
需要订阅 JoVE 才能查看此. 登录或开始免费试用。
Method Article
在活动场域中对“日常记忆”进行建模的行为任务,以培养啮齿动物的同种异体中心表示
摘要
在活动场馆中,这种优化的 “日常记忆 ”协议的目标是采用一个稳定的基地,鼓励使用异心空间表示。这种动物模型为未来使用行为和生理技术研究事件记忆的形成和保留提供了一个有效的试验台。
摘要
活动场为研究学习和记忆提供了一个最佳平台。本文中描述的食欲性日常记忆任务为研究啮齿动物的情景记忆和空间记忆提供了一种强大的协议,这专门促进了同种异体着点记忆表示。在编码阶段,大鼠被训练寻找和挖掘食物,经过一段时间的延迟,大鼠可以选择在正确的位置找到奖励食物颗粒。在该协议中,有两个关键要素促进了以同种异体为中心的策略的使用:1) 大鼠在会话内和会话之间从不同的起始位置开始,2) 部署了一个稳定的基地,大鼠必须携带食物进食。通过这些修改,我们有效地鼓励啮齿动物使用同种异体中心空间表示来执行任务。此外,该任务为受试者内实验设计提供了一个很好的范例,并允许实验者纵不同的条件以减少可变性。与行为和生理技术结合使用,由此产生的啮齿动物模型为未来记忆形成和保留的研究提供了一个有效的试验台。
引言
为了研究学习和记忆的神经生物学,需要侵入性技术,这在人类中通常不可行。因此,一个多世纪以来,人们一直为实验动物设计行为协议来模拟各种形式的人类记忆。任务和设备的设计和选择是人类记忆有效模型成功的核心。已经开发了许多具有不同复杂性的范式,从简单的经典和工具条件协议 1,2,3 到 T 型迷宫 4、桡臂迷宫5、巴恩斯迷宫6、水迷宫7 和奶酪板迷宫8 等迷宫。然而,虽然这些任务捕捉了联想学习和空间导航的各个方面,但它们不能明确地用于研究瞬间事件的记忆表示(即情景式记忆)。而且,尽管新颖的物体识别9(new object recognition)和这种自发记忆任务的排列,如物体位置记忆10(object-place memory),为识别记忆提供了有价值的见解,但它们并没有测试对事件的显式回忆。为了满足这一需求,专门开发了事件竞技场,它的使用使对长期、成对关联记忆编码和回忆 11,12,13 以及发生在熟悉空间中的离散事件的编码和回忆 14,15,16,17,18 成为可能。后一个主题是本手稿的重点。
活动场馆是一个大型的方形开阔区域,啮齿动物在这里发生事件。竞技场的大小可以扩大以容纳大鼠或小鼠,并鼓励啮齿动物进入和探索。竞技场内发生的事件的一个典型示例是从特定位置的沙坑中寻找和检索食物。活动场地专为此类食欲任务而设计,其中大鼠或老鼠被训练寻找、寻找和挖掘食物。它利用了它们将食物带回黑暗环境的自然倾向,在这种情况下,该环境位于竞技场附近,然后它们在那里吃掉食物。经过最少的挖掘食物训练后,啮齿动物自然而然地接受了这项任务,并在编码试验和召回选择试验中表现良好,该试验在编码试验之后经过 30 分钟的短暂延迟。在选择试验中,有几个沙井(即挖掘位置)可用,但只有一个获得奖励。
可以在事件领域内执行不同的任务(例如,空间记忆、情景式记忆和配对关联学习)。鉴于对开发情景样记忆的有效模型的兴趣,开发了以下协议,其中可以找到食物的位置每天都会改变。在这项任务中,啮齿动物需要记住挖掘并成功取回食物奖励的事件最近发生在活动场地内的位置。下面概述的协议需要一个编码试验,其中大鼠每天在新的地方搜索沙井,然后在延迟后进行召回选择试验,其中最近编码的沙井位置得到奖励,而其他不同位置的替代沙井不包含可获取的食物。记住前一天食物的位置是没有帮助的:必须每天至少在一段时间内对正确的位置进行编码和记住。因此,我们引入了“日常记忆”一词来捕捉在这项任务中建模的记忆形式,我们作为人类每天都在使用它。人类日常记忆的一个例子是记住一个人把车停在购物中心的哪个地方(图 1A)或把眼镜放在房子周围。在这个协议中,所有场内和场外的线索都是稳定的,就像它们在我们日常生活的环境中(即家庭、办公室、停车场等)一样。因此,啮齿动物必须记住在熟悉的环境中最近发生某事的位置(图 1B)。该任务类似于水迷宫19 中的延迟匹配到地点 (DMP) 任务,但对其有所改进。作为一项食欲任务,它利用啮齿动物的自然行为来觅食20,而不是它们逃离水的愿望。然而,与水迷宫7 一样,没有区分正确位置和错误位置的局部线索;动物必须在不同的记忆延迟持续时间后使用回忆而不是识别来定位正确的 Sandwell 位置。

图 1:日常记忆。 (A) 人类的日常记忆。示意图显示停在停车场的绿色汽车。延误后,司机试图准确记住她把车停在哪里。(B) 动物的日常记忆。示意图显示了一只老鼠在活动场地内某个位置的沙井中挖掘和取回颗粒。延迟一段时间后,给大鼠一个选择试验,其中包含多个不正确的沙坑(灰色)和一个正确的沙坑(绿色)。 请单击此处查看此图的较大版本。
{kind=link}
活动舞台已经成功地用于对“日常记忆”的研究。这些记忆每天都会自动编码,保留在长期记忆中,但通常会在相对较短的时间内被遗忘。Bast 等人 14 显示了单调延迟依赖性事件记忆,其变化从短间隔后的优秀记忆到 24 小时后的机会水平。然而,可以通过后编码新颖性或多次编码试验延长试验间隔15,17 来成功增强记忆的保留。
活动场地用途广泛且相对无压力;不使用厌恶刺激。竞技场的大小及其容纳的任务可以适应大鼠14,15 和小鼠16。此外,作为一项陆基任务,它与 watermaze21 不同,适用于生理记录和钙成像研究。此外,根据 3R(减少、细化、替代)的原则,采用事件领域的研究需要更少的动物来获得统计功效,因为受试者内实验设计是可行的(其中每只动物作为自己的药物干预、光遗传学刺激等的对照)并且不需要厌恶刺激来激励。尽管初始训练需要更多的时间并且比新奇识别任务发生的时间更多,但一旦动物达到稳定、渐近的任务执行水平,药物、载体控制或光遗传学刺激等作可能会穿插相对较少的额外训练课程17.此外,表示的不同方面在事件领域受到直接实验控制,例如解决任务时采用的空间表示的性质。
表征问题涉及老鼠在记住最近事件发生的地方时所采用的心理框架18.他们记得食物的位置,还是只记得如何到达食物?大鼠可以使用同种异体中心(类似地图)或自我中心(以身体为中心)空间表示来解决竞技场内的食欲任务18。但是,为了控制和识别每个实验对象在执行任务时采用的空间策略,有不同的训练方案能够选择性地促进仅使用一种空间表示。通常,当老鼠将食物奖励带回它们开始当天试验的同一位置时,会采用基于自我中心的表示,这允许在来回奔跑期间有多次机会记住奖励位置。无论开始位置是每天更改还是保持不变,都可以采用此空间策略。相比之下,当需要老鼠将食物奖励带到竞技场一侧的固定基地位置时,倾向于同种异体表示,这与不断变化的起始位置不同。就大脑的存储容量而言,同种异体表示有很多优点。
在本文中,我们概述了 home-base 协议,该协议鼓励仅使用同种异体中心表示。我们为这项任务提供了具有代表性的结果,这些结果清楚地说明了在学习和记忆研究中使用这种 “日常记忆 ”的啮齿动物模型的优势,并强调了如何促进情景式空间记忆的同种异体中心表示。
研究方案
本文中描述的方法已获得爱丁堡大学伦理审查委员会的批准;它们符合 1986 年英国动物(科学程序)法案和 1986 年 11 月 24 日欧洲共同体理事会指令 (86/609/EEC) 的规定,该指令规定了实验动物的维护和在科学实验中的使用。
注意:下面概述的方案的实验对象是李斯特帽大鼠,但它可以适应其他啮齿动物品系。
1. 动物处理、住房和食品控制
- 让 Lister 罩雄性大鼠在到达后安定下来 1 周。在此期间,每天在笼子里轻轻抚摸和挠痒痒来照顾它们。一旦它们安顿下来,开始每天拿起它们大约 5 分钟。
- 在抵达时和每周每 2-3 天记录每只大鼠的体重。调整它们的食物摄入量,使每只大鼠的体重逐渐减少到其自由进食体重的 85%-90% 左右;这可以使用自由喂养大鼠的既定生长曲线来估计。在整个实验过程中,将大鼠保持在食物限制的体重范围内。
- 将大鼠置于 12 小时(亮)/12 小时(关灯)光照周期中,并在光照阶段(上午 7 点至晚上 7 点)进行所有实验。
2. 设置设备
- 实验室、控制室和活动竞技场
- 实验室和控制室是一个房间分成两部分,或者两个相邻的房间,由窗帘或门隔开,是本实验所必需的。
注意:这种分离将防止实验者在动物执行和学习这项复杂的行为任务时影响或扰乱动物。 - 将一个房间专门用于活动场地、静态环境线索和实验程序(即实验室),并使用另一个房间记录实验者对大鼠的表现(即控制室)。
注意:活动场地是一个方形的开阔场地区域,可以研究活动地点的关联(图 2A)。“事件竞技场”这个名字来源于这样一个事实,即这个设备是一个竞技场(即一个开放但受限的空间),“事件”在其中发生(例如,在沙井中挖掘埋藏的食物颗粒;图 2B)17,18.为了防止实验者过度劳累和受伤(即背部劳损),竞技场被抬高到地板以上 (~1 m)。 - 使用透明有机玻璃,搭建一个方形(160 厘米 x 160 厘米)的竞技场。竞技场的地板由一个 7 x 7 网格组成,由 49 块可移动的白色有机玻璃砖(20 厘米 x 20 厘米;图 2A,B)。修改五个带有中心孔(直径 6 厘米)的额外图块:这些图块将保持竞技场内的沙坑。这些修改后的切片的五个位置按每个会话的 sandwell 地图中概述的配置排列。
注意:五个 sandwell 位置的配置会随着每个新会话而变化(请参阅步骤 4.2)。 - 将活动场馆放置在指定的实验室中。设置场馆内提示:将两个具有不同触觉表面的独特地标(例如,一堆胶合的高尔夫球(30 厘米(高)x 11 厘米(宽)x 11 厘米(长))和一个黑色水瓶(22 厘米(高)x 9 厘米(深)))放置在场馆内的两个位置:第 4 行、第 2 列和第 4 行, 第 6 列(图 2C)。
- 在整个实验过程中,保持用于竞技场内提示的物体及其位置不变,但每天用 70% 乙醇清洁它们。
- 设置 3D 场馆外提示:在活动场馆的周边放置独特的地标(例如,带图案的球形灯笼(40 厘米 (d));红星灯笼(60 厘米 (W));蓝色灯笼(70 cm (h) x 35 cm (w))))和图案 - 位于实验室的中央(图 2D)。
- 在整个实验过程中,保持用于场外提示的对象及其位置不变。
- 实验室和控制室是一个房间分成两部分,或者两个相邻的房间,由窗帘或门隔开,是本实验所必需的。
- 黑匣子
- 为了让动物能够进入竞技场,用黑色有机玻璃建造四个相同的黑盒子(每个盒子长:30 厘米,宽:25 厘米,高:35 厘米; 图 2E)。每个黑匣子都应该在单长表面上有一个遥控滑动门。这将使实验者能够控制老鼠进入竞技场。
注意:黑色有机玻璃营造出黑暗的内部,与开阔场地活动场馆的明亮环境相比,老鼠更喜欢这种内部。 - 将这些黑框放在竞技场四面墙的中间位置。这些黑匣子将通过它们相对于实时视频源顶部的位置来识别 - 由摄像机捕获并由控制室中的计算机接收 - 使用活动场地的北(上)、东(右)、南(下)和西(左)基点。
- 允许老鼠从三个黑匣子之一进入竞技场,这被称为 startbox(例如,东、南和西; 图 2A,橙色矩形)。使用剩余的黑框(例如,北; 图 2A,蓝色矩形)作为大本营,老鼠将进入该基地吃掉它们从竞技场中取回的食物奖励(即颗粒)。
注意:任何黑盒位置(即北、东、南、西)都可以指定为大本营,但必须在整个实验中保持不变:其位置的稳定性对于成功鼓励同种异体中心空间表示至关重要。 - 在启动箱和基地中,放置两个小的、透明的、平坦的井,一个用于水,一个用于食物颗粒(在基地的情况下,这仅用于习惯阶段的奖励)并将锯末放入每个启动箱和基地。
- 为了让动物能够进入竞技场,用黑色有机玻璃建造四个相同的黑盒子(每个盒子长:30 厘米,宽:25 厘米,高:35 厘米; 图 2E)。每个黑匣子都应该在单长表面上有一个遥控滑动门。这将使实验者能够控制老鼠进入竞技场。
- 桑德韦尔
- 使用内径 (d) 为 6 cm、总深度 (h) 为 6 cm 的透明丙烯酸塑料制作沙坑,用于隐藏老鼠取回的食物奖励(即定位、挖掘并带到基地食用)。在距离顶部 4 cm 处插入一个球形多孔塑料碗。将沙坑插入竞技场内经过改造的瓷砖中。
注意:塑料碗为大鼠可以访问的奖励颗粒创建了一个可接近的部分(6 厘米(d) x 4 cm(h)),以及一个大鼠无法接近的部分(6 cm(d) x 2 cm(h))(图3A,B)。 - 在奖励沙井中,对于编码和召回选择试验,在可接近的部分放置四个 0.5 g 颗粒,在不可接近的部分放置八个食物颗粒(图 3C)。在无奖励的沙井中,将 12 个颗粒放置在无法接近的部分(图 3C)。
注意:奖励和非奖励沙井总共包含 12 个颗粒,并填充有专门制备的沙子,将颗粒隐藏在沙坑中。 - 用沙子和马萨拉粉的混合物(2.5 克马萨拉/2.5 公斤沙子)填充沙坑,以掩盖食物颗粒散发的任何气味。在每次会议开始时新鲜准备沙子/马萨拉混合物(图 3D)。
注意:步骤 2.3.2 和 2.3.3 旨在掩盖编码和选择试验期间从沙井发出的任何嗅觉伪影。这将确保大鼠寻找正确的沙坑位置以及它们由此产生的任务执行完全由它们对挖出食物的位置的记忆来指导,而不是由奖励沙井发出的任何气味线索指导,这可能会揭示食物奖励的存在。 - 在探针试验中,测试大鼠对先前奖励的单个沙井位置的记忆(即包含食物颗粒),将竞技场内的所有五个沙坑都设置为非奖励(即,可访问部分没有食物颗粒可用);包括正确的 Sandwell 位置。
注意:竞技场内的所有沙坑在其不可接近的部分都包含相同数量的弹丸 (n = 12)。
- 使用内径 (d) 为 6 cm、总深度 (h) 为 6 cm 的透明丙烯酸塑料制作沙坑,用于隐藏老鼠取回的食物奖励(即定位、挖掘并带到基地食用)。在距离顶部 4 cm 处插入一个球形多孔塑料碗。将沙坑插入竞技场内经过改造的瓷砖中。
- 整体设置和软件
- 使用壁挂式卤素灯 (115-125 lux) 将实验室的照明保持在中等亮度,并将室温保持在 19 至 23 °C 之间。
- 在实验室的活动场地上方安装电荷耦合器件相机,以记录和监测大鼠的运动和行为(图 4A)。该摄像机为相邻的控制室提供实时馈送,用于自定义视频捕获和自定义计算机软件(由爱丁堡大学的 P. A. Spooner 开发)。
- 使用用于为大鼠计时的定制计算机软件监测大鼠的运动(图 4B)。该程序控制每个黑匣子的门,允许实验人员从相邻的控制室远程管理大鼠进出竞技场的通道。记录每只动物的延迟以找到正确的沙坑,以及在选择和探测试验期间在每个沙坑挖掘所花费的时间。

图 2:活动场地和提示。 (A) 显示活动场地的示意图(缩写:N= 北,E= 东,S= 南,W= 西)。(B) 具有场内和场外提示的赛事场馆。(C) 两个 3D 场内球杆(从左到右):高尔夫球堆和圆柱形黑色瓶子。(D) 几个 3D 场馆外提示(从左到右):图案球形灯笼;红星灯笼;蓝灯笼。(E) 四个黑匣子中的一个位于每个活动场馆墙壁的中间位置。其中三个黑匣子用作起始框,在每次试验开始时为大鼠提供起始位置。第四个黑匣子是一个大本营,老鼠在这里吃掉它们从竞技场中取回的食物奖励。 请单击此处查看此图的较大版本。
{kind=link}

图 3:沙井。 (A) 显示空沙井的示意图,其中标记了可访问和不可访问的部分。(B) 一个空的沙井,有一个可进入的部分和不可进入的部分。(C) 示意图,说明奖励(左)和非奖励(右)砂井中的颗粒排列。奖励和非奖励沙井总共包含 12 个颗粒,并填充有专门制备的沙子,将颗粒隐藏在沙井中。(D) 显示奖励沙井准备工作的一系列照片,包括将颗粒正确放置在可进入的部分(步骤 1-4)。 请单击此处查看此图的较大版本。
{kind=link}

图 4:活动场地的实验装置。 (A) 显示实验室和控制室的实验装置的示意图。(B) 屏幕截图,显示通过自定义计算机软件查看的实验室实时馈送。定制的计算机软件允许实验人员远程控制启动箱门并提供其他测量。 请单击此处查看此图的较大版本。
{kind=link}
3. 习惯协议
注意:在适应过程中,老鼠被训练寻找沙井,挖掘食物奖励,并探索活动场地。
- 学会挖掘食物奖励
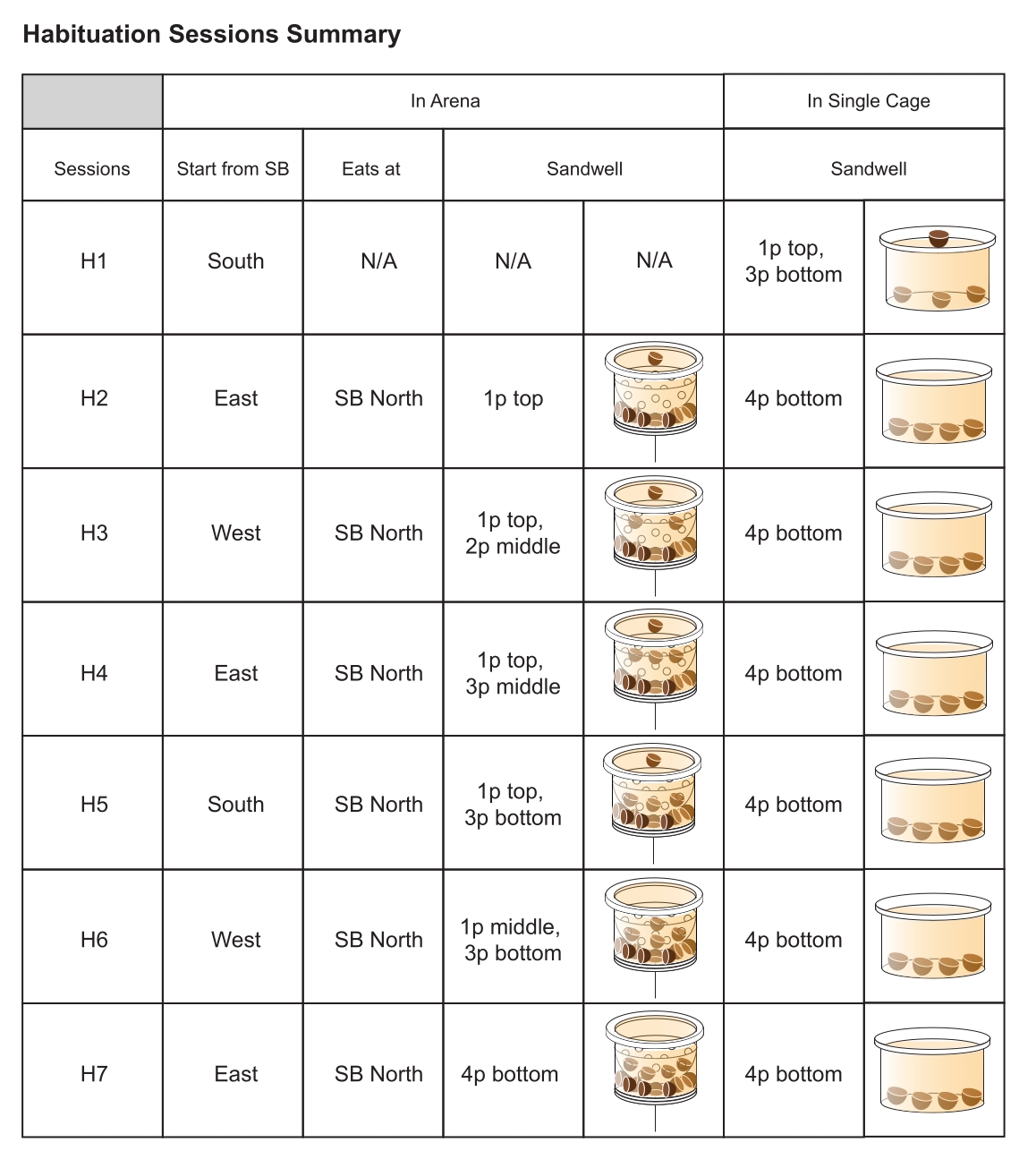
- 将一个装满沙子的小容器放在一个单独的笼子里。对于第一次(即 H1),在沙子表面下方添加三个 0.5 克的食物颗粒,并将一个食物颗粒放在上面。此后,将所有四个颗粒置于表面下(图 5)。
- 将每只大鼠放入带有一个沙坑的单独笼子中,为每只大鼠刷新沙坑。将老鼠留在单独的笼子里,直到它挖掘并成功从沙井中取出所有四个颗粒。
- 习惯化 第一节
- 将大鼠放入其中一个启动箱(例如,南)中 30 秒,然后将颗粒(0.5 克)放入基地(例如,北)指定用于食物的小平基井中。放置的食物颗粒作为奖励,鼓励老鼠去基地吃东西。
- 打开 startbox 门(例如,South)。当老鼠离开启动箱,进入竞技场并开始探索这个新环境时,关上门。
- 如果大鼠在 5 分钟内没有离开启动箱,则以两种方式之一进行干预。要么通过在启动箱门外拿着画笔来鼓励老鼠进入竞技场。一旦老鼠感兴趣,就将刷子进一步移动到竞技场中,让老鼠跟随它。一旦老鼠进入竞技场并与 startbox 门保持安全距离,关闭 startbox 门。
- 或者,将老鼠从 startbox 中取出,关闭 startbox 门,然后将老鼠放在竞技场中,直接位于它本应进入的 startbox 门外。如果大鼠没有动力并且没有很好地完成任务(例如,让启动箱无人协助、有效挖掘等),请检查大鼠的体重并计算其自由进食重量 (%)。
注意:如果自由摄食体重远高于 85%,大鼠可能并不饿;在这种情况下,其每日总食品津贴 (G) 可能需要进一步限制。 - 老鼠探索竞技场 10 分钟后,打开北黑匣子门(即 home-base)。如果老鼠在门打开后 5 分钟内没有进入,请将老鼠带出竞技场,关上 North 黑匣子的门,并将老鼠放在基地中。
- 在老鼠吃完训练开始时放置在基地中的颗粒后,将老鼠放回它的家笼中。
- 习惯化会议 2
- 在活动场地中放置一个沙坑,将所有四个食物颗粒(0.5 克/颗粒)埋在沙子表面下。此后每个会话更改此奖励沙井的位置。
注意:由于食物颗粒较大(0.5 克),大鼠更愿意将它们带到它们认为安全的环境(即黑暗环境)食用22。 - 将一个提示颗粒放入选定的启动框(即东)中指定用于食物的平坦井中,然后是一只老鼠。
- 当老鼠吃完提示颗粒时 - 大约 45 秒后 - 打开启动箱门(例如,东)。
- 一旦老鼠进入竞技场并与门保持安全距离,就关闭启动箱门。如果老鼠没有从启动箱中出来,请参考步骤 3.2.3-3.2.4。
- 让老鼠在沙坑中寻找第一个颗粒。要成功取回食物奖励,它必须在竞技场内现在存在的单个沙坑中挖掘。
- 一旦老鼠取回第一个颗粒,打开家门(即北)。然后老鼠应该找到并进入大本营吃掉它的奖励。如果老鼠开始在竞技场内吃颗粒,请轻轻地将其引导回基地吃掉颗粒。
注意:这很关键,因为必须鼓励每只老鼠在基地内进食;如果没有适当的训练,他们往往会回到 Trial 的 StartBox,他们从那里进入竞技场,吃东西。 - 老鼠在本垒完成第一颗子弹后,让它离开本垒并重新进入竞技场取回第二颗子弹。
- 取回第二个颗粒后,让老鼠再次找到基地以吃掉食物奖励。一旦老鼠进入大本营,就关上北方黑匣子的门。
- 老鼠吃完 Home-Base 中的第二个颗粒后,轻轻地将其从 Home-Base 中取出,然后将老鼠放回笼子里。
- 在活动场地中放置一个沙坑,将所有四个食物颗粒(0.5 克/颗粒)埋在沙子表面下。此后每个会话更改此奖励沙井的位置。
- 第 3-7 节
- 重复习惯化会话 2(步骤 3.3.1-3.3.9)五次,每次将颗粒埋在沙坑中更深(图 5)。在习惯化结束时,鼓励所有老鼠快速跑到竞技场内有奖励的沙井,连续收集可用的食物颗粒并将它们带回基地食用。

图 5:习惯化会话的设计。 从左列到右列:习惯化阶段 (H1-H7);用于每个会话的启动框(例如,H1:南启动框 (SB));老鼠需要吃食物奖励的地点(即 North home-base);可访问的 Pellet 在奖励的 Sandwell 中的位置(书面和插图形式;p = Pellet),该位置将放置在每个会话的指定 Sandwell 位置;颗粒在单个笼子中平基沙井中的位置(以书面和插图形式),旨在促进挖掘行为并加强大鼠在沙井中挖掘和获得食物奖励之间的联系。最后两列是指单个笼子(竞技场外)中的沙井。缩写: N/A= 不适用 请单击此处查看此图的较大版本。
{kind=link}
4. 主要训练方案
注意:每个主要训练课程包括两个记忆编码试验(E1、E2),然后在短暂的延迟(~30 分钟)后,进行一次召回选择试验 (C1)。在所有试验中,大鼠都需要从奖励的沙井中连续取回两个颗粒。找到每个颗粒后,老鼠应该找到并进入基地吃掉这个食物奖励。正确(即奖励)沙井的位置在所有大鼠的会话中被平衡(图 5)。
- 平衡措施
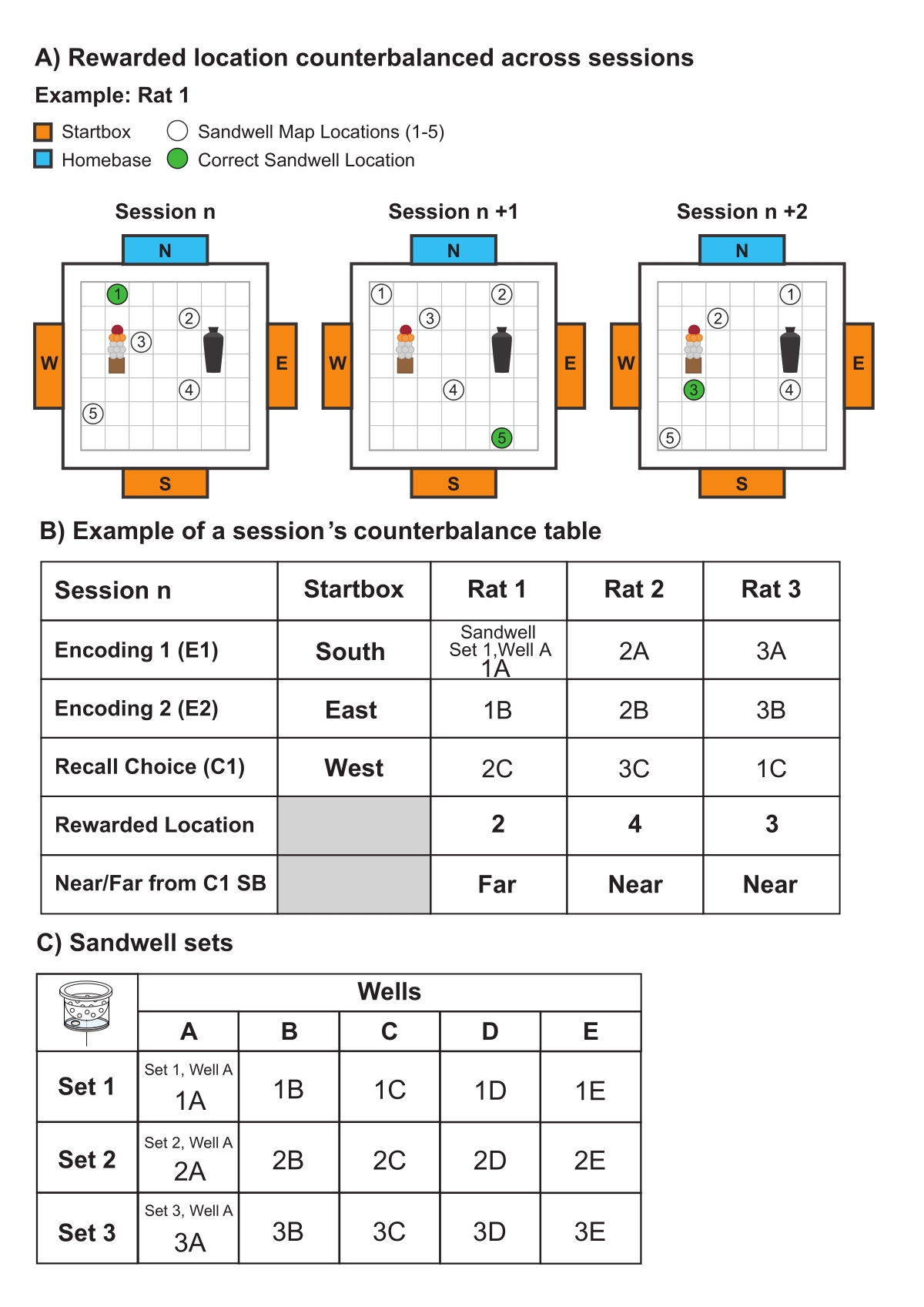
- 小心地平衡跨会话使用的 sandwell 位置和 startbox 顺序的顺序(图 6)。在每次会议之前,准备位置图(图 6A);为每只大鼠确定正确的 Sandwell 位置,该位置必须在会话中显着变化(图 6B);并创建平衡表(图 6B、C)和记录表(补充图 1)。
- 生产三个沙井组,每组有五个沙井(图 6C)。每组需要 5 个沙坑,因为在每次 Recall Choice 试验期间,竞技场中使用了 5 个 Sandwell 位置(1 个正确和 4 个错误),并且需要 3 个 Sandwell 集,以便每次试验使用的 Sandwell 可以在每个会话中交替。
- 在每次会话期间,使用一个 sandwell 集进行大鼠的编码试验(图 6B,C;编码 1:集合 1,井 A;编码 2:组 1,井 B)和另一个不同的沙井组(图 6B,C;召回选择:设置 2,井 C) 进行召回选择试验。
- 在每次会话中,为每只大鼠使用不同的沙井集组合(图 6B、C),并在会话中交替使用每只大鼠的沙井集组合。
- 编码试用
- 根据位置图和平衡表,将奖励沙井放在正确的位置(图 7)。切勿将竞技场内球杆位置、中心图块或四个启动框正前方的三个图块用作沙坑位置。
- 将一个颗粒,后跟一只大鼠,放入指定用于编码试验 1 (E1) 的启动框(例如,East)中;Pellet 将充当任务的提示。在开始试验之前,留出足够的时间 (~30 s) 让大鼠吃掉这个 Cue 颗粒。
- 按屏幕上的 Start 开始 按钮在内部视频捕获系统上录制试用版。
注意:重要的是要维护大鼠编码试验的记录,以便 (1) 研究透明度(即每只动物任务表现的原始证据),(2) 重新评分,以及 (3) 未来参考(即,探索和收集数据用于其他绩效衡量标准)。 - 使用自定义计算机软件远程打开 startbox 的门(图 4B)。
- 当老鼠进入竞技场时,在自定义计算机软件中启动计时器,并关闭启动箱门。
- 给老鼠 200 秒的时间寻找正确的沙坑,挖掘并取回它的第一个颗粒。如果大鼠在 200 秒后仍未找到正确的沙坑或它的第一个颗粒,请从沙子下方取回其中一个颗粒并将其放在顶部。如果大鼠未能访问正确的沙坑并在 200 秒后取回该颗粒,请使用刷子轻轻地将其引导至正确的沙坑。
- 一旦找到食物奖励,老鼠应该把它带到基地(例如,北方黑匣子)并在里面吃掉它。如果老鼠没有找到并进入基地,而是选择吃掉竞技场内的第一个颗粒,请迅速将老鼠从竞技场中取出并放置在基地中。
- 在基地吃完它的第一个颗粒后,让老鼠从基地进入竞技场,并从正确的沙坑中找到它的第二个颗粒。
- 取回它的第二个颗粒后,让老鼠找到并进入基地吃掉它。
- 一旦老鼠安全进入,就关上家门,给它足够的时间吃第二粒。
- 在自定义计算机软件上停止自定义视频捕获录制和计时器。按屏幕上 的 Stop 停止 自定义视频捕获软件上的按钮。然后,点击 Stop 停止 自定义计算机软件屏幕上计时器上的按钮。
- 当老鼠进食时,用浸泡在 70% 乙醇溶液中的布擦拭竞技场地板。在每次试用之间执行此作。
- 准备正确的沙井以进行编码试验 2 (E2),并将其放置在活动场地内的正确位置。
- 从 home-base 中取出 rat 并将其放入指定用于 E2 的启动框(例如 West)中。
注意:使用备用启动框对于有效鼓励大鼠仅使用同种异体中心空间解决方案来执行任务至关重要,因为大鼠不能依赖竞技场的静态视点或遵循它们之前的路径来成功找到正确的沙坑。相反,他们需要注意场内和场外的线索,这促进了同种异体中心编码。 - 重复步骤 4.3.2 到 4.3.12,然后将大鼠放回其家笼中。
- Recall Choice 试验
注意:每只大鼠的召回选择试验在第二次编码试验 (E2) 后 30-40 分钟进行,并向大鼠展示一个包含五个沙坑的竞技场。- 将奖励的沙井放置在分配给会话的正确位置,而四个未奖励的沙井放置在分配给会话和相关大鼠的四个错误位置(图 8A)。召回选择试验中 5 个沙坑的 Sandwell 位置图在每次会话中都会发生变化,并在会话之间进行平衡。
- 将包含四个可触及颗粒的沙井放在正确的位置。将四个额外的沙井(每个沙井都没有奖励,并且在可访问部分不包含颗粒)放在会话的沙井位置地图设置的错误位置。
- 将 cue pellet 和 rat 放入召回选择试验的启动框中(例如,South)。确保此起始位置(例如,C1:西)与两个编码试验中使用的位置(例如,E1:东,E2:南)不同。
- 使用内部视频捕获系统开始录制试用版。
注意:重要的是要保留大鼠召回选择试验的记录,以便 (1) 研究透明度(即每只动物任务表现的原始证据,可以作为论文补充材料的一部分提交),(2) 重新评分,以及 (3) 未来参考(即,探索和收集数据用于其他绩效测量)。 - 在自定义计算机软件上,选择要在此特定会话中使用的 sandwells(sandwell timers)匹配的计时器(图 4B)。
- 一旦老鼠吃掉了球杆颗粒(~30 秒),使用定制的计算机软件打开门。当老鼠离开启动箱时,关闭启动箱门,并在自定义计算机软件上启动计时器。
- 当老鼠在沙井中挖掘时,单击屏幕上的 Sandwell 图标以记录在每个沙井中挖掘所花费的时间。继续记录大鼠在访问的每个沙井中的挖掘时间,直到召回选择试验结束。
- 然后让老鼠找到并进入大本营吃掉这个食物奖励。
- 使用与编码试验相同的程序(参见步骤 4.2),在召回选择试验中从正确的沙井中检索第二个沉淀。
- 不要点击并记录老鼠在寻找第二个颗粒期间访问的每个沙井中的挖掘时间。仅记录在老鼠成功找到奖励的沙井并取回其第二个颗粒之前访问的沙井的顺序(使用分配给每个沙坑位置的数字 1-5)。这需要专注。

图 6:代表性的平衡。 (A) 示意图说明了大鼠(例如,大鼠 1)遇到的沙井位置图和正确的沙井位置在会话中如何变化。(B) 一个会话的平衡表示例(例如,会话 1)。在单个会话中的每个试验使用不同的启动框(即,编码从南启动框 (SB) 开始的试验 1 (E1)),但它们对每只动物的使用顺序相同(例如,大鼠 1-3)。用于正确位置的沙井(例如,位置 2、4、3)及其相关集合,在召回选择试验期间完全使用,在每次会话的试验(例如,编码 1、编码 2、召回选择)和执行任务的动物(例如,大鼠 1-3)之间被平衡。(C) 概述会话内和会话之间平衡的 Sandwell 集的表格。总共有 15 个沙井和三组(第 1-3 组)沙井,每组包含五口井 (A-E)。每只大鼠在每个编码和召回选择试验中使用不同的孔。例如,如图 6B 所示,大鼠 1 将在编码试验 1 中使用 Sandwell 1A,在编码试验 2 中使用 Sandwell 1B,在召回选择试验中使用 Sandwell 2C。 请单击此处查看此图的较大版本。
{kind=link}
5. 召回探针测试
- 使用与召回选择试验相同的设置,不同之处在于五个沙井中的任何一个都没有可接近的颗粒,包括会话之前奖励的正确沙坑位置(图 8A、B)。
注意:与召回选择试验一样,在探针测试期间,所有五个沙井都可用,大鼠可以自由挖掘它们选择的任何沙坑;然而,没有一个沙井包含可访问的食物奖励——相反,所有 12 个颗粒都存在于每个沙井的不可接近的部分(图 3C)。 - 将五个不包含可接近颗粒的沙坑放在竞技场中会话的沙井地图中提供的位置(图 8)。
- 将老鼠放入带有提示颗粒的启动箱中。使用同一会话的两个编码试验中未使用的起始位置。
- 在自定义计算机软件上设置沙井计时器以与会话的沙井图相对应(图 4B)。确保在自定义计算机软件上设置的 sandwell 计时器与会话的 sandwell 映射正确对应。
- 开始在内部视频捕获系统中录制探针试用。
注意:重要的是要保留大鼠召回探针试验的记录,以便 (1) 研究透明度(即每只动物任务表现的原始证据,可以作为论文补充材料的一部分提交),(2) 重新评分,以及 (3) 未来参考(即,探索和收集数据用于其他绩效衡量标准)。 - 一旦大鼠完成颗粒,使用自定义计算机软件远程打开启动箱门(图 4B)。
- 一旦老鼠进入竞技场并与门保持安全距离,关闭启动箱门并在自定义计算机软件中启动计时器。
- 在 120 秒探针试验期间,通过单击访问的每个沙井并按住鼠标,记录大鼠在 120 秒探针试验期间访问的每个沙井的挖掘时间和延迟,只要大鼠继续挖掘。这个 120 秒的倒计时从老鼠挖第一个沙井开始。
- 通过在 60 秒和 120 秒的时间戳处截取自定义计算机软件的屏幕截图,记录 60 秒和 120 秒的挖掘时间和延迟。
- 在 120 秒探针试验结束后,将三个颗粒放入正确的沙坑中(即编码试验中奖励沙坑的位置)以防止内存下降。大鼠需要取回这三个颗粒中的两个。一旦取回颗粒,老鼠就需要找到并进入基地吃掉它。
- 在 120 秒探针测试后,按下自定义计算机软件上的 屏幕上的停止 按钮。清除文件名,只记下老鼠的延迟,以检索放置在现在奖励的正确沙井位置的第一个和第二个颗粒。
注意:在正确位置优先挖掘用作内存索引:在编码试验中体验到的日常事件的良好记忆(即,遇到会话的正确 Sandwell 位置)表示在正确位置挖掘所花费的时间比在错误位置挖掘所花费的平均时间长。 - 在训练开始时安排召回率探测测试,以检查性能是否处于机会水平。此后,将探针测试安排在特定的时间间隔(例如,每六次会话),或仅在大鼠达到稳定的任务表现时安排:为了保证探针测试,它们的平均性能指数 (%) 需要连续三个会话达到 60% 或以上。平均性能指数在步骤 7.3.1 中定义。
6. 非编码对照测试
注意:非编码试验是一种控制措施,用于确定大鼠是否使用嗅觉伪影,而不是它们对正确沙坑位置的记忆来执行任务。顾名思义,“非编码对照测试”是指在召回选择试验之前没有进行编码试验;仅进行 Recall Choice 试验。预期是,在不允许对日常记忆事件的位置进行编码的情况下,大鼠在选择试验中的表现将处于机会水平。如果不是这种情况,并且大鼠在非编码对照测试中表现良好,则可能需要重新设计沙井及其可接近和不可接近的隔室。
- 按照第 4.3 节(步骤 4.3.1 到 4.3.10)中的说明执行召回选择试验。
7. 性能测量
注意:测量了几个参数, 补充图 1 显示了一个示例数据表。
- 沙井的选择
注意:选择定义为大鼠在召回选择和召回探针试验期间挖掘的沙井数量,直到并包括正确的沙坑。选择的最大可能值为 5,因为总共有 5 个沙井。- 在实验的每次试验(召回选择试验和召回探针试验)中,确定大鼠做出的选择数量:它是将前爪放在沙坑上还是放入沙坑中。如果老鼠跑过,或者只是在沙坑附近快速嗅探,这不被认为是一种选择。
- 在极少数情况下,当难以从视频监视器中判断老鼠是否做出选择(如上所述)时,请在试验结束时检查是否有任何挖掘痕迹:即沙子是否在沙井周围移动。如果有挖掘的证据,无论多么轻微,都要考虑这是一个选择。在沙坑前暂停而不挖掘仅被视为一次访问,不应被视为一种选择。
- 错误
注意:错误定义为大鼠在找到正确的沙坑之前访问的错误沙井(无奖励)的数量。选择定义为大鼠在召回选择和召回探针试验期间挖掘的沙井数量,直到并包括正确的沙井。最大错误数为 4,因为总共有 5 个沙井。- 使用以下公式计算误差:
错误 =(选项 - 1) - 当老鼠再次访问错误的沙井时,不要将此视为另一个错误,因为总共有五个沙井,因此最大错误数为 4。
- 使用以下公式计算误差:
- 性能指标 (PI)
注意:性能指数定义为大鼠在召回选择试验中找到正确的沙井之前所犯的错误数。使用 5 个沙井时,最多可能会发生 4 个错误。因此,五个沙井之间的机会水平是两个错误(即 50%)。- 使用以下公式计算性能指标:

- 当老鼠再次访问错误的沙井时,不要将此视为另一个错误,因为总共有五个沙井,因此最大错误数为 4。
- 使用以下公式计算性能指标:
- 延迟
注意:延迟定义为在正确的沙井开始挖掘之前经过的时间。- 测量从 rat 离开 startbox 到到达正确的 sandwell 的延迟。使用自定义计算机软件监控和记录延迟。
- 挖掘时间
- 在召回探针试验中测量大鼠在每个沙井(正确和错误的沙井)中的挖掘时间。
注意:对日常事件的良好记忆是由大鼠在 120 秒探针试验中挖掘正确的沙坑 (n = 1) 的比例大于它们在错误的沙坑中挖掘的平均时间 (n = 4) 来定义的。 - 使用以下公式计算正确和不正确:


- 在召回探针试验中测量大鼠在每个沙井(正确和错误的沙井)中的挖掘时间。
8. 避免无意的偏见
注意:在整个协议中实施了以下控制措施,以确保此日常记忆任务的可重复性和可靠性。
- 在会话之间平衡 Sandwell 位置。这避免了对活动竞技场的特定方面产生任何奖励偏差。
- 平衡沙井组以及这些组中的沙井,在每个会话中,跨会话和大鼠在正确的位置使用。这阻止了大鼠尝试遵循前一只大鼠试验中残留的任何残留气味试验。
- 在每次试验之间用浸泡在 70% 乙醇溶液中的布擦拭活动场地的地板;这将防止前一个 RAT 的路径影响后续任务性能。
结果
这种稳定的 home-base 协议已被用于成功训练大鼠使用同种异体表示来学习这项日常记忆任务。该协议中有两个重要元素。首先,动物在会话内和会话之间从不同的黑盒(例如,东、南和西)开始(图 7A)。每个会话有两个编码试验和一个召回选择试验(在某些情况下,或探测试验而不是选择试验),所有这些都从备用启动框开始。这鼓励动物在进入?...
讨论
人类会自动对日常生活中的单个事件进行编码。我们很容易回忆起一些事件,而忘记其他事件。上述类似情景的日常记忆协议为希望研究啮齿动物中此类记忆(情景记忆)的研究人员提供了一种强大的方法。由于该任务涉及从指定位置寻找和检索食物颗粒的日常行为,因此利用了啮齿动物觅食的自然本能。这项任务建立在一个合理的假设之上,即每天在一个特定但不断变...
披露声明
作者没有需要披露的利益冲突。
致谢
这项工作得到了医学研究委员会计划资助、欧洲研究委员会 (ERC-2010-AdG-268800-NEUROSCHEMA)、惠康信托高级研究者资助 (207481/Z/17/Z) 的支持。
材料
| Name | Company | Catalog Number | Comments |
| Camera | CCTVFirst | N/A | |
| Event Arena | University of Edinburgh (designed and built in house) | University of Edinburgh (designed and built in house) | Event arena for everyday memory task |
| Lister-hooded rats | Charles River UK | 603 | |
| Multitimer Labview | University of Edinburgh (designed and built in house) | University of Edinburgh (designed and built in house) | |
| Pneumatics, frames, screws of event arena | RS Components Ltd. | University of Edinburgh (P. Spooner) | Tools for building event arena |
| Sandwells | Adam Plastics (http://www.adamplastics.co.uk) | University of Edinburgh (P. Spooner) | Sandwells for arena |
| Startboxes | Adam Plastics (http://www.adamplastics.co.uk) | University of Edinburgh (P. Spooner) | |
| Video recording | Windows 10 computers with OBS software, Blackmagic Decklink Mini Recorder cards | N/A |
参考文献
- Pavlov, I. P. The work of digestive glands. Bristol Medico-Chirurgical Journal. 21 (80), 158-159 (1903).
- Thorndike, E. L. Animal intelligence: An experimental study of the associative processes in animals. Psychological Review. 5 (5), 551-553 (1898).
- Dickinson, A., Mackintosh, N. J. Reinforcer specificity in the enhancement of conditioning by posttrial surprise. Journal of Experimental Psychology: Animal Behaviour Processes. 5 (2), 162-177 (1979).
- Tolman, E. C., Gleitman, H. Studies in spatial learning: VII. Place and response learning under different degrees of motivation. Journal of Experimental Psychology. 39 (5), 653-659 (1949).
- Olton, D. S., Samuelson, R. J., Wagner, A. R. Remembrance of places passed: Spatial memory in rats. Journal of Experimental Psychology: Animal Behaviour Processes. 2 (2), 97-116 (1976).
- Barnes, C. A. Memory deficits associated with senescence: A neurophysiological and behavioral study in the rat. Journal of Comparative and Physiological Psychology. 93 (1), 74-104 (1979).
- Morris, R. G. M., Garrud, P., Rawlins, J. N. P., O'Keefe, J. Place navigation impaired in rats with hippocampal lesions. Nature. 297 (5868), 681-683 (1982).
- Kesner, R. P., Farnsworth, G., Kametani, H. Role of parietal cortex and hippocampus in representing spatial information. Cerebral Cortex. 1 (5), 367-373 (1991).
- Ennaceur, A., Delacour, J. A new one-trial test for neurobiological studies of memory in rats. 1: Behavioural data. Behavioural Brain Research. 31 (1), 47-59 (1988).
- Ennaceur, A., Neave, N., Aggleton, J. P. Spontaneous object recognition and object location memory in rats: the effects of lesions in the cingulate cortices, the medial prefrontal cortex, the cingulum bundle and the fornix. Experimental Brain Research. 113 (3), 509-519 (1997).
- Day, M., Langston, R. F., Morris, R. G. M. Glutamate-receptor-mediated encoding and retrieval of paired-associate learning. Nature. 424 (6945), 205-209 (2003).
- Tse, D., et al. Schemas and memory consolidation. Science (American Association for the Advancement of Science). 316 (5821), 76-82 (2007).
- Bethus, I., Tse, D., Morris, R. G. M. Dopamine and memory: modulation of the persistence of memory for novel hippocampal NMDA receptor-dependent paired associates. The Journal of Neuroscience. 30 (5), 1610-1618 (2010).
- Bast, T., da Silva, B. M., Morris, R. G. M. Distinct contributions of hippocampal NMDA and AMPA receptors to encoding and retrieval of one-trial place memory. The Journal of Neuroscience. 25 (25), 5845-5856 (2005).
- Wang, S. -. H., Redondo, R. L., Morris, R. G. M. of synaptic tagging and capture to the persistence of long-term potentiation and everyday spatial memory. Proceeding of the National Academy of Sciences. 107 (45), 19537-19542 (2010).
- Takeuchi, T., et al. Locus coeruleus and dopaminergic consolidation of everyday memory. Nature. 537 (7620), 357-362 (2016).
- Nonaka, M., et al. Everyday memory: towards a translationally effective method of modelling the encoding, forgetting and enhancement of memory. The European Journal of Neuroscience. 46 (4), 1937-1953 (2017).
- Broadbent, N., et al. A stable home-base promotes allocentric memory representations of episodic-like everyday spatial memory. The European Journal of Neuroscience. 51 (7), 1539-1558 (2020).
- Steele, R. J., Morris, R. G. M. Delay-dependent impairment of a matching-to-place task with chronic and intrahippocampal infusion of NMDA-antagonist D-AP5. Hippocampus. 9 (2), 118-136 (1999).
- Whishaw, I. Q., Coles, B. L. K., Bellerive, C. H. M. Food carrying: a new method for naturalistic studies of spontaneous and forced alternation. Journal of Neuroscience Methods. 61 (1), 139-143 (1995).
- Morris, R. G. M. Spatial localization does not require the presence of local cues. Learning and Motivation. 12 (2), 239-260 (1981).
- Whishaw, I. Q., Nicholson, L., Oddie, S. D. Food-pellet size directs hoarding in rats. Bulletin of the Psychonomic Society. 27 (1), 57-59 (1989).
- Dix, S. L., Aggleton, J. P. Extending the spontaneous preference test of recognition: Evidence of object-location and object-context recognition. Behavioural Brain Research. 99 (2), 191-200 (1999).
- Langston, R. F., Wood, E. R. Associative recognition and the hippocampus: differential effects of hippocampal lesions on object-place, object-context and object-place-context memory. Hippocampus. 20 (10), 1139-1153 (2010).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。