут как паразитические взрослые в тонком кишечнике7. Таким образом, многие желудочно-кишечные нематоды, вероятно, используют общее сенсорное поведение и методы уклонения от иммунитета. В результате знания, полученные от Strongyloides , дополнят результаты у других менее генетически поддающихся лечению нематод и приведут к более полному пониманию этих сложных и важных паразитов.
Этот протокол микроинъекции описывает метод введения ДНК в Strongyloides свободноживущих взрослых самок для создания трансгенного и мутантного потомства. Описаны требования к поддержанию штамма, включая сроки развития взрослых червей для микроинъекций и сбор трансгенного потомства. Протоколы и демонстрация полной техники микроинъекции, наряду с протоколами культивирования и скрининга трансгенного потомства, включены вместе со списком всего необходимого оборудования и расходных материалов.
Access restricted. Please log in or start a trial to view this content.
протокол
ПРИМЕЧАНИЕ: Песчанки использовались для прохождения S. stercoralis, а крысы использовались для прохождения S. ratti. Все процедуры были одобрены Управлением по надзору за исследованиями на животных UCLA (Протокол No 2011-060-21A), которое придерживается стандартов AAALAC и Руководства по уходу и использованию лабораторных животных. Следующие задачи должны быть выполнены по крайней мере за один день до микроинъекции: культивирование червей, подготовка микроинъекционных прокладок, создание конструкций для смеси микроинъекций и распространение бактерий (E. coli HB101) на пластины8 6 см Nematode Growth Media (NGM). Свободно живущим самкам требуется как минимум 24 ч после фекального сбора при 25 °C, чтобы развиться в молодых людей, прежде чем они могут быть микроинъектированы. Микроинъекционные прокладки должны быть полностью сухими. Бактериальные пластины должны высохнуть и установить небольшой газон.
1. Подготовка микроинъекционных слайдов: не менее чем за один день до инъекции
ПРИМЕЧАНИЕ: Черви устанавливаются на микроинъекционные крышки с сухими агаровыми прокладками для инъекций.
- Установите тепловой блок на 90 °C.
- Добавьте 5 мл ddH2O, затем 100 мг агарозы в боросиликатную стеклянную трубку.
- Нагрейте смесь агарозы в трубке над пламенем до тех пор, пока агароза не растворится.
- Поместите трубку в тепловой блок, установленный при 90 °C, чтобы поддерживать агарозу в жидком состоянии.
- Капните ~180 мкл раствора агарозы на крышку с помощью стеклянного пипетки Пастера или пипетки с пластиковым наконечником. Сразу же опустите сверху второй обшивку, чтобы сплющить агарозу в тонкую прокладку.
- Через 5-10 секунд снимите верхнюю крышку, раздвинув их друг от друга. Определите, на каком слайде находится агаровая подушка, и положите ее лицевой стороной вверх.
- Выберите крошечный кусочек стеклянного осколка из разбитого чехла и осторожно вдавите его в агар у верхнего края прокладки с помощью щипцов (дополнительный рисунок S1).
- Продолжайте делать микроинъекционные прокладки с раствором агарозы.
- Высушите подушечки агарозы ночью на скамейке или в духовке. Хранить в коробке с крышкой.
ПРИМЕЧАНИЕ: Агарозные прокладки могут использоваться до 2 месяцев, но используются только для одного запуска инъекций.
2. Культивирование стронгилоидов для получения глистов для микроинъекции: за 1 - 2 дня до инъекции
ПРИМЕЧАНИЕ: Протокол поддержания штамма можно найти в Дополнительном материале, который включает подробное описание того, как заражать песчанок и крыс нематодами и собирать нематод из фекалий инфицированных животных.
- За два дня до дня инъекции поместите инфицированных животных 9,10 в клетки для сбора на ночь.
- На следующее утро соберите зараженный кал и сделайте фекально-угольные пластиныпо 9,10.
- Поместите пластину при температуре 25 °C в течение 24 часов, чтобы свободно живущие черви развились в молодых людей.
- В ночь перед днем инъекций поместите неинфицированных животных-хозяев в клетки для сбора.
- В день инъекции соберите неинцифицированный кал для послеинъекционного культивирования.
3. Изготовление микроинъекционной смеси: до или в день инъекции
ПРИМЕЧАНИЕ: Микроинъекционная смесь состоит из интересующих плазмид, разбавленных до желаемой концентрации в червячном буферном физиологическом растворе (BU) (50 мМ Na2HPO4, 22 мМ KH2PO4, 70 мМ NaCl)11.
- Определяют концентрацию плазмидных запасов и желаемую концентрацию в микроинъекционной смеси (таблица 1).
| Микроинъекционная смесь: репортерная конструкция | |||
| Компонент | Концентрация запасов | Количество | Конечная концентрация |
| pMLC30 gpa-3::gfp | 300 нг/мкл | 1.7 мкл | 50 нг/мкл |
| БУ | на | 8.3 мкл | на |
| итог | 10 мкл | 50 нг/мкл | |
| Микроинъекционная смесь: мутагенез CRISPR/Cas9 | |||
| Компонент | Концентрация запасов | Количество | Конечная концентрация |
| pMLC47 tax-4 sgRNA | 300 нг/мкл | 2.7 мкл | 80 нг/мкл |
| плазмида pEY11 Ss-tax-4 HDR | 400 нг/мкл | 2.0 мкл | 80 нг/мкл |
| плазмида pPV540 strCas9 | 350 нг/мкл | 1.1 мкл | 40 нг/мкл |
| БУ | на | 4.2 мкл | на |
| итог | 10 мкл | 200 нг/мкл | |
| Микроинъекционная смесь: интеграция piggyBac | |||
| Компонент | Концентрация запасов | Количество | Конечная концентрация |
| pMLC30 gpa3::gfp | 300 нг/мкл | 2.0 мкл | 60 нг/мкл |
| pPV402 транспозазна плазмида | 450 нг/мкл | 0.9 мкл | 40 нг/мкл |
| БУ | на | 7.1 мкл | на |
| итог | 10 мкл | 100 нг/мкл | |
Таблица 1: Примеры микроинъекционных смесей. Плазмиды и концентрации для трех примеров микроинъекционных смесей: один для репортера gpa-3::GFP построен10, один для CRISPR/Cas9-опосредованного разрушения локуса Ss-tax-4 14,15 и один для piggyBac-опосредованной интеграции Ss-gpa-3::GFP конструкции 13,17,18. strCas9 обозначает ген Cas9, оптимизированный для кодона Strongyloides. Конечные концентрации обычно используются в микроинъекционных смесях strongyloides.
- Разводят плазмиды в БУ до общего объема 10-20 мкл.
- Прокрутите смесь через фильтрующую колонну при 5000 × г в течение 1-2 мин.
- Немедленно используйте микроинжекционную смесь или храните ее при -20 °C для будущего использования.
4. Соберите стронгилоиды для микроинъекции: утро дня инъекции
- Установите аппарат Baermann с 1 фекально-угольной пластиной стронгилоидов молодых взрослых (рисунок 2).
ПРИМЕЧАНИЕ: Фекально-древесноугольная пластина может содержать некоторые инфекционные личинки. Средства индивидуальной защиты состоят из лабораторного халата, перчаток и средств защиты глаз. Кожа не должна подвергаться воздействию между перчаткой и рукавом лабораторного халата.

Рисунок 2: Аппарат Baermann, используемый для сбора паразитических червей из культур10. Содержимое фекально-угольной пластины помещают в верхнюю часть столба с теплой водой. Черви мигрируют в воду и собираются на дне воронки. (A) Для установки аппарата Baermann подставка для воронки Baermann прижимается к скамье с помощью C-зажима. Резиновая трубка, прикрепленная к концу воронки, закрывается зажимами, а под трубкой помещается ведро для капель. В стеклянную воронку добавляют теплую воду. (B) Пластиковый держатель кольца для фекально-угольной смеси затем облицовывается 3 кусочками лабораторных тканей (слева). Деревянная палочка или депрессор языка (посередине) используется для переноса содержимого фекально-угольной пластины (справа) в держатель пластикового кольца. (C) Крупный план нижней части держателя пластикового кольца для смеси фекалий и древесного угля с изображением двойного слоя нейлонового тюля, выстилающего нижнюю часть держателя. (D) Затем держатель фекально-древесного угля помещается в верхнюю часть стеклянной воронки. (E) Лабораторную ткань смачивают водой и закрывают поверх фекально-угольной смеси. Добавляется больше теплой воды, чтобы в основном погрузить фекально-древесный уголь. (F) Полная установка Baermann, с фекально-угольной культурой, погруженной в теплую воду. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
- Установите стеклянную воронку с резиновой коллекторной трубкой на кольцевую подставку с помощью уплотнительного кольца и закрепите ее зажимом. Закройте коллекторную трубку 2 зажимами (рисунок 2А).
- Поместите под воронку ведро для ловли капель.
- Добавьте теплую (приблизительно 40 °C) воду в воронку на 5 см ниже обода. Убедитесь, что система не протекает.
- Выровняйте держатель Baermann, сито, изготовленное из 2 пластиковых колец с 2 слоями нейлоновой тюлевой сетки, закрепленной между ними, с 3 перекрывающимися кусками лабораторной ткани. Добавьте фекально-древесную смесь в держатель Baermann (рисунок 2B,C).
- Поместите держатель Baermann с фекально-угольной смесью в воронку. Сложите ткани вокруг фекально-угольной смеси и добавьте достаточно воды, чтобы погрузить большую часть фекально-древесного угля. Не заполняйте выше 2 см от края воронки (рисунок 2D,E).
- Сверху на воронку накройте 15-сантиметровую пластиковую крышку чашки Петри, чтобы она содержала запах. Пометьте воронку по мере необходимости (рисунок 2F).
- Подождите от 30 мин до 1 ч, чтобы собрать червей из аппарата Baermann.
- Держите трубку центрифуги объемом 50 мл под резиновой трубкой в нижней части воронки. Осторожно откройте зажимы на дне, чтобы дозировать 30-40 мл воды, содержащей червей, в трубку объемом 50 мл.
- Перенесите 15 мл воды Baermann, содержащей червей, в 15 мл центрифужной трубки. Вращайте трубку центрифуги объемом 15 мл в течение 1 мин при температуре ~750 × г (медленно). В качестве альтернативы, дайте червям самотеком осесть в течение 10-15 минут.
- Удалите супернатант до ~ 2 мл и выбросьте супернатант в контейнер для жидких отходов с йодом, чтобы убить любых червей.
- Добавьте больше воды Baermann в трубку для сбора 15 мл и повторите отжим. Удалите супернатант до ~2 мл и выбросьте, как показано на шаге 4.11.
- Повторяйте шаги 4.11 и 4.12 до тех пор, пока все черви не будут собраны в трубку центрифуги объемом 15 мл. После финального отжима удалите как можно больше воды.
- Осмотрите гранулы червей (40-100 мкл) на дне трубки. Если червей не видно, подождите еще 1-2 часа и попробуйте собрать больше червей из аппарата Baermann.
- Перенесите червей в как можно меньшее количество воды на пластину NGM 6 см 2% с газоном E . coli HB101. Используйте эту пластину в качестве исходной пластины для микроинъекции.
- Выбросьте фекально-древесноугольную смесь, обработав ее разбавленным йодом (50% разбавление йода Люголя в воде), завернув ее в полиэтиленовую пленку для улавливания капель и поместив в контейнер для биологически опасных отходов.
- Добавьте 10 мл разбавленного йода в ведро для улавливания и слейте в него лишнюю воду из Baermann.
- Промыть многоразовые компоненты (воронку, ведро для улавливания, пластиковый держатель с тюлем, пластиковую крышку и зажимы) 10% отбеливателем и тщательно промыть.
5. Вытягивание и загрузка микроинъекционных игл: непосредственно перед инъекцией
- Подготовьте микроинъекционные иглы, вытягивая стеклянные капиллярные трубки с помощью съемника игл.
ПРИМЕЧАНИЕ: Примеры настроек для коммерческого съемника иглы, оснащенного 3-мм платиновой/иридиевой нитью: Тепло = 810-820, Тяга = 800-820, микрометр = 2,5. - Просмотрите наконечники под рассекающим микроскопом. Если иглы имеют нужную форму (рисунок 3А-Ф), вытяните 4-6 игл (2-3 капиллярных трубки). Чтобы достичь правильной формы иглы, измените настройки по мере необходимости: отрегулируйте настройки Heat или Pull на 10 и тяните новые иглы, пока форма конуса и вала не станет более подходящей.

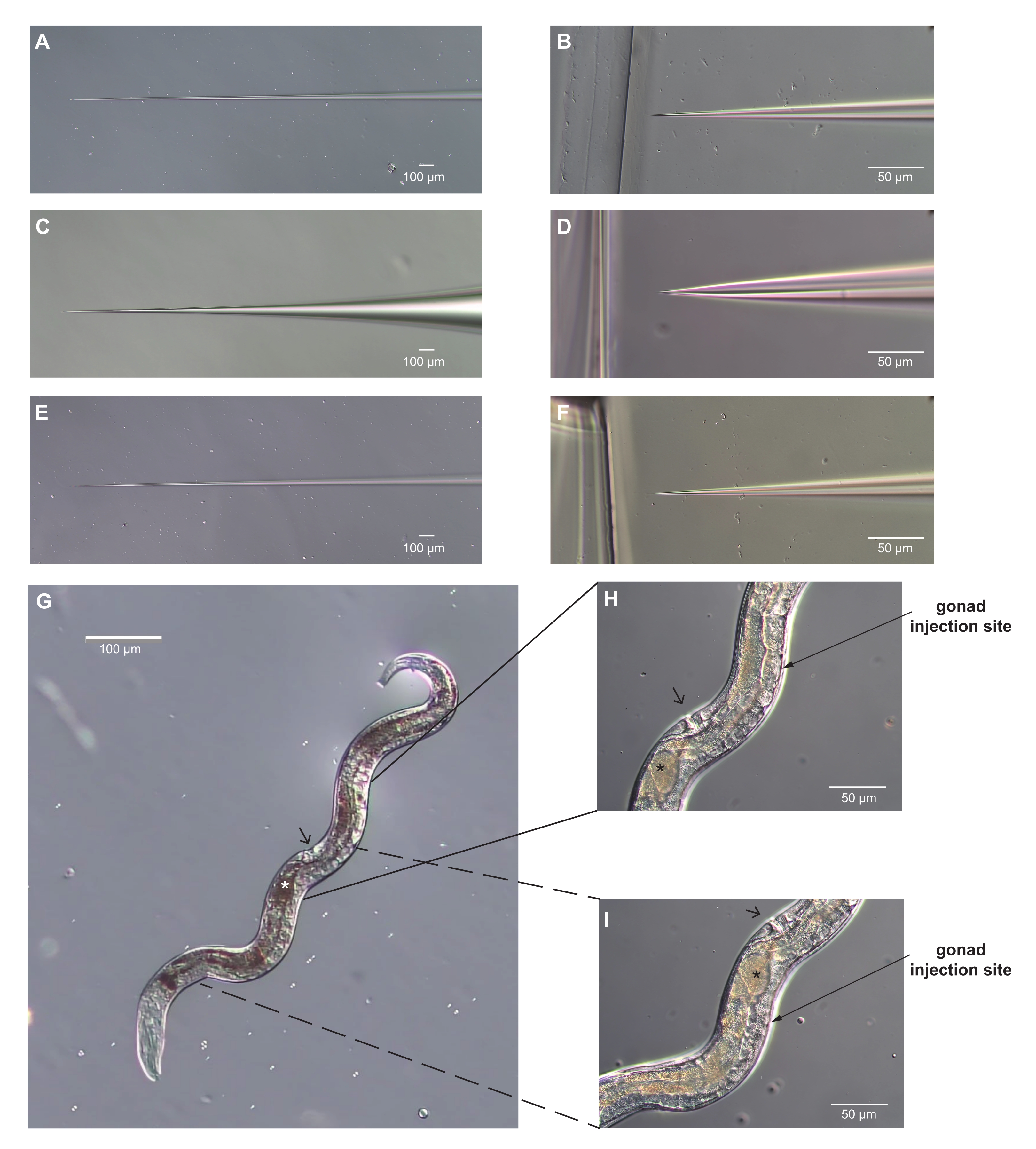
Рисунок 3: Микроинъекционные иглы и Strongyloides stercoralis взрослой самки с оптимальными участками для микроинъекции идентифицированы. (A-F) Изображения микроинъекционных игл. (А-Б) Конус вала (A) и наконечник (B) иглы, которая имеет правильную форму для микроинъекции. Наконечник достаточно острый, чтобы проткнуть кутикулу, и достаточно узкий, чтобы не нанести чрезмерных повреждений. (С-Д) Конус вала (C) и наконечник (D) микроинъекционной иглы, которая имеет неправильную форму для микроинъекции. Наконечник слишком тупой и широкий, и нанесет чрезмерный ущерб червю. (Е-Ф) Конус вала (E) и наконечник (F) иглы, которая, вероятно, будет слишком длинной и тонкой, чтобы работать для микроинъекции. Наконечник в F очень похож на наконечник в D. Однако вал более узкий и слишком гибкий, чтобы эффективно пробить кутикулу. Кроме того, очень тонкие иглы легко засоряются. (G) Изображение всего червя, правильно расположенного для микроинъекции, предполагая, что игла входит справа. Передний находится внизу и влево; вульва обозначается наконечником стрелы. Гонад видна вдоль правой стороны самки. У этой самки есть только одно яйцо в матке (обозначено звездочкой). (Н, И) Увеличенные виды участков микроинъекций. Угол стрелки приближается к углу инъекционной иглы. Вульва может быть использована в качестве ориентира; он находится на противоположной стороне червя от рук гонады. Рукава гонады изгибаются вокруг кишечника, а концы с делящимися ядрами находятся напротив вульвы. H) задняя рука гонады; (I) передняя рука. Можно вводить одну или обе руки. Для H, I, условности такие же, как в G. Шкала стержней = 50 мкм (B, D, F, H, I); 100 мкм (A, C, E, G). Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
- Храните вытянутые иглы в пластиковой чашке Петри 15 см с куском свернутой ленты, чтобы закрепить иглы и избежать скопления пыли на кончиках.
- Поместите каплю микроинъекционной смеси объемом 0,7 мкл на открытый конец вала. Повесьте иглу перпендикулярно полке с помощью свернутого куска ленты, чтобы заполнить конический вал смесью в течение 10 минут. Подготовьте по 2 иглы за один раз на случай, если первая не подействует.
6. Подготовка микроскопа и разрыв иглы
ПРИМЕЧАНИЕ: Микроинъекция использует перевернутый микроскоп с 5x и 40x объективами, оснащенный микроинжекторной установкой для управления движением иглы. Перевернутый микроскоп должен быть помещен на тяжелый стол или антивибрационный воздушный стол для снижения вибрационного шума. Игольчатый держатель микроинжектора подключен к газообразному азоту, который подает давление, необходимое для доставки смеси микроинъекций. Меньший рассекающий микроскоп поблизости используется для переноса червей.
- Установите давление в бензобаке на ~40-60 фунтов на квадратный дюйм для разрыва иглы и на ~30-50 фунтов на квадратный дюйм для микроинъекции, в зависимости от потока жидкости.
- На рассекающем микроскопе накройте осколок стекла на крышке микроинжекционной прокладки галогенуглеродным маслом с помощью стандартной платиновой червячной кирки.
- Поместите крышку микроинъекционной прокладки на микроинъекционный прицел и найдите осколок стекла, покрытый маслом. Выровняйте осколок стекла таким образом, чтобы край был перпендикулярен направлению иглы, чтобы служить поверхностью, используемой для разрыва иглы.
- Убедитесь, что игла не имеет пузырьков или мусора в коническом валу, используя рассекающий микроскоп. Затем закрепите иглу на 1-1,5 см в герметичный держатель.
- Расположите кончик иглы в центре поля зрения микроскопа на глаз. Затем при небольшом увеличении расположите кончик иглы в поле зрения, перпендикулярно стороне осколка стекла.
- Переключитесь на высокую мощность и выровняйте кончик иглы с краем стекла, приближаясь, но не касаясь его.
ПРИМЕЧАНИЕ: При вытягивании иглы сращиваются закрытыми. - Чтобы разбить кончик иглы, чтобы позволить жидкости течь, осторожно постучите ею по стороне куска стекла, применяя непрерывное давление от газа (дополнительный рисунок S1). Как только жидкость начнет течь, проверьте форму наконечника и убедитесь, что он острый с легко протекающей жидкостью.
ПРИМЕЧАНИЕ: Если жидкость течет слишком быстро или конец слишком тупой, черви будут повреждены во время микроинъекции (Видео 1 и Рисунок 3A-F). - Когда жидкость хорошо вытечет из иглы, переместите микроинъекционный слайд в рассекающий прицел и поместите капли 1-2 мкл галокарбонового масла на агаровую прокладку для размещения червей.
- Перенесите 20-30 молодых взрослых стронгилоидов на 2% пластину NGM без бактерий в течение не менее 5 минут, чтобы удалить лишние поверхностные бактерии и выбрать отдельных червей для микроинъекции. Добавляйте больше червей в пластину NGM по мере необходимости во время инъекции.
7. Микроинъекция стронгилоидов
- Используйте небольшое количество галогенуглеродного масла на червячной кирке, чтобы выбрать молодую взрослую самку Strongyloides с 1-4 яйцами в ее гонаде из 2% пластины NGM без бактерий.
- Переведите червя в крошечную каплю масла на агаровой прокладке. Используя червячную кирку, аккуратно расположите червя, чтобы он не сворачивался, а гонада была видна и легко доступна. Обратите внимание на направление гонады (рисунок 3G).
- Поместите червя в поле зрения микроинъекционного микроскопа. Убедитесь, что гонад находится на той же стороне, что и игла, и расположена так, чтобы игла соприкасалась с гонадой под небольшим углом (рисунок 3H, I).
- Поднесите кончик иглы в сторону червя в той же фокальной плоскости. Цельтесь в руку гонады около середины червя. Используйте микроинжектор, чтобы аккуратно вставить иглу в гонаду (видео 2).
- Немедленно надавите на иглу, чтобы аккуратно заполнить всю руку гонады раствором ДНК. Определите на глаз, когда было введено достаточно жидкости (видео 2).
ПРИМЕЧАНИЕ: Для заполнения гонады может потребоваться до 2 с. - Извлеките иглу и проверьте, закрывается ли рана.
ПРИМЕЧАНИЕ: Червь слишком поврежден, чтобы произвести потомство, если гонада выступает через стенку тела (Дополнительное видео S1). - Повторите с другим рукавом гонады, если он виден.
- Закончив инъекцию, быстро убедитесь, что игла не засорена, прикладывая давление кончиком иглы на агаровую прокладку. Перенесите слайд с введенным червем на рассекающий микроскоп.
- Чтобы восстановить введенного червя, сначала поместите несколько капель BU на червя, чтобы сбросить его с агаровой подушки.
- Соберите небольшое количество бактерий HB101 на червячной кирке. Прикоснитесь к червю с прилипшими бактериями на червячной кирке, чтобы удалить его из жидкости.
- Осторожно перенесите червя на восстановительную пластину, 2% пластину NGM, содержащую газон HB101.
ПРИМЕЧАНИЕ: Червь должен начать ползать в течение нескольких минут. - После того, как несколько самок были введены, добавьте несколько неинъектированных самцов из исходной пластины.
ПРИМЕЧАНИЕ: Минимум один самец на пять самок является хорошим исходным уровнем; избыток самцов является предпочтительным. - Повторяйте все шаги до тех пор, пока не будет введено достаточное количество самок для эксперимента.
- Оставьте взрослых на восстановительной пластине не менее чем на 1 ч после инъекции, чтобы позволить червям восстановиться и спариваться.
8. Извлечение и культивирование введенных стронгилоидов
- Собирайте фекалии на ночь у незараженных животных-хозяев, используя тот же протокол, что и для инфицированных животных.
- Смешайте непораженный кал с небольшим количеством древесного угля (отношение кала к древесному углю примерно 2 к 1 для этих пластин).
- Вылейте небольшое количество фекально-угольной смеси в 6 см чашку Петри, выстланную влажной фильтровальной бумагой. Следите за тем, чтобы смесь не касалась крышки блюда.
- Залить восстановительную пластину БУ. Используя пипетку, установленную на 3 мкл, перенесите червей к фекалиям в фекально-угольной пластине. Поместите червей непосредственно на кал, а не на древесный уголь.
- Убедитесь, что взрослые находятся на фекально-угольной пластине, используя рассекающий прицел.
- Чтобы культивировать червей, поместите пластину в увлажненную камеру, т. е. пластиковую коробку с плотно прилегающей крышкой, выстланную влажными бумажными полотенцами.
ПРИМЕЧАНИЕ: Через 2 дня будет смесь личиночных стадий. Через 5 дней большая часть личинок превратится в iL3s; останется несколько молодых личинок. Через 7 дней все личинки должны быть iL3s.
9. Сбор и скрининг личинок F 1 для восстановления трансгенов/нокаутов
- Используя установку Baermann, соберите личинок из мелкомасштабных фекально-угольных культивирующих пластин после инъекции. Чтобы получить как можно больше личинок, подождите не менее 2 часов, прежде чем извлекать червей из аппарата Baermann.
- Сконцентрируйте личинок в 15 мл центрифужной трубки, как на шагах 4.10-4.14, и переложите личинок в небольшое часовое стекло с BU.
- Если личинки будут использоваться для поведенческих экспериментов, используйте для скрининга 2% пластины NGM с густым газоном HB101.
- Перенесите 20-30 личинок на газон HB101.
ПРИМЕЧАНИЕ: Бактерии замедлят движение личинок. - Под флуоресцентным рассекающим микроскопом идентифицируют личинки, выражающие интересующий трансген. Используйте червячную кирку, чтобы выбрать трансгенных личинок и переместить их в небольшое часовое стекло с BU.
- Используйте новую пластину HB101 для скрининга еще одной небольшой партии личинок. Когда будет собрано достаточное количество личинок для экспериментального использования, обработайте пластины HB101 и избыточных червей разбавленным йодом (50% йода Люголя, разбавленного в воде) и выбросьте их как биологически опасные отходы. В качестве альтернативы, убейте лишних червей с помощью концентрированного очистителя питомника, содержащего алкилбензилхлориды аммония.
- Используйте червей немедленно или оставьте их в неглубоком часовом стакане в небольшом количестве БУ на ночь.
ПРИМЕЧАНИЕ: Черви могут стать гипоксическими, если жидкость слишком глубокая. Возможно, что оставление личинок в БУ на ночь может повлиять на определенное поведение; поэтому используют личинок для поведенческих экспериментов в течение 6 ч.
- Перенесите 20-30 личинок на газон HB101.
- Если личинки будут использоваться для микроскопии, а не поведенческих анализов, то обездвижить червей никотиновым параличом обратимо для скрининга.
- Используя лезвие бритвы, нанесите сетку на пластиковое дно 10-сантиметровой хемотаксисной пластины12 , чтобы было легче отслеживать расположение червей на пластине.
- Опустите ~3 мкл личинок в БУ в квадрат на сетке. Заполните столько квадратов, сколько необходимо. Не используйте те, которые находятся вблизи краев пластины, так как личинки могут ползти по бокам пластины.
- Добавьте 15-20 мкл капель 1% никотина в воду к каплям червя.
ПРИМЕЧАНИЕ: Через 4 минуты черви будут парализованы. - Скрининг червей с помощью флуоресцентного рассекающего микроскопа.
- Используйте червячную кирку для переноса трансгенных личинок в небольшое часовое стекло с 1-2 мл BU.
ПРИМЕЧАНИЕ: Личинки будут парализованы в течение нескольких часов и могут быть легко установлены на предметных стеклах микроскопа для микроскопии. Если оставить на ночь в BU, iL3s восстановится и может быть использован для некоторых анализов или инфекции хозяина млекопитающих. Тем не менее, никотиновый паралич и ночная инкубация в БУ могут влиять на определенное поведение.
Access restricted. Please log in or start a trial to view this content.
Результаты
Если эксперимент был успешным, личинкиF1 будут выражать трансгенный и/или мутантный фенотип, представляющий интерес (рисунок 4). Тем не менее, скорость трансформации сильно варьируется и зависит от конструкций, здоровья червей, условий культивирования после инъекц...
Access restricted. Please log in or start a trial to view this content.
Обсуждение
Этот протокол микроинъекции подробно описывает методы введения конструкций для трансгенеза и CRISPR/Cas9-опосредованного мутагенеза в S. stercoralis и S. ratti. Как для S. stercoralis , так и для S. ratti выживаемость после инъекции и скорость трансгенеза или мутагенеза зависят от нескольки...
Access restricted. Please log in or start a trial to view this content.
Раскрытие информации
Авторы заявляют об отсутствии конфликта интересов.
Благодарности
pPV540 и pPV402 были добрыми подарками от доктора Джеймса Лока из Университета Пенсильвании. Мы благодарим Астру Брайант за полезные комментарии к рукописи. Эта работа финансировалась премией Burroughs-Wellcome Fund Investigators in the Pathogenesis of Disease Award, премией Howard Hughes Medical Institute Faculty Scholar Award и National Institutes of Health R01 DC017959 (E.A.H.).
Access restricted. Please log in or start a trial to view this content.
Материалы
| Name | Company | Catalog Number | Comments |
| (−)-Nicotine, ≥99% (GC), liquid | Sigma-Aldrich | N3876-5ML | nicotine for paralyzing worms |
| 3" iron C-clamp, 3" x 2" (capacity x depth) | VWR | 470121-790 | C-clamp to secure setup to bench top |
| Agarose LE | Phenix | RBA-500 | agarose for slides |
| Bone char, 4 lb pail, 10 x 28 mesh | Ebonex | n/a | charcoal for fecal-charcoal cultures |
| Bone char, granules, 10 x 28 mesh | Reade | bonechar10x28 | charcoal for fecal-cultures (alternative to the above) |
| Coarse micromanipulator | Narishige | MMN-1 | coarse micromanipulator |
| Corning Costar Spin-X centrifuge tube filters | Fisher | 07-200-385 | microfilter column |
| Cover glass, 48 x 60 mm, No. 1 thickness | Brain Research Lab | 4860-1 | coverslips (48 x 60 mm) |
| Deep Petri dishes, heavy version with 6 vents, 100 mm diameter | VWR | 82050-918 | 10 cm Petri dishes (for fecal-charcoal cultures) |
| Eisco retort base w/ rod | Fisher | 12-000-101 | stand for Baermann apparatus |
| Eppendorf FemtoJet microinjector microloader tips | VWR | 89009-310 | for filling microinjection needles |
| Fisherbrand absorbent underpads | Fisher | 14-206-62 | bench paper (for prepping) |
| Fisherbrand Cast-Iron Rings | Fisher | 14-050CQ | Baermann o-ring |
| Fisherbrand tri-cornered polypropylene beakers | Fisher | 14-955-111F | Plastic beaker (for mixing) |
| Fisherbrand tri-cornered polypropylene beakers | Fisher | 14-955-111F | Plastic beaker (for catch bucket/water bucket) |
| Fisherbrand tri-cornered polypropylene beakers | Fisher | 14-955-111F | Plastic beaker (x2) (to make holder) |
| Gorilla epoxie in syringe | McMaster-Carr | 7541A51 | glue (to attach tubing) |
| Halocarbon oil 700 | Sigma-Aldrich | H8898-50ML | halocarbon oil |
| High-temperature silicone rubber tubing for food and beverage, 1/2" ID, 5/8" OD | McMaster-Carr | 3038K24 | tubing (for funnel) |
| KIMAX funnels, long stem, 60° Angle, Kimble Chase | VWR | 89001-414 | Baermann funnel |
| Kimberly-Clark Professional Kimtech Science benchtop protectors | Fisher | 15-235-101 | bench paper (for prepping) |
| Leica stereomicroscope with fluorescence | Leica | M165 FC | GFP stereomicroscope for identifying and sorting transgenic worms |
| microINJECTOR brass straight arm needle-holder | Tritech | MINJ-4 | microinjection needle holder |
| microINJECTOR system | Tritech | MINJ-1 | microinjection system |
| Mongolian Gerbils | Charles River Laboratories | 213-Mongolian Gerbil | gerbils for maintenance of S. stercoralis, male 4-6 weeks |
| Nasco Whirl-Pak easy-to-close bags, 18 oz | VWR | 11216-776 | Whirl-Pak sample bags |
| Nylon tulle (mesh) | Jo-Ann Fabrics | zprd_14061949a | nylon mesh for Baermann holder |
| Platinum wire, 36 Gauge, per inch | Thomas Scientific | 1233S72 | platinum/iridium wire for worm picks |
| Puritan tongue depressors, 152 mm (L) x 17.5 mm (W) | VWR | 62505-007 | wood sticks (for mixing samples) |
| QIAprep Spin Miniprep Kit (250) | QIAGEN | 27106 | QIAGEN miniprep kit |
| Rats-Long Evans | Envigo | 140 HsdBlu:LE Long Evans | rats for maintenance of S. ratti, female 4-6 weeks |
| Rats-Sprague Dawley | Envigo | 002 Hsd:Sprague Dawley SD | rats for maintenance of S. ratti, female 4-6 weeks |
| Really Useful Boxes translucent storage boxes with lids, 1.6 L capacity, 7-5/8" x 5-5/16" x 4-5/16" | Office Depot | 452369 | plastic boxes for humidified chamber |
| Shepherd techboard, 8 x 16.5 inches | Newco | 999589 | techboard |
| Stainless steel raised wire floor | Ancare | R20SSRWF | wire cage bottoms |
| StalkMarket compostable cutlery spoons, 6", white, pack of 1,000 | Office Depot | 9587303 | spoons |
| Stender dish, stacking type, 37 x 25 mm | Carolina (Science) | 741012 | watch glasses (small, round) |
| Stereomicroscope | Motic | K-400 LED | dissecting prep scope |
| Storage tote, color clear/white, outside height 4-7/8 in, outside length 13-5/8 in, Sterilite | Grainger | 53GN16 | plastic boxes for humidified chamber |
| Sutter P-30 micropipette puller | Sutter | P-30/P | needle puller with platinum/iridium filament |
| Syracuse watch glasses | Fisher | S34826 | watch glasses (large, round) |
| Thermo Scientific Castaloy fixed-angle clamps | Fisher | 05-769-2Q | funnel clamps (2x) |
| Three-axis hanging joystick oil hydrolic micromanipulator | Narishige | MM0-4 | fine micromanipulator |
| United Mohr pinchcock clamps | Fisher | S99422 | Pinch clamps (2x) |
| Vented, sharp-edge Petri dishes (60 mm diameter) | Tritech Research | T3308P | 6 cm Petri dishes (for small-scale fecal-charcoal cultures) |
| VWR light-duty tissue wipers | VWR | 82003-820 | lining for Baermann holder |
| watch glass, square, 1-5/8 in | Carolina (Science) | 742300 | watch glasses (small, square) |
| Whatman qualitative grade plain circles, grade 1, 5.5 cm diameter | Fisher | 09-805B | filter paper (for 6 cm Petri dishes) |
| Whatman qualitative grade plain circles, grade 1, 9 cm diameter | Fisher | 09-805D | filter paper (for 10 cm Petri dishes) |
| World Precision Instrument borosilicate glass capillary, 1.2 mm x 4 in | Fisher | 50-821-813 | glass capillaries for microinjection needles |
| X-Acto Knives, No. 1 Knife With No. 11 Blade | Office Depot | 238816 | X-Acto knives without blades to hold worm picks |
| Zeiss AxioObserver A1 | Zeiss | n/a | inverted microscope |
Ссылки
- Krolewiecki, A. J., et al. A public health response against Strongyloides stercoralis: time to look at soil-transmitted helminthiasis in full. PLoS Neglected Tropical Diseases. 7 (5), 2165(2013).
- Buonfrate, D., et al. The global prevalence of Strongyloides stercoralis infection. Pathogens. 9 (6), 468(2020).
- Castelletto, M. L., Gang, S. S., Hallem, E. A. Recent advances in functional genomics for parasitic nematodes of mammals. Journal of Experimental Biology. 223, Pt Suppl 1 206482(2020).
- Evans, T. C., et al. Transformation and microinjection. WormBook. , ed. The C. elegans Research Communnity (2006).
- Lok, J. B., Unnasch, T. R., et al. Transgenesis in animal parasitic nematodes: Strongyloides spp. and Brugia spp. WormBook. , ed. The C. elegans Research Communnity (2013).
- Shao, H. G., Li, X. S., Lok, J. B. Heritable genetic transformation of Strongyloides stercoralis by microinjection of plasmid DNA constructs into the male germline. International Journal for Parasitology. 47 (9), 511-515 (2017).
- Schafer, T. W., Skopic, A. Parasites of the small intestine. Current Gastroenterology Reports. 8 (4), 312-320 (2006).
- Stiernagle, T. Maintenance of C. elegans. The C. elegans Research Community, WormBook. , (2006).
- Gang, S. S., et al. Targeted mutagenesis in a human-parasitic nematode. PLoS Pathogens. 13 (10), 1006675(2017).
- Lok, J. B. Strongyloides stercoralis: a model for translational research on parasitic nematode biology. The C. elegans Research Community, WormBook. , (2007).
- Hawdon, J. M., Schad, G. A. Long-term storage of hookworm infective larvae in buffered saline solution maintains larval responsiveness to host signals. Proceedings of the Helminthological Society of Washington (USA). 58 (1), 140-142 (1991).
- Bargmann, C. I., Hartwieg, E., Horvitz, H. R. Odorant-selective genes and neurons mediate olfaction in C. elegans. Cell. 74 (3), 515-527 (1993).
- Junio, A. B., et al. Strongyloides stercoralis: cell- and tissue-specific transgene expression and co-transformation with vector constructs incorporating a common multifunctional 3' UTR. Experimental Parasitology. 118 (2), 253-265 (2008).
- Gang, S. S., et al. Chemosensory mechanisms of host seeking and infectivity in skin-penetrating nematodes. Proceedings of the National Academy of Sciences of the United States of America. 117 (30), 17913-17923 (2020).
- Bryant, A. S., et al. A critical role for thermosensation in host seeking by skin-penetrating nematodes. Current Biology. 28 (14), 2338-2347 (2018).
- Lok, J. B. Nucleic acid transfection and transgenesis in parasitic nematodes. Parasitology. 139 (5), 574-588 (2012).
- Shao, H., et al. Transposon-mediated chromosomal integration of transgenes in the parasitic nematode Strongyloides ratti and establishment of stable transgenic lines. PLoS Pathogens. 8 (8), 1002871(2012).
- Lok, J. piggyBac: a vehicle for integrative DNA transformation of parasitic nematodes. Mobile Genetic Elements. 3 (2), 24417(2013).
- Li, X., et al. Successful transgenesis of the parasitic nematode Strongyloides stercoralis requires endogenous non-coding control elements. International Journal for Parasitology. 36 (6), 671-679 (2006).
- Bryant, A. S., Hallem, E. A. The Wild Worm Codon Adapter: a web tool for automated codon adaptation of transgenes for expression in non-Caenorhabditis nematodes. G3. 3 (7), (2021).
- Crane, M., et al. In vivo measurements reveal a single 5'-intron is sufficient to increase protein expression level in Caenorhabditis elegans. Scientific Reports. 9 (1), 9192(2019).
- Han, Z., et al. Improving transgenesis efficiency and CRISPR-associated tools through codon optimization and native intron addition in Pristionchus nematodes. Genetics. 216 (4), 947-956 (2020).
- Adams, S., Pathak, P., Shao, H., Lok, J. B., Pires-daSilva, A. Liposome-based transfection enhances RNAi and CRISPR-mediated mutagenesis in non-model nematode systems. Scientific Reports. 9 (1), 483(2019).
- Dulovic, A., Puller, V., Streit, A. Optimizing culture conditions for free-living stages of the nematode parasite Strongyloides ratti. Experimental Parasitology. 168, 25-30 (2016).
- Harvey, S. C., Gemmill, A. W., Read, A. F., Viney, M. E. The control of morph development in the parasitic nematode Strongyloides ratti. Proceedings of the Royal Society B: Biological Sciences. 267 (1457), 2057-2063 (2000).
- Kim, A., Pyykko, I. Size matters: versatile use of PiggyBac transposons as a genetic manipulation tool. Molecular and Cellular Biochemistry. 354 (1-2), 301-309 (2011).
- Lok, J. B., Shao, H., Massey, H. C., Li, X. Transgenesis in Strongyloides and related parasitic nematodes: historical perspectives, current functional genomic applications and progress towards gene disruption and editing. Parasitology. 144 (3), 327-342 (2017).
- Farboud, B., Meyer, B. J. Dramatic enhancement of genome editing by CRISPR/Cas9 through improved guide RNA design. Genetics. 199 (4), 959-971 (2015).
- Cheong, M. C., et al. Identification of a nuclear receptor/coactivator developmental signaling pathway in the nematode parasite Strongyloides stercoralis. Proceedings of the National Academy of Sciences of the United States of America. 118 (8), 2021864118(2021).
- Nolan, T. J., Megyeri, Z., Bhopale, V. M., Schad, G. A. Strongyloides stercoralis: the first rodent model for uncomplicated and hyperinfective strongyloidiasis, the Mongolian gerbil (Meriones unguiculatus). Journal of Infectious Diseases. 168 (6), 1479-1484 (1993).
- Li, X., et al. Transgenesis in the parasitic nematode Strongyloides ratti. Molecular and Biochemical Parasitology. 179 (2), 114-119 (2011).
- Viney, M. E. Exploiting the life cycle of Strongyloides ratti. Parasitology Today. 15 (6), 231-235 (1999).
- Stoltzfus, J. D., Massey, H. C., Nolan, T. J., Griffith, S. D., Lok, J. B. Strongyloides stercoralis age-1: a potential regulator of infective larval development in a parasitic nematode. PLoS ONE. 7 (6), 38587(2012).
- Castelletto, M. L., Massey, H. C., Lok, J. B. Morphogenesis of Strongyloides stercoralis infective larvae requires the DAF-16 ortholog FKTF-1. PLoS Pathogens. 5 (4), 1000370(2009).
- Douglas, B., et al. Transgenic expression of a T cell epitope in Strongyloides ratti reveals that helminth-specific CD4+ T cells constitute both Th2 and Treg populations. PLoS Pathogens. 17 (7), 1009709(2021).
Access restricted. Please log in or start a trial to view this content.
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены